
Colletotrichum siamense Strain LVY 9 Causing Spot Anthracnose on Winterberry Holly in China

Ministry of Agriculture Key Lab of Molecular Biology of Crop Pathogens and Insects, Institute of Biotechnology, College of Agriculture & Biotechnology, Zhejiang University, Hangzhou 310058, China
*
Author to whom correspondence should be addressed.
Microorganisms 2023, 11(4), 976; https://doi.org/10.3390/microorganisms11040976
Submission received: 6 March 2023
/
Revised: 30 March 2023
/
Accepted: 6 April 2023
/
Published: 9 April 2023
(This article belongs to the Special Issue Restoring the Integrated Behaviour of the Soil-Plant-Microbe System)
Abstract
:Winterberry holly (Ilex verticillata) is an economically valuable landscaping ornamental plant. Serious outbreaks have been reported, in its leaf tips curl upward, irregular black brown spots appear on leaves, and extensive defoliation is commonly observed. The incidence in Hangzhou was estimated at 50% and resulted in large economic losses for growers in 2018. Samples were collected from the main cultivation area in Zhejiang Province. In total, 11 fungal isolates were obtained from diseased leaves through a single-spore purification method, and isolate LVY 9 exhibited strong pathogenicity. Based on morphology and molecular phylogenetic analyses based on multilocus sequence typing of the glyceraldehyde-3-phosphate dehydrogenase (GAPDH), internal transcribed spacer (ITS) regions, actin (ACT), calmodulin (CAL), and chitin synthase (CHS-1) genes, we identified the pathogen as Colletotrichum siamense, causative agent of anthracnose of winterberry holly.
1. Introduction
Winterberry holly (Ilex verticillata) is a deciduous shrub that is especially valued for its masses of glossy, bright red berries in winter. Also known as Black Alder, its branches are often used for decoration during the holiday season [1]. Winterberry holly is widely cultivated in Europe, the United States, and other countries for its unique ornamental qualities and high adaptability. To date, there are more than 30 varieties cultivated and sold in Europe and America, such as “Oosterwijk”, “Winter Red”, “Berry heavy”, and “A. Gray”, among many others [2,3].
In China, in addition to their desirable floral qualities and broad marketability, especially in the Spring Festival [4]. The vivid red berries symbolize good luck and auspicious harvest. The cultivation area of winterberry holly has been rapidly expanding since 2006 in Zhejiang, Shandong, Fujian, Henan, Jilin, and Gansu provinces, as well as in other regions of China [3,5].
Fruit rot of deciduous holly (Ilex spp. L.) was recently reported as an emerging threat in nurseries in the mid-western and eastern USA [6]. Disease symptoms including early defoliation and withered or rotten fruit were observed throughout all of the fruit development and maturation period [7]. Previous work has indicated that multiple fungal pathogens such as Alternaria alternata, Diaporthe ilicicola, as well as other minor pathogens such as A. arborescens, Botryosphaeriaceae, Colletotrichum fioriniae, C.nymphaeae, D.eres, and Epicoccum nigrum can cause primary and/or secondary infections during the growing season through inoculation by wind, rain splash, and pollinators [8,9]. To date, there have been very few reports describing diseases of winterberry holly, so the relevant disease resistance mechanisms remain unclear.
In 2018, growers reported the occurrence of a new anthracnose disease of winterberry holly in Zhejiang province, China, with primary symptoms including black or brown leaves, irregular leaf spots, and sparse white mycelium. The incidence of diseased leaves ranged from 10% to 50%, severely affecting the aesthetic quality. It is of great economic significance determine the pathogenic agent underlying this disease so that a timely and effective management strategy can be deployed. Here, in this work, we isolate the causative agent of this new disease, and identify it as Colletotrichum siamense.
2. Materials and Methods
2.1. Sample Collection and Pathogen Isolation
The symptomatic samples were collected from Zhejiang province in 2018 (Figure 1). Diseased leave tissues were surface-sterilized with 75% ethanol for 30 s, and then 0.1% mercuric chloride solution for 3 min, rinsed five times by sterile distilled water, and air dried; the axenic tissues were subsequently cut into about 25 mm2 pieces and plated on potato dextrose agar (PDA) medium containing 0.2 mg/mL−1 ampicillin sodium. The plates were incubated at 28 °C in the dark for 7–14 days. Pure mycelia were obtained by hyphae tip separation and stored in PDA slants covered by paroline at 4 °C [10,11].
2.2. Pathogenicity Test
Two inoculation methods were used for the pathogenicity assay. The vitro leaf method was described by Lin and Dissanayake [8,9,12]. One-month-old healthy leaves were collected from potted plants (A.Gray), surface-sterilized with 75% ethanol and immersed in sterile water-Tween 80 solutions (0.3% v/v) 3 times before inoculation. About 5 mm-diameter mycelial plugs were picked up from the culture medium and placed on left and right sides of the same leaf, then those leaves were incubated in a 10 cm2 petri dish containing a wet sterile filter paper, inoculated with sterile PDA as controls. There were 3 replicates of each isolate and the experiment was repeated three times. The dishes were placed in a growth chamber under artificial light (12/12 h light/dark) at 28 °C for 7 days. The second inoculation technique was described by Weir [13]: 1 × 106 conidia/mL of the spore suspensions were prepared in sterile distilled water. Leaves were wounded evenly by the sterilized needle, and then the front and back of the leaves of the biennial plants were sprayed with the suspensions. Sterile distilled water was used as control. Then, all plants were maintained in a greenhouse under 100% relative humidity at temperatures between 30 °C (day) and 28 °C (night) with natural photoperiods. The pathogen was re-isolated from leaf spots, and its identity confirmed by morphological characteristics, which is consistent with the Koch’s postulates.
2.3. Morphological Characteristics
Mycelia discs (7 mm in diameter) from 7-day-old PDA cultures were sub-cultured on PDA medium in incubator, at 28 °C. The mycelium growth was measured every day for 7 consecutive days, and the colony features (size and color) were recorded after 7 days [14]. The mycelia were removed from the surface of the plate to enhance the sporulation under a mixture of fluorescent white and UV light with 12 h dark at 20 °C for 15–20 days [13,15,16]. The conidia were examined visually and microscopically for morphological features, the mycelia (7 mm diameter) with conidia were observed in SEM (Hitachi Model SU-8010) [17,18,19].
Appressorium was produced using a slide culture technique [13]. A mycelium colony on PDA with 1 cm2 was placed in an empty petri dish (9 cm) and immediately covered with a sterile cover slip, then transferred into an empty square petri dish (10 cm) containing sterile water. After 14 days, the cover slip was removed, and a drop of lactic acid was added on a glass slide [13,14].
2.4. Phylogenetic Analysis
2.4.1. Genomic DNA Extraction and PCR Amplification
Genomic DNA was extracted using Ezup Column Fungi Genomic DNA Purification Kit (Sangon Biotech (Shanghai, China) Co., Ltd.) following the manufacturer’s instructions. The PCR reaction mixture contained 9 μL of sterile water, 13 μL of 2 × PCR Master Mix (TSINGKE, Beijing, China), 1 μL of each primer (10 μM), 1μL of genomic DNA. PCR was conducted in Applied Biosystems LongGene Thermal Cycler (LongGene Scientific Instruments (Hangzhou, China) Co., Ltd.) under the following conditions for the ITS: 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 30 s, annealing at 52 °C for 30 s, and extension at 72 °C for 1 min; and then followed by a final extension step at 72 °C for 10 min. Amplification programs for the other genes were the same except the annealing temperatures: ACT (58 °C ), CAL (59 °C ), CHS-1 (58 °C ), GAPDH (60 °C). The primers and references are summarized in Table 1.
2.4.2. Fungal Isolates Phylogenetic Analysis
The obtained sequences were edited by BioEdit 7.1.3.0 [24] and multiple alignments were generated with MAFFT 7.273 [25]. To construct the phylogenetic tree, available data for species of Colletotrichum were downloaded from the GenBank database (Table A1). Maximum likelihood (ML) analyses were performed using IQ-TREE [26,27] with the concatenation of the ACT, CAL, CHS-1, GAPDH, and ITS regions. The best evolutionary model was selected through Model Finder [28], as follows: TIM2 + F + R4 model for ACT, TIM + F + R3 for CAL, TIM + F + R3 for CHS-1, HKY + F + R2 for GAPDH and TIM2 + F + R3 for ITS. Confidence of the branch points was evaluated using 5000 bootstrap replicates. The ML values equal to or above 75% values were shown on a tree for significantly supported nodes. Bayesian Inference (BI) was applied to generate a phylogeny tree by MrBayes ver.3.2.6 [29]. GTR + F + I + G4 model was used to the analyses of the gene of ACT, CAL, CHS1, ITS and the HKY + F + G4 model for GAPDH following Ronquist and Huelsenbeck [29] for BI analyses with 200,000 replicates and the phylogenetic tree was sampled every 100 generations. The first 500 trees were removed for the burn-in phase. The consensus of the remaining trees was constructed with resulting node frequencies that were treated as Bayesian posterior probabilities (PP). BI posterior probability (BI-PP) values equal to or above 0.95 were determined to be significant. The tree was rooted with the outgroup, Monilochaetes infuscans.
3. Results
3.1. The Strain LVY 9 Was Pathogen of Anthracnose on Winterberry Holly through Koch’s Postulates
We identified and isolated eleven isolates (LVY 1–11) from symptomatic leaves of Ilexverticillata (Hangzhou city, Zhejiang province, China) based on morphological features and confirmation by Sanger sequencing. These isolates belonged to genera Colletotrichum, Alternaria, Botryosphaeria, Leptosphaeria, Cercospora, and Emericella, and subsequent pathogenicity tests indicated that only inoculation with Colletotrichum (isolate LVY 9) resulted in symptom formation on leaves in vitro. These symptoms included brown necrotic lesions with dense whitish-grey aerial mycelia, and a few bright orange conidial masses near the point of inoculation at 7 days post infection (dpi) (Figure 2a,b). The mycelia are grayish brown with white edge and reserve is turquoise after strain LVY 9 on PDA for 7 days. Colonies were round, swell, neat edge, with cottony radial growth of white. In particular, large clusters of bright orange conidia were produced on PDA for 30 d. These characteristics are very similar to those of Colletotrichum gloeosporioides complex. Following spray inoculations, similar symptoms appeared at 30 dpi in winterberry nursery fields (Figure 2c). In diseased plants, leaf tips of inoculated plants exhibited an obvious curly phenotype (Figure 2d). In addition, we observed irregular gray black spots on the leaves. We were able to successfully re-isolate the Colletotrichum isolates from these diseased plants, thereby fulfilling Koch’s postulates.
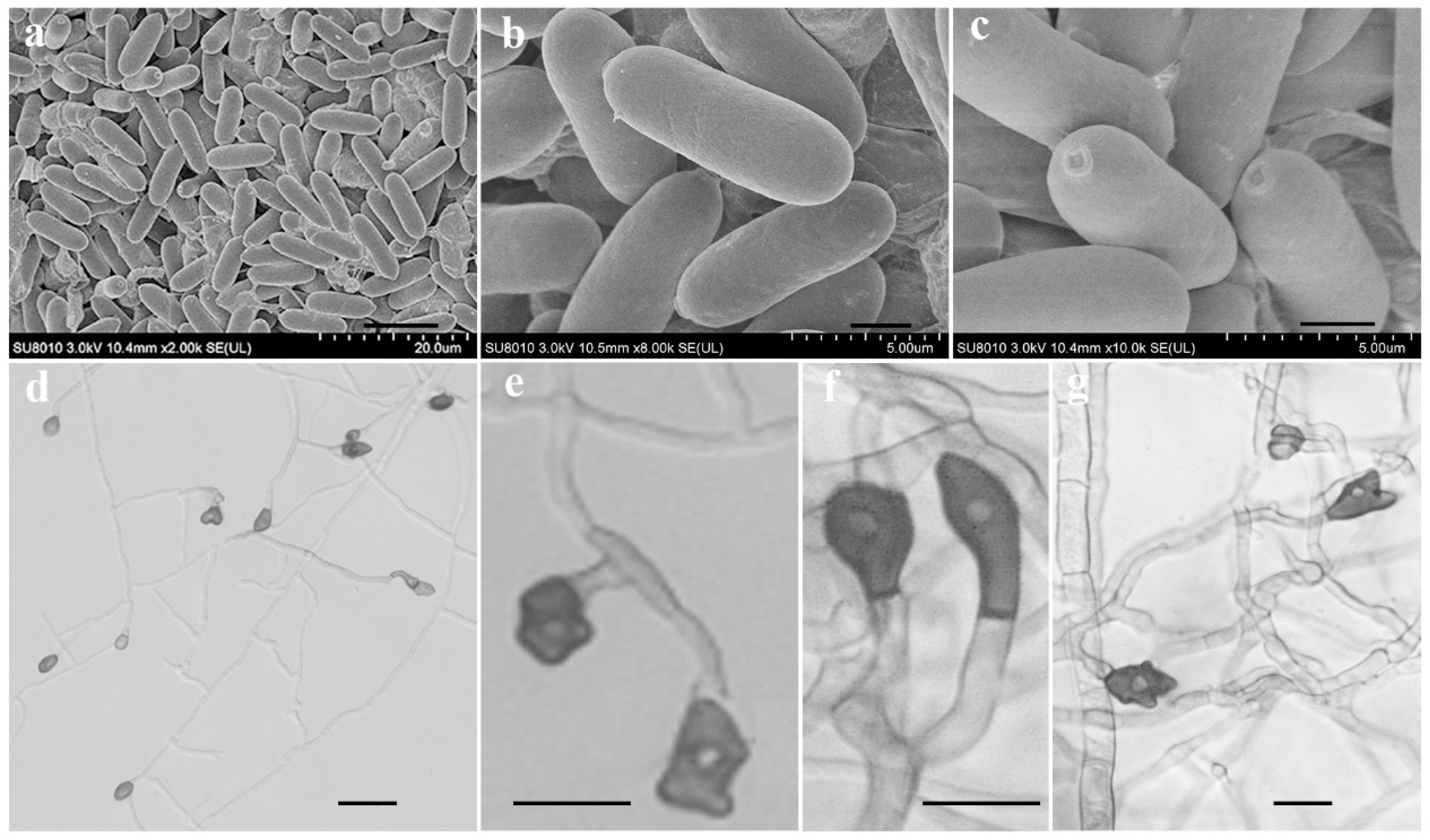
To characterize the microscopic features of the pathogen, in addition to its symptoms and colony morphology, we selected a representative isolate for morphological characterization by SEM and light microscopy. On PDA medium, isolate LVY 9 colonies first appeared white, then became gray to dark grey, with an average growth rate of approximately 10.72 mm/day. Bright orange conidia and conidiophores were produced under continuous UV + fluorescent white lights with a 12:12 h light: dark cycle (Figure 2g). Scanning electron microscopy revealed that conidial dimensions were 7.8–12.7 × 2. 6–3.6 µm, and conidia were smooth-walled, hyaline, and cylindrical, with obtuse to slightly rounded ends (Figure 3a–c). Under light microscopy, appressoria (5.8–10.7 × 3.9–7.7 µm) were observed to form mostly from mycelium on slide cultures, and appeared brown to dark brown, ovoid, clavate, and slightly irregular in shape (Figure 3d–h). Morphological features of the isolate LVY 9 were highly similar to those of species belonging to the Colletotrichum gloeosporioides complex [13,30,31,32].
3.2. Strain LVY 9 Was Identified as Colletotrichum siamense by Phylogenetic Analyses
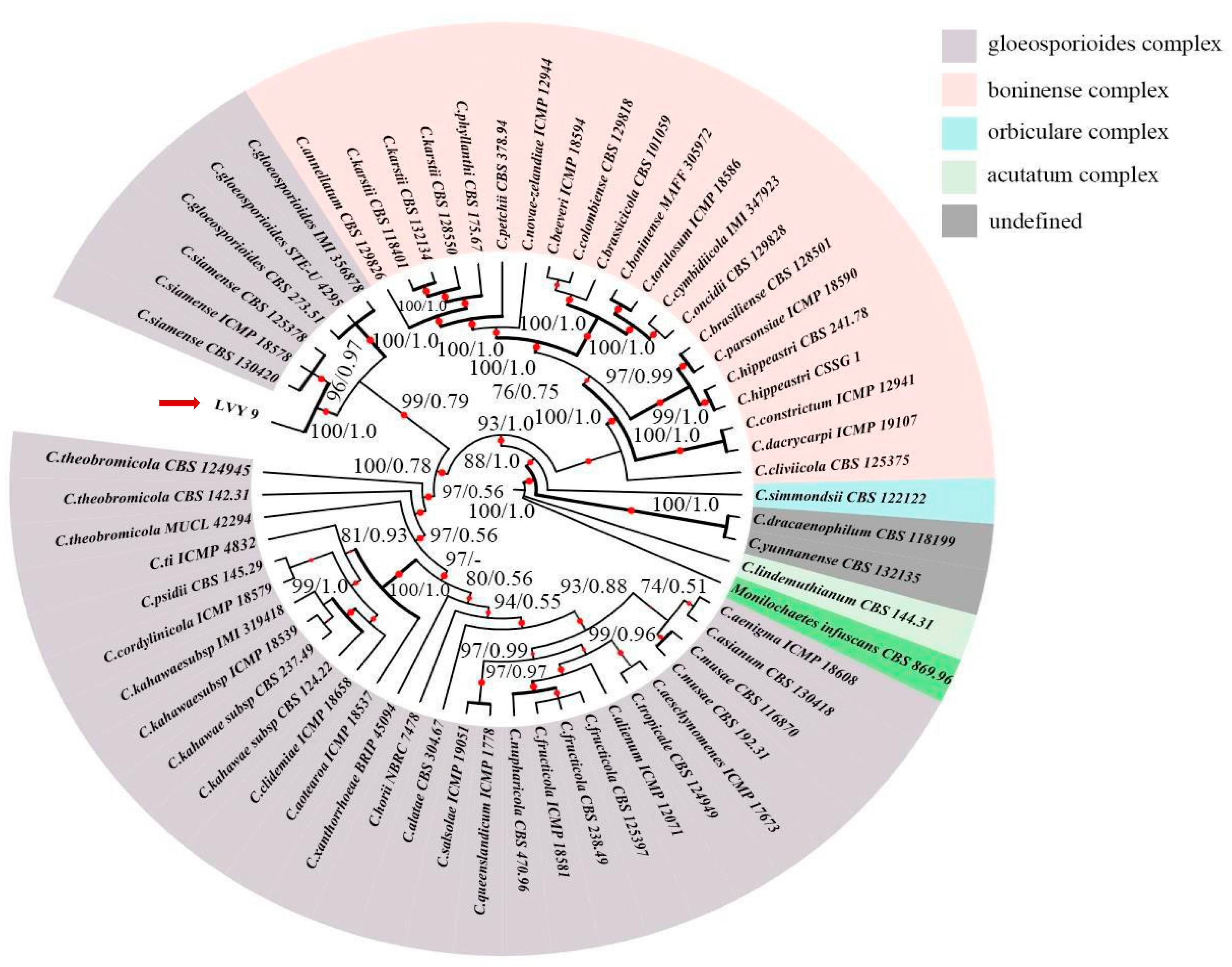
For rigorous molecular identification, we used partial sequences from ACT (GenBank number: OQ652092), CAL (GenBank number: OQ652091), GAPDH (GenBank number: OQ652089), CHS-1 (GenBank number: OQ652090), and ITS (GenBank number: OQ651128) from isolate LVY9 for species level identification of the pathogen (see Table 1 for primers and source studies). Maximum likelihood-based phylogenetic reconstruction of these concatenated sequences (2370 nucleotides) including ACT (1–311 bp), CAL (312–1119 bp), CHS-1 (1120–1420 bp), GADPH (1421–1736 bp), and ITS (1737–2370 bp) was congruent with that of a Bayesian inference (BI) tree for the five concatenated loci. Relationships among almost all of the reference isolates could be clearly distinguished at the species level. Furthermore, phylogenetic analysis indicated that isolate LVY 9 clustered with C. siamense CBS 130,420, C. siamense ICMP 18,587, and C. siamense CBS 125,378, thus forming a distinct clade, which was highly supported in both ML-BS (100%) and BI-PP (1.0) models (Figure 4). Based on these findings, we concluded that the LVY9 isolate was a strain of C. siamense, the previously reported causative agent of anthracnose in winterberry holly.
4. Discussion
The asexual genus Colletotrichum is largely comprised of economically and agriculturally destructive plant pathogens [33], causing major losses in yield and productivity to a wide range of fruit, vegetable and ornamental crops such as strawberry, mango, chilli, pear, eggplant, cowpea, mandevilla and rhododendron [18,34,35,36,37,38]. Based on morphological studies and phylogenetic analyses using actin (ACT), calmodulin (CAL), chitin synthase 1 (CHS-1), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), internal transcribed spacers (ITS), and β-Tubulin 2 (TUB2), Damm and colleagues [15] effectively separated the C. acutatum species complex into 30 species, while Weir and coworkers [13] distinguished 22 species and one subspecies within the C. gloeosporioides species complex. In particular, C. siamense was reported to cause anthracnose on persea americana, pistacia vera, coffea arabica, vitis vinifera, malus domestica, and hymenocallis Americana [13]. Chunhua indicated that the 13 isolates of C. gloeosporioides species complex from the rubber tree in Hainan Province were identified as C.siamense and C. fructicola [39]. Simlarly, C. siamense was involved in walnut, pyrus spp., litchi pepper, amorphophallus konjac, photinia × fraseri in China [40,41,42,43,44]. In this work, we first report the presence of C. siamense, the causative pathogen for anthracnose of the ornamental plant winterberry holly, in China.
In the Colletotrichum gloeosporioides complex, C. gloeosporioides, C. siamense, and C. fructicola exhibit similar morphological characteristics including conidial size, shape, and appressoria formation. C. siamense is genetically close to C. fructicola, but C. fructicola has slightly longer and narrower cylindrical or subcylindrical conidia that have irregularly shaped, crenate, brown to dark brown appressoria, and branched hyphae [36,45]. However, the conidia of C. siamense are fusiform with obtuse to slightly rounded ends, occasionally oblong and ovoid. Previous reports also confirm that this species exhibits regular- to slightly irregular-shaped appressoria [14]. In contrast, C. gloeosporioides conidia appear cylindrical, although slightly tapered with obtuse or slightly rounded to oblong ends. The C. gloeosporioides appressoria are circular to slightly irregular, thus differing from those of C. siamense [14,46]. In this study, the shapes of the conidia and appressoria of isolates matched those of C. siamense, described by Prihastuti [14]. Moreover, the size of appressoria was similar to that of C. siamense, although the conidia described by Sharma [36,47] were smaller. Mycelial growth rates of isolates obtained in this study were also slower than those of the strain of C. siamense described by Yaowen [48]. We speculated that these morphological differences may be due (at least in part) to responses to the environment, growth media, and host.
Since morphological identification is important but not definitive for Colletotrichum, sequence analysis and infection behavior are also used to discriminate down to species level. As mentioned in previous studies, species in the C. gleosporioide species complex are genetically distinct from those in the C. boninense complex, but have highly similar micro-morphologiesm [13,15,16,49]. Initial sequence analyses relying on ITS were unable to satisfactorily distinguish among Colletotrichum species due to their high level of evolutionary conservation. In this work, we therefore used several genes to identify the LVY9 strain that were previously reported to successfully resolve Colletotrichum species [18,50]. Notably, ITS can separate C. gloeosporioides from all other Colletotrichum species, but cannot reliably separate C. siamense from C. alienum, C. fructicola, or C. tropicale. These species are best distinguished using CAL or TUB2 [13]. The causative agent of leaf spots on Sterculia nobilis in China was identified as C. siamense using a combination of ITS, ACT, GAPDH, CAL, CHS-1, and TUB2 genes [48]. In this study, we used a combinations of five genes (ACT, CAL, CHS-1, GAPDH and ITS) to identify the isolates obtained from winterberry holly, thus providing strong molecular evidence for the identification of the isolates as C. siamense.
C. siamense was originally described as a pathogen of coffee berries in Thailand [14]. This species has since been confirmed to infect more than 60 plant species worldwide [13,15,16]. This paper presents the first report describing C. siamense as the causal agent of anthracnose of winterberry holly in Zhejiang, China. As winterberry holly is commonly used as a bonsai or cut flower, the disease directly reduces its ornamental value. In addition, the occurrence of disease may harm the introduction and cultivation of winterberry holly. This identification of the disease-causing species facilitates the establishment of control measures, not only for winterberry holly, but for all of the known hosts of this pathogen. This study also provides a basis for future studies of the molecular mechanisms of this pathogenic interaction, identification of disease-resistant varieties, and creation of stable resistant materials.
5. Conclusions
According to Koch’s rule, The strain LVY9 was elected as the pathogen causing anthracnose of Winterberry Holly in Zhejiang Province, China. The strain LVY9 was identified as the C. gloeosporioides complex by observing the morphology features of mycelia, conidia and appressorium. The strain LVY9 was identified as C. siamense by further analysis of the phylogenetic tree that combined Actin, Calmodulin, Glyceraldehyde-3- Phosphate dehydrogenase, Chitin synthase and Internal transcribed spacer genes. So, we concluded that C. siamense was causative agent of anthracnose in winterberry holly.
Author Contributions
L.F. and Y.Z. conceived the idea and drafted the manuscript. L.F., Y.Z., B.M. and W.C. contributed in writing up and revision. L.F. and B.M. performed formatting. All the authors read the final version and approved its submission. All authors have read and agreed to the published version of the manuscript.
Funding
The funding support from the Foundation of Study on the main diseases and control techniques for winterberry holly in China (2019-KYY-516108-0007).
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful for technical support and practical experimental shared by Youxiang Yu which is a staff of HangZhou Runtu Horticulture Technology Co., Ltd. In addition, we thank Seyed Ali Mirghasempour khoshkroudi and Yunfang He for proofreading the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
GenBank accession numbers of the accepted Colletotrichum species.
| Colletotrichum Species | Culture Collection | GenBank Accession Number | References | ||||
|---|---|---|---|---|---|---|---|
| ITS | GAPDH | CAL | ACT | CHS-1 | |||
| C.aenigma | ICMP 18,608 * | JX010244 | JX010044 | JX009683 | JX009443 | JX009774 | [13] |
| C.aeschynomenes | ICMP 17,673 * ATCC 201,874 | JX010176 | JX009930 | JX009721 | JX009483 | JX009799 | [13] |
| C.alatae | CBS 304.67 * ICMP 17,919 | JX010190 | JX009990 | JX009738 | JX009483 | JX009799 | [13] |
| C.alienum | ICMP 12,071 * | JX010251 | JX010028 | JX009654 | JX009572 | JX009882 | [13] |
| C.annellatum | CBS 129,826 CH1 * | JQ005222 | JQ005309 | JQ005743 | JQ005570 | JQ005396 | [16] |
| C.aotearoa | ICMP 18,537 * | JX010205 | JX010005 | JX009611 | JX009564 | JX009853 | [13] |
| C.asianum | ICMP 18,580 * CBS 130,418 | FJ972612 | JX010053 | FJ917506 | JX009584 | JX009867 | [13] |
| C.beeveri | ICMP 18,594 * CBS 125,827 | JQ005171 | JQ005258 | JQ005692 | JQ005519 | JQ005345 | [16] |
| C.boninense | MAFF 305,972 * CBS 123,755 | JQ005153 | JQ005240 | JQ005674 | JQ005501 | JQ005327 | [13] |
| C.brasiliense | CBS 128,501 PAS12 * | JQ005235 | JQ005322 | JQ005756 | JQ005583 | JQ005409 | [16] |
| C.brassicicola | CBS 101,059 LYN 16,331 * | JQ005172 | JQ005259 | JQ005693 | JQ005520 | JQ005346 | [16] |
| C.clidemiae | ICMP 18,658 * | JX010265 | JX009989 | JX009645 | JX009537 | JX009877 | [13] |
| C.cliviae | CBS 125,375 * | GQ485607 | GQ856756 | GQ849464 | GQ856777 | GQ856722 | [17] |
| C.colombiense | CBS 129,818 G2 * | JQ005174 | JQ005261 | JQ005695 | JQ005522 | JQ005348 | [16] |
| C.constrictum | ICMP 12,941 * CBS 128,504 | JQ005238 | JQ005325 | JQ005759 | JQ005586 | JQ005412 | [16] |
| C.cordylinicola | ICMP 18,579 MFLUCC 090,551 * | JX010226 | JX009975 | HM470237 | HM470234 | JX009864 | [13] |
| C.cymbidiicola | IMI 347,923 * | JQ005166 | JQ005253 | JQ005687 | JQ005514 | JQ005340 | [16] |
| C.dacrycarpi | ICMP 19,107 * CBS 130,241 | JQ005236 | JQ005323 | JQ005757 | JQ005584 | JQ005410 | [16] |
| C.dracaenophilum | CBS 118,199 * | JX519222 | JX546707 | - | JX519238 | JX519230 | [17] |
| C.fructicola | CBS 125,397 * ICMP 18,646 | JX010173 | JX010032 | JX009674 | JX009581 | JX009874 | [13] |
| C.fructicola | CBS 238.49 * ICMP 17,921 | JX010181 | JX009923 | JX009671 | JX009495 | JX009839 | [13] |
| C.fructicola | ICMP 18,581 * CBS 130,416 | JX010165 | JX010033 | FJ917508 | FJ907426 | JX009866 | [13] |
| C.gloeosporioides | CBS 273.51 * ICMP 19,121 | JX010148 | JX010054 | JX009745 | JX009558 | JX009903 | [13] |
| C.gloeosporioides | STE-U4295 * CBS 112,999 | JQ005152 | JQ005239 | JQ005673 | JQ005500 | JQ005326 | [16] |
| C.gloeosporioides | IMI 356,878 * ICMP 17,821 CBS 112,999 | JX010152 | JX010056 | JX009731 | JX009531 | JX009818 | [13] |
| C.hippeastri | CSSG 1 * CBS 125,376 | JQ005231 | JQ005318 | JQ005752 | JQ005579 | JQ005405 | [16] |
| C.hippeastri | CBS 241.78 | JX010293 | JX009932 | JX009740 | JX009485 | JX009838 | [13] |
| C.horii | NBRC 7478 * ICMP 10,492 | GQ329690 | GQ32961 | JX009604 | JX009438 | JX009752 | [13] |
| C.kahawae subsp | CBS 124.22* ICMP 19,122 | JX010228 | JX009950 | JX009744 | JX009536 | JX009902 | [13] |
| C.kahawae subsp | IMI 319,418 * ICMP 17,816 | JX010231 | JX010012 | JX009642 | JX009452 | JX009813 | [13] |
| C.kahawae subsp | ICMP 18,539 * | JX010230 | JX009966 | JX009635 | JX009523 | JX009800 | [13] |
| C.kahawae subsp | CBS 237.49 * ICMP 17,922 | JX010238 | JX010042 | JX009636 | JX009450 | JX009840 | [13] |
| C.karstii | CBS 132,134 * | HM585409 | HM858391 | HM582013 | HM581995 | HM582023 | [16] |
| C.karstii | CBS 128,550 ICMP 17,896 | JQ005219 | JQ005306 | JQ005740 | JQ005567 | JQ005393 | [16] |
| C.karstii | CBS 118,401 | JQ005192 | JQ005279 | JQ005713 | JQ005540 | JQ05366 | [16] |
| C.lindemuthianum | CBS 144.31 | JQ005779 | JX546712 | - | JQ005842 | JQ005800 | [17] |
| C.musae | CBS 192.31 ICMP 17,923 | JX010143 | JX009929 | JX009690 | JX009587 | JX009841 | [13] |
| C.musae | CBS 116,870 * ICMP 19,119 | JX010145 | JX010047 | JX009687 | JX009551 | JX009849 | [13] |
| C.novae-zelandiae | ICMP 12,944 * CBS 128,505 | JQ005228 | JQ005315 | JQ005749 | JQ005576 | JQ005402 | [16] |
| C.nupharicola | CBS 470.96 * ICMP 18,187 | JX010187 | JX009972 | JX009663 | JX009437 | JX009835 | [13] |
| C.oncidii | CBS 129,828 * | JQ005169 | JQ005256 | JQ005690 | JQ005517 | JQ005343 | [16] |
| C.parsonsiae | ICMP 18,590 * CBS 128,525 | JQ005223 | JQ005320 | JQ005754 | JQ005581 | JQ005407 | [16] |
| C.petchii | CBS 378.94 * | JQ005223 | JQ005310 | JQ005744 | JQ005571 | JQ005397 | [16] |
| C.phyllanthi | CBS 175.67 MACS 271 * | JQ005221 | JQ005308 | JQ005742 | JQ005569 | JQ005395 | [16] |
| C.psidii | CBS 145.29 * ICMP 19,120 | JX010219 | JX009967 | JX009743 | JX009515 | JX009901 | [13] |
| C.queenslandicum | ICMP 1778 * | JX010276 | JX009934 | JX009691 | JX009447 | JX009899 | [13] |
| C.salsolae | ICMP 19,051 * | JX010242 | JX009916 | JX009696 | JX009562 | JX009863 | [13] |
| C.siamense | CBS 125,378 * ICMP 18,642 | JX010278 | JX010019 | JX009709 | GQ85675 | GQ856730 | [13] |
| C.siamense | CBS 130,420 * ICMP 19,118 | HM131511 | HM13147 | JX009713 | HM13157 | JX009895 | [13] |
| C.siamense | ICMP 18,578 * CBS 130,417 | JX010171 | JX009924 | FJ917505 | FJ907423 | JX009865 | [13] |
| C.simmondsii | CBS 122,122, BRIP 28,519 * | JQ948276 | JQ948606 | FJ917510 | JQ949588 | JQ948937 | [16] |
| C.theobromicola | CBS 124,945 * ICMP 18,649 | JX010294 | JX010006 | JX009591 | JX009444 | JX009869 | [13] |
| C.theobromicola | CBS 142.31 * ICMP 17,927 | JX010286 | JX010024 | JX009582 | JX009516 | JX009830 | [13] |
| C.theobromicola | MUCL 42,294 * ICMP 17,957 | JX010289 | JX009962 | JX009597 | JX009575 | JX009821 | [13] |
| C.ti | ICMP 4832 * | JX010269 | JX009952 | JX009649 | JX009520 | JX009898 | [17] |
| C.torulosum | ICMP 18,586 * CBS 128,544 | JQ005164 | JQ005251 | JQ005685 | GU27899 | GU228291 | [17] |
| C.tropicale | CBS 124,949 * ICMP 18,653 | JX010264 | JX010007 | JX009719 | JX009489 | JX009870 | [13] |
| C.xanthorrhoeae | BRIP 45,094 * ICMP 17,903 | JX010261 | JX009927 | JX009653 | JX009478 | JX009823 | [13] |
| C.yunnanense | CBS 132,135 * | JX546804 | JX546706 | - | JX519239 | JX519231 | [17] |
Note: * refers to ex-type strains of Colletotrichum species
References
- Xiang, W.; Xu, Z.; Lin, L.; Yongjun, Z.; Weisong, M. Monitoring system based on WSN for fresh cut branches of Ilex verticillata in the whole cold chain. Trans. Chin. Soc. Agric. Mach. 2017, 48, 394–400. [Google Scholar] [CrossRef]
- Juanjuan, M.; Bin, Z.; Yin, C.; Xichen, L.; Jie, Y.; Qian, C. Physiological responses of seedlings of four Ilex verticillata varieties to low temperature stress and a comparison of their cold resistance. J. Nanjing For. Univ. Nat. Sci. Ed. 2020, 44, 34–40. [Google Scholar] [CrossRef]
- Weili, W.; Liping, H.; Minfen, Y.; Huwei, Y.; Daoliang, Y.; Liang, Z.; Youxiang, Y.; Jun, L.; Peng, Z.; Bingsong, Z. Phylogenetic relationships among 12 cultivars of Ilex verticillata based on ISSR molecular markers. J. Zhejiang Agric. For. Univ. 2018, 35, 612–617. [Google Scholar] [CrossRef]
- Yuzhen, Y.; Yunxia, Z.; Zhihao, Z.; Bingqing, A. Diurnal changes of photosynthesis and relationship with the eco-physiological factors of IIex verticillata. J. Northwest Norm. Univ. Nat. Sci. 2016, 52, 88–92. [Google Scholar] [CrossRef]
- Dengfeng, S.; Bin, W.; Chuntao, H.; Minghua, L.; Tonghui, Y.; Jianhong, Z. Transcriptome analysis of Ilex verticillata leaf treated by different concentrations acetic acid solution. Mol. Plant Breed. 2020, 18, 401–408. [Google Scholar] [CrossRef]
- Lin, Z.; Ziqian, Y.; Jianhua, D.; Youxiang, Y.; Yichen, L. Regional introduction experiment of Ilex verticillate ‘Oosterwijk’. J. For. Eng. 2015, 29, 80–82. [Google Scholar] [CrossRef]
- Lin, S.; Taylor, N.J.; Peduto Hand, F. Identification and characterization of fungal pathogens causing fruit rot of deciduous holly. Plant Dis. 2018, 102, 2430–2445. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Peduto Hand, F. Investigations on the timing of fruit infection by fungal pathogens causing fruit rot of deciduous holly. Plant Dis. 2019, 103, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Peduto Hand, F. Determining the sources of primary and secondary inoculum and seasonal inoculum dynamics of fungal pathogens causing fruit rot of deciduous holly. Plant Dis. 2019, 103, 951–958. [Google Scholar] [CrossRef]
- Qing, B.; Lifeng, Z.; Xiaoren, C.; Ni, H.; Wenxing, X.; Guoping, W. Biological and molecular characterization of five Phomopsis species associated with pear shoot canker in China. Plant Dis. 2015, 99, 1704–1712. [Google Scholar] [CrossRef] [Green Version]
- Noireung, P.; Phoulivong, S.; Fang, L.; Cai, L.; Eric, H.C.M.; Ekachai, C.; Jones, E.B.G.; Ali, H.B.; Hyde, D.K. Novel species of Colletotrichum revealed by morphology and molecular analysis. Cryptogam. Mycol. 2012, 33, 347–362. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Liu, M.; Zhang, W.; Chen, Z.; Udayanga, D.; Chukeatirote, E.; Li, X.H.; Yan, J.Y.; Hyde, K.D. Morphological and molecular characterisation of Diaporthe species associated with grapevine trunk disease in China. Fungal Biol. 2015, 119, 283–294. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S. The Colletotrichum boninense species complex. Stud. Mycology. 2012, 73, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhang, W.; Zhao, W.S.; Yan, J.Y. Notes on currently accepted species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum-current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- Xinlei, F.; Yingmei, L.; Rong, M.; Chenming, T. Morphological and phylogenetic studies of Cytospora (Valsaceae, Diaporthales) isolates from Chinese scholar tree, with description of a new species. Mycoscience 2014, 55, 252–259. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulate. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Whitem, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–332. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinfotmatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Schena, L.; Mosca, S.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Agosteo, G.; Sergeeva, V.; Lio, G.M. Species of the Colletotrichum gloeosporioides and C. boninense complexes associated with olive anthracnose. Plant Pathol. 2014, 63, 437–446, Corpus ID: 59138976. [Google Scholar] [CrossRef]
- Huang, F.; Chen, G.; Hou, X.; Fu, Y.; Cai, L.; Hyde, K.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74, Corpus ID: 15163531. [Google Scholar] [CrossRef]
- Rojas, E.; Rehner, S.; Samuels, G.; Bael, S.V.; Herre, E.; Cannon, P.; Chen, R.; Pang, J.; Wang, R.; Zhang, Y.; et al. Colletotrichum gloeosporioides associated with Theobroma cacao and other plants in Panamá: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338, Corpus ID: 44822023. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-kosack, K.E.; Dipietro, A.; Spanu, P.D.; Foster, G.D. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Crouch, J.A.; Beirn, L.A.; Cortese, L.M.; Bonos, S.A.; Clarke, B.B. Anthracnose disease of switchgrass caused by the novel fungal species Colletotrichum navitas. Mycol. Res. 2009, 113, 1411–1421. [Google Scholar] [CrossRef]
- Nam, M.; Park, M.; Lee, H.D.; Yu, S. Taxonomic Re-evaluation of Colletotrichum gloeosporioides Isolated from Strawberry in Korea. Plant Pathol. J. 2013, 29, 317–322, Corpus ID: 2295330. [Google Scholar] [CrossRef]
- Sharma, G.; Shenoy, B.D. Colletotrichum fructicola and C. siamense are involved in chilli anthracnose in India. Arch. Phytopathol. Plant Prot. 2014, 47, 1179–1194. [Google Scholar] [CrossRef]
- Li, H.N.; Jiang, J.J.; Hong, N.; Wang, G.P.; Xu, W.X. First Report of Colletotrichum fructicola causing bitter rot of Pear (Pyrus bretschneideri) in China. Plant Dis. 2013, 97, 1000. [Google Scholar] [CrossRef]
- Kyoko, W.; Hiromi, I.; Takashi, S. Anthracnose of genus Mandevilla caused by Colletotrichum truncatum and C. siamense in Japan. Fungal Dis. 2016, 82, 33–37. [Google Scholar] [CrossRef]
- Chunhua, L.; Huan, Y.; Xiaoyu, Z.; Xiaohan, P.; Wenbo, L.; Weiguo, L.; Fucong, Z. Identification and phylogenetic analysis of anthracnose pathogen Colletotrichum siamense and C. fructicola isolated from Rubber Tree in Hainan. Chin. J. Trop. Crops 2018, 39, 129–136. [Google Scholar] [CrossRef]
- Qinghai, W.; Kun, F.; Dewei, L.; Shanguang, N.; Xiaoqin, W. Walnut anthracnose caused by Colletotrichum siamense in China. Australas. Plant Pathol. 2017, 46, 585–595. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Wang, G.P. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Pers. Mol. Phylogeny Evol. Fugal 2019, 42, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Ling, J.F.; Song, X.B.; Xi, P.G.; Cheng, B.P.; Cui, Y.P.; Chen, X.; Peng, A.T.; Zhang, L.H. Identification of Colletotrichum siamense causing litchi pepper spot disease in mainland China. Plant Pathol. 2019, 68, 1533–1542. [Google Scholar] [CrossRef]
- Wu, J.P.; Zhou, J.; Jiao, Z.B. Amorphophallus konjac anthracnose caused by Colletotrichum siamense in China. J. Appl. Microbiol. 2020, 128, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shengfeng, M.; Haodong, C.; Yanjun, L. First report of the Colletotrichum siamense species complex causing anthracnose on Photinia × fraseri Dress in China. Plant Dis. 2020, 105, 510. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, Y.; Li, N.; Ni, D.; Yang, Y.; Wang, X. Differences in the characteristics and pathogenicity of Colletotrichum camelliae and C. fructicola isolated from the tea plant [Camellia sinensis (L.) O. Kuntze]. For. Microbiol. 2018, 9, 30–60. [Google Scholar] [CrossRef]
- Yang, S.Y.; Su, S.C.; Liu, T.; Fan, G.; Wang, J. First report of anthracnose caused by Colletotrichum gloeosporioides on pistachio (Pistacia vera) in China. Plant Dis. 2011, 95, 1314. [Google Scholar] [CrossRef]
- Sharma, G.; Pinnaka, A.K.; Shenoy, B.D. Resolving the Colletotrichum siamense species complex using ApMat marker. Fungal Divers. 2015, 71, 247–264. [Google Scholar] [CrossRef]
- Yaowen, Z.; Rong, S.; Yixue, M.; Qiqin, L.; Wei, L.; Gaoqing, Y. Colletotrichum siamense: A novel leaf pathogen of Sterculia nobilis Smith detected in China. For. Pathol. 2020, 50, e12575. [Google Scholar] [CrossRef]
- Afanador-Kafuri, L.; Minz, D.; Maymon, M.; Freeman, S. Characterization of Colletotrichum isolates from tamarillo, passiflora, and mango in colombia and identification of a unique species from the genus. Phytopathology 2003, 93, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Gazis, R.; Rehner, S.; Chaverri, P. Species delimitation in fungal endophyte diversity studies and its implications in ecological and biogeographic inferences. Mol. Ecol. 2011, 20, 3001–3013. [Google Scholar] [CrossRef]
Figure 1.
Symptoms of diseased winterberry holly. (a) Symptomatic leaves change from green to yellowish green, and gradually become grayish brown Coalesced irregular leaf spots eventually resulted in early plant defoliation. The arrow points to the spot. (b,c) A large area of irregular black–brown leaf spots with dense white mycelia on winterberry holly’s defoliation. The arrow points to white mycelia.
Figure 1.
Symptoms of diseased winterberry holly. (a) Symptomatic leaves change from green to yellowish green, and gradually become grayish brown Coalesced irregular leaf spots eventually resulted in early plant defoliation. The arrow points to the spot. (b,c) A large area of irregular black–brown leaf spots with dense white mycelia on winterberry holly’s defoliation. The arrow points to white mycelia.

Figure 2.
Symptoms of winterberry holly leaves after incubation with isolate LVY 9. In vitro inoculation, brown necrotic lesions with bright orange conidial masses appear (b) compared with control (a) after 7 days. The arrow points to orange conidia (b). In spray inoculation, grayish brown spots appear on tips of leaves with obvious curly phenotype compared with control (c) after 30 days. The arrow points to irregular spots (d). The mycelia are grayish brown with white edge (e) and reserve is turquoise (d) after strain LVY 9 on PDA for 7 d. Orange masses of conidia released from pycnidia after strain LVY 9 under a mixture of fluorescent white and UV light on PDA 20 d (g,h). The arrow points to orange conidia (g). Strain LVY 9 resembles Colletotrichum gloeosporioides by SEM and light microscopy.
Figure 2.
Symptoms of winterberry holly leaves after incubation with isolate LVY 9. In vitro inoculation, brown necrotic lesions with bright orange conidial masses appear (b) compared with control (a) after 7 days. The arrow points to orange conidia (b). In spray inoculation, grayish brown spots appear on tips of leaves with obvious curly phenotype compared with control (c) after 30 days. The arrow points to irregular spots (d). The mycelia are grayish brown with white edge (e) and reserve is turquoise (d) after strain LVY 9 on PDA for 7 d. Orange masses of conidia released from pycnidia after strain LVY 9 under a mixture of fluorescent white and UV light on PDA 20 d (g,h). The arrow points to orange conidia (g). Strain LVY 9 resembles Colletotrichum gloeosporioides by SEM and light microscopy.

Figure 3.
Conidia and appressorium characteristics of isolate LVY 9 on PDA. Conidia are smooth-walled, hyaline, and cylindrical, with obtuse to slightly rounded ends (a–c). Bars = 10, 2, 1 µm, respectively; appressoria appear brown to dark brown, ovoid, clavate, and slightly irregular in shape(d–g). Bars = 20, 10, 10, 5 µm, respectively.
Figure 3.
Conidia and appressorium characteristics of isolate LVY 9 on PDA. Conidia are smooth-walled, hyaline, and cylindrical, with obtuse to slightly rounded ends (a–c). Bars = 10, 2, 1 µm, respectively; appressoria appear brown to dark brown, ovoid, clavate, and slightly irregular in shape(d–g). Bars = 20, 10, 10, 5 µm, respectively.

Figure 4.
Maximum likelihood (ML) tree generated from the combined ACT, CAL, CHS-1, GAPDH and ITS sequence data of 59 taxa of Colletotrichum, of which 21 are in C.boninense complex, 34 are in C.gloeosporioides complex, 1 is in C.orbiculare complex, 1 is in C.acutatum complex, 2 are undefined. Clades with >75% ML_BS (left) and 0.50 BI-PP (right) are indicated by the corresponding support values. Parsimony and likelihood bootstrap support values ≥ 95% are indicated at the nodes and branches with Bayesian posterior probabilities above 0.95 given in bold. The ex-type strains are in bold. Dashes indicate support values lower than 75% ML-BS and 0.50 BI-PP. The tree is rooted with Monilochaetes infuscans CBS 869.96. The red arrow points to strain LVY9.
Figure 4.
Maximum likelihood (ML) tree generated from the combined ACT, CAL, CHS-1, GAPDH and ITS sequence data of 59 taxa of Colletotrichum, of which 21 are in C.boninense complex, 34 are in C.gloeosporioides complex, 1 is in C.orbiculare complex, 1 is in C.acutatum complex, 2 are undefined. Clades with >75% ML_BS (left) and 0.50 BI-PP (right) are indicated by the corresponding support values. Parsimony and likelihood bootstrap support values ≥ 95% are indicated at the nodes and branches with Bayesian posterior probabilities above 0.95 given in bold. The ex-type strains are in bold. Dashes indicate support values lower than 75% ML-BS and 0.50 BI-PP. The tree is rooted with Monilochaetes infuscans CBS 869.96. The red arrow points to strain LVY9.

Table 1.
Primers used in this study, with sequences and sources.
| Gene | Product Name | Primers | Sequence (5′–3′) | Reference |
|---|---|---|---|---|
| ACT | Actin | ACT-512F ACT-783R | ATGTGCAAGGCCGGTTTCGC TACGAGTCCTTCTGGCCCAT | [20] |
| CAL | Calmodulin | CL1C CL2C | GAATTCAAGGAGGCCTTCTC CTTCTGCATCATGAGCTGGAC | [13] |
| GAPDH | Glyceraldehyde-3- Phosphate dehydrogenase | GDF GDR | GCCGTCAACGACCCCTTCATTGA GGGTGGAGTCGTACTTGAGCATGT | [21] |
| CHS-1 | Chitin synthase | CHS-79F CHS-345R | TGGGGCAAGGATGCTTGGAAGAAG TGGAAGAACCATCTGTGAGAGTTG | [20] |
| ITS | Internal transcribed spacer | ITS-1F ITS-4 | CTTGGTCATTTAGAGGAAGTAA TCCTCCGCTTATTGATATGC | [22] [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Feng, L.; Zhang, Y.; Chen, W.; Mao, B. Colletotrichum siamense Strain LVY 9 Causing Spot Anthracnose on Winterberry Holly in China. Microorganisms 2023, 11, 976. https://doi.org/10.3390/microorganisms11040976
AMA Style
Feng L, Zhang Y, Chen W, Mao B. Colletotrichum siamense Strain LVY 9 Causing Spot Anthracnose on Winterberry Holly in China. Microorganisms. 2023; 11(4):976. https://doi.org/10.3390/microorganisms11040976
Chicago/Turabian StyleFeng, Lin, Yahui Zhang, Weiliang Chen, and Bizeng Mao. 2023. "Colletotrichum siamense Strain LVY 9 Causing Spot Anthracnose on Winterberry Holly in China" Microorganisms 11, no. 4: 976. https://doi.org/10.3390/microorganisms11040976
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.