
Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries


1
School of Ecology and Nature Conservation, Beijing Forestry University, P.O. Box 61, Beijing 100083, China
2
Forest Pest Control and Quarantine Station of Hebei Province, Shijiazhuang 050081, China
3
Faculty of Agriculture, University of Mauritius, Reduit 80837, Mauritius
4
Charles Regnaud Street, Curepipe 74319, Mauritius
5
Department of Health Sciences, Faculty of Medicine and Health Sciences, University of Mauritius, Reduit 80837, Mauritius
*
Authors to whom correspondence should be addressed.
Life 2021, 11(7), 657; https://doi.org/10.3390/life11070657
Submission received: 16 April 2021
/
Revised: 15 June 2021
/
Accepted: 24 June 2021
/
Published: 5 July 2021
(This article belongs to the Special Issue Biodiversity and Ecology of Fungi in Terrestrial and Marine Ecosystems)
Abstract
:Botryosphaeriaceous fungi cause stem blight, canker and dieback in woody plants. During a survey on the fungal pathogens associated with blueberries in China, 135 blighted, cankered or dead blueberry branches were collected from Fujian and Shandong Provinces. Based on the morphological characterization and phylogenetic analyses of a concatenated ITS rDNA, tef1-α, TUB, and RPB2 loci, five new species of Lasiodiplodia, viz., L. clavispora, L. fujianensis, L. henanica, L. nanpingensis and L. paraphysoides were recognized. Detailed descriptions and illustrations, as well as multigene phylogenies, are provided in this paper. The diversity of plant pathogens on agriculturally and economically important plants is higher than anticipated.
1. Introduction
Blueberries (Vaccinium spp.) are perennial shrub fruit trees. The fruits are well known and are widely consumed for their protective properties against heart diseases and cancer, they can help to maintain bone strength and mental health and can regulate blood pressure [1]. Blueberries are widely distributed in temperate regions, such as North America, Europe, Canada and Northern China [2,3,4,5,6,7,8,9,10,11,12,13]. Due to their health benefits and economic value, blueberries have been commercially cultivated worldwide, particularly in the USA, Canada and European countries [14,15]. Blueberry cultivation started in 1981 in China, and productivity has reached 43,244 tons per year [16].
Botryosphaeriaceous fungi are a group of economically important plant pathogens [17,18,19,20]. They cause stem blight, canker or dieback in a wide range of hosts, including blueberries [8,9,11,15,21,22,23,24,25]. In the USA, blueberry stem blight, caused by Botryosphaeria ribis, has been a major disease in commercial plantations in North Carolina [26,27]. Pathogenicity studies conducted show that stem dieback is caused by B. dothidea and canker by Lasiodiplodia corticis in blueberries in North Carolina. Neofusicoccum parvum was identified as the causal agent for blueberry stem blight and dieback in California and Mexico [2,4]. In Florida, the blueberry stem blight and dieback caused by Neofusicoccum ribis and Lasiodiplodia theobromae led to huge economic losses and were one of the most severe diseases in the local blueberry planting industry [8,9,28]. The incidence of blueberry stem blight and canker caused by Neofusicoccum parvum has been a limiting factor for blueberry production in Chile [29]. The incidence of blueberry blight and crown rot caused by N. ribis and L. theobromae was so severe in New Zealand that it resulted in an annual loss of about USD 500,000 due to yield losses and replanting [6,8,9,28]. Neofusicoccum parvum and N. austral caused blueberry stem dieback and canker in Spain [3,30]. Many more botryosphaeriaceous species have been reported to cause blueberry stem dieback or canker worldwide, such as Botryosphaeria corticis, Lasiodiplodia mediterranea, L. pseudotheobromae, Macrophomina phaseolina, Neofusicoccum arbuti, N. austral, N. kwambonambiense, N. macroclavatum, N. occulatum and N. ribis [7,8,9,15,29,31,32,33,34].
Blueberry cultivation started in 1981 in China and, subsequently, the blueberry stem diseases caused by botryosphaeriaceous fungi received more and more attention. For instance, some studies first reported that blueberry bud and stem blight or dieback were caused by Neofusicoccum vitifusiforme in Yunnan Province in China [5,12]. In addition, N. parvum caused blueberry stem blight in highbush blueberries (Vaccinium corymbosum) in Yunnan Province [12]. In Shandong Province, it was reported that blueberry stem blight and dieback were caused by Lasiodiplodia pseudotheobromae (current name L. chinensis) [22,35]. It was noticed that botryosphaeriaceous fungi cause blueberry stem blight or dieback in eight provinces in China, and three species were recognized: Botryosphaeria dothidea, Lasiodiplodia theobromae and Neofusicoccum parvum [11]. Furthermore, it was also noticed that L. theobromae and N. parvum are more virulent than Botryosphaeria dothidea [11]. A new fungus has been described, viz. L. vaccini, which causes blueberry stem blight in the greenhouses of blueberry plantations in rural areas of Beijing [13]. The pathogenicity of Botryosphaeriaceae was discussed by Manawasinghe et al. [36].
During a survey on the fungal pathogens associated with blueberries in China, several species of Lasiodiplodia have been identified, and five of them are described as new to science. A concatenated DNA dataset from ITS rDNA and tef1-α, TUB, and RPB2 loci have been analyzed, and the phylogenetic relationships of these novel species have been established.
2. Methods and Materials
2.1. Sample Collections and Fungal Isolation
One hundred and thirty-five blighted, cankered or dead blueberry branches were collected from Fujian (69 samples) and Shandong (66 samples) Provinces in China from April to November 2018. Diseased or dead twigs of blueberries (ca. 30 cm) were cut for sampling, from which the fungal strains were isolated. Wood segments (0.5 × 0.5 × 0.2 cm) were cut from the diseased lesion boundary or dead tissues and were subsequently surface sterilized and incubated in malt extract agar (MEA) at 28 °C for fungal strain isolation [13,37,38,39]. The isolates were kept at ambient temperatures (about 26–28 °C) and grown in the dark.
2.2. Morphological Characterization
Fungal colonies were initially identified based on morphological characteristics. Fungal isolates were transferred to synthetic nutrient-poor agar (SNA) with sterilized pine needles for 3 weeks in order to induce sporulation. The pycnidia produced on the pine needles were morphologically described following the work by Dou et al. [35,40]. Microscopic observations were made from material mounted in water. Measurements of paraphyses, conidiogenous cells and conidia were made in water. For each new species, the measurements of 20 paraphyses, 20 conidiogenous cells and 50 conidia were taken under a Nikon Eclipse E600 microscope. Fungal isolates and herbarium specimens were deposited at the China General Microbiological Culture Collection Center (CGMCC) and the Mycological Herbarium of the Institute of Microbiology, Chinese Academy of Sciences (HMAS). The new species were established based on the guidelines outlined by Jeewon and Hyde [41].
2.3. DNA Extraction, PCR Amplification
DNA was extracted with the CTAB plant genome DNA fast extraction kit (Aidlab Biotechnologies Co, Ltd., Beijing, China) from the mycelium grown on MEA. PCR amplifications were performed using the Easy Taq PCR Super Mix kit (Beijing Transgen Biotech Co., Ltd., Beijing, China). The internal transcribed spacers of rDNA (ITS) were amplified and sequenced with the primers ITS-1 and ITS-4 [42]. The translation elongation factor-1α (tef1-α) was amplified and sequenced with primers EF1-688F and EF1-1251R [43]. The TUB gene was amplified and sequenced with primers Bt2a and Bt2b [44]. The RPB2 were amplified and sequenced using primers RPB2-LasF and RPB2-LasR [45]. PCR amplification and sequencing followed the protocol outlined by Zhang et al. [46]. PCR amplifications were performed using the Easy Taq PCR Super Mix kit (Beijing Transgen Biotech Co., Ltd., Beijing, China). For the ITS regions, the following PCR profile was used: 95 °C for 3 min, followed by 34 cycles of denaturation at 95 °C for 1 min, annealing at 52°C for 30 s and elongation at 72 °C for 1 min, with a final extension step of 72 °C for 10 min. The PCR profiles for the tef1-α, TUB and RPB2 genes were same, except that 35 cycles of denaturation were used and the annealing temperature was 55 °C.
2.4. Sequence Alignment and Phylogenetic Analysis
The concatenated loci of ITS, tef1-α, TUB and RPB2 were used to infer the phylogenetic relationships of taxa within Lasiodiplodia. Alignments were conducted in MEGA v. 6, and phylogenetic analyses performed in PAUP v. 4.0b10 and MrBayes v. 3.1.2 [47,48,49]. Prior to phylogenetic analysis, ambiguous sequences at the start and end were deleted and gaps manually adjusted in order to optimize the alignments. Maximum Parsimony (MP) was used to conduct heuristic searches, as implemented in PAUP with the default options method [50]. Analyses were conducted under different parameters of maximum parsimony criteria [50]. Clade stability was assessed in a bootstrap analysis with 1000 replicates, random sequence additions with maxtrees set to 1000 and other default parameters, as implemented in PAUP. For the MrBayes analysis, the best-fit model of nucleotide evolution (GTR+I+G) was selected by the Akaike information criterion [51] in MrModeltest v. 2.3. The metropolis-coupled Markov Chain Monte Carlo (MCMCMC) approach was used to calculate posterior probabilities [47]. Bayesian inference (BI) analysis with MrBayes revealed that the Markov chain Monte Carlo (MCMC) steady state was reached after fewer than 19,820,000 generations (the average standard deviation of split frequencies was constantly below 0.01). A conservative burn-in of 198,200 trees was chosen, and a full analysis of 20,000,000 generations was carried out with sampling every 100 generations. Trees were viewed in TREEVIEW [52]. The nucleotide sequences generated in this paper were deposited in GenBank (Table 1). Trees and alignments were deposited in TreeBase (http://purl.org/phylo/treebase/phylows/study/TB2:S24322?x-access-code=1443788eea51ad240fcd94b3927ffb1a&format=html, accessed on 15 June 2021).
3. Results
For Lasiodiplodia, the concatenated ITS, tef1-α, TUB, and RPB2 DNA sequence dataset comprises 1974 bp with 335 parsimony-informative characters. A MP tree (TL = 890 steps, CI = 0.609, RI = 0.870, RC = 0.530, HI = 0.391) generated based on a heuristic search with the random addition of taxa (1000 replicates) is shown in Figure 1.
Taxonomy
Lasiodiplodia clavispora Y. Zhang ter., Y. Wang, sp. nov. (Figure 2).
MycoBank: MB 830994.
The etymology of the name reflects the clavate conidia.
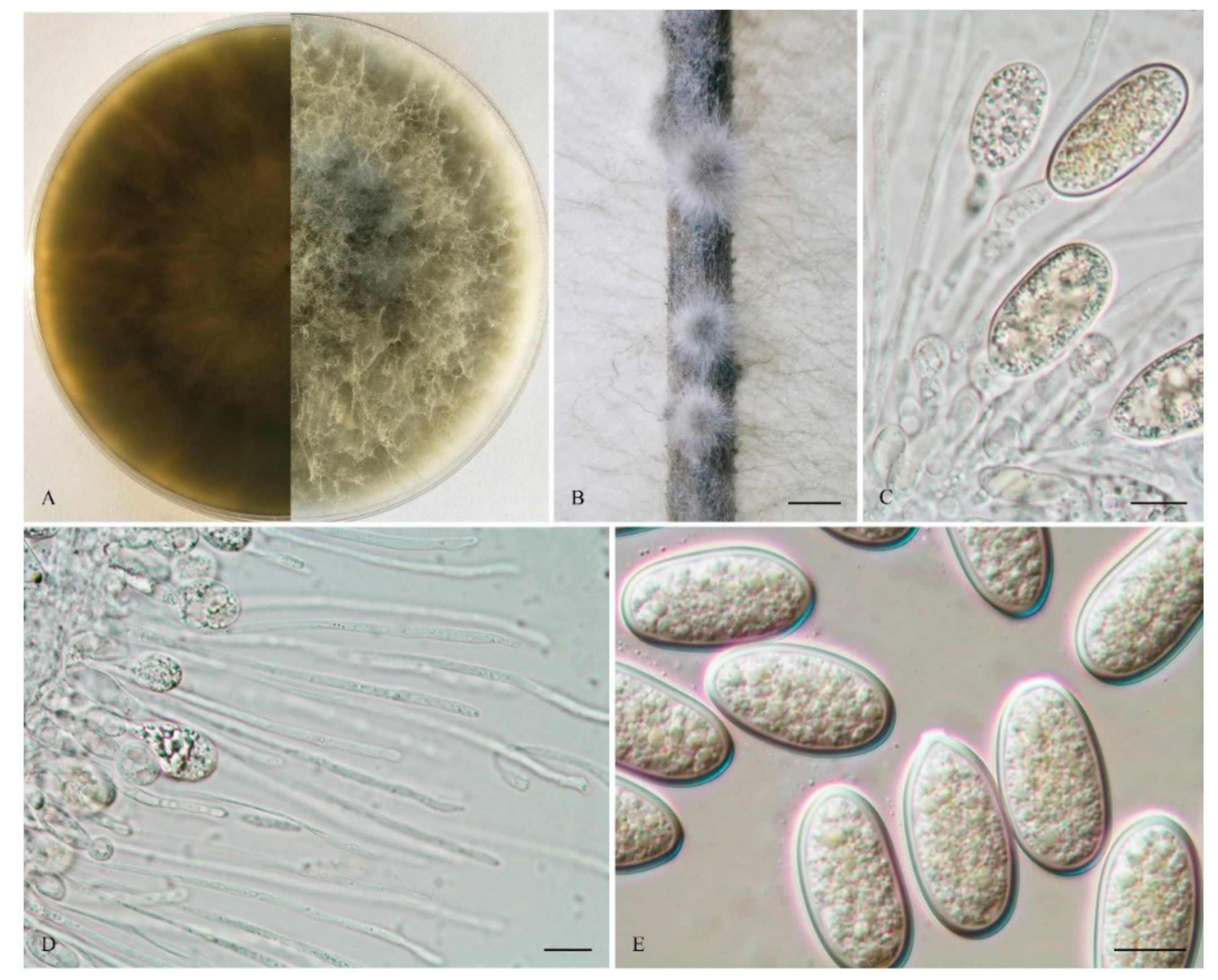
The sexual stage was not observed. Conidiomata were stromatic, produced on both sterilized pine needles and SNA within 10 days, it was semi-immersed, uniloculate and rarely multiloculate, black, covered by greyish brown mycelium, and up to 570 μm diam when there was uniloculate. Paraphyses were filiform, arising from the conidiogenous layer, extending above the level of developing conidia, up to 100 μm long and 3 μm wide, cylindrical, thin walled, aseptate, hyaline, tip rounded, and unbranched. Conidiophores were reduced to conidiogenous cells. Conidiogenous cells were holoblastic, hyaline, discrete, smooth, and thin-walled, (9.5–) 11–18 (–19) × 2.5–5 μm (mean of 50 conidiogenous cells = 14.3 × 3.8 μm, L/W ratio = 4). Conidia were hyaline, with a wall of 1–2 μm thick, clavate, narrowly ellipsoid to narrowly ovoid with a round apex and had a slightly tapered base, (28–) 29–36 (–38) × 12–15 μm (mean of 50 conidia = 31.7 × 13.8 μm, L/W ratio = 2.3, range from 2.0 to 3.0), no pigmented conidia observed after 15 days. Spermatia were not observed.
Culture characteristics: Colonies on MEA were initially white with moderately dense aerial mycelia reaching the lid of the plate and became olive grey on the surface after 5 d, with the reverse side of the colonies being pale grey to grey. Colonies reached 18 mm on MEA after 24 h in the dark at 28 °C, and were more than 55 mm after 48 h.
Materials examined: CHINA, Fujian province, Nanping, Jianyang district, from blighted stems of Vaccinium uliginosum Linn., 1 April 2018, L. Zhao (Holotype: HMAS 255607, ex-type isolate: CGMCC 3.19594; Paratype: HMAS 255612, isolate: CGMCC 3.19595).
Notes: Phylogenetically, L. clavispora is closely related to L. gonubiensis (PP/MP = 1.00/100, Figure 1). Lasiodiplodia clavispora (CGMCC 3.19594) differs from its closest phylogenetic neighbor L. gonubiensis (CMW14077) (Figure 1) by 14 bp in tef1-α (0.72 %) (Table 2). In addition, a conidial size of L. clavispora also differs from L. gonubiensis (12–15 vs. (14–) 16–18.5 (–21) μm) [37].
Lasiodiplodia fujianensis Y. Zhang ter., Y. Wang, sp. nov (Figure 3).
MycoBank: MB 830996.
The etymology is in reference to the location, Fujian province, where the species was first reported.
The sexual stage was not observed. Conidiomata were stromatic, produced on both sterilized pine needles and SNA within 10 days, semi-immersed, uniloculate, black, covered by greyish brown mycelium, and were up to 1.3 mm in diameter. Paraphyses were filiform, arising from the conidiogenous layer that extended above the level of developing conidia and were up to 95 μm long and 3 μm wide, aseptate, hyaline, tip rounded, and unbranched. Conidiophores reduced to conidiogenous cells. Conidiogenous cells were holoblastic, hyaline, discrete, smooth, and thin walled, (11–) 12–18.5 (–20) × (3–) 4–8 (–8.5) μm (mean of 50 conidiogenous cells = 14.9 × 5.4 μm, L/W ratio = 2.9). Conidia were hyaline, with a 1–2 μm thick wall, ellipsoid with a round apex and round base, and occasionally truncated at the base, (22–) 23–29 (–30) × (12–) 13–15 (–16) μm (mean of 50 conidia = 26.2 × 14.5 μm, L/W ratio = 1.8, range from 1.5 to 2.2), with pigmented conidia observed after 15 days. Spermatia were not observed.
Culture characteristics: Colonies on MEA were initially white with moderately dense aerial mycelia reaching the lid of the plate and became ash-grey on the surface after 5 d, with the reverse side of the colonies being pale grey to grey. Colonies reached 45 mm on MEA after 24 h in the dark at 28 °C, and more than 90 mm after 48 h.
Materials examined: China, Fujian Province, Nanping, Jianyang district, from blighted stems of Vaccinium uliginosum, 1 April 2018, L. Zhao (Holotype: HMAS 255606, ex-type isolate: CGMCC 3.19593).
Notes: Phylogenetically, L. fujianensis is basal to L. thailandica and L. iraniensis (Figure 1). Lasiodiplodia fujianensis (CGMCC 3.19593), however, differs from L. thailandica (CPC 22755) (Figure 1) by 16 bp in tef1-α (1.09 %, gaps included) (Table 2). Morphologically, L. fujianensis also differs from L. thailandica in the size of the conidiomata and conidiogenous cells (310–330 × 300–370µm and 8–9 × 2–4 µm, respectively [53]. In addition, the aseptate paraphyses of L. fujianensis also make it morphologically different from L. thailandica (1–3-septate).
Mycobank: MB 817650.
The etymology is in reference to the location, Henan province, where the species were reported.
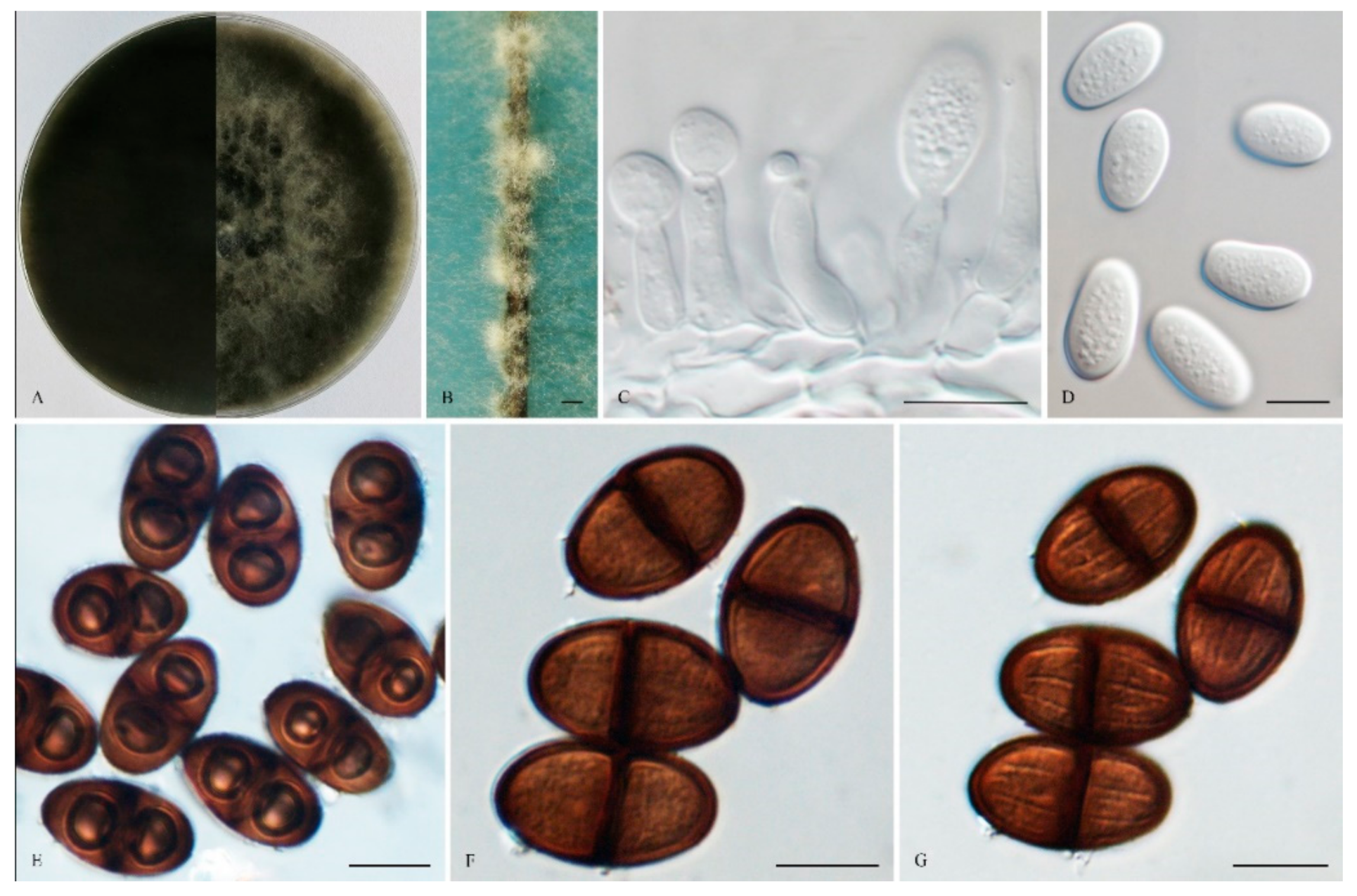
The sexual stage was not observed. Conidiomata were stromatic, produced on both sterilized pine needles on SNA within 14 days, and were semi-immersed or superficial, mostly solitary, globose, smooth, mostly non-papillate, iron grey to black, covered by brown mycelium, and up to 520 μm in diameter. Paraphyses were filiform and arose from the conidiogenous layer, extending above the level of developing conidia, and were up to 105 μm long and 4 μm wide, cylindrical, thin-walled, initially aseptate, which became up to 1–3-septate when mature, hyaline, apex rounded, occasionally basal cells swollen, and unbranched. Conidiophores were reduced to Conidiogenous cells. Conidiogenous cells were holoblastic, hyaline, discrete, smooth, thin-walled, and were cylindrical to ampulliform, (8–) 9–16 × 3 –5 (–7) μm (mean of 50 conidiogenous cells = 12.1 × 4.0 μm, L/W ratio = 2.95). Conidia were initially hyaline, with a 1 μm thick wall, ovoid to cylindrical, turning brown with a median septum and longitudinal striations when mature, and sometimes with two vacuoles, (14–) 19–26 (–27) × 10–13 (–15) μm (mean of 100 conidia = 22.1 × 12.0 μm, L/W ratio = 1.86, by range from 1.17 to 2.6). Spermatia were not observed.
Culture characteristics: Colonies on MEA were initially white with moderately dense aerial mycelia reaching the lid of the plate and became dark grey to black on the surface after 7 d, with the reverse side of the being colonies dark black. Colonies reached 26 mm on MEA after 24 h in the dark at 28 °C, and more than 65 mm after 48 h.
Specimens examined: China, Shandong province, Qingdao, Huangdao district, were from blighted stems of Vaccinium uliginosum, 17 November 2018, Y. Zhang and L. Zhao (Holotype: HMAS 247961, ex-type isolate: CGMCC 3. 19176). Henan province, Puyang city, Qingfeng, a farmer orchard was from cankered stems of Morus alba Linn. var. alba, 11 November 2014, Z. P. Dou & W. He (Paratype: HMAS 255410, isolate: CGMCC 3.17969).
Notes: Phylogenetically, L. henanica is basal to the clade and comprised of L. citricola, L. paraphysoides, L. aquilariae, L. euphorbicola, L. parva, L. hormozganensis and L. laeliocattleyae. Morphologically, L. henanica differs from L. hormozganensis in having smaller-sized conidiomata (up to 520 μm vs. up to 950 μm) [54]. In addition, the presence of vacuoles in the conidia of L. henanica also makes it different from L. hormozganensis and L. laeliocattleyae [54,55]. The broader 1–3-septate paraphyses of L. henanica are also distinguishable from L. laeliocattleyae (up to 3 μm, aseptate) [55].
Lasiodiplodia nanpingensis Y. Zhang ter., Y. Wang, sp. nov. (Figure 5).
MycoBank: MB 830997.
The etymology of the name reflects Nanping, where this species was first reported.
The sexual stage was not observed. Conidiomata were stromatic, it was produced on both sterilized pine needles and SNA within 7 days, and was solitary, scattered or in small groups (up to 5), semi-immersed or superficial, uniloculate, black, covered by greyish brown mycelium, and up to 640 μm diam. Paraphyses were filiform, arising from the conidiogenous layer, extending above the level of developing conidia, up to 102 μm long and 3.5 μm wide, and was aseptate, hyaline, tip rounded, and branched. Conidiophores was reduced to conidiogenous cells. Conidiogenous cells were holoblastic, hyaline, discrete, smooth, and thin walled, 9–16 (–19) × 3–6 (–7) μm (mean of 50 conidiogenous cells = 13.0 × 4.6 μm, L/W ratio = 2.97). Conidia were hyaline, with a 1 μm thick wall, ellipsoid with round apexes and was rarely irregular, (20–) 21–26 (–28) × 13–16 (–17) μm (mean of 50 conidia = 23.9 × 14.8 μm, L/W ratio = 1.6, range from 1.4 to 1.9). Spermatia were not observed.
Culture characteristics: Colonies on MEA were initially white with moderately dense aerial mycelia reaching the lid of the plate and becoming ash-grey on the surface after 5 d, with the reverse side of the colonies being pale grey to grey. Colonies reached 17 mm on MEA after 24 h in the dark at 28 °C, and more than 60 mm after 48 h.
Materials examined: China, Fujian province, Nanping, Jianyang district, from blighted stems of Vaccinium uliginosum, 1 April 2018, L. Zhao (Holotype: HMAS 255608, ex-type isolate: CGMCC 3.19596; Paratype: HMAS 255609, isolate: CGMCC 3.19597).
Notes: Phylogenetically, the clade comprising L. curvata, L. exigua, L. mahajangana, L. nanpingensis and L. irregularis received moderate bootstrap support (PP/MP = 0.95/59) (Figure 1). It can also be noted that our two strains of L. nanpingensis constituted a strongly supported independent subclade. Morphologically, the deeply curved conidia of L. curvata distinguished it from L. nanpingensis [56]. The larger-sized conidiomata of L. nanpingensis also differed from L. irregularis (up to 640 μm vs. up to 400 μm). In addition, the branched and aseptate paraphyses of L. nanpingensis made the latter distinct from the unbranched and 1-septate paraphyses of L. irregularis [56], as well as from L. mahajangana [57]. The larger-sized conidiomata and conidia of L. nanpingensis also differed from L. mahajangana [57]. Lasiodiplodia nanpingensis became longer and had slender paraphyses, which were different from those of L. exigua (up to 102 × 3.5 μm vs. up to 66 × 5 μm) [58].
Lasiodiplodia paraphysoides Z. P. Dou, Y. Wang, Y. Zhang ter sp. nov. (Figure 6).
Mycobank: MB 817655.
The etymology is in reference to the long and multiseptate paraphyses.
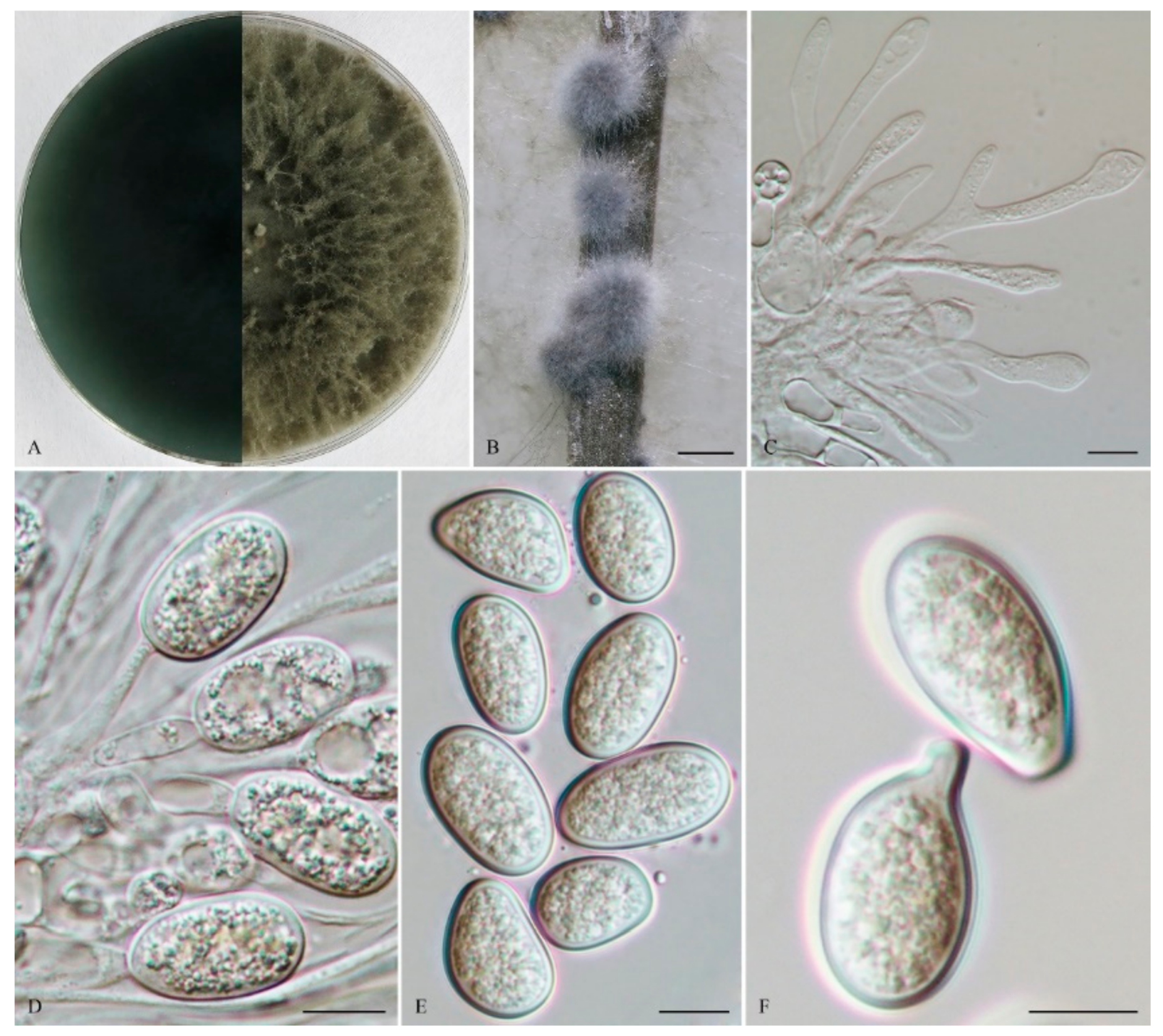
The sexual stage was not observed. Conidiomata were stromati, produced on both sterilized pine needles on SNA within 14 days, and were solitary, globose, semi-immersed or superficial, uniloculate, dark brown to black, covered with brown mycelium, up to 1.8 mm diam, and often had a long papilla, which was up to 383 μm long and 113 μm wide. Paraphyses were filiform, arising from the conidiogenous layer, extending above the level of developing conidia, up to 125 μm long and 7 μm wide, and were cylindrical, thin-walled, hyaline, tip rounded, initially aseptate, becoming up to 1–2-septate when mature, branched, occasionally basal, and were middle or apical swollen cells. Conidiophores were reduced to conidiogenous cells. Conidiogenous cells were holoblastic, hyaline, discrete, smooth, thin-walled, and were cylindrical to ampulliform, (8–) 10–16 (–18) × 3–7 μm (mean of 50 conidiogenous cells = 13.0 × 4.7 μm, L/W ratio =2.92). Conidia were initially hyaline, aseptate, with a 1 μm thick wall, and ellipsoid to ovoid with a round apex and round base, straight to obvious curved, turning brown with a median septum and longitudinal striations when mature, 1-septate, verruculose, (20–) 21–25 (–30) × (10–) 12–15 (–17) μm (mean of 50 conidia = 23.0 × 13.7 μm, L/W ratio = 1.69, range from 1.38 to 2.31), conidia sometimes germinating before septum formed or after pigmented. Spermatia were not observed.
Culture characteristics: Colonies on MEA were initially white with moderately dense aerial mycelia reaching the lid of the plate, and became dark grey on the surface after 7 d, with the reverse sides of the colonies dark grey to dark bluish grey. Colonies reached 20.5 mm on MEA after 24 h in the dark at 28 °C.
Specimens examined: China, Shandong province, Qingdao, Huangdao district, from blighted stems of Vaccinium uliginosum, 17 November 2018, Y. Zhang and L. Zhao (Holotype: HMAS 247959, ex-type isolate: CGMCC 3. 19174; Paratype: HMAS 247960, isolate: CGMCC 3. 19175).
Notes: Phylogenetically, L. paraphysoides was closely related to L. citricola and an unidentified taxon, Lasiodiplodia sp. Lasiodiplodia paraphysoides (CGMCC 3. 19174) and differred from its closest phylogenetic neighbor L. citricola (IRAN1522C) (Figure 1) by unique fixed alleles in two loci based on alignments of the separate loci deposited in TreeBASE (S25538), by 4 bp in tef1-α (0.72 %, gaps included) (Table 2). Morphologically, the long papilla of the conidiomata of L. paraphysoides delineated itself from the non-papillate conidiomata of L. citricola [54]. Furthermore, the conidiogenous cells of L. citricola had 1–2 annellations, which also differed from the holoblastic conidiogenous cells of L. paraphysoides [54].
4. Discussion
In this study, we recovered five new species of Lasiodiplodia associated with stem blight and/or canker of blueberries, namely, L. clavispora, L. fujianensis, L. henanica, L. nanpingensis and L. paraphysoides, and they were characterized in terms of their morphology and their phylogenetic relationships to other species of Lasiodiplodia. Phylogenetically, each of these five newly described species formed a well-supported subclade close to other species (Figure 1). Species of Lasiodiplodia were mostly differentiated based on the morphology of the conidia (especially dimensions) and paraphyses [17,35]. In this study, we attempted to use other morphological characters, such as the dimensions and papillate nature of conidiomata, as well as annelations of conidiogenous cells, but to what extent these are phylogenetically significant warrants further investigation.
Geographically, Lasiodiplodia tends to be distributed in tropical or subtropical areas or in warm temperate areas associated with various stem diseases of woody substrates [8,9,11,22,33,35]. For instance, L. mediterranea and L. pseudotheobromae have been reported as canker-causing agents of grapevine and other woody hosts in Italy, Algeria and Tunisia [58]. The stem blight and crown rot of blueberry caused by L. theobromae have been reported in Florida in the USA, as well as in Zhejiang Province and Shanghai in China [8,9,10,11]. The cane dieback of blueberry caused by L. mediterranea has been reported in Washington in the USA [33]. In China, blueberry stem blight and dieback caused by L. chinensis have been reported in Shandong Province [40,56]. The stem blight of blueberry caused by L. vaccinii was reported in a greenhouse plantation in Beijing, where it was warm with high humidity [13]. All the five species of Lasiodiplodia newly described in this study were from Fujian and Shandong Province, which belong to subtropical or warm temperate areas in China. The distribution of Lasiodiplodia spp seems largely influenced by environmental conditions, such as temperature, humidity, elevation, as well as the prevalence of alternative hosts instead of their host associations [28,59].
We also compared our species with newly described species recently published by de Silva et al. [60]. From a phylogenetic perspective, our new species are quite different, except for one, L. fujianensisis. The latter is basal to L. thailandica and L. iraniensis, which are known species. de Silva et al. [60] also reported that their new species, L. endophytica from Magnolia plant, are phylogenetically closely related to L. thailandica and L. iraniensis albeit in a distinct independent lineage with weak support. To avoid any ambiguous taxonomic interpretation in connection with the identification of L. fujianensisis, we compared DNA base pair differences with L. endophytica. DNA sequences from the TEF protein coding region for L. endophytica is quite short (271 bp) and we still found two major differences, which supports that our species is different. With respect to the DNA sequences from the Beta tubulin gene region, L. fujianensisis was 100% similar to L. endophytica. Could this be pointing to the fact that these two taxa are conspecific? This might be true, but we compared existing DNA sequences of the Beta tubulin from other published species, such as L. pseudotheobromae, L. jatrophicola, L. vitis and L. iraniensis and found that they are identical to L. fujianensisis and L. endophytica. The taxonomy of Lasioplodia has been rather controversial [35]. While some are proponents of a taxonomy based on morphological characteristics, others argue that more protein genes should be included in the taxonomy, especially at the species level. However, the protein genes might not be useful, at least in some fungal groups, because they have reached saturation and are possibly less informative than has been anticipated. Mycologists also encounter difficulties when analyzing DNA sequence data for many bitunicate fungi. In this case, even with L. endophytica, de Silva et al. [60] demonstrated that single gene phylogenies could reveal extensive incongruence (Figure 1, Figure 2 and Figure 3), which can be found in the supplementary information provided by de Silva et al. [60]. We could not compare the morphs of L. fujianensisis to L. endophytica as the latter was isolated as an endophyte and did not produce any fruiting bodies in culture. There is also a need to update the name of the GB accession numbers of MK501838, MK584572, and MK550606 as these are labelled as “Lasiodiplodia sp. NIS-2019a isolate”, but we presume that it should be Lasiodiplodia endophytica.
Author Contributions
Conceptualization, Y.Z. and R.J.; methodology, Y.W., Y.Z. and R.J.; software, Y.W.; validation, Y.W., Y.Z. and R.J.; formal analysis, Y.W., Y.Z. and R.J.; investigation, Y.W., Y.Z. and R.J.; resources, Y.W., Y.Z. and R.J.; data curation, Y.W., Y.Z. and R.J.; writing—original draft preparation, Y.W., Y.Z., V.B., S.R. and R.J.; writing—review and editing, Y.W., Y.Z., V.B., S.R. and R.J.; visualization, Y.W., Y.Z., V.B., S.R. and R.J.; supervision, Y.Z. and R.J.; project administration, Y.Z. and R.J.; funding acquisition, Y.Z. and R.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (General Program, 31971658, 31770015, 31370063), NSFC Projects of International Cooperation and Exchanges (3155461143028). Rajesh Jeewon also acknowledges the contribution of the Mauritius Research Innovation Council (grant number MRC/RUN1705).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
DNA sequence data generated in this study have been submitted to Genbank. Dataset used to generate phylogenies has been submitted to TREEBASE with accession number: 24322.
Acknowledgments
Y. Zhang and R. Jeewon thank the Beijing Forestry University and University of Mauritius respectively for research support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shi, H.Z.; Liu, H.M. The current research situation of Blueberry. J. Jiangsu For. Sci. Technol. 2009, 36, 48–51. [Google Scholar]
- Boyzo, M.J.; Rebollar, A.A.; Silva, R.H.V.; Ramirez, M.G. First Report of Neofusicoccum parvum causing stem blight and dieback of blueberry in Mexico. Plant Dis. 2016, 100, 2524. [Google Scholar]
- Castillo, S.; Borrero, C.; Castaño, R.; Rodríguez, A.; Avilés, M. First report of canker disease caused by Neofusicoccum parvum and N australe on blueberry bushes in spain. Plant Dis. 2013, 97, 1112. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.T.; Rooneylatham, S.; Wright, A.F. First report of stem blight of blueberry in California caused by Neofusicoccum parvum. Plant Dis. 2014, 98, 1280. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Qiu, X.L.; Yi, K.S.; Yu, X.F.; Yu, L.; Kong, C.S.; Qiu, X.L.; Yi, K.S.; Yu, X.F.; Yu, L. First Report of Neofusicoccum vitifusiforme Causing Blueberry Blight of Blueberry in China. Plant Dis. 2010, 94, 1373. [Google Scholar] [CrossRef]
- Sammonds, J.; Billones, R.; Ridgway, H.; Walter, M.; Jaspers, M. Survey of blueberry farms for Botryosphaeria dieback and crown rot pathogens. New Zealand Plant Prot. 2009, 62, 238–242. [Google Scholar] [CrossRef] [Green Version]
- Scarlett, K.A.; Shuttleworth, L.A.; Collins, D.; Rothwell, C.; Guest, D.I.; Daniel, R. Botryosphaeriales associated with stem blight and dieback of blueberry (Vaccinium spp.) in New South Wales and Western Australia. Australas. Plant Pathol. 2019, 48, 45–57. [Google Scholar] [CrossRef]
- Wright, A.F.; Harmon, P.F. First Report of Lasiodiplodia theobromae Associated with Stem Blight of Southern Highbush Blueberries in Florida. Plant Dis. 2009, 93, 962. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.F.; Harmon, P.F. Identification of Species in the Botryosphaeriaceae Family Causing Stem Blight on Southern Highbush Blueberry in Florida. Plant Dis. 2010, 94, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.N. Etiology, Molecular Detection and Control of Blueberry Stem Blight in China. Ph.D. Thesis, Hebei Agricultural University, Baoding, China, 2016. [Google Scholar]
- Xu, C.N.; Zhang, H.J.; Zhou, Z.S.; Hu, T.; Wang, S.T.; Wang, Y.N.; Cao, K.Q. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Yu, L.; Impaprasert, R.; Zhao, J.R.; Xu, S.G.; Wu, X. Stem dieback of highbush blueberries caused by Neofusicoccum parvum in China. New Dis. Rep. 2016, 27, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, Y.; He, W.; Zhang, Y. Stem blight of blueberry caused by Lasiodiplodia vaccinii sp. nov. in China. Plant Dis. 2013, 13, 2041–2050. [Google Scholar] [CrossRef]
- Evans, E.A.; Ballen, F.H. An overview of US blueberry production, trade, and consumption, with special reference to Florida. Food Resour. Econ. 2017. Available online: https://edis.ifas.ufl.edu/fe952 (accessed on 15 September 2020).
- Popović, T.; Blagojević, J.; Aleksić, G.; Jelušić, A.; Krnjajić, S.; Milovanović, P. A blight disease on highbush blueberry associated with Macrophomina phaseolina in Serbia. Can. J. Plant Pathol. 2018, 40, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.D.; Sun, H.Y.; Chen, L. China Blueberry Industry Development Report. China Fruits 2016, 5, 1–10. Available online: http://caod.oriprobe.com/articles/49349906/wo_guo_lan_zuo_chan_ye_fa_zhan_bao_gao_.htm (accessed on 15 October 2020).
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.; Groenewald, J.; Crous, P. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Perera, R.H.; De Silva, N.I.; Maharachchikumburua, S.S.N.; Samarakoon, M.C.; Ekanayake, A.H.; et al. One stop shop III: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 51–75 (2019). Fungal Divers. 2019, 98, 77–160. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Ghobad-Nejhad, M.; Wanasinghe, D.N.; Liu, N.; Phillips, A.J.L.; Oliveira-Filho, J.R.C.; Da Silva, G.A.; Gibertoni, T.B.; et al. One stop shop II: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 26–50 (2019). Fungal Divers. 2019, 94, 41–129. [Google Scholar] [CrossRef]
- Poll, J.T.K.; Wood, F.H. Blueberries in New Zealand. Acta Hortic. 1985, 165, 35–46. [Google Scholar] [CrossRef]
- Wang, J.N.; Zhao, H.H.; Yu, Y.Y.; Li, X.D.; Liang, C.; Li, B.D. The pathogen causing Lasiodiplodia twig blight of blueberry. Mycosystema 2016, 35, 657–665. [Google Scholar]
- Milholland, R.D. Factors affecting sporulation and infection by the blueberry stem canker fungus, Botryosphaeria corticis. Phytopathology 1972, 62, 137–139. [Google Scholar] [CrossRef]
- Milholland, R.D. Histopathology and pathogenicity of Botryosphaeria dothidea on blueberry stems. Phytopathology 1972, 62, 654–660. [Google Scholar] [CrossRef]
- Phillips, A.J.; Oudemans, P.V.; Correia, A.; Alves, A. Characterisation and epitypification of Botryosphaeria corticis, the cause of blueberry cane canker. Fungal Divers. 2006, 21, 141–155. [Google Scholar]
- Creswell, T.C.; Milholland, R.D. Spore release and infection periods of Botryosphaeria dothidea on blueberry in North Carolina. Plant Dis. 1988, 72, 342. [Google Scholar] [CrossRef]
- Witcher, W.; Clayton, C.N. Blueberry stem blight caused by Botryosphaeria dothidea. Phytopathology 1962, 53, 705–712. [Google Scholar]
- Tennakoon, K.M.S.; Ridgway, H.J.; Jaspers, M.V.; Jones, E.E. Factors affecting Neofusicoccum ribis, infection and disease progression in blueberry. European journal of plant pathology. Eur. J. Plant Pathol. 2018, 151, 87–99. [Google Scholar]
- Espinoza, J.G.; Briceño, E.X.; Chávez, E.R.; Úrbez-Torres, J.R.; Latorre, B.A. Neofusicoccum spp. associated with stem canker and dieback of blueberry in Chile. Plant Dis. 2009, 93, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Borrero, C.; Castaño, R.; Avilés, M. First report of Pestalotiopsis clavispora (Neopestalotiopsis clavispora) causing canker and twig dieback on blueberry bushes in Spain. Plant Dis. 2009, 102, 1178. [Google Scholar] [CrossRef]
- Choi, I.Y. First report of bark dieback on blueberry caused by Botryosphaeria dothidea in Korea. Plant Dis. 2011, 95, 227. [Google Scholar] [CrossRef]
- Daniel, R.; Scarlett, K.; Rothwell, C.T.; Shuttleworth, L.; Guest, D.I. Botryosphaeriaceae associated with stem blight and dieback of blueberry (Vaccinium corymbosum) in Australia. In Proceedings of the International Congress of Plant Pathology (ICPP) 2018: Plant Health in A Global Economy, Boston, MA, USA, 29 July–3 August 2018; Available online: https://apsnet.confex.com/apsnet/ICPP2018/meetingapp.cgi/Paper/10147 (accessed on 15 August 2020).
- Wiseman, M.S.; Serdani, M.; Putnam, M.L. A New Cane Dieback Disease of Northern Highbush Blueberry in the United States Caused by Lasiodiplodia mediterranea. Plant Dis. 2017, 101, 1317. [Google Scholar] [CrossRef]
- Lopes, A.; Barradas, C.; Phillips, A.J.; Alves, A. Diversity and phylogeny of Neofusicoccum species occurring in forest and urban environments in Portugal. Mycosphere 2016, 7, 906–920. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Does morphology matter in taxonomy of Lasiodiplodia? An answer from Lasiodiplodia hyalina sp nov. Mycosphere 2017, 8, 1014–1027. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.; Hyde, K.D.; Chethana, K.W.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X.H. Mycosphere Essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Gryzenhout, M.; Wingfield, M.J. Lasiodiplodia gonubiensis sp. nov., a new Botryosphaeria anamorph from native Syzygium cordatum in South Africa. Stud Mycol. 2004, 50, 313–322. [Google Scholar]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jeewon, R.; Yeung, Q.S.Y.; Hyde, K.D. Novel phylogenetic group within Thozetella (Chaetosphaeriaceae): A new taxon based on morphology and DNA sequence analyses. Can. J. Microbiol. 2009, 55, 680–687. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Lasiodiplodia chinensis sp. nov., a new holomorphic species from China. Mycosphere 2017, 8, 521–532. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Elsevier: Berlin/Heidelberg, Germany, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–3. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruywagen, E.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Phylogenetic species recognition and hybridisation in Lasiodiplodia: A case study on species from baobabs. Fungal Biol. 2017, 121, 420–436. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, H.K.; Fournier, J.; Crous, P.W.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Towards a phylogenetic clarification of Lophiostoma/Massarina and morphologically similar genera in the Pleosporales. Fungal Divers. 2009, 38, 225–251. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. Paup*: Phylogenetic analysis using parsimony (*and other methods) 4.0. B5. Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jeewon, R.; Fournier, J.; Hyde, K.D. Multi-gene phylogeny and morphotaxonomy of Amniculicola lignicola: A novel freshwater fungus from France and its relationships to the Pleosporales. Mycol. Res. 2008, 112, 1186–1194. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Page, R.D. Tree View: An application to display phylogenetic trees on personal computers. Bioinformatics 1996, 12, 357–358. [Google Scholar] [CrossRef] [Green Version]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia 2015, 34–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, S.; Zhao, L.; Sun, X.; He, W.; Zhang, Y.; Dai, Y.C. Lasiodiplodia spp. associated with Aquilaria crassna in Laos. Mycol. Prog. 2019, 18, 683–701. [Google Scholar] [CrossRef]
- Begoude, B.D.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in Cameroon, South Africa and Madagascar. Mycol Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Boutiti, M.Z.; Jamâa, M.B.; Phillips, A.J. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- de Silva, N.I.; Phillips, A.J.; Liu, J.K.; Lumyong, S.; Hyde, K.D. Phylogeny and morphology of Lasiodiplodia species associated with Magnolia forest plants. Sci. Rep. 2019, 9, 1. Available online: https://www.nature.com/articles/s41598-019-50804-x (accessed on 15 October 2020). [CrossRef] [PubMed]
Figure 1.
Maximum parsimony tree obtained from combined sequence ITS nrDNA, tef1-α, TUB and RPB2 dataset of Lasiodiplodia species. Designated out-group taxon is Diplodia mutila (CMW 7060) and D. seriata (CBS 112555). Bayesian posterior probabilities (PP) support the above 0.7 and maximum parsimony (MP) support values above 50%, are shown on nodes (PP/MP). * represents either PP or MP support values which are below 0.7 (PP) and 50% (MP) respectively. Ex-type strains are printed in bold face and new isolates in red bold face.
Figure 1.
Maximum parsimony tree obtained from combined sequence ITS nrDNA, tef1-α, TUB and RPB2 dataset of Lasiodiplodia species. Designated out-group taxon is Diplodia mutila (CMW 7060) and D. seriata (CBS 112555). Bayesian posterior probabilities (PP) support the above 0.7 and maximum parsimony (MP) support values above 50%, are shown on nodes (PP/MP). * represents either PP or MP support values which are below 0.7 (PP) and 50% (MP) respectively. Ex-type strains are printed in bold face and new isolates in red bold face.

Figure 2.
Lasiodiplodia clavispora (From holotype HMAS 255607). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C,D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, aseptate conidia. Scale bars: B = 1 mm; C–E = 10 µm.
Figure 2.
Lasiodiplodia clavispora (From holotype HMAS 255607). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C,D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, aseptate conidia. Scale bars: B = 1 mm; C–E = 10 µm.

Figure 3.
Lasiodiplodia fujianensis (From holotype HMAS 255606). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Conidia developing on conidiogenous cells between paraphyses. (D). Aseptate and unbranched paraphyses. (E). Hyaline and aseptate conidia. Scale bars: B = 1 mm; C–G = 10 µm.
Figure 3.
Lasiodiplodia fujianensis (From holotype HMAS 255606). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Conidia developing on conidiogenous cells between paraphyses. (D). Aseptate and unbranched paraphyses. (E). Hyaline and aseptate conidia. Scale bars: B = 1 mm; C–G = 10 µm.

Figure 4.
Lasiodiplodia henanica (from holotype HMAS 247961). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Conidia developing on conidiogenous cells. (D). Hyaline and immature conidia with granular content. (E). Conidia with two vacuoles. (F,G) Pigmented, 1-septate conidia in two different focal planes to show the longitudinal striations. Scale bars: B = 1 mm; C–G = 10 µm.
Figure 4.
Lasiodiplodia henanica (from holotype HMAS 247961). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Conidia developing on conidiogenous cells. (D). Hyaline and immature conidia with granular content. (E). Conidia with two vacuoles. (F,G) Pigmented, 1-septate conidia in two different focal planes to show the longitudinal striations. Scale bars: B = 1 mm; C–G = 10 µm.

Figure 5.
Lasiodiplodia nanpingensis (from holotype HMAS 255608). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Developing, aseptate and branched paraphyses. (D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, aseptate conidia. (F). Germinating conidia. Scale bars: B = 1 mm; C–F = 10 µm.
Figure 5.
Lasiodiplodia nanpingensis (from holotype HMAS 255608). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Developing, aseptate and branched paraphyses. (D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, aseptate conidia. (F). Germinating conidia. Scale bars: B = 1 mm; C–F = 10 µm.

Figure 6.
Lasiodiplodia paraphysoides (From holotype HMAS 247959). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Septate or aseptate, unbranched or branched paraphyses. (D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, immature and germinating conidia. (F,G). Pigmented, 1-septate conidia in two different focal planes to show the longitudinal striations. (H). Germinating pigmented conidia. Scale bars: B = 1 mm; C–H = 10 µm.
Figure 6.
Lasiodiplodia paraphysoides (From holotype HMAS 247959). (A). Culture grown on MEA. (B). Conidiomata developing on pine needles in culture. (C). Septate or aseptate, unbranched or branched paraphyses. (D). Conidia developing on conidiogenous cells between paraphyses. (E). Hyaline, immature and germinating conidia. (F,G). Pigmented, 1-septate conidia in two different focal planes to show the longitudinal striations. (H). Germinating pigmented conidia. Scale bars: B = 1 mm; C–H = 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Isolates used in the phylogenetic analysis of Lasiodiplodia spp. and their GenBank accession numbers. Newly generated sequences are indicated in bold. * Type collections.
Table 1.
Isolates used in the phylogenetic analysis of Lasiodiplodia spp. and their GenBank accession numbers. Newly generated sequences are indicated in bold. * Type collections.
| Species | Cultures | Host | Locality | Longitude and Latitude | GenBank | |||
|---|---|---|---|---|---|---|---|---|
| ITS | tef1-α | TUB | RPB2 | |||||
| L. aquilariae | CGMCC 3.18471 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783442 | KY848600 | N/A | KY848562 |
| L. avicenniae | CMW41467 | Avicennia marina | South Africa | 25°44′ S, 28°15′ E * | KP860835 | KP860680 | KP860758 | KU587878 |
| L. avicenniae | LAS199 | Avicennia marina | South Africa | 25°44′ S, 28°15′ E * | KU587957 | KU587947 | KU587868 | KU587880 |
| L. brasiliensis | CMM 2321 | Carica papaya | Brazil * | 15°47′ S, 47°55′ W | KY783475 | KY848612 | KY848556 | KY848595 |
| L. brasiliensis | CMM 4015 | Mangifera indica | Brazil * | 15°47′S, 47°55′W | JX464063 | JX464049 | N/A | N/A |
| L. brasiliensis | CMW 35884 | Adansonia madagascariensis | Madagascar * | 18°52′ S, 47°29′ E | KU887094 | KU886972 | KU887466 | KU696345 |
| L. bruguierae | CMW41470 | Bruguiera gymnorrhiza | South Africa * | 25°44′ S, 28°15′ E | KP860833 | KP860678 | KP860756 | KU587875 |
| L. bruguierae | CMW42480 | Bruguiera gymnorrhiza | South Africa * | 25°44′ S, 28°15′ E | KP860832 | KP860677 | KP860755 | KU587876 |
| L. caatinguensis | CMM1325 | Citrus sinensis | Brazil * | 15°47′ S, 47°55′ W | KT154760 | KT008006 | KT154767 | N/A |
| L. caatinguensis | IBL381 | Spondias purpurea | Brazil * | 15°47′ S, 47°55′ W | KT154757 | KT154751 | KT154764 | N/A |
| L. chinensis | CGMCC3.18044 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | KX499875 | KX499913 | KX499988 | KX499951 |
| L. chinensis | CGMCC3.18061 | unknown | Hainan, China | 20°0′ N, 110°12′ E | KX499889 | KX499927 | KX500002 | KX499965 |
| L. chinensis | CGMCC3.18066 | Hevea brasiliensis | Hainan, China | 20°0′ N, 110°12′ E | KX499899 | KX499937 | KX500012 | KX499974 |
| L. chinensis | CGMCC3.18067 | Sterculia lychnophora | Hainan, China | 20°0′ N, 110°12′ E | KX499901 | KX499939 | KX500014 | KX499976 |
| L. citricola | IRAN1521C | Citrus sp. | Iran * | 33°05′ N, 43°06′ E | GU945353 | GU945339 | KU887504 | KU696350 |
| L. citricola | IRAN1522C | Citrus sp. | Iran * | 33°05′ N, 43°06′ E | GU945354 | GU945340 | KU887505 | KU696351 |
| L. clavispora | CGMCC 3.19594 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802166 | N/A | MK816339 | MK809507 |
| L. clavispora | CGMCC 3.19595 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802165 | N/A | MK816338 | MK809506 |
| L. crassispora | CMW 13488 | Eucalyptus urophylla | Venezuela * | 10°28′ N, 66°53′ W | DQ103552 | DQ103559 | KU887507 | KU696352 |
| L. crassispora | WAC12533 | Santalum album | Australia * | 32° S, 150° E | DQ103550 | DQ103557 | KU887506 | KU696353 |
| L. curvata | CGMCC 3.18456 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783437 | KY848596 | KY848529 | KY848557 |
| L. curvata | CGMCC 3.18476 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783443 | KY848601 | KY848532 | KY848563 |
| L. euphorbicola | CMM 3609 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KU887149 | KU887026 | KU887455 | KU696346 |
| L. euphorbicola | CMW 33350 | Adansonia digitata | Botswana * | 24°36′ S, 25°40′ E | KU887187 | KU887063 | KU887494 | KU696347 |
| L. euphorbicola | CMW 36231 | Adansonia digitata | Zimbabwe * | 17°49′ S, 31°03′ E | KF234543 | KF226689 | KF254926 | N/A |
| L. exigua | BL184 | Retama raetam | Tunisia * | 34°44′ N, 10°44′ E | KJ638318 | KJ638337 | N/A | N/A |
| L. exigua | CBS 137785 | Retama raetam | Tunisia * | 34°44′ N, 10°44′ E | KJ638317 | KJ638336 | KU887509 | KU696355 |
| L. fujianensis | CGMCC 3.19593 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802164 | MK887178 | MK816337 | MK809505 |
| L. gilanensis | IRAN 1501C | Unknown | Iran * | 33°05′ N, 43°06′ E | GU945352 | GU945341 | KU887510 | KU696356 |
| L. gilanensis | IRAN 1523C | Unknown | Iran * | 33°05′ N, 43°06′ E | GU945351 | GU945342 | KU887511 | KU696357 |
| L. gonubiensis | CMW 14077 | Syzygium cordatum | South Africa * | 25°44′ S, 28°15′ E | AY639595 | DQ103566 | DQ458860 | KU696359 |
| L. gonubiensis | CMW 14078 | Syzygium cordatum | South Africa * | 25°44′ S, 28°15′ E | AY639594 | DQ103567 | EU673126 | KU696358 |
| L. gravistriata | CMM 4564 | Anacardium humile | Brazil * | 15°47′ S, 47°55′ W | KT250949 | KT250950 | N/A | N/A |
| L. gravistriata | CMM 4565 | Anacardium humile | Brazil * | 15°47′ S, 47°55′ W | KT250947 | KT266812 | N/A | N/A |
| L. henanica | CGMCC 3.19176 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729351 | MH729357 | MH729360 | MH729354 |
| L. hormozganensis | IRAN 1498C | Mangifera indica | Iran * | 33°05′ N, 43°06′ E | GU945356 | GU945344 | KU887514 | KU696360 |
| L. hormozganensis | IRAN 1500C | Olea sp. | Iran * | 33°05′ N, 43°06′ E | GU945355 | GU945343 | KU887515 | KU696361 |
| L. iraniensis | CMM 3610 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234544 | KF226690 | KF254927 | N/A |
| L. iraniensis | CMW 36237 | Adansonia digitata | Mozambique * | 25°56′ S, 32°35′ E | KU887121 | KU886998 | KU887499 | KU696348 |
| L. iraniensis | CMW 36239 | Adansonia digitata | Mozambique * | 25°56′ S, 32°35′ E | KU887123 | KU887000 | KU887501 | KU696349 |
| L. iraniensis | IRAN 1502C | Juglans sp. | Iran * | 33°05′ N, 43°06′ E | GU945347 | GU945335 | KU887517 | KU696362 |
| L. iraniensis | IRAN 1520C | Salvadora persica | Iran * | 33°05′ N, 43°06′ E | GU945348 | GU945336 | KU887516 | KU696363 |
| L. irregularis | CGMCC 3.18468 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783472 | KY848610 | KY848553 | KY848592 |
| L. laeliocattleyae | BOT 29 | Mangifera indica | Egypt * | 30°03′ N, 31°14′ E | JN814401 | JN814428 | N/A | N/A |
| L. laeliocattleyae | CBS 130992 | Mangifera indica | Egypt * | 30°03′ N, 31°14′ E | JN814397 | JN814424 | KU887508 | KU696354 |
| L. laosensis | CGMCC 3.18464 | Aquilaria crassna | Laos | 17°59′N, 102°34′E | KY783471 | KY848609 | KY848552 | KY848591 |
| L. laosensis | CGMCC 3.18473 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783450 | KY848603 | KY848536 | KY848570 |
| L. lignicola | CBS 134112 | dead wood | Thailand * | 13°43′ N, 100°28′ E | JX646797 | KU887003 | JX646845 | KU696364 |
| L. lignicola | CGMCC 3.18460 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783462 | N/A | N/A | KY848582 |
| L. lignicola | CGMCC 3.18483 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783449 | N/A | N/A | KY848569 |
| L. lignicola | MFLUCC 11-0656 | dead wood | Thailand * | 13°43′ N, 100°28′ E | JX646798 | N/A | JX646846 | N/A |
| L. macroconidica | CGMCC 3.18479 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783438 | KY848597 | KY848530 | KY848558 |
| L. macrospora | CMM3833 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234557 | KF226718 | KF254941 | N/A |
| L. mahajangana | CMW 27801 | Terminalia catappa | Madagascar * | 18°52′ S, 47°29′ E | FJ900595 | FJ900641 | FJ900630 | KU696365 |
| L. mahajangana | CMW 27818 | Terminalia catappa | Madagascar * | 18°52′ S, 47°29′ E | FJ900596 | FJ900642 | FJ900631 | KU696366 |
| L. margaritacea | CBS 122065 | Adansonia gibbosa | Western Australia * | 31°56′ S, 115°55′ E | EU144051 | EU144066 | N/A | N/A |
| L. margaritacea | CBS 122519 | Adansonia gibbosa | Western Australia * | 31°56′ S, 115°55′ E | EU144050 | EU144065 | KU887520 | KU696367 |
| L. mediterranea | CBS 137783 | Quercus ilex | Italy * | 41°54′ N, 12°18′ E | KJ638312 | KJ638331 | KU887521 | KU696368 |
| L. mediterranea | CBS 137784 | Vitis vinifera | Italy * | 41°54′ N, 12°18′ E | KJ638311 | KJ638330 | KU887522 | KU696369 |
| L. microcondia | CGMCC 3.18485 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783441 | KY848614 | N/A | KY848561 |
| L. missouriana | UCD 2193MO | Vitis sp. | USA * | 38° N, 97° W | HQ288225 | HQ288267 | HQ288304 | KU696370 |
| L. missouriana | UCD 2199MO | Vitis sp. | USA * | 38° N, 97° W | HQ288226 | HQ288268 | HQ288305 | KU696371 |
| L. nanpingensis | CGMCC 3.19596 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802167 | N/A | MK816340 | MK809508 |
| L. nanpingensis | CGMCC 3.19597 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802168 | N/A | MK816341 | MK809509 |
| L. paraphysoides | CGMCC 3.19174 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729349 | MH729355 | MH729358 | MH729352 |
| L. paraphysoides | CGMCC 3.19175 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729350 | MH729356 | MH729359 | MH729353 |
| L. parva | CBS 456.78 | Cassava field-soil | Colombia, USA | 34°0′ N, 81°1′ W | EF622083 | EF622063 | KU887523 | KU696372 |
| L. parva | CBS 494.78 | Cassava field-soil | Colombia, USA | 34°0′ N, 81°1′ W | EF622084 | EF622064 | EU673114 | KU696373 |
| L. plurivora | STE-U 4583 | Vitis vinifera | South Africa * | 25°44′ S, 28°15′ E | AY343482 | EF445396 | KU887525 | KU696375 |
| L. plurivora | STE-U 5803 | Prunus salicina | South Africa * | 25°44′ S, 28°15′ E | EF445362 | EF445362 | EF445362 | EF445362 |
| L. pontae | CMM1277 | Spondias purpurea | Brazil * | 15°47′ S, 47°55′ W | KT151794 | KT151791 | KT151797 | N/A |
| L. pseudotheobromae | CBS 116459 | Gmelina arborea | Costa Rica * | 9°55′ N, 84°3′ W | EF622077 | EF622057 | EU673111 | KU696376 |
| L. pseudotheobromae | CGMCC 3.18047 | Pteridium aquilinum | China * | 39°54′ N, 116°23′ E | KX499876 | KX499914 | KX499989 | KX499952 |
| L. pseudotheobromae | CGMCC 3.18451 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783468 | KY848621 | N/A | KY848588 |
| L. pseudotheobromae | CGMCC 3.18452 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783467 | KY848620 | KY848549 | KY848587 |
| L. pseudotheobromae | CGMCC 3.18453 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783460 | KY848618 | KY848545 | KY848580 |
| L. pseudotheobromae | CGMCC 3.18457 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783436 | KY848613 | N/A | N/A |
| L. pseudotheobromae | CGMCC 3.18461 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783446 | N/A | N/A | KY848566 |
| L. pseudotheobromae | CGMCC 3.18465 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783445 | N/A | N/A | KY848565 |
| L. pseudotheobromae | CGMCC 3.18466 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783444 | KY848615 | KY848533 | KY848564 |
| L. pseudotheobromae | CGMCC 3.18470 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783458 | N/A | N/A | KY848578 |
| L. pseudotheobromae | CGMCC 3.18474 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783452 | N/A | KY848538 | KY848572 |
| L. pseudotheobromae | CGMCC 3.18475 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783459 | KY848617 | KY848544 | KY848579 |
| L. pyriformis | CBS 121770 | Acacia mellifera | Namibia * | 22°33′S, 17°04′E | EU101307 | EU101352 | KU887527 | KU696378 |
| L. pyriformis | CBS 121771 | Acacia mellifera | Namibia * | 22°33′ S, 17°04′ E | EU101308 | EU101353 | KU887528 | KU887528 |
| L. rubropurpurea | WAC 12535 | Eucalyptus grandis | Australia * | 32° S, 151° E | DQ103553 | DQ103571 | EU673136 | KU696380 |
| L. rubropurpurea | WAC 12536 | Eucalyptus grandis | Australia * | 32° S, 152° E | DQ103554 | DQ103572 | KU887530 | KU696381 |
| L. sterculiae | CBS342.78 | Sterculia oblonga | Germany * | 52°31′ N, 13°26 E | KX464140 | KX464634 | KX464908 | KX463989 |
| L. subglobosa | CMM3872 | Jatropha curcas | Brazil * | 15°47′S, 47°55′W | KF234558 | KF226721 | KF254942 | N/A |
| L. subglobosa | CMM4046 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234560 | KF226723 | KF254944 | N/A |
| L. tenuiconidia | CGMCC 3.18449 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783466 | KY848619 | N/A | KY848586 |
| L. thailandica | CBS 138653 | Phyllanthus acidus | Thailand * | 13°43′ N, 100°28′ E | KM006433 | KM006464 | N/A | N/A |
| L. thailandica | CBS 138760 | Mangifera indica | Thailand * | 13°43′ N, 100°28′ E | KJ193637 | KJ193681 | N/A | N/A |
| L. theobromae | CBS 111530 | Fruit along coral reef coast | Papua New Guinea * | 9°25′ S, 147°22′ E | EF622074 | EF622054 | KU887531 | KU696382 |
| L. theobromae | CBS 164.96 | Unknown | Unknown | – | AY640255 | AY640258 | KU887532 | KU696383 |
| L. tropica | CGMCC 3.18477 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783454 | KY848616 | KY848540 | KY848574 |
| L. venezuelensis | WAC 12539 | Acacia mangium | Venezuela * | 10°28′ N, 66°53′ W | DQ103547 | DQ103568 | KU887533 | KU696384 |
| L. venezuelensis | WAC 12540 | Acacia mangium | Venezuela * | 10°28′ N, 66°53′ W | DQ103548 | DQ103569 | KU887534 | KU887534 |
| L. viticola | UCD 2553AR | Vitis sp. | USA * | 38° N, 97° W | HQ288227 | HQ288269 | HQ288306 | KU696385 |
| L. viticola | UCD 2604MO | Vitis sp. | USA * | 38° N, 97° W | HQ288228 | HQ288270 | HQ288307 | KU696386 |
| L. vitis | CBS 124060 | Vitis vinifera | Italy * | 41°54′ N, 12°18′ E | KX464148 | KX464642 | KX464917 | KX463994 |
| Diplodia mutila | CMW 7060 | Fraxinus excelsior | Netherlands * | 52°22′ N, 4°51′ E | AY236955 | AY236904 | AY236933 | EU339574 |
| D. seriata | CBS 112555 | Vitis vinifera | Portugal | 38°43′ N, 9°7′ W | AY259094 | AY573220 | DQ458856 | N/A |
Table 2.
Tef1-α position of mismatch of L. clavispora, L. gonubiensis, L. fujianensis, L. thailandica, L. paraphysoides and L. citricola.
Table 2.
Tef1-α position of mismatch of L. clavispora, L. gonubiensis, L. fujianensis, L. thailandica, L. paraphysoides and L. citricola.
| Species | Base Pair Difference | Nucleotides Difference (tef1-α) |
|---|---|---|
| L. clavispora and L. gonubiensis | A instead of G | 30 |
| T instead of G | 33 | |
| T instead of gap | 35, 36, 37 | |
| G instead of gap | 38, 42 | |
| C instead of gap | 39, 40, 41 | |
| C instead of T | 44, 48, 105 | |
| G instead of A | 121 | |
| L. fujianensis and L. thailandica | T instead of C | 4 |
| A instead of G | 7 | |
| C instead of A | 27 | |
| gap instead of C | 71, 74, 76 | |
| gap instead of A | 72 | |
| gap instead of G | 73, 75, 78 | |
| gap instead of T | 77 | |
| C instead of T | 92, 153, 296 | |
| C instead of G | 185 | |
| G instead of C | 495 | |
| L. paraphysoides and L. citricola | gap instead of A | 9 |
| T instead of C | 111 | |
| gap instead of G | 197 | |
| A instead of G | 248 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Zhang, Y.; Bhoyroo, V.; Rampadarath, S.; Jeewon, R. Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries. Life 2021, 11, 657. https://doi.org/10.3390/life11070657
AMA Style
Wang Y, Zhang Y, Bhoyroo V, Rampadarath S, Jeewon R. Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries. Life. 2021; 11(7):657. https://doi.org/10.3390/life11070657
Chicago/Turabian StyleWang, Yu, Ying Zhang, Vishwakalyan Bhoyroo, Sillma Rampadarath, and Rajesh Jeewon. 2021. "Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries" Life 11, no. 7: 657. https://doi.org/10.3390/life11070657
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.