
Fungal Communities in Re-Emerging Fraxinus excelsior Sites in Lithuania and Their Antagonistic Potential against Hymenoscyphus fraxineus
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
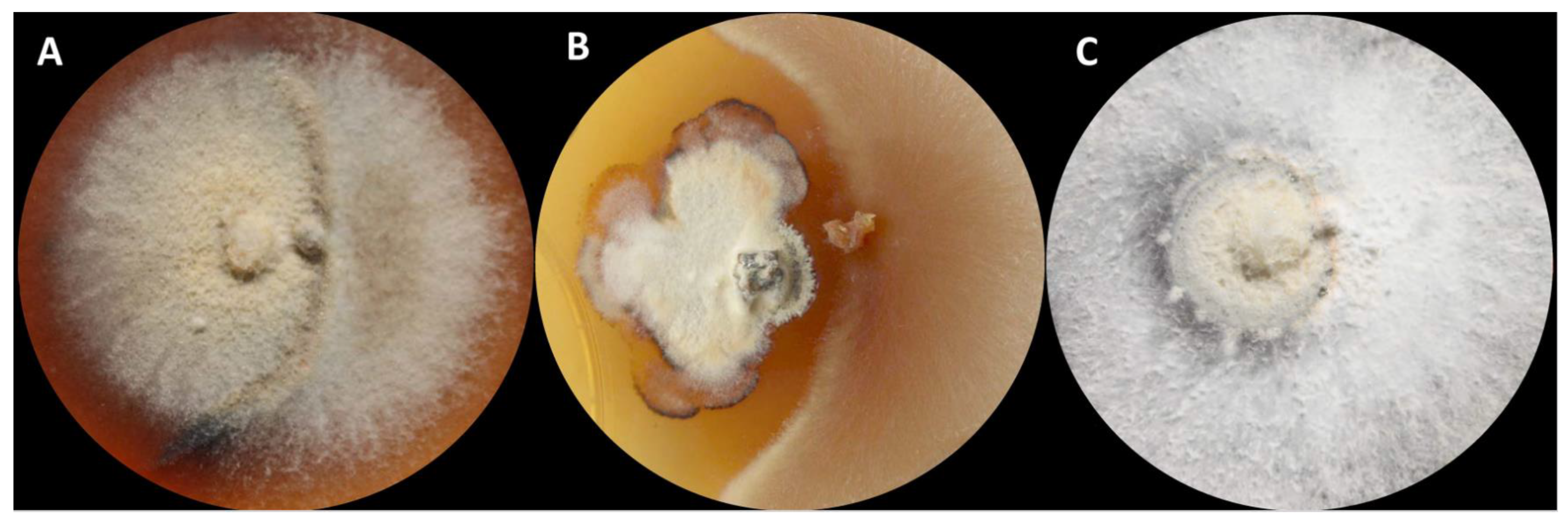
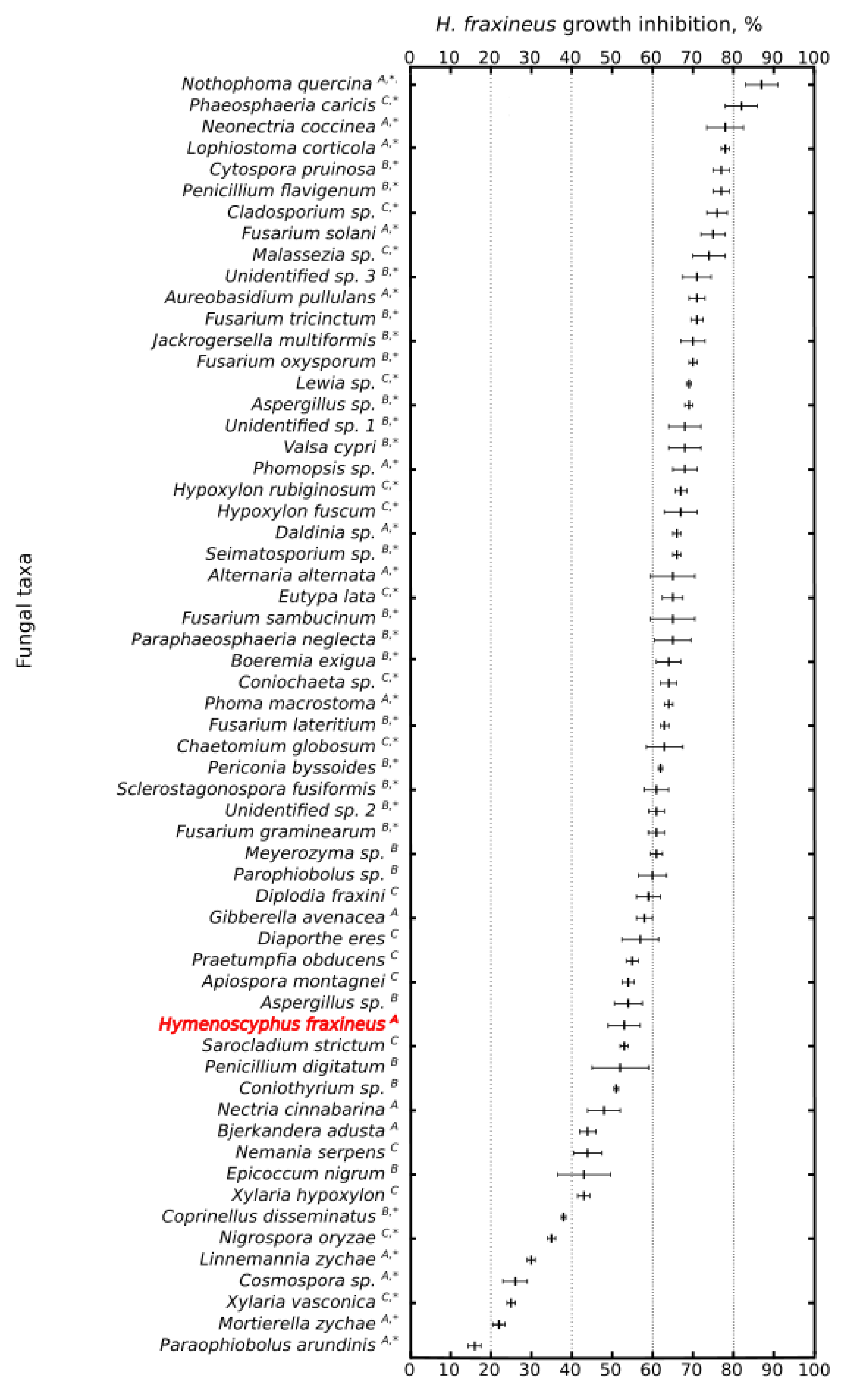
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CABI. Invasive Species Compendium—Hymenoscyphus Fraxineus (Ash Dieback). Available online: https://www.cabi.org/isc/datasheet/108083#tosummaryOfInvasiveness (accessed on 10 June 2022).
- Baral, H.O.; Queloz, V.; Hosoya, T. Hymenoscyphus fraxineus, the correct scientific name for the fungus causing ash dieback in Europe. IMA Fungus 2014, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Pautasso, M.; Aas, G.; Queloz, V.; Holdenrieder, O. European ash (Fraxinus excelsior) dieback—A conservation biology challenge. Biol. Conserv. 2013, 158, 37–49. [Google Scholar] [CrossRef]
- Anonymous. Lietuvos Miškų Ūkio Statistika [Lithuanian Statistical Yearbook of Forestry]; Valstybinė miškų tarnyba, Lututė: Kaunas, Lithuania, 2001; pp. 1–112. [Google Scholar]
- Anonymous. Lietuvos Miškų Ūkio Statistika [Lithuanian Statistical Yearbook of Forestry]; Valstybinė miškų tarnyba, Lututė: Kaunas, Lithuania, 2021; pp. 1–184. [Google Scholar]
- Grigaliūnas, K. Lithuanian State Forest Service, Kaunas, Lithuania. Personal Communication. 22th May 2021.
- Stenlid, J. Population structure of Heterobasidion annosum as determined by somatic incompatibility, sexual incompatibility, and isoenzyme patterns. Can. J. Bot. 1985, 6, 2268–2273. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Fungi and oomycetes in the irrigation water of forest nurseries. Forests 2020, 11, 459. [Google Scholar] [CrossRef]
- Kowalski, T.; Bilański, P. Fungi detected in the previous year’s leaf petioles of Fraxinus excelsior and their antagonistic potential against Hymenoscyphus fraxineus. Forests 2021, 12, 1412. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Havrdová, L.; Černý, K.; Zahradník, D.; Koukol, O. Fungal endophytes in ash shoots—Diversity and inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Ihrmark, K.; Stenlid, J. Investigations concerning the role of Chalara fraxinea in declining Fraxinus excelsior. Plant Pathol. 2009, 58, 284–292. [Google Scholar] [CrossRef]
- Davydenko, K.; Vasaitis, R.; Stenlid, J.; Menkis, A. Fungi in foliage and shoots of Fraxinus excelsior in eastern Ukraine: A first report on Hymenoscyphus pseudoalbidus. For. Pathol. 2013, 43, 462–467. [Google Scholar] [CrossRef]
- Cross, H.; Sønstebø, J.H.; Nagy, N.E.; Timmermann, V.; Solheim, H.; Børja, I.; Kauserud, H.; Carlsen, T.; Rzepka, B.; Wasak, K.; et al. Fungal diversity and seasonal succession in ash leaves infected by the invasive ascomycete Hymenoscyphus fraxineus. New Phytol. 2017, 213, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, A.; Murphy, B.R.; Hodkinson, T.R. Assessing genotypic and environmental effects on endophyte communities of Fraxinus (ash) using culture dependent and independent DNA sequencing. J. Fungi 2021, 7, 565. [Google Scholar] [CrossRef] [PubMed]
- Bakys, R.; Pliūra, A.; Bajerkevičienė, G.; Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Menkis, A. Mycobiota associated with symptomatic and asymptomatic Fraxinus excelsior in Post-Dieback Forest Stands. Forests 2022, 13, 1609. [Google Scholar] [CrossRef]
- Yun, Y.H.; Oh, M.H. First Report of Nothophoma quercina causing shoot canker on Chaenomeles sinensis in Korea. Plant Dis. 2016, 100, 2533–2534. [Google Scholar] [CrossRef]
- Răut, I.; Călin, M.; Capră, L.; Gurban, A.-M.; Doni, M.; Radu, N.; Jecu, L. Cladosporium sp. isolate as fungal plant growth promoting agent. Agronomy 2021, 11, 392. [Google Scholar] [CrossRef]
- Almatar, M.; Makky, E.A. Cladosporium cladosporioides from the perspectives of medical and biotechnological approaches. 3 Biotech 2016, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Rashmi, R.; Siddalingamurthy, K.R. Cladosporium tenuissiumum—A new producer of xyloglucanase–specific endoglucanase. Int. J. Pharma Bio Sci. 2013, 4, 1291–1297. [Google Scholar]
- Ferreira-Pinto, M.M.; Moura-Guedes, M.C.; Santos, M.R.; Silva, M.J.; Barreiro, M.G.; Santos, M. The Antagonistic activity of Aureobasidium pullulans to reduce blue mold in ‘Rocha’ pear. Acta Hortic. 2008, 800, 921–928. [Google Scholar] [CrossRef]
- Becker, R.; Ulrich, K.; Behrendt, U.; Kube, M.; Ulrich, A. Analyzing ash leaf-colonizing fungal communities for their biological control of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 590944. [Google Scholar] [CrossRef] [PubMed]
- Agostinelli, M.; Nguyen, D.; Witzell, J.; Cleary, M. Mycobiome of Fraxinus excelsior with different phenotypic susceptibility to ash dieback. Front. For. Glob. Change 2021, 4, 580514. [Google Scholar] [CrossRef]
- Agan, A.; Drenkhan, R.; Adamson, K.; Tedersoo, L.; Solheim, H.; Børja, I.; Matsiakh, I.; Timmermann, V.; Nagy, N.E.; Hietala, A.M. The relationship between fungal diversity and invasibility of a foliar niche—The case of ash dieback. J. Fungi 2020, 6, 150. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Site | Latitude, N | Longitude, E | Tree Age, y | Tree Species Composition, % * | Mean Annual Temperature, °C | Mean Annual Precipitation, mm | Stand Origin ** |
|---|---|---|---|---|---|---|---|
| U1 (Žeimelis) | 56°16′37.1″ | 24°02′33.0″ | 10–25 | 70F,10P,10B,10U | 6.3 | 620 | S |
| U2 (Kėdainiai) | 55°13′35.4″ | 23°57′58.5″ | 15–20 | 90F,10P | 6.7 | 600 | S |
| U3 (Šakiai) | 55°01′49.8″ | 23°00′20.2″ | 15–20 | 100F | 7.0 | 650 | S |
| U4 (Telšiai) | 56°03′43.4″ | 22°25′08.3″ | 17 | 70F,30Q | 6.3 | 800 | P |
| U5 (Šakiai) | 55°00′50.2″ | 23°04′00.9″ | 15–25 | 70F,10P,10B,10S | 7.0 | 650 | S |
| U6 (Dubrava) | 54°44′20.6″ | 23°47′38.8″ | 17 | 100F | 6.8 | 650 | P |
| U7 (Telšiai) | 56°03′33.8″ | 22°25′37.6″ | 17 | 70F,30Q | 6.3 | 800 | P |
| U8 (Pakruojis) | 56°00′38.4″ | 23°55′48.5″ | 15–20 | 90F,10P | 6.5 | 600 | S |
| U9 (Jonava) | 55°09′11.1″ | 24°06′56.8″ | 7–10 | 70F,30Q | 6.7 | 620 | S |
| U10 (Dubrava) | 54°45′07.4″ | 23°47′53.5″ | 15–17 | 80F,10P,10B | 6.8 | 650 | S |
| Fungal Taxa | No. of Pure Cultures/% | |
|---|---|---|
| Shoots | Leaves | |
| Alternaria alternata | 87/10.9 | 71/22.5 |
| Apiospora montagnei | 65/8.2 | 44/13.9 |
| Aspergillus sp. | 2/0.3 | -/- |
| Aspergillus sp. | 6/0.8 | -/- |
| Aureobasidium pullulans | 6/0.8 | 43/13.6 |
| Bjerkandera adusta | 12/1.5 | -/- |
| Boeremia exigua | 14/1.8 | 12/3.8 |
| Chaetomium globosum | 24/3.0 | -/- |
| Cytospora pruinosa | 2/0.3 | -/- |
| Cladosporium allicinum | 6/0.8 | 7/2.2 |
| Coniochaeta sp. | 4/0.5 | -/- |
| Coniothyrium sp. | 3/0.4 | -/- |
| Coprinellus disseminatus | 9/1.1 | -/- |
| Cosmospora sp. | 1/0.1 | 3/0.9 |
| Daldinia sp. | 2/0.3 | -/- |
| Diaporthe eres | 21/2.6 | 4/1.2 |
| Diplodia fraxini | 11/11.4 | 3/0.9 |
| Epicoccum nigrum | 49/6.2 | 14/4.4 |
| Eutypa lata | 21/2.6 | 12/3.8 |
| Fusarium graminearum | 3/0.4 | -/- |
| Fusarium lateritium | 7/0.9 | 4/1.2 |
| Fusarium oxysporum | 3/0.4 | 2/0.6 |
| Fusarium sambucinum | 2/0.3 | -/- |
| Fusarium solani | 1/0.1 | 1/0.3 |
| Fusarium tricinctum | 2/0.3 | 2/0.6 |
| Gibberella avenacea | 54/6.8 | 24/7.6 |
| Hymenoscyphus fraxineus | 1/0.1 | -/- |
| Hypoxylon fuscum | 19/2.4 | 3/0.9 |
| Hypoxylon rubiginosum | 13/1.6 | 5/1.5 |
| Jackrogersella multiformis | 2/0.3 | -/- |
| Lewia sp. | 16/2.0 | -/- |
| Linnemannia zychae | 1/0.1 | -/- |
| Lophiostoma corticola | 4/0.5 | -/- |
| Malassezia sp. | 52/6.5 | 7/2.2 |
| Meyerozyma sp. | 3/0.4 | -/- |
| Mortierella zychae | 6/0.8 | 14/4.3 |
| Nectria cinnabarina | 3/0.4 | -/- |
| Nemania serpens | 7/0.9 | 2/0.6 |
| Neonectria coccinea | 5/0.6 | -/- |
| Nigrospora oryzae | 12/1.5 | 3/0.9 |
| Nothophoma quercina | 1/0.1 | -/- |
| Paraophiobolus arundinis | 2/0.3 | -/- |
| Paraphaeosphaeria neglecta | 3/0.4 | 4/1.2 |
| Penicillium digitatum | 20/2.5 | -/- |
| Penicillium flavigenum | 6/0.8 | -/- |
| Periconia byssoides | 17/2.1 | 7/2.2 |
| Phaeosphaeria caricis | 3/0.4 | 4/1.2 |
| Phoma macrostoma | 67/8.4 | 11/3.4 |
| Phomopsis sp. | 15/1.9 | -/- |
| Praetumpfia obducens | 3/0.4 | -/- |
| Sarocladium strictum | 44/5.5 | 7/2.2 |
| Sclerostagonospora fusiformis | 3/0.4 | -/- |
| Seimatosporium sp. | 2/0.3 | -/- |
| Valsa cypri | 15/0.9 | -/- |
| Xylaria hypoxylon | 2/0.3 | 2/0.6 |
| Xylaria vasconica | 7/0.9 | 1/0.3 |
| Unidentified sp. | 5/0.6 | -/- |
| Unidentified sp. | 1/0.1 | -/- |
| Unidentified sp. | 2/0.3 | -/- |
| Total | 779/100 | 316/100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakys, R.; Bajerkevičienė, G.; Pliūra, A.; Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Mishcherikova, V.; Menkis, A. Fungal Communities in Re-Emerging Fraxinus excelsior Sites in Lithuania and Their Antagonistic Potential against Hymenoscyphus fraxineus. Microorganisms 2022, 10, 1940. https://doi.org/10.3390/microorganisms10101940
Bakys R, Bajerkevičienė G, Pliūra A, Marčiulynas A, Marčiulynienė D, Lynikienė J, Mishcherikova V, Menkis A. Fungal Communities in Re-Emerging Fraxinus excelsior Sites in Lithuania and Their Antagonistic Potential against Hymenoscyphus fraxineus. Microorganisms. 2022; 10(10):1940. https://doi.org/10.3390/microorganisms10101940
Chicago/Turabian StyleBakys, Remigijus, Gintarė Bajerkevičienė, Alfas Pliūra, Adas Marčiulynas, Diana Marčiulynienė, Jūratė Lynikienė, Valeriia Mishcherikova, and Audrius Menkis. 2022. "Fungal Communities in Re-Emerging Fraxinus excelsior Sites in Lithuania and Their Antagonistic Potential against Hymenoscyphus fraxineus" Microorganisms 10, no. 10: 1940. https://doi.org/10.3390/microorganisms10101940