
Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species

1
Plant Research Team, Animal and Plant Research Department, Nakdonggang National Institute of Biological Resources, Sangju 37242, Republic of Korea
2
Department of Agriculture, Forestry and Bioresources, Plant Genomics & Breeding Institute, Research Institute of Agriculture and Life Science, College of Agriculture & Life Sciences, Seoul National University, Seoul 08826, Republic of Korea
*
Author to whom correspondence should be addressed.
Genes 2023, 14(10), 1914; https://doi.org/10.3390/genes14101914
Submission received: 13 September 2023
/
Revised: 3 October 2023
/
Accepted: 6 October 2023
/
Published: 7 October 2023
(This article belongs to the Special Issue Plant Plastid Genome)
Abstract
:Potamogetonaceae are aquatic plants divided into six genera. The largest genus in the family is Potamogeton, which is morphologically diverse with many hybrids and polyploids. Potamogetonaceae plastomes were conserved in genome size (155,863 bp–156,669 bp), gene contents (113 genes in total, comprising 79 protein-coding genes and 30 tRNA and 4 rRNA genes), and GC content (36.5%). However, we detected a duplication of the trnH gene in the IR region of the Potamogeton crispus and P. maakianus plastomes. A comparative analysis of Alismatales indicated that the plastomes of Potamogetonaceae, Cymodaceae, and Ruppiaceae have experienced a 6-kb inversion of the rbcL-trnV region and the ndh complex has been lost in the Najas flexilis plastome. Five divergent hotspots (rps16-trnQ, atpF intron, rpoB-trnC, trnC-psbM, and ndhF-rpl32) were identified among the Potamogeton plastomes, which will be useful for species identification. Phylogenetic analyses showed that the family Potamogetonaceae is a well-defined with 100% bootstrap support and divided into two different clades, Potamogeton and Stuckenia. Compared to the nucleotide substitution rates among Alismatales, we found neutral selection in all plastid genes of Potamogeton species. Our results reveal the complete plastome sequences of Potamogeton species, and will be helpful for taxonomic identification, the elucidation of phylogenetic relationships, and the plastome structural analysis of aquatic plants.
1. Introduction
Potamogetonaceae is an aquatic family comprising six genera (Althenia, Groenlandia, Lapilaena, Zannichillia, Stuckenia, and Potamogeton) and 110 species [1]. Potamogeton L. is used as food and a habitat for aquatic animals [2,3,4,5], and is divided into two subgenera Potamogeton and Coleogetonia [4]. However, the subgenus Coleogetonia was previously treated as an independent genus by Haynes [6], and was also treated as a synonym of genus Stuckenia Börner by Holub [7]. Potamogeton and Stuckenia differ in leaf shape, peduncle anatomy, and ploidy level [6,7]. Potamogeton species have highly similar morphological characteristics, such as leaves, seeds, and pollen, as well as various leaf shapes depending on the growing conditions [8,9,10]. Moreover, many Potamogeton species have undergone hybridization and polyploidization [3,11,12,13,14,15]. Hence, Potamogeton species are difficult to delimit taxonomically via morphological characteristics. Kaplan [11] reported that Potamogeton species comprise at least 50 hybrids worldwide. A previous molecular study using a 5S nuclear ribosomal array (5S-NTS) and plastid non-coding regions (psbA-trnH region and trnL intron) showed that Potamogeton was divided into two major groups: broad-leaved species and narrow-leaved species [16]. However, Iida et al. [17] indicated that the two major groups were not supported by the plastid non-coding region (trnT-L intron), and they proposed two alternative groups based on the shape of the submerged leaves and the anatomical features of the stem, and on linear submerged leaves and the presence of sub-epidermal bundles [17]. In general, the relationships among Potamogeton species are still unclear [16,17,18,19,20].
Plastids are important organelles for photosynthesis in plants, algae, and cyanobacteria [21,22]. Most plastid genomes (plastomes) have maternal inheritance and are often used in evolutionary and hybrid studies because of their unique characteristics [23,24]. The plastomes in angiosperms are highly conserved in terms of size, structure, order, and content. They usually comprise a pair of inverted repeat (IR) regions, a large single-copy (LSC) region, and a small single-copy (SSC) region. The genome size usually ranges from 150–160 kb, and comprises 113 genes, including, 79 protein-coding genes, 29 tRNA genes, and 4 rRNA genes [22,25]. However, recent studies have reported many variations, including rearrangement, inversion, repositioning, gene deletion, and IR, in the chloroplast genomes of the IR-lacking clade (IRLC) [26,27,28,29,30], Geraniaceae [31,32,33], Campanulaceae [34,35,36], and Orobanchaceae [37,38,39]. Therefore, plastomes have been widely used for phylogenomics [40,41,42], the development of molecular markers [43,44,45], and evolutionary studies [33,46,47,48].
Only two plastomes of Potamogetonaceae (Potamogeton perfoliatus and Struckenia pectiatus) have been reported so far [49,50], and Luo et al. [49] detected a 6 kb inversion including that in rbcL, atpB, atpE, trnM-CAU, and trnV-UAC in the P. perfoliatus plastome. In this study, we generated the complete plastomes of five Potamogeton species. We aimed to (1) characterize Potamogetonaceae plastomes, (2) identify divergent hotspot regions in the plastomes among Potamogetonaceae species, (3) compare plastomes among Alismatales species, (4) perform plastid phylogenomics within Potamogetonaceae and Alismatales, and (5) determine the nucleotide substitution rates among Alismatales.
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
Fresh leaves of Potamogeton species were sampled from a natural population in South Korea. All voucher specimens were deposited at the Nakdonggang National Institute of Biological Resources (NNIBR). Total genomic DNA was extracted using the DNeasy Plant Mini Kit (Qiagen Inc., Valencia, CA, USA).
2.2. DNA Sequencing, Assembly, and Annotation
Genomic DNA was sequenced using the Illumina Truseq Nano DNA kit (Illumina, San Diego, CA, USA) in accordance with the manufacturer’s protocol. Approximately 8.0 GB of raw data were generated for each species. P. perfoliatus, P. maackianus, P. crispus, P. wrightii, and P. distinctus were sequenced to produce 26,702,712–40,111,878 total reads from the 150 bp paired-end sequences. The raw reads were assembled using GetOrganelle software v. 1.7.6.1. [50]. The coverge of P. perfoliatus, P. maackianus, P. crispus, P. wrightii, and P. distinctus was 2271X, 3381X, 2169X, 1123X, and 1012X, respectively. Coding genes and tRNA were annotated using GeSeq [51] and tRNAscan-SE v. 2.0. [52], respectively. OrganellarGenomeDRAW (OGDRAW) v. 1.3.1 [53] was used to draw circular maps of the plastomes of Potamogeton species. All plastomes were submitted to GenBank under the accession numbers listed in Table 1.
2.3. Comparative Genomics, Divergence Hotspot and Repeat Analysis
The six completed chloroplast genome sequences were aligned using MAFFT [54]. Nucleotide diversity (Pi) was determined using DnaSP v. 6.0 [55]. The step size was set to 200 bp and the window length to 600 bp.
Repeat sequences, such as forward, palindromic, reverse, and complement sequences, were analyzed using REPuter [56] with a Hamming distance of 3 and a minimum repeat size of 30 bp. The simple sequence repeats (SSRs) were detected using MISA [57]. SSRs with a minimum number of repetitions of 10, 5, 4, 3, 3, and 3 for mono-, di-, tri-, tetra-, penta-, and hexa-nucleotides, respectively, were detected.
2.4. Phylogenetic and Substitution Rate Analysis
The plastomes of 20 Alismatales species including six Potamogeton species (two of P. perfoliatus) and one outgroup (Acorus gramineus) were used. The 65 shared-protein coding genes were aligned using MAFFT v.7.222 [54]. A maximum likelihood (ML) tree was constructed on Geneious Prime using RAxML v. 8.2.11 [58] and the GTRGAMMA model with 1000 bootstrap replicates. The Bayesian inference (BI) method for phylogenies was implemented with MrBayes [59]. Markov chain Monte Carlo (MCMC) analysis was run for one million generations. The trees were sampled every 1000 generations and the initial 25% were discarded as burn-in. The remaining trees were used to build a majority-rule consensus tree.
The dN and dS rates were estimated for each of the 48 shared protein-coding genes (>200 bp) using CODEML in PAML v. 4.8 [60]. The phylogenetic tree generated in the previous section was used as the constraint tree for all rate comparisons. Codon frequencies were determined in PAML using the F3 × 4 model, and gapped regions were excluded with the “clean data = 1” parameter option. The transition/transversion ratio and dN/dS values were estimated using the initial values of 2.0 and 0.4, respectively.
3. Results
3.1. Genome Features of Potamogetonaceae Species
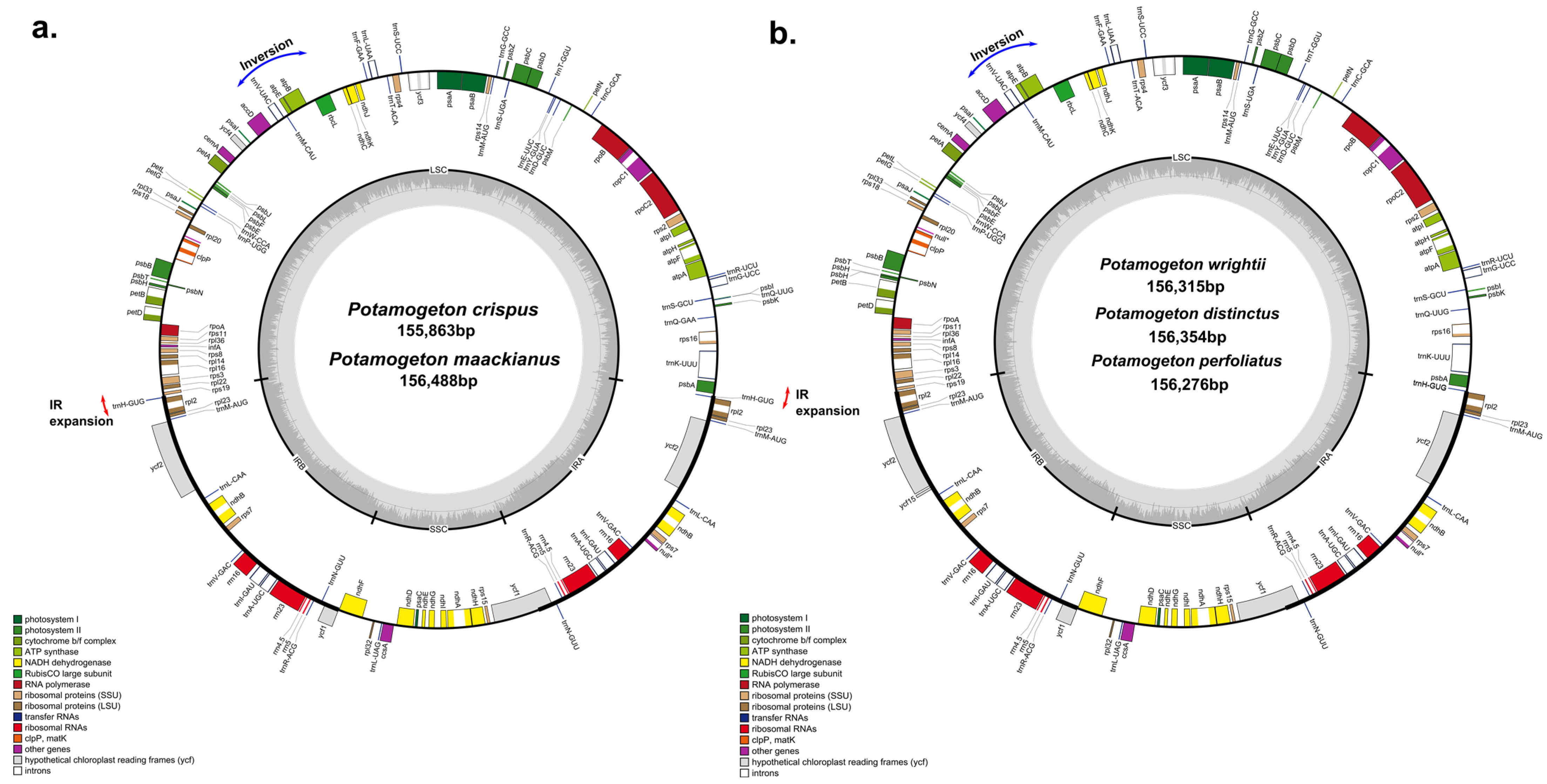
The plastomes of seven Potamogetonaceae species (six Potamogeton and one Stuckenia species) ranged from 155,863 bp (P. crispus) to 156,669 bp (S. pectinata). The seven Potamogetonaceae plastomes displayed a typical quadripartite structure, consisting of a pair of IRs (25,585–26,073 bp) separated by LSC (86,191–86,898 bp) and SSC (18,182–18,286 bp) regions (Figure 1, Table 1). The overall GC content was consistent (36.5%) in the seven Potamogetonaceae species. The seven Potamogetonaceae plastomes contained 113 genes, i.e., 79 protein-coding genes, 30 tRNA genes, and 4 rRNA genes. The IR regions of P. wrightii, P. distinctus, and P. perfoliatus had 17 genes (trnN-GUU, trnR-ACG, trnA-GAU, trnI-GAU, trnV-GAC, trnL-CAA, trnM-AUG, rrn4.5, rrn5, rrn23, rrn16, rpl2, rpl23, rps7, ndhB, ycf1, and ycf2), whereas those of P. crispus, P. maackianus, and S. pectinatus had 18 genes due to IR expansion, in which trnH gene was included in the IR region of the three plastomes.
The boundaries between the IR and single-copy (SC) regions of the six Potamogeton plastomes and the single Stuckenia plastome were compared (Figure 2). While the IRb/SSC boundary was similar in all Potamogeton species, the LSC/IRb boundary was located between rps19 and rpl2 regions in three species (P. distincus, P. wirghtii, and P. perfoliatus), whereas it was between rps19 and trnH in the P. crispus, P. maackianus, and S. pectinatus plastomes. Five species (P. distincus, P. wirghtii, P. perfoliatus, P. maackianus, and S. pectinatus) had overlapping ycf1 and ndhF genes, and the overlapped region between ycf1 and ndhF ranged from 8 to 29 bp in length. The IRb/SSC boundary in P. crispus did not overlap the ycf1 and ndhF genes. The SSC/IRa boundary was located in the ycf1 gene in all Potamogeton species. The IRa/LSC boundary was located at the trnH gene in three species (P. wrightii, P. distinctus, and P. perfoliatus) and three species (P. crispus, P. maackianus and S. pectinatus) had trnH genes in the IRa region. The 6 kb inversion previously reported from P. perfoliatus was found in all Potamogetonaceae plastomes (Figure 1).
3.2. Repeat and Simple Sequence Repeat (SSR) Analysis
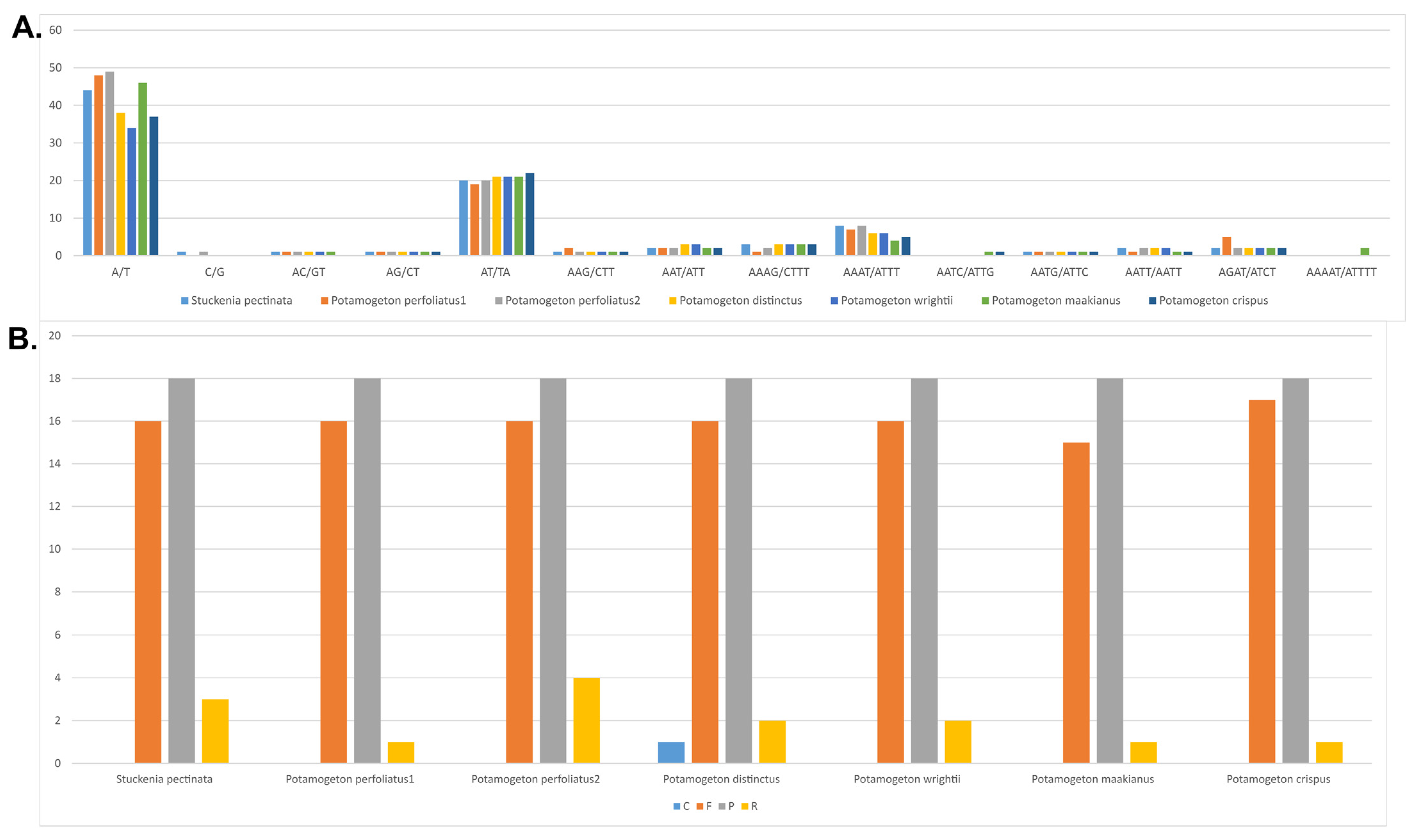
In total, 86, 90, 79, 75, 76, 86, and 88 SSRs were identified in the plastomes of P. perfoliatus (NC_029814), P. perfoliatus, P. distinctus, P. wrightii, P. crispus, P. maakianus, and S. pectinata, respectively (Figure 3A). Most SSRs were mononucleotide A/T repeats in all Potamogetonaceae plastomes. The plastomes of P. perfoliatus, P. maackianus, and S. pectinate had more mononucleotide repeats than did those of P. distinctus, P. wrightii, and P. crispus.
Four types of repeats (forward, reverse, complement, and palindromic) were found in the Potamogetonaceae plastomes (Figure 3B). The number of tandem repeats ranged from 34 (P. maakianus) to 38 (P. perfoliatus). Five Potamogeton species had 16 forward repeats, whereas P. cirspus had 17. Eighteen palindromic repeats were found in all the Potamogetonaceae plastomes. Three species (P. crispus, P. maakianus, and S. pectinate) had one reverse repeat, two species (P. wrightii and P. distinctus) had two reverse repeats, and P. perfoliatus had four reverse repeats in their plastomes. Complement repeats were found only in the P. distinctus plastome.
3.3. Divergence Regions in the Potamogetonaceae Plastomes
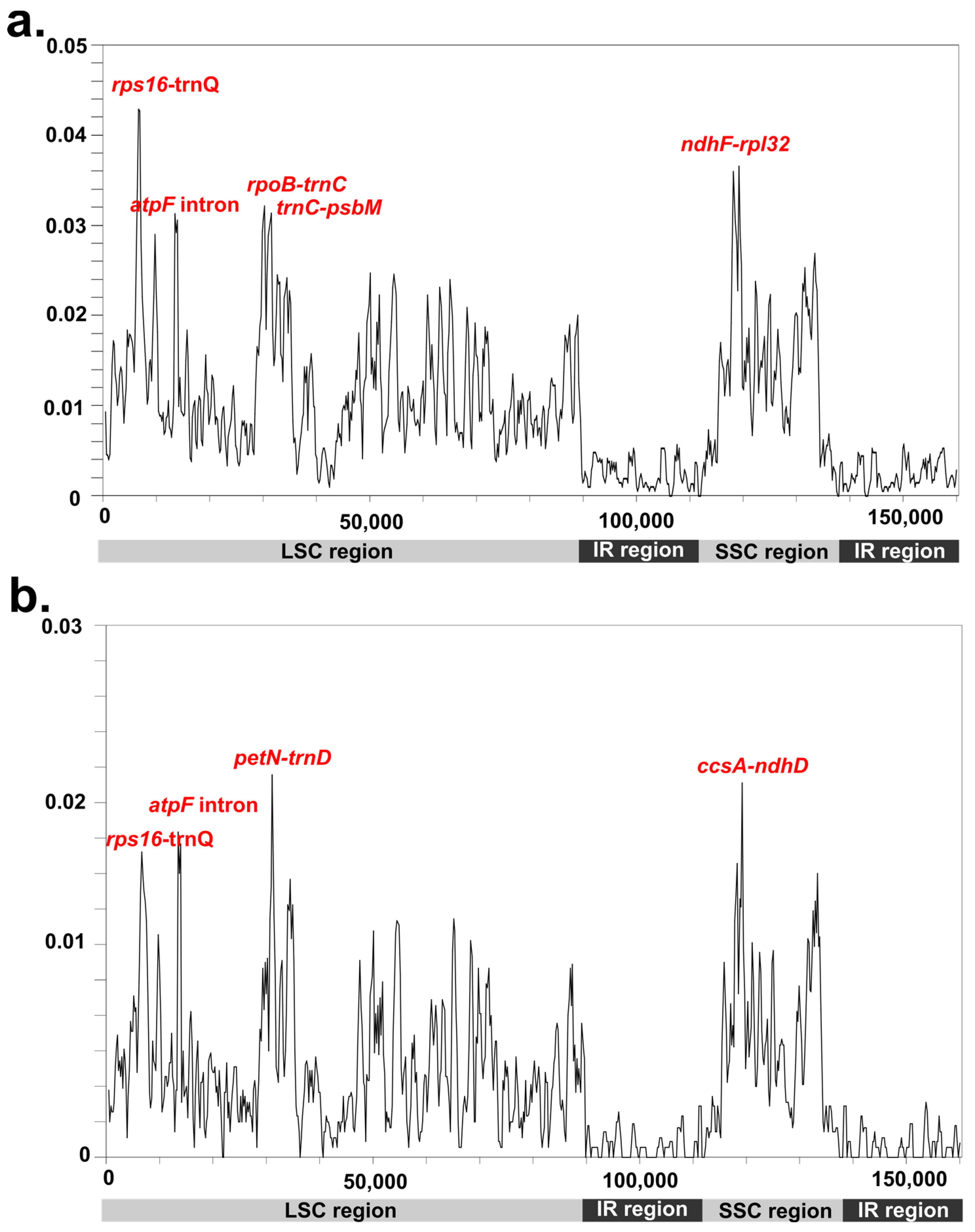
Whole plastomes within the family Potamogetonaceae (genera Potamogeton and Stuckenia) and within the genus Potamogeton were compared. The LSC and SSC regions showed a much higher variation than did the IR region. Among the Potamogetonaceae plastomes, the values ranged from 0 to 0.04286 (Figure 4a). Five regions, rps16-trnQ, rpoB-trnC, trnC-psbM, ndhF-rpl32, and the atpF intron showed higher nucleotide diversity (Pi) values than did other regions in their plastomes. Among the five Potamogeton plastomes, the Pi values ranged from 0 to 0.02156 (Figure 4b) and five regions (rps16-trnQ, the atpF intron, petN-trnD, ccsA-ndhD, and ycf1) were found to be divergence regions.
3.4. Comparative Analyses of the Plastomes among Alismatales
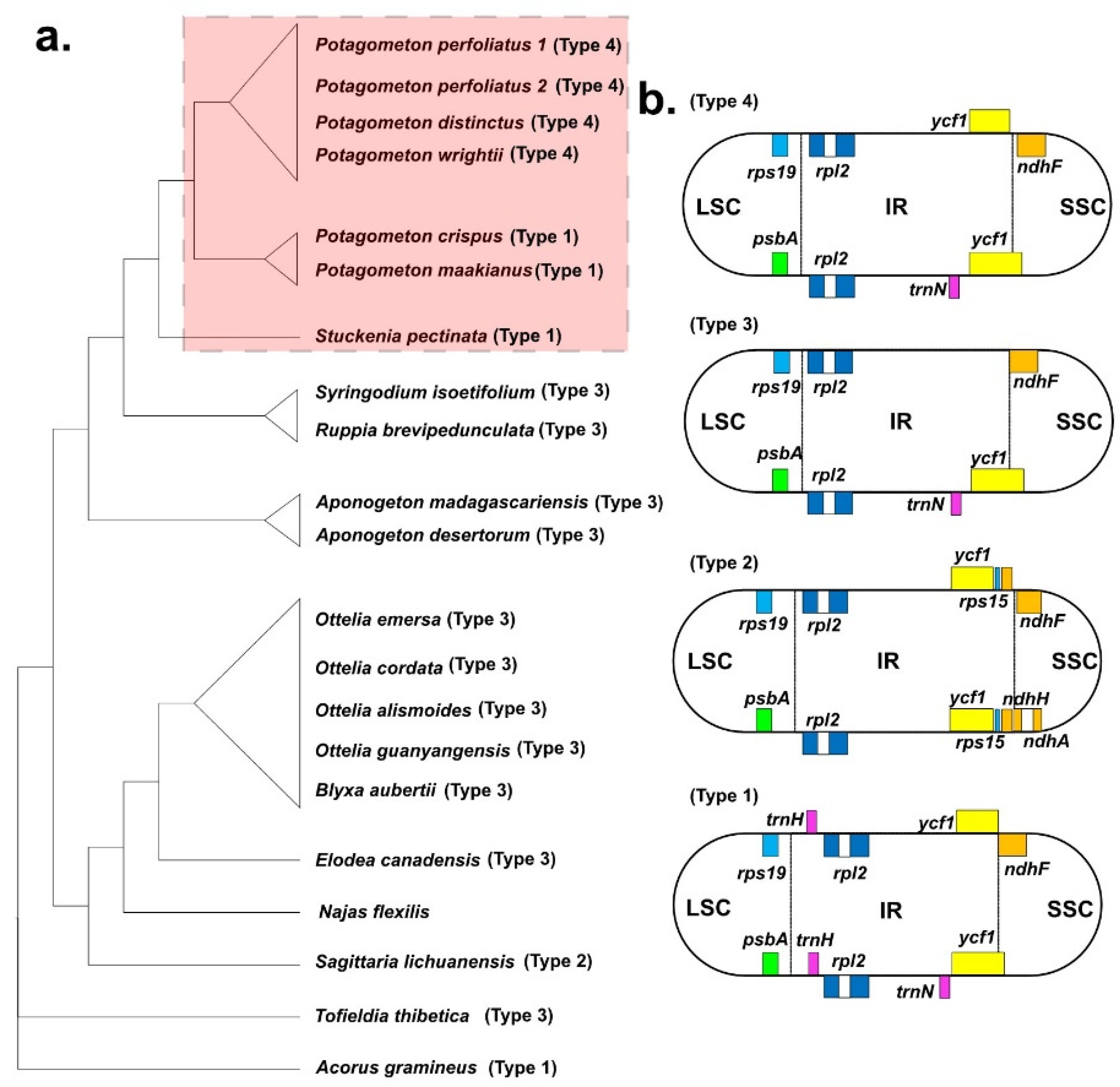
Twenty complete plastomes of Alismatales ranged from 154,516 bp (Aponogeton desertorum) to 179,007 bp (Sagittaria lichunanensis) (Table 1). The S. lichunanensis plastome had the longest LSC and a shorter SSC region compared with those of the plastomes of other species. The overall GC content was comparable, ranging from 36.8% (S. lichuanensis) to 38.2% (Najas flexilis). Most plastomes of Alismatales contained 113 genes (79 protein-coding genes, 30 tRNA genes, and 4 rRNA genes). However, the plastid NAD(P)H dehydrogenase (NDH) complex was lost in the N. flexilis plastome. The genome structure and gene order of the Alismatales plastomes are conserved. However, the S. lichiuanensis plastome revealed an inversion of the psbK-trnS region, and a 6 kb inversion was detected in the plastomes of Sytingodium isoetifolium, and Ruppia brevipedunculata (Figure 1 and Figure 5). Four types of IR/SC junctions were found in Alismatales (Figure 4). Most Alismatales plastomes had the type 3 junction. The LSC/IRa and LSC/IRb junctions were located in the rps19-rpl2 region and the psbA-rpl2 region, respectively. The SSC/IRa and SSC/IRb junctions were located in the ψycf1 and the ndhF-ycf1 region, respectively.
3.5. Phylogenetic Analysis
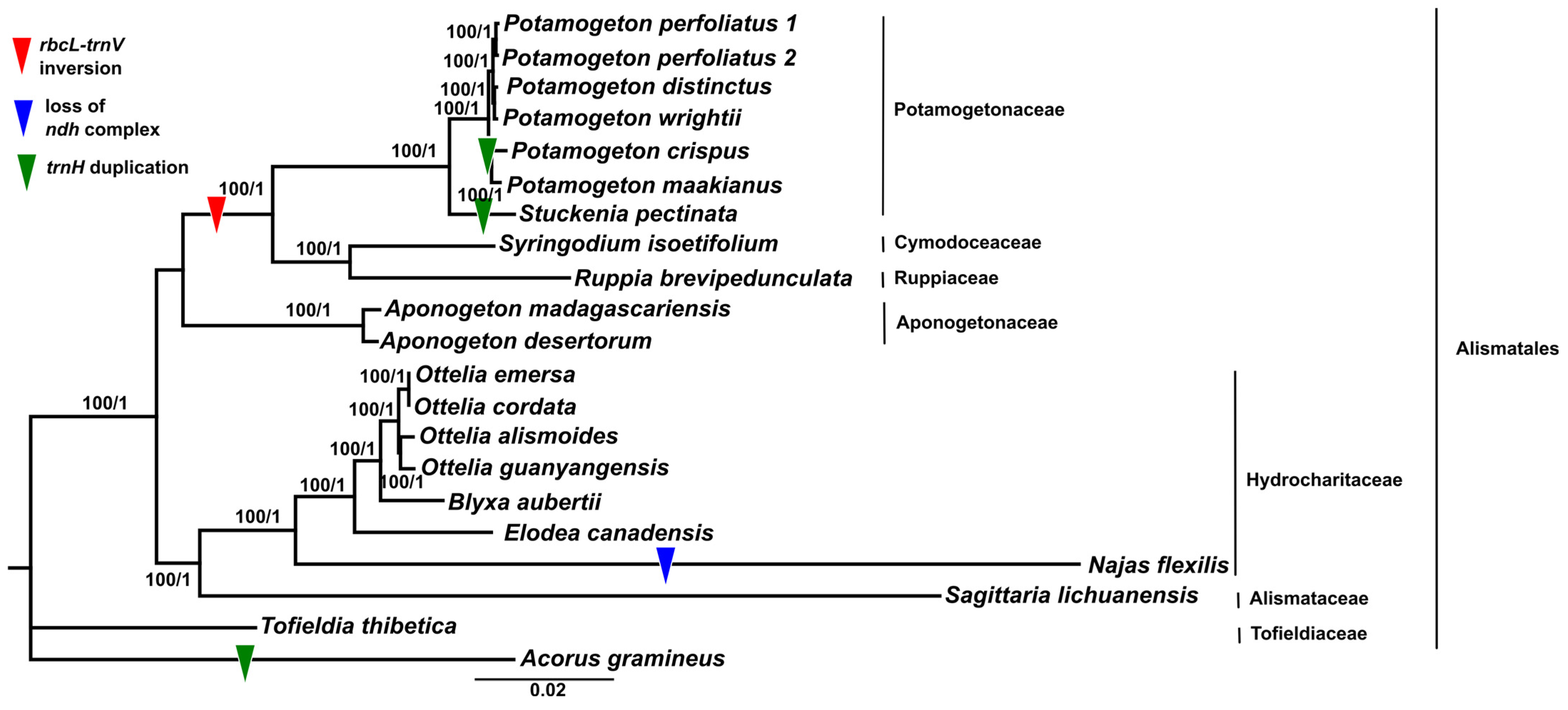
Due to the numerous gene losses in the cp genomes of Najas flexilis (13 genes including 11 ndh genes, infA, and psbH) and A. gramineus (accD), in total, 65 shared protein-coding genes were used to reconstruct the phylogenetic relationships of Alismatales (Figure 6; Supplementary Material Table S1). The topologies obtained from the ML and BI trees were consistent. As a result, Alismatales was divided into two groups: (1) Alismataceae (Sagittaria) and Hydrocharitaceae (Najas, Elodea, Blyxa, and Ottelia), and (2) Aponogetonaceae (Aponogeton), Ruppiaceae (Ruppia), Cymodaceae (Syringodium), and Potamogetonaceae (Stuckenia and Potamogeton). Both groups were strongly supported by a 100% bootstrap value. The genus Potamogeton was sister to the genus Stuckenia. Within the clade Potamogeton, P. maakianus and P. crispus formed a subclade, and P. wrightii, P. distinctus, and P. perfoliatus formed another subclade. The monophyly of the genus Potagometon and their subclades was supported by 100% bootstrap supporting values.
3.6. Nucleotide Substitution Rate Analyses
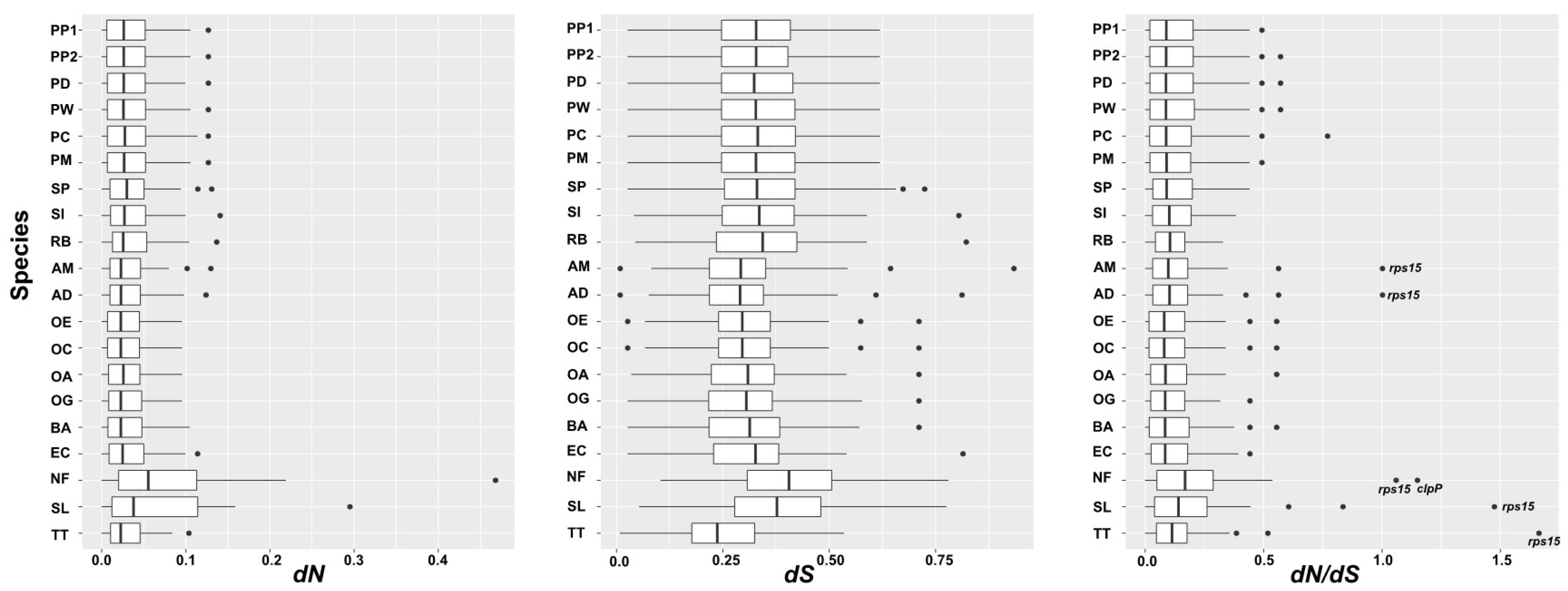
In total, 48 shared protein-coding genes of 21 Alismatales plastomes, including thpse of 7 Potamogetonaceae were used to estimate the synonymous (dS) and nonsynonymous (dN) nucleotide substitution rates (Figure 7). The mean dS of the Alismatales plastomes was higher than the mean dN. While the dN/dS ratio of most genes was less than 1, the dN/dS ratios of the rps15 gene in the plastomes of Tofieldia thibetica, S. lichuanensis, N. flexilis, A. desertorum, and A. madagascariensis were 1.663, 1.4746, 1.0597, 1.002, and 1.002, respectively, and that of ClpP in N. flexilis was 1.149. All Potamogetonaceae plastomes including Stuckia and Potamogeton showed a dN/dS ratio of less than 1. The dS of the Potamogetonaceae plastomes ranged from 0 to 0.77, the dN ranged from 0 to 0.1366, and the dN/dS ratio ranged from 0.0263 to 0.822.
4. Discussion
The six Potamogeton plastomes measured approximately 156 kb in length and consistent gene content (79 protein-coding genes, 30 tRNA genes, and 4 rRNA genes). Luo et al. [49] found a 6 kb inversion in the P. perfoliatus plastome. By comparing the genome structure of the Potamogetonaceae (Potamogeton and Stuckenia) and Alismatale plastomes, we found the 6 kb inversion in all Potamogetonaceae plastomes (Figure 1 and Figure 6). The 6 kb inversion was also detected in the Syringodium (Cymodaceae) and Ruppia (Ruppiaceae) plastomes (Figure 5). Previous studies [61,62,63,64] suggested that inversion is likely caused by the intramolecular recombination of the repeats, and Fullerton et al. [65] suggested that G + C content affects the plastome structure. However, we did not find any evidence that the repeats or G + C content were associated with the 6 kb inversion in the Alismatales plastomes (Figure 2, Table 1).
IR expansion and construction in plastomes have been reported from diverse angiosperm lineages, such as Passifloraceae, Fabiaceae, Geraniaceae, Campanulaceae, and Poaceae [31,35,66,67,68]. The IR/SC junctions of the Potamogetonaceae plastomes can be divided into two types: (1) trnH in the LSC/IR junction type and (2) trnH in the IR region type (Figure 2 and Figure 4). trnH duplication was previously reported in Elaeagnaceae [69] and monocots [70], and it was hypothesized to have IR expansion [70]. First, double-strand break (DSB) events occur within the IR regions, and then the free 3′ end of the broken strand is repaired against the homologous sequence in the IR regions. We speculated that the IRb region was expanded to the trnH gene, which was duplicated in the IRa region via a copy correction mechanism. The S. lichuanensis plastome showed different IR expansions, with DSB events occurring within the IRa region and an expansion of the IRa region to the SSC (ndhH) region. Subsequently, a duplication of the ndhH gene in the newly repaired IRb was achieved.
Previous studies have conducted phylogenetic analyses of the Alismatales [49,71,72]. We reconstructed a phylogenetic tree of 21 taxa in Alismatales based on 48 shared protein-coding gene sequences. Our results showed that Alismatales was divided into two groups: (1) the petaloid clade (Hydrocharitaceae and Alismataceae) and (2) the tepaloid clade (Potamogetonaceae, Cymodaceae, Ruppiaceae, and Aponogetonaceae). This result was consistent with the results from previous studies [71,72].
Previous studies have suggested that the genus Stuckenia should be distinguished from the genus Potamogeton [6,16,73], whereas Wiegleb and Kaplan [11] did not support this. Our study revealed that the genus Potamogeton is monophyletic and sister to Stuckenia pectinate (Figure 6). This result also supports the taxonomic treatment of two independent genera, Stuckenia and Potamogeton. The phylogenetic relationship of Potamogeton was not resolved in previous studies [16,17,20,74]. All molecular phylogenetic studies of Potamogeton used a few plastid genes, including psbA-trnH, trnT-L, the trnL intron, trnL-trnF, rbcL, and nrDNA ITS regions, resulting in uncertain species delimitations of Potamogeton species. For example, Iida et al. [17] showed that the genus Potamogeton could be divided into two groups. However, they failed to distinguish between Potamogeton gramineus and P. perfoliatus, because these two species formed a clade together. Moreover, Aykurt et al. [20] suggested a sister relationship between P. perfoliatus and P. nodosus, whereas P. perfoliatus formed a clade with P. richardsonii and the clade was sister to the clade of P. wrightii, P. distinctus, P. illinoensis, and P. nodosus [74]. Our study showed that P. perfoliatus was sister to the clade of P. wrightii and P. distinctus (Figure 6). Due to the insertion in the trnL-trnF region, P. crispus was distant from P. maackianus, but these two species were shown to have a sister relationship in the phylogeny by Ito et al. [74]. In this study, the sister relationship between P. crispus and P. maackianus was reconstructed and supported the findings of the previous study [74]. These differences may have been caused by the misidentification of the species owing to the similar morphological characteristics and hybridization of the species. Alternatively, they may have been caused by the insufficient molecular data for the phylogenetic reconstruction of the genus Potamogeton. Our study suggested five regions for the phylogenetic reconstruction and species identification of the genus Potamogeton. The five regions, rps16-trnQ, the atpF intron, petN-trnD, ccsA-ndhD, and ycf1 have not been used for Potamogeton so far, but the five regions will be used as valuable resources for determining the taxonomy and phylogenetics of the genus Potamogeton.
Synonymous and nonsynonymous rates provide evidence to understand the evolutionary forces in a gene [75]. The dN/dS ratio indicates the selection pressures. If the dN/dS ratio is higher than 1, the gene is under a positive selection, whereas if the ratio is less than 1, the gene is under a purifying selection [76]. We found two genes, rps15 (T. thibetica, S. lichuanensis, N. flexilis, A. desertorum, and Aponogeton madagascariensis) and clpP (N. flexilis), which are under positive selection in the Alismatales plastomes (Figure 7). It has been reported that the substitution rates of the gene in the IR regions were relatively lower than those in the two SC (LSC and SSC) regions. However, the genes in the expanded IR regions did not show any reduction in substitution rates [33,47,77]. Our study also showed that the substitution rates of the rps15 gene, which was relocated from the SSC to the IR regions in the S. lichuanensis plastome, were higher than those of the genes in the IR region (Figure 7). The Potamogeton plastomes exhibited a purifying selection (dN/dS < 1) for all genes.
5. Conclusions
Our study provides five newly assembled plastomes of the Potamogeton species. Comparative genomics of the Potamogeton plastomes showed that their genomes were conserved in genome size (155,863 bp–156,488 bp) and GC content (36.5%). However, IR boundary variation, such as trnH duplication, was detected in the P. crispus and P. maakianus plastomes. Five regions (rps16-trnQ, the atpF intron, rpoB-trnC, trnC-psbM, and ndhF-rpl32) were identified for phylogenetic and taxonomic studies of Potamogeton. Comparative genomics of the Alismatales plastomes showed that the Potamogetonaceae, Cymodaceae, and Ruppiaceae plastomes had the 6 kb inversion, and that the trnH duplication had occurred in the IR region of the Stuckenia pectinate plastome. Our phylogenomic studies using 48 shared protein-coding genes showed that Potamogetonaceae (Potamogeton and Stuckenia) was monophyletic. The synonymous and non-synonymous rates showed that the genes of the Potamogeteon plastomes were under purifying selection (dN/dS < 1).
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/genes14101914/s1. Table S1: The list of genes in 21 Alismatales chloroplast genomes.
Author Contributions
Conceptualization, K.C.; methodology, K.C., Y.H. and J.-K.H.; software, K.C. and J.-S.K.; validation, K.C. and J.-S.K.; formal analysis, K.C. and J.-S.K.; investigation, K.C., Y.H. and J.-K.H.; resources, K.C., Y.H. and J.-K.H.; data curation, K.C.; writing—original draft preparation, K.C.; writing—review and editing, K.C., Y.H., J.-K.H. and J.-S.K.; visualization, K.C.; supervision, K.C.; project administration, K.C.; funding acquisition, K.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant [NNIBR202301102] from the Nakddongang National Institute of Biological Resources, funded by the Ministry of Environment, Republic of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the newly sequenced sequences in this study are available from the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gob/ (accessed on 9 April 2023); accession numbers: OQ561449-OQ561453; see Table 1). Information for other samples used for phylogenetic analysis downloaded from GenBank can be found in Table 1.
Acknowledgments
We thank the Plant Research team members in Nakdonggang National Institute of Biological Resources for their assistance in sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201. [Google Scholar] [CrossRef]
- Hagström, J.O. Critical Researches on the Potamogeton; Kungliga Svenska Vetenskapsakademiens Handlinger; Almqvist & Wiksells: Stockholm, Sweden, 1916; Volume 55, pp. 1–281. [Google Scholar]
- Wiegleb, G. A redescription of Potamogeton distinctus including remarks on the taxonomy of the Potamogeton nodosus group. Plant Syst. Evol. 1990, 169, 245–259. [Google Scholar] [CrossRef]
- Wiegleb, G. The importance of stem anatomical characters for the systematics of the genus Potamogeton L. Flora 1990, 184, 197–208. [Google Scholar] [CrossRef]
- Xu, Z.; Chang, L. (Eds.) Potamogetonaceae. In Identification and Control of Common Weeds; Springer: Singapore, 2017; Volume 3, pp. 733–737. [Google Scholar]
- Les, D.H.; Haynes, R.R. Coleogeton (Potamogetonaceae), a new genus of Pondweeds. Novon 1996, 6, 389. [Google Scholar] [CrossRef]
- Holub, J. Stuckenia Börner 1912—The correct name for Coleogeton (Potamogetonaceae). Preslia 1997, 69, 361–366. [Google Scholar]
- Kaplan, Z. Phenotypic plasticity in Potamogeton (Potamogetonaceae). Folia Geobot. 2002, 37, 141–170. [Google Scholar] [CrossRef]
- Kaplan, Z.; Symoens, J.-J. Taxonomy, distribution and nomenclature of three confused broad-leaved Potamogeton species occurring in Africa and on surrounding islands. Bot. J. Linn. Soc. 2005, 148, 329–357. [Google Scholar] [CrossRef]
- Wafai, B.A.; Khuroo, A.A. Fruit morphology of the genus Potamogeton L. in Kashmir Himalaya and its utility in taxonomic delimitation. J. Asia-Pac. Biodivers. 2017, 10, 274–278. [Google Scholar]
- Wieglet, G.; Kaplan, Z. An account of the species of Potamogeton L. (Potamogetonaceae). Folia Geobot. 1998, 33, 241–316. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J. Erroneous identities of Potamogeton hybrids corrected by molecular analysis of plants from type clones. Taxon 2011, 60, 758–766. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J.; Hellquist, C. Potamogeton × jacobsii (Potamogetonaceae) from New South Wales, Australia—The first Potamogeton hybrid from the Southern Hemisphere. Telopea 2011, 13, 245–256. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J. Molecular identification of hybrids from a former hot spot of Potamogeton hybrid diversity. Aquat. Bot. 2013, 105, 34–40. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J.; Bambasová, V.; Hellquist, C.B. The endangered Florida pondweed (Potamogeton floridanus) is a hybrid: Why we need to understand biodiversity thoroughly. PLoS ONE 2018, 13, e0195241. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, C.; De Laet, J.; Haynes, R.R.; Aagesen, L.; Keener, B.R.; Albert, V.A. Molecular phylogenetics of an aquatic plant lineage, Potamogetonaceae. Cladistics 2006, 22, 568–588. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Kosuge, K.; Kadono, Y. Molecular phylogeny of Japanese Potamogeton species in light of noncoding chloroplast sequences. Aquat. Bot. 2004, 80, 115–127. [Google Scholar] [CrossRef]
- Wang, Q.D.; Zhang, T.; Wang, J.B. Phylogenetic relationships and hybrid origin of Potamogeton species (Potamogetonaceae) distributed in China: Insights from the nuclear ribosomal internal transcribed spacer sequence (ITS). Plant Syst. Evol. 2007, 267, 65–78. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Yang, C.-F.; Chen, J.-M.; Guo, Y.-H. Identification of hybrids in broad-leaved Potamogeton species (Potamogetonaceae) in China using nuclear and chloroplast DNA sequence data. Plant Syst. Evol. 2010, 287, 57–63. [Google Scholar] [CrossRef]
- Aykurt, C.; Fehrer, J.; Sarı Yol, D.; Kaplan, Z.; Bambasova, V.; Deniz, İ.G.; Aydemir, E.; İmir, N. Taxonomic treatment and phylogenetic analysis of the family Potamogetonaceae in Turkey. Taxon 2020, 69, 1172–1190. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Dobrogojski, J.; Adamiec, M.; Luciński, R. The chloroplast genome: A review. Acta Physiol Plant. 2020, 42, 98. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Yang, C.-F.; Chen, J.-M.; Guo, Y.-H.; Kadiri, A.B. Using DNA-based techniques to identify hybrids among linear-leaved Potamogeton plants collected in China. J. Syst. Evol. 2010, 48, 265–270. [Google Scholar] [CrossRef]
- Li, S.; Duan, W.; Zhao, J.; Jing, Y.; Feng, M.; Kuang, B.; Wei, N.; Chen, B.; Yang, X. Comparative analysis of chloroplast genome in Saccharum spp. and related members of ‘Saccharum Complex’. Int. J. Mol. Sci. 2022, 23, 7661. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Cai, Z.; Guisinger, M.; Kim, H.G.; Ruck, E.; Blazier, J.C.; McMurtry, V.; Kuehl, J.V.; Boore, J.; Jansen, R.K. Extensive reorganization of the plastid genome of Trifolium subterraneum (Fabaceae) is associated with numerous repeated sequences and novel DNA insertions. J. Mol. Evol. 2008, 67, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-S.; Choi, B.-H. The distinct plastid genome structure of Maackia fauriei (Fabaceae: Papilionoideae) and its systematic implications for genistoids and tribe Sophoreae. PLoS ONE 2017, 12, e0173766. [Google Scholar] [CrossRef]
- Duan, L.; Li, S.J.; Su, C.; Sirichamorn, Y.; Han, L.N.; Ye, W.; Loc, P.K.; Wen, J.; Compton, J.A.; Schrire, B.; et al. Phylogenomic framework of the IRLC legumes (Leguminosae subfamily Papilionoideae) and intercontinental biogeography of tribe Wisterieae. Mol. Phylogenet. Evol. 2021, 163, 107235. [Google Scholar] [CrossRef]
- Wu, S.; Chen, J.; Li, Y.; Liu, A.; Li, A.; Yin, M.; Shrestha, N.; Liu, J.; Ren, G. Extensive genomic rearrangements mediated by repetitive sequences in plastomes of Medicago and its relatives. BMC Plant Biol. 2021, 21, 421. [Google Scholar] [CrossRef]
- Son, O.; Choi, K.S. Characterization of the chloroplast genome structure of Gueldenstaedtia verna (Papilionoideae) and comparative analyses among IRLC species. Forests 2022, 13, 1942. [Google Scholar] [CrossRef]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The complete chloroplast genome sequence of Pelargonium × hortorum: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef]
- Blazier, C.J.; Guisinger, M.M.; Jansen, R.K. Recent loss of plastid-encoded ndh genes within Erodium (Geraniaceae). Plant Mol. Biol. 2011, 76, 263–272. [Google Scholar] [CrossRef]
- Weng, M.-L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef]
- Cosner, M.E.; Jansen, R.K.; Palmer, J.D.; Downie, S.R. The highly rearranged chloroplast genome of Trachelium caeruleum (Campanulaceae): Multiple inversions, inverted repeat expansion and contraction, transposition, insertions/deletions, and several repeat families. Curr. Genet. 1997, 31, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Cosner, M.E.; Raubeson, L.A.; Jansen, R.K. Chloroplast DNA rearrangements in Campanulaceae: Phylogenetic utility of highly rearranged genomes. BMC Evol. Biol. 2004, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Cheon, K.-S. Complete chloroplast genome sequence of Adenophora racemosa (Campanulaceae): Comparative analysis with congeneric species. PLoS ONE 2021, 16, e0248788. [Google Scholar] [CrossRef]
- Wicke, S. Genomic Evolution in Orobanchaceae. In Parasitic Orobanchaceae; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 267–286. [Google Scholar]
- Cusimano, N.; Wicke, S. Massive intracellular gene transfer during plastid genome reduction in nongreen Orobanchaceae. New Phytol. 2016, 210, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Park, S. Complete plastid and mitochondrial genomes of Aeginetia indica reveal intracellular gene transfer (IGT), horizontal gene transfer (HGT), and cytoplasmic male sterility (CMS). Int. J. Mol. Sci. 2021, 22, 6143. [Google Scholar] [CrossRef]
- Davis, C.C.; Xi, Z.; Mathews, S. Plastid phylogenomics and green plant phylogeny: Almost full circle but not quite there. BMC Biol. 2014, 12, 11. [Google Scholar] [CrossRef]
- Henriquez, C.L.; Arias, T.; Pires, J.C.; Croat, T.B.; Schaal, B.A. Phylogenomics of the plant family Araceae. Mol. Phylogenet. Evol. 2014, 75, 91–102. [Google Scholar] [CrossRef]
- Li, H.T.; Luo, Y.; Gan, L.; Ma, P.F.; Gao, L.M.; Yang, J.B.; Cai, J.; Gitzendanner, M.A.; Fritsch, P.W.; Zhang, T.; et al. Plastid phylogenomic insights into relationships of all flowering plant families. BMC Biol. 2021, 19, 232. [Google Scholar] [CrossRef]
- Wu, F.H.; Chan, M.T.; Liao, D.C.; Hsu, C.T.; Lee, Y.W.; Daniell, H.; Duvall, M.R.; Lin, C.S. Complete chloroplast genome of Oncidium Gower Ramsey and evaluation of molecular markers for identification and breeding in Oncidiinae. BMC Plant Biol. 2010, 10, 68. [Google Scholar] [CrossRef]
- Li, C.; Zheng, Y.; Huang, P. Molecular markers from the chloroplast genome of rose provide a complementary tool for variety discrimination and profiling. Sci. Rep. 2020, 10, 12188. [Google Scholar] [CrossRef]
- Li, B.; Lin, F.; Huang, P.; Guo, W.; Zheng, Y. Development of nuclear SSR and chloroplast genome markers in diverse Liriodendron chinense germplasm based on low-coverage whole genome sequencing. Biol. Res. 2020, 53, 21. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.N.; Ruhlman, T.A.; Weng, M.L.; Khiyami, M.A.; Sabir, J.S.; Hajarah, N.H.; Alharbi, N.S.; Rabah, S.O.; Jansen, R.K. Plastome-wide nucleotide substitution rates reveal accelerated rates in Papilionoideae and correlations with genome features across Legume subfamilies. J. Mol. Evol. 2017, 84, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Ha, Y.-H.; Gil, H.-Y.; Choi, K.; Kim, D.-K.; Oh, S.-H. Two Korean endemic Clematis chloroplast genomes: Inversion, reposition, expansion of the inverted repeat region, phylogenetic analysis, and nucleotide substitution rates. Plants 2021, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Claude, S.-J.; Park, S.; Park, S. Gene loss, genome rearrangement, and accelerated substitution rates in plastid genome of Hypericum ascyron (Hypericaceae). BMC Plant Biol. 2022, 22, 135. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, P.-F.; Li, H.-T.; Yang, J.-B.; Wang, H.; Li, D.-Z. Plastid phylogenomic analyses resolve Tofieldiaceae as the root of the early diverging monocot order Alismatales. Genome Biol Evol. 2016, 8, 932–945. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Tian, J.-J.; Zhang, X.; Liu, Z.-D.; Shen, S.-K. Characterization of the complete chloroplast genome sequence of submerged macrophyte Stuckenia pectinata (Potamogetonaceae) and its phylogenetic position. Mitochondrial DNA Part B 2020, 5, 327–328. [Google Scholar] [CrossRef]
- Rogalski, M.; Ruf, S.; Bock, R. Tobacco plastid ribosomal protein S18 is essential for cell survival. Nucleic Acids Res. 2006, 34, 4537–4545. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S.; Lin, C.-P.; Hsu, C.-Y.; Wang, R.-J.; Chaw, S.-M. Comparative chloroplast genomes of Pinaceae: Insights into the mechanism of diversified genomic organizations. Genome Biol. Evol. 2011, 3, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Ruhlman, T.A.; Zhang, J.; Blazier, J.C.; Sabir, J.S.M.; Jansen, R.K. Recombination-dependent replication and gene conversion homogenize repeat sequences and diversify plastid genome structure. Am. J. Bot. 2017, 104, 559–572. [Google Scholar] [CrossRef]
- Fullerton, S.M.; Bernardo Carvalho, A.; Clark, A.G. Local rates of recombination are positively correlated with GC content in the human genome. Mol. Biol. Evol. 2001, 18, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wicke, S.; Wang, H.; Jin, J.J.; Chen, S.Y.; Zhang, S.D.; Li, D.Z.; Yi, T.S. Plastid genome evolution in the early-diverging Legume subfamily Cercidoideae (Fabaceae). Front. Plant Sci. 2018, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Cauz-Santos, L.A.; da Costa, Z.P.; Callot, C.; Cauet, S.; Zucchi, M.I.; Bergès, H.; van den Berg, C.; Vieira, M.L.C. A repertory of rearrangements and the loss of an inverted repeat region in Passiflora chloroplast genomes. Genome Biol. Evol. 2020, 12, 1841–1857. [Google Scholar] [CrossRef]
- Chen, N.; Sha, L.N.; Wang, Y.L.; Yin, L.J.; Zhang, Y.; Wang, Y.; Wu, D.D.; Kang, H.Y.; Zhang, H.Q.; Zhou, Y.H.; et al. Variation in plastome sizes accompanied by evolutionary history in monogenomic Triticeae (Poaceae: Triticeae). Front. Plant Sci. 2021, 12, 741063. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Son, O.; Park, S. The chloroplast genome of Elaeagnus macrophylla and trnH duplication event in Elaeagnaceae. PLoS ONE 2015, 10, e0138727. [Google Scholar] [CrossRef]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.G.; Barrett, C.F.; Soto Gomez, M.; Lam, V.K.; Henriquez, C.L.; Les, D.H.; Davis, J.I.; Cuenca, A.; Petersen, G.; Seberg, O.; et al. Plastid phylogenomics and molecular evolution of Alismatales. Cladistics 2016, 32, 160–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Lu, B.; Morales-Briones, D.F.; Moody, M.L.; Liu, F.; Hu, G.W.; Huang, C.H.; Chen, J.M.; Wang, Q.F. Phylogenomic analyses of Alismatales shed light into adaptations to aquatic environments. Mol. Biol. Evol. 2022, 39, msac079. [Google Scholar] [CrossRef]
- Haynes, R.R.; Les, D.H.; Král, M.; Kral, M. Two New Combinations in Stuckenia, the correct Name for Coleogeton (Potamogetonaceae). Novon 1998, 8, 241. [Google Scholar] [CrossRef]
- Ito, Y.; Robledo, G.L.; Iharlegui, L.; Tanaka, N. Phylogeny of Potamogeton (Potamogetonaceae) revisited: Implications for hybridization and Introgression in Argentina. Bull. Natl. Sci. Mus. 2016, 42, 131–141. [Google Scholar]
- Clegg, M.T.; Gaut, B.S.; Learn, G.H.; Morton, B.R. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, D.S.; Messer, P.W.; Hershberg, R.; Petrov, D.A. Strong purifying selection at synonymous sites in D. melanogaster. PLoS Genet. 2013, 9, e1003527. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Y.; Yang, J.-X.; Bai, M.-Z.; Zhang, G.-Q.; Liu, Z.-J. The chloroplast genome evolution of Venus slipper (Paphiopedilum): IR expansion, SSC contraction, and highly rearranged SSC regions. BMC Plant Biol. 2021, 21, 248. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gene map of Potamogeton plastid genomes. (a) Plastid genomes of P. crispus and P. maackianus. (b) Plastid genomes of P. wrightii, P. distinctus and P. perfoliatus. Genes drawn inside the circle are transcribed clockwise, and those outside are transcribed counterclockwise. The darker gray in the inner circle corresponds to GC contents.
Figure 1.
Gene map of Potamogeton plastid genomes. (a) Plastid genomes of P. crispus and P. maackianus. (b) Plastid genomes of P. wrightii, P. distinctus and P. perfoliatus. Genes drawn inside the circle are transcribed clockwise, and those outside are transcribed counterclockwise. The darker gray in the inner circle corresponds to GC contents.

Figure 2.
Comparison of junction between large single-copy (LSC), small single-copy (SSC), and IR regions among seven Potamogetonaceae plastomes.
Figure 2.
Comparison of junction between large single-copy (LSC), small single-copy (SSC), and IR regions among seven Potamogetonaceae plastomes.

Figure 3.
Analyses of repeat sequences in seven Potamogetonaceae plastomes. (A) Frequency of SSRs (simple sequence repeats). (B) Frequency of repeat sequences. C, complementary repeats; F, forward repeats; P, palindromic repeats; R, reverse repeats.
Figure 3.
Analyses of repeat sequences in seven Potamogetonaceae plastomes. (A) Frequency of SSRs (simple sequence repeats). (B) Frequency of repeat sequences. C, complementary repeats; F, forward repeats; P, palindromic repeats; R, reverse repeats.

Figure 4.
Comparison of the nucleotide variability (Pi) values (a) compared among Potamogetonaceae species and (b) compared among Potamogeton species.
Figure 4.
Comparison of the nucleotide variability (Pi) values (a) compared among Potamogetonaceae species and (b) compared among Potamogeton species.

Figure 5.
(a) The phylogenetic trees were constructed based on 65 coding genes of 21 Alismatales plastomes. (b) Types of junction between large single-copy (LSC), small single-copy (SSC), and IR regions in the Alismatales plastomes.
Figure 5.
(a) The phylogenetic trees were constructed based on 65 coding genes of 21 Alismatales plastomes. (b) Types of junction between large single-copy (LSC), small single-copy (SSC), and IR regions in the Alismatales plastomes.

Figure 6.
Phylogenetic tree constructed using the maximum likelihood (ML) and Baysian inference (BI) methods based on 65 plastid protein-coding genes. The number above the lines indicates bootstrap values/BI posterior probabilities.
Figure 6.
Phylogenetic tree constructed using the maximum likelihood (ML) and Baysian inference (BI) methods based on 65 plastid protein-coding genes. The number above the lines indicates bootstrap values/BI posterior probabilities.

Figure 7.
Nonsynonymous (dN) and synonymous (dS) substitution rates and dN/dS values of 48 plastid protein-coding genes across Alismatales. PP1, Potamogeton perfoliatus1; PP2, Potamogeton perfoliatus 2; PD, Potamogeton distinctus; PW, Potamogeton wrightii; PC, Potamogeton crispus; PM, Potamogeton maackianus; SP, Stuckenia pectinate; SI, Syringodium isoetifolium; RB, Ruppia brevipdeunculata; AM, Aponogeton madagascariensis; AD, Aponogeton desertorum; OE, Ottelia emersa; OC, Ottelia cordata; OA, Ottelia alismoides; OG, Ottelia guanyangensis; BA, Blyxa aubertii; EC, Eldea canadensis; NF, Najas flexilis; SL, Sagittaria lichuanensis; TT, Tofieldia thibetica.
Figure 7.
Nonsynonymous (dN) and synonymous (dS) substitution rates and dN/dS values of 48 plastid protein-coding genes across Alismatales. PP1, Potamogeton perfoliatus1; PP2, Potamogeton perfoliatus 2; PD, Potamogeton distinctus; PW, Potamogeton wrightii; PC, Potamogeton crispus; PM, Potamogeton maackianus; SP, Stuckenia pectinate; SI, Syringodium isoetifolium; RB, Ruppia brevipdeunculata; AM, Aponogeton madagascariensis; AD, Aponogeton desertorum; OE, Ottelia emersa; OC, Ottelia cordata; OA, Ottelia alismoides; OG, Ottelia guanyangensis; BA, Blyxa aubertii; EC, Eldea canadensis; NF, Najas flexilis; SL, Sagittaria lichuanensis; TT, Tofieldia thibetica.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of the plastid genome features of Potagometon species.
| Order | Family | Species | Length (bp) | GC Contents | NCBI Accession Number | |||
|---|---|---|---|---|---|---|---|---|
| Total | LSC | SSC | IR | |||||
| Acorales | Acoraceae | Acorus gramineus | 152,849 | 82,977 | 18,228 | 25,822 | 38.7% | NC_026299 |
| Alismatales | Tofieldiaceae | Tofieldia thibetica | 155,512 | 84,584 | 18,151 | 26,388 | 37.4% | NC_029813 |
| Alismataceae | Sagittaria lichuanensis | 179,007 | 99,125 | 13,278 | 33,302 | 36.8% | NC_029815 | |
| Hydrocharitaceae | Najas flexilis | 156,366 | 88,697 | 15,266 | 31,201 | 38.2% | NC_021936 | |
| Eldea canadensis | 156,700 | 86,194 | 17,808 | 26,349 | 37.0% | NC_018541 | ||
| Blyxa aubertii | 158,187 | 87,799 | 18,804 | 25,792 | 36.5% | MK940507 | ||
| Ottelia guanyangensis | 157,362 | 87,230 | 19,004 | 25,564 | 36.7% | MK940522 | ||
| Ottelia alismoides | 157,880 | 87,699 | 19,067 | 25,557 | 36.6% | MK940517 | ||
| Ottelia cordata | 157,896 | 87,665 | 19,121 | 25,555 | 36.6% | MK940519 | ||
| Ottelia emersa | 157,896 | 87,665 | 19,121 | 25,555 | 36.6% | MK940520 | ||
| Aponogetonaceae | Aponogeton desertorum | 154,516 | 85,760 | 19,890 | 24,433 | 36.9% | MK570533 | |
| Aponogeton madagascariensis | 155,669 | 86,896 | 19,869 | 24,452 | 36.9% | MK570534 | ||
| Cymodoceaceae | Ruppia brevipedunculata | 158,943 | 88,857 | 19,130 | 25,478 | 35.8% | NC_051974 | |
| Syringodium isoetifolium | 159,333 | 89,055 | 19,160 | 25,559 | 35.9% | MZ325253 | ||
| Potamogetoncaceae | Stuckenia pectinata | 156,669 | 86,285 | 18,237 | 26,073 | 36.5% | MN661144 | |
| Potamogeton perfoliatus1 | 156,226 | 86,764 | 18,238 | 25,612 | 36.5% | NC_029814 | ||
| Potamogeton perfoliatus2 | 156,276 | 86,821 | 18,231 | 25,612 | 36.5% | This study (OQ561452) | ||
| Potamogeton maackianus | 156,488 | 86,833 | 18,221 | 25,717 | 36.5% | This study (OQ561451) | ||
| Potamogeton crispus | 155,863 | 86,191 | 18,182 | 25,745 | 36.5% | This study (OQ561449) | ||
| Potamogeton wrightii | 156,315 | 86,827 | 18,282 | 25,603 | 36.5% | This study (OQ561453) | ||
| Potamogeton distinctus | 156,354 | 86,898 | 18,286 | 25,585 | 36.5% | This study (OQ561450) | ||
LSC, large single-copy region; SSC, small single-copy region; IR, inverted-repeat region.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Choi, K.; Hwang, Y.; Hong, J.-K.; Kang, J.-S. Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species. Genes 2023, 14, 1914. https://doi.org/10.3390/genes14101914
AMA Style
Choi K, Hwang Y, Hong J-K, Kang J-S. Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species. Genes. 2023; 14(10):1914. https://doi.org/10.3390/genes14101914
Chicago/Turabian StyleChoi, KyoungSu, Yong Hwang, Jeong-Ki Hong, and Jong-Soo Kang. 2023. "Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species" Genes 14, no. 10: 1914. https://doi.org/10.3390/genes14101914
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.