
Control Strategies of Clubroot Disease Caused by Plasmodiophora brassicae

1
Faculty of Agricultural and Environmental Sciences, University of Rostock, Satower Str. 48, 18059 Rostock, Germany
2
Faculty of Agriculture and Food Sciences, University of Applied Sciences Neubrandenburg, Brodaer Str. 2, 17033 Neubrandenburg, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(3), 620; https://doi.org/10.3390/microorganisms10030620
Submission received: 3 January 2022
/
Revised: 17 February 2022
/
Accepted: 11 March 2022
/
Published: 14 March 2022
(This article belongs to the Special Issue Plant Pathogens in a Global Change Context)
Abstract
:The clubroot disease caused by the soil-borne pathogen Plasmodiophora brassicae is one of the most important diseases of cruciferous crops worldwide. As with many plant pathogens, the spread is closely related to the cultivation of suitable host plants. In addition, temperature and water availability are crucial determinants for the occurrence and reproduction of clubroot disease. Current global changes are contributing to the widespread incidence of clubroot disease. On the one hand, global trade and high prices are leading to an increase in the cultivation of the host plant rapeseed worldwide. On the other hand, climate change is improving the living conditions of the pathogen P. brassicae in temperate climates and leading to its increased occurrence. Well-known ways to control efficiently this disease include arable farming strategies: growing host plants in wide crop rotations, liming the contaminated soils, and using resistant host plants. Since chemical control of the clubroot disease is not possible or not ecologically compatible, more and more alternative control options are being investigated. In this review, we address the challenges for its control, with a focus on biological control options.
Keywords:
Brassicaceae; biological control; canola; clubroot; lime fertilizer; oilseed rape; soilborne disease1. Introduction
Clubroot caused by the soil-living, obligate biotrophic protist Plasmodiophora brassicae Woronin belongs to the most devastating diseases of cruciferous crops worldwide [1,2]. As Plasmodiophoridae the pathogen belongs to the Rhizaria—a group of protists [3,4]. Together with the protist groups Stramenopiles (also called Heterokonta) and Alveolata, the Rhizaria represent the eukaryotic supergroup SAR, which is a highly diverse group of eukaryotic organisms [5]. Clubroot causes root tumors which lead to the disruption of water and nutrient uptake. As a consequence, infection can result in wilting and stunting. Yield losses of oilseed rape from 10% yield reduction to a total yield loss occur including plant losses and reduced seeds per pod [6]. The development of disease and cellular changes of host plants after infection were recently described in detail by [7,8].
Successful management of the disease is difficult as chemical control of clubroot is not allowed or not successful. In soil, the pathogen survives as extremely robust, thick-walled resting spores. Those spores can be easily transported from field to field via infested soil on machinery, by animals, water, or wind [9]. In this way, the spread of the disease can occur rapidly within a region. Spreading over continents already took place very early on. In the 19th century, clubroot was first described in Russia [10]. It is assumed that clubroot arrived with immigrants, colonization movement, and early settlers from Europe to North America [7], South America [11], and Australia [12], who probably carried the pathogen with infested food and fodder or to which contaminated soil adhered. At present, clubroot is reported from all continents (except Antarctica) and more than 75 countries [13]. It is very likely that the disease is distributed throughout the world wherever cruciferous crops are grown or cruciferous vegetation is available. Regions where the pathogen occurs with high population densities are mainly humid, temperate areas [14]. The expansion of cropping areas and proximal crop rotation contribute to an increase in clubroot [1].
An important factor in clubroot management is the deployment of effective host plant resistance. Not all known R genes are active against all races and resistance can be eroded by new pathogen races. In this review, we do not address the details of resistance to clubroot; instead, we want to focus on the control options available in agricultural practice. Crop management practices are widely used to combat the disease. However, control measures such as crop rotation and raising soil pH are not enough in reducing the disease. To improve control strategies, in recent years, research on biological control measures has emerged as an increasingly important role in clubroot management. In addition, strengthening of the plants by plant growth stimulants has gained increasing significance.
In this review, we summarize the different options for combating clubroot, focusing on biological control.
2. Environmental Parameters Influencing Plasmodiophora brassicae Development
If clubroot host plants are repeatedly grown in the field, the resting spores of the pathogen can accumulate in the soil [15]. Those spores are robust and can persist in the soil for many years, leading to an infestation of cabbage plants even after several years of cultivation [14]. Wallenhammar [16] calculated that the level of infestation in the field only decreases below the detection level after 17.3 years. However, a more than 2-year cultivation break and a diverse rotation pattern can reduce resting spores in soil [17,18].
In order to combat the pathogen P. brassicae, knowledge of the optimal environmental conditions that lead to an outbreak of the disease is a prerequisite. The causal agent of clubroot disease P. brassicae was described in 1878 by Woronin [10]; and more than 50 years later its lifecycle was shown for the first time [19]. Since then, the environmental conditions that are optimal for the disease and by which it is promoted have been studied in detail (for a review see: [14,20]).
Temperature and soil moisture play important roles in the development of the disease. The strongest infection rate and symptom severity occur at 21–25 °C [21,22,23]. At temperatures significantly below 20 °C clubroot severity is considerably reduced [21,22,23]. High soil moisture, especially during the first two weeks after inoculation, or during the first and second infections, is necessary for the successful development of the disease [20,21,24]. Moreover, wet soil conditions favor the spread of motile zoospores [25]. The soil type has a weak influence on the infestation intensities. However, it was shown that sandy soils and soils with low humic content are suppressive to disease [26] and that clubroot severity was lower in sand than in loamy soils or clay [16,27,28].
Another important factor for clubroot development is the soil pH. A low pH value (pH 5 to <7) in the soil promotes spore germination [29,30] and usually results in a more severe infestation. As the pH value increases, from about 7.2 onwards, there is hardly any new infection in most cases [31,32,33].
In conclusion, the most conducive conditions for clubroot development are high summer temperatures combined with light acidic soils and good soil moisture during the first weeks after sowing. It can be assumed that this will have considerable regional and between-year effects in the occurrence of clubroot disease.
3. Agricultural Practices
In general, chemical control of soil-borne diseases is difficult and cost intensive and is also not allowed in many countries because of the ecological impact. Of the currently worldwide registered chemicals, the oomycete fungicides fluazinam and cyazofamid reduce clubroot [34,35]. They are registered for cabbage crops in some countries, but they are not allowed in EU countries for clubroot control. The use of these agents is difficult and expensive because they have to be drenched into the soil to be effective against clubroot. Therefore, appropriate crop management practices are important measures for clubroot control.
3.1. Plant Resistance
Sowing of resistant host plants is an effective way to suppress the disease. Several resistant loci have been identified by quantitative trait locus mapping in Brassica napus and B. rapa (for a review see: [36,37,38]). The most widely used resistance loci originate from B. rapa; however, these loci do not confer resistance to all P. brassicae pathotypes or, even more alarming, pathotypes can overcome the resistance. Therefore, resistance breeding remains an important tool to combat P. brassicae [7,39].
3.2. Crop Rotation and Tillage
Due to the high market prices of rapeseed, the area under cultivation has increased and the crop rotation has narrowed [40]. This led to increasing problems with the pathogen and, therefore, another important factor in the control of clubroot is the use of crop rotation. More than 2 years’ break or diverse crop rotations can reduce severe clubroot epidemics [15,40]. Moreover, also the preceding crop might have an influence on the clubroot infection. A recent study compared different preceding crops before oilseed rape was planted. The results yielded a 40 and 50% reduction in clubroot disease index and incidence rate, respectively, when soybean was planted before oilseed rape [18]. In addition, microbiome analyses of soil showed that within the soybean—oilseed rape soil—more bacteria and fungi with known biocontrol functions were detectable than in maize—or rice—oilseed rape soil [18]. It has recently been shown that resting spores are highly sensitive to UV light [41]. Therefore, tillage measures, that bring resting spores to the soil surface for exposure to sunlight, could be an effective way to support clubroot management.
3.3. Field Sanitation
An aspect often overseen by practitioners is that resting spores are very easily carried with soil particles, e.g., with machinery, with boots, with surface water, or with animals, thus, spreading the disease. Furthermore, it has also been shown that resting spores are transported by wind-borne dust or soil erosion from field to field [42]. Therefore, activities that can reduce the transport of spores from field to field as sanitization of farmers’ machinery or measures that help prevent soil erosion thus prevent initial infestation of fields [43,44].
3.4. Soil pH
In addition, pH management by liming is an efficient way of controlling clubroot disease. Although liming is traditionally used as a control measure, it has been discussed as highly controversial [47,48,49]. One reason is that the term “liming” means the application of different formulations of lime. Mostly lime with varying proportions of calcium carbonate (often mixed with Mg2+) [50,51] cyanamide [33,48,52] is used. Moreover, a combination of calcium carbonate and calcium sulfate [53] or calcium hydroxide [48] is applied. Only rarely has calcium oxide, also called burnt lime or quicklime, been applied [12,54]. Few studies have attempted to compare different types of lime [26]. Besides the calcium concentration, other factors such as the amount of lime, date of application, and soil pH may have effects on clubroot development. However, Niwa et al. [55] showed that it is not the calcium effect that is decisive but rather the soil pH. Germination of the resting spores is drastically reduced by a neutral soil pH. Because of the diversity of substances and soil differences, comparability of most field trials or greenhouse studies exploring the effect of lime on clubroot is difficult, and therefore, results are highly diverse.
With the aim of recognizing the effect of different lime fertilizers on clubroot development in oilseed rape, we conducted a greenhouse experiment with up to 37 one-week-old seedlings per lime type under strictly controlled conditions as described in [6]. We used a variety of lime fertilizers commonly applied in practice (Table 1). Before planting, the substrate was treated with the lime formulations. Rough-grained lime materials were finely crushed and passed through a 2 mm-mesh sieve before being added. Calcium oxide was added at a rate of 10 tonnes CaO ha−1, all other fertilizers at 13 tonnes ha−1. The P. brassicae isolates were sampled on heavily infested fields in North Germany and propagated in susceptible oilseed rape cultivar Avatar (Norddeutsche Pflanzenzucht Hans-Georg Lembke KG) as described in [6]. We used three isolates from different fields in Northern Germany to test the liming effect on a broader basis. Resting spores were isolated from homogenized clubbed roots and spore concentration was estimated microscopically with a hemocytometer after the spores were stained with Evan’s blue [56]. Spore concentration was adjusted to a value of 2 × 106–107 spores per ml. One milliliter of the suspension was placed directly to the seedling.
To determine soil pH, mixed soil samples (20 mL) were prepared from all pots of each variant weekly. To each soil sample, 50 mL CaCl2 (0.01 M) was added and mixed thoroughly. After 2 h at room temperature, the samples were filtered and the pH values were determined. Six to seven weeks after inoculation, roots were removed from the pots, gently washed, and the disease rating was recorded on a 4-classes scale from 0 to 3 and combined as the disease severity index (DSI) [6,57].
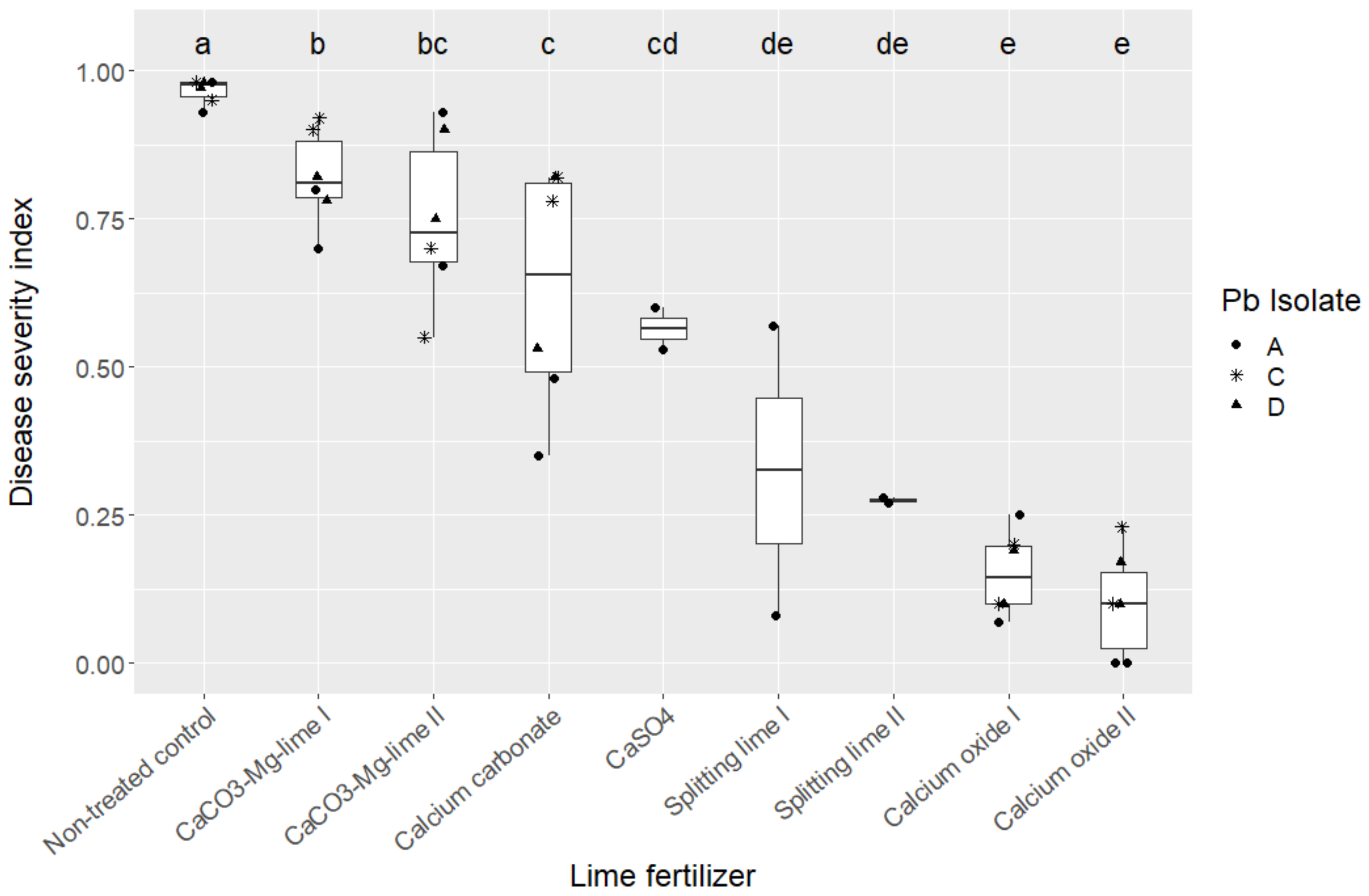
Significant differences between the lime fertilizers on the development of clubroot were found by multiple comparisons of treatments by means using Tukey’s honestly significant difference post hoc test on ranks of data. Pearson’s correlation coefficient was used to test the association between DSI and pH value of the soil. Statistical analysis was performed using the software R version 3.6.1 (R core Team, 2019) and basic packages were complemented by the packages “agricolae” [58]. The results clearly show that the plants that grew in the most alkaline soil had the least clubroot damage (Figure 1, Table 2). Crucial is a soil pH > 0.68 during the seedling stage of the plants when they are particularly susceptible to infection. Due to the rapid reaction with water, calcium oxide reaches this pH value very quickly and thus contributes to the reduction in clubroot severity, while the other lime fertilizers do not raise the soil pH so early after application. A similar result was shown by Fox et al. [59], who used calcium hydroxide as lime, which is produced by the reaction of calcium oxide with water.
4. Clubroot Control Using Beneficial Microorganisms
The lack of effective control measures against P. brassicae makes it necessary to explore other, novel control options. The application of biological control measures could help to reduce soil-borne pathogens in particular. However, the complex life cycle of P. brassicae makes it difficult to apply biological control mechanisms against this pathogen. At least three phases can be used for control: (i) germination of the resting spores and/or secondary spores, which initiate (ii) primary infection of the root hairs and secondary infection of the root cortex; (iii) antagonism/competition against the developing pathogen within the host root tissue. In addition, resistance induction in host plants and changes in microbial communities in the rhizosphere soil could be biological control options [60].
Biocontrol agents that have been explored are bacteria or fungi including oomycetes. The mechanisms mostly are parasitism, antagonism by toxic/antibiotic secondary metabolites, and/or competition. Many studies have illustrated the biological control potential against soil fungi in sensu stricto. This refers to the direct antagonistic or inhibitory effect on the pathogen and not to an indirect effect such as plant growth promotion effect or induction of plant resistance [61,62].
Organisms such as, e.g., Trichoderma spp. and Bacillus subtilis sensu lato, are commercially employed in many control agents against a diverse group of plant pathogens [63,64,65,66,67]. There are numerous examples that illustrate that excellent control results can be achieved in in vitro trials [62,68]. Whereas in field trials, those successful control results often cannot be confirmed [69]. Therefore, for a successful control option, detailed research work must be preceded.
4.1. Antagonistic Bacteria
Bacteria of the Bacillus subtilis species complex are well studied for their biocontrol activity against plant pathogens. These bacteria have the potential to produce many hydrolytic enzymes and diverse secondary metabolites with antimicrobial properties [64,65]. One very well-characterized biological control agent patented strain in China is B. subtilis XF-1. Like other Bacillus strains, it produces fengycins, which are a group of nonribosomal lipopeptides. These metabolites have fungitoxic activities and are involved in the biocontrol effect of many Bacillus species (reviewed by [70,71]). Resting spores of P. brassicae directly treated with fengycins collapsed, and the cell contents leaked out [72]. Irrespective of this, the mode of action of fengycin was demonstrated with the B. subtilis strain NCD-2, which showed a reducing effect on clubroot; whereas, by using fengycin, defect mutants showed no effect against the clubroot pathogen [73]. In another experiment, Chinese cabbage seeds were soaked in a fengycin-producing B. subtilis XF-1 bacterial suspension and the bacterial culture disease incidence was reduced by 40% and 69%, respectively [74]. The authors demonstrated that the B. subtilis XF-1 treatment at an early stage of seedling development had the most positive effect. In the field, B. subtilis XF-1 reduced the disease index by about 17% [75]. We can therefore assume that in heavily infested fields, a reduction in this small amount is not useful to farmers.
The B. amyloliquefaciens strain QST713 (formerly B. subtilis strain) is registered for commercial uses (Serenade®) in many countries [76]. In Canada, it was tested against P. brassicae; however, in the field, the control success is limited [77]. Detailed greenhouse studies showed that the biofungicide Serenade® applicated as soil drench reduced the clubroot disease incidence substantially [28,78,79]. Recently, besides a new strain of B. amyloliquefaciens, another member of the genus, Bacillus, B. velezensis, was described as a biocontrol agent against P. brassicae [68].
Several species of the bacterial genus Lysobacter are known for their activity against soil pathogens. These bacteria synthesize many hydrolytic enzymes and antimicrobial compounds and several commercial preparations are available against soilborne fungal pathogens [80]. By screening bacterial strains from vegetable rhizosphere soil [81], Lysobacter antibioticus strains were isolated whose culture filtrates reduced clubroot severity on Chinese cabbage after application as a soil drench or seed treatment. Another Streptomyces strain, S. platensis 3–10, was used to optimize the culture medium and reached an inhibition of resting spore germination up to 80% [82]. Recently, a strain of Bacillus cereus, MZ-12, isolated from the rhizosphere soil of symptomless B. campestris (pak choi) showed an inhibitory effect on germination of resting spores. Co-inoculation of the pak choi plants with P. brassicae spores and MZ-12 resulted in a 64% reduction in clubroot gall formation [83].
It has been shown repeatedly that the results of in vitro and greenhouse experiments cannot be achieved with field experiments. For example, the bacterial strain Zhihengliuella aestuarii B18 isolated from rhizosphere of Brassica juncea showed a control efficiency of 63.4% against clubroot in greenhouse tests, whereas the control effect in the field was only 49.7% [84]
In addition to free-living microorganisms in the rhizosphere or epiphytic-living microorganisms, endophytic-living microorganisms can also contribute to biological control. Ahmed et al. [85] provide an overview of research carried out with endophytic bacteria and fungi as biocontrol agents. Mostly, endophytic bacteria derived from the rhizosphere enter the plant and colonize the plant tissue without any negative effect on the plant [86]. In many cases, this form of bacterial colonization contributes to the promotion of plant growth by different mechanisms [87]. However, the antagonistic activity by endophytic actinobacteria against clubroot has been reported by Lee et al. [88]. They isolated 81 actinobacterial strains from the surface-sterilized root tissue of Chinese cabbage. Among them, they selected three strains that showed in vivo biocontrol activities against P. brassicae. Two of these strains were identified as Microbispora rosea, the third strain as Streptomyces olivochromogenes [88]. Wang et al. [89] tested 63 actinobacteria strains isolated from the rhizosphere of Chinese cabbage by measuring the inhibition of the germination of P. brassicae resting spores. This resulted in six strains that were used in greenhouse and field trials against clubroot. The strain A316 showed high control values of 73.69% in a glasshouse experiment and 65.91% in a field experiment [89].
A recent paper by Wei et al. [90] presents a detailed analysis of bacterial metabolites with biocontrol functions against clubroot. The work shows that co-culturing bacterial species of different genera produces more relevant metabolites than culturing bacterial species of the same genera. The results reveal that bacterial interactions between genera promote the production of biocontrol active substances.
Increasingly, microbiome studies of the rhizosphere are showing that the microbial communities are complex and highly variable [87]. Recently, a study was presented comparing heavily P. brassicae-contaminated soils with weakly contaminated soils. The results showed that the bacterial communities of both soil types differed significantly [91]. Furthermore, the study showed that certain groups of bacteria were mainly found in weakly contaminated soils [91]. Such studies suggest that individual bacterial species cannot generally act as control organisms. Instead, in each soil type, the composition of microbial communities differs and one must assume and consider, for further work, that groups of organisms act together.
4.2. Antagonistic Fungi
The soil-borne ascomycetous fungus Phoma glomerata (current name: Didymella glomerata) has been described as a plant pathogen [92,93] as well as a potential biocontrol agent [94,95]. The strain P. glomerata no. 324 produces the secondary metabolite epoxydon. Although this substance showed very weak antifungal activity, it reduced clubroot symptoms on Chinese cabbage after spraying over the infested soil (30 mL extract solution containing 250 µg mL−1 per 180 mL pot) completely [96]. However, this work was conducted in the 1990s and has never been commercialized.
The fungus Clonostachys rosea f. sp. catenulata (syn. Gliocladium catenulatum) is widely known as a biocontrol agent. In many countries, it is commercialized as the biofungicide Prestop® with control activity against several soil-borne plant pathogens in various crops [97]. Neither the fungus C. rosea nor the biofungicide Prestop® had any effect on the germination and viability of the resting spores of P. brassicae [98]. However, soil drench treatments on B. napus seedlings 7 to 14 days after seeding resulted in a reduction in clubroot severity of about 90%. The fungus colonized the plant root system and, in this way, suppressed clubroot. In addition, it appeared to induce plant resistance since some induced plant resistance-associated genes were up-regulated [98].
The fungal genus Trichoderma comprises several species that are well studied as biological control agents against various plant pathogens [66,67]. In greenhouse pot experiments, the control efficiency of T. harzianum strain T4 against P. brassicae in Chinese cabbage was about 79% [99]. Another work showed the control effect of T. harzianum strain LTR-2 in Chinese cabbage in the field. The disease incidence was lowered from 96.7% (untreated control) to 51.3% (seeds treated with spores of T. harzianum LTR-2) [100].
Endophytic (mutualistic) fungi grow within their host plants tissue without causing visible disease symptoms [101]. They may have beneficial effects on the plant via plant growth promotion or by suppressing plant pathogens or pests [102,103]. The ascomycetous soil fungus Heteroconium chaetospira has been isolated from cabbage roots and was described as an endophytic root fungus growing throughout the cortical cells [104,105]. In greenhouse experiments, H. chaetospira reduced clubroot in Chinese cabbage plants by 90 to 100% after inoculation with low to moderate P. brassicae resting spore concentrations (up to 105 spores per g of soil). Severely diseased plants after inoculation with 106 spores per g of soil could not be protected by the endophytic fungus [106]. In field experiments, the disease reduction was lower; however, there was no reduction effect at high soil moisture [106]. Further investigations showed that H. chaetospira induced resistance reactions in Brassica napus plants. The phenyalanine ammonia lyase (PAL) activity, which was increased by H. chaetospira, and the upregulated transcript levels of several genes known to be involved in inducing plant resistance (ethylene/jasmonic acid synthesis, PR-2 protein, auxin biosynthesis) served as indicators for the resistance reaction [107]. Another endophytic fungal genus associated with plant roots is Acremonium. The species A. alternatum can colonize root cells of Brassicaceae. When Arabidopsis plants were co-inoculated with A. alternatum and P. brassicae, the disease index of clubroot was reduced by up to 50% [108].
5. Conclusions
Clubroot management has always been a challenge for farmers and chemical as well as classical agronomic measures have not been fully successful. In the last 10 to 20 years, a great wealth of work has been carried out on the biological control of clubroot. Some of the tested biofungicides appear promising for controlling the pathogen. However, to date, none of these approaches has become established in practice. Several studies showed that under a high disease pressure, the control activity of biocontrol agents is too weak [77,106]. Nevertheless, it is important to pursue these efforts further. There are a number of microbial control agents against clubroot, but our knowledge of the modes of action is too low (competition, hyperparasitism, antibiosis, mixed modes with induced plant resistance?). Research into the mode of action is required to bring more agents to the approval stage. However, even more complex is the nature of soil microbial interactions and the role of microorganisms of the rhizosphere for plant health is still wildly unknown [109]. The host plant-associated microbiomes of the rhizosphere affect the development of soil pathogens and the diseases they transmit. In the past, this interaction has often been neglected in research. Instead, the focus was on individual pathogens and their host plants in their particular environment [110]. Recently, the number of studies on the effect of soil microbiome has increased [111]. Raaijmakers and Mazzola [112] discussed that the functional similarity of immune-suppressed soils across many agroecosystems suggests that it may be possible to develop disease-suppressive soil microbiomes using a universal approach. Microbiome engineering is being vigorously discussed as the biocontrol method of the future [113]. It is becoming increasingly clear that the composition of the rhizosphere microbiome is important [114]. It has been shown that the bacterial diversity of the seed microbiome of oilseed rape differs depending on the cultivar [115]. Should there be less emphasis on pathogen control in the future and more focus on strengthening and compensating plants and influencing plant development by plant growth-promoting rhizobacteria? [116]. The composition of the microbiome could play an important role here. Lebreton et al. [117] showed that microorganism communities of healthy and clubroot-diseased plants differ considerably. Much attention should therefore be paid to comparing different rhizosphere microbiomes in order to identify important microorganisms. Can we compose a suitable microbiome? We are still far from an effective clubroot biological control option, but it is becoming possible that interactions of microbial communities could make a general contribution to the control of soil-borne plant diseases [118].
Author Contributions
Conceptualization, C.S. and B.S.; investigation, S.R.; data curation, B.S.; writing—original draft preparation, C.S. and B.S.; writing—review and editing, C.S. and B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Ingolf Gliege, Rosa Minderlen and Diana Mercedes Sicard for technical assistance. Many thanks to Norddeutsche Pflanzenzucht Hans-Georg Lembke KG, Holtsee, Germany, for providing oilseed rape seeds and to Vereinigte Kalkwerke Dammann GmbH & Co. KG, Sassnitz, Germany, for providing lime fertiliser.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Zheng, X.; Koopmann, B.; Ulber, B.; Von Tiedemann, A. A global survey on diseases and pests in oilseed rape—Current challenges and innovative strategies of control. Front. Agron. 2020, 2, 590908. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.-Y. Phylogeny and classification of phylum Cercozoa (Protozoa). Protist 2003, 154, 341–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burki, F.; Kudryavtsev, A.; Matz, M.V.; Aglyamova, G.V.; Bulman, S.; Fiers, M.; Keeling, P.J.; Pawlowski, J. Evolution of Rhizaria: New insights from phylogenomic analysis of uncultivated protists. BMC Evol. Biol. 2010, 10, 377. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.; Lewis, R. Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: Contrasting cell organisation of sister phyla Cercozoa and Retaria. Protoplasma 2018, 255, 1517–1574. [Google Scholar] [CrossRef] [Green Version]
- Strehlow, B.; de Mol, F.; Struck, C. Risk potential of clubroot disease on winter oilseed rape. Plant Dis. 2015, 99, 667–675. [Google Scholar] [CrossRef]
- Hwang, S.-F.; Strelkov, S.E.; Feng, J.; Gossen, B.D.; Howard, R.J. Plasmodiophora brassicae: A review of an emerging pathogen of the Canadian canola (Brassica napus) crop. Mol. Plant Pathol. 2012, 13, 105–113. [Google Scholar] [CrossRef]
- Malinowski, R.; Truman, W.; Blicharz, S. Genius architect or clever thief-how Plasmodiophora brassicae reprograms host development to establish a pathogen-oriented physiological sink. Mol. Plant Microbe Interact. 2019, 32, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Botero-Ramirez, A.; Hwang, S.-F.; Strelkov, S.E. Plasmodiophora brassicae inoculum density and spatial patterns at the field level and relation to soil characteristics. Pathogens 2021, 10, 499. [Google Scholar] [CrossRef]
- Woronin, M. Plasmodiophora brassicae, Urheber der Kohlpflanzen-Hernie. Jahrb. Wiss. Bot. 1878, 11, 548–574. [Google Scholar]
- Botero, A.; García, C.; Gossen, B.D.; Strelkov, S.E.; Todd, C.D.; Bonham-Smith, P.C.; Pérez-López, E. Clubroot disease in Latin America: Distribution and management strategies. Plant Pathol 2019, 68, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Donald, E.C.; Porter, I.J. Clubroot in Australia: The history and impact of Plasmodiophora brassicae in Brassica crops and research efforts directed towards its control. Can. J. Plant Pathol. 2014, 36, 66–84. [Google Scholar] [CrossRef]
- EPPO Global Database. Plasmodiophora Brassicae—Distribution. Available online: https://gd.eppo.int/taxon/PLADBR/distribution (accessed on 21 December 2021).
- Dixon, G.R. Plasmodiophora brassicae in its Environment. J. Plant Growth Regul. 2009, 28, 212–228. [Google Scholar] [CrossRef]
- Ernst, T.W.; Kher, S.; Stanton, D.; Rennie, D.C.; Hwang, S.F.; Strelkov, S.E. Plasmodiophora brassicae resting spore dynamics in clubroot resistant canola (Brassica napus) cropping systems. Plant Pathol. 2019, 68, 399–408. [Google Scholar] [CrossRef]
- Wallenhammar, A.-C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Peng, G.; Pageau, D.; Strelkov, S.E.; Gossen, B.D.; Hwang, S.-F.; Lahlali, R. A >2-year crop rotation reduces resting spores of Plasmodiophora brassicae in soil and the impact of clubroot on canola. Eur. J. Agron. 2015, 70, 78–84. [Google Scholar] [CrossRef]
- Yang, X.; Huang, X.; Wu, W.; Xiang, Y.; Du, L.; Zhang, L.; Liu, Y. Effects of different rotation patterns on the occurrence of clubroot disease and diversity of rhizosphere microbes. J. Integr. Agric. 2020, 19, 2265–2273. [Google Scholar] [CrossRef]
- Cook, W.R.I.; Schwartz, E.J., VI. The life-history, cytology and method of infection of Plasmodiophora brassiae Woron., the cause of finger-and-toe disease of cabbages and other crucifers. Philos. Trans. R. Soc. Lond. B 1930, 218, 450–461. [Google Scholar]
- Gossen, B.D.; Deora, A.; Peng, G.; Hwang, S.-F.; McDonald, M.R. Effect of environmental parameters on clubroot development and the risk of pathogen spread. Can. J. Plant Pathol. 2014, 36, 37–48. [Google Scholar] [CrossRef]
- Thuma, B.A.; Rowe, R.C.; Madden, L.V. Relationships of soil temperature and moisture to clubroot (Plasmodiophora brassicae) severity on radish in organic soil. Plant Dis. 1983, 67, 758–762. [Google Scholar] [CrossRef]
- Sharma, K.; Gossen, B.D.; McDonald, M.R. Effect of temperature on primary infection by Plasmodiophora brassicae and initiation of clubroot symptoms. Plant Pathol. 2011, 60, 830–838. [Google Scholar] [CrossRef]
- Gossen, B.D.; Adhikari, K.C.; McDonald, M.R. Effects of temperature on infection and subsequent development of clubroot under controlled conditions. Plant Pathol. 2012, 61, 593–599. [Google Scholar] [CrossRef]
- Gravot, A.; Lemarié, S.; Richard, G.; Lime, T.; Lariagon, C.; Manzanares-Dauleux, M.J. Flooding affects the development of Plasmodiophora brassicae in Arabidopsis roots during the secondary phase of infection. Plant Pathol. 2016, 65, 1153–1160. [Google Scholar] [CrossRef]
- Dobson, R.; Gabrielson, R.L.; Baker, A.S. Soil water matric potential requirements for root hair and cortical infection of Chinese cabbage by Plasmodiophora brassicae. Phytopathology 1982, 72, 1598–1600. [Google Scholar] [CrossRef]
- Murakami, H.; Tsushima, S.; Kuroyanagi, Y.; Shishido, Y. Reduction of resting spore density of Plasmodiophora brassicae and clubroot disease severity by liming. Soil Sci. Plant Nutr. 2002, 48, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Strehlow, B.; de Mol, F.; Struck, C. Standorteigenschaften und Anbaumanagement erklären regionale Unterschiede im Kohlherniebefall in Deutschland: Location and crop management affect clubroot severity in Germany. Gesunde Pflanz. 2014, 66, 157–164. [Google Scholar] [CrossRef]
- Gossen, B.D.; Kasinathan, H.; Deora, A.; Peng, G.; McDonald, M.R. Effect of soil type, organic matter content, bulk density and saturation on clubroot severity and biofungicide efficacy. Plant Pathol. 2016, 65, 1238–1245. [Google Scholar] [CrossRef]
- Niwa, R.; Nomura, Y.; Osaki, M.; Ezawa, T. Suppression of clubroot disease under neutral pH caused by inhibition of spore germination of Plasmodiophora brassicae in the rhizosphere. Plant Pathol. 2008, 57, 445–452. [Google Scholar] [CrossRef]
- Rashid, A.; Ahmed, H.U.; Xiao, Q.; Hwang, S.F.; Strelkov, S.E. Effects of root exudates and pH on Plasmodiophora brassicae resting spore germination and infection of canola (Brassica napus L.) root hairs. Crop Protect. 2013, 48, 16–23. [Google Scholar] [CrossRef]
- Macfarlane, I. A solution-culture technique for obtaining root hair or primary, infection by Plasmodiophora brassicae. J. Gen. Microbiol. 1958, 18, 720–732. [Google Scholar] [CrossRef] [Green Version]
- Myers, D.F.; Campbell, R.N. Lime and the control of clubroot of Crucifers: Effects of pH, calcium, magnesium, and their interaction. Phytopathology 1985, 75, 670–673. [Google Scholar] [CrossRef]
- Donald, E.C.; Porter, I.J. A sand–solution culture technique used to observe the effect of calcium and pH on root hair and cortical stages of infection by Plasmodiophora brassicae. Austral. Plant Pathol. 2004, 33, 585. [Google Scholar] [CrossRef]
- Liao, J.; Luo, L.; Zhang, L.; Wang, L.; Shi, X.; Yang, H.; Tan, S.; Tan, L.; Liu, X.; Wang, D.; et al. Comparison of the effects of three fungicides on clubroot disease of tumorous stem mustard and soil bacterial community. J. Soils Sediments 2022, 22, 256–271. [Google Scholar] [CrossRef]
- Donald, C.; Porter, I. Integrated control of Clubroot. J. Plant Growth Regul. 2009, 28, 289–303. [Google Scholar] [CrossRef]
- Diederichsen, E.; Frauen, M.; Linders, E.G.A.; Hatakeyama, K.; Hirai, M. Status and perspectives of clubroot resistance breeding in crucifer crops. J. Plant Growth Regul. 2009, 28, 265–281. [Google Scholar] [CrossRef]
- Piao, Z.; Ramchiary, N.; Lim, Y.P. Genetics of clubroot resistance in Brassica species. J. Plant Growth Regul. 2009, 28, 252–264. [Google Scholar] [CrossRef]
- Rahman, H.; Peng, G.; Yu, F.; Falk, K.C.; Kulkarni, M.; Selvaraj, G. Genetics and breeding for clubroot resistance in Canadian spring canola (Brassica napus L.). Can. J. Plant Pathol. 2014, 36, 122–134. [Google Scholar] [CrossRef]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y. An update on the arsenal: Mining resistance genes for disease management of Brassica crops in the genomic era. Hortic. Res. 2020, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Hegewald, H.; Wensch-Dorendorf, M.; Sieling, K.; Christen, O. Impacts of break crops and crop rotations on oilseed rape productivity: A review. Eur. J. Agron. 2018, 101, 63–77. [Google Scholar] [CrossRef]
- Zahr, K.; Sarkes, A.; Yang, Y.; Ahmed, H.; Zhou, Q.; Feindel, D.; Harding, M.W.; Feng, J. Plasmodiophora brassicae in its environment: Effects of temperature and light on resting spore survival in soil. Phytopathology 2021, 111, 1743–1750. [Google Scholar] [CrossRef]
- Rennie, D.C.; Holtz, M.D.; Turkington, T.K.; Leboldus, J.M.; Hwang, S.-F.; Howard, R.J.; Strelkov, S.E. Movement of Plasmodiophora brassicae resting spores in windblown dust. Can. J. Plant Pathol. 2015, 37, 188–196. [Google Scholar] [CrossRef]
- Howard, R.J.; Strelkov, S.E.; Harding, M.W. Clubroot of cruciferous crops—new perspectives on an old disease. Can. J. Plant Pathol. 2010, 32, 43–57. [Google Scholar] [CrossRef]
- Hwang, S.-F.; Howard, R.J.; Strelkov, S.E.; Gossen, B.D.; Peng, G. Management of clubroot (Plasmodiophora brassicae) on canola (Brassica napus) in western Canada. Can. J. Plant Pathol. 2014, 36, 49–65. [Google Scholar] [CrossRef]
- Ren, L.; Xu, L.; Liu, F.; Chen, K.; Sun, C.; Li, J.; Fang, X. Host range of Plasmodiophora brassicae on cruciferous crops and weeds in China. Plant Dis. 2016, 100, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani-Noor, N.; Rodemann, B. Reducing the build-up of Plasmodiophora brassicae inoculum by early management of oilseed rape volunteers. Plant Pathol. 2018, 67, 426–432. [Google Scholar] [CrossRef]
- Campbell, R.N.; Greathead, A.S.; Myers, D.F.; de Boer, G.J. Factors related to control of clubroot of Crucifers in the salinas valley of California. Phytopathology 1985, 75, 665–670. [Google Scholar] [CrossRef]
- Tremblay, N.; Bélec, C.; Coulombe, J.; Godin, C. Evaluation of calcium cyanamide and liming for control of clubroot disease in cauliflower. Crop Protect. 2005, 24, 798–803. [Google Scholar] [CrossRef]
- Knox, O.; Oghoro, C.O.; Burnett, F.J.; Fountaine, J.M. Biochar increases soil pH, but is as ineffective as liming at controlling clubroot. J. Plant Pathol. 2015, 97, 149–152. [Google Scholar]
- McGrann, G.R.; Gladders, P.; Smith, J.A.; Burnett, F. Control of clubroot (Plasmodiophora brassicae) in oilseed rape using varietal resistance and soil amendments. Field Crop Res. 2016, 186, 146–156. [Google Scholar] [CrossRef]
- Wakeham, A.; Faggian, R.; Kennedy, R. Assessment of the response of Plasmodiophora brassicae in contaminated horticultural land, using lime-based fertilizer concentrations. Food Energy Secur. 2017, 6, 84. [Google Scholar] [CrossRef]
- Dixon, G.R. Managing clubroot disease (caused by Plasmodiophora brassicae Wor.) by exploiting the interactions between calcium cyanamide fertilizer and soil microorganisms. J. Agric. Sci. 2017, 155, 527–543. [Google Scholar] [CrossRef]
- Webster, M.A.; Dixon, G.R. Calcium, pH and inoculum concentration influencing colonization by Plasmodiophora brassicae. Mycol. Res. 1991, 95, 64–73. [Google Scholar] [CrossRef]
- Neuweiler, R.; Heller, W.E.; Kraus, J. Bekämpfung der Kohlhernie durch gezielte Düngungsmassnahmen. (Preventive effects of different fertilization strategies against the clubroot disease (Plasmodiophora brassicae)). AgrarForschung 2009, 16, 360–365. [Google Scholar]
- Niwa, R.; Kumei, T.; Nomura, Y.; Yoshida, S.; Osaki, M.; Ezawa, T. Increase in soil pH due to Ca-rich organic matter application causes suppression of the clubroot disease of crucifers. Soil Biol. Biochem. 2007, 39, 778–785. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kochi, S.; Kunita, H.; Ito, S.; Kameya-Iwaki, M. Biological mode of action of the fungicide, flusulfamide, against Plasmodiophora brassicae (clubroot). Eur. J. Plant Pathol. 1999, 105, 577–584. [Google Scholar] [CrossRef]
- Diederichsen, E.; Sacristan, M.D. Disease response of resynthesized Brassica napus L. lines carrying different combinations of resistance to Plasmodiophora brassicae Wor. Plant Breeding 1996, 115, 5–10. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, R Package Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 21 December 2021).
- Fox, N.M.; Hwang, S.-F.; Manolii, V.P.; Turnbull, G.; Strelkov, S.E. Evaluation of lime products for clubroot (Plasmodiophora brassicae) management in canola (Brassica napus) cropping systems. Can. J. Plant Pathol. 2021, 44, 21–38. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, Y.; Gan, G.; Li, W.; Wan, W.; Jiang, Y.; Yang, T.; Zhang, Y.; Xu, Y.; Wang, Y.; et al. Exploring rhizo-microbiome transplants as a tool for protective plant-microbiome manipulation. ISME COMMUN. 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Pliego, C.; Ramos, C.; de Vicente, A.; Cazorla, F.M. Screening for candidate bacterial biocontrol agents against soilborne fungal plant pathogens. Plant Soil 2011, 340, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P. Screening for novel biocontrol agents applicable in plant disease management—A review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.-J.; Qiao, J.; Gao, X.; Borriss, R. Novel routes for improving biocontrol activity of Bacillus based bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma-plant-pathogen interactions for better development of biocontrol applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef]
- Zhu, M.; He, Y.; Li, Y.; Ren, T.; Liu, H.; Huang, J.; Jiang, D.; Hsiang, T.; Zheng, L. Two new biocontrol agents against clubroot caused by Plasmodiophora brassicae. Front. Microbiol. 2019, 10, 3099. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Li, X.-Y.; Mao, Z.-C.; Wang, Y.-H.; Wu, Y.-X.; He, Y.-Q.; Long, C.-L. Diversity and active mechanism of fengycin-type cyclopeptides from Bacillus subtilis XF-1 against Plasmodiophora brassicae. J. Microbiol. Biotechnol. 2013, 23, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, J.; Dong, L.; Li, X.; Asif, M.; Guo, Q.; Jiang, W.; Ma, P.; Zhang, L. Fengycin produced by Bacillus subtilis NCD-2 is involved in suppression of clubroot on Chinese cabbage. Biol. Control 2019, 136, 104001. [Google Scholar] [CrossRef]
- He, P.; Cui, W.; Munir, S.; He, P.; Li, X.; Wu, Y.; Yang, X.; Tang, P.; He, Y. Plasmodiophora brassicae root hair interaction and control by Bacillus subtilis XF-1 in Chinese cabbage. Biol. Control 2019, 128, 56–63. [Google Scholar] [CrossRef]
- Liu, C.; Yang, Z.; He, P.; Munir, S.; Wu, Y.; Ho, H.; He, Y. Deciphering the bacterial and fungal communities in clubroot-affected cabbage rhizosphere treated with Bacillus subtilis XF-1. Agr. Ecosyst. Environ. 2018, 256, 12–22. [Google Scholar] [CrossRef]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.-T.; Waidyarathne, P.; Kloepper, J.; de La Fuente, L. Strain-specific quantification of root colonization by plant growth promoting rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in non-sterile soil and field conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; McGregor, L.; Lahlali, R.; Gossen, B.D.; Hwang, S.F.; Adhikari, K.K.; Strelkov, S.E.; McDonald, M.R. Potential biological control of clubroot on canola and crucifer vegetable crops. Plant Pathol. 2011, 60, 566–574. [Google Scholar] [CrossRef]
- Lahlali, R.; Peng, G.; McGregor, L.; Gossen, B.D.; Hwang, S.F.; McDonald, M. Mechanisms of the biofungicide Serenade (Bacillus subtilis QST713) in suppressing clubroot. Biocontrol Sci. Technol. 2011, 21, 1351–1362. [Google Scholar] [CrossRef]
- Lahlali, R.; Peng, G.; Gossen, B.D.; McGregor, L.; Yu, F.Q.; Hynes, R.K. Evidence that the biofungicide Serenade (Bacillus subtilis) suppresses clubroot on canola via antibiosis and induced host resistance. Phytopathology 2013, 103, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Expósito, R.G.; Postma, J.; Raaijmakers, J.M.; de Bruijn, I. Diversity and activity of Lysobacter species from disease suppressive soils. Front. Microbiol. 2015, 6, 1243. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zhang, L.; He, Y.; Liu, F.; Li, M.; Wang, Z.; Ji, G. Isolation and characterization of bacterial isolates for biological control of clubroot on Chinese cabbage. Eur. J. Plant Pathol. 2014, 140, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, Q.; Lyu, A.; Zhang, J.; Wu, M.; Chen, S.; Chen, W.; Li, G.; Yang, L. Optimization of the cultural medium and conditions for production of antifungal substances by Streptomyces platensis 3-10 and evaluation of its efficacy in suppression of clubroot disease (Plasmodiophora brassicae) of oilseed rape. Biol. Control 2016, 101, 59–68. [Google Scholar] [CrossRef]
- Arif, S.; Liaquat, F.; Yang, S.; Shah, I.H.; Zhao, L.; Xiong, X.; Garcia, D.; Zhang, Y. Exogenous inoculation of endophytic bacterium Bacillus cereus suppresses clubroot (Plasmodiophora brassicae) occurrence in pak choi (Brassica campestris sp. chinensis L.). Planta 2021, 253, 25. [Google Scholar] [CrossRef]
- Luo, Y.; Dong, D.; Gou, Z.; Wang, X.; Jiang, H.; Yan, Y.; Wu, C.; Zhou, C. Isolation and characterization of Zhihengliuella aestuarii B18 suppressing clubroot on Brassica juncea var tumida Tsen. Eur. J. Plant Pathol. 2018, 150, 213–222. [Google Scholar] [CrossRef]
- Ahmed, A.; Munir, S.; He, P.; Li, Y.; He, P.; Yixin, W.; He, Y. Biocontrol arsenals of bacterial endophyte: An imminent triumph against clubroot disease. Microbio.l Res. 2020, 241, 126565. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.O.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Park, D.J.; Kim, C.J.; Kim, J.-C. Isolation and characterization of endophytic actinomycetes from Chinese cabbage roots as antagonists to Plasmodiophora brassicae. J. Microbiol. Biotechnol. 2008, 18, 1741–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, Y.; Lin, S.; Liu, F.; Song, Q.; Peng, Y.; Zhao, L. A strain of Streptomyces griseoruber isolated from rhizospheric soil of Chinese cabbage as antagonist to Plasmodiophora brassicae. Ann. Microbiol. 2012, 62, 247–253. [Google Scholar] [CrossRef]
- Wei, L.; Yang, J.; Ahmed, W.; Xiong, X.; Liu, Q.; Huang, Q.; Ji, G. Unraveling the association between metabolic changes in inter-genus and intra-genus bacteria to mitigate clubroot disease of Chinese cabbage. Agronomy 2021, 11, 2424. [Google Scholar] [CrossRef]
- Da Saraiva, A.L.R.F.; Da Bhering, A.S.; Carmo, M.G.F.; Andreote, F.D.; Dias, A.C.; Da Coelho, I.S. Bacterial composition in brassica-cultivated soils with low and high severity of clubroot. J. Phytopathol. 2020, 168, 613–619. [Google Scholar] [CrossRef]
- Thomidis, T.; Michailides, T.J.; Exadaktylou, E. Phoma glomerata (Corda) Wollenw. & Hochapfel a new threat causing cankers on shoots of peach trees in Greece. Eur. J. Plant Pathol. 2011, 131, 171–178. [Google Scholar] [CrossRef]
- Tran, H.S.; You, M.P.; Lanoiselet, V.; Khan, T.N.; Barbetti, M.J. First Report of Phoma glomerata associated with the Ascochyta blight complex on field pea (Pisum sativum) in Australia. Plant Dis. 2014, 98, 427. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef]
- Sullivan, R.F.; White, J.F. Phoma glomerata as a mycoparasite of powdery mildew. Appl. Environ. Microbiol. 2000, 66, 425–427. [Google Scholar] [CrossRef] [Green Version]
- Arie, T.; Kobayashi, Y.; Okada, G.; Kono, Y.; Yamaguchi, I. Control of soilborne clubroot disease of cruciferous plants by epoxydon from Phoma glomerata. Plant Pathol. 1998, 47, 743–748. [Google Scholar] [CrossRef]
- Sun, Z.-B.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, M.-H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Lahlali, R.; Peng, G. Suppression of clubroot by Clonostachys rosea via antibiosis and induced host resistance. Plant Pathol. 2014, 63, 447–455. [Google Scholar] [CrossRef]
- Yu, X.-X.; Zhao, Y.-T.; Cheng, J.; Wang, W. Biocontrol effect of Trichoderma harzianum T4 on brassica clubroot and analysis of rhizosphere microbial communities based on T-RFLP. Biocontrol Sci. Technol. 2015, 25, 1493–1505. [Google Scholar] [CrossRef]
- Li, J.; Philp, J.; Li, J.; Wei, Y.; Li, H.; Yang, K.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D.; et al. Trichoderma harzianum inoculation reduces the incidence of clubroot disease in Chinese cabbage by regulating the rhizosphere microbial community. Microorganisms 2020, 8, 1325. [Google Scholar] [CrossRef] [PubMed]
- Card, S.; Johnson, L.; Teasdale, S.; Caradus, J. Deciphering endophyte behaviour: The link between endophyte biology and efficacious biological control agents. FEMS Microbiol. Ecol. 2016, 92, fiw114. [Google Scholar] [CrossRef] [Green Version]
- Van Wees, S.C.M.; van der Ent, S.; Pieterse, C.M.J. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Narisawa, K.; Tokumasu, S.; Hashiba, T. Suppression of clubroot formation in Chinese cabbage by the root endophytic fungus, Heteroconium chaetospira. Plant Pathol. 1998, 47, 206–210. [Google Scholar] [CrossRef]
- Hashiba, T.; Narisawa, K. The development and endophytic nature of the fungus Heteroconium chaetospira. FEMS Microbiol. Lett. 2005, 252, 191–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narisawa, K.; Shimura, M.; Usuki, F.; Fukuhara, S.; Hashiba, T. Effects of pathogen density, soil moisture, and soil pH on biological control of clubroot in Chinese cabbage by Heteroconium chaetospira. Plant Dis. 2005, 89, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahlali, R.; McGregor, L.; Song, T.; Gossen, B.D.; Narisawa, K.; Peng, G. Heteroconium chaetospira induces resistance to clubroot via upregulation of host genes involved in jasmonic acid, ethylene, and auxin biosynthesis. PLoS ONE 2014, 9, e94144. [Google Scholar] [CrossRef] [Green Version]
- Jäschke, D.; Dugassa-Gobena, D.; Karlovsky, P.; Vidal, S.; Ludwig-Müller, J. Suppression of clubroot (Plasmodiophora brassicae) development in Arabidopsis thaliana by the endophytic fungus Acremonium alternatum. Plant Pathol. 2010, 59, 100–111. [Google Scholar] [CrossRef]
- Baldrian, P. The known and the unknown in soil microbial ecology. FEMS Microbiol. Ecol. 2019, 95, fiz005. [Google Scholar] [CrossRef]
- Jeger, M.; Beresford, R.; Bock, C.; Brown, N.; Fox, A.; Newton, A.; Vicent, A.; Xu, X.; Yuen, J. Global challenges facing plant pathology: Multidisciplinary approaches to meet the food security and environmental challenges in the mid-twenty-first century. CABI Agric. Biosci. 2021, 2, 1–18. [Google Scholar] [CrossRef]
- Sessitsch, A.; Mitter, B. 21st century agriculture: Integration of plant microbiomes for improved crop production and food security. Microb. Biotechnol. 2015, 8, 32–33. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Soil immune responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef]
- Mueller, U.G.; Sachs, J.L. Engineering microbiomes to improve plant and animal health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Rybakova, D.; Mancinelli, R.; Wikström, M.; Birch-Jensen, A.-S.; Postma, J.; Ehlers, R.-U.; Goertz, S.; Berg, G. The structure of the Brassica napus seed microbiome is cultivar-dependent and affects the interactions of symbionts and pathogens. Microbiome 2017, 5, 104. [Google Scholar] [CrossRef] [PubMed]
- Verbon, E.H.; Liberman, L.M. Beneficial microbes affect endogenous mechanisms controlling root development. Trends Plant Sci. 2016, 21, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebreton, L.; Guillerm-Erckelboudt, A.-Y.; Gazengel, K.; Linglin, J.; Ourry, M.; Glory, P.; Sarniguet, A.; Daval, S.; Manzanares-Dauleux, M.J.; Mougel, C. Temporal dynamics of bacterial and fungal communities during the infection of Brassica rapa roots by the protist Plasmodiophora brassicae. PLoS ONE 2019, 14, e0204195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial interactions within multiple-strain biological control agents impact soil-borne plant disease. Front. Microbiol. 2020, 11, 585404. [Google Scholar] [CrossRef]
Figure 1.
Effect of lime fertilizer on the disease severity index (DSI) of Plasmodiophora brassicae on the susceptible oilseed rape cv “Avatar”. Boxes and whiskers indicate interquartile ranges and subsequent 1.5-fold interquartile ranges. Different letters differ significantly at p < 0.05 according to Tukey’s honestly significant difference post hoc test on ranks of data.
Figure 1.
Effect of lime fertilizer on the disease severity index (DSI) of Plasmodiophora brassicae on the susceptible oilseed rape cv “Avatar”. Boxes and whiskers indicate interquartile ranges and subsequent 1.5-fold interquartile ranges. Different letters differ significantly at p < 0.05 according to Tukey’s honestly significant difference post hoc test on ranks of data.


{kind=link}
Table 1.
Characteristics of the lime fertilizers used.
| Lime Fertilizer | Components (%) | CaO Equivalent (%) | pH of Soil (6 dpi) | ||||
|---|---|---|---|---|---|---|---|
| CaCO3 | CaO | MgO | SO3 | SO4 | |||
| Calcium carbonate | 80 | --- | --- | --- | --- | 45 | 6.44 |
| CaCO3-Mg-lime I | 80 | --- | 5 | --- | --- | 48 | 5.42 |
| CaCO3-Mg-lime II | 50 | --- | 35 | --- | --- | 19 | 6.39 |
| CaSO4 | 68 | --- | 1–2 | --- | 4.5 | 38 | 6.58 |
| Splitting lime I 1 | 80 + 75 | --- | --- | +25 | --- | 22.5 + 22.5 | 6.71 |
| Splitting lime II 1 | 80 + 75 | --- | --- | +25 | --- | 45 + 45 | 6.72 |
| Calcium oxide I | --- | 38 | 3 | --- | --- | 38 | 7.00 |
| Calcium oxide II | --- | 90 | 1 | --- | 0.3 | 90 | 7.34 |
| Non-treated control | --- | --- | --- | --- | --- | --- | 4.90 |
1 additional application of SO3-containing lime 6 days after inoculation.
Table 2.
Disease severity index (DSI). Standard deviation (SD), number of plants tested (N), infestation frequency (IF).
Table 2.
Disease severity index (DSI). Standard deviation (SD), number of plants tested (N), infestation frequency (IF).
| Lime Fertilizer | DSI | SD | N | IF (%) |
|---|---|---|---|---|
| Calcium carbonate | 0.63 | 0.28 | 120 | 86.67 |
| CaCO3-Mg-lime 1 | 0.75 | 0.24 | 120 | 95.83 |
| CaCO3-Mg-lime 2 | 0.82 | 0.25 | 160 | 96.67 |
| CaSO4 | 0.57 | 0.28 | 40 | 82.50 |
| Splitting lime 1 1 | 0.33 | 0.27 | 40 | 47.50 |
| Splitting lime 2 1 | 0.28 | 0.08 | 40 | 40.00 |
| Calcium oxide 1 | 0.15 | 0.25 | 120 | 29.60 |
| Calcium oxide 2 | 0.10 | 0.15 | 120 | 23.33 |
| Non-treated control | 0.97 | 0.08 | 120 | 100.00 |
1 Additional application of SO3-containing lime 6 days after inoculation.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Struck, C.; Rüsch, S.; Strehlow, B. Control Strategies of Clubroot Disease Caused by Plasmodiophora brassicae. Microorganisms 2022, 10, 620. https://doi.org/10.3390/microorganisms10030620
AMA Style
Struck C, Rüsch S, Strehlow B. Control Strategies of Clubroot Disease Caused by Plasmodiophora brassicae. Microorganisms. 2022; 10(3):620. https://doi.org/10.3390/microorganisms10030620
Chicago/Turabian StyleStruck, Christine, Stefanie Rüsch, and Becke Strehlow. 2022. "Control Strategies of Clubroot Disease Caused by Plasmodiophora brassicae" Microorganisms 10, no. 3: 620. https://doi.org/10.3390/microorganisms10030620
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.