Reproductive Parameters and Host Specificity of Eurytoma erythrinae (Hymenoptera: Eurytomidae), a Biological Control Agent of the Erythrina Gall Wasp, Quadrastichus erythrinae (Hymenoptera: Eulophidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Propagation
2.2. EGW Propagation
2.3. Origin of the Parasitoid Colony
2.4. Eurytoma Erythrinae Propagation
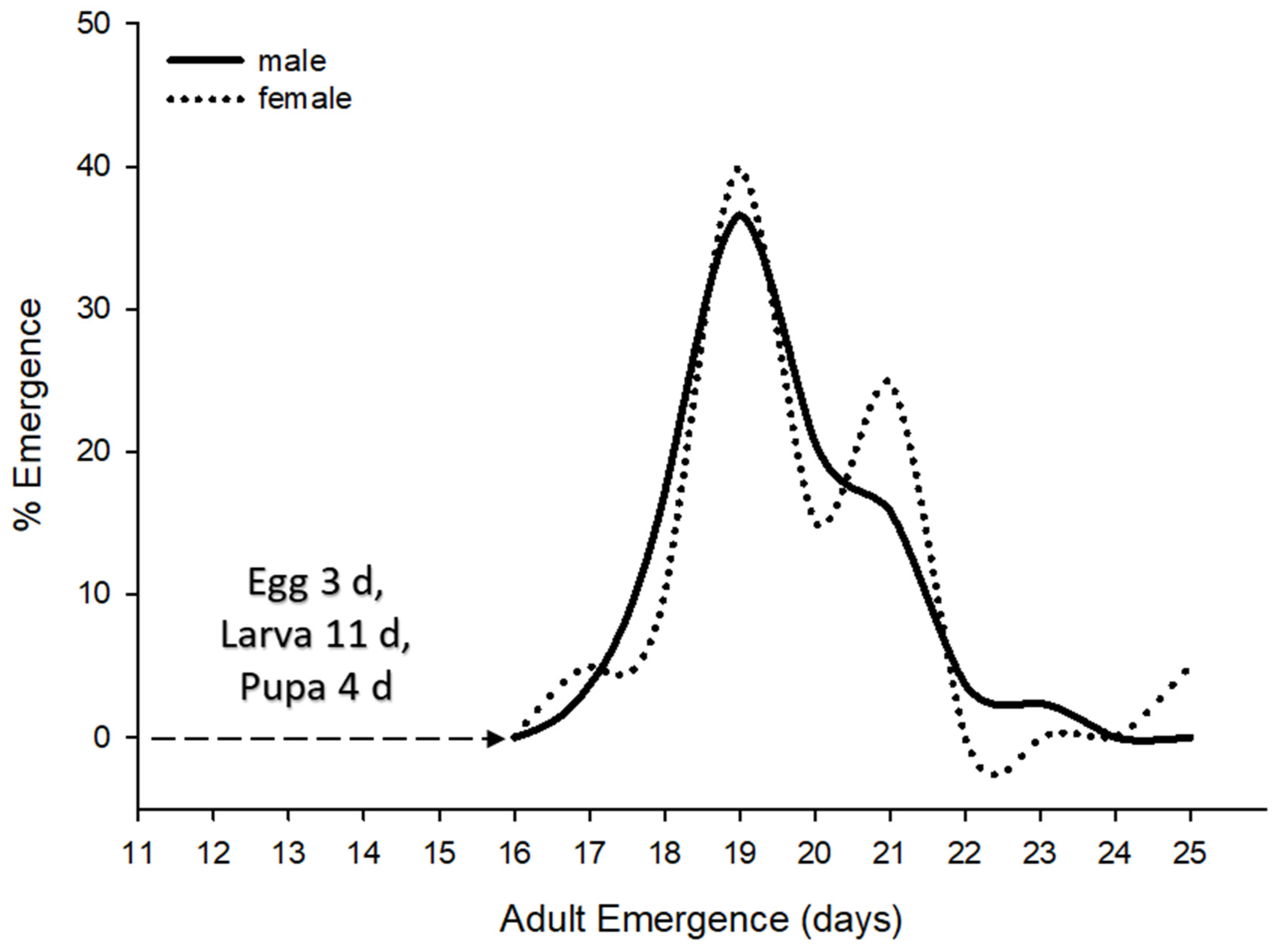
2.5. Life History Study
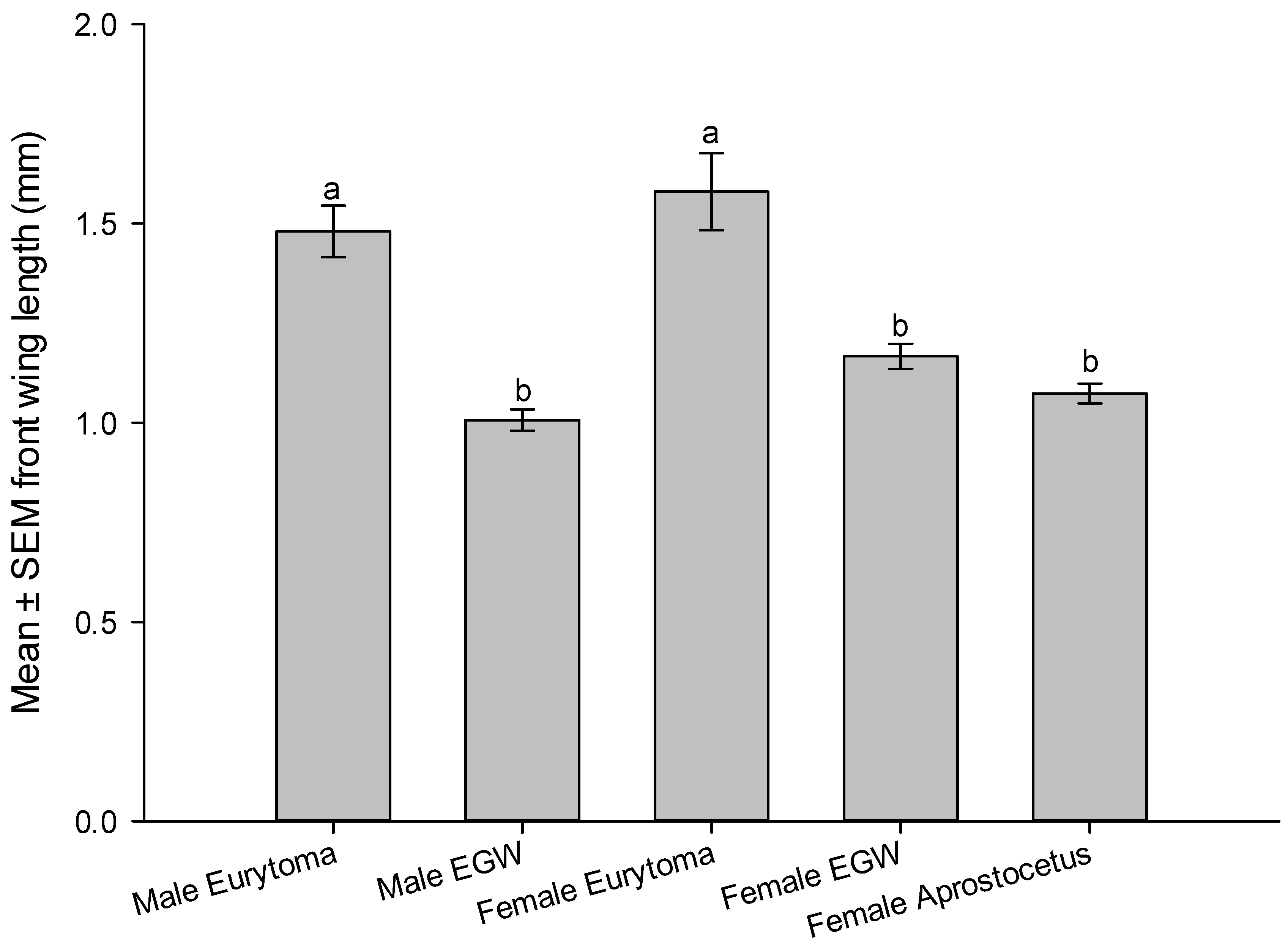
2.6. Longevity Study and Size of Wasps
2.7. Fecundity
2.8. Host Specificity Testing
2.9. Colonization Records on the Islands
2.10. Statistical Analysis and Vouchers
3. Results
3.1. Life History
3.2. Longevity
3.3. Reproductive Attributes
3.4. Host Specificity Testing
3.5. Records of Field Releases
4. Discussion

5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, I.K.; Delvare, G.; LaSalle, J. A new species of Quadrastichus (Hymenoptera: Eulophidae): A gall-inducing pest on Erythrina spp. (Fabaceae). J. Hymenopt. Res. 2004, 13, 243–249. [Google Scholar]
- Yang, M.; Tung, G.; LaSalle, J.; Wu, M. Outbreak of erythrina gall wasp on Erythrina spp. (Fabaceae) in Taiwan. Plant Prot. Bull. 2004, 46, 391–396. [Google Scholar]
- Heu, R.; Tsuda, D.; Nagamine, W.; Yalemar, J.; Suh, T. Erythrina Gall Wasp, Quadrastichus erythrinae Kim (Hymenoptera: Eulophidae). Hawaii Department of Agriculture, New Pest Advisory, 2006, No. 05-03. Available online: http://www.hawaiiag.org/hdoa/npa/npa05-03-EGW.pdf (accessed on 26 October 2007).
- Nami, U.; Takumi, U.; Yukawa, J. Detection of an invasive gall-inducing pest, Quadrastichus erythrinae (Hymenoptera: Eulophidae), causing damage to Erythrina variegata L. (Fabaceae) in Okinawa Prefecture, Japan. Entomol. Sci. 2007, 10, 209–212. [Google Scholar]
- Rubinoff, D.; Holland, B.S.; Shibata, A.; Messing, R.H.; Wright, M.G. Rapid invasion despite lack of genetic variation in the Erythrina gall wasp (Quadrastichus erythrinae Kim). Pac. Sci. 2010, 64, 23–31. [Google Scholar] [CrossRef]
- Wiley, J.; Skelley, P. Erythrina gall wasp, Quadrastichus erythrinae Kim, in Florida. Pest Alert, Florida Department of Agriculture and Consumer Services. 2006. Available online: http://www.doacs.state.fl.us/pi/enpp/ento/gallwasp.html (accessed on 2 November 2023).
- Palacios-Torres, R.E.; Malpica-Pita, J.; Bustamante-Ortiz, A.G.; Valdez-Carrasco, J.; Santos-Chávez, A.; Vega-Muñoz, R.; Vibrans-Lindemann, H. The Invasive Gall Wasp Quadrastichus erythrinae Kim in Mexico. Southwest. Entomol. 2017, 42, 1099–1102. [Google Scholar] [CrossRef]
- Medianero, E.; Zachrisson, B. Erythrina gall wasp, Quadrastichus erythrinae Kim, 2004 (Hymenoptera: Eulophidae: Tetrastichinae): A new pest in Central America. BioInvasions Rec. 2019, 8, 452–456. [Google Scholar] [CrossRef]
- Wagner, W.L.; Herbst, D.R.; Sohmer, S.H. Erythrina. In Manual of the Flowering Plants of Hawaii; University of Hawaii Press: Honolulu, HI, USA; Bishop Museum Press: Honolulu, HI, USA, 1990; Volume 2, pp. 671–672. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2023. Published on the Internet. Available online: http://www.plantsoftheworldonline.org/ (accessed on 22 October 2023).
- Neal, M.C. Gardens of Hawaii; Bishop Museum Press: Honolulu, HI, USA, 1965; p. 924. [Google Scholar]
- Hollier, D. The Seed Savers. Hana Hou! Mag. Hawaii. Airl. 2007, 10, 74–84. [Google Scholar]
- Dingeman, R. Victory at hand in war against invasive, tree-killing gall wasp. The Honolulu Advertiser, 26 March 2010. [Google Scholar]
- Rotar, P.P.; Joy, R.J.; Weissich, P.R. “Tropic Coral” tall erythrina. Univ. Haw. CTAHR Res. 1986, 72, 1–10. [Google Scholar]
- Messing, R.H.; Noser, S.; Hunkeler, J. Using host plant relationships to help determine origins of the invasive Erythrina gall wasp Quadrastichus erythrinae Kim (Hymenoptera: Eulophidae. Biol. Invasions 2009, 11, 2233–2241. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Z.; Yu, D.; Kang, L.; Yang, W.; Chen, Z.; Chen, X. Bionomics of the erythrina gall wasp, Quadristichus erythrinae Kim (Hymenoptera: Eulophidae). Acta Entomol. Sin. 2007, 50, 46–50. [Google Scholar]
- Doccola, J.; Smith, S.; Strom, B.; Medeiros, A.; von Allmen, E. Systemically Applied Insecticides for Treatment of Erythrina Gall Wasp, Quadrastichus erythrinae Kim (Hymenoptera: Eulophidae). Arboric. Urban For. 2009, 35, 173–181. [Google Scholar] [CrossRef]
- Xu, T.; Jacobsen, C.M.; Hara, A.H.; Li, J.; Li, Q.X. Efficacy of systemic insecticides on the gall wasp Quadrastichus erythrinae in wiliwili trees (Erythrina spp.). Pest Manag. Sci. 2009, 65, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Brannon, J. Dying Trees Cost $1M a Year. The Honolulu Advertiser. 2007. Available online: http://the.honoluluadvertiser.com/article/2007/Mar/23/ln/FP703230359.html (accessed on 21 October 2023).
- Gates, M.; Delvare, G. A new species of Eurytoma (Hymenoptera: Eurytomidae) attacking Quadrastichus spp. (Hymenoptera: Eulophidae) galling Erythrina spp. (Fabaceae), with a summary of African Eurytoma biology and species checklist. Zootaxa 2008, 1751, 1–24. [Google Scholar] [CrossRef]
- Ohl, M.; Thiele, K. Estimating body size in apoid wasps: The significance of linear variables in a morphologically diverse taxon (Hymenoptera, Apoidea). Zoosyst. Evol. 2007, 83, 110–124. [Google Scholar] [CrossRef]
- Yeager, J.F. Stimulation of isolated heart roach. J. Agric. Res. 1939, 59, 121–137. [Google Scholar]
- Nishida, G.M. Hawaiian Terrestrial Arthropod Checklist. Fourth Edition. Bish. Mus. Tech. Bull. 2002, 22, 313. [Google Scholar]
- Perkins, R.C.L.; Forel, A. Fauna Hawaiiensis. In Hymenoptera Aculeata; Sharp, D., Ed.; Cambridge University Press: London, UK, 1899; Volume 1, Part 1. [Google Scholar]
- Yoshimoto, C.M. Synopsis of Hawaiian Eulophidae including Aphelininae (Hym.: Chalcidoidea). Pac. Insects 1965, 7, 665–699. [Google Scholar]
- Boucek, Z. Australasian Chalcidoidea (Hymenoptera): A Biosystematic Revision of Genera of Fourteen Families, with Reclassification of Species; CAB International: Wallingford, UK, 1988; p. 832. [Google Scholar]
- JMP®, Version 11; 2023; SAS Institute Inc.: Cary, NC, USA, 1989–2023.
- Lotfalizadeh, H.; Delvare, G.; Rasplus, J.-Y. Phylogenetic analysis of Eurytominae (Chalcidoidea: Eurytomidae) based on morphological characters. Zool. J. Linn. Soc. 2007, 151, 441–510. [Google Scholar] [CrossRef]
- Noyes, J.S. Universal Chalcidoidea Database. World Wide Web Electronic Publication. 2020. Available online: https://www.nhm.ac.uk/chalcidoids (accessed on 10 October 2020).
- Gómez, J.F.; Nieves-Aldrey, J.L.; Nieves, M.H.; Stone, G.N. Comparative morphology and biology of terminal-instar larvae of some Eurytoma (Hymenoptera, Eurytomidae) species parasitoids of gall wasps (Hymenoptera, Cynipidae) in western Europe. Zoosystema 2011, 33, 287–323. [Google Scholar] [CrossRef]
- Vårdal, H.; Gómez, J.F.; Nieves-Aldrey, J.L. Ovarian egg morphology in chalcidoid wasps (Hymenoptera: Chalcidoidea) parasitizing gall wasps (Hymenoptera: Cynipidae). Graellsia 2016, 72, e044. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Rosenheim, J.A.; Kattari, D. Adult feeding and lifetime reproductive success in the parasitoid Aphytis melinus. Entomol. Exp. Appl. 1997, 83, 305–315. [Google Scholar] [CrossRef]
- Boulton, R.A.; Shuker, D.M. A sex allocation cost to polyandry in a parasitoid wasp. Biol. Lett. 2015, 11, 20150205. [Google Scholar] [CrossRef] [PubMed]
- Augustin, J.; Bourgeois, G.; Brodeur, J.; Boivin, G. Low and high temperatures decrease the mating success of an egg parasitoid and the proportion of females in the population. J. Therm. Biol. 2022, 110, 103382. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.M.; Wang, X. Male Impact on Female Reproductive Performance of the Larval Tephritid Parasitoid Diachasmimorpha tryoni (Hymenoptera: Braconidae). J. Econ. Entomol. 2021, 114, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Prinsloo, G.L.; Kelly, J.A. The tetrastichine wasps (Hymenoptera: Chalcidoidea: Eulophidae) associated with galls on Erythrina species (Fabaceae) in South Africa, with the description of five new species. Zootaxa 2009, 2083, 27–45. [Google Scholar] [CrossRef]
- Lin, S.F.; Tung, G.S.; Yang, M.M. The Erythrina Gall Wasp Quadrastichus erythrinae (Insecta: Hymenoptera: Eulophidae): Invasion History, Ecology, Infestation and Management. Forests 2021, 12, 948. [Google Scholar] [CrossRef]
- Lin, S.-F.; Tung, G.-S.; Yang, M.-M. Out of Africa: Origin of the Erythrina Gall Wasp Quadrastichus erythrinae (Hymenoptera: Chalcidoidea: Eulophidae). Formos. Entomol. 2021, 41, 26–36. [Google Scholar] [CrossRef]
- Day, M.D.; Cock, M.J.W.; Conant, P.; Cooke, B.; Furlong, M.J.; Paynter, Q.; Ramadan, M.M.; Wright, M.G. Biological control successes and failures: Oceania region. In Biological Control: Global Impacts, Challenges and Future Directions of Pest Management; Mason, P., Ed.; CSIRO Publishing: Clayton, Australia, 2021; pp. 342–376. [Google Scholar]
- Ramadan, M.M.; Kaufman, L.V.; Wright, M.G. Insect and weed biological control in Hawaii: Recent case studies and trends. Biol. Control 2023, 179, 105170. [Google Scholar] [CrossRef]
- Kaufman, L.V.; Yalemar, J.; Wright, M.G. Classical biological control of the erythrina gall wasp, Quadrastichus erythrinae, in Hawaii: Conserving an endangered habitat. Biol. Control 2020, 142, 104161. [Google Scholar] [CrossRef]
- Bell, R.C.; Belmaker, A.; Couch, C.S.; Marchetto, K.M.; Simonis, J.L.; Thomas, Q.; Sparks, J. Effectiveness of Erythrina gall wasp biocontrol and implications for the recovery of threatened Wiliwili trees (Fabaceae: Erythrina sandwicensis). J. Torrey Bot. Soc. 2013, 140, 215–224. [Google Scholar] [CrossRef]
- R-5 and Southern Research Station U.S. Department of Agriculture Forest Service Forest Health Protection. Early Detection Pest Advisory 2007: Identifying and managing the Erythrina Gall Wasp. Science Update SRS-012. United States Department of Agriculture, Southern Research Station. Brochure. 2007. Available online: https://www.fs.usda.gov/research/treesearch/27989 (accessed on 25 October 2023).
- LaSalle, J.; Ramadan, M.; Kumashiro, B.R. A new parasitoid of the Erythrina Gall Wasp, Quadrastichus erythrinae Kim (Hymenoptera: Eulophidae). Zootaxa 2009, 2083, 19–26. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order and Family | Gall-Former (Scientific and Common Name) | Gall-Former Status and Source | Host Plant (Scientific, Common Name), Infested Plant Part Used for Testing |
|---|---|---|---|
| Hymenoptera: Agaonidae | Josephiella microcarpae Beardsley & Rasplus, Banyan gall wasp | Immigrant, Field-collected, Honolulu, Oahu | Ficus microcarpa Chinese banyan, cuttings |
| Hemiptera: Eriococcidae | Tectococcus ovatus Hempel biocontrol agent | Biocontrol agent, Lab-reared, USFS, HVNP and HDOA | Psidium cattleianum strawberry guava, whole plants |
| Hymenoptera: Eulophidae | Ophelimus sp. Eucalyptus gall wasp | Immigrant, Field-collected, Camp Maluhia, Maui Island | Eucalyptus sp. Eucalyptus, Cuttings |
| Hemiptera: Psyllidae | Trioza sp. Ohia psyllid | Endemic, Field-collected, Aiea and Manoa, Oahu Island | Metrosideros polymorpha Ohia, Cuttings |
| Diptera: Tephritidae | Eutreta xanthochaeta Aldrich Lantana gall fly | Biocontrol agent, Field-collected, Hauula, Oahu Island and lab-reared HDOA | Lantana camara Lantana, whole plants |
| Diptera: Tephritidae | Procecidochares alani Steyskal Hamakua pamakani gall fly | Biocontrol agent, Field-collected, Nuuanu, Oahu Island and lab-reared HDOA | Ageratina riparia Hamakua pamakani, whole plants |
| Diptera: Tephritidae | Procecidochares utilis Stone Maui pamakani gall fly | Biocontrol agent, Lab-reared, UH-Manoa and lab-reared HDOA | Ageratina adenophora Maui pamakani, whole plants |
| Parasitoid and Host | N | Longevity (Days) Mean ± SEM | Body Length a (mm) Mean ± SEM | |
|---|---|---|---|---|
| Laboratory Colony | Wild Wasps b | |||
| Female Eurytoma | 39 | 40.4 ± 2.2 | 2.1 ± 0.05 | 2.2 ± 0.08 a |
| Male Eurytoma | 92 | 20.5 ± 1.1 | 1.5 ± 0.04 | 2.04 ± 0.07 a |
| Mean comparison | T58 = 8.12, p = 0.0001 | T128 = 9.20, p = 0.0001 | ||
| Female EGW | 100 | 5.9 ± 0.3 | - | 1.6 ± 0.03 b |
| Male EGW | 100 | 7.8 ± 0.3 | - | 1.45 ± 0.07 b |
| Mean comparisons | T197 = 4.106, p = 0.0001 | - | F3,44 = 25.55, p < 0.0001 | |
| Reproductive Parameter | Mean ± SEM | Range | Unit |
|---|---|---|---|
| Oviposition period a | 37.7 ± 6.7 | 31–51 | days |
| Post-oviposition period | 13.7 ± 5.4 | 3–20 | days |
| Mature ovarian eggs at death | 2.3 ± 1.5 | 0–5 | number eggs |
| Female longevity | 51.3 ± 1.5 | 49–54 | days |
| Male longevity (n = 15) b | 24.6 ± 2.7 | 3–42 | days |
| Female progeny | 40.0 ± 15.3 | 20–70 | adult |
| Male progeny | 125.3 ± 41.8 | 82–209 | adult |
| Total progeny per female | 165.3 ± 39.3 | 105–239 | offspring |
| Daily progeny | 4.3 ± 0.5 | 3.4–4.9 | adult |
| Sex ratio (% females) | 25.9 ± 10.3 | 12.6–46.1 | % female |
| Sex ratio (% males) | 74.1 ± 10.3 | 53.9–87.4 | % male |
| Female lifecycle (n = 120) | 18.4 ± 0.1 | 15–24 | days |
| Male lifecycle (n = 376) | 18.2 ± 0.1 | 15–26 | days |
| Galled Plant Infested with Insect | Frequency of Visits by E. erythrinae (Mean ± SEM) | E. erythrinae Adult Emergence (Mean ± SEM) |
|---|---|---|
| Josephiella microcarpae | 0 b | 0 b |
| Q. erythrinae (control) | 7.6 ± 2.6 a | 25.0 ± 9.6 a |
| Tectococcus ovatus | 0.2 ± 0.2 b | 0 b |
| Q. erythrinae (control) | 10.6 ± 3.4 a | 62.2 ± 17.9 a |
| Ophelimus sp. | 1.0 ± 1.0 b | 0 b |
| Q. erythrinae (control) | 7.2 ± 1.4 a | 19.8 ± 3.1 a |
| Trioza sp. | 0 b | 0 b |
| Q. erythrinae (control) | 7.6 ± 1.2 a | 25.4 ± 9.8 a |
| Eutreta xanthochaeta | 0.8 ± 0.3 b | 0 b |
| Q. erythrinae (control) | 6.0 ± 0.7 a | 14.8 ± 5.8 a |
| Procecidochares alani | 0 b | 0 b |
| Q. erythrinae (control) | 7.0 ± 1.8 a | 14.5 ± 5.4 a |
| Procecidochares utilis | 0.2 ± 0.2 b | 0 b |
| Q. erythrinae (control) | 9.8 ± 3.2 a | 17.0 ± 6.0 a |
| Island | Erythrina Species Infested by the Gall Wasp | N 2 | Numbers of Eurytoma Released | Release Time (Month/Day/Year) | Dates of Recovery (Month/Day/Year) |
|---|---|---|---|---|---|
| Oahu | Erythrina sandwicensis O.Deg. | 1070 | 25 November 2008–23 August 2010 | 6 February 2009–23 August 2010 3 | |
| Erythrina crista-galli L. | 470 | ||||
| Erythrina variegata L. 1 | 130 | ||||
| Sub-total and % of total | 38 | 1670 (41.8%) | |||
| Maui | Erythrina sandwicensis | 380 | 17 December 2008–23 December 2009 | 25 February 2009–21 January 2010 | |
| Erythrina crista-galli | 60 | ||||
| Sub-total % of total | 10 | 440 (11.0%) | |||
| Hawaii | Erythrina sandwicensis. | 688 | 1 December 2008–27 January 2010 | 14 January 2009–27 January 2010 3 | |
| Erythrina crista-galli | 30 | ||||
| Erythrina variegata 1 | 330 | ||||
| Sub-total and % of total | 20 | 1048 (26.2%) | |||
| Kauai | Erythrina sandwicensis | 240 | 4 December 2008 | 3 March 2009–30 June 2010 | |
| Erythrina variegata | 30 | ||||
| Sub-total and % of total | 5 | 270 (6.7%) | |||
| Molokai | Erythrina sandwicensis | 390 | 14 April 2009–9 June 2009 | 10 February 2010–24 March 2010 | |
| Erythrina crista-galli | 60 | ||||
| Erythrina variegata 1 | 30 | ||||
| Sub-total and % of total | 9 | 480 (12.0%) | |||
| Lanai total and % of total | Erythrina sandwicensis | 2 | 90 (2.2%) | 12 May 2009 | 17 February 2010–7 April 2010 |
| Total released on all islands | 84 | 3998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagamine, W.T.; Yalemar, J.A.; Wright, M.G.; Ramadan, M.M. Reproductive Parameters and Host Specificity of Eurytoma erythrinae (Hymenoptera: Eurytomidae), a Biological Control Agent of the Erythrina Gall Wasp, Quadrastichus erythrinae (Hymenoptera: Eulophidae). Insects 2023, 14, 923. https://doi.org/10.3390/insects14120923
Nagamine WT, Yalemar JA, Wright MG, Ramadan MM. Reproductive Parameters and Host Specificity of Eurytoma erythrinae (Hymenoptera: Eurytomidae), a Biological Control Agent of the Erythrina Gall Wasp, Quadrastichus erythrinae (Hymenoptera: Eulophidae). Insects. 2023; 14(12):923. https://doi.org/10.3390/insects14120923
Chicago/Turabian StyleNagamine, Walter T., Juliana A. Yalemar, Mark G. Wright, and Mohsen M. Ramadan. 2023. "Reproductive Parameters and Host Specificity of Eurytoma erythrinae (Hymenoptera: Eurytomidae), a Biological Control Agent of the Erythrina Gall Wasp, Quadrastichus erythrinae (Hymenoptera: Eulophidae)" Insects 14, no. 12: 923. https://doi.org/10.3390/insects14120923