
The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines. Part I. Isolation, Identification and In Vitro Studies

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation
2.2. Identification of Endophytic Trichoderma Species
2.3. Mycelial Growth
2.4. Biocontrol Index of Different Trichoderma Species
2.5. Fungicide Tolerance Test
3. Results
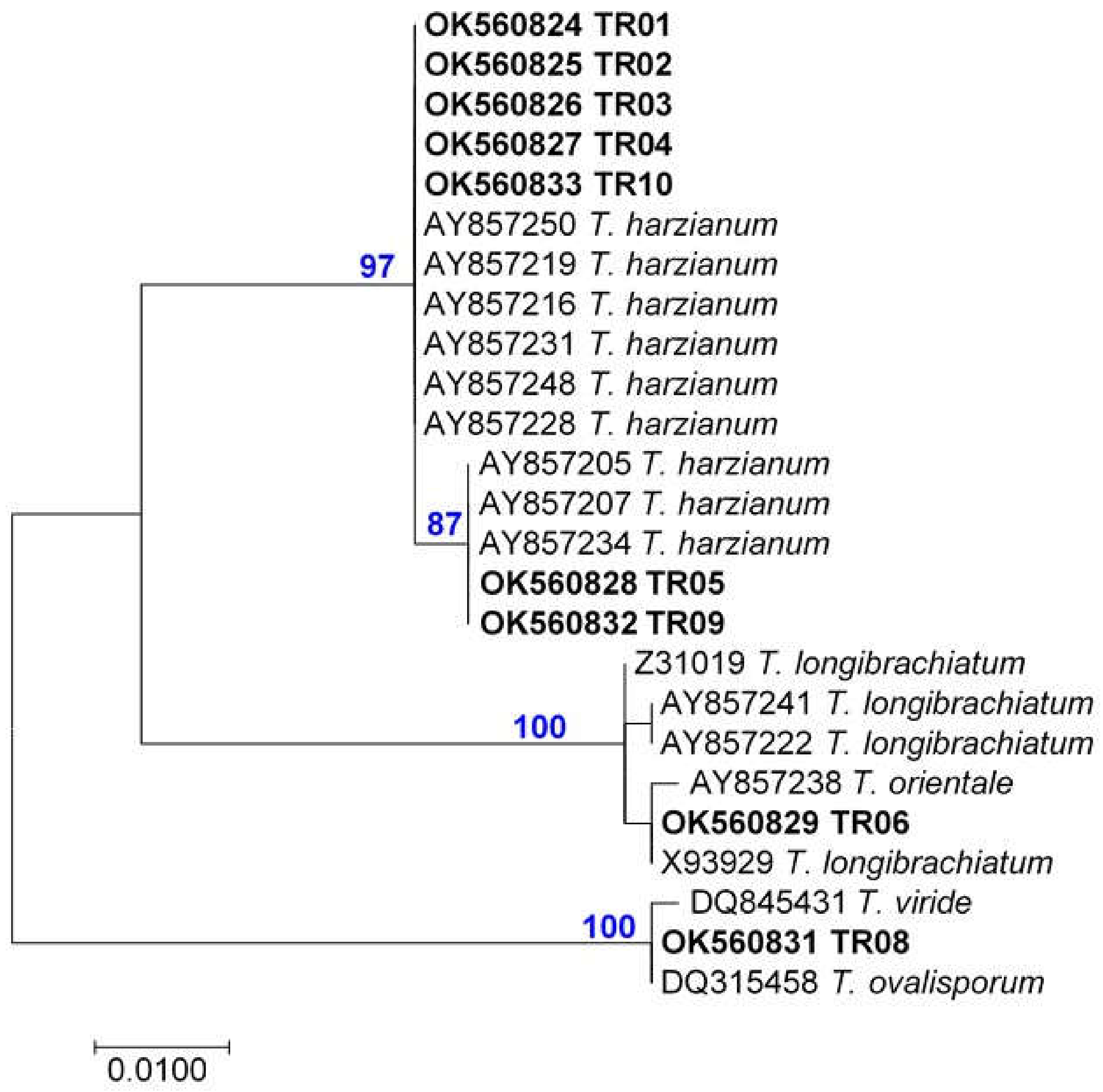
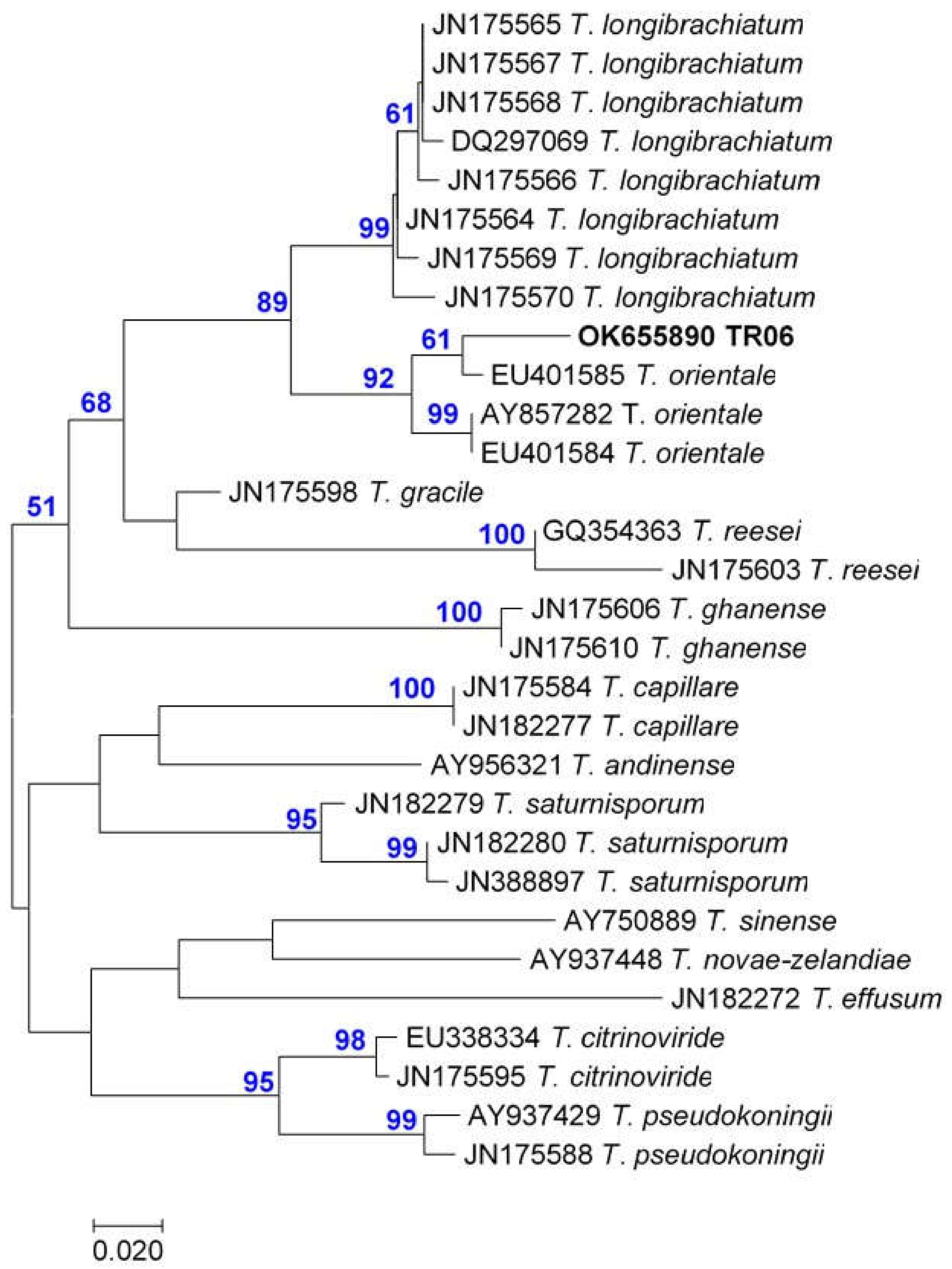
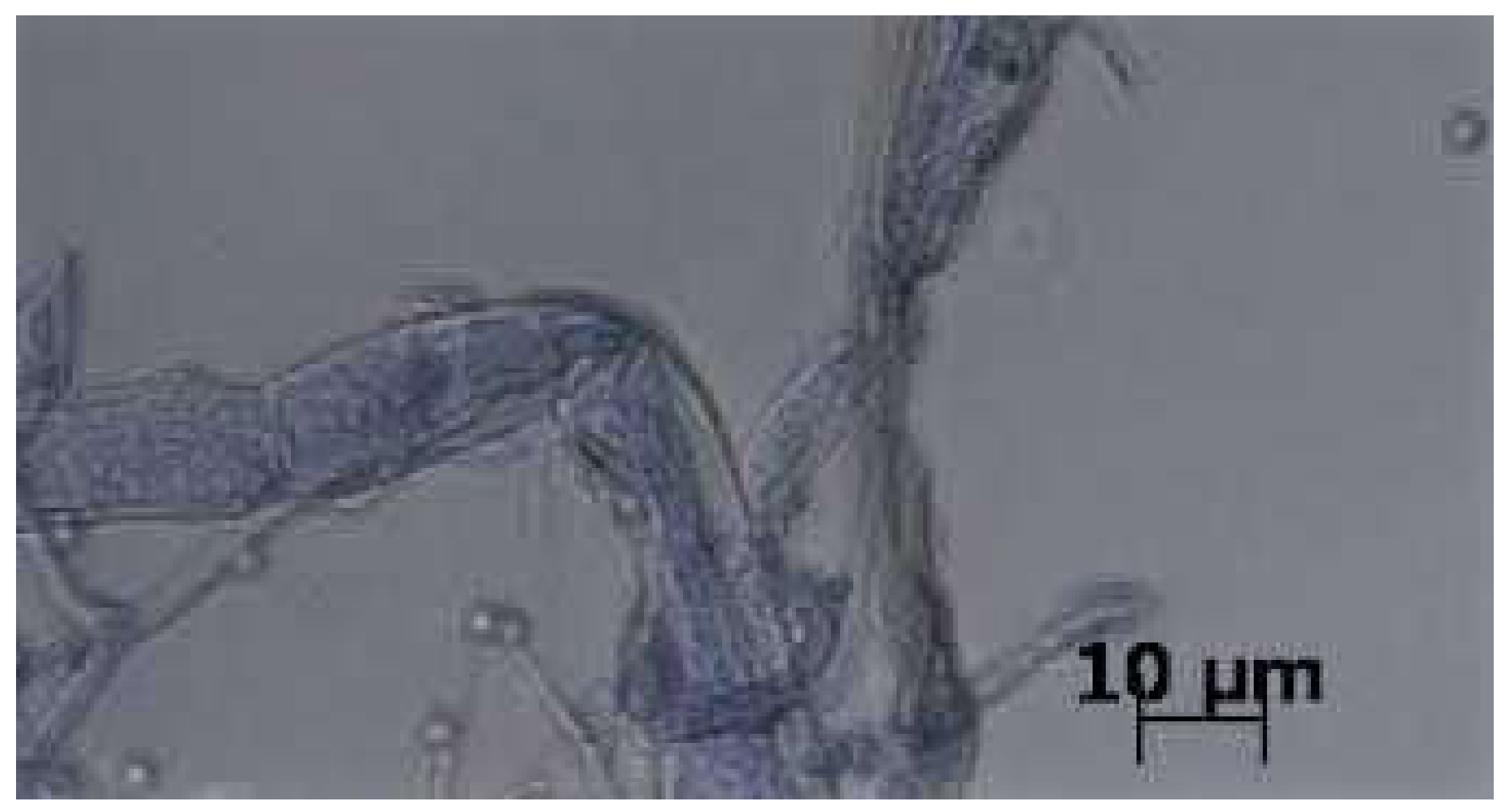
3.1. Strain Identification
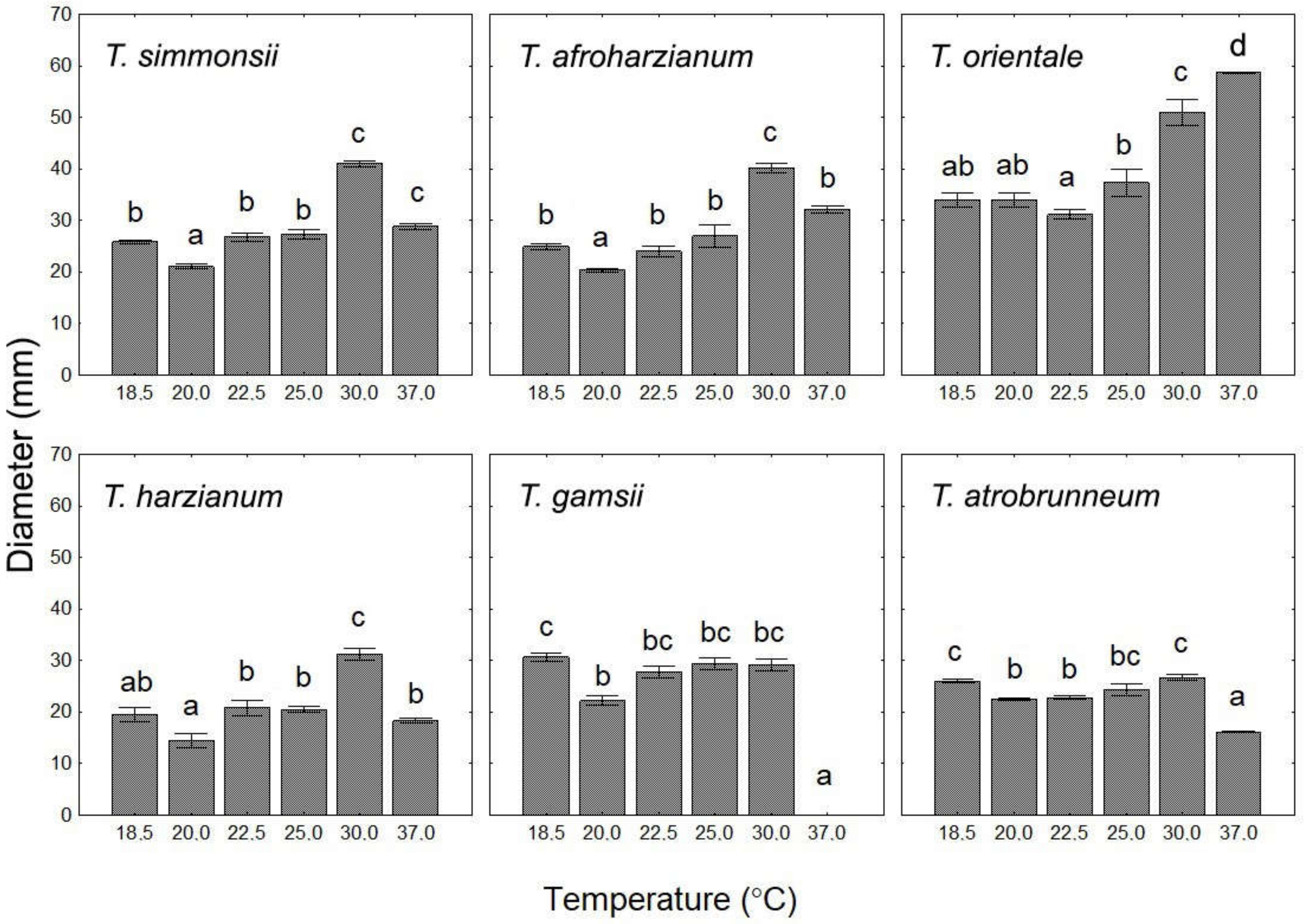
3.2. Growth Characteristics at Different Temperatures
3.3. Potential for Biocontrol
3.4. Pesticide Tolerance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaki, O.; Weekersb, F.; Thonarta, P.; Teschc, E.; Kuenemannc, P.; Jacquesa, P. Limiting factors of mycopesticide development. Biol. Control 2020, 144, 104220. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Chenthamara, K.; Zhang, J.; Atanasova, L.; Yang, D.; Miao, Y.; Rahimi, M.J.; Grujic, M.; Cai, F.; Pourmehdi, S.; et al. Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 2018, 14, e1007322. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.K.; Norris, R.V. A Biochemical study of some soil fungi with special reference to ammonia production. J. Indian Inst. Sci. 1928, 11, 141. [Google Scholar]
- Tjamos, E.C.; Papavizas, G.C.; Cook, R.J. Biological Control of Plant Diseases: Progress and Challenges for the Future; Plenum Press: New York, NY, USA, 1922; p. 222. [Google Scholar]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Druzhinina, I.S.; Komon-Zelazowska, M.; Kredics, L.; Hatvani, L.; Antal, Z.; Belayneh, T.; Kubicek, C.P. Alternative reproductive strategies of Hypocrea orientalis and genetically close but clonal Trichoderma longibrachiatum, both capable of causing invasive mycoses of humans. Microbiology 2008, 154, 3447–3459. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.; Howell, C.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Mulatu, A.; Alemu, T.; Megersa, N.; Vetukuri, R.R. Optimization of Culture Conditions and Production of Bio-Fungicides from Trichoderma Species under Solid-State Fermentation Using Mathematical Modeling. Microorganisms 2021, 9, 1675. [Google Scholar] [CrossRef]
- Druzhinina, I.; Kubicek, C.P. Species Concepts and Biodiversity in Trichoderma and Hypocrea: From Aggregate Species to Species Clusters? J. Zhejiang Univ. Sci. B. 2005, 6, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J. Trichoderma: Systematics, the sexual state, and ecology. Phytopathology 2006, 96, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Waghunde, R.R.; Shelake, R.M.; Sabalpara, A.N. Trichoderma: A significant fungus for agriculture and environment. Afr. J. Agric. Res. 2016, 11, 1952–1965. [Google Scholar]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: Authoritative guidelines on molecular identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Verma, M.; Brara, S.K.; Tyagia, R.D.; Surampalli, R.Y.; Valéroa, J.R. Antagonistic fungi, Trichoderma spp.: Panoply of biological control. Biochem. Eng. J. 2007, 37, 1–20. [Google Scholar] [CrossRef]
- Carro-Huerga, G.; Compant, S.; Gorfer, M.; Cardoza, R.E.; Schmoll, M.; Gutiérrez, S.; Casquero, P.A. Colonization of Vitis vinifera L. by the Endophyte Trichoderma sp. Strain T154: Biocontrol Activity Against Phaeoacremonium minimum. Front. Plant Sci. 2020, 11, 1170. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Tomaselli, E.; Pollard-Flamand, J.; Boulé, J.; Gerin, D.; Pollastro, S. Characterization of Trichoderma isolates from southern Italy, and their potential biocontrol activity against grapevine trunk disease fungi. Phytopathol. Mediterr. 2020, 59, 425–439. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a Newly Identified Endophytic Fungus, Trichoderma phayaoense for Plant Growth Promotion and Biological Control of Gummy Stem Blight and Wilt of Muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef]
- Silva-Valderrama, I.; Toapanta, D.; Miccono, M.A.; Lolas, M.; Díaz, G.A.; Cantu, D.; Castro, A. Biocontrol potential of grapevine endophytic and rhizospheric fungi against trunk pathogens. Front. Microbiol. 2021, 11, 614620. [Google Scholar] [CrossRef]
- Pesticide Info Database. Available online: https://www.pesticideinfo.org/ (accessed on 7 February 2021).
- BPDB: Bio-Pesticides DataBase. Available online: https://sitem.herts.ac.uk/aeru/bpdb (accessed on 7 February 2021).
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- NÉBIH (2009): 2009-ben Kiadott Növényvédő Szer Forgalomba Hozatali és Felhasználási Engedélyek. Available online: https://portal.nebih.gov.hu/-/novenyvedo-szer-okiratok-tara-2009 (accessed on 7 February 2021).
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [Green Version]
- Vargas, W.A.; Mandawe, J.C.; Kenerley, C.M. Plant-derived sucrose is a key element in the symbiotic association between Trichoderma virens and maize plants. Plant Physiol. 2009, 151, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, S.; Lin, F.; Kubicek, C.P.; Druzhinina, I.S. Trichoderma taxi sp. nov., an endophytic fungus from Chinese yew Taxus mairei. FEMS Microbiol. Lett. 2007, 270, 90–96. [Google Scholar] [CrossRef]
- Hanada, R.E.; de Jorge Souza, T.; Pomella, A.W.V.; Hebbar, K.P.; Pereira, J.O.; Ismaiel, A.; Samuels, G.J. Trichoderma martiale sp. nov., a new endophyte from sapwood of Theobroma cacao with a potential for biological control. Mycol. Res. 2008, 112, 1335–1343. [Google Scholar] [CrossRef]
- Mulaw, T.G.; Kubicek, C.P.; Druzhinina, I.S. The rhizosphere of Coffea arabica in its native highland forests of Ethiopia provides a niche for a distinguished diversity of Trichoderma. Diversity 2010, 2, 527–549. [Google Scholar] [CrossRef]
- Casieri, L.; Hofstetter, V.; Viret, O.; Gindro, K. Fungal communities living in the wood of different cultivars of young Vitis vinifera plants. Phytopathol. Mediterr. 2009, 48, 73–83. [Google Scholar]
- González, V.; Tello, M.L. The endophytic mycota associated with Vitis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.H.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Kovács, C.S.; Balling, P.; Bihari, Z.; Nagy, A.; Sándor, E. Incidence of grapevine trunk diseases is influenced by soil, topology and vineyard age, but not by Diplodia seriata infection rate in the Tokaj Wine Region, Hungary. Phytoparasitica 2017, 45, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Kulling, C.; Szakacs, G.; Kubicek, C.P. Molecular identification of Trichoderma species from Russia, Siberia and the Himalaya. Mycol. Res. 2000, 104, 1117–1125. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Váczy, K.Z.; Sándor, E.; Karaffa, L.; Fekete, E.; Fekete, É.; Árnyasi, M.; Czeglédi, L.; Kövics Gy, J.; Druzhinina, I.S.; Kubicek, C.P. Sexual Recombination in the Botrytis cinerea populations in Hungarian vineyards. Phytopathology 2008, 98, 1312–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Higgins, D.G.; Sharp, P.M. CLUSTAL: A package for performing multiple sequence alignment on a microcomputer. Gene 1988, 73, 237–244. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL-X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, K.B.; Nicholas, H.B.; Deerfield, D.W. GeneDoc: Analysis and visualization of genetic variation. EMBnet News 1997, 4, 14. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, A.; Leitgeb, B.; Kredics, L.; Manczinger, L.; Vágvölgyi, C. A novel, image analysis-based method for the evaluation of in vitro antagonism. J. Microbiol. Methods 2006, 65, 619–622. [Google Scholar] [CrossRef]
- Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions: The European Green Deal; COM(2019) 640 Final; European Commission: Brussels, Belgium, 2019; Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52019DC0640&print=true (accessed on 7 February 2021).
- Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions: A Farm to Fork Strategy; COM(2020) 381 Final; European Commission: Brussels, Belgium, 2020; Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52020DC0381&print=true (accessed on 7 February 2021).
- Leahy, J.; Mendelsohn, M.; Kough, J.; Jones, R.; Berckes, N. Biopesticide Oversight and Registration at the U.S. Environmental Protection Agency; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2014; pp. 3–18. [Google Scholar] [CrossRef]
- OECD. Annex to the Working Document on the Risk Assessment of Secondary Metabolites of Microbial Biocontrol Agents; ENV/JM/MONO(2018)33/ANN1; Series on Pesticides No. 98; OECD: Paris, France, 2018; Available online: https://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2018)33/ann1&doclanguage=en (accessed on 7 February 2021).
- Viterbo, A.; Inbar, J.; Hadar, Y.; Chet, I. Plant Disease Biocontrol and Induced Resistance via Fungal Mycoparasites. In A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research, 2nd ed.; The Mycota; Kubicek, C.P., Druzhinina, I.S., Eds.; Springer: Berlin, Germany, 2007; pp. 127–146. [Google Scholar]
- Kredics, L.; Antal, Z.; Dóczi, I.; Manczinger, L.; Kevei, F.; Nagy, E. Clinical importance of the genus Trichoderma. A review. Acta Microbiol. Immunol. Hung. 2003, 50, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Longa, C.M.O.; Pertot, I.; Tosi, S. Ecophysiological requirements and survival of a Trichoderma atroviride isolate with biocontrol potential. J. Basic Microbiol. 1998, 48, 269–277. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Prieto, P.; Rincón, A.; Gómez Rodríguez, M.V.; Valderrama, R.; Barroso, J.B.; Mercado-Blanco, J. Fate of Trichoderma harzianum in the olive rhizosphere: Time course of the root colonization process and interaction with the fungal pathogen Verticillium dahliae. BioControl 2016, 61, 269–282. [Google Scholar] [CrossRef]
- Bailey, B.A.; Melnick, R.L. The Endophytic Trichoderma. In Trichoderma: Biology and Applications; Mukherjee, P.K., Horwitz, B.K., Singh, U.S., Mukherjee, M., Schmoll, M., Eds.; CAB International: Wallingford, UK, 2013; pp. 152–172. [Google Scholar] [CrossRef]
- Tseng, Y.-H.; Rouina, H.; Groten, K.; Rajani, P.; Furch, A.C.U.; Reichelt, M.; Baldwin, I.T.; Nataraja, K.N.; Uma Shaanker, R.; Oelmüller, R. An endophytic Trichoderma strain promotes growth of its hosts and defends against pathogen attack. Front. Plant Sci. 2020, 11, 573670. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Komon-Zelazowska, M.; Druzhinina, I.S. Fungal genus Hypocrea/Trichoderma: From barcodes to biodiversity. J. Zhejiang Univ. Sci. B. 2008, 9, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J.; Chaverri, P.; Farr, D.F.; McCray, E.B. Trichoderma Online, Systematic Mycology 10 and Microbiology Laboratory; ARS, USDA: Washington, DC, USA, 2011. Available online: https://nt.ars-grin.gov/fungaldatabases (accessed on 7 February 2021).
- Kuhls, K.; Lieckfeldt, E.; Börner, T.; Guého, E. Molecular re-identification of human pathogenic Trichoderma isolates as Trichoderma longibrachiatum and Trichoderma citrinoviride. Med. Mycol. 1999, 37, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Jaklitsch, W.; Samuels, G.; Dodd, S.; Lu, B.; Druzhinina, I.S. Hypocrea rufa/Trichoderma viride: A reassessment, and description of five closely related species with and without warted conidia. Stud. Mycol. 2006, 56, 135–177. [Google Scholar] [CrossRef] [Green Version]
- Marik, T.; Tyagi, C.; Racić, G.; Rakk, D.; Szekeres, A.; Vágvölgyi, C.S.; Kredics, L. New 19-residue Peptaibols from Trichoderma Clade Viride. Microorganisms 2018, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J.; Hebbar, P.K. Trichoderma: Identification and Agricultural Properties; APS Press: St. Paul, MN, USA, 2015; p. 204. [Google Scholar]
- Chaverri, P.; Samuels, G.J. Evolution of habitat preference and nutrition mode in a cosmopolitan fungal genus with evidence of inter kingdom host jumps and major shifts in ecology. Evolution 2013, 67, 2823–2837. [Google Scholar]
- Fungicide Resistance Action Committee. FRAC Code List 2021, Fungal Control Agents Sorted by Cross Resistance Pattern and Mode of Action. FRAC: Brussels, Belgium, 2021; Available online: https://www.frac.info/docs/default-source/publications/frac-code-list/frac-code-list-2021--final.pdf?sfvrsn=f7ec499a_2 (accessed on 7 December 2021).
- McLean, K.L.; Hunt, J.; Stewart, A. Compatibility of the biocontrol agent Trichoderma harzianum C52 with selected fungicides. N. Z. Plant Prot. 2001, 54, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Bhai, S.R.; Thomas, J. Compatibility of Trichoderma harzianum (Rifai.) with fungicides, insecticides and fertilizers. Indian Phytopathol. 2010, 63, 145–148. [Google Scholar]
- Madhavi, G.B.; Bhattiprolu, S.L.; Reddy, V.B. Compatibility of biocontrol agent Trichoderma viride with various pesticides. J. Hortl. Sci. 2011, 6, 71–73. [Google Scholar]
- Vasundara, P.; Rangaswamy, V.; Johnson, M. Compatibility studies with fungicides, insecticides and their combinations on Trichoderma viridae in invitro conditions International. Int. J. Sci. Eng. Res. 2015, 6, 310–316. [Google Scholar]
- Wedajo, B. Compatibility Studies of Fungicides with Combination of Trichoderma species under in vitro conditions. Virol. Mycol. 2015, 4, 149. [Google Scholar] [CrossRef] [Green Version]
- Khirallah, W.; Mouden, N.; Selmaoui, K.; Achbani, E.; Benkirane, R.; Touhami, A.O.; Douira, A. Compatibility of Trichoderma spp. with some fungicides under in vitro conditions. Int. J. Recent Sci. Res. 2016, 7, 9060–9067. [Google Scholar]
- Silva, M.A.F.; Moura, K.E.; Moura, K.E.; Salomão, D.; Patricio, F.R.A. Compatibility of Trichoderma isolates with pesticides used in lettuce crop. Summa Phytopathol. 2018, 44, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Tomer, A.; Singh, R.; Prasad, D. Compatibility Trichoderma harzianum with systemic and two non systemic fungicides of in vitro. Asian J. Crop Sci. 2018, 10, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Maurya, S.; Rai, D.; Dubey, S.; Pal, R.K. Compatibility of Trichoderma harzianum with different fungicides under in vitro. Int. J. Chem. Stud. 2020, 8, 2946–2952. [Google Scholar] [CrossRef]
- Caron, J.; Thibodeau, P.O.; Bélanger, R.R. Sélection d’isolats de Trichoderma comme agent de lute biologique contre la moisissure grise (Botrytis cinerea) dans la production de la tomate de serre. In Rapport de Recherche, Club D’encadrement Technique Pro-Serre; SPSQ: Québec, QC, Canada, 1994; p. 48. [Google Scholar]
- Figueras-Roca, M.; Cristani, C.; Vannacci, G. Sensitivity of Trichoderma isolates and selected resistant mutants to DMI fungicides. Crop Prot. 1996, 15, 615–620. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trichoderma Species | Strain | Deponated Sequences/Genome | Product | Producer/Distributor | Reference/Database Information |
|---|---|---|---|---|---|
| Harzianum clade of Trichoderma | |||||
| T. afroharzianum (formerly: T. harzianum Rifai) | T-22 (KRL-AG2, ATCC20847) | https://genome.jgi.doe.gov/portal/pages/projectStatus.jsf?db=TriharT22_1 (accessed on 7 December 2021) | T-22 WP BW240 G a (PlantShield® HC Biological Fungicide; T-22® HC Biological Fungicide; RootShield® Home and Garden Biological Fungicide; Root Guardian; RootShield® Seed Treatment; RootShield® Seed Treatment Biological Fungicide; RootShield® AG; RootShield® AG Biological Fungicide) | Bio Works Inc., Victor, NY, USA | http://www.pesticideinfo.org (accessed on 7 February 2021) https://www3.epa.gov/pesticides/chem_search/ppls/068539-00010-20190430.pdf (accessed on 7 February 2021) https://mycocosm.jgi.doe.gov/TriharT22_1/TriharT22_1.info.html (accessed on 7 February 2021) |
| T-22™ HC | http://www.pesticideinfo.org (accessed on 7 February 2021) | ||||
| Trianum G Biological Fungicide (Trianum Granules Biological Fungicide; Trianum G) | Koppert Biological Systems, Inc., Howell, MI, USA | http://www.pesticideinfo.org (accessed on 7 February 2021) https://www3.epa.gov/pesticides/chem_search/ppls/089635-00003-20151005.pdf (accessed on 7 February 2021) | |||
| Trianum WG Biological Fungicide (Trianum P Biological Fungicide; Trianum P; Trianum Granules Biological Fungicide; T-22 WG Water Dispersible Granules-Biological Fungicide) | http://www.pesticideinfo.org (accessed on 7 February 2021) https://www3.epa.gov/pesticides/chem_search/ppls/089635-00002-20150930.pdf (accessed on 7 February 2021) | ||||
| T. atrobrunneum (formerly: T. harzianum Rifai) | ITEM 908 | PNRQ10000000 | Trianum P | Koppert Biological Systems, Inc., Howell, MI, USA | https://sitem.herts.ac.uk/aeru/bpdb/Reports/2034.htm (accessed on 7 February 2021) |
| Virens clade of Trichoderma | |||||
| T. virens | G-41 | ATCC 20609 | BW240 G a (RootShield® Plus Granules, RootShield® Plus Granules Biological Fungicide, TurfShield® PLUS G, TurfShield® PLUS G Biological Fungicide, TurfShield® PLUS Granules, TurfShield® PL US Granules Biological Fungicide) | Bio Works Inc., Victor, NY, USA | http://www.pesticideinfo.org (accessed on 7 February 2021) https://www.atcc.org/products/20906 (accessed on 7 February 2021) |
| Viride clade of Trichoderma | |||||
| T. asperellum | T34 | EU077228 EU077227 | T34 Biocontrol (Asperello T34 BIOCONTROL) | Biobest Biocontrol Technologies, Barcelona, Spain | https://www.pesticideinfo.org/ https://www3.epa.gov/pesticides/chem_search/ppls/087301-00001-20200403.pdf (accessed on 7 December 2021) |
| Trifender Pro | Kwizda Agro Hungary Kft., Budapest, Hungary | https://sitem.herts.ac.uk/aeru/bpdb/Reports/2043.htm (accessed on 07 December 2021) https://kwizda.hu/AGRO_HU/products/t/Trifender%20Pro/Product%20Management/Registration%20Report/Trifender%20Pro%20Okirat.pdf (accessed on 7 December 2021) | |||
| ICC012 | GQ351595 GQ351596 | Bioten ™ WP b (Tenet™ WP and Remedier WP and Tenet™ T&O) | Isagro USA, Inc., Morrisville, NC, USA | https://sitem.herts.ac.uk/aeru/bpdb/Reports/2043.htm (accessed on 7 December 2021) https://www3.epa.gov/pesticides/chem_search/ppls/080289-00009-20110217.pdf (accessed on 7 December 2021) | |
| T. atroviride | I-1237 | Esquive WP | Agrauxine, Marcq en Baroeul, France | https://sitem.herts.ac.uk/aeru/bpdb/Reports/2046.htm (accessed on 7 December 2021) https://www.efsa.europa.eu/en/efsajournal/pub/2706 (accessed on 7 December 2021) | |
| Tri-Soil | Agrauxine, Marcq en Baroeul, France | https://ephy.anses.fr/ppp/tri-soil (accessed on 7 December 2021) https://sitem.herts.ac.uk/aeru/bpdb (accessed on 7 December 2021) | |||
| SC1 | HV500891-500896 | VINTEC® | Bi-PA nv, Londerzeel, Belgium | https://www.pesticideinfo.org/ https://sitem.herts.ac.uk/aeru/bpdb/Reports/2799.htm (accessed on 7 December 2021) https://www3.epa.gov/pesticides/chem_search/ppls/092083-00002-20200805.pdf (accessed on 7 December 2021) | |
| T. gamsii (formerly: T. viride) | ICC080 | GQ351598 | Bioten ™ WP b (Tenet™ WP and Remedier WP and Tenet™ T&O) | Isagro USA, Inc., Morrisville, NC, USA | http://www.pesticideinfo.org (accessed on 7 December 2021) https://www3.epa.gov/pesticides (accessed on 7 December 2021) |
| Amplified Region | Primer | Reference | Annealing Temperature (°C) |
|---|---|---|---|
| ITS1/ITS2 | SR6R | White et al. [36] | 50 |
| LR1 | |||
| tef1 | EF1 728F | Carbone and Kohn [37] | 59 |
| EF1 986R |
| Reference Number * | Fungal Pathogen | Host | Accession Number ** |
|---|---|---|---|
| CBS 477.71 | Aphanomyces cochlioides | Beta vulgaris L. | HQ665241 |
| JT2015 | Botryosphaeria dothidea | Juglans regia L. | MN706192 |
| J2034 | Diaporthe eres | Juglans regia L. | MT111103 |
| HUT01 | Diplodia seriata | Vitis vinifera L. | KU377167 |
| R.3 | Eutypa lata | Vitis vinifera L. | OK178559 |
| B.CS.5.4.20.1.B | Neofusicoccum parvum | Vitis vinifera L. | OK178560 |
| CBS 337.29 | Pythium acantophoron | Ananas sativus (L.) Merr. | HQ665212 |
| Target | Pesticide | Active Ingredient | Tested Concentration of the Pesticide (mg/L or mL/L) |
|---|---|---|---|
| Downy mildew | Orvego | Ametoctradin | 399 |
| Dimethomorph | 299.25 | ||
| Powdery mildew | Rally Q SC | Myclobutanil, | 45 |
| Quinoxifen | 45 | ||
| Sercadis | Fluxapyroxad | 225 | |
| Talentum 20 EW | Myclobutanil | 80 | |
| Grey mold | Chorus 50 WG | Cyprodinil | 469 |
| Teldor 500 SC | Fenhexamid | 835 |
| Trichoderma Species | Strain No. | NCBI GenBank Accession No. | |
|---|---|---|---|
| ITS a | tef1b | ||
| Harzianum clade | |||
| T. afroharzianum | TR04 | OK560827 | OK655888 |
| T. atrobrunneum | TR09 | OK560832 | OK655893 |
| T. harzianum | TR07 | OK560830 | OK655891 |
| TR10 | OK560833 | OK655894 | |
| T. simmonsii | TR01 | OK560824 | OK655885 |
| TR02 | OK560825 | OK655886 | |
| TR03 | OK560826 | OK655887 | |
| TR05 | OK560828 | OK655889 | |
| Longibrachiatum Clade | |||
| T. orientale | TR06 | OK560829 | OK655890 |
| Viride Clade | |||
| T. gamsii | TR08 | OK560831 | OK655892 |
| (a) | |||||||
| 192 h | 336 h | ||||||
| Strains | Species | Mean | SE | Mean | SE | ||
| Harzianum Clade | |||||||
| T. afroharzianum | 16.17 | 2.20 | ab | 34.00 | 6.00 | ab | |
| T. atrobrunneum | 17.00 | 1.44 | a | 27.50 | 1.61 | a | |
| T. harzianum | 7.83 | 0.88 | c | 17.58 | 1.35 | d | |
| T. simmonsii | 14.92 | 0.64 | ab | 28.79 | 0.85 | a | |
| Longibrachiatum Clade | |||||||
| T. orientale | 12.67 | 0.17 | b | 22.67 | 0.44 | b | |
| Viride Clade | |||||||
| T. gamsii | 8.00 | 1.15 | c | 22.17 | 0.60 | bc | |
| TR01 | T. simmonsii | 15.33 | 0.44 | a | 30.33 | 0.93 | a |
| TR02 | T. simmonsii | 17.83 | 1.09 | b | 31.17 | 0.60 | a |
| TR03 | T. simmonsii | 13.83 | 0.33 | a | 29.17 | 0.60 | a |
| TR05 | T. simmonsii | 12.67 | 0.17 | c | 24.50 | 1.04 | b |
| TR07 | T. harzianum | 9.50 | 0.76 | a | 19.67 | 0.83 | a |
| TR10 | T. harzianum | 6.17 | 0.73 | b | 15.50 | 2.02 | a |
| (b) | |||||||
| 24 h | 54 h | ||||||
| Strains | Species | Mean | SE | Mean | SE | ||
| Harzianum Clade | |||||||
| T. afroharzianum | 17.06 | 0.87 | b | 56.42 | 3.81 | ab | |
| T. atrobrunneum | 18.29 | 0.41 | b | 51.25 | 2.42 | a | |
| T. harzianum | 13.85 | 0.49 | a | 47.55 | 2.31 | a | |
| T. simmonsii | 18.72 | 0.39 | bc | 60.13 | 2.04 | bc | |
| Longibrachiatum Clade | |||||||
| T. orientale | 26.00 | 0.80 | d | 72.17 | 3.65 | d | |
| Viride Clade | |||||||
| T. gamsii | 20.50 | 0.83 | c | 66.04 | 3.41 | c | |
| TR01 | T. simmonsii | 17.92 | 1.58 | 0.00 | 56.25 | ||
| TR02 | T. simmonsii | 18.58 | 1.59 | 0.00 | 57.54 | ||
| TR03 | T. simmonsii | 20.42 | 3.29 | 0.00 | 59.46 | ||
| TR05 | T. simmonsii | 17.96 | 3.34 | 0.00 | 67.25 | ||
| TR07 | T. harzianum | 14.13 | 2.11 | 0.00 | 47.79 | ||
| TR10 | T. harzianum | 13.58 | 2.76 | 0.00 | 47.31 | ||
| (c) | |||||||
| 24 h | 30 h | ||||||
| Strains | Species | Mean | SE | Mean | SE | ||
| Harzianum Clade | |||||||
| T. afroharzianum | 21.50 | 0.29 | c | 32.17 | 0.67 | c | |
| T. atrobrunneum | 13.17 | 0.17 | b | 16.17 | 0.17 | b | |
| T. harzianum | 14.17 | 0.53 | b | 18.33 | 0.44 | b | |
| T. simmonsii | 20.63 | 0.43 | c | 28.83 | 0.55 | c | |
| Longibrachiatum Clade | |||||||
| T. orientale | 39.83 | 0.33 | d | 58.67 | 0.17 | d | |
| Viride Clade | |||||||
| T. gamsii | 0.00 | 0.00 | a | 0.00 | 0.00 | a | |
| TR01 | T. simmonsii | 19.00 | 0.76 | 28.67 | 0.73 | ||
| TR02 | T. simmonsii | 21.83 | 0.67 | 30.00 | 0.29 | ||
| TR03 | T. simmonsii | 21.17 | 0.88 | 27.50 | 2.00 | ||
| TR05 | T. simmonsii | 20.50 | 0.29 | 29.17 | 0.67 | ||
| TR07 | T. harzianum | 14.00 | 1.00 | 18.50 | 0.87 | ||
| TR10 | T. harzianum | 14.33 | 0.60 | 18.17 | 0.44 | ||
| BCI (%) | ||
|---|---|---|
| Fungal Pathogen | TR04 | TR05 |
| Oomycota | ||
| Aphanomyces cochlioides | 90.37 (0.64) | 84.81 (0.64) |
| Pythium acantophoron | 100.00 (0.00) | 100.00 (0.00) |
| Ascomycota | ||
| Botryosphaeria dothidea | 100.00 (0.00) | 25.19 (0.64) |
| Diaporthe eres | 100.00 (0.00) | 100.00 (0.00) |
| Diplodia seriata | 100.00 (0.00) | 100.00 (0.00) |
| Eutypa lata | 100.00 (0.00) | 100.00 (0.00) |
| Neofusicoccum parvum | 95.19 (1.28) | 90.00 (1.11) |
| Mycelial Growth Inhibition (%) | ||
|---|---|---|
| Pesticide | TR04 | TR05 |
| Orvego | 0.00 (0.00) | 0.00 (0.00) |
| Rally Q SC | 28.82 (1.78) | 41.18 (2.10) |
| Sercadis | 0.00 (0.00) | 0.00 (0.00) |
| Talentum 20 EW | 57.6 (2.44) | 58.43 (0.61) |
| Chorus 50 WG | 43.33 (0.48) | 51.96 (0.98) |
| Teldor 500 SC | 0.00 (0.00) | 7.25 (1.88) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, C.; Csótó, A.; Pál, K.; Nagy, A.; Fekete, E.; Karaffa, L.; Kubicek, C.P.; Sándor, E. The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines. Part I. Isolation, Identification and In Vitro Studies. Pathogens 2021, 10, 1612. https://doi.org/10.3390/pathogens10121612
Kovács C, Csótó A, Pál K, Nagy A, Fekete E, Karaffa L, Kubicek CP, Sándor E. The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines. Part I. Isolation, Identification and In Vitro Studies. Pathogens. 2021; 10(12):1612. https://doi.org/10.3390/pathogens10121612
Chicago/Turabian StyleKovács, Csilla, András Csótó, Károly Pál, Antal Nagy, Erzsébet Fekete, Levente Karaffa, Christian P. Kubicek, and Erzsébet Sándor. 2021. "The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines. Part I. Isolation, Identification and In Vitro Studies" Pathogens 10, no. 12: 1612. https://doi.org/10.3390/pathogens10121612