Influence of Bacterial Secondary Symbionts in Sitobion avenae on Its Survival Fitness against Entomopathogenic Fungi, Beauveria bassiana and Metarhizium brunneum

 , , , , ,
, , , , ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Cultures
2.2. Exclusion of BSS
2.3. PCR Protocol for the Detection of BSS
2.4. Fungal Cultures
2.5. Bioassay
2.6. Fecundity Assay
2.7. Fungal Growth Assay
2.8. Statistical Analysis
3. Results
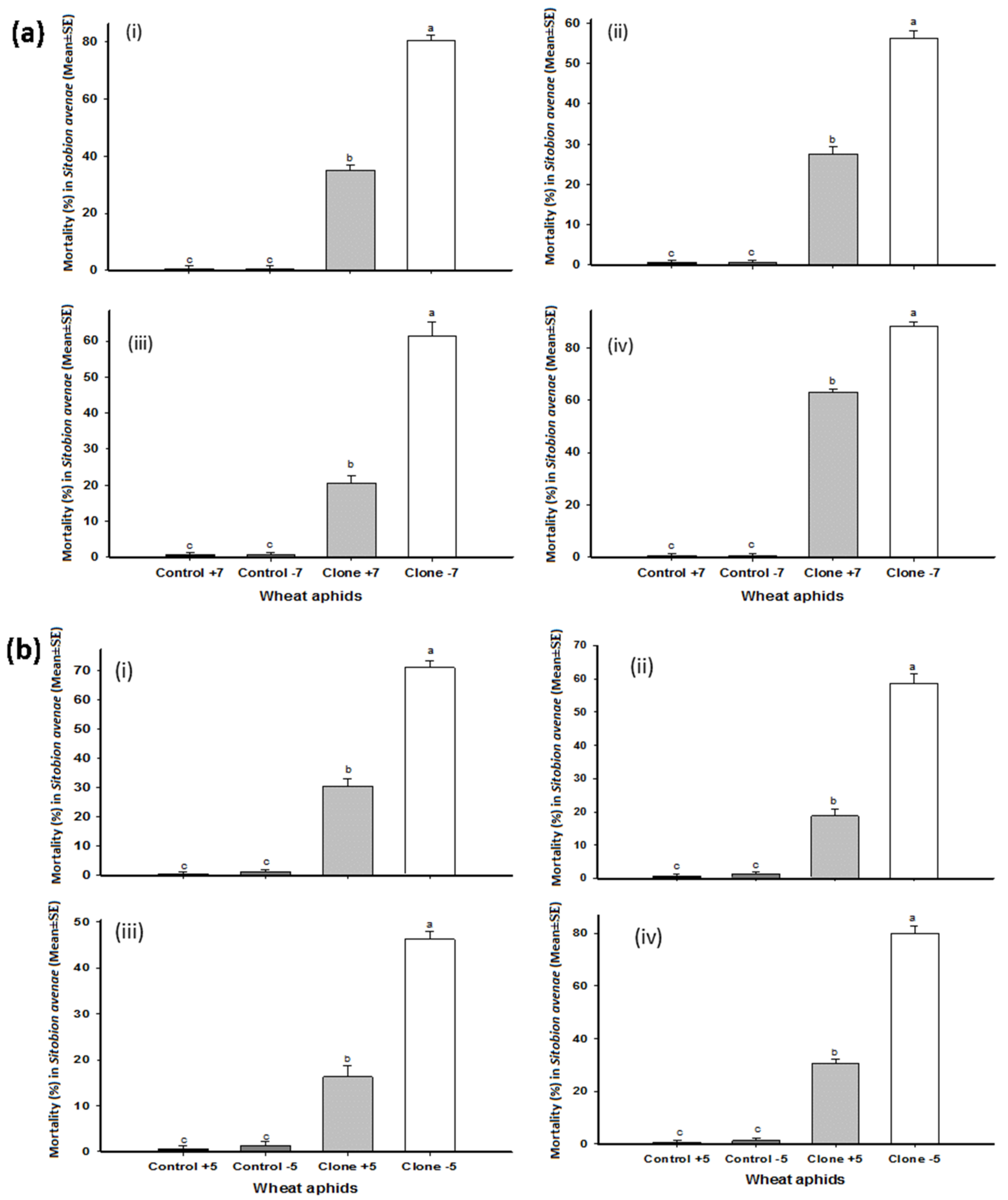
3.1. Total Mortality in Wheat Aphid
3.2. Fungal Radial Growth
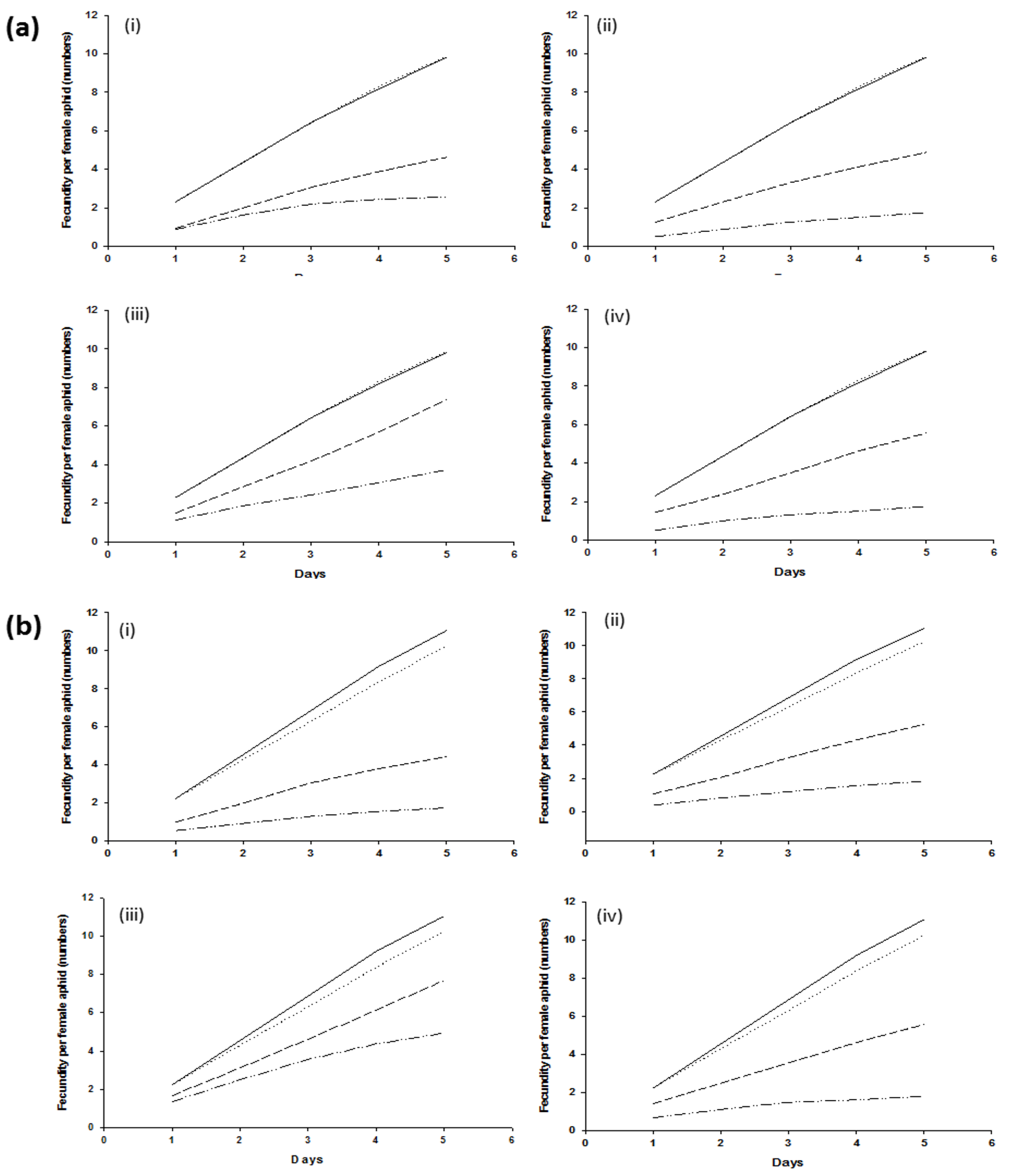
3.3. Fecundity Assay
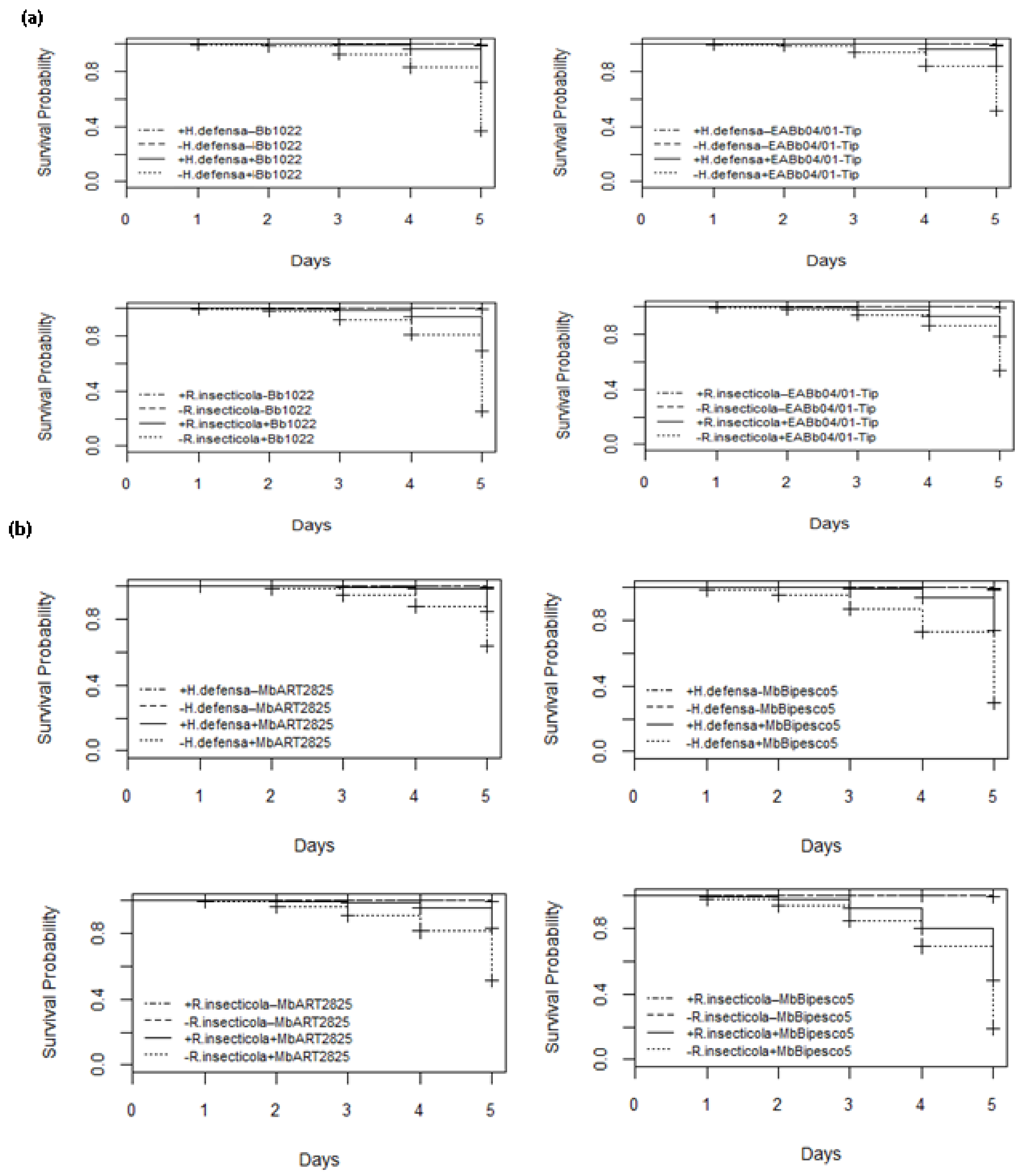
3.4. Infection Period
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louie, A. Relational biology of symbiosis. Axiomathes 2010, 20, 495–509. [Google Scholar] [CrossRef]
- Zientz, E.; Silva, F.J.; Gross, R. Genome interdependence in insect-bacterium symbioses. Genome Biol. 2001, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y. Endosymbiotic bacteria in insects: Their diversity and culturability. Microbes Environ. 2009, 24, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourtzis, K.; Miller, T.A. Insect Symbiosis, 1st ed.; Bourtzis, K., Miller, T.A., Eds.; CRC Press: Boca Raton, FL, USA, 2006; Volume 2, pp. 1–304. [Google Scholar]
- Moran, N.A. Bacterial menageries inside insects. Proc. Natl. Acad. Sci. USA 2001, 98, 1338–1340. [Google Scholar] [CrossRef] [Green Version]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A. Phloem-sap feeding by animals: Problems and solutions. J. Exp. Bot. 2006, 57, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Prado, S.S.; Almeida, R.P. Role of symbiotic gut bacteria in the development of Acrosternum hilare and Murgantia histrionica. Entomol. Exp. Appl. 2009, 32, 21–29. [Google Scholar] [CrossRef]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Dion, E.; Polin, S.E.; Simon, J.C.; Outreman, Y. Symbiont infection affects aphid defensive behaviours. Biol. Lett. 2011, 7, 743–746. [Google Scholar] [CrossRef]
- Montllor, C.B.; Maxmen, A.; Purcell, A.H. Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol. Entomol. 2002, 27, 189–195. [Google Scholar] [CrossRef]
- Russell, J.A.; Moran, N.A. Costs and benefits of symbiont infection in aphids: Variation among symbionts and across temperatures. Proc. R. Soc. B 2006, 273, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Coga, R.; Shibao, H.; Matsumoto, T.; Fukatsu, T. Diversity and geographic distribution of secondary endosymbiotic bacteria in natural populations of the pea aphid, Acyrthosiphon pisum. Mol. Ecol. 2002, 11, 2123–2135. [Google Scholar] [CrossRef]
- Ferrari, J.; Darby, A.C.; Daniell, T.J.; Godfray, C.H.; Douglas, A.E. Linking the bacterial community in pea aphids with host-plant use and natural enemy resistance. Ecol. Entomol. 2004, 29, 60–65. [Google Scholar] [CrossRef]
- McLean, A.; van Ash, M.; Ferrari, J.; Godfray, C.H. Effects of bacterial secondary symbionts on host plant use in pea aphids. Proc. R. Soc. B 2011, 278, 760–766. [Google Scholar] [CrossRef]
- Moran, N.A.; Russel, J.A.; Koga, R.; Fukatsu, T. Evolutionary relationships of three new species of Enterobacteriaceae living as symbionts of aphids and other insects. Appl. Environ. Microbiol. 2005, 71, 3302–3310. [Google Scholar] [CrossRef] [Green Version]
- Scarborough, C.L.; Ferrari, J.; Godfray, C.H. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef]
- Łukasik, P.; Guo, H.; van Asch, M.; Ferrari, J.; Godfray, C.H. Protection against a fungal pathogen conferred by the aphid facultative endosymbionts Rickettsia and Spiroplasma is expressed in multiple host genotypes and species and is not influenced by co-infection with another symbiont. J. Evol. Biol. 2013, 26, 2654–2661. [Google Scholar] [CrossRef]
- Parker, B.J.; Sprag, C.; Altincicek, B.; Gerardo, N.M. Symbiont-mediated protection against fungal pathogens in pea aphids: A role for pathogen specificity? Appl. Environ. Microbiol. 2013, 79, 2455–2458. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, S.H. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef]
- Costopoulos, K.; Kovacs, J.L.; Kamins, A.; Gerardo, N.M. Aphid facultative symbionts reduce survival of the predatory lady beetle Hippodamia convergens. BMC Ecol. 2014, 14, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haine, E.R. Symbiont-mediated protection. Proc. Biol. Sci. R. Soc. B 2008, 275, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, K.M.; Campos, J.; Moran, N.A.; Hunter, M.S. Population dynamics of defensive symbionts in aphids. Proc. Biol. Sci. R. Soc. B 2008, 275, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, J.; Vavre, F. Bacterial symbionts in insects or the story of communities affecting communities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1389–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, H. A crop loss model and economic thresholds for the grain aphid, Sitobion avenae (F.), in winter wheat in southern Sweden. Crop Prot. 2005, 24, 397–405. [Google Scholar] [CrossRef]
- Dedryver, C.-A.; Le Gallic, J.-F.; Haack, L.; Halkett, F.; Outreman, Y.; Simon, J.C. Seasonal and annual genotypic variation and the effect of climate on population genetic structure of the cereal aphid Sitobion avenae in northern France. Bull. Entomol. Res. 2008, 98, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Alkhedir, H.; Karlovsky, P.; Vidal, S. Effect of light intensity on colour morph formation and performance of the grain aphid Sitobion avenae F. (Homoptera: Aphididae). J. Insect Physiol. 2010, 56, 1999–2005. [Google Scholar] [CrossRef]
- Liu, X.; Yang, X.F.; Wang, C.Y.; Wang, J.Y.; Zang, H.; Ji, W.Q. Molecular mapping of resistance gene to English grain aphid (Sitobion avenae F.) in Triticum durum wheat line C273. Theor. Appl. Genet. 2012, 124, 287–293. [Google Scholar] [CrossRef]
- Ambethgar, V. Potential of entomopathogenic fungi in insecticide resistance management (IRM): A review. J. Biopest. 2009, 2, 177–193. [Google Scholar]
- Dewar, A.M.; Dewar, A.J.; Haylock, L.A.; Foster, S.P.; Williamson, M.S. Alternative insecticides to control cereal aphids, Sitobion avenae, that are resistant to pyrethroids. In Proceedings of the Crop Protection in Northern Britain, Dundee, UK, 25–26 February 2014; pp. 131–136. [Google Scholar]
- Mulé, R.; Sabella, G.; Robba, L.; Manachini, B. Systematic Review of the Effects of Chemical Insecticides on Four Common Butterfly Families. Front. Environ. Sci. 2017, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.I.; Kim, K. Selection of entomopathogenic fungi for aphid control. J. Biosci. Bioeng. 2007, 104, 498–505. [Google Scholar]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Goettel, M.; Eilenberg, J.; Glare, T. Entomopathogenic Fungi and their Role in Regulation of Insect Populations. In Insect Control: Biological and Synthetic Agents, 1st ed.; Lawrence, I., Gill, S., Eds.; Academic Press: London, UK, 2005; Volume 1, pp. 387–438. [Google Scholar]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]
- EC (European Commission). Commission Regulation (EU) 2021/2010 of 17 November 2021 amending Regulation (EC) No 1185/2009 of the European Parliament and of the Council concerning statistics on pesticides, as regards the list of active substances. Off. J. Eur. Union 2021, L 410, 1–47. [Google Scholar]
- Pathan, A.A.K.; Devi, K.U.; Vogel, H.; Reineke, A. Analysis of differential gene expression in the generalist entomopathogenic fungus Beauveria bassiana (Bals.) Vuillemin grown on different insect cuticular extracts and synthetic medium through cDNA-AFLPs. Fungal Genet. Biol. 2007, 44, 1231–1241. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Kim, J.J.; Jeong, G.; Han, J.H.; Lee, S. Biological control of aphid using fungal culture and culture filtrates of Beauveria bassiana. Mycobiology 2013, 41, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.A.; Dunlap, C.A.; Jaronski, S.T. Ecological considerations in producing and formulating fungal entomopathogens for use in insect biocontrol. Bio. Control 2010, 55, 129–145. [Google Scholar] [CrossRef]
- Alkhedir, H.; Karlovsky, P.; Vidal, S. Relationship between water soluble carbohydrate content, aphid endosymbionts and clonal performance of Sitobion avenae on cocksfoot cultivars. PLoS ONE 2013, 8, e54327. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, T.E. Removal of a specialization-associated symbiont does not affect aphid fitness. Ecol. Lett. 2004, 7, 461–468. [Google Scholar] [CrossRef]
- Simon, J.C.; Boutin, S.; Tsuchida, T.; Koga, R.; Le Gallic, J.F.; Frantz, A.; Outreman, Y.; Fukatsu, T. Facultative symbiont infections affect aphid reproduction. PLoS ONE 2011, 6, e21831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, R.; Tsuchida, T.; Sakurai, M.; Fukatsu, T. Selective elimination of aphid endosymbionts: Effects of antibiotic dose and host genotype, and fitness consequences. FEMS Microbiol. Ecol. 2007, 60, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Rangasamy, M.; Tan, S.Y.; Wang, H.; Siegfried, B.D. Evaluation of five methods for total DNA extraction from western corn rootworm beetles. PLoS ONE 2010, 5, e11963. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant-endophyte-herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Quesada-Moraga, E.; Muñoz-Ledesma, F.J.; Santiago-Álvarez, C. Systemic protection of Papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an endophytic strain of Beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckard, S.; Ansari, M.A.; Bacher, S.; Butt, T.M.; Enkerli, J.; Grabenweger, G. Virulence of in vivo and in vitro produced conidia of Metarhizium brunneum strains for control of wireworms. Crop Prot. 2014, 64, 137–142. [Google Scholar] [CrossRef]
- Kölliker, U.; Biasio, L.; Jossi, W. Potential control of Swiss wireworms with entomopathogenic fungi. Potential Control. Swiss Wireworms Entomopathog. Fungi 2011, 66, 517–520. [Google Scholar]
- Johnson, D.M.; White, R.L.; Pereira, R.M.; Geden, C.J. Beauveria bassiana culturing and harvesting for bioassays with house flies. J. Insect Sci. 2020, 20, 14. [Google Scholar] [CrossRef]
- Strasser, H.; Forer, A.; Schinner, F. Development of media for the selective isolation and maintenance of virulence of Beauveria brongniartii. In Proceedings of the 3rd International Workshop on Microbial Control of Soil Dwelling Pests, Lincoln, New Zealand, 21–23 February 1996. [Google Scholar]
- Statistix 8.1, (Version 8.1); Analytical Software: Tallahassee, FL, USA, 2003.
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Godfray, C.H. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecol. Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef]
- Castillo, M.A.; Moya, P.; Hernández, E.; Primo-Yufera, E. Susceptibility of Ceratitis capitata Wiedemann (Diptera, Tephritidae) to entomopathogenic fungi and their extracts. Biol. Control 2000, 19, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.K.; Vorburger, C.; Moran, N.A. Genomic basis of endosymbiont-conferred protection against an insect parasitoid. Genome Res. 2012, 22, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Guay, J.F.; Boudreault, S.; Michaud, D.; Cloutier, C. Impact of environmental stress on aphid clonal resistance to parasitoids: Role of Hamiltonella defensa bacterial symbiosis in association with a new facultative symbiont of the pea aphid. J. Insect Physiol. 2009, 55, 919–926. [Google Scholar] [CrossRef]
- Cayetano, L.; Vorburger, C. Genotype-by-genotype specificity remains robust to average temperature variation in an aphid/endosymbiont/parasitoid system. J. Evol. Biol. 2013, 26, 1603–1610. [Google Scholar] [CrossRef]
- Romo, C.M.; Tylianakis, J.M. Elevated temperature and drought interact to reduce parasitoid effectiveness in suppressing hosts. PLoS ONE 2013, 8, e58136. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.; Sieber, R.; Zimmermann, Y.S.; Vorburger, C. Development, specificity and sublethal effects of symbiont-conferred resistance to parasitoids in aphids. Funct. Ecol. 2012, 26, 207–215. [Google Scholar] [CrossRef]
- Hajek, A.E.; Delalibera, I. Fungal pathogens as classical biological control agents against arthropods. Bio. Control 2010, 55, 147–158. [Google Scholar] [CrossRef]

 ) shows control with BSS, (
) shows control with BSS, ( ) shows control without BSS, (
) shows control without BSS, ( ) shows treatment with BSS, and (
) shows treatment with BSS, and ( ) shows treatment without BSS.
) shows control with BSS, () shows control without BSS, () shows treatment with BSS, and () shows treatment without BSS.
) shows treatment without BSS.
) shows control with BSS, () shows control without BSS, () shows treatment with BSS, and () shows treatment without BSS.


{kind=link}
{kind=link}
{kind=link}
| Fungus Genotype | Strain | Geographic Origin | Insect Host | Reference |
|---|---|---|---|---|
| Beauveria bassiana | Bb1022 | Canada | Rhyacionia buoliana | Vidal & Jaber, 2015 [47] |
| EABb04/01-Tip | Spain | Iraella luteipes | Quesada-Moraga et al., 2009 [48] | |
| Metarrhizium brunneum | BIPESCO 5/F52 | Austria | Cydia pomonella | Eckard et al., 2014 [49] |
| ART2825 | Switzerland | Agriotes obscurus | Kölliker et al., 2011 [50] |
| Beauveria bassiana | Metarhizium brunneum | |||||||
|---|---|---|---|---|---|---|---|---|
| Days | Bb1022 | EABb 04/01-Tip | ART 2825 | BIPESCO 5 | ||||
| Clone +5 | Clone −5 | Clone +5 | Clone −5 | Clone +5 | Clone −5 | Clone +5 | Clone −5 | |
| 5 | 3.9 ± 0.2 a | 3.6 ± 0.1 a | 4.4 ± 0.2 a | 4.3 ± 0.1 a | 2.3 ± 0.2 b | 3.5 ± 0.1 a | 4.7 ± 0.2 a | 5.3 ± 0.2 a |
| 7 | 7.2 ± 0.1 a | 7.1 ± 0.1 a | 7.1 ± 0.2 a | 7.5 ± 0.1 a | 4.3 ± 0.3 b | 5.5 ± 0.2 a | 7.9 ± 0.4 b | 8.9 ± 0.3 a |
| 9 | 10.3 ± 0.2 a | 10.3 ± 0.2 a | 10.5 ± 0.2 a | 10.6 ± 0.2 a | 7.4 ± 0.4 a | 7.9 ± 0.2 a | 11.7 ± 0.4 a | 11.8 ± 0.5 a |
| Clone +7 | Clone −7 | Clone +7 | Clone −7 | Clone +7 | Clone −7 | Clone +7 | Clone −7 | |
| 5 | 3.3 ± 0.2 a | 3.3 ± 0.1 a | 4.2 ± 0.1 a | 4.5 ± 0.1 a | 2.9 ± 0.1 b | 3.5 ± 0.3 a | 4.1 ± 0.2 b | 5.2 ± 0.2 a |
| 7 | 6.7 ± 0.2 a | 6.7 ± 0.2 a | 7.2 ± 0.2 a | 7.2 ± 0.1 a | 4.5 ± 0.1 b | 5.8 ± 0.4 a | 7.6 ± 0.3 b | 8.3 ± 0.3 a |
| 9 | 9.3 ± 0.2 a | 9.6 ± 0.2 a | 9.8 ± 0.3 a | 9.9 ± 0.2 a | 6.9 ± 0.2 b | 8.1 ± 0.4 a | 10.9 ± 0.5 a | 11.3 ± 0.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.; Sajjad, A.; Shakeel, Q.; Farooqi, M.A.; Aqueel, M.A.; Tariq, K.; Ullah, M.I.; Iqbal, A.; Jamal, A.; Saeed, M.F.; et al. Influence of Bacterial Secondary Symbionts in Sitobion avenae on Its Survival Fitness against Entomopathogenic Fungi, Beauveria bassiana and Metarhizium brunneum. Insects 2022, 13, 1037. https://doi.org/10.3390/insects13111037
Ali S, Sajjad A, Shakeel Q, Farooqi MA, Aqueel MA, Tariq K, Ullah MI, Iqbal A, Jamal A, Saeed MF, et al. Influence of Bacterial Secondary Symbionts in Sitobion avenae on Its Survival Fitness against Entomopathogenic Fungi, Beauveria bassiana and Metarhizium brunneum. Insects. 2022; 13(11):1037. https://doi.org/10.3390/insects13111037
Chicago/Turabian StyleAli, Sajjad, Asif Sajjad, Qaiser Shakeel, M. Aslam Farooqi, M. Anjum Aqueel, Kaleem Tariq, Muhammad Irfan Ullah, Aamir Iqbal, Aftab Jamal, Muhammad Farhan Saeed, and et al. 2022. "Influence of Bacterial Secondary Symbionts in Sitobion avenae on Its Survival Fitness against Entomopathogenic Fungi, Beauveria bassiana and Metarhizium brunneum" Insects 13, no. 11: 1037. https://doi.org/10.3390/insects13111037