
Phylogenetic Revision and Patterns of Host Specificity in the Fungal Subphylum Entomophthoromycotina

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition of Fungal Strains
2.2. Phylogenetic Analysis
2.3. Parasitic Fungi and Their Insect Hosts
3. Results
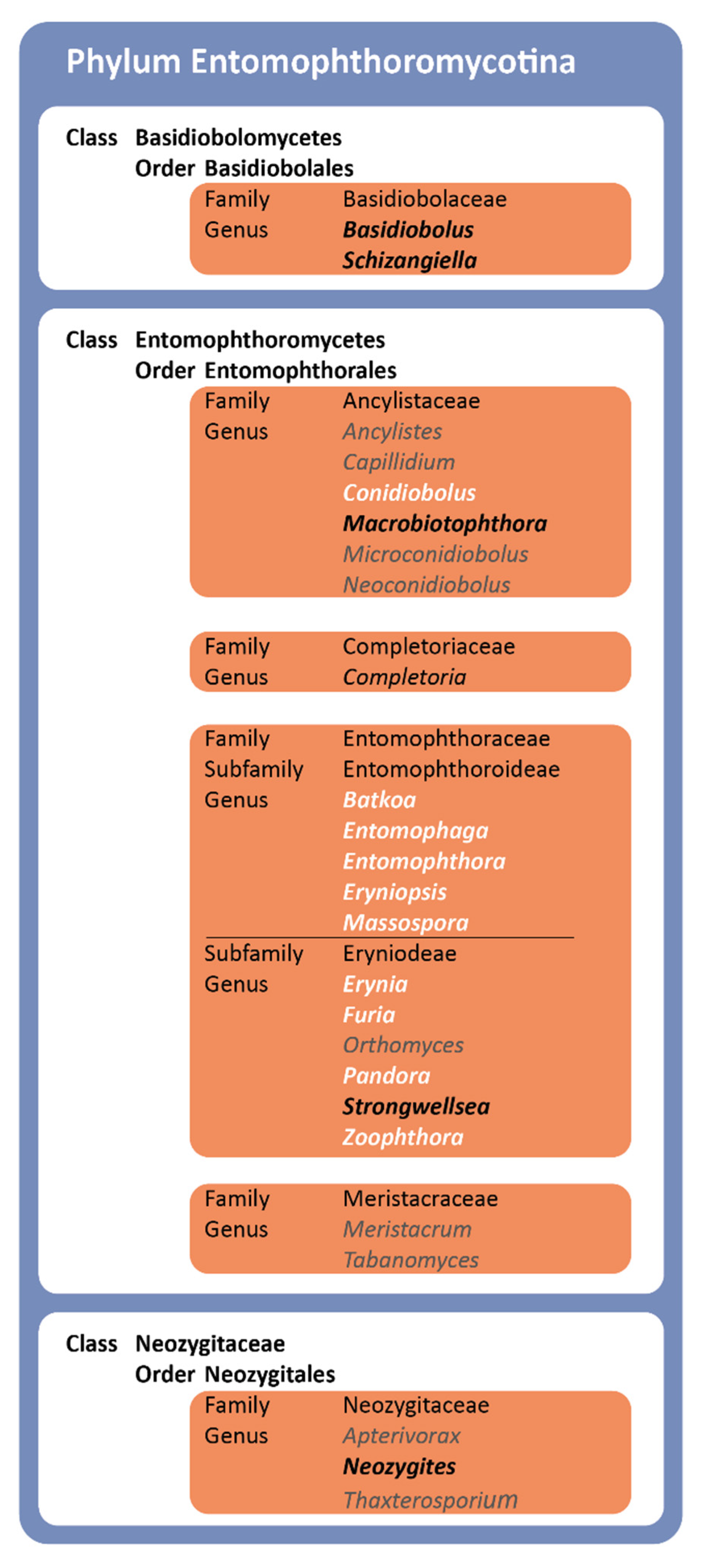
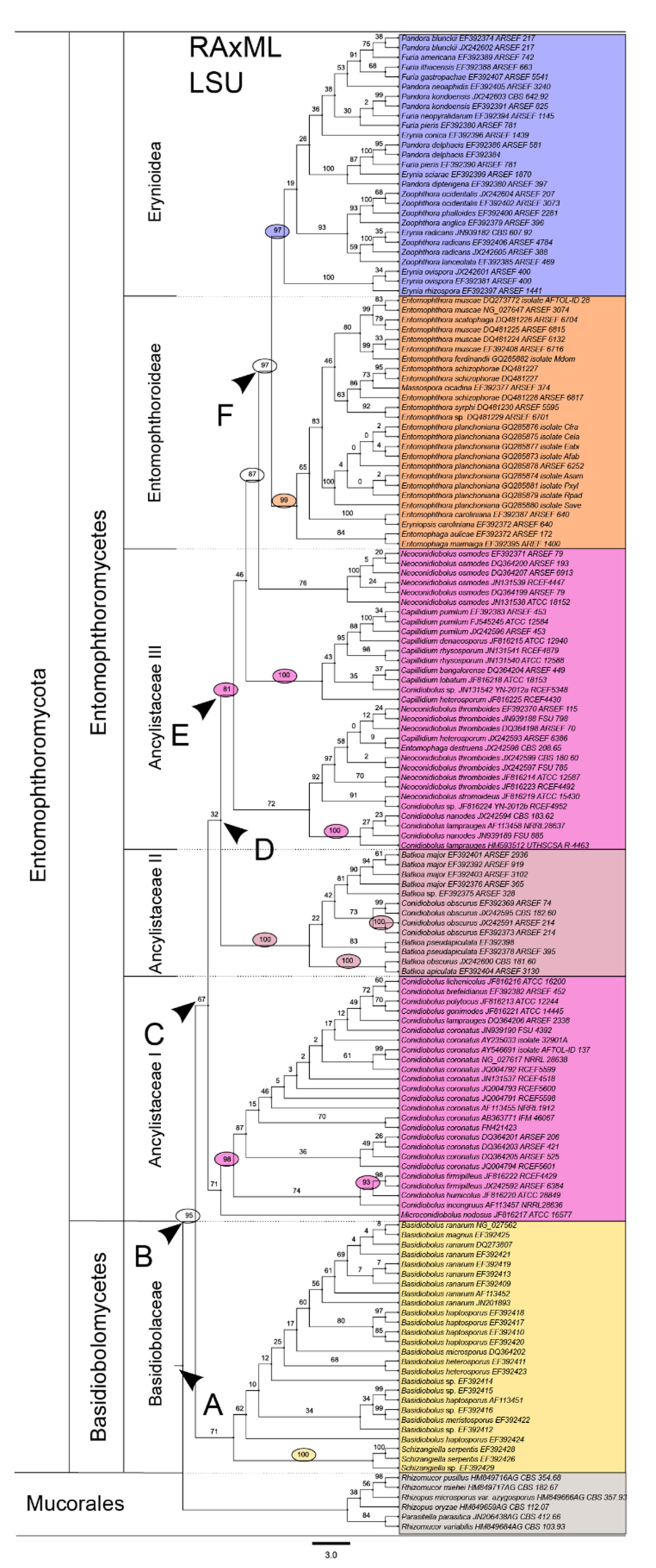
3.1. Revised Phylogeny of the Entomophthoromycotina
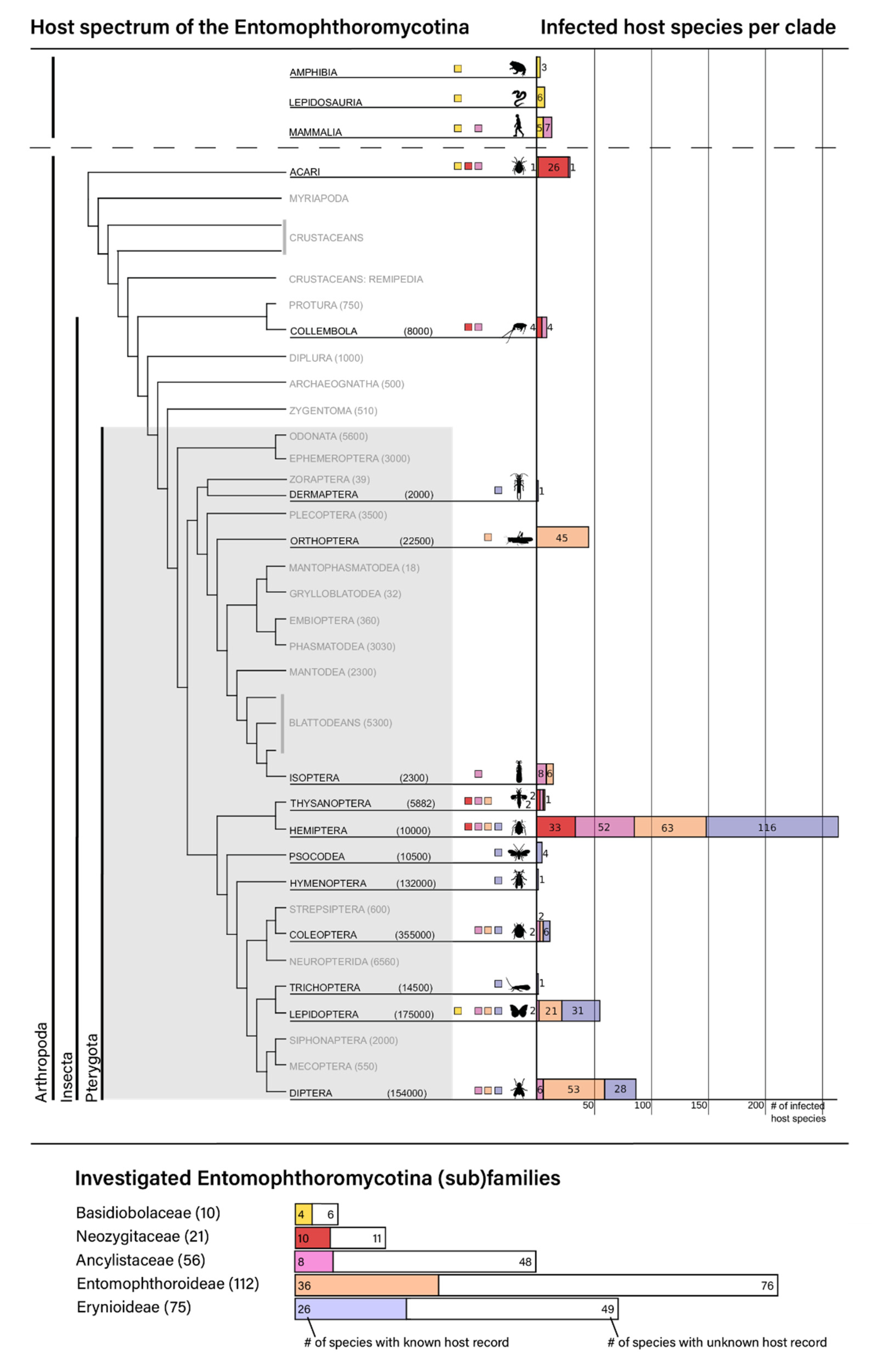
3.2. Patterns of Host Specificity of the Entomophthoromycotina
3.3. New Type Species for Conidiobolus: C. coronatus
| Type: | Conidiobolus Bref. 1884. |
| Synonymy: | Boudierella Costantin, Bull. Soc. mycol. Fr. 13: 40 (1897) [48]. Delacroixia Sacc. and P. Syd., Syll. fung. (Abellini) 14(1): 457 (1899) [49]. |
| Classification: | Entomophthoromycotina, Entomophthoromycetes, Entomophthorales, Ancylistaceae. |
- Comprises Conidiobolus clade C: Ancylistaceae I (see Figure 3).
- Note: Comparing Conidiobolus coronatus AF113418 with Macrobiotophthora vermicola AF052400 at the level of small subunit (18S, SSU) rDNA nucleotide sequences, the deviation was found to be high: identity = 824/967 (85%) and gaps = 34/967 (3%), which was lower than 90% SSU sequence similarity as proposed by [50] justifying classification in a new family.
- Lectotype: Conidiobolus utriculosus Bref. 1884 (Clements and Shear, Gen. Fungi: 239. 1931) [46].
- Lectotype: Costantin, Bull. Soc. mycol. Fr. 13: 40 (1897): see Pl. 4, Figures 1–10, ; Pl. 5, Figures 11–17 in [48].
- Type: JMRC:SF:11506 (metabolically inactive culture), Sweden, isol. M. Gustafsson (B42536, here designated).
- Isotypes: NRRL 28638; CBS 209.66; ATCC 28846.
- Whole genome sequence PRJNA67455 available from: http://www.ncbi.nlm.nih.gov/bioproject/PRJNA67455 (accessed on 23 February 2016), see also [52].
- Reference sequences: AF113418 (SSU), AF113456, AY546691 and NG_027617 (LSU), AY997041 (ITS1-5.8S-ITS2), DQ275337 (elongation factor 1-alpha), DQ294591 (RNA polymerase II subunit RPB1), DQ302779 (RNA polymerase II subunit RPB2).
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gryganskyi, A.P.; Humber, R.A.; Smith, M.E.; Hodge, K.; Huang, B. Phylogenetic lineages in Entomophthoromycota. Mol. Phylogen. Evol. 2013, 65, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Gryganskyi, A.P.; Humber, R.A.; Smith, M.E.; Miadlikowska, J.; Wu, S.; Voigt, K.; Walther, G.; Anishchenko, I.M.; Vilgalys, R. Molecular phylogeny of the Entomophthoromycota. Mol. Phylogen. Evol. 2012, 65, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Elya, C.; Lok, T.C.; Spencer, Q.E.; McCausland, H.; Martinez, C.C.; Eisen, M. Robust manipulation of the behavior of Drosophila melanogaster by a fungal pathogen in the laboratory. eLife 2018, 7, e34414. [Google Scholar] [CrossRef]
- White, M.M.; James, T.; O'Donnell, K.; Cafaro, M.; Tanabe, Y.; Sugiyama, J. Phylogeny of Zygomycota based on nuclear RNA. Mycologia 2006, 98, 872–884. [Google Scholar] [CrossRef]
- Carruthers, R.I.; Hural, K. Fungi as naturally occuring entomopathogens. UCLA Symp. Mol. Cell Biol. 1990, 112, 115–138. [Google Scholar]
- De Fine Licht, H.H.; Jensen, A.B.; Eilenberg, J. Comparative transcriptomics reveal host-specific nucleotide variation in entomophthoralean fungi. Mol. Ecol. 2017, 26, 2092–2110. [Google Scholar] [CrossRef]
- De Ruiter, J.; Arnbjerg-Nielsen, S.F.; Herren, P.; Høier, F.; De Fine Licht, H.H.; Jensen, K.H. Fungal artillery of zombie flies: Infectious spore dispersal using a soft water cannon. J. R. Soc. Interface 2019, 16, 20190448. [Google Scholar] [CrossRef]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; De Fine Licht, H.H.; et al. Psychoactive plant- and mushroom-associated alkaloids from two behavior modifying cicada pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef]
- Arnesen, J.A.; Małagocka, J.; Gryganskyi, A.; Grigoriev, I.V.; Voigt, K.; Stajich, J.E.; De Fine Licht, H.H. Early diverging insect-pathogenic fungi of the order Entomophthorales possess diverse and unique subtilisin-like serine proteases. G3 Genes Genomes Genet. 2018, 8, 3311–3319. [Google Scholar] [CrossRef] [Green Version]
- Humber, R.A. Evolution of entomopathogenicity in fungi. J. Invertebr. Pathol. 2008, 98, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Rothhardt, J.E.; Schwartze, V.U.; Voigt, K. Entomophthorales. In Molecular Detection of Human Fungal Pathogens; Liu, D., Ed.; Taylor & Francis CRC Press: Boca Raton, FL, USA, 2011; pp. 723–734. [Google Scholar]
- El-Shabrawi, M.H.; Arnaout, H.; Madkour, L.; Kamal, N.M. Entomophthoromycosis: A challenging emerging disease. Mycoses 2014, 57, 132–137. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.; Figueras, M.; Vitale, R. Atlas of Clinical Fungi: The Ultimate Benchtool for Diagnostics. Filamentous Ascomycetes C-Z, Part 2, 4th ed.; Foundation Atlas of Clinical Fungi: Hilversum, The Netherlands, 2020. [Google Scholar]
- Radhakrishnan, N.; Sachdeva, A.; Oberoi, J.; Yadav, S.P. Conidiobolomycosis in relapsed acute lymphoblastic leukemia. Pediatr. Blood Cancer 2009, 53, 1321–1323. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J. Taxonomy of fungi causing mucormycosis and entomophthoramycosis (zygomycosis) and nomenclature of the disease: Molecular mycologic perspectives. Clin. Infect. Dis. 2012, 54, S8–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durel, L.; Estrada-Peña, A.; Franc, M.; Mehlhorn, H.; Bouyer, J. Integrated fly management in European ruminant operations from the perspective of directive 2009/128/EC on sustainable use of pesticides. Parasitol. Res. 2015, 114, 379–389. [Google Scholar] [CrossRef]
- Stern, V.M.; Smith, R.F.; van den Bosch, R.; Hagen, K.S. The integrated control concept. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, S.E.; Ellsworth, P.C.; Frisvold, G.B. Economic value of biological control in Integrated Pest Management of managed plant systems. Annu. Rev. Entomol. 2015, 60, 621–645. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T.; et al. A genome-scale phylogeny of the kingdom fungi. Curr. Biol. 2021, 31, 1653–1665. [Google Scholar] [CrossRef]
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-y.; Voigt, K. Molecular characters of zygomycetous fungi. In Molecular Identification of Fungi; Gherbawy, Y., Voigt, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 461–488. [Google Scholar]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lucking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Nie, Y.; Yu, D.-S.; Wang, C.-F.; Liu, X.-Y.; Huang, B. A taxonomic revision of the genus Conidiobolus (Ancylistaceae, Entomophthorales): Four clades including three new genera. MycoKeys 2020, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Ze'ev, I.; Kenneth, R.G. Features-criteria of taxonomic value in the Entomophthorales: 1. A revision of the Batkoan classification. Mycotaxon 1982, 14, 393–455. [Google Scholar]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef] [PubMed]
- De Fine Licht, H.H.; Hajek, A.E.; Eilenberg, J.; Jensen, A.B. Utilizing genomics to study entomopathogenicity in the fungal phylum entomophthoromycota: A review of current genetic resources. In Advances in Genetics; Lovett, B., St. Leger, R.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 94, pp. 41–65. [Google Scholar]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Iriny, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 151–187. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Altschul, S.F.; Boguski, M.S.; Gish, W.; Wootton, J.C. Issues in searching molecular sequence databases. Nat. Genet. 1994, 6, 119–129. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, 20–25. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Henikoff, S.; Henikoff, J.G. Amino acid substitution matrices from protein blocks. Proc. Natl. Acad. Sci. USA 1992, 89, 10915–10919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Towns, J.; Cockerill, T.; Dahan, M.; Foster, I.; Gaither, K.; Grimshaw, A.; Hazlewood, V.; Lathrop, S.; Lifka, D.; Peterson, G.D.; et al. XSEDE: Accelerating scientific discovery. Comput. Sci. Eng. 2014, 16, 62–74. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Voigt, K.; A, M.; Wostemeyer, J. Phylogeny of zygomycetes: A molecular approach towards systematics of Mucorales. Cour. Forsch. Inst. Senckenberg 1999, 215, 207–213. [Google Scholar]
- Beutel, R.; Yavorskaya, M.; Mashimo, Y.; Fukui, M.; Meusemann, K. The phylogeny of Hexapoda (Arthropoda) and the evolution of megadiversity. Proc. Arthropod. Embryol. Soc. Jpn. 2017, 51, 1–15. [Google Scholar]
- Brefeld, O. Conidiobolus Utriculosus und Minor; Untersuchungen aus der Gesammtgebiete der Mykologie Arthur Felix: Leipzig, Germany, 1884; Volume 6, pp. 35–78. [Google Scholar]
- Clements, F.; Shear, C.L. The Genera of Fungi; H.W. Wilson Co.: New York, NY, USA, 1931. [Google Scholar]
- Mendoza, L.; Vilela, R.; Voelz, K.; Ibrahim, A.S.; Voigt, K.; Lee, S.C. Human fungal pathogens of Mucorales and Entomophthorales. Cold Spring Harb. Perspect. Med. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Costantin, S. Sur une Entomophthoree nouvelle. Bull. Soc. Mycol. Fr. 1897, 13, 38–43. [Google Scholar]
- Saccardo, P.; Sydow, P. Supplementum Universale, Pars IV. Syll. Fung. 1899, 14, 1–1316. [Google Scholar]
- Hoffmann, K.; Telle, S.; Walther, G.; Eckhart, M.; Kirchmair, M.; Prillinger, H.; Prazenica, A.; Newcombe, G.; Dölz, F.; Tamás, P.; et al. Diversity, genotypic identification, ultrastructural and phylogenetic characterization of zygomycetes from different ecological habitats and climatic regions: Limitations and utility of nuclear ribosomal DNA barcode markers. In Current Advances in Molecular Mycology; Gherbawy, Y.M., Mach, R.L., Rai, M., Eds.; Nova Science Pub: New York, NY, USA, 2008; pp. 263–312. [Google Scholar]
- Batko, A. Notes on entomophthoraceous fungi in Poland. Entomophaga 1964, 2, 129–131. [Google Scholar]
- Chang, Y.; Wang, S.; Sekimoto, S.; Aerts, A.L.; Choi, C.; Clum, A.; LaButti, K.M.; Lindquist, E.A.; Yee Ngan, C.; Ohm, R.A.; et al. Phylogenomic analyses indicate that early fungi evolved digesting cell walls of algal ancestors of land plants. Genome Biol. Evol. 2015, 7, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, Q.D.; Blackwell, M. Fungus-Insect Relationships; Columbia University Press: New York, NY, USA, 1984. [Google Scholar]
- Grimaldi, D.A.; Engel, M.S. Evolution of Insects; Cambridge University Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola); Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Berbee, M.L.; Taylor, J.W. Dating the molecular clock in fungi–how close are we? Fungal Biol. Rev. 2010, 24, 1–16. [Google Scholar] [CrossRef]
- Elya, C.; De Fine Licht, H.H. The genus Entomophthora: Bringing the insect destroyers into the twenty-first century. IMA Fungus 2021, 12, 34. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Möckel, L.; Meusemann, K.; Misof, B.; Schwartze, V.U.; De Fine Licht, H.H.; Voigt, K.; Stielow, B.; de Hoog, S.; Beutel, R.G.; Buellesbach, J. Phylogenetic Revision and Patterns of Host Specificity in the Fungal Subphylum Entomophthoromycotina. Microorganisms 2022, 10, 256. https://doi.org/10.3390/microorganisms10020256
Möckel L, Meusemann K, Misof B, Schwartze VU, De Fine Licht HH, Voigt K, Stielow B, de Hoog S, Beutel RG, Buellesbach J. Phylogenetic Revision and Patterns of Host Specificity in the Fungal Subphylum Entomophthoromycotina. Microorganisms. 2022; 10(2):256. https://doi.org/10.3390/microorganisms10020256
Chicago/Turabian StyleMöckel, Lars, Karen Meusemann, Bernhard Misof, Volker U. Schwartze, Henrik H. De Fine Licht, Kerstin Voigt, Benjamin Stielow, Sybren de Hoog, Rolf G. Beutel, and Jan Buellesbach. 2022. "Phylogenetic Revision and Patterns of Host Specificity in the Fungal Subphylum Entomophthoromycotina" Microorganisms 10, no. 2: 256. https://doi.org/10.3390/microorganisms10020256