

The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines, Part II, Grapevine Stimulation
 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichoderma Isolates and Inocula
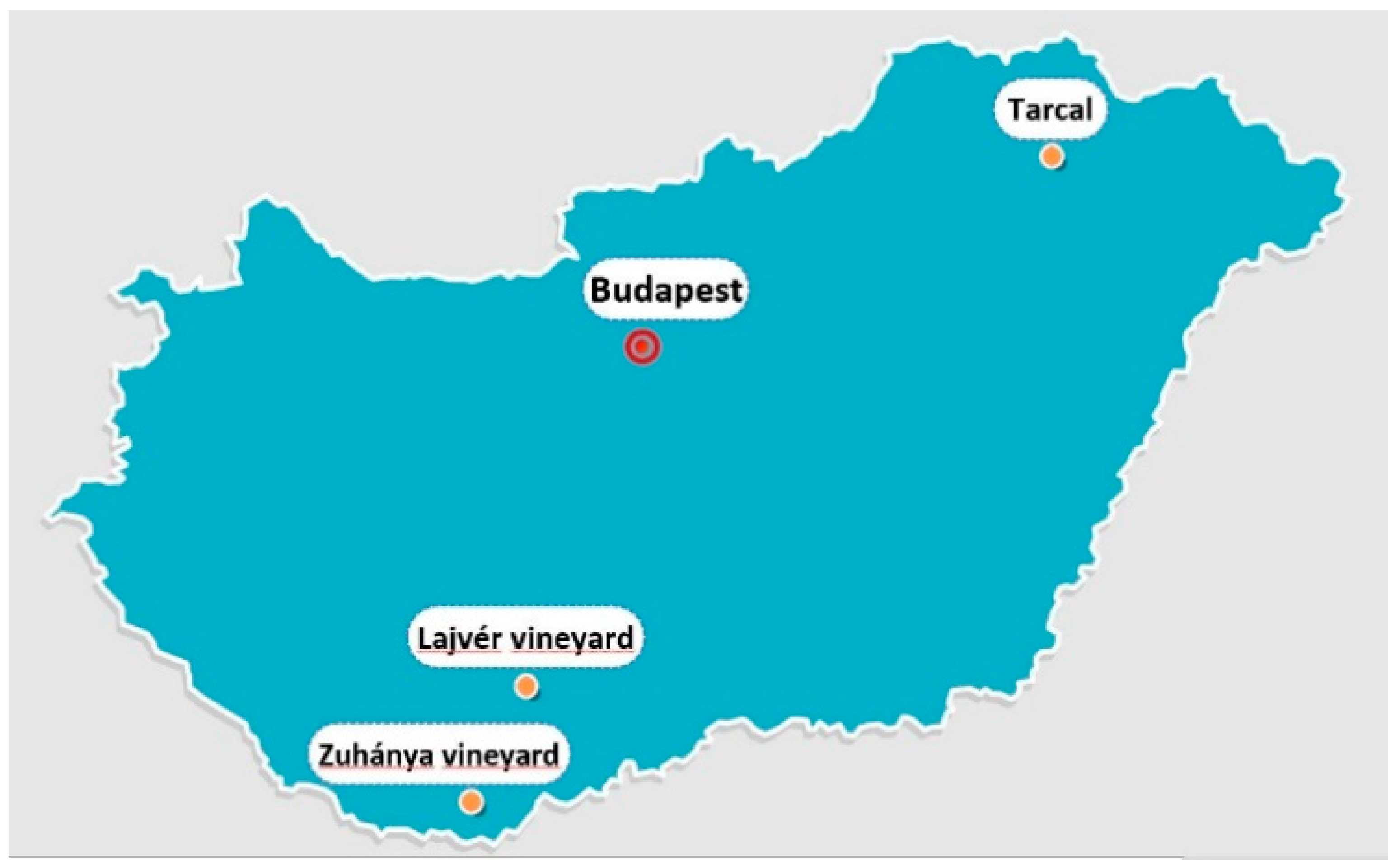
2.2. Experimental Fields and Treatments
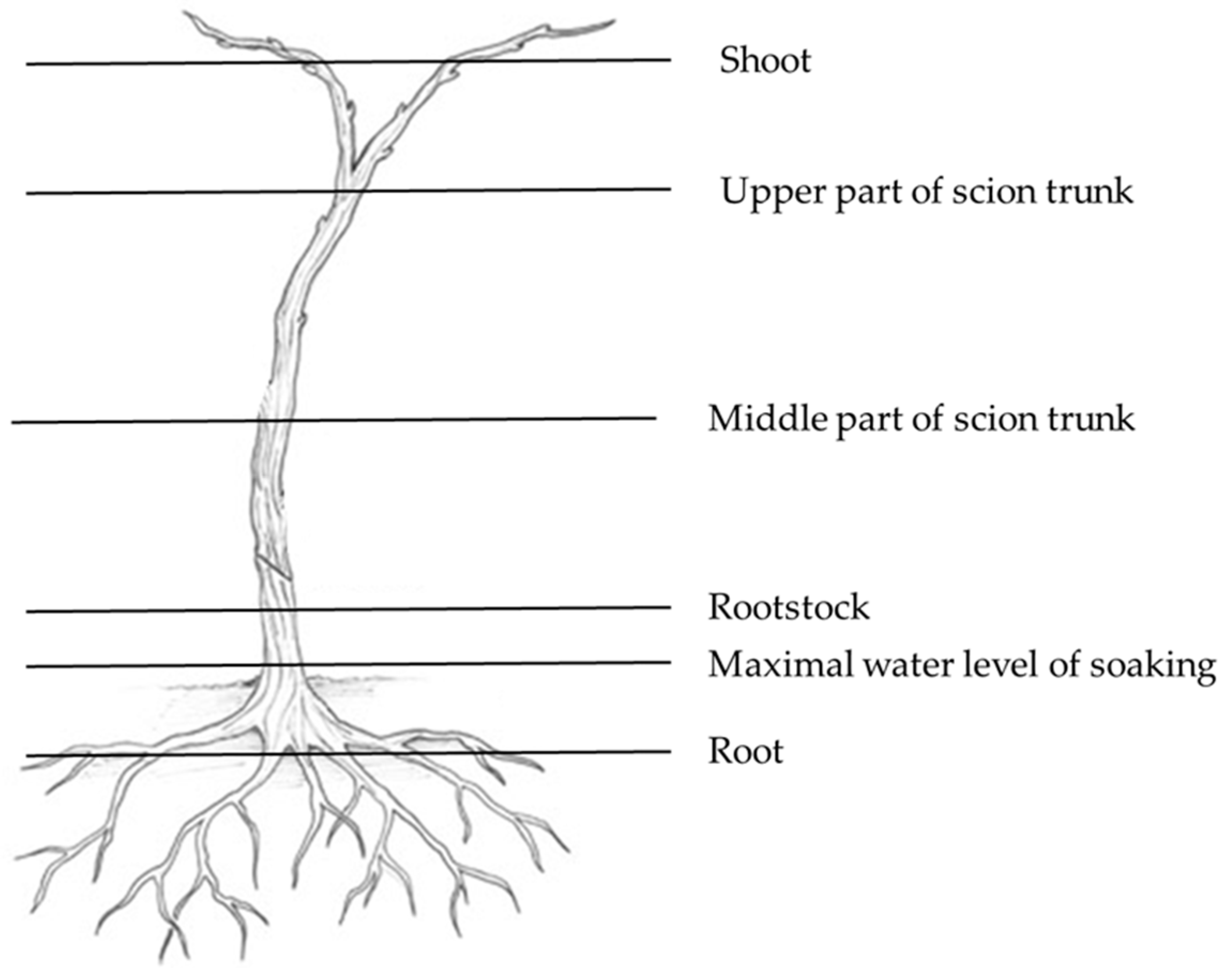
2.3. Isolation and Identification of Fungi from Woody Parts of Grapevine Plants

2.4. Determination of Lost Plant Percentage
2.5. Determination of the Effect of Trichoderma Treatment on Bud Burst
2.6. Determination of Grape Yield
2.7. Determination of Chemical Parameters from the Must
2.8. Statistical Analysis
3. Results
3.1. Potential Grape Wood Colonization by Trichoderma spp.
3.2. Loss of Planted Grafts
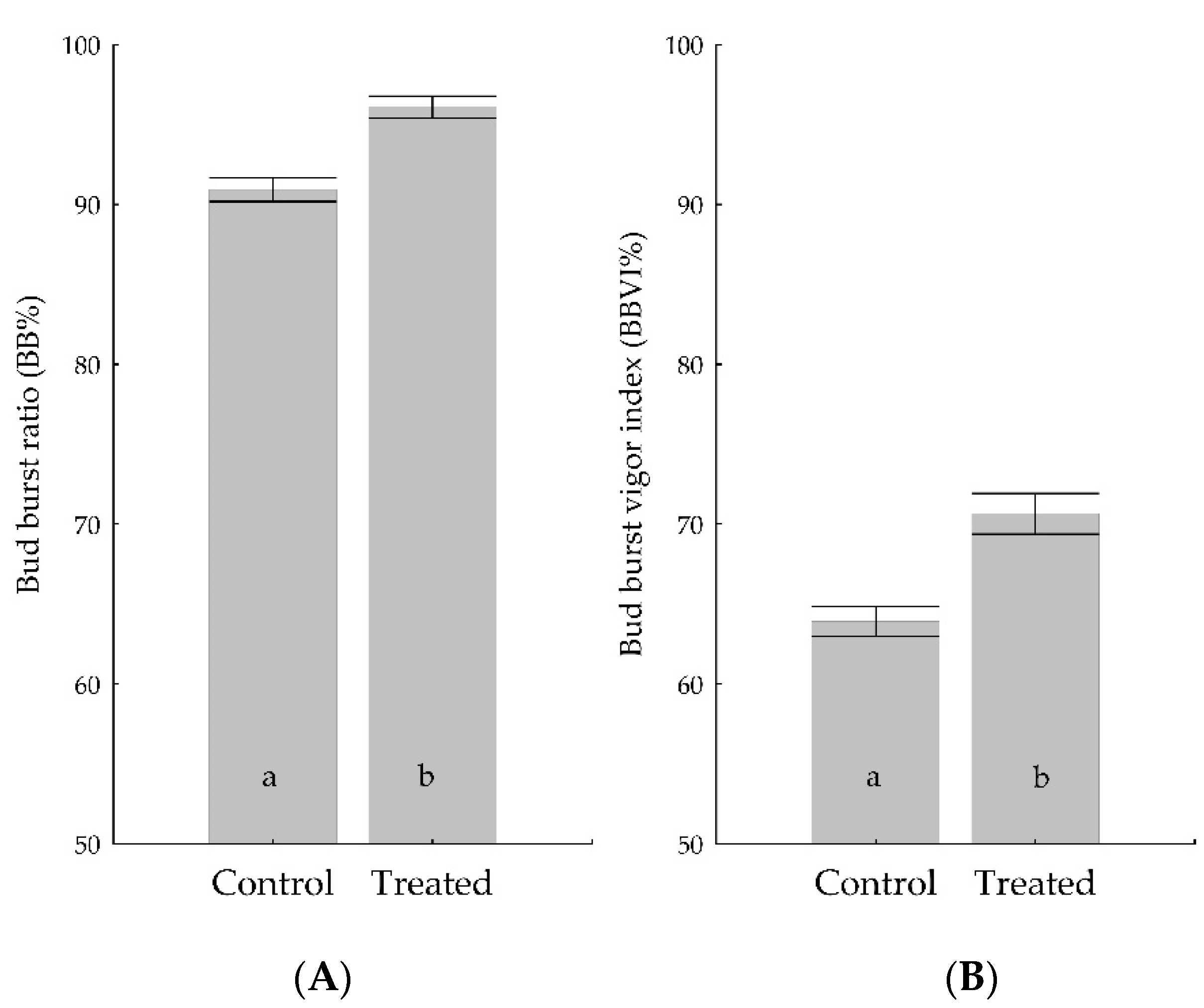
3.3. Bud Burst Ratio and Vigor Index
3.4. Quantity and Quality of Harvested Grapes
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El Gueddari, N.E.; Moerschbacher, B.M. Efficacy and risks of “biorationals“ in organic and integrated pest management. In Proceedings of the 8th International Symposium Plant Protection and Plant Health in Europe jointly organised by DPG, JKI and HU-Berlin, Braunschweig, Germany, 3 December 2017; Available online: https://plant-protection.net/fileadmin/Tagungen/PPPHE/2017/Documents/PPPHE2017_Report.pdf (accessed on 12 January 2022).
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multi level properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Pinto, C.; Custódio, V.; Nunes, M.; Songy, A.; Rabenoelina, F.; Courteaux, B.; Clément, C.; Gomes, A.C.; Fontaine, F. Understand the potential role of Aureobasidium pullulans, a resident microorganism from grapevine, to prevent the infection caused by Diplodia seriata. Front. Microbiol. 2018, 9, 3047. [Google Scholar] [CrossRef] [PubMed]
- Zhanbin, S.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, M. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Chenthamara, K.; Zhang, J.; Atanasova, L.; Yang, D.; Miao, Y.; Rahimi, M.J.; Grujic, M.; Cai, F.; Pourmehdi, S.; et al. Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 2018, 14, e1007322. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, D.L.; Baker, R. Effect of certain fungi on dwarf tomatoes grown under gnotobiotic conditions. Phytopathology 1967, 57, 1262–1263. [Google Scholar]
- Sofo, A.; Scopa, A.; Manfra, M. Trichoderma harzianum strain T-22 induces changes in phytohormone levels in cherry rootstocks (Prunus cerasus X P. canescens). Plant Growth Regul. 2011, 65, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Review. Phytoter. Res. 2013, 34, 2835–2842. [Google Scholar] [CrossRef]
- Rawal, R.; Scheerens, J.C.; Fenstemaker, S.M.; Francis, D.M.; Miller, S.A.; Benitez, M.-S. Novel Trichoderma isolates alleviate water deficit stress in susceptible tomato genotypes. Front. Plant Sci. 2022, 13, 869090. [Google Scholar] [CrossRef]
- Björkman, T.; Blanchard, L.M.; Harman, G.E. Growth enhancement of shrunken-2 sweet corn with Trichoderma harzianum 1295-22: Effect of environmental stress. J. Am. Soc. Hort. Sci. 1998, 123, 35–40. [Google Scholar] [CrossRef]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil. 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Wang, R.; Chen, D.; Khan, R.A.A.; Cui, J.; Hou, J.; Liu, T. A novel Trichoderma asperellum strain DQ-1 promotes tomato growth and induces resistance to gray mold caused by Botrytis cinerea. FEMS Microbiol. Lett. 2021, 368, fnab140. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, Z.; Zhang, Y.; Wang, Y.; Liu, Z. Biocontrol and growth-promoting effect of Trichoderma asperellum TaspHu1 isolate from Juglans mandshurica rhizosphere soil. Microbiol. Res. 2020, 242, 126596. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Chang, Y.-C.; Baker, R.; Kleifeld, O.; Chet, I. Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant. Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, Z.; Yang, X.; Ran, W.; Shen, O. Trichoderma harzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Chliyeh, M.; Chahdi, A.; Selmaoui, K.; Touhami, A.; Abdelkarim, F.-M.; El Modafar, C.; Abdelmajid, M.; Oukabli, A.; Benkirane, R.; Douira, A. Effect of Trichoderma harzianum and arbuscular mycorrhizal fungi against Verticillium wilt of Tomato. Int. J. Recent Sci. Res. 2014, 5, 449–459. [Google Scholar]
- Schubert, M.; Fink, S.; Schwarze, F.W.M.R. Evaluation of Trichoderma spp. as a biocontrol agent against wood decay fungi in urban trees. Biol. Control. 2008, 45, 111–123. [Google Scholar] [CrossRef]
- Rocha, S.L.; Evans, H.C.; Jorge, V.L.; Cardoso, L.A.; Pereira, F.S.; Rocha, F.B.; Barreto, R.W.; Hart, A.G.; Elliot, S.L. Recognition of endophytic Trichoderma species by leaf-cutting ants and their potential in a Trojan-horse management strategy. R. Soc. Open Sci. 2017, 4, 160628. [Google Scholar] [CrossRef] [Green Version]
- Holmes, K.A.; Schroers, H.-J.; Thomas, S.E.; Evans, H.C.; Samuels, G.J. Taxonomy and biocontrol potential of a new species of Trichoderma from the Amazon basin in South America. Mycol. Prog. 2004, 3, 199–210. [Google Scholar] [CrossRef]
- Rosmana, A.; Samuels, G.; Ismaiel, A.; Ibrahim, E.; Chaverri, P.; Herawati, Y.; Asman, A. Trichoderma asperellum: A dominant endophyte species in cacao grown in sulawesi with potential for controlling vascular streak dieback disease. Trop. Plant. Pathol. 2015, 40, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Di Marco, S.; Metruccio, E.G.; Moretti, S.; Nocentini, M.; Carella, G.; Pacetti, A.; Battiston, E.; Osti, F.; Mugnai, L. Activity of Trichoderma asperellum strain ICC 012 and Trichoderma gamsii strain ICC 080 toward diseases of esca complex and associated pathogens. Front. Microbiol. 2022, 12, 813410. [Google Scholar] [CrossRef] [PubMed]
- Pollard-Flamand, J.; Boulé, J.; Hart, M.; Úrbez-Torres, J.R. Biocontrol Activity of Trichoderma species isolated from grapevines in British Columbia against botryosphaeria dieback fungal pathogens. J. Fungi 2022, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Carro-Huerga, G.; Compant, S.; Gorfer, M.; Cardoza, R.E.; Schmoll, M.; Gutiérrez, S.; Casquero, P.A. Colonization of Vitis vinifera L. by the Endophyte Trichoderma sp. Strain T154: Biocontrol Activity Against Phaeoacremonium minimum. Front. Plant Sci. 2020, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- John, S.; Wicks, T.J.; Hunt, J.S.; Scott, E.S. Colonisation of grapevine wood by Trichoderma harzianum and Eutypa lata. Aust. J. Grape Wine Res. 2008, 14, 18–24. [Google Scholar] [CrossRef]
- Jaarsveld, W.J.; Halleen, F.; Bester, M.C.; Pierron, R.J.G.; Stempien, E.; Mostert, L. Investigation of Trichoderma species colonization of nursery grapevines for improved management of black foot disease. Pest Manag. Sci. 2020, 77, 397–405. [Google Scholar] [CrossRef]
- Hunt, J.S.; Gale, D.S.J.; Harvey, I.C. Evaluation of Trichoderma as biocontrol for protection against wood-invading fungi implicated in grapevine trunk diseases. Phytopathol. Mediterr. 2001, 40, 485–486. [Google Scholar]
- Berbegal, M.; Ramón-Albalat, A.; León, M.; Armengol, J. Evaluation of long term protection from nursery to vineyard provided by Trichoderma atroviride SC1 against fungal grapevine trunk pathogens. Pest. Manag. Sci. 2020, 76, 967–977. [Google Scholar] [CrossRef]
- Leal, C.; Richet, N.; Guise, J.F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar contributes to the beneficial effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to protect grapevine against Neofusicoccum parvum. Front. Microbiol. 2021, 12, 726132. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Latorre, B.; Agosin, E.; San Martin, R.; Riegel, D.G.; Nielsen, P.A.; Tronsmo, A.; Pearson, R.C. Biological and integrated control of Botrytis bunch rot of grape using Trichoderma spp. Biol. Control 1996, 7, 259–266. [Google Scholar] [CrossRef]
- Lazazzara, V.; Vicelli, B.; Bueschl, C.; Parich, A.; Pertot, I.; Schuhmacher, R.; Perazzoli, M. Trichoderma spp. volatile organic compounds protect grapevine plants by activating defense-related processes against downy mildew. Physiol. Plant. 2021, 172, 1950–1965. [Google Scholar] [CrossRef] [PubMed]
- Perazzolli, M.; Moretto, M.; Fontana, P.; Ferrarini, A.; Velasco, R.; Moser, C.; Delledonne, M.; Pertot, I. Downy mildew resistance induced by Trichoderma harzianum T39 in susceptible grapevines partially mimics transcriptional changes of resistant genotypes. BMC Genom. 2012, 13, 660. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, S.; Osti, F. Applications of Trichoderma to prevent Phaeomoniella chlamydospora infections in organic nurseries. Phytopathol. Mediterr. 2007, 46, 73–83. [Google Scholar] [CrossRef]
- Pascale, A.; Vinale, F.; Manganiello, G.; Nigro, M.; Lanzuise, S.; Ruocco, M.; Marra, R.; Lombardi, N.; Woo, S.; Lorito, M. Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Kovács, C.; Csótó, A.; Pál, K.; Nagy, A.; Fekete, E.; Karaffa, L.; Kubicek, C.P.; Sándor, E. The biocontrol potential of endophytic Trichoderma fungi isolated from Hungarian grapevines. Part I. Isolation, identification and in vitro studies. Pathogens 2021, 10, 1612. [Google Scholar] [CrossRef]
- Gaál, M.; Moriondo, M.; Bindi, M. Modelling the impact of climate change on the Hungarian wine regions using random forest. Appl. Ecol. Environ. Res. 2012, 10, 121–140. [Google Scholar] [CrossRef]
- Maul, E.; Töpfer, R. Vitis International Variety Catalogue. Available online: www.vivc.de (accessed on 15 June 2022).
- Hellman, E. Parts of the Grape Vine: Roots. Texas AgriLife Extension. 2019. Available online: https://grapes.extension.org/parts-of-the-grape-vine-roots/ (accessed on 18 September 2022).
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants; BBCH Monograph; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Stamp, J.A. The contribution of imperfections in nursery stock to the decline of young vines in California. Phytopathol. Mediterr. 2001, 40, 369–375. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, F.; Zhang, Y.; Zhang, J. Involvement of Trichoderma asperellum strain T6 in regulating iron acquisition in plants. J. Basic Microbiol. 2014, 54, 115–124. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef]
- Samolski, I.; Rincón, A.M.; Pinzón, L.M.; Viterbo, A.; Monte, E. The qid74 gene from Trichoderma harzianum has a role in root architecture and plant biofertilization. Microbiology 2012, 158, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Geilfus, C.-M.; Mühling, K.H.; Ludwig-Müller, J. The influence of salt stress on ABA and auxin concentrations in two maize cultivars differing in salt resistance. J. Plant Physiol. 2013, 170, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Doni, F.; Khadka, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. Microbiol. 2021, 130, 529–546. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.F.A. Evaluation of some biocontrol agents to control Thompson seedless grapevine powdery mildew disease. Egypt. J. Biol. Pest Control 2018, 28, 93. [Google Scholar] [CrossRef] [Green Version]
- Hickey, C. Dormant Spur and Cane Pruning Bunch Grapevines. UGA Cooperative Extension Bulletin. 2018. Available online: https://secure.caes.uga.edu/extension/publications/files/pdf/B%201505_3.PDF (accessed on 18 September 2022).
- Shukla, N.; Awasthi, R.P.; Rawat, L.; Kumar, J. Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 2012, 54, 78–88. [Google Scholar] [CrossRef]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Gomes, A.C.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2015, 144, 707–721. [Google Scholar] [CrossRef]
- Bernard, M.B.; Horne, P.A.; Hoffmann, A.A. Eriophyoid mite damage in Vitis vinifera (grapevine) in Australia: Calepitrimerus vitis and Colomerus vitis (Acari: Eriophyidae) as the common cause of the widespread ‘Restricted Spring Growth’ syndrome. Exp. Appl. Acarol. 2005, 35, 83–109. [Google Scholar] [CrossRef]
- Hlaszny, E.; Hajdu, E.; Bisztray, G.; Ladányi, M. Comparison of budburst models predictions for Kékfrankos. Appl. Ecol. Environ. Res. 2012, 10, 75–86. [Google Scholar] [CrossRef]
- Andreini, L.; Guarino, L.; Viti, R.; Scalabrelli, G. Evaluation of the effect of esca disease on bud break in Vitis vinifera L.: Possible relationship between cultivars and rootstocks. Vitis 2015, 52, 33–40. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Experimental Field | Trichoderma Strains | Inoculum Production Method | Spore Concentration (Spores mL−1) | |
|---|---|---|---|---|
| following Production | in Soaking Treatments | |||
| I (Siklós, Zuhánya) | T. simmonsii (TR05) | Culture on PDA medium | 1.5 × 109 | 106 |
| T. orientale (TR06) | 1.1 × 109 | |||
| T. gamsii (TR08) | 1.4 × 109 | |||
| II (Szálka, Lajvér) | T. afroharzianum (TR04) | Submerged liquid culture | 4.3 × 107 | 106 |
| T. simmonsii (TR05) | 6.7 × 107 | |||
| Experimental Field | Cultivar (Clone) * | Rootstock (Clone) | Planting Time |
|---|---|---|---|
| I (Siklós, Zuhánya) | Blaufraenkisch (Kt.1.) | 5BB (K21) | 17.04.–20.04.2015 |
| Blaufraenkisch (A4/1) | 5BB (We48) | ||
| Cabernet Sauvignon (337) | K5BB (ISV1) | ||
| Cabernet Franc (GM/Trv) | K5BB (101) | ||
| Cabernet Franc (E11) | K5BB (ISV1) | ||
| Cabernet Franc (ISV5) | K5BB (GM13) | ||
| II (Szálka, Lajvér) | Cabernet Sauvignon (E153) | K5BB (ISV1) | 02.05.–04.05.2017 |
| Scale | BBCH Value * | Description of Phenological Stage |
|---|---|---|
| 0 | 0 | The winter buds are dormant or aborted |
| 1 | 01–05 | Start of bud swelling to “wool stage” |
| 2 | 07–09 | Bud burst |
| 3 | 11–15 | Starting of leaf development |
| Experimental Field | ID of Isolates | Identified Trichoderma Strain | ID of Sequences | Plant Part of Re-Isolation | Date of Sampling for Re-Isolation | Elapsed Time between Treatment and Re-Isolation |
|---|---|---|---|---|---|---|
| I. | RIV3 | T. orientale | ON937623 | scion trunk (upper part) | October 2016 | 15 months |
| RIV6 | ON937624 | scion trunk (upper part) | October 2016 | 15 months | ||
| RIV7 | ON937625 | scion trunk (upper part) | October 2016 | 15 months | ||
| RIV41 | T. gamsii | ON931231 | root | August 2019 | 4 years | |
| RIV42 | ON931232 | root | August 2019 | 4 years | ||
| II. | RIS1 | T. afroharzianum | ON937626 | root | October 2017 | 5 months |
| RIS2 | ON937627 | rootstock | October 2017 | 5 months | ||
| RIS5 | ON937628 | root | October 2017 | 5 months | ||
| RIS6 | ON937629 | scion trunk (middle part) | October 2017 | 5 months |
| Cultivar | Experimental Field | n 1 | U-Test 2 | Loss of Planted Grafts (%) | |
|---|---|---|---|---|---|
| Control | Trichoderma3 | ||||
| All | I and II | 68 | p = 0.4945 | 5.33 ± 1.12 | 3.71 ± 0.54 |
| Cabernet Franc | I | 16 | p = 0.1770 | 1.88 ± 0.60 | 0.78 ± 0.36 |
| Blaufraenkisch | I | 30 | p = 0.1187 | 3.17 ± 0.68 | 4.83 ± 0.74 |
| Cabernet Sauvignon | II | 22 | p = 0.0943 | 9.46 ± 2.46 | 4.38 ± 1.03 |
| Cultivar | Experimental Field | Clone | n 1 | Treatment 2 | BB% ± SE 3 | BBVI% ± SE 3 |
|---|---|---|---|---|---|---|
| Cabernet Franc | I | E11 | 149 | Control | 94.97 ± 1.10a | 69.36 ± 1.83a |
| Trichoderma | 99.60 ± 0.40b | 90.74 ± 1.78b | ||||
| I | ISV5 | 98 | Control | 86.29 ± 2.52+ | 52.13 ± 2.26+ | |
| Trichoderma | 92.31 ± 1.98+ | 57.54 ± 2.22+ | ||||
| I | N101 | 142 | Control | 91.74 ± 1.48+ | 64.41 ± 1.95a | |
| Trichoderma | 96.50 ± 1.16+ | 71.05 ± 2.09b | ||||
| I | all | 389 | Control | 91.96 ± 0.91a | 63.96 ± 1.23a | |
| Trichoderma | 96.15 ± 0.81b | 73.28 ± 1.64b | ||||
| Blaufraenkisch | I | A4 | 150 | Control | 86.48 ± 1.98a | 52.49 ± 1.76a |
| Trichoderma | 92.43 ± 2.41b | 60.83 ± 2.32b | ||||
| I | Kt1 | 147 | Control | 88.96 ± 1.85a | 67.41 ± 2.08b | |
| Trichoderma | 95.60 ± 2.31b | 60.83 ± 4.11a | ||||
| I | all | Control | 87.72 ± 1.35a | 59.95 ± 1.46 | ||
| Trichoderma | 93.97 ± 1.67b | 60.83 ± 2.31 | ||||
| Cabernet Sauvignon | II | E153 | 98 | Control | 98.96 ± 1.04 | 80.21 ± 2.49 |
| Trichoderma | 100.00 ± 0.00 | 82.11 ± 2.49 |
| Must Parameter | Control | Trichoderma1 |
|---|---|---|
| Brix (°Bx) | 22.80 | 23.70 |
| Extract (g L−1) | 256.95 | 268.10 |
| Reducing sugar (g L−1) | 230.50 | 241.20 |
| Glucose (g L−1) | 109.65 | 114.90 |
| Fructose (g L−1) | 117.50 | 121.85 |
| Glycerol (g L−1) | 0.60 | 0.80 |
| pH | 3.39 | 3.42 |
| Total Acidity (g L−1) | 6.30 | 6.40 |
| Volatile Acidity (g L−1) | 0.09 | 0.11 |
| Potassium (mg L−1) | 1114.50 | 1153.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csótó, A.; Kovács, C.; Pál, K.; Nagy, A.; Peles, F.; Fekete, E.; Karaffa, L.; Kubicek, C.P.; Sándor, E. The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines, Part II, Grapevine Stimulation. Pathogens 2023, 12, 2. https://doi.org/10.3390/pathogens12010002
Csótó A, Kovács C, Pál K, Nagy A, Peles F, Fekete E, Karaffa L, Kubicek CP, Sándor E. The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines, Part II, Grapevine Stimulation. Pathogens. 2023; 12(1):2. https://doi.org/10.3390/pathogens12010002
Chicago/Turabian StyleCsótó, András, Csilla Kovács, Károly Pál, Antal Nagy, Ferenc Peles, Erzsébet Fekete, Levente Karaffa, Christian P. Kubicek, and Erzsébet Sándor. 2023. "The Biocontrol Potential of Endophytic Trichoderma Fungi Isolated from Hungarian Grapevines, Part II, Grapevine Stimulation" Pathogens 12, no. 1: 2. https://doi.org/10.3390/pathogens12010002