
Fungicolous Fungi on Pseudosclerotial Plates and Apothecia of Hymenoscyphus fraxineus and Their Biocontrol Potential

Department of Forest Ecosystems Protection, University of Agriculture in Krakow, Al. 29 Listopada 46, 31-425 Krakow, Poland
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(11), 2250; https://doi.org/10.3390/microorganisms10112250
Submission received: 30 September 2022
/
Revised: 28 October 2022
/
Accepted: 11 November 2022
/
Published: 14 November 2022
(This article belongs to the Special Issue Restoring the Integrated Behaviour of the Soil-Plant-Microbe System)
Abstract
:In the present work, research tasks were carried out in the search for fungi with potential biocontrol possibilities in relation to the ash dieback pathogen, Hymenoscyphus fraxineus. In the years 2012–2021, dead petioles of F. excelsior and F. mandshurica were collected, on which morphological structures of H. fraxineus showed unusual symptoms of dying (apothecia) and signs of colonization by other fungi (pseudosclerotial plates). Based on morphological and molecular phylogenetic data, 18 fungal taxa were identified. Thirteen of them belong to Ascomycota: Clonostachys rosea, Cl. solani, Cordyceps sp., Minimidochium sp., Nemania diffusa, Fusarium sp., Pestalotiopsis sp., Trichoderma atroviride, T. harzianum, T. polysporum, T. rodmanii, T. tomentosum, Trichoderma sp., and five other taxa are represented by Basidiomycota: Corticiales sp., Cyathus olla, Efibula sp., Gymnopus sp. and Polyporales sp. In 108 dual cultures in vitro, three different types of interactions were distinguished: (i) physical colony contact (5.6%), (ii) presence of an inhibition zone between the colonies (0.9%), and (iii) copartner overgrowth of H. fraxineus colonies and partial or complete replacement of the pathogen (93.5%). In the dual cultures, various morphological deformations of H. fraxineus hyphae were observed: the development of apical or intercalary cytoplasmic extrusions, development of internal hyphae of the test fungi in pathogens’ hyphae, the deformation and disruption of significant sections of H. fraxineus hyphae via lysis and mycoparasitism, complete desolation of H. fraxineus cells and breakdown of hyphae into short fragments, and disappearing of pigment in the affected hyphae of H. fraxineus. The inoculation tests performed in vivo or in glass Petrie dishes showed that all the identified taxa were able to lead to pathological changes in H. fraxineus apothecia, and the mycelium of some of them completely covered pseudosclerotial plates of H. fraxineus. It was emphasized in the discussion that such activity of these fungi in forest stands may contribute to the reduction in the H. fraxineus inoculum reservoir.
1. Introduction
Ash dieback caused by an alien acomycete Hymenoscyphus fraxineus has been the most dangerous Fraxinus excelsior disease in most European countries since the 1990s [1,2,3,4]. Trees of all age classes are at risk, but generally young plantations suffer more than older trees [5,6,7]. The disease leads to a significant reduction in the population of F. excelsior in forest ecosystems and urban green areas in many European countries [8,9]. In Great Britain, for example, the total economic cost of ash dieback is estimated at £ 14.8 billion [10].
Various silvicultural methods are used to reduce the losses caused by the disease, depending on the severity of damage, local climate, age, and stand type [2,6,11]. The possibilities of reduction in the amount of infectious material [12,13] and methods of eliminating the pathogen from seeds and seedlings were indicated [14]. Some fungicides proved to be effective against H. fraxineus. However, they cannot be used on a wider scale in forest stands due to ecological and public health considerations [15]. Despite the measures undertaken, the establishment of new ash plantations in many European countries is not recommended because of the high losses in ash stands [2,16,17].
These are the reasons why alternative disease management products need to be developed, that is, in order to offer environmentally friendly and economically feasible control of this dangerous tree disease. An important element in integrated disease management strategies could be biological control. It involves the use of any living organism to extinguish a certain plant pathogen by means of competition for resources or space, antibiosis, and parasitism [18,19]. Biocontrol agents or microbial antagonists prevent infection or the establishment of the pathogen in the host plant. Thus far, approximately 300 taxa belonging to 113 fungal genera have been identified as biological control agents for plant fungal pathogens [20].
Among the main beneficial fungi with biocontrol capacity are endophytes [21]. Recent studies, conducted mainly in vitro, show that the endophytes in ash leaves and shoots are very diverse and may have a high potential for antagonistic influence toward H. fraxineus [22,23,24,25,26]. The usefulness of Thielvia basicola, an endophyte in ash petioles, was confirmed in tests with callus cultures and in vitro regenerated ash seedlings [27]. The antagonistic potential of ash endophyte Hypoxylon rubiginosum, producing the antifungal beta-tubulin phomopsidin inhibitor, has also been demonstrated [28]. Schlegel et al. [29] found that exudates of ash endophytes, Boeremia exigua, Kretzschmaria deusta, Neofabraea alba, Paraconiothyrium sp., and Fraxinicola (Venturia) fraxini inhibited H. fraxineus ascospore germination. Barta et al. [30] observed that the growth of H. fraxineus was the most inhibited by four endophytic fungi from ash twigs, Fusarium lateritium, Didymella aliena, D. macrostoma, and Dothiorella gregaria. Two of them, F. lateritium and D. aliena, caused significant inhibition of the length of necroses caused by H. fraxineus after inoculation in the same trunks of F. excelsior in situ.
After infected ash leaves fall to the bottom of the stand, H. fraxineus begins the saprotrophic phase. In this stage, a large role is played by saprotrophs, which may limit the expansion of H. fraxineus through the competition for space or antibiosis. Kowalski and Bilański [31] found over 130 fungal taxa in the previous year ash petioles. They showed that the most common competitors for H. fraxineus in situ were: Cyathicula coronata, C. fraxinophila, Hypoderma rubi, Hymenoscyphus caudatus, H. scutula, Leptosphaeria sp., Pyrenopeziza petiolaris and Typhula sp. The highest antibiotic activity against H. fraxineus in dual cultures was shown in Pseudocoleophoma polygonicola and Malbranchea sp. [31].
Among the biocontrol agents, an important group of fungi used in the environmentally sustainable approach to management of plant diseases are mycoparasites [19,32,33]. Mycoparasitism is based on the physical penetration of the parasite into the host hyphae via the development of specific organs such as haustoria, or through its growth inside hyphal cells, and secretion of enzymes or secondary metabolites leading to the degradation of fungal structures followed by nutrient uptake from the host fungus [34,35,36]. The term mycoparasitism applies strictly to those relationships in which one living fungus acts as a nutrient source for another. However, fungicolous fungi have a stable but indeterminate connection with another fungus, though usually presenting a true parasitic relationship is a difficult task [34].
Mycoparasitism can be classified as a four-step process: (a) chemotropic growth of biocontrol fungus mycelium towards the target pathogen, (b) recognition, (c) direct attachment and coiling, and (d) penetration and digestion [35,37]. Mycoparasitic relationships can be biotrophic or necrotrophic. Biotrophic mycoparasites obtain nutrients from living cells without harming the mycohost in a distinct way. The necrotrophic parasite destroys the host cell after or slightly before invasion and utilizes nutrients from the dying or dead host. The invasion is often initiated by coiling of hyphae around the mycohost cells and results in rapid destruction. The necrotrophic mycoparasites are considered to have the best potential as biocontrol agents. Some of them have been used for the commercial control of plant diseases [34,37,38,39,40,41].
The mycoparasites represent different taxonomic units and differ in terms of the extent of the mycohosts. Mycoparasites that can infest a wide range of fungi are classified as generalist, and those that occur on only one or a small number of hosts are referred to as specialist [40]. The most commonly encountered and studied mycoparasitic fungi that infest the broad-spectrum of phytopathogens are mainly Clonostachys rosea and Trichoderma spp. [20,37]. Ampelomyces quisqualis has been considered a mycoparasite that can infect dozens of species of powdery mildew. However, recent studies show that it is genetically diverse and may include several cryptic species [39]. Additionally, Eudarluca caricis, which was assumed to be a nonspecific mycoparasite of rust fungi, according to new findings, is not genetically uniform and shows host specificity [42]. Paraphaeosphaeria (Coniothyrium) minitans is a mycoparasite of Sclerotinia spp. and other related species. In the fungus Sclerotinia sclerotiorum it parasitizes both the hypha and sclerotia [43]. The soilborne oomycete, Pythium oligandrum can parasitize on many pathogens such as Rhizoctonia solani, Fusarium oxysporum and Phytophthora megasperma [44]. Verticillium biguttatum is a mycoparasite of the dangerous soil-borne fungus Rhizoctonia solani and several other plant pathogens [45]. The basidiomycete, Xenolachne longicornis, is a known parasite of some ascomycetes of the Helotiaceae family, including Hymenoscyphus vernus [46,47]. Thus far, there are no reports of fungicolous or mycoparasitic fungi on ash dieback pathogen, H. fraxineus.
The subject of the present work was research on the occurrence of fungicolous and mycoparasitic fungi on ash dieback pathogen, H. fraxineus, with the assumption that determining the belonging of the encountered fungi to one of these groups separately will not be easy and at the current stage will have to be considered together. The aims of this study were: (i) identification of fungi occurring on prematurely dead apothecia and on psedosclerotial plates of H. fraxineus in situ, (ii) determining the types of in vitro interactions in dual cultures between these species and H. fraxineus, and (iii) conducting inoculation tests to evaluate the effect of these fungi on the overgrowth of pseudosclerotial plates and the death of the apothecia of H. fraxineus developed in the previous year ash petioles, which would be associated with a reduction in the inoculum of this pathogen.
2. Materials and Methods
2.1. Sampling and Isolation of Fungi
In the years 2012–2021, during other ash dieback studies, 45 petioles of F. excelsior and F. mandshurica from the leaf litter were collected (Figure 1 and Table 1), on which morphological structures of H. fraxineus (apothecia, pseudosclerotial plates) showed unusual symptoms and signs of other fungi. The apothecia were gray-brown with some deformations, not typical for H. fraxineus. However, the pseudosclerotial plates of H. fraxineus were overgrown by various forms of mycelium (Figure 2a–o).
Petioles were placed separately in plastic bags and transported to the laboratory, where the symptoms were described in detail and photomicrographs were taken. Subsequently, various attempts were made to isolate fungi colonizing pseudosclerotia and apothecia of H. fraxineus. If spore aggregates were present, they were transferred to a drop of sterilized distilled water with a sterile preparatory needle, and then spread on the surface of the agar medium in Petrie dishes. After the hymenium fragments were collected from the apothecium, the procedure was similar. Attempts were also made to obtain colonies of H. fraxineus from brown apothecia (from ascospores), according to the method used by Kraj and Kowalski [48]. In the remaining cases, different combinations of disinfection of sample fragments with 96% ethyl alcohol and/or sterile water (3 × 30 s) were used. After drying them on sterile blotting paper, very small pieces were laid out on medium agar. Mycelia of emerging colonies were regularly transferred on MEA in new Petrie dishes and incubated for 3–4 weeks at 20 °C in darkness.
From nine petioles, it was not possible to isolate cultures whose microscopic features of mycelium or spores would match those of the fungi inhabiting H. fraxineus morphological structures in situ. Thus, isolates obtained only from 36 samples were included in the analyzes and experiments.
Fungi were grown on malt extract agar [MEA; 20 g l-1 malt extract (Difco; Sparks, MD, USA), 15 g l-1 Difco agar] supplemented with 100 mg l-1 streptomycin sulfate to suppress bacterial growth. Morphological observations, as well as initial morphological identification, were performed as described by Kowalski et al. [49].
The nomenclature of the taxa followed the Index Fungorum (http://www.indexfungorum.org, accessed 1 September 2022). Representative fungal strains were stored on MEA slants at 5 °C in the culture collection of the Department of Forest Ecosystems Protection, University of Agriculture in Kraków.
2.2. DNA Extraction, PCR and Sequencing
To verify the morphology-based identification and to identify non-sporulating morphotypes, the nucleotide sequences of the ITS region of the ribosomal RNA (rRNA) gene cluster of representative cultures were determined.
DNA was extracted from 3-week-old fungal cultures using the Genomic Mini AX Plant Kit (A&A Biotechnology, Gdynia, Poland) according to the manufacturer’s instruction. The ITS region of the rRNA gene cluster was amplified for sequencing using primers ITS1-F [50], ITS4 [51] or ITS5 and ITS4 [51]. Polymerase chain reaction (PCR) amplification and sequencing reactions of the isolates were carried out according to the methods described in detail by Bilański and Kowalski [26].
Searches using the BLASTn algorithm were performed in order to find sequences from NCBI GenBank (http://www.ncbi.nlm.nih.gov, accessed 1 September 2022) [52] and were similar to those obtained in the present study. Selected reference sequences of taxa closely related to the isolates collected in this study were obtained from GenBank with the purpose of conducting phylogenetic analysis. All newly obtained sequences in this study were deposited in GenBank with the accession numbers presented in Table 1.
An ITS sequence matches of ≥98% were used to define species boundaries and matches ≥90% were used to define order boundaries [53]. If sequencing of the ITS region of the rRNA gene cluster was ambiguous, the taxonomy was determined by phylogenetic analysis with reference sequences from the NCBI GenBank.
The ITS region of the rRNA gene cluster sequences were aligned using MAFFT v.6 [54], with the E-INS-i option and the remaining default parameters. The BioEdit v.2.7.5 [55] program was used to manually check the correctness of the alignment obtained this way.
Phylogenetic analyses were performed for ITS dataset, using two different methods, i.e., maximum likelihood (ML) and Bayesian inference (BI). The best-fit evolutionary substitution model for the dataset was established for these methods using the corrected Akaike Information Criterion (AICc) in jModelTest v.2.1.10 [56,57]. The best model that met the assumed criteria was the GTR + I + G. ML analysis and was performed in PhyML 3.0 [58] with 1000 replicates of the bootstrap. The BI analysis was carried out in MrBayes v.3.2.6 [59]. The Markov chain Monte Carlo (MCMC) algorithm was run for 10 million generations using the best-fit model. Trees were sampled every 100 generations, resulting in 100,000 trees from both runs and discarding the first 25% as burn-in. The resulting phylogenetic tree (Figure 3) contain all the ITS region of the rRNA gene cluster sequences generated in this study.
2.3. Dual Culture Assay
Obtained fungal isolates (Table 1) were screened for their interaction with the colonies of three strains of H. fraxineus (HMC 20952, HMC 21508 and Js 9359, further described as Hf1, Hf2, and Hf3, respectively) by in vitro dual culture trials on malt extract agar (one dish per interaction). All of H. fraxineus strains originate from the previous year leaf petioles, with developed pseudosclerotial plates, collected in 2017 from the litter in ash stands in southern Poland. They showed different rates of growth on MEA; after three weeks, the average colony radii were: Hf1-3.5, Hf2-3.9, and Hf3-2.8 cm.
A plug of young, actively growing mycelium (8 mm diam.) of a H. fraxineus isolate was placed inside the plate, 25 mm from its margin; 7 days later, a similar mycelium plug of the test fungus was placed 40 mm from the H. fraxineus plug and 25 mm from the opposite margin of the dish. Two replicates for each H. fraxineus strain without other fungi served as control plates. All these plates were incubated in the dark at 20 °C. The interactions between the pathogen and test fungi were assessed after 3 weeks. Any earlier or later (up to 6 weeks) changes in the cultures were also noted. The width of the inhibition zone (mm) was measured along the axis joining the plugs of both fungi.
Three types of pathogen-test fungus interactions were considered, following Bilański and Kowalski [26]: type A, mutual direct contact, when both colonies meet along the contact line, without inhibition zone; type B, with an inhibition zone between colonies; and type C, the test fungus overgrows and covers the colony of H. fraxineus. Within type C, fungi that had only overgrown the H. fraxineus colonies weakly (<1.5 cm from the edge of the colony at the time of evaluation) have been noted separately. When a test fungus overgrew and covered the H. fraxineus colony, its efficacy against H. fraxineus was checked using re-isolations (6 inocula taken from each combination). Microscopic observations of H. fraxineus mycelium in the impact zone were performed to verify the occurrence of morphological deformations in comparison to the control cultures. This was done for at least one isolate of each fungal taxon.
2.4. Inoculation of Ash Petioles
Inoculations were performed in the second half of July, for some isolates in 2019, and for others in 2021. About 250 previous year leaf petioles of F. excelsior with pseudosclerotial plates and apothecia of H. fraxineus in ash stands in Miechów-Domiarki and Ojców were collected each year (Figure 1). The experiments were performed in vitro and in vivo (Supplementary Table S1). In both situations, a set of five petioles, with a total of more than 20 apothecia of H. fraxineus, was used to study the interaction with a given fungal species. These petioles were stacked side by side in parallel. In 2019, on each set of petioles, in their basal and apical parts, four pieces of agar (approx. 2 cm × 0.5 cm) overgrown with about 14-day-old mycelium of a given taxon were placed across the petioles. There was one set per tested interaction used. The controls were petioles without fungal inoculum.
Sterile glass Petri dishes (20 cm diam., 2.8 cm height) were used for the in vitro test. A sterile blotting paper was placed at the bottom of these dishes and was wetted with sterile water at the beginning and approximately every 7 days. Then, three glass rods (5 mm diameter) were placed in parallel to each dish on the blotting paper, approx. 5 cm apart from each other. A set of five petioles was placed perpendicularly on these glass rods, so that the petioles would not lay directly on the blotting paper. After inoculation, Petrie dishes were placed in plastic boxes in a room (approx. 18 °C) with diffused daylight.
The in vivo test was carried out on a fenced area in the form of urban greenery (over 400 m2). This site was partly (about 30%) moderately shaded by coniferous and deciduous trees and bushes. The ground was mostly covered with grass with extensive patches of moss. Sets of five petioles were put in different, partially shaded places with previously cut down grass. After inoculation, each set was covered with two living, densely needled shoots, about 30 cm long (Thuja sp., Juniperus sabina).
In 2021, the test was carried out according to the same methodological assumptions, with one difference regarding the inoculum. In order to obtain inoculum, the fungi have been grown for two weeks in darkness at room temperature on MEA. Subsequently, sterile 3-cm-long fragments of ash petioles were placed on the colonies and incubated for two additional weeks. Four such fragments, overgrown by the test fungus, were placed in parallel between the petioles so that they adhered to them as closely as possible.
The inoculated petioles were observed every 5–7 days with the use of a magnifying glass, and the observed changes have been noted. A final evaluation of the condition of the pseudosclerotial plates and apothecia of H. fraxineus was done after 17 to 21 days. Mycelium (spores) on pseudosclerotial plates and changes, indicating death or inactivation of apothecia, have been noted. To estimate the intensity of this phenomenon, the following scale was adopted: scarce (<10% of dying apothecia), moderately numerous (11–50%), and very numerous (>50%). The share of a given taxon in the process of colonization of petioles and death of apothecia was confirmed by re-isolation (six fragments for each taxon) and through microscopic examination.
3. Results
3.1. Fungi Occurring on Pseudosclerotial Plates and Apothecia of H. fraxineus
A total of 36 petioles colonized by H. fraxineus with developed pseudosclerotial plates and apothecia were analyzed. They were mostly petioles of F. excelsior (31), while a few (5) belonged to F. mandshurica. Overall, on eleven petioles, apothecia occurred either singularly or in groups that showed dying symptoms. Both the stipe and receptacle were brown in such apothecia. The disc was sometimes wrinkled and smaller compared to the live neighboring white apothecia (Figure 2a–f and Table 1).
However, on 25 petioles, on pseudosclerotial plates of H. fraxineus, various forms of mycelium were present, sometimes with spores (Figure 2g–o and Table 1). In total, 18 fungal taxa that colonized the morphological structures of H. fraxineus were identified (Figure 3 and Table 1). Thirteen of them belong to Ascomycota: Clonostachys rosea (5 samples), Cl. solani (2), Cordyceps sp. (3), Minimidochium sp. (1), Nemania diffusa (1), Fusarium sp. (4), Pestalotiopsis sp. (2), Trichoderma atroviride (3), T. harzianum (2), T. polysporum (1), T. rodmanii (1), T. tomentosum (1), and Trichoderma sp. 1 (3) (Table 1). Whereas five taxa are represented by Basidiomycota: Corticiales sp. (1), Cyathus olla (1), Efibula sp. (1), Gymnopus sp. (1), and Polyporales sp. (3) (Figure 3 and Table 1). The currently isolated Fusarium sp. differed morphologically and genetically from the endophytic Fusarium species (FeC23, FeC44) and saprotrophic Fusarium species (FeF132, FeF133) isolated from ash petioles in South Poland (Figure 3).
Four taxa, Corticiales sp., Minimidochium sp., T. harzianum and T. rodmanii, were found only on apothecia, while the rest were found either only on pseudosclerotial plates or on both types of substrate (Table 1). On one petiole, next to the T. polysporum mycelium, dead apothecium of H. fraxineus was present, which was also colonized by this fungus species (Figure 2n).
The apothecia were only clearly overgrown with mycelium when they were colonized by T. atroviride and T. harzianum (Figure 2d,f), while on the remaining apothecia, the mycelium or spore aggregates were poor (Figure 2a–c,e). On most pseudosclerotial plates, the mycelium was spread (Figure 2g,i–m), less often fine powdery clusters of spores (Cordyceps sp.), or in the case of T. polysporum and Trichoderma sp. 1, the cushion-shaped mycelium with spores (Figure 2n,o) were present. Efibula sp. produced distinct mycelial cords (Figure 1h). Conidia in situ or in vitro were produced by: Cl. rosea, Cl. solani, Cordyceps sp., Fusarium sp., Pestalotiopsis sp., and Trichoderma species, while sexual morphs were not observed in any taxon.
3.2. Competition Test in Dual Cultures
Overall, the observations were made in 108 dual cultures in vitro (3 H. fraxineus strains × 36 fungal isolates). In dual cultures, at the time of evaluation, three different types of interactions were distinguished: (i) physical colony contact, (ii) presence of an inhibition zone between the colonies, and (iii) copartner overgrowth of H. fraxineus colonies (Table 2). The frequency of these types varied considerably. The physical contact of the colonies of both partners (type A) was observed only in 5.6% of the dual cultures, in those variants where the copartners were two Cordyceps sp. isolates (Table 2, Figure 4a). In the following two weeks, these isolates also slightly overgrew the colonies of H. fraxineus and produced conidia on them. An inhibition zone (type B) was only visible in one dual culture, between H. fraxineus (Hf1) and Minimidochium sp. (Figure 4b and Table 2). Such a zone was present earlier (after 2 weeks) in all three dual cultures with this fungus. However, Minimidochium sp. in dual cultures with two H. fraxineus strain (Hf2 and Hf3) started to produce a delicate mycelium that outgrew the inhibition zone and started to slightly overgrow the pathogen colony. The development of this mycelium only in a limited zone may indicate chemotropism (Figure 4c and Table 2).
In a third type of interaction (type C), the H. fraxineus colony became overgrown by the test fungus. It happened in 101 (93.5%) of the dual cultures (Table 2). The intensity of their mycelium development on the pathogen colony was varied. To a small extent (less than 1.5 cm from the edge of the colony), H. fraxineus was overgrown in 26 dual cultures by: Corticiales sp., Cordyceps sp. Fe446K, Minimidochium sp., Fusarium sp. and Pestalotiopsis sp. (Figure 4d,e and Table 2). Even after the evaluation period, the development of their mycelium was relatively slow. However, the colonies of H. fraxineus were covered more intensively by: Cl. rosea, Cl. solani, Cy. olla, Efibula sp., Gymnopus sp., Polyporales sp., N. diffusa and Trichoderma species (Figure 4f–l and Table 2). Four of them: Cy. olla, Polyporales sp., T. atroviride and Trichoderma sp. 1 complete replaced H. fraxineus colonies until evaluation, the others did so one–three weeks after evaluation. Mycelia of Cy. olla, Efibula sp. and Polyporales sp. after contact with H. fraxineus showed a change in structure; mycelium was cord-like with a fan-shaped pattern (Figure 4g,i,k). After re-cultivation from the overgrowth zone, only the test fungal colonies were detected.
In 32 dual cultures (10 fungal taxa) showing a type C interaction, when the colonies of test fungi approached the colony of H. fraxineus, their growth was stopped, a narrow inhibition zone was formed, then this zone became overcrossed and their growth continued (Figure 4e,f and Table 2). In addition, it was observed that, in six dual cultures, H. fraxineus produced a narrow blackish zone on the side of the copartner’s interaction (Figure 4i and Table 2). In this zone, the melanized hyphae of H. fraxineus showed a black-brown color instead of rusty brown. However, among the test fungi, a wide black-brown zone was observed only in Cy. olla colonies (Figure 4h).
In the dual cultures, various morphological deformations of H. fraxineus hyphae (compared to the control colonies) were observed (Figure 4m–t and Table 2). The most common modifications included: (i) the development of apical or intercalary cytoplasmic extrusions (Figure 4m); (ii) the lysis of hyphae in the zone of physical contact of the copartners, sometimes leading to the complete destruction of H. fraxineus cells and breakdown of hyphae into short fragments (Figure 4o,p); and (iii) the deformation and disruption of significant sections of H. fraxineus hyphae via lysis and mycoparasitism. In addition, the production of appressoria by T. tomentosum and the haustoria-like structures by T. harzianum (Figure 4t) was observed. These pathological changes were particularly visible in the thick and pigmented H. fraxineus hyphae. Inside such hyphae, the hyaline hyphae of the mycoparasite were visible (Figure 4m,q–t). The pigment disappeared in the affected hyphae of H. fraxineus (Figure 4q–t). It is extremely important that Cordyceps sp. (Fe 446K), Efibula sp. and Polyporales sp. are able to attack melanized hyphae of H. fraxineus forming a black zone in dual cultures (Figure 4i,n), because melanized hyphae form, in situ, a pseudosclerotial plate of H. fraxineus.
3.3. Inoculation Test
Petioles inoculations were performed in vivo and/or in glass Petrie dishes (Figure 5a–t and Supplementary Material Table S1). The test showed that all the fungi used were able to lead to pathological changes in H. fraxineus apothecia that were not found in apothecia not affected by fungi.
Fungal taxa colonized the ash petioles in a significantly diverse manner. Fungi such as: Cl. rosea, Cl. solani, Cordyceps sp., Corticiales sp., Fusarium sp., Minimidochium sp. and Pestalotiopsis sp. developed rather sparse filamentous or floccose mycelium on the surface of the pseudosclerotium (Figure 5a–t). As soon as such hyphae reached the apothecia, brown spots began to appear on the hymenium (Figure 5a,n). With time, the apothecia turned completely brown and numerous hyphae appeared on some of them, and others, Cl. rosea, Cl. solani and Cordyceps sp., also produced conidiophores and conidia (Figure 5b–d). Some fungi (Cordyceps sp., Fusarium sp.) developed filamentous hyphae, especially between apothecia, showing some form of chemotropism (Figure 5e,m). Four fungal taxa: Cy. olla, Efibula sp., Gymnopus sp. and Polyporales sp., developed abundant mycelium on both petioles and apothecia (Figure 5g–j,o), which led to the rapid death of the apothecia (Figure 5h–j,o).
Efibula sp. quickly spread to subsequent petioles due to the production of string-like mycelium (Figure 5i). Trichoderma species spread relatively quickly within individual petioles, producing mainly cobweb or threadlike mycelium (Figure 5p–t and Supplementary Material Table S1). Only T. polysporum produced cushion-like mycelium on petioles and in the vicinity of apothecia (Figure 5s). T. harzianum and Trichoderma sp. 1 formed abundant clusters of mycelium on colonized apothecia (Figure 5r,t). Atypical development of new H. fraxineus apothecia on petioles inoculated with T. atroviride (377E, 664K) and Cl. solani was observed. Only stipes mostly developed excessively, reaching dimensions of 3–6 × 1.2–1.8 mm. If an apothecial disc developed, it only reached about 2.5 mm in diameter (Figure 3q).
The rate of colonization of petioles by inoculated fungi influenced the number of apothecia of H. fraxineus showing pathological changes. Slowly spreading fungi were only able to colonize a few apothecia (<10%) (Supplementary Material Table S1).
4. Discussion
In the life cycle of H. fraxineus, an important role is played by the saprotrophic stage, which begins after the leaves fall to the bottom of the stand. On leaf residues, especially petioles and thicker veins, the pathogen produces black pseudosclerotial plates. Only on these pseudosclerotia does H. fraxineus produce huge amounts of apothecia with ascospores capable of infecting live ash [1,60,61,62]. Current research has shown that both of these H. fraxineus structures can be colonized by different fungi. Such apothecia were characterized, among other things, by the fact that they turned brown prematurely and declined. Assessment of the condition of apothecia must be very careful, as typical white apothecia of H. fraxineus, without the participation of other fungi, turn ochraceous to dark brown with age [63]. The research revealed 18 fungal taxa that are able to colonize pseudosclerotia and apothecia of H. fraxineus. Only a few of these taxa have so far been found on ash petioles studied in Poland. There were only two species, Clonostachys rosea and Nemania diffusa, living, symptomless petioles in tree crown and in dead petioles lying at the bottom of the stands [26,31]. This indicates that the pseudosclerotia or apothecia of H. fraxineus were colonized predominantly by other fungal species than those colonizing symptomless or dead petioles. Clear differences between these fungi could be found on the basis of the interactions in dual cultures in vitro. Currently, in 93.5% of cases, an overgrowth of H. fraxineus colonies by the test fungi was found. In the case of endophytes and saprotrophs from ash petioles, the share of this type of interaction was below 10%, and the most common type of interaction with H. fraxineus was the creation of an inhibition zone [26,31]. It can be assumed that the dual-culture assay showed different strategies used by various fungal species to colonize various substrates and to obtain the nutrients that are essential for their survival [64].
The deformations currently observed in the mycelia of H. fraxineus strains indicate a destructive effect of the test fungi in dual cultures on the pathogen. This effect is very often proved to be lethal to numerous hyphae of H. fraxineus, which was associated with apical or intercalar cytoplasmic extrusions. This phenomenon has been observed many times in fungi and is believed to be the result of increasing pressure within hyphal cells affected by toxic organic compounds [26,65]. Other phenomena observed in H. fraxineus mycelium, such as local lysis and hyphae fragmentation, loss of cytoplasm and cell walls, development of appresorium-like and haustoria-like structures, development of test fungus hyphae closely adjacent to the pathogen’s hyphae and their development inside pathogen hyphae are characteristic for mycoparasites [35,66,67]. These changes led to such a state that re-isolations of fungi from covered H. fraxineus colonies indicated the successful replacement of H. fraxineus by the numerous test fungi. The results of the in-dual cultures tests turned out to be largely consistent with the results of the inoculation test performed.
All isolates used in test inoculations were able to make destructive changes in single or multiple apothecia of H. fraxineus, leading to their premature death. The observed differences in the development of mycelium on the pseudosclerotial plate on the tested petioles compared to the petioles sampled in situ can be explained by various external conditions, especially temperature and humidity, which largely influence the development of mycelium [65]. Petioles were collected in situ at different times of year, from May to October, and it is not known how long they were colonized by fungi. In the experiment, the period of fungal development after inoculation was limited to a few weeks.
The species of fungi found in the present research differ significantly in terms of lifestyle and ecological role in forest ecosystems, agriculture, or horticulture. The findings of some of them as factors that may affect the reduction of H. fraxineus inoculum puts their significance in a completely new light. The best-known biocontrol agents are species of the genus Clonostachys and Trichoderma.
Cl. rosea strains are widely distributed all over the world. They appeared most commonly in soil and different plants organs, such as roots, leaves and flowers [37]. Cl. rosea is a destructive parasite against several plant pathogens affecting different crops and forest species [37,68]. For example, it killed spores and vegetative cells of Ceratocystis fimbriata [69], parasites on F. circinatum hyphae [68], Rhizoctonia solani hyphae and sclerotia [70,71] and the Crinipellis roreri pseudostroma, a dangerous causal agent of frosty pod rot on Theobroma gileri and T. cacao [72,73]. Cl. rosea can cause decomposition of fungal cell walls due to the ability to produce appropriate enzymes [37]. In the interaction with fungal pathogens, secondary metabolite production often plays a significant role as well [37,40,64]. Several studies have shown that one possible way in which Cl. rosea can reduce the disease incidence is by induced systemic resistance [74]. This has recently been shown for P. radiata-F. circinatum pathosystem [68]. While Cl. rosea represents a huge resource with many possibilities, experience in applying of Cl. rosea is lacking and requires more research [37].
Fungal species from the genus Trichoderma (sexual morph Hypocrea) are widely distributed in nature, especially in soil [75]. According to Thambugala et al. [20], Trichoderma is the fungal genus with the greatest biocontrol potential. Twenty-five Trichoderma species have been used in this capacity against several phytopathogens, such as Botrytis cinerea, Fusarium spp., Pythium spp., Rhizoctonia solani, Sclerotium rolfsii, and Sclerotinia sclerotiorum [20,67,76]. The main biocontrol strategies developed by Trichoderma against fungal pathogens are mycoparasitism, competition, antibiosis and systemic induced resistance [67,76]. Lytic enzymes synthesized by Trichoderma species, particularly chitinases, β-1,3-glucanases, and proteases are believed to be responsible for their mycoparasitic activity, leading to the degradation of the cell walls of fungal pathogens. Moreover, numerous Trichoderma species synthesize a large number of different secondary metabolites that are involved in mycoparasitic action [76,77,78]. Trichoderma-based products are marketed worldwide and used for crop protection against numerous disease-causing agents. The living fungal spores are applied in a variety of preparations used for spraying leaves, covering wounds after pruning, treating seeds and propagating material, or by introducing them into the soil before sowing or transplanting plants [40].
The present results support the argument that H. fraxineus may also be included in the fungal pathogens that can be suppressed by numerous Trichoderma species. This is indicated by both the dual cultures assay and the inoculations test. The mycelium of Trichoderma spp. was found on ash petioles in situ, mainly in the period from May to August, i.e., before or during the fructification of H. fraxineus, which should be assessed as a positive phenomenon. The appearance of Trichoderma in the final stage of petioles degradation would be of less importance for the reduction of the pathogen’s inoculum.
Minimidochium species are known mainly as saprotrophic fungi that colonize leaf litter or as endophytes [79,80]. Some species can produce extracellular laccases, which are responsible for the breakdown of lignin in white rot fungi [81]. The observed antibiotic activity of Minimidochium sp., lysis of hyphal wall and the possible effect on the death of H. fraxineus apothecia are a new aspect for this genus. Studies on the isolate Minimidochium sp. Jp49, strongly antagonistic to H. fraxineus, showed that it did not damage callus cultures and seedlings of F. excelsior and F. pennsylvanica [27].
Fusarium species exhibit diverse lifestyles. They can occur as saprotrophs, endophytes or plant pathogens, causing serious damage to crop and forest plants. Torbati et al. [82] also found a fungicolous Fusarium species associated with smut fungi. This genus also includes mycoparasitic species, which for a long time have been destroying known pathogens such as Botrytis cinerea, Cercospora spp. and Rhizoctonia solani [83,84,85,86]. The current results justify including Fusarium sp. to colonize the structure of H. fraxinus in the group of mycotrophic fungi. It is a species that is taxonomically different from those found in Poland as endophytes or saprotrophs on ash petioles. Its virulence has not been known. If found to be non-pathogenic, it could be considered as an interesting biocontrol agent against H. fraxineus.
Cordyceps species (Cordycipitaceae) are known primarily as important entomopathogenic fungi with a worldwide distribution. Numerous studies show that entomopathogenic fungi play very diverse functions in nature. They can act as endophytes, antagonists to plant pathogens or plant growth stimulants. Great research interest currently lies in the possibility of using entomopathogenic fungi as endophytes for the biological control of fungal diseases as well as insect pests [87,88]. In the course of the current research, it could be observed that, when Cordyceps sp. reached H. fraxineus structures, it proliferated and formed the next generation of spores, thus negatively affecting the pathogen. In dual cultures, despite limited expansion on H. fraxineus mycelium, some Cordyceps sp. isolates were capable of damaging H. fraxineus hyphae, which should be considered as a new aspect in the ecological role of this fungus.
Pestalotiopsis species show different modes of life. Among them there are pathogens that cause diseases in a variety of plants, endophytes and saprotrophs in leaves, bark and twigs. Moreover, species of Pestalotiopsis have been found to produce very numerous secondary metabolites that may be applied in medicine, agriculture and industry [89,90]. Only in recent years, Pestalotiopsis sp. was detected as a mycoparasite of the plant pathogen Aecidium wenshanense [91]. This study reveals a large difference between the mechanism of mycoparasitism of Pestalotiopsis and that of Trichoderma [91]. In Pestalotiopsis sp., the number of mycoparasitism related β-1,3-glucanases was greater than that in Trichoderma. Recent studies have shown that another fungicolous Pestalotiopsis species produces three new sesquiterpenoids related to the caryophyllene-derived punctaporonins [92]. The obtained results suggest that the Polish strain of Pestalotiopsis exhibits mycotrophic activity in relation to H. fraxineus.
Nemania diffusa, like Cl. rosea, was found in ash petioles, both as endophyte and saprotroph [26,31]. It cannot be ruled out that its mycelium colonizing petioles in the saprotrophic stage can grow on pseudosclerotial plates of H. fraxineus without the need for external infection. The results show that its positive role does not have to only be limited to competition for space inside petioles. N. diffusa produces enzymes such as phenoloxidases, peroxidase, acid phosphatase and laccase, which have the ability to neutralize fungitoxic compounds and take part in the decomposition of lignocellulosic substrates [93]. There is also a possibility that they can allow for the destruction of fungal cell walls, as observed in dual cultures in vitro.
Mycoparasitism by basidiomycetous fungi has been known for a long time [94]. The observed interactions in dual cultures and on inoculated petioles seem to confirm such possibilities of the test fungi in relation to H. fraxineus. Their antagonistic activity is probably favored by the relatively quick production of mycelium, which abundantly covers ash petioles. They are ecologically important fungi, because they produce enzymes to degrade lignocellulosic material and they may be used as an inoculant to accelerate plant residue decomposition. Identified Basidiomycota species show significant differences in terms of the occurrence and lifestyle. For example, Efibula is a genus characterized by resupinate basidiomata classified currently in the family Irpicaceae. They are typically found growing on the underside of dead trees, causing a white rot. E. tuberculata is a plant pathogen infecting plane trees [95]. In Brazil a representative of this genus was found to be a frequent endophyte in petioles of the medicinal plant Vochysia divergens [90]. Gymnopus species are cap-forming fungi that grow on the ground or on dead logs and can create mycorrhiza. Species of this genus belong to the most common endophytes in Orchidaceae [80]. Unlike Collybia or Psathyrella, there are no known mycotrophic species among Gymnopus [96]. Cyathus olla, a bird’s nest fungus, is the most abundant species of Cyathus found in Europe, and has been reported in other regions of the world. It is a saprotroph, considered to be an aggressive colonizer due to the formation of mycelial cords that invade the substrate. It also spreads through the peridioles and basidiospores. In the last two decades, the role of Cy. olla in the biological control of stubble-borne diseases of canola (Brassica napus, B. rapa) has been studied [97,98]. This species could play a similar role for ash dieback by reducing the inoculum reservoir of H. fraxineus.
When evaluating the results of the inoculations test, it should be considered that the inoculum was added only locally on the petioles, so the fungi needed some time before they came into contact with H. fraxineus apothecia. Application of spores or fragmented mycelium as a spray directly on the apothecia and along the entire petioles would probably have a more damaging effect on H. fraxineus. Observations during the experiment have been conducted for less than a month. However, the impact on the apothecia of H. fraxineus in nature may be much greater, as new generations of apothecia forming during the growing season may be invaded by the inoculum formed in large numbers on the originally attacked apothecia. Besides, H. fraxineus persists for several years on the petioles, which increases the overall amount of litter inoculum [61]. Mycoparastic fungi may reduce this persistence as they can grow from old petioles to freshly fallen leaves into litter. This may increase the importance of such fungi.
From the fungal taxa briefly presented above, it appears that forest stands (soil, leaf litter or dead wood) are their natural environment. If we were to develop an appropriate methodology and introduce the inoculum of some of these fungi as biocontrol agents, it would not involve the risk of introducing foreign organisms into the environment. Some of these taxa, however, would require further detailed identification and assessment of virulence. The conducted experiments did not provide a final solution, but presented the fungal taxa, within which it is possible to search for an optimal organism for the biocontrol of ash dieback pathogen, H. fraxineus. It can be assumed that there would be different applicability of the identified biocontrol fungi against H. fraxineus. In numerous places in forests and in larger ash groups in urban green areas, where leaf removal proposed by Noble et al. [13] is impossible, biocontrol fungi could be used to reduce the inoculum reservoir of H. fraxineus in the litter. Basidiomycetous species appear to be particularly promising for this purpose [94,95,96,97]. It is possible that the mass colonization of ash petioles in a given year will result in the colonization of freshly fallen leaves in the following years. This would lead to a long-term inoculum reduction. The identified biocontrol fungi could also be used to prevent infections at the base of the stems of especially valuable trees, which often opens the way for subsequent attacks by Armillaria species [99,100]. For this purpose, mycoparasitic fungi belonging to Ascomycota could be more useful [20,37].
5. Conclusions
Use of beneficial microorganisms in biological control could become an important factor in integrated disease management strategies with regard to ash dieback. Current studies have shown that certain fungi are able to colonize apothecia and pseudosclerotial plates of ash dieback pathogen, H. fraxineus, contributing to the reduction of its inoculum. However, this phenomenon occurs relatively rarely in natural conditions. Morphological structures of H. fraxineus can be colonized by fungi such as Clonostachys rosea and Trichoderma species, which are known mycoparasites in the broad spectrum of mycohosts, as well as by more specialized ascomycetous fungi, such as Cordyceps, Fusarium, Minimidochium and Pestalotiopsis. An important role should be attributed to the presently found fungi belonging to Basidiomycota: Corticiales sp., Cyathus olla, Efibula sp., Gymnopus sp., and Polyporales sp., which grow intensively on ash petioles and synthesize enzymes that enable them to intensively decompose lignocellulosic substrates. The conducted research provides indications concerning the potential of many of the identified species of fungi for their use against H. fraxineus. A very positive aspect is the fact that these are fungi that occur naturally in forests, soil, leaf litter or dead wood.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10112250/s1, Table S1: Symptoms on apothecia and pesudosclerotial plates of Hymenoscyphus fraxineus up to three weeks after inoculation of ash petioles with test fungi.
Author Contributions
Conceptualization, T.K. and P.B.; methodology, T.K. (mycological aspects) and P.B. (molecular and statistical aspects); investigation, T.K. and P.B.; formal analysis, T.K. and P.B.; data curation, T.K. and P.B.; writing—original draft preparation, T.K. and P.B.; writing—review and editing, T.K. and P.B.; software, P.B.; supervision, T.K. and P.B.; visualization, P.B.; project administration and funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted in a project no. 2016/21/B/NZ9/01226, financed by the National Science Centre, Poland.
Data Availability Statement
The data presented in this study are available in Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Enderle, R. An overview of ash (Fraxinus spp.) and the ash dieback disease in Europe. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pastircáková, K.; Adamcíková, K.; Barta, M.; Pažitnỳ, J.; Hot’ka, P.; Sarvašová, I.; Horáková, M.K. Host range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests 2020, 11, 596. [Google Scholar] [CrossRef]
- Marçais, B.; Kosawang, C.; Laubray, S.; Kjær, E.; Kirisits, T. Ash dieback. In Forest Microbiology Vol. 2 Forest Tree Health; Asiegbu, F., Kovalchuk, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 215–237. ISBN 9780323984485. [Google Scholar]
- Kowalski, T.; Czekaj, A. Symptomy chorobowe i grzyby na zamierających jesionach (Fraxinus excelsior L.) w drzewostanach Nadleśnictwa Staszów. Disease symptoms and fungi on dying ash trees (Fraxinus excelsior L.) in Staszów Forest District stands. Leśne Pract. Badaw. 2010, 71, 357–368. [Google Scholar]
- Skovsgaard, J.P.; Wilhelm, G.J.; Thomsen, I.M.; Metzler, B.; Kirisits, T.; Havrdová, L.; Enderle, R.; Dobrowolska, D.; Cleary, M.; Clark, J. Silvicultural strategies for Fraxinus excelsior in response to dieback caused by Hymenoscyphus fraxineus. For. Int. J. For. Res. 2017, 90, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T.; Freinschlag, C. Ash dieback caused by Hymenoscyphus pseudoalbidus in a seed plantation of Fraxinus excelsior in Austria. J. Agric. Ext. Rural Dev. 2012, 4, 184–191. [Google Scholar] [CrossRef]
- Matisone, I.; Matisons, R.; Laiviņš, M.; Gaitnieks, T. Statistics of ash dieback in Latvia. Silva Fenn. 2018, 52, 6. [Google Scholar] [CrossRef] [Green Version]
- Coker, T.L.R.; Rozsypálek, J.; Edwards, A.; Harwood, T.P.; Butfoy, L.; Buggs, R.J.A. Estimating mortality rates of European ash (Fraxinus excelsior) under the ash dieback (Hymenoscyphus fraxineus) epidemic. Plants People Planet 2019, 1, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Hill, L.; Jones, G.; Atkinson, N.; Hector, A.; Hemery, G.; Brown, N. The £15 billion cost of ash dieback in Britain. Curr. Biol. 2019, 29, R315–R316. [Google Scholar] [CrossRef]
- Havrdová, L.; Zahradník, D.; Romportl, D.; Pešková, V.; Černý, K. Environmental and silvicultural characteristics influencing the extent of ash dieback in forest stands. Balt. For. 2017, 23, 168–182. [Google Scholar]
- Bartha, B.; Mayer, A.; Lenz, H.D. Acceleration of ash petiole decomposition to reduce Hymenoscyphus fraxineus apothecia growth—A feasible method for the deprivation of fungal substrate. Balt. For. 2017, 23, 82–88. [Google Scholar]
- Noble, R.; Woodhall, J.W.; Dobrovin-Pennington, A.; Perkins, K.; Somoza-Valdeolmillos, E.; Gómez, H.L.; Lu, Y.; Macarthur, R.; Henry, C.M. Control of Hymenoscyphus fraxineus, the causal agent of ash dieback, using composting. For. Pathol. 2019, 49, e12568. [Google Scholar] [CrossRef]
- Hauptman, T.; Piškur, B.; de Groot, M.; Ogris, N.; Ferlan, M.; Jurc, D. Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. For. Pathol. 2013, 43, 360–370. [Google Scholar] [CrossRef]
- Hauptman, T.; Celar, F.; de Groot, M.; Jurc, D. Application of fungicides and urea for control of ash dieback. iFor.-Biogeosci. For. 2015, 8, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Gil, W.; Kowalski, T.; Kraj, W.; Zachara, T.; Łukaszewicz, J.; Paluch, R.; Nowakowska, J.A.; Oszako, T. Ash dieback in Poland—History of the phenomenon and possibilities of its limitation. In Dieback of European Ash (Fraxinus spp.): Consequences and Guidelines for Sustainable Management; Vasaitis, R., Enderle, R., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2017; pp. 176–184. [Google Scholar]
- Rozsypálek, J.; Dvořák, M.; Longauer, R.; Botella, L.; Prouza, M.; Palovčíková, D.; Jankovský, L. Ash and ash dieback in the Czech Republic. In Dieback of European Ash (Fraxinus spp.): Consequences and Guidelines for Sustainable Management; Vasaitis, R., Enderle, R., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2017; pp. 79–88. [Google Scholar]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major biological control strategies for plant pathogens. Pathogens 2022, 11, 273. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Omidvari, M.; Abbaszadeh-Dahaji, P.; Omidvar, R.; Kariman, K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Schulz, B.; Haas, S.; Junker, C.; Andrée, N.; Schobert, M. Fungal endophytes are involved in multiple balanced antagonisms. Curr. Sci. 2015, 109, 39–45. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Havrdová, L.; Černý, K.; Zahradník, D.; Koukol, O. Fungal endophytes in ash shoots—Diversity and inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Kosawang, C.; Amby, D.B.; Bussaban, B.; McKinney, L.V.; Xu, J.; Kjær, E.D.; Collinge, D.B.; Nielsen, L.R. Fungal communities associated with species of Fraxinus tolerant to ash dieback, and their potential for biological control. Fungal Biol. 2018, 122, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.; Ulrich, K.; Behrendt, U.; Kube, M.; Ulrich, A. Analyzing ash leaf-colonizing fungal communities for their biological control of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 590944. [Google Scholar] [CrossRef] [PubMed]
- Bilański, P.; Kowalski, T. Fungal endophytes in Fraxinus excelsior petioles and their in vitro antagonistic potential against the ash dieback pathogen Hymenoscyphus fraxineus. Microbiol. Res. 2022, 257, 126961. [Google Scholar] [CrossRef]
- Nawrot-Chorabik, K.; Marcol-Rumak, N.; Latowski, D. Investigation of the biocontrol potential of two ash endophytes against Hymenoscyphus fraxineus using in vitro plant–fungus dual cultures. Forests 2021, 12, 1750. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.-P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Schlegel, M.; Dubach, V.; von Buol, L.; Sieber, T.N. Effects of endophytic fungi on the ash dieback pathogen. FEMS Microbiol. Ecol. 2016, 92, fiw142. [Google Scholar] [CrossRef]
- Barta, M.; Pastirčáková, K.; Ostrovský, R.; Kobza, M.; Kádasi Horáková, M. Culturable endophytic fungi in Fraxinus excelsior and their interactions with Hymenoscyphus fraxineus. Forests 2022, 13, 1098. [Google Scholar] [CrossRef]
- Kowalski, T.; Bilański, P. Fungi detected in the previous year’s leaf petioles of Fraxinus excelsior and their antagonistic potential against Hymenoscyphus fraxineus. Forests 2021, 12, 1412. [Google Scholar] [CrossRef]
- Baker, R. Mycoparasitism: Ecology and physiology. Can. J. Plant Pathol. 1987, 9, 370–379. [Google Scholar] [CrossRef]
- Parratt, S.R.; Laine, A.-L. The role of hyperparasitism in microbial pathogen ecology and evolution. ISME J. 2016, 10, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Jeffries, P. Biology and ecology of mycoparasitism. Can. J. Bot. 1995, 73, 1284–1290. [Google Scholar] [CrossRef]
- Steyaert, J.M.; Ridgway, H.J.; Elad, Y.; Stewart, A. Genetic basis of mycoparasitism: A mechanism of biological control by species of Trichoderma. N. Zeal. J. Crop Hortic. Sci. 2003, 31, 281–291. [Google Scholar] [CrossRef]
- Daguerre, Y.; Siegel, K.; Edel-Hermann, V.; Steinberg, C. Fungal proteins and genes associated with biocontrol mechanisms of soil-borne pathogens: A review. Fungal Biol. Rev. 2014, 28, 97–125. [Google Scholar] [CrossRef]
- Sun, Z.-B.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, M.-H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, R.D. Mycoparasitism of soilborne plant pathogen. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 275–293. [Google Scholar]
- Kiss, L. A review of fungal antagonists of powdery mildews and their potential as biocontrol agents. Pest Manag. Sci. 2003, 59, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Zhao, H.; Yang, X.; Qiang, C.; Cheng, J.; Xie, J.; Chen, T.; Jiang, D.; Fu, Y. Functional analysis of the melanin-associated gene CmMR1 in Coniothyrium minitans. Front. Microbiol. 2018, 9, 2658. [Google Scholar] [CrossRef]
- Nischwitz, C.; Newcombe, G.; Anderson, C.L. Host specialization of the mycoparasite Eudarluca caricis and its evolutionary relationship to Ampelomyces. Mycol. Res. 2005, 109, 421–428. [Google Scholar] [CrossRef]
- Li, G.Q.; Huang, H.C.; Miao, H.J.; Erickson, R.S.; Jiang, D.H.; Xiao, Y.N. Biological control of sclerotinia diseases of rapeseed by aerial applications of the mycoparasite Coniothyrium minitans. Eur. J. Plant Pathol. 2006, 114, 345–355. [Google Scholar] [CrossRef]
- Brožová, J. Exploitation of the mycoparasitic fungus Pythium oligandrum in plant protection. Plant Prot. Sci. 2002, 38, 29–35. [Google Scholar]
- McQuilken, M.P.; Gemmell, J. Enzyme Production by the Mycoparasite Verticillium biguttatum against Rhizoctonia solani. Mycopathologia 2004, 157, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Grauwinkel, B. Xenolachne longicornis Hauerslev 1977—Ein Parasit auf Hymenoscyphus vernus. Rheinland-Pfälzisches Pilzjournal 1993, 3, 3–5. [Google Scholar]
- Van Vooren, N.; Gaignon, M. Xenolachne longicornis, une espèce rare de Tremellaceae parasite d’ascomycètes inoperculés. Publ. Société linnéenne Lyon 2002, 71, 376–379. [Google Scholar] [CrossRef]
- Kraj, W.; Kowalski, T. Genetic variability of Hymenoscyphus pseudoalbidus on ash leaf rachises in leaf litter of forest stands in Poland. J. Phytopathol. 2014, 162, 218–227. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Collado, J.; Platas, G.; Paulus, B.; Bills, G.F. High-throughput culturing of fungi from plant litter by a dilution-to-extinction technique. FEMS Microbiol. Ecol. 2007, 60, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, T.; Białobrzeski, M.; Ostafińska, A. The occurrence of Hymenoscyphus pseudoalbidus apothecia in the leaf litter of Fraxinus excelsior stands with ash dieback symptoms in southern Poland. Acta Mycol. 2013, 48, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T. Ascocarp formation of Hymenoscyphus fraxineus on several-year-old pseudosclerotial leaf rachises of Fraxinus excelsior. For. Pathol. 2015, 45, 254–257. [Google Scholar] [CrossRef]
- Hietala, A.M.; Børja, I.; Cross, H.; Nagy, N.E.; Solheim, H.; Timmermann, V.; Vivian-Smith, A. Dieback of European ash: What can we learn from the microbial community and species-specific traits of endophytic fungi associated with ash? In Endophytes of Forest Trees. Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 229–258. ISBN 978-3-319-89832-2. [Google Scholar]
- Baral, H.-O.; Bemmann, M. Hymenoscyphus fraxineus vs. Hymenoscyphus albidus—A comparative light microscopic study on the causal agent of European ash dieback and related foliicolous, stroma-forming species. Mycology 2014, 5, 228–290. [Google Scholar] [CrossRef]
- Wicklow, D.T. Interference competition. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 265–274. [Google Scholar]
- Walker, G.M.; White, N.A. Introduction to Fungal Physiology. In Fungi; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–35. [Google Scholar]
- Huang, H.C.; Kokko, E.G. Penetration of Hyphae of Sclerotinia sclerotiorum by Coniothyrium minitans Without the Formation of Appressoria. J. Phytopathol. 1988, 123, 133–139. [Google Scholar] [CrossRef]
- Kishan, G. Deciphering the mechanism of mycoparasitism of Sclerotinia sclerotiorum by Trichoderma spp. Int. J. Pure Appl. Biosci. 2017, 5, 1246–1250. [Google Scholar] [CrossRef]
- Moraga-Suazo, P.; Sanfuentes, E.; Le-Feuvre, R. Induced systemic resistance triggered by Clonostachys rosea against Fusarium circinatum in Pinus radiata. For. Res. 2016, 5, 174. [Google Scholar] [CrossRef] [Green Version]
- Barnett, H.L.; Lilly, V.G. A destructive mycoparasite, Gliocladium roseum. Mycologia 1962, 54, 72–77. [Google Scholar] [CrossRef]
- Jager, G.; Hoopen, A.; Velvis, H. Hyperparasites of Rhizoctonia solani in Dutch potato fields. Netherlands J. Plant Pathol. 1979, 85, 253–268. [Google Scholar] [CrossRef]
- Salamone, A.L.; Gundersen, B.; Inglis, D.A. Clonostachys rosea, a potential biological control agent for Rhizoctonia solani AG-3 causing black scurf on potato. Biocontrol Sci. Technol. 2018, 28, 895–900. [Google Scholar] [CrossRef]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Ten Hoopen, G.M.; Rees, R.; Aisa, P.; Stirrup, T.; Krauss, U. Population dynamics of epiphytic mycoparasites of the genera Clonostachys and Fusarium for the biocontrol of black pod (Phytophthora palmivora) and moniliasis (Moniliophthora roreri) on cocoa (Theobroma cacao). Mycol. Res. 2003, 107, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Lahoz, E.; Contillo, R.; Porrone, F. Induction of systemic resistance to Erysiphe orontii Cast in tobacco by application on roots of an isolate of Gliocladium roseum Bainier. J. Phytopathol. 2004, 152, 465–470. [Google Scholar] [CrossRef]
- Loc, N.H.; Huy, N.D.; Quang, H.T.; Lan, T.T.; Thu Ha, T.T. Characterisation and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology 2020, 11, 38–48. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Sivan, A.; Chet, I. Degradation of fungal cell walls by lytic enzymes of Trichoderma harzianum. Microbiology 1989, 135, 675–682. [Google Scholar] [CrossRef] [Green Version]
- De La Cruz, J.; Hidalgo-Gallego, A.; Lora, J.M.; Benitez, T.; Pintor-Toro, J.A.; Llobell, A. Isolation and characterization of three chitinases from Trichoderma harzianum. Eur. J. Biochem. 1992, 206, 859–867. [Google Scholar] [CrossRef]
- Sutton, B.C. Minimidochium setosum n. gen., n. sp. and Dinemasporium aberrans n. sp. from West Africa. Can. J. Bot. 1969, 47, 2095–2100. [Google Scholar] [CrossRef]
- Rashmi, M.; Kushveer, J.; Sarma, V. A worldwide list of endophytic fungi with notes on ecology and diversity. Mycosphere 2019, 10, 798–1079. [Google Scholar] [CrossRef]
- Saparrat, M.; Arambarri, A.; Balatti, P. Growth and extracellular laccase production in liquid cultures of Minimidochium parvum LPS # 548 Strain. Boletín Soc. Argent. Botánica 2007, 42, 39–44. [Google Scholar]
- Torbati, M.; Arzanlou, M.; Sandoval-Denis, M.; Crous, P.W. Multigene phylogeny reveals new fungicolous species in the Fusarium tricinctum species complex and novel hosts in the genus Fusarium from Iran. Mycol. Prog. 2019, 18, 119–133. [Google Scholar] [CrossRef]
- Rathaiah, Y.; Pavgi, M.S. Fusarium semitectum mycoparasitic on Cercosporae. J. Phytopathol. 1973, 77, 278–281. [Google Scholar] [CrossRef]
- Arora, D.K.; Dwivedi, R.S. Mycoparasitism of Fusarium spp. on Rhizoctonia solani Kühn. Plant Soil 1980, 55, 43–53. [Google Scholar] [CrossRef]
- Chaturvedi, A.P.; Dwivedi, R.S. Mycoparasitic behaviour of Fusarium oxysporum Schlechtendahl towards Aspergillus luchuensis Inui. Plant Soil 1985, 84, 419–422. [Google Scholar] [CrossRef]
- Vajna, L. Phytopathogenic Fusarium oxysporum Schlecht, as a necrotrophic mycoparasite. J. Phytopathol. 1985, 114, 338–347. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Vega, F. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.-D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palácio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the Brazilian biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yu, J.; Ma, C.; Kong, L.; He, C.; Li, J. Genomic analysis of the mycoparasite Pestalotiopsis sp. PG52. Polish J. Microbiol. 2021, 70, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.H.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T. Pestaloporonins: Caryophyllene-derived sesquiterpenoids from a fungicolous isolate of Pestalotiopsis sp. Org. Lett. 2015, 17, 4284–4287. [Google Scholar] [CrossRef]
- Tang, D.; Zhang, G.; Wang, Y.; Zhang, M.; Wang, Y.; Yu, H. Characterization of complete mitochondrial genome of Nemania diffusa (Xylariaceae, Xylariales) and its phylogenetic analysis. Mitochondrial DNA Part B 2020, 5, 459–460. [Google Scholar] [CrossRef] [Green Version]
- Griffith, N.T.; Barnett, H.L. Mycoparasitism by Basidiomycetes in culture. Mycologia 1967, 59, 149–154. [Google Scholar] [CrossRef]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.-H.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef]
- Reyes, R.G.; Umagat, M.R.; Garcia, B.L.; Barza, A.J.J.; Ryo, S.; Nobuo, M.; Noriko, M.; Fumio, E. A New Record of the Mycoparasitic Habit of Collybia reinakeana RGR-FE-NSC Strain against Aspergillus flavus, Fusarium oxysporum and Cladosporium sphaerospermum. Int. J. Pharm. Res. Allied Sci. 2017, 6, 29–32. [Google Scholar]
- Shinners, T.C.; Tewari, J.P. Morphological and RAPD Analyses of Cyathus olla from crop residue. Mycologia 1998, 90, 980. [Google Scholar] [CrossRef]
- Shinners-Carnelley, T.C.; Szpacenko, A.; Tewari, J.P.; Palcic, M.M. Enzymatic activity of Cyathus olla during solid state fermentation of canola roots. Phytoprotection 2005, 83, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Chandelier, A.; Gerarts, F.; San Martin, G.; Herman, M.; Delahaye, L. Temporal evolution of collar lesions associated with ash dieback and the occurrence of Armillaria in Belgian forests. For. Pathol. 2016, 46, 289–297. [Google Scholar] [CrossRef]
- Enderle, R.; Sander, F.; Metzler, B. Temporal development of collar necroses and butt rot in association with ash dieback. iFor.—Biogeosci. For. 2017, 10, 529–536. [Google Scholar] [CrossRef]
Figure 1.
Locations of sampling sites in Poland: 1—Brody, 2—Gorce Konina, 3—Kowary, 4—Kraków Młynówka, 5—Kraków Zakrzówek, 6—Miechów Domiarki, 7—Myślenice, 8—Ojców, 9—Rogów, 10—Stary Sącz, and 11—Trzęsacz.
Figure 1.
Locations of sampling sites in Poland: 1—Brody, 2—Gorce Konina, 3—Kowary, 4—Kraków Młynówka, 5—Kraków Zakrzówek, 6—Miechów Domiarki, 7—Myślenice, 8—Ojców, 9—Rogów, 10—Stary Sącz, and 11—Trzęsacz.

Figure 2.
Previous year ash petioles with developed in situ apothecia and pseudosclerotial plates of Hymenoscyphus fraxineus: (a–f) apothecia with degrading symptoms colonized by: (a) Clonostachys rosea Fe886F, (b) Minimidochium sp.1 Fe173F, (c) Fusarium sp. 1 Fm409E, (d) Trichoderma atroviride Fe377E, (e) Trichoderma harzianum Fm506K (f) Trichoderma harzianum Fe412E; (g–o) pseudosclerotial plate covered with mycelium of: (g) Cyathus olla Fe141F, (h) Efibula sp. 1 Fe287F, (i) Gymnopus sp. 1 Fe1035F, (j) Polyporales sp. 1 Fe302F, (k) Polyporales sp. 1 Fe1011F, (l) Pestalotiopsis sp. 1 Fe447K, (m) Trichoderma atroviride Fe57K, (n) Trichoderma polysporum Fe166F—mycelium and dead apothecium (arrow), and (o) Trichoderma sp. 1 Fe462K; scale bar = 2 mm.
Figure 2.
Previous year ash petioles with developed in situ apothecia and pseudosclerotial plates of Hymenoscyphus fraxineus: (a–f) apothecia with degrading symptoms colonized by: (a) Clonostachys rosea Fe886F, (b) Minimidochium sp.1 Fe173F, (c) Fusarium sp. 1 Fm409E, (d) Trichoderma atroviride Fe377E, (e) Trichoderma harzianum Fm506K (f) Trichoderma harzianum Fe412E; (g–o) pseudosclerotial plate covered with mycelium of: (g) Cyathus olla Fe141F, (h) Efibula sp. 1 Fe287F, (i) Gymnopus sp. 1 Fe1035F, (j) Polyporales sp. 1 Fe302F, (k) Polyporales sp. 1 Fe1011F, (l) Pestalotiopsis sp. 1 Fe447K, (m) Trichoderma atroviride Fe57K, (n) Trichoderma polysporum Fe166F—mycelium and dead apothecium (arrow), and (o) Trichoderma sp. 1 Fe462K; scale bar = 2 mm.

Figure 3.
Phylogram obtained from Maximum Likelihood (ML) analyses of the ITS data for the isolated fungal taxa. Sequences obtained during this study are presented in bold type. The Bootstrap values ≥ 75% for ML and Maximum Parsimony (MP) analyses are presented at nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analyses. * Bootstrap values < 75%. The scale bar corresponds to expected number of substitutions per nucleotide site. Cyathus olla was used as the outgroup taxon.
Figure 3.
Phylogram obtained from Maximum Likelihood (ML) analyses of the ITS data for the isolated fungal taxa. Sequences obtained during this study are presented in bold type. The Bootstrap values ≥ 75% for ML and Maximum Parsimony (MP) analyses are presented at nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analyses. * Bootstrap values < 75%. The scale bar corresponds to expected number of substitutions per nucleotide site. Cyathus olla was used as the outgroup taxon.

Figure 4.
Various types of interactions observed in dual cultures (a–l) and morphological deformations (m–t) (MEA, 3 weeks, 20 °C), Hymenoscyphus fraxineus (either Hf1, Hf2 or Hf3) from the left, test fungus from the right: (a) type A interaction, Cordyceps sp. 1 Fe842F, (b) type B interaction, Minimidochium sp. 1 (Hf1, Fe173F), (c–l) type C interaction: (c) Minimidochium sp. 1 (Hf3, Fe173F), (d) Fusarium sp. 1 Fe167F, (e) Pestalotiopsis sp. 1 Fe447K, (f) Clonostachys solani Fe26K, (g,h) Cyathus olla Fe141F (averse and reverse–dark zone), (i) Efibula sp. 1 Fe287F, (j) Gymnopus sp. 1 Fe1035F, (k) Polyporales sp. 1 Fe1037F, (l) Trichoderma atroviride Fe377E, (m–t) morphological deformations in H. fraxineus hyphae caused by test fungi (thin arrow–H. fraxineus, thick arrow–test fungus): (m) Trichoderma rodmanii Fe56K–cytoplasmic extrusion, internal hypha, (n) Efibula sp. 1 Fe287F–the deformation and disruption of significant sections of H. fraxineus hyphae from the melanized zone, (o) Clonostachys solani Fe888F–internal hypha, (p) Cordyceps sp. 1 Fe446K–local lysis and disintegration of pigmented H. fraxineus hypha, (q) Corticiales sp. 1 Fe18K–lysis and disintegration of pigmented H. fraxineus hypha, disappearance of the pigment, (r) Fusarium sp. 1 Fe410E–internal hypha, disintegration of pigmented H. fraxineus hypha, disappearance of the pigment, (s) Nemania diffusa Fe831E–internal hyphae, lysis and deformations of pigmented H. fraxineus hypha, disappearance of the pigment, and (t) Trichoderma harzianum Fe412E–disappearance of the pigment, the haustoria-like structures; scale bar (m–t) = 25 µm.
Figure 4.
Various types of interactions observed in dual cultures (a–l) and morphological deformations (m–t) (MEA, 3 weeks, 20 °C), Hymenoscyphus fraxineus (either Hf1, Hf2 or Hf3) from the left, test fungus from the right: (a) type A interaction, Cordyceps sp. 1 Fe842F, (b) type B interaction, Minimidochium sp. 1 (Hf1, Fe173F), (c–l) type C interaction: (c) Minimidochium sp. 1 (Hf3, Fe173F), (d) Fusarium sp. 1 Fe167F, (e) Pestalotiopsis sp. 1 Fe447K, (f) Clonostachys solani Fe26K, (g,h) Cyathus olla Fe141F (averse and reverse–dark zone), (i) Efibula sp. 1 Fe287F, (j) Gymnopus sp. 1 Fe1035F, (k) Polyporales sp. 1 Fe1037F, (l) Trichoderma atroviride Fe377E, (m–t) morphological deformations in H. fraxineus hyphae caused by test fungi (thin arrow–H. fraxineus, thick arrow–test fungus): (m) Trichoderma rodmanii Fe56K–cytoplasmic extrusion, internal hypha, (n) Efibula sp. 1 Fe287F–the deformation and disruption of significant sections of H. fraxineus hyphae from the melanized zone, (o) Clonostachys solani Fe888F–internal hypha, (p) Cordyceps sp. 1 Fe446K–local lysis and disintegration of pigmented H. fraxineus hypha, (q) Corticiales sp. 1 Fe18K–lysis and disintegration of pigmented H. fraxineus hypha, disappearance of the pigment, (r) Fusarium sp. 1 Fe410E–internal hypha, disintegration of pigmented H. fraxineus hypha, disappearance of the pigment, (s) Nemania diffusa Fe831E–internal hyphae, lysis and deformations of pigmented H. fraxineus hypha, disappearance of the pigment, and (t) Trichoderma harzianum Fe412E–disappearance of the pigment, the haustoria-like structures; scale bar (m–t) = 25 µm.

Figure 5.
Symptoms of colonization of pseudosclerotial plates and apothecia of H. fraxineus for up to three weeks after inoculation of ash petioles with test fungi: (a) Clonostachys rosea Fe213F, (b) Clonostachys rosea Fe886F, (c) Clonostachys solani Fe26K, (d) Cordyceps sp. 1 Fe446K, (e) Cordyceps sp. 1 Fe842F, (f) Corticiales sp. 1 Fe18K, (g) Cyathus olla Fe141F, (h) Cyathus olla Fe141F, (i) Efibula sp. 1 Fe287F–arrows show dead apothecia, (j) Gymnopus sp. 1 Fe1035F, (k) Minimidochium sp. 1 Fe173F, (l) Nemania diffusa Fe831E–arrow shows dead apothecium, (m) Fusarium sp. 1 Fm409E, (n) Pestalotiopsi sp. 1 Fe447K, (o) Polyporales sp. 1 Fe1011F, (p) Trichoderma atroviride Fe57K, (q) Trichoderma atroviride Fe377K–arrows show undeveloped and deformed apothecia, (r) Trichoderma harzianum Fe412E, (s) Trichoderma polysporum Fe166F, and (t) Trichoderma sp. 1 Fe462K; scale bar = 2 mm.
Figure 5.
Symptoms of colonization of pseudosclerotial plates and apothecia of H. fraxineus for up to three weeks after inoculation of ash petioles with test fungi: (a) Clonostachys rosea Fe213F, (b) Clonostachys rosea Fe886F, (c) Clonostachys solani Fe26K, (d) Cordyceps sp. 1 Fe446K, (e) Cordyceps sp. 1 Fe842F, (f) Corticiales sp. 1 Fe18K, (g) Cyathus olla Fe141F, (h) Cyathus olla Fe141F, (i) Efibula sp. 1 Fe287F–arrows show dead apothecia, (j) Gymnopus sp. 1 Fe1035F, (k) Minimidochium sp. 1 Fe173F, (l) Nemania diffusa Fe831E–arrow shows dead apothecium, (m) Fusarium sp. 1 Fm409E, (n) Pestalotiopsi sp. 1 Fe447K, (o) Polyporales sp. 1 Fe1011F, (p) Trichoderma atroviride Fe57K, (q) Trichoderma atroviride Fe377K–arrows show undeveloped and deformed apothecia, (r) Trichoderma harzianum Fe412E, (s) Trichoderma polysporum Fe166F, and (t) Trichoderma sp. 1 Fe462K; scale bar = 2 mm.


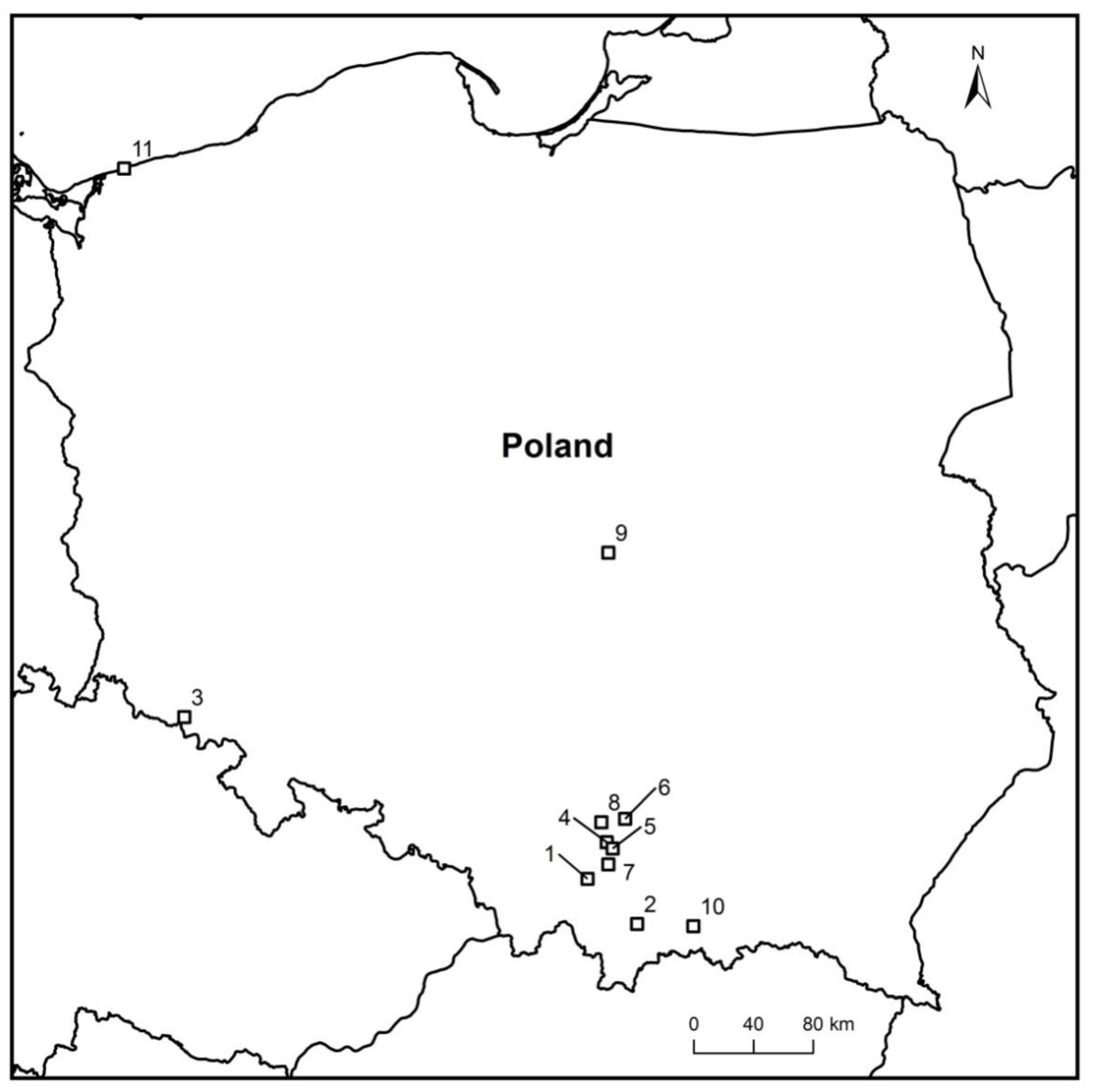{kind=link}
{kind=link}
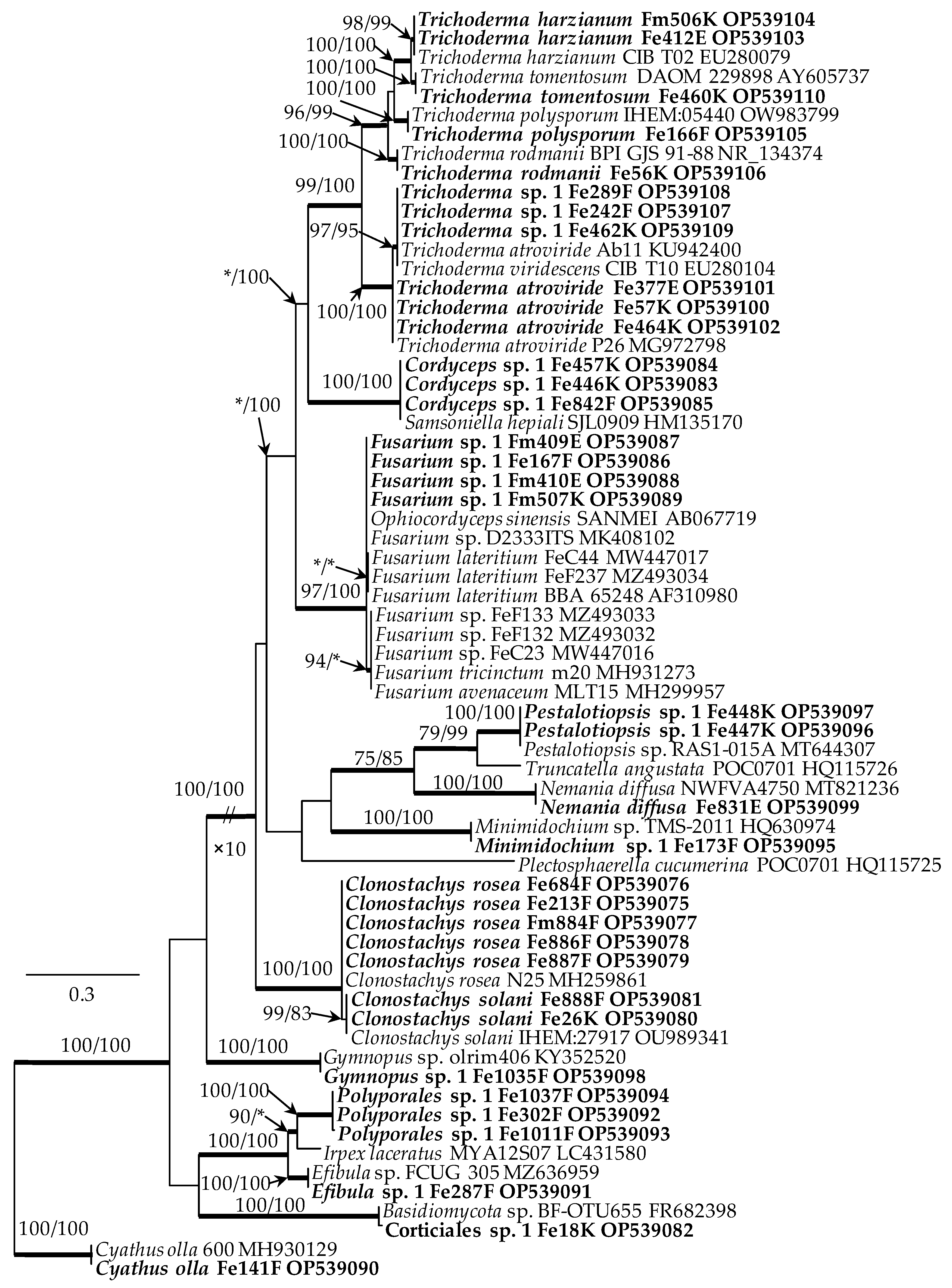{kind=link}
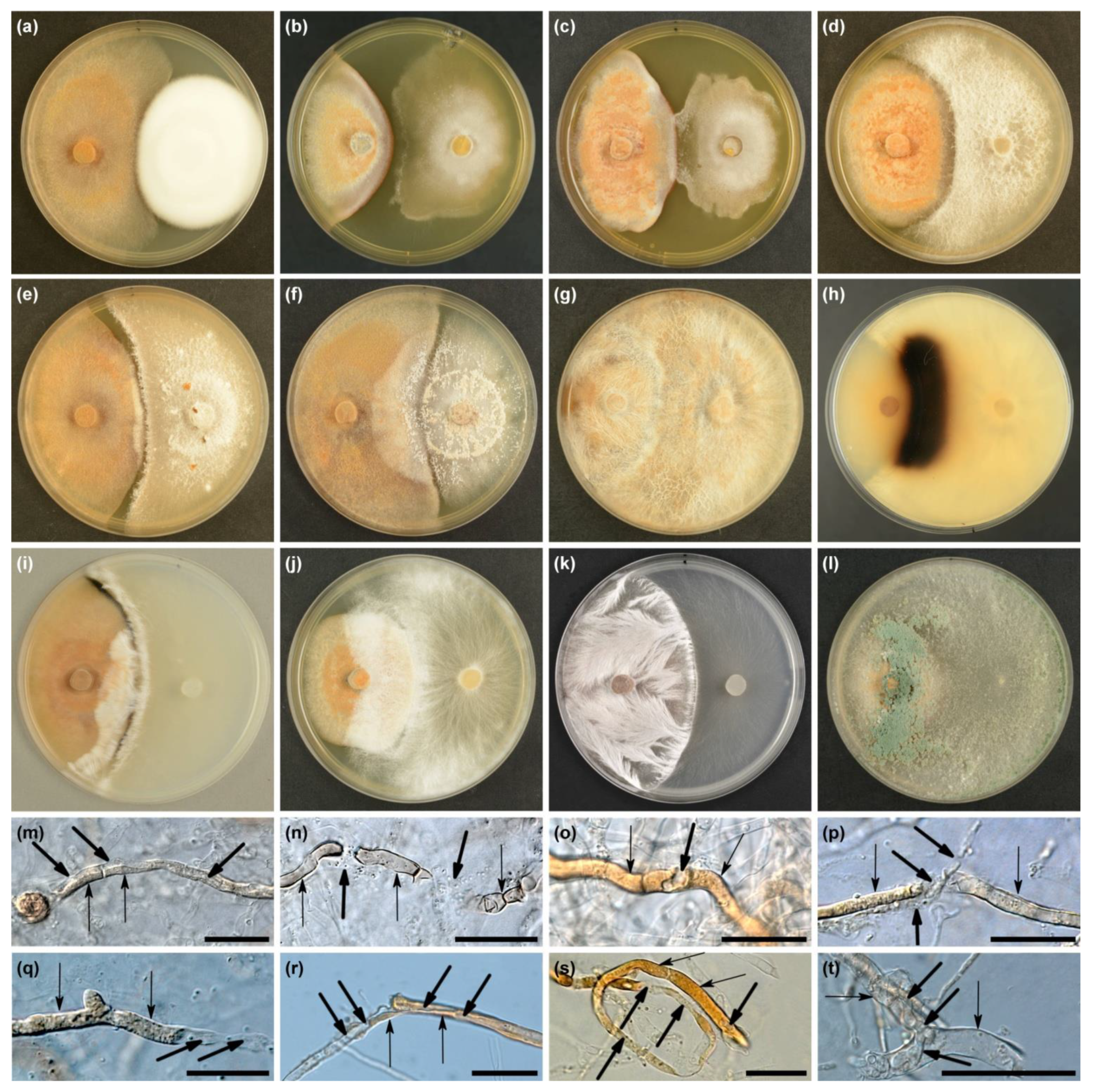{kind=link}
{kind=link}
Table 1.
Fungi isolated from apothecia and pseudosclerotial plates of Hymenoscyphus fraxineus with symptoms of colonization by other fungi. Sequences were deposited in GenBank. Reference sequences from GenBank were obtained using BLAST.
Table 1.
Fungi isolated from apothecia and pseudosclerotial plates of Hymenoscyphus fraxineus with symptoms of colonization by other fungi. Sequences were deposited in GenBank. Reference sequences from GenBank were obtained using BLAST.
| Fungi | Strain Number | Sub. 1 | Host 2 | Locality | Collection Date | ITS Accession | BLAST | Identities % |
|---|---|---|---|---|---|---|---|---|
| Clonostachys rosea | Fe213F | Pp | Fex | Ojców | 17 October 2017 | OP539075 | MH259861 | 99.83 |
| Fe684F | Pp | Fex | Miechów Domiarki | 12 September 2018 | OP539076 | MH259861 | 99.83 | |
| Fm884F | Ap | Fm | Rogów | 6 August 2016 | OP539077 | MH259861 | 99.83 | |
| Fe886F | Ap | Fex | Stary Sącz | 10 July 2014 | OP539078 | MH259861 | 99.83 | |
| Fe887F | Ap | Fex | Kowary | 6 August 2016 | OP539079 | MH259861 | 99.83 | |
| Clonostachys solani | Fe26K | Pp | Fex | Ojców | 26 September 2019 | OP539080 | OU989341 | 100 |
| Fe888F | Pp | Fex | Brody | 10 October 2018 | OP539081 | OU989341 | 100 | |
| Corticiales sp. 1 | Fe18K | Ap | Fex | Kowary | 6 August 2016 | OP539082 | FR682398 | 99.54 |
| Cordyceps sp. 1 | Fe446K | Pp | Fex | Miechów Domiarki | 28 August 2018 | OP539083 | HQ115725 | 100 |
| Fe457K | Pp | Fex | Miechów Domiarki | 4 October 2013 | OP539084 | HQ115726 | 100 | |
| Fe842F | Pp | Fex | Myślenice | 5 June 2019 | OP539085 | HM135170 | 100 | |
| Cyathus olla | Fe141F | Pp | Fex | Miechów Domiarki | 8 June 2017 | OP539090 | MH930129 | 99.87 |
| Efibula sp. 1 | Fe287F | Pp | Fex | Myślenice | 1 December 2017 | OP539091 | MZ636959 | 99.83 |
| Fusarium sp. 1 | Fe167F | Pp | Fex | Brody | 10 June 2017 | OP539086 | AB067719 | 100 |
| Fm409E | Ap | Fm | Rogów | 12 July 2016 | OP539087 | MK408102 | 100 | |
| Fm410E | Ap | Fm | Rogów | 12 July 2016 | OP539088 | MK408102 | 100 | |
| Fm507K | Ap | Fm | Rogów | 12 July 2016 | OP539089 | MK408102 | 100 | |
| Gymnopus sp. 1 | Fe1035F | Pp | Fex | Myślenice | 20 August 2019 | OP539098 | KY352520 | 100 |
| Minimidochium sp. 1 | Fe173F | Ap | Fex | Kraków Zakrzówek | 18 July 2017 | OP539095 | HQ630974 | 100 |
| Nemania diffusa | Fe831E | Pp | Fex | Kraków Młynówka | 12 May 2017 | OP539099 | MT821236 | 100 |
| Pestalotiopsis sp. 1 | Fe447K | Pp | Fex | Ojców | 07 May 2021 | OP539096 | MT644307 | 100 |
| Fe448K | Pp | Fex | Ojców | 07 May 2021 | OP539097 | MT644307 | 100 | |
| Polyporales sp. 1 | Fe302F | Pp | Fex | Miechów Domiarki | 20 October 2017 | OP539092 | LC431580 | 91.95 |
| Fe1011F | Pp | Fex | Myślenice | 2 August 2019 | OP539093 | LC431580 | 91.95 | |
| Fe1037F | Pp | Fex | Myślenice | 2 August 2019 | OP539094 | LC431580 | 91.95 | |
| Trichoderma atroviride | Fe57K | Pp | Fex | Brody | 10 May 2017 | OP539100 | MG972798 | 100 |
| Fe377E | Pp | Fex | Ojców | 4 May 2016 | OP539101 | MG972798 | 100 | |
| Fe464K | Pp | Fex | Ojców | 10 May 2017 | OP539102 | MG972798 | 100 | |
| Trichoderma harzianum | Fe412E | Ap | Fex | Gorce Konina | 27 August 2013 | OP539103 | EU280079 | 100 |
| Fm506K | Ap | Fm | Rogów | 12 July 2016 | OP539104 | EU280079 | 100 | |
| Trichoderma polysporum | Fe166F | Pp | Fex | Kraków Młynówka | 17 July 2016 | OP539105 | OW983799 | 99.84 |
| Trichoderma rodmanii | Fe56K | Ap | Fex | Trzęsacz | 18 August 2012 | OP539106 | NR_134374 | 100 |
| Trichoderma tomentosum | Fe460K | Pp | Fex | Kraków Młynówka | 17 July 2016 | OP539110 | AY605737 | 100 |
| Trichoderma sp. 1 | Fe242F | Pp | Fex | Miechów Domiarki | 5 May 2017 | OP539107 | KU942400 | 100 |
| Fe289F | Pp | Fex | Miechów Domiarki | 20 November 2017 | OP539108 | KU942400 | 100 | |
| Fe462K | Pp | Fex | Myślenice | 26 July 2019 | OP539109 | EU280104 | 100 |
1 Substrate: Ap—Apothecium with degrading symptoms, Pp—pseudosclerotial plate overgrown with hyphae or with clusters of conidiophores and conidia. 2 Host: Fex—Fraxinus excelsior, Fm—Fraxinus mandshurica.
Table 2.
Types of interactions of Hymenoscyphus fraxineus with test fungi in dual cultures in vitro and morphophysiological deformations in the hyphae of H. fraxineus.
Table 2.
Types of interactions of Hymenoscyphus fraxineus with test fungi in dual cultures in vitro and morphophysiological deformations in the hyphae of H. fraxineus.
| Fungal Species | Strain Number | H. fraxineus Strains Interaction Type 1 | Symptoms of Mycoparasitism, Deformations in H. fraxineus Hyphae 2 | ||||
|---|---|---|---|---|---|---|---|
| Hf1 | Hf2 | Hf3 | ME | MH | ML | ||
| Clonostachys rosea | Fe213F | C | C | Cs (B) | + | + | + |
| Fe684F | C | C | C | ||||
| Fm884F | C (B) | C | C | ||||
| Fe886F | C | C | C | ||||
| Fe887F | C | C | C | ||||
| Clonostachys solani | Fe26K | C | C | C | + | + | + |
| Fe888F | C | C | C | ||||
| Corticiales sp. 1 | Fe18K | Cs | Cs | Cs (B) | + | + | + |
| Cordyceps sp. 1 | Fe446K | Cs | Cs; Mz | Cs | + | + | + |
| Fe457K | A | A | A | – | – | – | |
| Fe842F | A | A | A | ||||
| Cyathus olla | Fe141F | C | C | C | + | + | + |
| Efibula sp. 1 | Fe287F | C (B); Mz | C (B); Mz | C (B) | + | + | + |
| Fusarium sp. 1 | Fe167F | Cs | Cs | Cs | + | + | + |
| Fm409E | Cs | Cs (B) | Cs (B) | ||||
| Fm410E | Cs | Cs (B) | Cs (B) | ||||
| Fm507K | Cs | Cs | Cs | ||||
| Gymnopus sp. 1 | Fe1035F | C | C | C | + | + | + |
| Minimidochium sp. 1 | Fe173F | B | Cs (B) | Cs (B) | + | – | + |
| Nemania diffusa | Fe831E | C | C | C (B) | + | + | + |
| Pestalotiopsis sp. 1 | Fe447K | Cs (B) | Cs (B) | Cs (B) | + | + | + |
| Fe448K | Cs (B) | Cs (B) | Cs (B) | ||||
| Polyporales sp. 1 | Fe302F | C (B) | C (B); Mz | C (B) | + | + | + |
| Fe1011F | C (B) | C (B) | C (B) | ||||
| Fe1037F | C (B) | C (B); Mz | C (B); Mz | ||||
| Trichoderma atroviride | Fe57K | C | C | C | + | + | + |
| Fe377E | C | C | C | ||||
| Fe464K | C | C (B) | C | ||||
| Trichoderma harzianum | Fe412E | C | C | C | + | + | + |
| Fm506K | C | C | C | ||||
| Trichoderma polysporum | Fe166F | C | C | C | + | + | – |
| Trichoderma rodmanii | Fe56K | C (B) | C (B) | C (B) | + | + | + |
| Trichoderma tomentosum | Fe460K | C | C | C | + | + | + |
| Trichoderma sp. 1 | Fe242F | C | C | C | + | + | – |
| Fe289F | C | C | C | ||||
| Fe462K | C | C | C | ||||
1 A—physical contact of mycelia; B—inhibition zone; C—overgrowth of H. fraxineus colony by test fungus; Cs—slow overgrowth, up to 1.5 cm from the edge of the H. fraxineus colony; C (B)—test fungus overgrew H. fraxineus colony but there was a very narrow inhibition zone; Mz—melanized blackish zone at the edge of H. fraxineus colony. 2 ME—apical and intercalar cytoplasmic extrusion; MH—internal hyphae of mycoparasite in hyphae of H. fraxineus, the deformation and disruption of significant sections of hyphae; ML—H. fraxineus: lysis of hyphae, desolation of hyphal cells, breakdown of hyphae into short fragments, the disappearance of the pigment.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kowalski, T.; Bilański, P. Fungicolous Fungi on Pseudosclerotial Plates and Apothecia of Hymenoscyphus fraxineus and Their Biocontrol Potential. Microorganisms 2022, 10, 2250. https://doi.org/10.3390/microorganisms10112250
AMA Style
Kowalski T, Bilański P. Fungicolous Fungi on Pseudosclerotial Plates and Apothecia of Hymenoscyphus fraxineus and Their Biocontrol Potential. Microorganisms. 2022; 10(11):2250. https://doi.org/10.3390/microorganisms10112250
Chicago/Turabian StyleKowalski, Tadeusz, and Piotr Bilański. 2022. "Fungicolous Fungi on Pseudosclerotial Plates and Apothecia of Hymenoscyphus fraxineus and Their Biocontrol Potential" Microorganisms 10, no. 11: 2250. https://doi.org/10.3390/microorganisms10112250
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.