
Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture


1
Centre for Environmental and Marine Studies (CESAM), Department of Biology, University of Aveiro, Campus Universitário de Santiago, 3810-193 Aveiro, Portugal
2
Division of Microbiology, Department of Pathology, Faculty of Medicine, University of Porto, 4200-319 Porto, Portugal
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(12), 2453; https://doi.org/10.3390/microorganisms10122453
Submission received: 17 November 2022
/
Revised: 6 December 2022
/
Accepted: 8 December 2022
/
Published: 12 December 2022
(This article belongs to the Special Issue Microorganisms as Biocontrol Agents in Plant Pathology)
Abstract
:Plant pathogens are responsible for causing economic and production losses in several crops worldwide, thus reducing the quality and quantity of agricultural supplies. To reduce the usage of chemically synthesized pesticides, strategies and approaches using microorganisms are being used in plant disease management. Most of the studies concerning plant-growth promotion and biological agents to control plant diseases are mainly focused on bacteria. In addition, a great portion of registered and commercialized biopesticides are bacterial-based products. Despite fungal endophytes having been identified as promising candidates for their use in biological control, it is of the utmost importance to develop and improve the existing knowledge on this research field. The genus Diaporthe, encompasses plant pathogens, saprobes and endophytes that have been screened for secondary metabolite, mainly due to their production of polyketides and a variety of unique bioactive metabolites with agronomic importance. Some of these metabolites exhibit antifungal and antibacterial activity for controlling plant pathogens, and phytotoxic activity for the development of potential mycoherbicides. Moreover, species of Diaporthe are reported as promising agents in the development of biofertilizers. For this reason, in this review we summarize the potential of Diaporthe species to produce natural products with application in agriculture and describe the benefits of these fungi to promote their host plant’s growth.
1. Introduction
In 1807, Bénédict Prévost found that germination of spores from Tilletia caries was inhibited by metallic copper when placed in the soil, thus describing it as the first compound with fungicidal properties [1]. The first organic fungicide was synthesized in the early 20th century. After that, several fungicides such as 2-methoxyethyl silicate and 2-hydroxyphenyl mercury, effective against the fungal species Fusarium spp. and Dreschlera spp., started also to be commercialized [2]. Nevertheless, the excessive use of agrochemicals has contributed to the environmental pollution (e.g., long degradation period), undesirable effects on human health (e.g., carcinogenicity) and the development of pathogen resistance [3]. Therefore, alternative methods for the safe control of plant pathogens and weed managements such as the use of biocontrol microorganisms and the application of naturally sourced metabolites have received increasing attention in the past decade [4,5].
Microorganisms are known for their ability to synthesize secondary metabolites, which exhibit promising bioactivities for the development of agrochemicals. Many natural antifungal fungicides have been obtained from microbial resources [6]. For instance, kasugamycin isolated from Streptomyces kasugaensis is widely used to control leaf spot, fire blight, rice blast and bacterial diseases in several crops [7]. The polyoxins, produced by Streptomyces cacaoi, are effective for rice fungal diseases as well as for the gray mold disease of fruits (Botrytis cinerea) [8]. Moreover, the antifungal antibiotic validamycin produced by Streptomyces hygroscopicus var. limoneus is commonly used to control sheath blight of rice plants caused by Rhizoctonia solani [9]. It is also well-stablished that several fungal genera may confer herbicidal activities by producing competent phytotoxins, such as species of the genus Colletotrichum and Xylaria [10].
The application of endophytic fungi to promote a sustainable agriculture has also been of interest, due to their role as plant-growth promoters [11]. This role is based on recognized mechanisms, such as the increase in nutrient and water acquisition and the production of plant hormones, leading to an increase in resistance to biotic and abiotic stresses [12]. Recent research has also demonstrated that the use of bacteria and fungi as biological control agents is advantageous to control plant diseases, thus improving agricultural yields [13]. The application of fungal biological control agents has largely increased due to their high reproductive rate (sexually and asexually), and their being target specific [14].
The genus Diaporthe comprises plant pathogens and endophytes, and it is a source of secondary metabolites. These have been explored for their potential applications in health care (e.g., antioxidant and anti-inflammatory properties), pharmacology (e.g., clinical toxicology assessment) and biomedicine (e.g., development of drugs) [15,16]. However, there is still a lack of information on the phytotoxins produced by species of Diaporthe, which should be explored given their potential application in agriculture as promising candidates for the development of natural herbicides [15]. Moreover, endophytic Diaporthe species are also reported as producing antimicrobial compounds to control plant pathogens, and as promising agents in the development of biofertilizers to promote plant growth [17]. Therefore, the main goal of this review is to summarize the potential benefits of species of Diaporthe as biocontrol agents, and as promising sources for the development of antimicrobials and mycoherbicides to assist in a sustainable agriculture.
2. Material and Methods
2.1. Criteria Used for Considering Studies
This review was aimed to summarize and gather current knowledge from published scientific data concerning the importance of endophytic species of Diaporthe as biological control agents. Notwithstanding the recognition of Diaporthe as the most common genera of endophytic fungi, this research field still requires up-to-date review papers. Considering this, the literature review was organized and compiled to deepen the knowledge and identify the possibility of using endophytic Diaporthe as crucial elements for the development of biopesticides and biofertilizers to assist in a sustainable agriculture. The importance and suitability of multi-omics approaches on species of Diaporthe was also a key point discussed in this review. Multi-omics are important tools for unraveling functions and beneficial properties of endophytes and their metabolites. Moreover, such approaches are also crucial to unveil metabolic pathways towards plant growth and tolerance to environmental stresses. The assemblage of all published data covered in this review represents a step closer to sustainable and ecological agricultural production.
2.2. Search Strategy for Identification of Studies
The literature review was carried out mainly in Web of Science, Scopus and Google Scholar databases between 10 August 2022 to 15 October 2022. The main keywords used for this review were used alone or in combination as follows: endophyte, endophytic, Diaporthe, Phomopsis, omics, genomics, transcriptomics, metabolomics, proteomics, fungi, fungus, biocontrol, antagonism, microorganisms, biofertilizers, plant-microbe interactions, biopesticides, sustainable agriculture, agrochemicals, environment, phytotoxins and plant promoters. Only articles, reviews, book chapters and books written in the English language were included. Articles were also manually screened for additional references.
All scientific literature was taken into consideration with a special focus on publications from the last decade (2012–2022), which represents more than 77% of the references used (136 out of 176). However, to show evidence of the historical background of some topics covered in this review, some publications prior to the 2000s, dated between 1874 and 1999, were also selected which correspond to 7% of all references used (12 out of 176).
3. Fungal Endophytes and Their Benefits for Plants
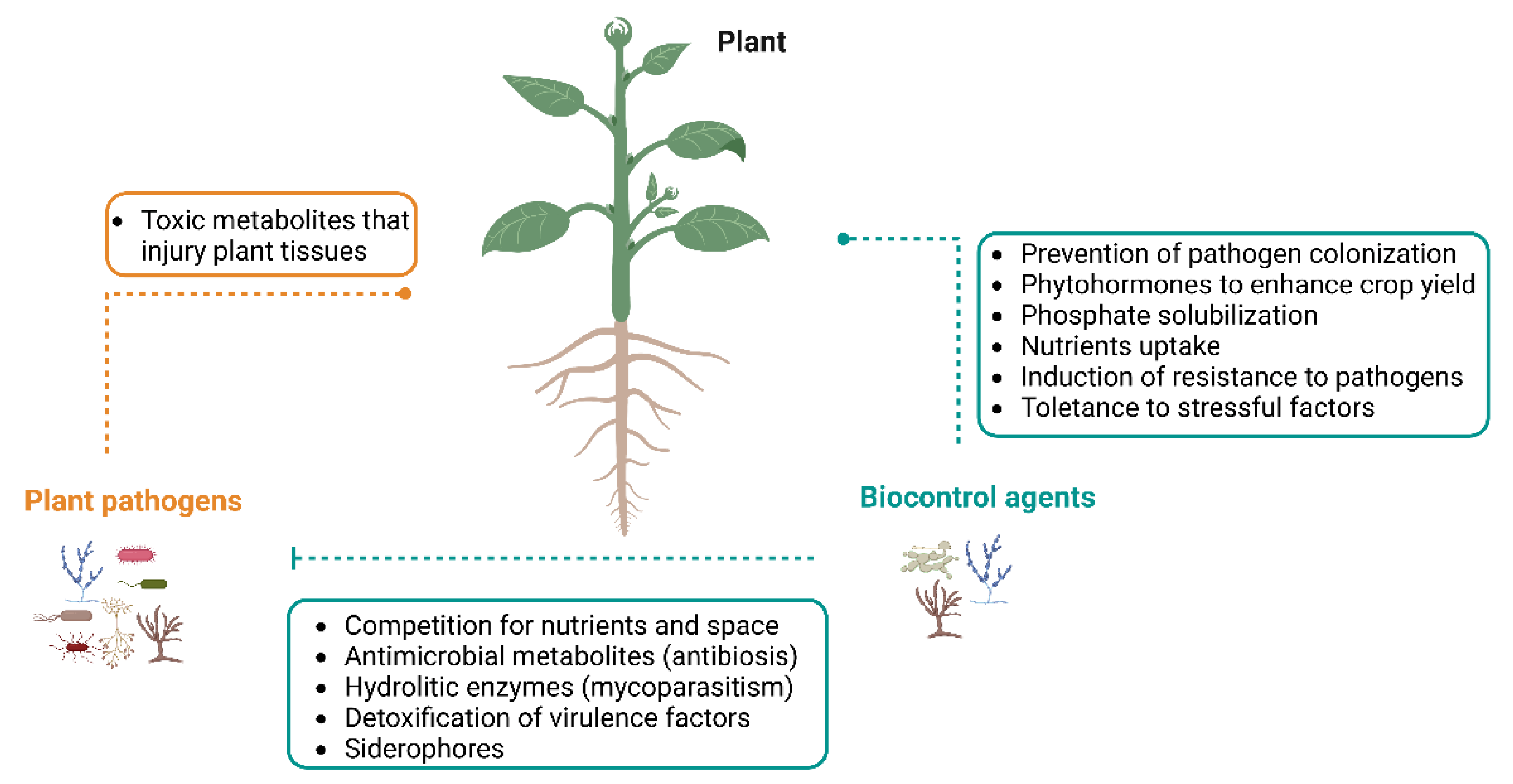
Although the term “endophyte” was originally introduced by de Bary in 1866, the most used definition of endophytes was proposed by Petrini in 1991 [18]. It refers to a group of organisms “inhabiting plant organs that at some time in their life can colonize internal plant tissues without causing apparent harm to the host” [19]. These endophytes are usually fungi or bacteria that are present in the phyllosphere, endosphere or rhizosphere. These microorganisms live in the tissues of plants without causing any symptoms of disease, leading to beneficial effects for the hosts (Figure 1) by:
In this regard, some fungal species have been studied due to their ability to promote plant growth. For instance, Fusarium equiseti increased the herbage yield of Trifolium subterraneum (subclover) by facilitating nitrogen uptake, while Sporormiella intermedia increased the mineral uptake of calcium, copper and zinc in subclover, thus enhancing the nutritional value of forage [23]. Similarly, Baron et al. [24] used Aspergillus sydowii to inoculate maize plants (Zea mays). The authors stated that those plants inoculated with the fungus accumulated significantly higher amounts of phosphorous in their tissues. The endophyte Colletotrichum tropicale can also enhance the nitrogen uptake and change its distribution in cacao plants [25]. Trichoderma asperellum was also reported to significantly increase seed vigor and the yield of Sorghum bicolor roots [26].
Moreover, Khan et al. [27] detected gibberellin production by Penicillium citrinum. These authors have thus demonstrated that the P. citrinum improved the length of seedlings in the sandy plant Atriplex gmelinii, thus promoting its growth. Baron et al. [28] also demonstrated that the fungal species Purpureocillium lilacinum, P. lavendulum and Metarhizium marquandii are able to produce indole acetic acid and to solubilize phosphorous. The authors showed that these strains were able to promote the availability of phosphorous and nitrogen in soybean, bean and maize plants. In another study, Ismail et al. 2020 [29] reported that soybean plants inoculated with the endophyte Aspergillus niger showed tolerance to high temperatures. The inoculation with this fungal endophyte promoted and increased plant height, biomass and chlorophyll content, as well as to reduced lipid peroxidation during heat stress [29].
The intensive use of chemical fungicides to suppress the growth of plant pathogens over a long period have led to pesticide-related pollution, resistant microbial strains, chemical consumption through bioaccumulation, biodiversity losses and the elimination of natural/beneficial microorganisms [30]. Considering that the most current strategies contained in the United Nations 2030 Agenda (17 Sustainable Development Goals) aim at achieving sustainable development, the biopesticides application creates an equilibrium between economic productivity and environmental protection that is crucial to sustainable agriculture [30,31]. In this regard, the growing search for new biopesticides to replace synthetic chemicals is supported by its low toxicity properties, eco-friendliness, specificity, biodegradability, low post-harvest contamination and compatibility in integrated pest management [32].
The drawbacks of biopesticides usage are defined as the high cost of commercial products, standard method of preparations and dose determination of active substances [32]. Nevertheless, the application of antagonistic endophytic fungi as biocontrol agents, has drawn special attention for being a sustainable option for the management of some plant diseases, thus resulting in minimal impact on the environment [4,17,33]. The main interaction between endophytic fungi and pathogens is the limitation of mycelium growth by contact, or through the formation of inhibition zones in dual culture [34]. Such facts indicate that the endophytes that act as biocontrol agents harbor multiple mechanisms to control the pathogens (Figure 1) by:
Since early times, man has attempted to increase and improve crop production and to control plant diseases by using antagonistic microorganisms [39]. For instance, Roberts, in 1874 [40], introduced the term antagonism in microbiology after showing the antagonistic action between the fungus Penicillium glaucum and a bacterial strain. Later in 1921, Hartley inoculated forest nursery soils with antagonistic fungi to control damping-off caused by Pythium debaryanum [41]. In 1941, Weindling [42] noted that species of Trichoderma produced an antifungal compound, the gliotoxin, that was toxic to plant pathogens including Rhizoctonia solani and Sclerotinia americana. This study conducted by Weindling [42] was the first to record the use of gliotoxin in plant disease control [43]. Since the discovery of penicillin by Alexander Fleming in 1928 with pharmaceutical application, the studies on the discovery of biological control agents against plant pathogens have been increasing, attempting to unveil secondary metabolites with promising applications in agriculture [14,44].
It is noteworthy that endophytic fungi produce large numbers of metabolites with different chemical structures from, including alkaloids, terpenoids, benzopyranones or quinones [45]. These compounds are crucial for agricultural application once they exhibit promising bioactivities such as antifungal, antibacterial, herbicidal and other agricultural activities [3,16]. For instance, the fungal genus Xylaria associated with the Azadirachta indica plant produces antifungal compounds with activities against Aspergillus niger and Fusarium avenaceum [46,47]. Sangeetha et al. [48] demonstrated that species of Trichoderma may produce antifungal compounds due to their biocontrol potential against Colletotrichum musae, Fusarium verticillioides and Lasiodiplodia theobromae (causing postharvest crown rot of banana). Griseofulvin, a secondary metabolite initially isolated from the fungus Penicillium griseofulvum, has drawn special attention due to many reports of antifungal activities against plant pathogenic fungi such as Cytospora sp., Cladosporium gloeosporioides, Botrytis cinerea, Alternaria solani and Fusarium solani [49]. Therefore, endophytic fungi are promising leads for the discovery of novel secondary metabolites with potential for agricultural applications as biocontrol agents, biostimulants, biofertilizers and bioherbicides [3,4,13,50].
4. Species of Diaporthe as Benefit Microorganisms to Agriculture
4.1. Production of Antimicrobial Compounds
Species of the genus Diaporthe can switch between lifestyles, meaning that the same species can be found on the same or other hosts as phytopathogens or as endophytes in asymptomatic tissues [51,52]. For example, D. eres is a pathogen that is present on a wide range of hosts, including economically important fruit trees (e.g., apple, blueberry, hazelnut) [53,54,55] and ornamental plants (e.g., Allium giganteum, Magnolia soulangeana) [56], and as an endophyte on Prunus domestica [17]. Moreover, D. limonicola can be found on Citrus grandis leaves as an endophyte [57], or as a pathogen causing dieback on lemon trees in Europe [58].
Despite its known record as a plant pathogen, Diaporthe is recognized as one of the most frequently isolated genera occurring as endophytes in the stems and leaves of several hosts in tropical and temperate ecosystems [55,56,59]. Due to the high number of species of Diaporthe as endophytes, and given their potential as producers of secondary metabolites, these species have been widely investigated for the production of valuable compounds with different bioactivities [17]. A recent review by Xu et al. [16] summarized a total of 335 bioactive secondary metabolites isolated from species of Diaporthe and Phomopsis-like species. These metabolites were classified into polyketides, terpenoids, steroids, macrolides, ten-membered lactones, alkaloids, flavonoids and fatty acids. Polyketides are the main chemical population (64%), and their bioactivities involve antitumor (e.g., clavaric acid), antioxidant (e.g., pyranonigrin E; diportharine A; phochrodine D) [60,61], cytotoxic (e.g., diaporthelactone, phomopsidone A, phomaspyrone A-E) [62,63], anti-bacterial (e.g., phomosine A, 3-Hydroxypropionic acid) [64,65], anti-fungal (e.g., (+)-2,2′-Epicytoskyrin A, phomopsolide A-C) [66,67,68], antimalarial (e.g., epoxycytochalasin H) [69] and anti-inflammatory activities (e.g., Biatriosporin N) [70]. Considering the several compounds produced by species of the genus Diaporthe, Xu et al. [16] have stated that this genus is a promising source for the discovery of small molecules for drug candidates.
In agriculture, several studies have reported that species of the genus Diaporthe (including Phomopsis-like species) also exhibit antibacterial and antifungal activity against plant pathogenic microorganisms (Table 1). For instance, Abramczyk et al. [17] showed that D. eres from Prunus dulcis exhibited antifungal activity against Trichothecium roseum, F. avenaceum and A. alternata. Moreover, Endophytic species of Diaporthe, isolated from Pachystachys lutea, were effective against F. oxysporum and Colletotrichum sp. [34]. The antifungal activity of D. citri, isolated from Mikania glomerata, was also verified against F. solani and Didymella bryoniae [71]. Verma et al. [72] also demonstrated that under dual culture techniques, D. melonis and D. longicolla showed antagonism to Corynespora cassiicola and F. solani with an inhibition halo percentage ranging from 52–64% against C. cassiicola and 28–52% against F. solani.
Moreover, two derivatives of the phytotoxin alternariol, alternariol 4,10-dimethyl ether and alternariol methyl ether, were isolated from Diaporthe phragmitis (syn. D. eres), a kiwi endophytic fungus [73]. The authors showed that this endophytic fungus exhibited antibacterial activity against Pseudomonas syringae pv. actinidiae which causes kiwi cankers. Carvalho et al. [74] have also shown that seven strains of D. miriciae and two strains of a Diaporthe sp. produced cytochalasins H and J. After a bioassay-directed fractionation to isolate cytochalasins, the authors evaluated the compounds for activities against the fungal plant pathogens Colletotrichum fragariae, Botrytis cinerea, F. oxysporum, Phomopsis obscurans (syn. Paraphomopsis obscurans) and Phomopsis viticola (syn. D. rudis). The cytochalasins H and J exhibited effective activities against Paraphomopsis obscurans, associated with strawberry leaf blight and D. rudis, a grapevine pathogen [74]. Therefore, these studies suggest that Diaporthe fungal endophytes could be used as biocontrol agents and offer insights for the screening and isolation of antimicrobial compounds for the further development of new agrochemicals.

{kind=link}
{kind=link}
Table 1.
Antimicrobial activity of Diaporthe against fungal and bacterial plant pathogens.
| Host Plant | Endophyte | Antimicrobial Activity | References |
|---|---|---|---|
| Aconitum carmichaelii | Diaporthe sp. | Antifungal activity against the rice blast fungus Magnaporthe oryzae | [75] |
| Aconitum carmichaeli | D. amygdali | Antifungal activity against plant pathogenic fungi: Fusarium graminearum, Verticillium albo-atrum and Microdochium nivale | [76] |
| Actinidia chinensis | D. phragmatis (syn. D. eres) | Inhibitory activity against Pseudomonas syringae pv. actinidiae, the causal agent of kiwi canker disease | [73] |
| Balanophora polyandra | D. foeniculina | Antibacterial potential against plant pathogenic bacteria: Ralstonia solanacearum, Pseudomonas lachrymans and Xanthomonas vesicatoria | [77] |
| Cistus salvifolius | Diaporthe sp. | Antifungal activity against pathogens of agricultural importance: Phytophthora infestans, Botrytis cinerea and Septoria tritici | [78] |
| Copaifera pubiflora and Melocactus ernestii | D. miriciae | Antifungal activity against the grapevine pathogen D. rudis | [74] |
| Endodesmia calophylloides | Diaporthe sp. | Inhibition of zoospores of grapevine pathogen Plasmopara viticola | [79] |
| Espeletia sp. | D. phaseolorum | Antifungal activity against the plant pathogen Phytophthora infestans | [80] |
| Gossypium hirsutum and G. arboreum | D. longicolla and D. melonis | Antifungal activity against P. citricarpa | [72] |
| Gossypium hirsutum | Phomopsis-like species | Antifungal activity against Sclerotinia sclerotiorum, F. oxysporum, B. cinerea, Bipolaris sorokiniana, Gaeumannomyces graminis var. tritici, and Rhizoctonia cerealis | [81] |
| Maytenus ilicifolia | D. endophytica | Antifungal activity against the citrus pathogen Phyllosticta citricarpa | [82,83] |
| Mikania glomerata | D. citri | Antifungal activity against F. solani and Didymella bryoniae | [71] |
| Pachystachys lutea | Diaporthe sp. | Antifungal activity against the pathogenic fungi F. oxysporum and Colletotrichum sp. | [34] |
| Prunus domestica | D. eres | Antifungal activity against plant pathogenic fungi: Trichothecium roseum, F. avenaceum and Alternaria alternata | [17] |
| Rhizophora mucronata | Diaporthe sp. | Antifungal activity against pathogens of agricultural importance: Verticillium dahlia, Botrytis cinerea and Sclerotinia sclerotiorum | [84] |
| Schinus terebinthifolius | D. terebinthifolii | Antifungal activity against the citrus pathogen Phyllosticta citricarpa | [82,83] |
| Solanum lycopersicum | D. phaseolorum | Inhibitory activity against bacterial spot of tomato (Xanthomonas vesicatoria) | [85] |
| Vochysia divergens and Stryphnodendron adstringens | Diaporthe cf. heveae | Antifungal activity against P. citricarpa and Colletotrichum abscissum | [86] |
4.2. Phytotoxins as Potential Mycoherbicides
Weeds hamper the growth of several crops once they compete with the plants for water and nutrients resulting in enormous production losses [87]. Moreover, weeds can act as a host for insects and pathogens (fungi and bacteria), which can cause serious damage to crop plants [88]. Therefore, weeds management is a crucial agricultural practice to avoid significant yield losses [89]. However, chemical herbicides can have negative side effects, such as surface and ground water contamination, leaving herbicide residues in the food chain and, decreasing the soil microbial communities and earthworm populations, thus suppressing nutrient availability and soil biodiversity. [87,90]. Therefore, the use of herbicides should be minimal and effective to reduce their impact on human health and the environment. Mitigation of herbicides should be considered an important achievement for a sustainable agriculture. [90]. Such facts drive a growing search for new herbicides with low toxicity profiles, which is a step closer for human safety and environmental health [87,91]. One important aspect to be highlighted is that bioherbicides do not need to cause the death of weeds to increase crop productivity; they can suppress weed populations, which is a strategy with low risk when compared with the application of chemical herbicides [92].
Among the several microorganisms producing phytotoxins, some of them exhibit a potential for the production of molecules with herbicidal activity. Accumulating evidence on molecular biology and natural products demonstrates that many fungal species are a promising source of natural phytotoxins (e.g., Alternaria, Colletotrichum, Chondrostereum, Lasiodiplodia, Sclerotinia, Xylaria) [10,93]. For example, phytotoxic compounds isolated from the endophytic Xylaria feejeensis exhibited a toxic effect on the photosynthesis machinery of spinach chloroplasts [94].
Species of the genus Diaporthe (including Phomopsis-like species) are also producers of phytotoxins, showing interesting results (Table 2) [92,95,96,97,98,99]. For instance, Cimmino et al. [100] tested the fungal phytotoxin phomentrioloxin produced by D. gulyae and verified a suppression in the growth of the annual weed Carthamus lanatus. Brun et al. [92] have also demonstrated that biomolecules from D. schini caused yellowing lesions and a decrease in the height of the grass species Echinochloa crusgalli and Lolium multiflorum. Kongiidiazadione, isolated from D. kongii, showed a phytotoxic activity on the leaves of tomato plants but caused clear and significant necrosis on H. annuus [101]. Additionally, an endophytic Phomopsis sp. (Diaporthe) was reported by Yang et al. [102] as the producer of phytotoxins such as cytochalasins (H, N, and epoxycytochalasin H) and a nonenolide compound ((6S,7R,9R)6,7-Dihydroxy-9-propylnon-4-eno-9-lactone). The authors demonstrated that the above-mentioned compounds showed phytotoxic effects on Medicago sativa, Trifolium hybridum and Buchloe dactyloides by decreasing the germination and radicle growth [102]. Such examples are evidence that species of Diaporthe are a source of phytotoxic compounds. Therefore, it is crucial to analyze, screen and isolate herbicidal compounds from these species to be applied in the development of new mycoherbicides.
4.3. Plant-Growth Promoters
Chemical fertilizers are used to boost agricultural productivity but can cause negative environmental impacts [115]. In addition, environmental stresses and climate changes scenarios (e.g., drought, high soil salinity) are major limitations to plant growth and yield, which can cause production losses [116,117]. For this reason, to counteract potential losses, endophytic fungi are an alternative to increase agricultural productivity. Endophytic microorganisms, which inhabit plant tissues, are capable of increasing agricultural productivity by increasing access to nutrients (e.g., nitrogen, phosphorus, potassium, zinc, iron), production of phytohormones or by increasing the water acquisition rates [115,118]. Among endophytic fungi used as biofertilizers, Epichloë bromicola, for instance, increases seed germination and growth capacity of wild barley (Hordeum brevisubulatum) when exposed to salinity stress [6]. Additionally, Piriformospora indica, an endophyte with high economic importance, are able to enhance nutrient uptake and to modulate the phytohormones involved in the growth and development of Hordeum vulgare [119].
Species of the genus Diaporthe are also well documented promising agents in the development of biofertilizers due to their plant-growth properties (Table 3). For example, the generalist fungus Phomopsis liquidambaris (syn. D. liquidambaris) is also able to establish symbiosis with rice and peanut plants by colonizing their roots and conferring tolerance to abiotic stress, as well as promoting plant host growth [120,121]. This species is also reported to induce rice resistance to Fusarium graminearum, the causal agent of rice spikelet rot disease [122]. Recently, Aldana et al. [123] reported that root and shoot biomass of the hybrid Triticum durum × Hordeum chilense increased by up to 30% after inoculation with a strain of Diaporthe sp. The authors have also shown increased concentrations of calcium, magnesium, sulphur, iron and boron in inoculated plants. Additionally, the inoculation of Triticum durum × Hordeum chilense with Diaporthe sp. strain EB4 under salinity conditions, heightened proline, gibberellins, and indole 3-acetic acid and increased nutrient uptake in roots, thus resulting in an enhanced growth [124]. Moreover, Ważny et al. [125] stated that the inoculation of Noccaea goesingensis with Diaporthe eres improved the biomass of the plant and increased nickel accumulation. The authors have thus proposed D. eres as a nickel uptake stimulating microorganism, which might be potentially used as a biofertilizer and in the bioremediation of metal contaminated soils.
5. Omics to Explore the Secondary Metabolism of Diaporthe
5.1. Genomics
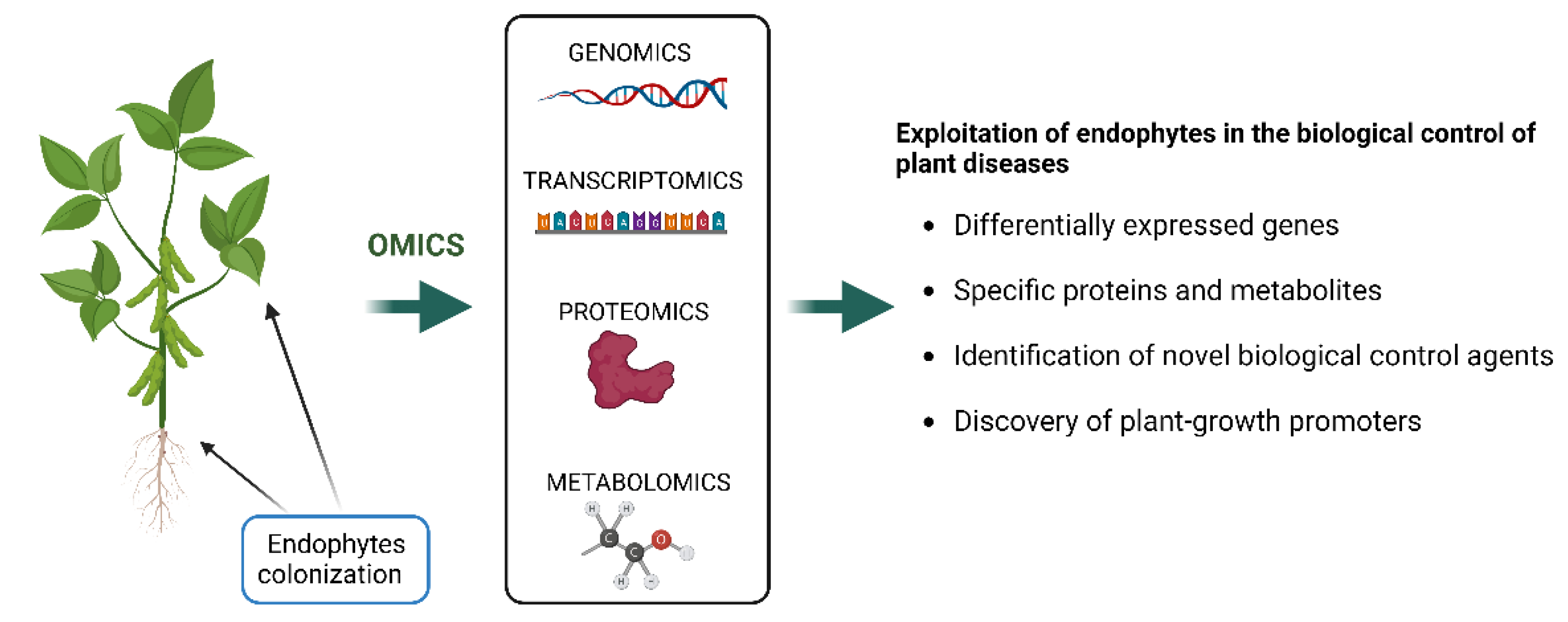
The application of omics approaches including genomics, transcriptomics, proteomics and metabolomics, opens up a new opportunity for the discovery of novel genes and their functions, novel pathways and metabolic network [134] (Figure 2).
Next-generation sequencing has been widely used to identify and characterize genes involved in plant–endophyte interactions [135]. Genome analysis has revealed genes responsible for nitrogen fixation, nutrition acquisition, and hormones biosynthesis as well as has elucidated the adaptative genomic signatures for bioactive secondary metabolites [136]. Recent genome mining has also offered in-depth information to search for natural products from fungi, and their biosynthetic gene clusters (BGCs) involved in different biosynthetic pathways [137,138]. Since 2011, researchers have used the “antibiotics and secondary metabolite analysis shell–antiSMASH” for their microbial genome mining tasks [139]. In fungi, less than 3% of the biosynthetic space of fungal genomes has been linked to the production of secondary metabolites, which are encoded by BGCs. The most common BGCs include non-ribosomal peptide (NRPSs), polyketide (PKSs) and terpene synthases [140]. However, the number of genomes available from the genus Diaporthe hampers the identification of BGCs and thus the discovery of bioactive compounds from these species. A search at NCBI (National Center for Biotechnology Information) (https://www.ncbi.nlm.nih.gov/, accessed on 19 June 2022) and JGI Genome Portal (Joint Genome Institute, Berkeley, CA, USA) (https://genome.jgi.doe.gov/portal/, accessed on 19 June 2022), unveiled 20 species of Diaporthe that have completed or are undergoing genome projects.
In this review, species with genomes available at the NCBI database were screened for BGCs (Table 4) and we identified some important compounds with 100% similarity with known BGCs with potential agricultural application The BGCs were predicted using the web-based application antiSMASH v.5.0, using the strictness ‘relaxed’ option [140].
Alternariol is classified as a phytotoxic and antifungal compound produced by species of the genus Alternaria, an important contaminant in cereals [141]. Nevertheless, its complete BGC has been detected in the genomes of D. amygdali, D. destruens, D. capsici, D. citri, D. citrichinensis, D. eres (syn. D. phragmitis) and D. eres (syn. D. vaccinii). This suggests the ability of these species to efficiently produce alternariol, and consequently their status as promising sources for the development of biopesticides. Moreover, (-)-Mellein is a phenolic compound initially isolated from Aspergillus melleus that showed antimicrobial activity [142]. Therefore, it is suggested that this compound could be screened and isolated from species of Diaporthe and produced to inhibit the growth of competitors and thus take on an agricultural application.
Fusicoccin A is a remarkable phytotoxin produced by the fungal species Phomopsis amygdali (syn. D. amygdali) [109,130]. However, it has been shown that it can be more effective than the growth-promoting hormone auxin [131]. Balio et al. [143] were prompted to investigate the effect of Fusicoccin in plants and carried out a pilot-scale study for the production of cultures of the fungus [143]. The quantities of the toxin available unveiled the physiological effects enhanced by Fusicoccin A such as an increase in tissue growth, nutrient uptake and the breaking seed coat dormancy [143,144]. Considering that the BGC that encode for this toxin was detected in the genome of D. amygdali with 100% similarity (Table 4), we can suggest that besides its reported phytotoxic activity, Fusicoccin can be used as a plant-growth promoter [15,130].
Asperlactone belongs the methylsalicylic acid (MSA) type polyketide group and has strong antibacterial and antifungal activities [145,146]. It has been reported that asperlactone also presented ovicidal activities [147] against Nezara viridula, a threatening pest for agriculture [148], and Tribolium castaneum, a significant global pest of stored food [149]. Considering that D. longicolla contains the complete BGC that encode for asperlactone, it is suggested that this species is able to produce this compound and should thus be regarded as a promising source for the development of novel bioinsecticides.
However, the presence of the complete BGC involved in the biosynthesis of one particular compound does not necessarily imply the production of that compound. Mainly because sometimes BGCs may remain silent under laboratory conditions [150]. Therefore, to bridge this flaw, altering cultivation parameters or adding chemical elicitors to activate silent BGCs can elucidate the hidden reservoir of complex chemical diversity. Such an outcome is readily achieved by the one strain many compounds (OSMAC) approach [151,152]. For instance, Zhang et al. [150] used the OSMAC approach on Aspergillus fumigatus LN-4 and Xylaria sp. XC-1 for the isolation of anticancer compounds and phytotoxins, respectively, by altering the cultivation parameters.
Table 4.
Some secondary compounds produced by species of Diaporthe which are 100% identical to known BGCs.
Table 4.
Some secondary compounds produced by species of Diaporthe which are 100% identical to known BGCs.
| Compound | Compound Nature | Biological Function | Species | References |
|---|---|---|---|---|
| ACR-Toxin I | Polyketide | Phytotoxin produced by the plant pathogenic fungus A. alternata, causing lemon leaf spot disease. | D. ampelina D. helianthi | [153,154] |
| ACT-Toxin II | Polyketide | Toxin causing brown spot disease on tangerine, produced by A. alternata | D. eres D. capsici D. citrisiana D. vaccinii (syn. D. eres) | [155] |
| Alternariol | Polyketide | Metabolite produced by Alternaria spp. that exhibits both phytotoxic and antifungal activity (e.g., zoosporicidal potential) | D. amygdali D. destruens D. capsici D. citri D. citrichinensis D. phragmitis (syn. D. eres) D. vaccinii (syn. D. eres) | [141] |
| Asperlactone | Polyketide | Methylsalicylic acid produced by Aspergillus westerdijkiae, antibacterial, antifungal and insecticidal activities | D. longicolla | [145,146] |
| Enniatin | Non-ribosomal peptide | Toxin produced by Fusarium spp., as a contaminant in cereals. It is also known as antibacterial, antifungal, and herbicidal. | D. citrichinensis | [156,157] |
| Fusarin | Polyketide | Mycotoxin produced mainly by fungi of the genus Fusarium, which can infect agriculturally important crops. | D. amygdali D. aspalathi D. helianthi | [158,159] |
| Fusicoccin | Terpene | Phytotoxin produced by D. amygdali, with plant-growth promoting potential | D. amygdali | [131] |
| (-)-Mellein | Phenolic compound | Metabolite firstly isolated from Alternaria melleus, with phytotoxic and antimicrobial activity | D. capsici D. citri D. citrichinensis D. destruens D. phragmitis (syn. D. eres) D. vaccinii (syn. D. eres) D. longicolla | [142] |
Besides the presence of BGCs that have 100% homology with known compounds, it also detected some incomplete BGCs. For instance, on D. amygdali CAA958 and D. eres (syn. D. vaccinii) CBS 160.32 genomes, the BGC encoding for betaenones (phytotoxic polyketides) was detected. By analyzing the BGCs, we found that both species contain four genes encoding for enzymes involved in betaenone synthesis but were lacking a dehydrogenase and a FAD-dependent oxidase [160].
The BGC encoding for chaetoglobosin X, a hybrid polyketide with antifungal activity, was detected on the genome of D. citrichinensis ZJUD34. From the 11 genes involved in the biosynthesis of this compound, only 7 genes were detected such as PN3-12 (C6 zinc finger protein), PN3-13 (hybrid NRPS/PKS), PN3-14 (enoyl reductase), PN3-15 (hypothetical protein), PN3-16 (FAD-linked oxidoreductase), PN3-17 (P450 monooxygenase) and PN3-19 (P450 oxygenase). Given that four of these genes present in D. citrichinensis are identified as being involved in the biosynthesis of chaetoglobosins [161], it is suggested that this species may have the potential to produce a related compound.
Nevertheless, on both betaenone and chaetoglobosin BGCs, the absent genes needed to complement the incomplete cluster may be silent or truncated, or even located on different fragmented contigs. The absence of those necessary genes for the core cluster suggests that many BGCs may not be functional and thus require additional experimental validation [162].
5.2. Transcriptomics
While genomics provides the whole genomic sequencing information, transcriptomics identifies the genes with changed expressions under a particular condition. It also involves the comparative analysis of the transcriptomes of groups and aids to understand the response of microbial communities upon changing environments [135,163].
The regulation of transcripts can be achieved by transcriptomic analysis. For example, mRNA sequencing is a valuable approach to understand differences in plants’ response in the presence and absence of endophytes [135]. In this regard, Ważny et al. [125] compared the transcriptomes of the hyperaccumulating plants Noccaea caerulescens and N. goesingensis, inoculated with Phomopsis columnaris (syn. D. eres), with uninoculated controls. The authors showed that the presence of the fungus enhanced uptake and accumulation of nickel, as well as that several genes involved in plant stress protection and metal uptake were upregulated.
Moreover, to understand the mechanisms of plant colonization by Phomopsis liquidambaris (syn. D. liquidambaris) under low nitrogen conditions, Zhou et al. [122] performed a transcriptomic analysis by the RNA-Seq technique. The authors compared the transcriptome profiles of inoculated and non-inoculated rice plants, and observed that gibberellin and auxin related genes, as well as genes encoding plant defense-related endopeptidase inhibitors, were upregulated on inoculated plants. Such results using transcriptomics might be useful to provide a better understanding regarding the molecular mechanisms of plant-endophyte interaction, as well as a deeper knowledge of fungal endophytes that promote plant growth under different conditions.
5.3. Proteomics
Nowadays, mass spectrometry (MS)-based proteomics is an effective tool to map the proteome of fungal endophytes, providing a solid basis for understanding the mechanisms involved in plant–microbe interactions (e.g., biological pathways and posttranslational modifications) [164]. The protein content of uninoculated and endophyte-inoculated plants can be predicted and assessed to investigate which specific proteins are involved in the relationship between these two groups. For instance, Yuan et al. [165] unveiled the transcriptome and proteome of Atractylodes lancea inoculated with and without the endophytic fungus Gilmaniella sp. AL12. In the study, the authors observed an upregulation of proteins involved in carbon fixation, carbohydrate metabolism and energy metabolism, thus suggesting that Gilmaniella sp. may improve the biomass of the herbal medicine A. lancea [165].
Nevertheless, as far as we know, no proteomic studies have been conducted in the genus Diaporthe. As the proteome profiling allows us to examine the structures, functions and interactions of dynamic proteins in an organism under a specific condition [135,166], in-depth studies should be undertaken to reveal the mechanism of the interactions between species of endophytic Diaporthe and their hosts.
5.4. Metabolomics
The discovery of new secondary metabolites that are involved in cellular functions and microbial networking can assist in the development of alternative antibiotics through microbial metabolite profiling [167]. The analysis of the diverse metabolites has been performed using mainly the following techniques: CE-MS (capillary electrophoresis), LC-MS and GC-MS (liquid and gas chromatography with mass spectrometry) and NMR (nuclear magnetic resonance spectroscopy) [168].
Several studies demonstrate that the genus Diaporthe have the potential to produce a wide range of metabolites with several biological activities with applications mainly in pharmaceuticals and biomedicine [169]. For instance, Kemkuignou et al. [169] used metabolomics approaches, mainly the High-Resolution ElectroSpray Ionization Mass Spectrometry (HR-ESIMS) to screen for the presence of metabolites from the species D. breyniae. The authors extracted and isolated the polyketides fusaristatin G and H, and the cytochalasan phomopchalasin N. Kemkuignou et al. [169] have also demonstrated the antimicrobial activity of these compounds against the fungus Mucor hiemalis, the bacteria Staphylococcus aureus and the yeast Schizosaccharomyces pombe. Moreover, through semi-preparative high-performance liquid chromatography (HPLC), Yedukondalu et al. [170] extracted and separated the compounds diapolic acid A-B and xylarolide from D. terebinthifolii. The authors demonstrated that diapolic A-B exhibited strong antibacterial activity against the human pathogen Yersinia enterocolitica, while xylarolide was effective against Candida albicans and showed potent cytotoxicity against the breast cancer cell line. Moreover, through the NMR approach, Mandavid et al. [171] isolated the polyketide mycoepoxydiene from the fungus D. pseudomangiferae. This metabolite was shown to exhibit cytotoxic activity against some human cancer cell lines (e.g., uterine cervical carcinoma, breast cancer and lung fibroblast) [171].
Although metabolomics is useful in showing the relationship among plants and growth promoters [172], this approach has been less frequently employed in determining functional traits in endophytic microbes [173]. For instance, Cao et al. [174] used mass spectrometry to unveil the metabolic profiles of the endophytic fungus Neotyphodium lolii and its host ryegrass (Lolium perenne). The authors verified changes in the metabolome of inoculated plants and reported the presence of key compounds such as peramine and perloline (alkaloids that increase the capacity of plants to resist the environmental stresses).
Similarly, our understanding of the functional secondary metabolites of species of Diaporthe on microbial interactions in plants are still scarce. Lacerda et al. [127] applied the NMR technique to report the metabolomic changes in Combretum lanceolatum plants inoculated with the endophytic fungus Diaporthe phaseolorum. The authors demonstrated that this fungus affected the metabolic pathways of the plant aerial parts, improving the biosynthesis of primary metabolites involved in plant self-defense such as threonine, malic acid and N-acetyl-mannosamine [127]. Therefore, to better understand the mechanisms of plant–Diaporthe interactions, studies using integrated analysis of genomics, transcriptomics, proteomics and metabolomics could be crucial to analyze key genes, specific proteins and metabolites that are differentially expressed. The exploitation of endophytes is surely important for the identification of biological control agents, as well as of plant-growth promoters, to assist in a sustainable agriculture (Figure 2).
6. Conclusions and Future Prospects
Regardless of the recognized benefits of endophytic fungi on plants and their potential in both biocontrol and biofertilization, they have been rarely studied regarding their application in agriculture. Nevertheless, due to the actual climate change scenarios (e.g., drought and high levels of soil salinity), it is crucial to understand the impacts of these environmental stresses on agriculture, as well as to unravel adaptation patterns of the endophytic community. Moreover, an effective utilization of endophytic fungi aids in promoting a sustainable agriculture for a safe environment and a positive impact on human health.
In this review, our results demonstrate that phytotoxic compounds, antibacterial and antifungal metabolites from species of the genus Diaporthe are promising leads for the development of new biopesticides. This study also suggests that species of this genus could be used as biofertilizers, given their ability as plant-growth promoters and stress tolerance-enhancers. However, a deeper understanding is needed not only to unveil which adaptation patterns of Diaporthe are triggered for their adaptation to changing climatic conditions, but also which adapted community might be applied as tolerance-enhancer treatment. This can be achieved through omics approaches that have revealed an enormous potential to unravel the functions of endophytes and their metabolites in plant disease control, as well as metabolic pathways towards plant growth and tolerance to environmental stresses. Moreover, multi-omics offer a valuable framework that allows a detailed analysis of the biological mechanisms of endophytic Diaporthe to design strategies highlighting their beneficial properties as control agents and plant-growth promoters. Therefore, a detailed understanding of species of Diaporthe, their mechanisms of action and bioactive metabolites, will likely provide a strong basis for developing:
- (1)
- Reliable tools to enhance plant health and growth;
- (2)
- Novel strategies for mitigating the impacts of climate changes;
- (3)
- An ecological and sustainable agriculture.
In recent years, accumulating evidence has provided important advances regarding biological control agents for the development of commercialized bacterial and fungal-based biopesticides to control plant diseases. However, the implementation of large-scale studies to expand the knowledge on the usage of biopesticides is still hampered by the high cost of commercial products, the standard methods of preparations, the dose determination of active substances and the susceptibility of biopesticides to environmental conditions. In this regard, and taking into consideration the possibility of using endophytic Diaporthe as promising leads for the development of biopesticides and biofertilizers, some strategies should be adopted to improve the performance of these endophytic fungi. For instance, the development of specific delivery systems such as biopriming, encapsulation or foliar spraying should be favored to support the success of biocontrol and biofertilization programs. Moreover, the development of effective microbial consortium composed of endophytic fungi such as the species of Diaporthe could also be a promising strategy, not only to ensure the microbial diversity in the soil, but also in the phylosphere; phylosphere colonization is of paramount importance to ensure crop development and plant health management, regulating plant physiology under climate change scenarios.
Author Contributions
Conceptualization: S.H.; Data curation: S.H.; Investigation: M.F.M.G. and S.H.; Methodology: M.F.M.G. and S.H.; Writing—original draft: S.H.; Writing—review and editing: M.F.M.G. and S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Portuguese Foundation for Science and Technology (FCT/MCTES) that finances CESAM (Centre for Environmental and Marine Studies) (UIDP/50017/2020 + UIDB/50017/2020 + LA/P/0094/2020) through national funds, and the PhD grant to Sandra Hilário (SFRH/BD/137394/2018).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Large, E.C. Control of Potato Blight (Phytophthora infestans) by Spraying with Suspensions of Metallic Copper. Nature 1943, 151, 80–81. [Google Scholar] [CrossRef]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Li, X.Q.; Zhao, D.L.; Zhang, P. Antifungal secondary metabolites produced by the fungal endophytes: Chemical diversity and potential use in the development of biopesticides. Front. Microbiol. 2021, 12, 689527. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.H.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef]
- Wang, Z.; Li, C.; White, J. Effects of Epichloë endophyte infection on growth, physiological properties and seed germination of wild barley under saline conditions. J. Agron. Crop Sci. 2019, 206, 43–51. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, C.; Xiang, X.; Zhang, Q.; Zhao, W.; Wei, G.; Hu, A. Uptake and transport of antibiotic kasugamycin in castor bean (Ricinus communis L.) seedlings. Front. Microbiol. 2022, 13, 948171. [Google Scholar] [CrossRef]
- Osada, H. Discovery and applications of nucleoside antibiotics beyond polyoxin. J. Antibiot. 2019, 72, 855–864. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A. Streptomyces-based cell factories for production of biomolecules and bioactive metabolites. In Microbial Cell Factories Engineering for Production of Biomolecules; Academic Press: Cambridge, MA, USA, 2021; pp. 183–234. [Google Scholar]
- Chakraborty, A.; Ray, P. Mycoherbicides for the Noxious Meddlesome: Can Colletotrichum be a Budding Candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef]
- Suebrasri, T.; Harada, H.; Jogloy, S.; Ekprasert, J.; Boonlue, S. Auxin-producing fungal endophytes promote growth of sunchoke. Rhizosphere 2020, 16, 100271. [Google Scholar] [CrossRef]
- Domka, A.M.; Rozpaadek, P.; Turnau, K. Are fungal endophytes merely mycorrhizal copycats? The role of fungal endophytes in the adaptation of plants to metal toxicity. Front. Microbiol. 2019, 10, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Y.; Luan, P.; Wang, K.; Li, G.; Yin, Y.; Yang, Y.; Zhang, Q.; Liu, Y. Antifungal Activity and Plant Growth-Promoting Properties of Bacillus mojovensis B1302 against Rhizoctonia cerealis. Microorganisms 2022, 10, 1682. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Crit. Rev. Environ. Sci. Technol. 2022, 52, 395–435. [Google Scholar] [CrossRef]
- Xu, T.C.; Lu, Y.H.; Wang, J.F.; Song, Z.Q.; Hou, Y.G.; Liu, S.S.; Liu, C.S.; Wu, S.H. Bioactive secondary metabolites of the genus Diaporthe and anamorph Phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef]
- Abramczyk, B.; Marzec-Grządziel, A.; Grządziel, J.; Król, E.; Gałązka, A.; Oleszek, W. Biocontrol Potential and Catabolic Profile of Endophytic Diaporthe eres Strain 1420S from Prunus domestica L. in Poland—A Preliminary Study. Agronomy 2022, 12, 165. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, e173. [Google Scholar]
- Chitnis, V.R.; Suryanarayanan, T.S.; Nataraja, K.N.; Prasad, S.R.; Oelmüller, R.; Shaanker, R.U. Fungal endophyte-mediated crop improvement: The way ahead. Front. Plant Sci. 2020, 11, 1588. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: A tool for plant growth promotion and sustainable agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Proc. Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Jung, H.-Y.; Lee, J.-H.; Lee, I.-J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- García-Latorre, C.; Rodrigo, S.; Santamaria, O. Effect of fungal endophytes on plant growth and nutrient uptake in Trifolium subterraneum and Poa pratensis as affected by plant host specificity. Mycol. Prog. 2021, 20, 1217–1231. [Google Scholar] [CrossRef]
- Baron, N.C.; Costa, N.T.A.; Mochi, D.A.; Rigobelo, E.C. First report of Aspergillus sydowii and Aspergillus brasiliensis as phosphorus solubilizers in maize. Ann Microbiol. 2018, 68, 863–870. [Google Scholar] [CrossRef]
- Christian, N.; Herre, E.A.; Clay, K. Foliar endophytic fungi alter patterns of nitrogen uptake and distribution in Theobroma cacao. New Phytol. 2019, 222, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Rajini, S.B.; Nandhini, M.; Udayashankar, A.C.; Niranjana, S.R.; Lund, O.S.; Prakash, H.S. Diversity, plant growth-promoting traits, and biocontrol potential of fungal endophytes of Sorghum bicolor. Plant Pathol. 2020, 69, 642–654. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Baron, N.C.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef]
- Ismail, H.M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.J. Aspergillus niger boosted heat stress tolerance in sunflower and soybean via regulating their metabolic and antioxidant system. J. Plant Interact. 2020, 15, 223–232. [Google Scholar] [CrossRef]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in sustainable agriculture: A critical sustainable development driver governed by green chemistry principles. Front. Sustain. Food Syst. 2021, 5, 619058. [Google Scholar] [CrossRef]
- Adhikari, K.; Bhandari, S.; Niraula, D.; Shrestha, J. Use of neem (Azadirachta indica A. Juss) as a biopesticide in agriculture: A review. J. Agric. Appl. Biol. 2021, 1, 100–117. [Google Scholar] [CrossRef]
- Kalpana, T.; Anil, K. A review of biopesticides and their plant phytochemicals information. Ann. Romanian Soc. Cell Biol. 2021, 25, 3576–3588. [Google Scholar]
- Santos, C.M.; da Silva, R.A.; Garcia, A.; Polli, A.D.; Polonio, J.C.; Azevedo, J.L.; Pamphile, J.A. Enzymatic and antagonist activity of endophytic fungi from Sapindus saponaria L. (Sapindaceae). Acta Biol. Colomb. 2019, 24, 322. [Google Scholar] [CrossRef]
- Ribeiro, A.D.S.; Polonio, J.C.; Costa, A.T.; Dos Santos, C.M.; Rhoden, S.A.; Azevedo, J.L.; Pamphile, J.A. Bioprospection of culturable endophytic fungi associated with the ornamental plant Pachystachys lutea. Curr. Microbiol. 2018, 75, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Landum, M.; Felix, M.; Alho, J.; Garcia, R.; Cabrita, M.J.; Rei, F.; Varanda, C.M. Antagonistic activity of fungi of Olea europaea L. against Colletotrichum acutatum. Microbiol. Res. 2016, 183, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Rakshith, D.; Santosh, P.; Satish, S. Isolation and characterization of antimicrobial metabolite producing endophytic Phomopsis sp. from Ficus pumila Linn. (Moraceae). Int. J. Chem. Anal. Sci. 2013, 4, 156–160. [Google Scholar] [CrossRef]
- Deketelaere, S.; Tyvaert, L.; França, S.C.; Höfte, M. Desirable traits of a good biocontrol agent against Verticillium wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Sharma, M. Approaches and Trends in Plant Disease Management; Scientific Publishers: Jodhpur, India, 2014; p. 429. [Google Scholar]
- Roberts, W. Studies on biogenesis. Philos. Trans. R. Soc. 1874, 164, 466. [Google Scholar]
- Baker, K.F. Evolving concepts of biological control of plant pathogens. Annu. Rev. Phytopathol. 1987, 25, 67–85. [Google Scholar] [CrossRef]
- Weindling, R. Experimental consideration of the mold toxins of Gliocladium and Trichoderma. Phytopathology 1941, 31, 991. [Google Scholar]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A Source of Potential Antifungal Compounds. J. Fungi 2018, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; He, J.; Li, X.-N.; Huang, R.; Song, F.; Chen, Y.-W.; Miao, C.-P. Guaiane sesquiterpenes and isopimarane diterpenes from an endophytic fungus Xylaria sp. Phytochemistry 2014, 105, 197–204. [Google Scholar] [CrossRef]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Biocontrol with Trichoderma species for the management of postharvest crown rot of banana. Phytopathol. Mediterr. 2009, 48, 214–225. [Google Scholar]
- Bai, Y.B.; Gao, Y.Q.; Nie, X.D.; Tuong, T.M.; Li, D.; Gao, J.M. Antifungal activity of griseofulvin derivatives against phytopathogenic fungi in vitro and in vivo and three-dimensional quantitative structure-activity relationship analysis. J. Agric. Food Chem. 2019, 67, 6125–6132. [Google Scholar] [CrossRef]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Lupo, S. Diversity of fungal latent pathogens and true endophytes associated with fruit trees in Uruguay. J. Phytopathol. 2018, 166, 633–647. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Arciuolo, R.; Camardo Leggieri, M.; Chiusa, G.; Castello, G.; Genova, G.; Spigolon, N.; Battilani, P. Ecology of Diaporthe eres, the causal agent of hazelnut defects. PLoS ONE 2021, 16, e0247563. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Renderos, W.; Bevis, E.; Hebb, J.; Abbasi, P.A. Diaporthe eres causes stem cankers and death of young apple rootstocks in Canada. Can. J. Plant Pathol. 2020, 42, 218–227. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 609387. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by Diaporthe spp. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Chepkirui, C.; Stadler, M. The genus Diaporthe: A rich source of diverse and bioactive metabolites. Mycol. Prog. 2017, 16, 477–494. [Google Scholar] [CrossRef]
- Chen, H.; Huang, M.; Li, X.; Liu, L.; Chen, B.; Wang, J.; Lin, Y. Phochrodines A-D, first naturally occurring new chromenopyridines from mangrove entophytic fungus Phomopsis sp. 33#. Fitoterapia 2018, 124, 103–107. [Google Scholar]
- Sharma, V.; Singamaneni, V.; Sharma, N.; Kumar, A.; Arora, D.; Kushwaha, M.; Bhushan, S.; Jaglan, S.; Gupta, P. Valproic acid induces three novel cytotoxic secondary metabolites in Diaporthe sp., an endophytic fungus from Datura inoxia Mill. Bioorganic Med. Chem. Lett. 2018, 28, 2217–2221. [Google Scholar] [CrossRef]
- Song, H.C.; Qin, D.; Han, M.J.; Wang, L.; Zhang, K.; Dong, J.Y. Bioactive 2-pyrone metabolites from an endophytic Phomopsis asparagi SWUKJ5.2020 of Kadsura angustifolia. Phytochem. Lett. 2017, 22, 235–240. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, L.; Yang, L.; Huang, Y.; Li, S.; Shen, Y. Phomopsidone A, a novel depsidone metabolite from the mangrove endophytic fungus Phomopsis sp. A123. Fitoterapia 2014, 96, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Farooq, U.; Hussain, H.; Ahmed, I.; Rheinheimer, J.; Draeger, S.; Schulz, B.; van Ree, T. Phomosines H-J, novel highly substituted biaryl ethers, isolated from the endophytic fungus Phomopsis sp. from Ligustrum vulgare. Nat. Prod. Commun. 2011, 6, 1907–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastianes, F.L.S.; Cabedo, N.; El Aouad, N.; Valente, A.M.M.P.; Lacava, P.T.; Azevedo, J.L.; Pizzirani-Kleiner, A.A.; Cortes, D. 3-Hydroxypropionic acid as an antibacterial agent from endophytic fungi Diaporthe phaseolorum. Curr. Microbiol. 2012, 65, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.L.; Mottier, N.; Jeanneret-Gris, J.; Christen, D.; Tabacchi, R.; Abou-Mansour, E. Differential production of phytotoxins from Phomopsis sp. from grapevine plants showing esca symptoms. J. Agric. Food Chem. 2014, 62, 8602–8607. [Google Scholar] [CrossRef]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef]
- Wulansari, D.; Julistiono, H.; Nurkanto, A.; Agusta, A. Antifungal activity of (+)-2,20-epicytoskyrin A and its membrane-disruptive action. Makara J. Sci. 2016, 20, 160–166. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Cantrell, C.L.; Wedge, D.E.; Gonçalves, V.N.; Jacob, M.R.; Khan, S.; Rosa, C.A.; Rosa, L.H. Antimycobacterial and antimalarial activities of endophytic fungi associated with the ancient and narrowly endemic neotropical plant Vellozia gigantea from Brazil. Mem. Inst. Oswaldo Cruz 2017, 112, 692–697. [Google Scholar] [CrossRef]
- Liu, Y.; Ruan, Q.; Jiang, S.; Qu, Y.; Chen, J.; Zhao, M.; Yang, B.; Liu, Y.; Zhao, Z.; Cui, H. Cytochalasins and polyketides from the fungus Diaporthe sp. GZU-1021 and their anti-inflammatory activity. Fitoterapia 2019, 137, 104187. [Google Scholar] [CrossRef]
- Polonio, J.C.; Almeida, T.; Garcia, A.; Mariucci, G.; Azevedo, J.; Rhoden, S.; Pamphile, J.A. Biotechnological prospecting of foliar endophytic fungi of guaco (Mikania glomerata Spreng.) with antibacterial and antagonistic activity against phytopathogens. Genet. Mol. Res. 2015, 14, 7297–7309. [Google Scholar] [CrossRef]
- Verma, P.; Hiremani, N.S.; Gawande, S.P.; Sain, S.K.; Nagrale, D.T.; Narkhedkar, N.G.; Prasad, Y.G. Modulation of plant growth and antioxidative defense system through endophyte biopriming in cotton (Gossypium spp.) and non-host crops. Heliyon 2022, 8, e09487. [Google Scholar] [CrossRef]
- Yu, J.J.; Yang, H.X.; Zhang, F.L.; He, J.; Li, Z.H.; Liu, J.K.; Feng, T. Secondary metabolites from cultures of the kiwi-associated fungus Diaporthe phragmitis and their antibacterial activity assessment. Phytochem. Lett. 2021, 46, 143–148. [Google Scholar] [CrossRef]
- Carvalho, C.D.; Ferreira-D’Silva, A.; Wedge, D.E.; Cantrell, C.L.; Rosa, L.H. Antifungal activities of cytochalasins produced by Diaporthe miriciae, an endophytic fungus associated with tropical medicinal plants. Can. J. Microbiol. 2018, 64, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ma, K.X.; Xie, X.S.; Wang, T.; Wu, S.H. Secondary metabolites of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2015, 51, 392–394. [Google Scholar] [CrossRef]
- Ma, K.X.; Shen, X.T.; Huang, R.; Wang, T.; Xie, X.S.; Liu, S.W.; Wu, S.H.; He, J. Bioactive metabolites produced by the endophytic fungus Phomopsis sp. YM355364. Nat. Prod. Commun. 2014, 9, 669–670. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, W.; Wang, X.; Shahid, H.; Yang, Y.; Wang, Y.; Wang, S.; Shan, T. Diversity and communities of culturable endophytic fungi from the root holoparasite Balanophora polyandra Griff. and their antibacterial and antioxidant activities. Ann. Microbiol. 2022, 72, 19. [Google Scholar] [CrossRef]
- Hussain, H.; Ahmed, I.; Schulz, B.; Draeger, S.; Krohn, K. Pyrenocines J-M: Four new pyrenocines from the endophytic fungus, Phomopsis sp. Fitoterapia 2012, 83, 523–526. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Islam, M.T.; Facey, P.; Douanla-Meli, C.; von Tiedemann, A.; Laatsch, H. Depsidones and other constituents from Phomopsis sp. CAFT69 and its host plant Endodesmia calophylloides with potent inhibitory effect on motility of zoospores of grapevine pathogen Plasmopara viticola. Phytochem. Lett. 2012, 3, 657–664. [Google Scholar] [CrossRef]
- Prada, H.; Avila, L.; Sierra, R.; Bernal, A.; Restrepo, S. Morphological and molecular characterization of the antagonistic interaction between the endophyte Diaporthe sp. isolated from frailejon (Espeletia sp.) and the plant pathogen Phytophthora infestans. Rev. Iberoam. Micol. 2009, 26, 198–201. [Google Scholar] [CrossRef]
- Fu, J.; Zhou, Y.; Li, H.F.; Ye, Y.H.; Guo, J.H. Antifungal metabolites from Phomopsis sp. By254, an endophytic fungus in Gossypium hirsutum. Afr. J. Microbiol. Res. 2011, 5, 1231–1236. [Google Scholar]
- Santos, P.J.; Savi, D.C.; Gomes, R.R.; Goulin, E.H.; Da Costa Senkiv, C.; Tanaka, F.A.; Almeida, Á.M.; Galli-Terasawa, L.; Kava, V.; Glienke, C. Diaporthe endophytica and D. terebinthifolii from medicinal plants for biological control of Phyllosticta citricarpa. Microbiol. Res. 2016, 186–187, 153–160. [Google Scholar] [CrossRef]
- Tonial, F.; Maia, B.H.; Sobottka, A.M.; Savi, D.C.; Vicente, V.A.; Gomes, R.R.; Glienke, C. Biological activity of Diaporthe terebinthifolii extracts against Phyllosticta citricarpa. FEMS Microbiol. Lett. 2017, 364, fnx026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiono, Y.; Sasaki, T.; Shibuya, F.; Yasuda, Y.; Koseki, T.; Supratman, U. Isolation of a phomoxanthone A derivative, a new metabolite of tetrahydroxanthone, from a Phomopsis sp. isolated from the mangrove, Rhizhopora mucronata. Nat. Prod. Commun. 2013, 8, 1735–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, T.S. Bioactive metabolites from tomato endophytic fungi with antibacterial activity against tomato bacterial spot disease. Rhizosphere 2021, 17, 100292. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palacio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the Brazilian biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.; Santos, S. Sustainable approach to weed management: The role of precision weed management. Agronomy 2022, 12, 118. [Google Scholar] [CrossRef]
- Esposito, M.; Crimaldi, M.; Cirillo, V. Drone and sensor technology for sustainable weed management: A review. Chem. Biol. Technol. Agric. 2021, 8, 18. [Google Scholar] [CrossRef]
- Bjawa, A.A.; Mahajan, G.; Chauhan, B.S. Nonconventional weed management strategies for modern agriculture. Weed Sci. 2015, 63, 723–747. [Google Scholar] [CrossRef]
- Mia, M.J.; Massetani, F.; Murri, G.; Neri, D. Sustainable alternatives to chemicals for weed control in the orchard—A Review. Hortic. Sci. 2020, 47, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bastos, B.D.O.; Deobald, G.A.; Brun, T.; Dal Prá, V.; Junges, E.; Kuhn, R.C.; Pinto, A.K.; Mazutti, M.A. Solid-state fermentation for production of a bioherbicide from Diaporthe sp. and its formulation to enhance the efficacy. Biotech 2017, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Brun, T.; Rabuske, J.E.; Luft, L.; Confortin, T.C.; Todero, I.; Aita, B.C.; Zabot, G.L.; Mazutti, M.A. Powder containing biomolecules from Diaporthe schini for weed control. Environ. Technol. 2022, 43, 2135–2144. [Google Scholar] [CrossRef]
- Cimmino, A.; Cinelli, T.; Masi, M.; Reveglia, P.; da Silva, M.A.; Mugnai, L.; Michereff, S.J.; Surico, G.; Evidente, A. Phytotoxic lipophilic metabolites produced by grapevine strains of Lasiodiplodia species in Brazil. J. Agric. Food Chem. 2017, 65, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Macías-Rubalcava, M.L.; García-Méndez, M.C.; King-Díaz, B.; Macías-Ruvalcaba, N.A. Effect of phytotoxic secondary metabolites and semisynthetic compounds from endophytic fungus Xylaria feejeensis strain SM3e-1b on spinach chloroplast photosynthesis. J. Photochem. Photobiol. B 2017, 166, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and phomentrioloxins B and C Produced by Diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.R.C.; Baldoni, D.B.; Lima, J.; Porto, V.; Marcuz, C.; Ferraz, R.C.; Kuhn, R.C.; Jacques, R.J.S.; Guedes, J.V.C.; Mazutti, M.A. Bioherbicide production by Diaporthe sp. isolated from the Brazilian Pampa biome. Biocatal. Agric. Biotechnol. 2015, 4, 575–578. [Google Scholar] [CrossRef]
- Souza, A.R.C.; Baldoni, D.B.; Lima, J.; Porto, V.; Marcuz, C.; Machado, C.; Ferraz, R.C.; Kuhn, R.C.; Jacques, R.J.S.; Guedes, J.V.C.; et al. Selection, isolation, and identification of fungi for bioherbicide production. Braz. J. Microbiol. 2016, 48, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Pes, M.P.; Mazutti, M.A.; Almeida, T.C.; Curioletti, L.E.; Melo, A.A.; Guedes, J.V.C.; Kuhn, R.C. Bioherbicide based on Diaporthe sp. secondary metabolites in the control of three tough weeds. Afr. J. Agric. Res. 2016, 11, 4242–4249. [Google Scholar]
- Briscoe, W.E. Identification of a Phytotoxic Fungus and an Investigation into the Isolation of Its Phytotoxic Constituents. Ph.D. Tesis, University of Mississippi, Oxford, MS, USA, 2014. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Boari, A.; Troise, C.; Motta, A.; Vurro, M.; Ash, C.; Evidente, A. Phomentrioloxin, a fungal phytotoxin with potential herbicidal activity, and its derivatives: A structure–activity relationship study. J. Agric. Food Chem. 2013, 61, 9645–9649. [Google Scholar] [CrossRef]
- Evidente, M.; Boari, A.; Vergura, S.; Cimmino, A.; Vurro, M.; Ash, G.; Superchi, S.; Evidente, A. Structure and absolute configuration of kongiidiazadione, a new phytotoxic 3-substituted-5-diazenylcyclopentendione produced by Diaporthe Kongii. Chirality 2015, 27, 557–562. [Google Scholar] [CrossRef]
- Yang, Z.; Ge, M.; Yin, Y.; Chen, Y.; Luo, M.; Chen, D. A novel phytotoxic nonenolide from Phomopsis sp. HCCB03520. Chem. Biodivers. 2012, 9, 403–408. [Google Scholar] [CrossRef]
- Meepagala, K.M.; Briscoe, W.E.; Techen, N.; Johnson, R.D.; Clausen, B.M.; Duke, S.O. Isolation of a phytotoxic isocoumarin from Diaporthe eres-infected Hedera helix (English ivy) and synthesis of its phytotoxic analogs. Pest Manag. Sci. 2018, 74, 37–45. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Solfrizzo, M.; Tosi, L.; Zazzerini, A.; Fanizzi, F.P.; Visconti, A. Isolation and characterization of phytotoxic compounds produced by Phomopsis helianthi. Nat. Toxins 1999, 7, 119–127. [Google Scholar] [CrossRef]
- Reveglia, P.; Pacetti, A.; Masi, M.; Cimmino, A.; Carella, G.; Marchi, G.; Mugnai, L.; Evidente, A. Phytotoxic metabolites produced by Diaporthe eres involved in cane blight of grapevine in Italy. Nat. Prod. Res. 2021, 35, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Srikitikulchai, P.; Srichomthong, K.; Lumyong, S. Oblongolides from the endophytic fungus Phomopsis sp. BCC 9789. J. Nat. Prod. 2010, 73, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Hussain, H.; Schulz, B.; Draeger, S.; Padula, D.; Pescitelli, G.; Van Ree, T.; Krohn, K. Three new antimicrobial metabolites from the endophytic fungus Phomopsis sp. Eur. J. Chem. 2011, 15, 2867–2873. [Google Scholar] [CrossRef]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Ogilvie, K.K.; Watson, A.K. Phytotoxic metabolites of Phomopsis convolvulus, a host-specific pathogen of field bindweed. Can. J. Chem. 1992, 70, 2276–2284. [Google Scholar] [CrossRef] [Green Version]
- Ballio, A.; Bottalico, A.; Framondino, M.; Graniti, A.; Randazzo, G. Fusicoccin: Structure-phytotoxicity relationships. Phytopathol. Mediterr. 1971, 10, 26–32. [Google Scholar]
- Evidente, A.; Lanzetta, R.; Abouzeid, M.A.; Corsaro, M.M.; Mugnai, L.; Surico, G. Foeniculoxin, a new phytotoxic geranylhydroquinone from Phomopsis foeniculi. Tetrahedron 1994, 50, 10371–10378. [Google Scholar] [CrossRef]
- Turner, N.C.; Graniti, A. Fusicoccin: A fungal toxin that opens stomata. Nature 1969, 223, 1070–1071. [Google Scholar] [CrossRef]
- Culvenor, C.C.J.; Edgar, J.A.; Mackay, M.F.; Gorst-Allman, C.P.; Marasas, W.F.O.; Steyn, P.S.; Vleggaar, R.; Wessels, P.L. Structure elucidation and absolute configuration of phomopsin A, a hexapeptide mycotoxin produced by Phomopsis leptostromiformis. Tetrahedron 1989, 45, 2351–2372. [Google Scholar] [CrossRef]
- Mazars, C.; Rossignol, M.; Auriol, P.; Klaebe, A. Phomozin, a phytotoxin from Phomopsis helianthi, the causal agent of stem canker of sunflower. Phytochemistry 1990, 29, 3441–3444. [Google Scholar] [CrossRef]
- Lopes, M.J.D.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful plant growth-promoting microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci. Hort. 2020, 263, 1–10. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Ali Babar, M. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poveda, J.; Eugui, D.; Abril-Urías, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92, fiw194. [Google Scholar] [CrossRef]
- Du, Y.C.; Kong, L.J.; Cao, L.S.; Zhang, W.; Zhu, Q.; Ma, C.Y.; Sun, K.; Dai, C.C. Endophytic Fungus Phomopsis liquidambaris Enhances Fe Absorption in Peanuts by Reducing Hydrogen Peroxide. Front. Plant Sci. 2022, 13, 872242. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Dai, C.C.; Li, X.; Tian, L.S.; Wang, X.X. Extensive host range of an endophytic fungus affects the growth and physiological functions in rice (Oryza sativa L.). Symbiosis 2007, 43, 21–28. [Google Scholar]
- Zhou, J.; Huang, P.W.; Li, X.; Vaistij, F.E.; Dai, C.C. Generalist endophyte Phomopsis liquidambaris colonization of Oryza sativa L. promotes plant growth under nitrogen starvation. Plant Mol. Biol. 2022, 6, 703–715. [Google Scholar] [CrossRef]
- Aldana, B.R.V.; Arellano, J.B.; Cuesta, M.J.; Mellado-Ortega, E.; González-Blanco, V.; Zabalgogeazcoa, I. Screening fungal endophytes from a wild grass for growth promotion in tritordeum, an agricultural cereal. Plant Sci. 2021, 303, 110762. [Google Scholar] [CrossRef]
- Toghueo, R.M.; Zabalgogeazcoa, I.; Pereira, E.C.; de Aldana, B.R.V. A Diaporthe fungal endophyte from a wild grass improves growth and salinity tolerance of tritordeum and perennial ryegrass. Front. Plant Sci. 2022, 13, 896755. [Google Scholar] [CrossRef] [PubMed]
- Ważny, R.; Rozpądek, P.; Jędrzejczyk, R.J.; Domka, A.; Nosek, M.; Kidd, P.; Turnau, K. Phytohormone based biostimulant combined with plant growth promoting endophytic fungus enhances Ni phytoextraction of Noccaea goesingensis. Sci. Total Environ. 2021, 789, 147950. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, A.S.; Ray, J.G. Applications of endophytic-fungal-isolates from velamen root of wild orchids in floriculture. Braz. J. Biol. 2019, 6, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, J.W.; Siqueira, K.A.; Vasconcelos, L.G.; Bellete, B.S.; Dall’Oglio, E.L.; Sousa Junior, P.T.; Faraggi, T.M.; Vieira, L.C.; Soares, M.A.; Sampaio, O.M. Metabolomic Analysis of Combretum lanceolatum Plants Interaction with Diaporthe phaseolorum and Trichoderma spirale Endophytic Fungi through 1H-NMR. Chem. Biodivers. 2021, 18, 2100350. [Google Scholar] [CrossRef] [PubMed]
- Serrano, L.; Lacerda, J.W.; Moura, M.S.; Ali, A.; de Vasconcelos, L.G.; Junior, P.T.S.; Bellete, B.S.; Soares, M.A.; Vieira, L.C.; Sampaio, O.M. Metabolomics Analysis of C. lanceolatum Roots in the Presence of Its Endophytic Fungi. J. Braz. Chem. Soc. 2022, 18, e2100350. [Google Scholar] [CrossRef]
- da Silva Santos, S.D.S.; da Silva, A.A.D.; Polonio, J.C.; Polli, A.D.; Orlandelli, R.C.; dos Santos Oliveira, J.A.D.S.; Brandão Filho, J.U.T.; Azevedo, J.L.; Pamphile, J.A. Influence of plant growth-promoting endophytes Colletotrichum siamense and Diaporthe masirevici on tomato plants (Lycopersicon esculentum Mill.). Mycology 2022, 13, 257–270. [Google Scholar] [CrossRef]
- Ballio, A.; Michelis, M.I.; Lado, P.; Randazzo, G. Fusicoccin structure-activity relationships: Stimulation of growth by cell enlargement and promotion of seed germination. Physiol. Plant. 1981, 52, 471–475. [Google Scholar] [CrossRef]
- Marra, A.; Camoni, A.L.; Visconti, A.; Fiorillo, A.; Evident, A. The Surprising story of Fusicoccin: A wilt-inducing phytotoxin, a tool in plant physiology and a 14-3-3-targeted drug. Biomolecules 2021, 11, 1393. [Google Scholar] [CrossRef]
- Chithra, S.; Mathew, J.; Radhakrishnan, E.K. Endophytic Phomopsis sp. colonization in Oryza sativa was found to result in plant growth promotion and piperine production. Physiol. Plant. 2017, 160, 437–446. [Google Scholar]
- Bhattacharjee, S.; Roy Das, A.; Saha, A.K.; Das, P. Fungal endophytes from medicinal plants: Growth promotion in Oryza sativa L. and Cicer arietinum L. Vegetos 2019, 32, 381–386. [Google Scholar] [CrossRef]
- Yang, D.; Du, X.; Yang, Z.; Liang, Z.; Guo, Z.; Liu, Y. Transcriptomics, proteomics, and metabolomics to reveal mechanisms underlying plant secondary metabolism. Eng. Life Sci. 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, J.; Chen, C.; Mo, X.; Tan, Q.; He, Y.; Wang, Z.; Yin, J.; Zhou, G. The Multifunctions and Future Prospects of Endophytes and Their Metabolites in Plant Disease Management. Microorganisms 2022, 10, 1072. [Google Scholar] [CrossRef] [PubMed]
- Lubna, L.; Asaf, S.; Jan, R.; Khan, A.L.; Bilal, S.; Asif, S.; Al-Harrasi, A.; Kim, K.M. Unraveling the Genome Sequence of Plant Growth Promoting Aspergillus niger (CSR3) Provides Insight into the Synthesis of Secondary Metabolites and Its Comparative Genomics. J. Fungi 2022, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559–577. [Google Scholar] [CrossRef]
- Mohana, N.C.; Rao, H.Y.; Rakshith, D.; Mithun, P.R.; Nuthan, B.R.; Satish, S. Omics based approach for biodiscovery of microbial natural products in antibiotic resistance era. J. Genet. Eng. Biotechnol. 2018, 16, 1–8. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Robey, M.T.; Caesar, L.K.; Drott, M.T.; Keller, N.P.; Kelleher, N.L. An interpreted atlas of biosynthetic gene clusters from 1,000 fungal genomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2020230118. [Google Scholar] [CrossRef]
- Grover, S.; Lawrence, C.B. The Alternaria alternata mycotoxin alternariol suppresses lipopolysaccharide-induced inflammation. Int. J. Mol. Sci. 2017, 18, 1577. [Google Scholar] [CrossRef] [Green Version]
- Reveglia, P.; Masi, M.; Evidente, A. Melleins—Intriguing natural compounds. Biomolecules 2020, 10, 772. [Google Scholar] [CrossRef]
- Ballio, A.; Carilli, A.; Santurbano, B.; Tuttobello, L. Produzione di fusicoccina in scala pilota. Ann. Ist. Sup. Sanità 1968, 4, 317–332. [Google Scholar]
- Marrè, E. Fusicoccin: A tool in plant physiology. Annu. Rev. Plant Physiol. 1979, 30, 273–288. [Google Scholar] [CrossRef]
- Bacha, N.; Dao, H.P.; Atoui, A.; Mathieu, F.; O’Callaghan, J.; Puel, O.; Liboz, T.; Dobson, A.D.W.; Lebrihi, A. Cloning and characterization of novel methylsalicylic acid synthase gene involved in the biosynthesis of isoasperlactone and asperlactone in Aspergillus westerdijkiae. Fungal Genet. Biol. 2009, 46, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Balcells, M.; Sala, N.; Sanchis, V.; Canela, R. Bactericidal and fungicidal activity of Aspergillus ochraceus metabolites and some derivatives. Pestic. Sci. 1998, 53, 9–14. [Google Scholar] [CrossRef]
- Balcells, M.; Canela, R.; Coll, J.; Sanchís, V.; Torres, M. Effect of fungal metabolites and some derivatives against Tribolium castaneum (Herbst) and Nezara viridula (L.). Pest. Sci. 1995, 45, 319–323. [Google Scholar] [CrossRef]
- Giacometti, R.; Jacobi, V.; Kronberg, F.; Panagos, C.; Edison, A.S.; Zavala, J.A. Digestive activity and organic compounds of Nezara viridula watery saliva induce defensive soybean seed responses. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pointer, M.D.; Gage, M.J.; Spurgin, L.G. Tribolium beetles as a model system in evolution and ecology. Heredity 2021, 126, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, S.Q.; Tang, H.Y.; Li, X.J.; Zhang, L.; Xiao, J.; Gao, Y.Q.; Zhang, A.L.; Gao, J.M. Potential allelopathic indole diketopiperazines produced by the plant endophytic Aspergillus fumigatus using the one strain–many compounds method. J. Agric. Food Chem. 2013, 61, 11447–11452. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, J.; Sun, Q.Q.; Qin, J.C.; Pescitelli, G.; Gao, J.M. Characterization of cytochalasins from the endophytic Xylaria sp. and their biological functions. J. Agric. Food Chem. 2014, 62, 10962–10969. [Google Scholar] [CrossRef]
- Izumi, Y.; Ohtani, K.; Miyamoto, Y.; Masunaka, A.; Fukumoto, T.; Gomi, K.; Tada, Y.; Ichimura, K.; Peever, T.L.; Akimitsu, K. A polyketide synthase gene, ACRTS2, is responsible for biosynthesis of host-selective ACR-toxin in the rough lemon pathotype of Alternaria alternata. Mol. Plant Microbe Interact. 2012, 25, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Masunaka, A.; Ohtani, K.; Peever, T.L.; Timmer, L.W.; Tsuge, T.; Yamamoto, M.; Yamamoto, H.; Akimitsu, K. An isolate of Alternaria alternata that is pathogenic to both tangerines and rough lemon and produces two host-selective toxins, ACT-and ACR-toxins. Phytopathology 2005, 95, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Hostselective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Pinson-Gadais, L.; Richard-Forget, F. Fusarium mycotoxins enniatins: An updated review of their occurrence, the producing Fusarium species, and the abiotic determinants of their accumulation in crop harvests. J. Agric. Food Chem. 2020, 68, 4788–4798. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A review of the mycotoxin enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Kleigrewe, K.; Söhnel, A.C.; Humpf, H.U. A new high-performance liquid chromatography–tandem mass spectrometry method based on dispersive solid phase extraction for the determination of the mycotoxin fusarin C in corn ears and processed corn samples. J. Agric. Food Chem 2011, 59, 10470–10476. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, W.; Lin, Y.; Lin, Y.; Tang, Z.; Wang, J.; Tao, J.; Tang, W.; Liu, W. Characterization of a carboxyl methyltransferase in Fusarium graminearum provides insights into the biosynthesis of fusarin A. Org. Biomol. Chem. 2021, 19, 6638–6643. [Google Scholar] [CrossRef]
- Ugai, T.; Minami, A.; Fujii, R.; Tanaka, M.; Oguri, H.; Gomi, K.; Oikawa, H. Heterologous expression of highly reducing polyketide synthase involved in betaenone biosynthesis. Chem. Commun. 2015, 51, 1878–1881. [Google Scholar] [CrossRef]
- Qi, J.; Jiang, L.; Zhao, P.; Chen, H.; Jia, X.; Zhao, L.; Dai, H.; Hu, J.; Liu, C.; Shim, S.H.; et al. Chaetoglobosins and azaphilones from Chaetomium globosum associated with Apostichopus japonicus. Appl. Microbiol. Biotechnol. 2020, 104, 545–1553. [Google Scholar] [CrossRef]
- Moolhuijzen, P.M.; Muria-Gonzalez, M.J.; Syme, R.; Rawlinson, C.; See, P.T.; Moffat, C.S.; Ellwood, S.R. Expansion and conservation of biosynthetic gene clusters in pathogenic Pyrenophora spp. Toxins 2020, 12, 242. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.; Sharma, T.K.; Dhar, M. “Omics” tools for better understanding the plant–endophyte interactions. Front. Plant Sci. 2016, 7, 955. [Google Scholar] [CrossRef] [Green Version]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. Shotgun proteomics as a powerful tool for the study of the proteomes of plants, their pathogens, and plant–pathogen interactions. Proteomes 2022, 10, 5. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, W.; Sun, K.; Tang, M.-J.; Chen, P.-X.; Li, X.; Dai, C.-C. Comparative Transcriptomics and Proteomics of Atractylodes lancea in Response to Endophytic Fungus Gilmaniella sp. AL12 Reveals Regulation in Plant Metabolism. Front. Microbiol. 2019, 10, 1208. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, A.; Kataoka, R. Metabolite profiling reveals a complex response of plants to application of plant growth-promoting endophytic bacteria. Microbiol. Res. 2020, 234, 126421. [Google Scholar] [CrossRef] [PubMed]
- Spina, R.; Saliba, S.; Dupire, F.; Ptak, A.; Hehn, A.; Piutti, S.; Poinsignon, S.; Leclerc, S.; Bouguet-Bonnet, S.; Laurain-Mattar, D. Molecular identification of endophytic bacteria in Leucojum aestivum in vitro culture, NMR-based metabolomics study and LC-MS analysis leading to potential Amaryllidaceae alkaloid production. Int. J. Mol. Sci. 2021, 22, 1773. [Google Scholar] [CrossRef] [PubMed]
- Kemkuignou, B.M.; Schweizer, L.; Lambert, C.; Anoumedem, E.G.M.; Kouam, S.F.; Stadler, M.; Marin-Felix, Y. New polyketides from the liquid culture of Diaporthe breyniae sp. nov.(Diaporthales, Diaporthaceae). MycoKeys 2022, 90, 85–118. [Google Scholar] [CrossRef]
- Yedukondalu, N.; Arora, P.; Wadhwa, B.; Malik, F.A.; Vishwakarma, R.A.; Gupta, V.K.; Riyaz-Ul-Hassan, S.; Ali, A. Diapolic acid A–B from an endophytic fungus, Diaporthe terebinthifolii depicting antimicrobial and cytotoxic activity. J. Antibiot. 2017, 70, 212–215. [Google Scholar] [CrossRef]
- Mandavid, H.; Rodrigues, A.M.; Espindola, L.S.; Eparvier, V.; Stien, D. Secondary metabolites isolated from the amazonian endophytic fungus Diaporthe sp. SNB-GSS10. J. Nat. Prod. 2015, 78, 1735–1739. [Google Scholar] [CrossRef]
- Gupta, S.; Schillaci, M.; Roessner, U. Metabolomics as an emerging tool to study plant–microbe interactions. Emerg. Top. Life Sci. 2022, 6, 175–183. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Meta-omics of endophytic microbes in agricultural biotechnology. Biocatal. Agric. Biotechnol. 2022, 42, 102332. [Google Scholar] [CrossRef]
- Cao, M.; Koulman, A.; Johnson, L.J.; Lane, G.A.; Rasmussen, S. Advanced data-mining strategies for the analysis of direct-infusion ion trap mass spectrometry data from the association of perennial ryegrass with its endophytic fungus, Neotyphodium lolii. Plant Physiol. 2008, 146, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of possible modes of action of endophytic fungi as biological control agents and plant-growth promoters. Beneficial microorganisms can exhibit direct antagonism against plant pathogens (inhibitor green line), as well as promote plant growth (dotted green line). Plant pathogens can also produce toxins to injure the plant (dot orange line). The figure was created with BioRender.com (accessed on 20 October 2022).
Figure 1.
Overview of possible modes of action of endophytic fungi as biological control agents and plant-growth promoters. Beneficial microorganisms can exhibit direct antagonism against plant pathogens (inhibitor green line), as well as promote plant growth (dotted green line). Plant pathogens can also produce toxins to injure the plant (dot orange line). The figure was created with BioRender.com (accessed on 20 October 2022).

Figure 2.
Schematic representation of omics approaches’ suitability for the exploitation of fungal endophytes for their application as biocontrol agents and plant-growth promoters. The figure was created with BioRender.com (accessed on 20 October 2022).
Figure 2.
Schematic representation of omics approaches’ suitability for the exploitation of fungal endophytes for their application as biocontrol agents and plant-growth promoters. The figure was created with BioRender.com (accessed on 20 October 2022).

Table 2.
Overview of some secondary metabolites isolated from species of Diaporthe and Phomopsis-like species with phytotoxic activity.
Table 2.
Overview of some secondary metabolites isolated from species of Diaporthe and Phomopsis-like species with phytotoxic activity.
| Compound | Strain | Host | References |
|---|---|---|---|
| 3,4-Dihydro-8-hydroxy-3,5-dimethylisocoumarin | D. eres | Hedera helix | [103] |
| (6S,7R,9R)6,7-Dihydroxy-9-propylnon-4-eno-9-lactone | Phomopsis sp. (syn. Diaporthe sp.) | Achyranthes bidentata | [100,102] |
| 4,6-dihydroxymellein | P. helianthi (syn. D. helianthi) | Helianthus annus | [104] |
| 4-Hydroxybenzaldehyde | D. eres | Vitis vinifera | [105] |
| 4-Hydroxybenzoic acid | D. eres | Vitis vinifera | [105] |
| 8-hydroxy-3,7-dimethylisochroman-1-one | D. eres | Hedera helix | [99] |
| 5-(hydroxymethyl) mellein | Phomopsis sp. (syn. Diaporthe sp.) | Musa acuminata Cistus monspeliensis | [106,107] |
| 2-(4-hydroxyphenyl)-ethanol | D. eres | Hedera helix | [99,103] |
| 5-methylmellein | Phomopsis sp. (syn. Diaporthe sp.) | Musa acuminata Cistus monspeliensis | [106,107] |
| 3-Nitropropionic acid | D. gulyae | Carthamus lanatus | [95] |
| alternariol 4,10-dimethyl | P. phragmitis (syn. D. eres) | Actinidia chinensis | [73] |
| Altersolanol A | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [108] |
| Altersolanol J | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [108] |
| Convolvulanic acid A | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Convolvulanic acid B | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Convolvulol | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| p-Cresol | D. eres | Vitis vinifera | [105] |
| Cytochalasin H, N | Phomopsis sp. (syn. Diaporthe sp.) D. miriciae | Achyranthes bidentata, Copaifera pubiflora and Melocactus ernestii | [74,102] |
| dideacetylfusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Epoxycytochalasin H | Phomopsis sp. (syn. Diaporthe sp.) | Achyranthes bidentata | [102] |
| etheralternariol methyl ether | P. phragmitis (syn. D. eres) | Actinidia chinensis | [73] |
| Foeniculoxin | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [111] |
| Fusicoccin | P. amygdali (syn. D. amygdali) | Prunus dulcis Prunus persica | [112] |
| Gulypyrone A | D. gulyae | Carthamus lanatus | [95] |
| Gulypyrone B | D. gulyae | Carthamus lanatus | [95] |
| isofusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Kongiidiazadione | D. kongii | Carthamus lanatus | [101] |
| monodeacetylfusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Nectriapyrone | D. kongii D. eres P. foeniculi (syn. D. angelicae) | Carthamus lanatus Vitis vinifera Foeniculum vulgare | [101,105] |
| Phomentrioloxin B | D. gulyae | Carthamus lanatus | [95,100] |
| Phomopsolide B | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsolidone A | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsolidone B | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsin A | P. leptostromiformis (syn. D. toxica) | Lupinus sp. | [113] |
| Phomozin | P. helianthi (syn. D. helianthi) | Helianthus annus | [114] |
| α-pyrone convolvulopyrone | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Sydowinin A | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Sydowinol | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Tyrosol | D. eres | Vitis vinifera Hedera helix | [103,105] |
Table 3.
Benefits of endophytic Diaporthe to agriculture.
| Host Plant | Endophyte | Benefits | References |
|---|---|---|---|
| Acampe praemorsa | D. eucalyptorum | Increase the fresh-weight and enhance the growth of ornamental orchids (Dendrobium sp.) | [126] |
| Combretum lanceolatum | D. phaseolorum | Improve the biosynthesis of primary metabolites involved in Combretum lanceolatum self-defence | [127,128] |
| Festuca rubra | Diaporthe sp. | Improve nutrient uptake, growth, and salinity tolerance of Lolium perenne and the hybrid from Triticum durum × Hordeum | [124] |
| Helianthus tuberosus | D. phaseolorum | Enhance the growth and increases chlorophyll content of sunchoke plants (Helianthus tuberosus) | [11] |
| Justicia brandegeana | D. masirevicii | Growth-promoting effects on tomato plants and suppression of F. oxysporum symptoms in this host | [129] |
| Lactuca sativa | P. amygdali (syn. D. amygdali) | Cell enlargement, cotyledon growth and seed germination | [130,131] |
| Noccaea goesingensis | D. eres | Improve the biomass Noccaea goesingensis and increase nickel accumulation. Potentially used as biofertilizer. | [124] |
| Oryza sativa | P. liquidambaris (syn. D. liquidambaris) | Enhance growth of rice plants under nitrogen-deficient conditions, and induces rice resistance to rice spikelet rot disease caused by Fusarium graminearum | [121] |
| Piper nigrum | Phomopsis sp. (syn. Diaporthe sp.) | Enhance growth of rice plants due to the production of gibberellins and indole acetic acid | [132] |
| Terminalia arjuna Phlogacanthus thyrsiflorus | D. phaseolorum | Increase root length and enhance plant growth of Cicer arietinum | [133] |
| Triticum durum × Hordeum chilense | Diaporthe sp. | Increase concentrations of calcium, magnesium, sulphur, iron, and boron, thus increasing root and shoot biomass of T. durum × H. chilense | [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hilário, S.; Gonçalves, M.F.M. Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture. Microorganisms 2022, 10, 2453. https://doi.org/10.3390/microorganisms10122453
AMA Style
Hilário S, Gonçalves MFM. Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture. Microorganisms. 2022; 10(12):2453. https://doi.org/10.3390/microorganisms10122453
Chicago/Turabian StyleHilário, Sandra, and Micael F. M. Gonçalves. 2022. "Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture" Microorganisms 10, no. 12: 2453. https://doi.org/10.3390/microorganisms10122453
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.