

Bacillus Species: Excellent Biocontrol Agents against Tomato Diseases
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Tomato Diseases and Pathogens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Causative Agent | Symptoms | Epidemiology | Reference |
|---|---|---|---|---|
| Early blight | Alternaria solani | Black-brown, concentric lesions on the leaves, stems, and fruits; yellowing. | It survives on plant debris, seeds, volunteer Solanaceous crops, and soil; favoured by high humidity and temperatures of 24–29 °C. | [22] |
| Septoria leaf spot | Septoria lycopersici | Tan-to-grey spots with dark margins on the leaves. | It survives on plant debris, seeds, volunteer Solanaceous crops, and weeds; favoured by high humidity and temperatures above 25 °C. | [23] |
| Grey mould | Botrytis cinerea | Brown lesions on the leaves and stems; pale or white rings on the fruits. | It survives on plant debris, seeds, soil, and various hosts; favoured by high humidity and temperatures of 18–23 °C. | [28] |
| Fusarium vascular wilt | Fusarium oxysporum f. sp. lycopersici | Yellowing; wilting; browning; stunted growth. | It survives in the soil and on plant debris, seeds, and seedlings; favoured by high humidity and temperatures of 20–30 °C. | [29] |
| Fusarium crown and root rot | F. oxysporum f. sp. radicis-lycopersici | Yellowing; wilting; browning; stunted growth; stem and root discoloration. | It survives in the soil and on plant debris, seeds, and seedlings; favoured by high humidity and temperatures of 10–20 °C. | [33] |
| Verticillium wilt | Verticillium dahliae | Yellowing; wilting; stunted growth; v-shaped lesions on the leaves; stem discoloration. | It survives in the soil and on plant debris; favoured by high humidity and temperatures of 21–30 °C. | [35] |
| Late blight | Phytophthora infestans | Green-black lesions on the leaves; dark spots on the fruits. | It survives on plants, tubers, soil, seeds, Solanaceous crops, and weeds; favoured by high humidity and temperatures of 10–20 °C. | [39] |
| Bacterial speck | Pseudomonas syringae pv. tomato | Brown-black spots on the leaves; stunted growth; dark specks on the fruits. | It survives on plant debris, soil, seeds, and weeds; favoured by high humidity and temperatures of 13–25 °C. | [45] |
| Bacterial wilt and canker | Clavibacter michiganensis subsp. michiganensis | Wilting; yellowing; stunted growth; stem discoloration; white spots with a necrotic centre on the fruits (bird’s eye). | It survives on plant debris, soil, seeds, and weeds; favoured by high humidity and temperatures of 24–28 °C. | [48] |
| Bacterial spot | Xanthomonas campestris pv. vesicatoria | Elliptical, dark, chlorotic spots on the leaves, stems, and fruits. | It survives on plant debris, soil, and seeds; favoured by high humidity and temperatures of 23–30 °C. | [50] |
| Bacterial wilt | Ralstonia solanacearum | Wilting; root rot; stem discoloration and decay. | It survives on plant debris, soil, seeds, seedlings, and weeds; favoured by high humidity and temperatures above 29 °C. | [53] |
3. Bacillus Species: General Characteristics and Benefits of Application
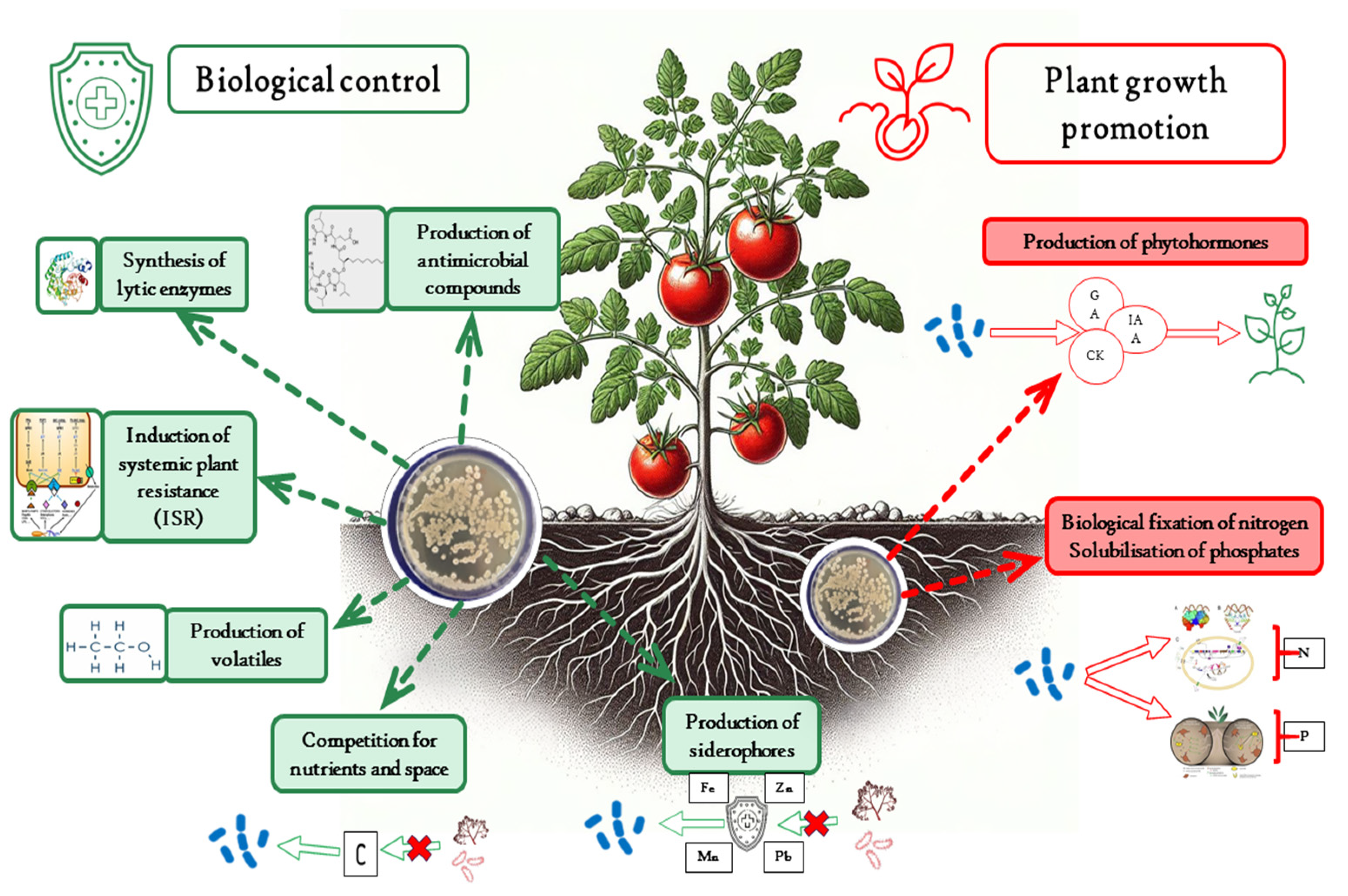
4. Mechanisms of Biological Control
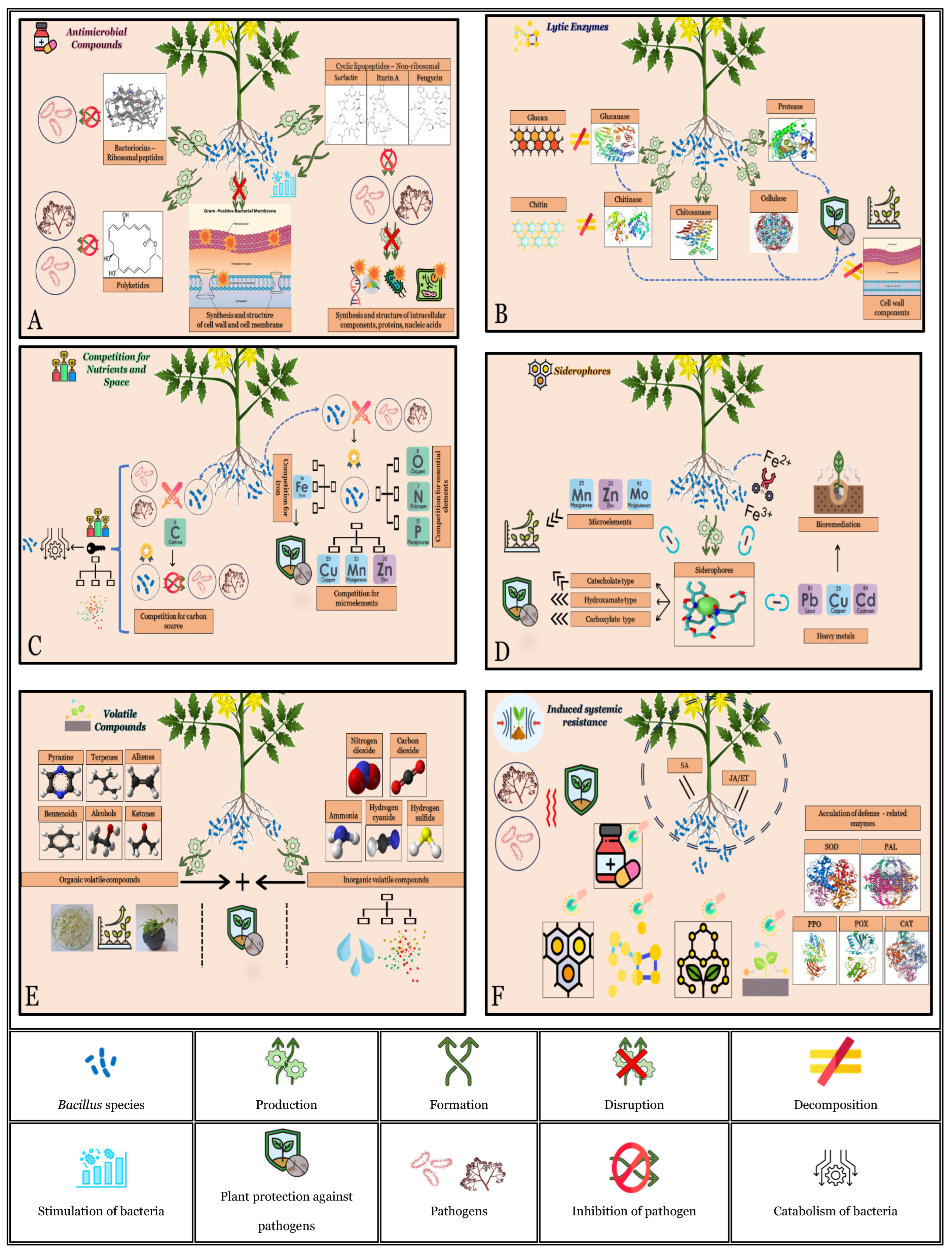
4.1. Antimicrobial Compounds
4.2. Lytic Enzymes
4.3. Competition for Nutrients and Space
4.4. Siderophores
4.5. Volatile Compounds
| Bacillus Strain | Part of Tomato | Pathogen/Disease | Mode of Action | Reference |
|---|---|---|---|---|
| B. pumilus PTB180 B. subtilis PTB185 | Leaves | B. cinerea/grey mould | Surfactin/surfactin, iturin, fengycin | [90] |
| B. velezensis NKMV-3 | Leaves | A. solani/early blight | Surfactin, iturin, fengycin | [91] |
| B. amyloliquefaciens XJ-BV2007 | Fruits | A. alternata/black spot | Fengycin | [92] |
| B. amyloliquefaciens PPL | Plants | Fusarium oxysporum f. sp. lycopersici/Fusarium wilt | Fengycin | [93] |
| B. subtilis MB14 B. amyloliquefaciens MB101 | Roots | R. solani/root rot | Surfactin, fengycin bacillomycin, iturin | [94] |
| B. amyloliquefaciens Oj-2.16 | Plants | V. dahliae/verticillium wilt | Surfactin, iturin, fengycin, bacilysin | [95] |
| B. methylotrophicus DR-08 | Plants | R. solanacearum/bacterial wilt | Difficidin, oxydifficidin | [96] |
| B. amyloliquefaciens DSBA-11 | Plants | R. solanacearum/bacterial wilt | Difficidin, macrolactin, bacillaene | [98] |
| B. velezensis SDTB038 | Plants | Fusarium oxysporum f. sp. radicis-lycopersici/Fusarium crown and root rot wilt | Macrolactin H, bacillaene, fengycin, difficidin, bactin, bacilysin, surfactin | [99] |
| B. subtilis V26 | Fruits | B. cinerea/grey mould | Chitosanase | [104] |
| B. velezensis KS04AU | Roots | F. oxysporum f. sp. radicis-lycopersici/Fusarium rot | Chitinase, cellulase, amylase, protease, lipase, phytase | [105] |
| Several Bacillus spp. | Plants | Fusarium oxysporum f. sp. lycopersici/Fusarium wilt | Cellulase, protease | [106] |
| B. velezensis GF267 | Plants | X. perforans/bacterial spot | Competition for nutrients and space | [110] |
| B. amyloliquefaciens CM-2 and T-5 | Plants | R. solanacearum/bacterial wilt | Competition for nutrients and space | [111] |
| B. amyloliquefaciens SQYUV 162 | Plants | R. solanacearum/bacterial wilt | Competition for nutrients and space | [112] |
| B. amyloliquefaciens SG08-09 B. subtilis SG09-12 | Plants | B. cinerea/grey mould | Siderophores, protease, cellulase, ammonia, IAA | [117] |
| B. velezensis RC116 | Plants | R. solanacearum/bacterial wilt | Protease, amylase, lipase, siderophores, IAA | [118] |
| B. amyloliquefaciens S1 | Plants | C. michiganensis ssp. michiganensis/bacterial canker | Siderophores, chitinase, cellulase, protease, lipase | [119] |
| B. subtilis EB-28 | Leaves | B. cinerea/grey mould | Hydrogen sulphide | [125] |
| B. subtilis BS-01 | Leaves | A. solani/early blight | Volatile organic compounds | [127] |
| B. tequilensis XK29 | Fruits | B. cinerea/grey mould | Volatile compounds | [128] |
| B. subtilis BS 21-1 | Plants | B. cinerea/Botrytis rot | Induced resistance | [129] |
| B. aryabhattai SRB02 | Plants | F. oxysporum f. sp. lycopersici/Fusarium wilt | Induced resistance | [15] |
| B. subtilis OTPB1 | Leaves | P. infestans/late blight A. solani/early blight | Induced resistance | [130] |
| B. amyloliquefaciens FZB42 | Stems | S. sclerotiorum/Sclerotinia rot | Induced resistance | [131] |
| B. cabrialesii BH5 | Leaves | B. cinerea/grey mould | Induced resistance | [132] |
| B. velezensis YYC | Leaves | P. solanacearum/bacterial wilt | Induced resistance | [133] |
| B. subtilis CBR05 | Leaves | X. campestris pv. vesicatoria/ bacterial spot | Induced resistance | [134] |
| B. subtilis SR22 | Roots | R. solani/Rhizoctonia rot | Induced resistance | [135] |
4.6. Induced Resistance
5. Combined Strategies for Tomato Disease Management
| Bacillus Strain | Combination with Antagonists/Fungicide | Pathogen/Disease | Reference |
|---|---|---|---|
| B. subtilis | Plant nutrients (NPK, Zn, Mg, B) | A. solani/early blight | [141] |
| B. subtilis | NPK fertiliser Trichoderma asperellum | Pythium aphanidermatum/damping off | [142] |
| B. amyloliquefaciens | Cattle manure compost Amino acid fertiliser | R. solanacearum/bacterial wilt | [143] |
| B. subtilis subsp. subtilis | Oak-bark compost | P. infestans/late blight | [144] |
| B. methylotrophicus | Fungicide fluopimomide | B. cinerea/grey mould | [145] |
| B. subtilis B-001 | Bactericide Saisentong | R. solanacearum/bacterial wilt | [146] |
| B. subtilis B. amyloliquefaciens B. pumilus | Acibenzolar-S-methyl hymexazol | F. oxysporum f. sp. radicis-lycopersici/Fusarium crown and root rot | [147] |
| Bacillus spp. B. subtilis | Reynoutria sachalinensis Malaleuca alternifolia Harpin αβ proteins Bee honey | Grey mould/B. cinerea Powdery mildew/Leveillula taurica | [148] |
| B. amyloliquefaciens | Peppermint oil | F. oxysporum f. sp. lycopersici/Fusarium wilt | [149] |
| B. subtilis B. thuringiensis | Enterobacter cloacae Pythium oligandrum | R.solani/Rhizoctonia root rot | [150] |
| B. amyloliquefaciens | Trichoderma asperellum | X. perforans/bacterial spot | [152] |
| B. subtilis | Trichoderma spp. Pseudomonas fluorescens | A.solani/early blight | [153] |
| Bacillus sp. B. velezensis | Funneliformis mosseae Glomus fasciculatum | Fusarium oxysporum f. sp. lycopersici/Fusarium wilt | [154] |
| B. subtilis PPB9 | Stenotrophomonas maltophila PPB3 | Sclerotium rolfsii/Southern blight | [155] |
| B. velezensis | P. fluorescens | R. solanacearum/bacterial wilt | [156] |
| B. velezensis | P. fluorescens | R.solani/Rhizoctonia root rot | [157] |
| B. subtilis | Trichoderma viride | Fusarium solani/Fusarium root rot | [158] |
6. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Database. Food and Agriculture Organization Statistics. Available online: https://www.fao.org/faostat/en/ (accessed on 15 November 2023).
- Ali, M.-Y.; Sina, A.A.I.; Khandker, S.S.; Neesa, L.; Tanvir, E.M.; Kabir, A.; Khalili, M.I.; Gan, S.H. Nutritional composition and bioactive compounds in tomatoes and their impact on human health and disease: A review. Foods 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandić, A.; Noris, E.; Matić, S. A Review of the most common and economically important diseases that undermine the cultivation of tomato crop in the Mediterranean Basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- Montenegro, I.; Madrid, A.; Cuellar, M.; Seeger, M.; Alfaro, J.F.; Besoain, X.; Martínez, J.P.; Ramirez, I.; Olguín, Y.; Valenzuela, M. Biopesticide activity from drimanic compounds to control tomato pathogens. Molecules 2018, 23, 2053. [Google Scholar] [CrossRef]
- Attia, M.S.; El-Wakil, D.A.; Hashem, A.H.; Abdelaziz, A. Antagonistic effect of plant growth-promoting fungi against fusarium wilt disease in tomato: In vitro and in vivo study. Appl. Biochem. Biotechnol. 2022, 194, 5100–5118. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Gao, Z.; Yang, W. Breeding for resistance to tomato bacterial diseases in China: Challenges and prospects. Hortic. Plant J. 2018, 4, 193–207. [Google Scholar] [CrossRef]
- Butsenko, L.; Pasichnyk, L.; Kolomiiets, Y.; Kalinichenko, A. The effect of pesticides on the tomato bacterial speck disease pathogen Pseudomonas syringae pv. tomato. Appl. Sci. 2020, 10, 3263. [Google Scholar] [CrossRef]
- Ma, M.; Taylor, P.W.J.; Chen, D.; Vaghefi, N.; He, J.Z. Major soilborne pathogens of field processing tomatoes and management strategies. Microorganisms 2023, 11, 263. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Influence of synthetic fertilizers and pesticides on soil health and soil microbiology. In Agrochemicals Detection, Treatment and Remediation; Butterworth-Heinemann: Oxford, UK, 2020; pp. 25–54. [Google Scholar]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Jacobsen, B.J.; Zidack, N.K.; Larson, B.J. The role of Bacillus-based biological control agents in integrated pest management systems: Plant diseases. Phytopathology 2004, 94, 1272–1275. [Google Scholar] [CrossRef] [PubMed]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef] [PubMed]
- Taha, N.A.; Elsharkawy, M.M.; Shoughy, A.A.; El-Kazzaz, M.K.; Khedr, A.A. Biological control of postharvest tomato fruit rots using Bacillus spp. and Pseudomonas spp. Egypt. J. Biol. Pest Control 2023, 33, 106. [Google Scholar] [CrossRef]
- Syed Nabi, R.B.; Shahzad, R.; Tayade, R.; Shahid, M.; Hussain, A.; Ali, M.W.; Yun, B.W. Evaluation potential of PGPR to protect tomato against Fusarium wilt and promote plant growth. PeerJ 2021, 9, e11194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, Y.; Liang, F.; Ma, Y.; Wu, X. An extract produced by Bacillus sp. BR3 influences the function of the GacS/GacA two-component system in Pseudomonas syringae pv. tomato DC3000. Front. Microbiol. 2019, 10, 2005. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B.; Zitter, T.A.; Momol, M.T.; Miller, S.A. Compendium of Tomato Diseases and Pests, 2nd ed.; APS Press: St. Paul, MN, USA, 2016; pp. 15–119. [Google Scholar] [CrossRef]
- Ahmed, F.A.; Sipes, B.S.; Alvares, A.M. Postharvest diseases of tomato and natural products for disease management. Afr. J. Agric. Res. 2017, 12, 684–691. [Google Scholar] [CrossRef]
- Rodrigues, M.H.P.; Furlong, E.B. Fungal diseases and natural defense mechanisms of tomatoes (Solanum lycopersicum): A review. Physiol. Mol. Plant Pathol. 2022, 122, 101906. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R.; Kashyap, P.L.; Srivastava, A.K. Rapid detection and quantification of Alternaria solani in tomato. Sci. Hortic. 2013, 151, 184–189. [Google Scholar] [CrossRef]
- Mugao, L. Morphological and molecular variability of Alternaria solani and Phytophthora infestans causing tomato blights. Int. J. Microbiol. 2023, 2023, 8951351. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Ávila, M.C.R.; Lourenço, V.L., Jr.; Quezado-Duval, A.M.; Becker, W.F.; Abreu-Tarazi, M.F.; Borges, L.C.; Nascimento, A.D.R. Field validation of TOMCAST modified to manage Septoria leaf spot on tomato in the central-west region of Brazil. Crop Prot. 2020, 138, 105333. [Google Scholar] [CrossRef]
- Silva, B.N.; Picanco, B.B.M.P.; Hawerroth, C.; Silva, L.C.; Rodrigues, F.A. Physiological and biochemical insights into induced resistance on tomato against septoria leaf spot by a phosphite combined with free amino acids. Physiol. Mol. Plant Pathol. 2022, 120, 101854. [Google Scholar] [CrossRef]
- Mani, S.D.; Pandey, S.; Govindan, M.; Muthamilarasan, M.; Nagarathnam, R. Transcriptome dynamics underlying elicitor-induced defense responses against Septoria leaf spot disease of tomato (Solanum lycopersicum L.). Physiol. Mol. Biol. Plants 2021, 27, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Barquero, M.; González-Andrés, F. Insight into the microbiological control strategies against Botrytis cinerea using systemic plant resistance activation. Agronomy 2020, 10, 1822. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Zhang, Z.; Li, B.; Qin, G.; Tian, S. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef]
- Fillinger, S.; Elad, Y. Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Springer: New York, NY, USA, 2016. [Google Scholar]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.P.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, A.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Ponsankar, A.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Pandiyan, R.; Karthi, S.; Kalaivani, K.; Chellappandian, M.; Narayanaswamy, R.; Thanugaivel, A.; Patcharin, K.; et al. Systematic induced resistance in Solanum lycopersicum (L.) against vascular wilt pathogen (Fusarium oxysporum f. sp. lycopersici) by Citrullus colocynthis and Trichoderma viride. PLoS ONE 2023, 18, e0278616. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Dinesh, K.; Nirmala, N.S.; Kumar, A.; Chakraborti, D.; Bhattacharyya, A. Insight into tomato plant immunity to necrotrophic fungi. Curr. Res. Biotechnol. 2023, 6, 100144. [Google Scholar] [CrossRef]
- Li, X.; Wang, Q.; Li, H.; Wang, X.; Zhang, R.; Yang, X.; Jiang, Q.; Shi, Q. Revealing the mechanisms for linalool antifungal activity against Fusarium oxysporum and its efficient control of Fusarium wilt in tomato plants. Int. J. Mol. Sci. 2023, 24, 458. [Google Scholar] [CrossRef]
- Aravena, R.; Besoain, X.; Riquelme, N.; Salinas, A.; Valenzuela, M.; Oyanedel, E.; Barros, W.; Olguin, Y.; Madrid, A.; Alvear, M.; et al. Antifungal nanoformulation for biocontrol of tomato root and crown rot caused by Fusarium oxysporum f. sp. radicis-lycopersici. Antibiotics 2021, 10, 1132. [Google Scholar] [CrossRef]
- Acharya, B.; Ingram, T.W.; Oh, Y.; Adhikari, T.B.; Dean, R.A.; Louws, F.J. Opportunities and challenges in studies of host-pathogen interactions and management of Verticillium dahliae in tomatoes. Plants 2020, 9, 1622. [Google Scholar] [CrossRef]
- Retief, E.; Lamprecht, S.; McLeod, A. Characterisation and pathogenicity of Verticillium dahliae isolates associated with Verticillium wilt of tomato in the Limpopo Province of South Africa. J. Plant Pathol. 2023, 105, 1465–1481. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef]
- Buhtz, A.; Hohe, A.; Schwarz, D.; Grosch, R. Effects of Verticillium dahliae on tomato root morphology considering plant growth response and defence. Plant Pathol. 2017, 66, 667–676. [Google Scholar] [CrossRef]
- Garcia, P.G.; Neves dos Santos, F.; Zanotta, S.; Eberlin, M.N.; Carazzone, C. Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants. Molecules 2018, 23, 3330. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, P.; Singh, P.; Kethiravan, D.; Ramathani, I.; Ramakrishnan, N. Late blight in tomato: Insights into the pathogenesis of the aggressive pathogen Phytophthora infestans and future research priorities. Planta 2021, 253, 119. [Google Scholar] [CrossRef]
- Maxim, A.; Albu, V.C.; Vodnar, D.C.; Mihăiescu, T.; Mang, Ș.M.; Camele, I.; Trotta, V.; Bonomo, M.G.; Mihalescu, L.; Sandor, M.; et al. Assessment of Tomato (Solanum lycopersicum) Landraces for Their Agronomic, Biochemical Characteristics and Resistance to Phytophthora infestans. Agronomy 2023, 13, 21. [Google Scholar] [CrossRef]
- Skliros, D.; Papazoglou, P.; Gkizi, D.; Paraskevopoulou, E.; Katharios, P.; Goumas, D.E.; Tjamos, S.; Flemetakis, E. In planta interactions of a novel bacteriophage against Pseudomonas syringae pv. tomato. Appl. Microbiol. Biotechnol. 2023, 107, 3801–3815. [Google Scholar] [CrossRef] [PubMed]
- Basim, H.; Basim, E.; Yilmaz, S.; Dickstein, E.R.; Jones, J.B. An outbreak of bacterial speck caused by Pseudomonas syringae pv. tomato on tomato transplants grown in commercial seedling companies located in the Western Mediterranean Region of Turkey. Plant Dis. 2004, 88, 1050. [Google Scholar] [CrossRef] [PubMed]
- Santamaría-Hernando, S.; López-Maroto, Á.; Galvez-Roldán, C.; Munar-Palmer, M.; Monteagudo-Cascales, E.; Rodríguez-Herva, J.J.; Tino Krell, T.; López-Solanilla, E. Pseudomonas syringae pv. tomato infection of tomato plants is mediated by GABA and l-Pro chemoperception. Mol. Plant Pathol. 2022, 23, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Chai, A.; Ben, H.Y.; Guo, W.T.; Shi, Y.X.; Xie, X.W.; Li, L.; Li, B.J. Quantification of viable cells of Pseudomonas syringae pv. tomato in tomato seed using propidium monoazide and a real-time PCR assay. Plant Dis. 2020, 104, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Zhao, L.; Gao, Y.G.; Xia, Y. Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae. Plants 2023, 12, 1765. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Q.; Zhou, P.; Zhao, W.; Sun, X. Evaluation of droplet digital PCR for the detection of black canker disease in tomato. Plant Dis. 2022, 106, 395–405. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.M.; Bagy, H.M.M.K.; Hashem, M.; Alamri, S.A.M.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt. J. Biol. Pest Control 2019, 29, 54. [Google Scholar] [CrossRef]
- Tripathi, R.; Vishunavat, K.; Tewari, R. Morphological and molecular characterization of Clavibacter michiganensis subsp. michiganensis causing bacterial canker in tomatoes. Physiol. Mol. Plant Pathol. 2022, 119, 101833. [Google Scholar] [CrossRef]
- Malliarakis, D.; Pagoulatou, M.G.; Mpalantinaki, E.; Trantas, E.; Ververidis, F.; Goumas, D.E. Phylogenetic diversity of Clavibacter michiganensis subsp. michiganensis isolates causing bacterial canker of tomato in Greece. J. Plant Pathol. 2023, 105, 1403–1419. [Google Scholar] [CrossRef]
- Abrahamian, P.; Klein-Gordon, J.M.; Jones, J.B.; Vallad, G.E. Epidemiology, diversity, and management of bacterial spot of tomato caused by Xanthomonas perforans. Appl. Microbiol. Biotechnol. 2021, 105, 6143–6158. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, T.K.; Woudt, B.; Lastdrager, J.; Berendsen, S.; Koenraadt, H. Development and validation of real-time PCR tests for specific identification of Xanthomonas species causing bacterial spot disease on tomato (Solanum lycopersicum) and pepper (Capsicum annuum). EPPO Bull. 2023, 53, 416–424. [Google Scholar] [CrossRef]
- Bernal, E.; Rotondo, F.; Roman-Reyna, V.; Klass, T.; Timilsina, S.; Minsavage, G.V.; Iruegas-Bocardo, F.; Goss, E.M.; Jones, J.B.; Jacobs, J.M.; et al. Migration Drives the Replacement of Xanthomonas perforans races in the absence of widely deployed resistance. Front. Microbiol. 2022, 13, 826386. [Google Scholar] [CrossRef]
- Bamazi, B.; Banito, A.; Ayisah, K.D.; Sikirou, R.; Paret, M.L.; Kunwar, S.; Kamde, K.; Tchalla, P.; Afole, S.L.N.A. Distribution and incidence of tomato bacterial wilt caused by Ralstonia solanacearum in the Central Region of Togo. Plant Health Prog. 2022, 23, 235–240. [Google Scholar] [CrossRef]
- Mekonnen, H.; Kibret, M.; Assefa, F. Plant growth promoting rhizobacteria for biocontrol of tomato bacterial wilt caused by Ralstonia solanacearum. Int. J. Agron. 2022, 2022, 1489637. [Google Scholar] [CrossRef]
- Rivera-Zuluaga, K.; Hiles, R.; Barua, P.; Caldwell, D.; Iyer-Pascuzzi, A.S. Getting to the root of Ralstonia invasion. Sem. Cell Dev. Biol. 2023, 148–149, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, D.; Kim, B.S.; Iyer-Pascuzzi, A.S. Ralstonia solanacearum differentially colonizes roots of resistant and susceptible tomato plants. Phytopathology 2017, 107, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Kruidhof, H.M.; Elmer, W.H. Cultural methods for greenhouse pest and disease management. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M., Albajes, R., Nicot, P., Eds.; Springer: Cham, Switzerland, 2020; pp. 285–330. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Gardener, B.M.; Miller, S.A. Soil Borne Disease Management in Organic Vegetable Production. Org. Agric. 2012. Available online: https://eorganic.org/node/7581 (accessed on 15 November 2023).
- Lee, Y.; Choi, C.; Kim, S.H.; Yun, J.; Chang, S.; Kim, Y.S.; Hong, J. Chemical pesticides and plant essential oils for disease control of tomato bacterial wilt. Plant Pathol. J. 2012, 28, 32–39. [Google Scholar] [CrossRef]
- Lima, G.P.P.; Gómez, H.A.G.; Seabra Junior, S.; Maraschin, M.; Tecchio, M.A.; Borges, C.V. Functional and nutraceutical compounds of tomatoes as affected by agronomic practices, postharvest management, and processing methods: A mini review. Front. Nutr. 2022, 9, 868492. [Google Scholar] [CrossRef]
- Kumari, D.; Gopireddy, B.M.; Kumari, A.; Vijaya, M.; Suresh, V.; Nayak, H. Integrated disease management in tomato. J. Entomol. Zool. Stud. 2020, 8, 281–284. [Google Scholar]
- Ayaz, M.; Li, C.-H.; Ali, Q.; Zhao, W.; Chi, Y.-K.; Shafiq, M.; Ali, F.; Yu, X.-Y.; Yu, Q.; Zhao, J.-T.; et al. Bacterial and fungal biocontrol agents for plant disease protection: Journey from lab to field, current status, challenges, and global perspectives. Molecules 2023, 28, 6735. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef]
- Stenberg, J.A.; Sundh, I.; Becher, P.G.; Björkman, C.; Dubey, M.; Egan, P.A.; Friberg, H.; Gil, J.F.; Jensen, D.F.; Jonsson, M.; et al. When is it biological control? A framework of definitions, mechanisms, and classifications. J. Pest Sci. 2021, 94, 665–676. [Google Scholar] [CrossRef]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogen using microbial antagonists. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as biological control agents of plant diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef] [PubMed]
- Tsotetsi, T.; Nephali, L.; Malebe, M.; Tugizimana, F. Bacillus for plant growth promotion and stress resilience: What have we learned? Plants 2018, 11, 2482. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, D.R.; Perkins, J.B. The genus Bacillus. In Practical Handbook of Microbiology; CRC Press: Boca Raton, FL, USA, 2008; pp. 310–319. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2019, 128, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Piggot, P.; Hilbert, D. Sporulation of Bacillus subtilis. Curr. Opin. Microbiol. 2004, 7, 579–586. [Google Scholar] [CrossRef]
- Haque, M.M.; Mosharaf, M.K.; Khatun, M.; Haque, M.A.; Biswas, M.S.; Islam, M.S.; Islam, M.M.; Shozib, H.B.; Miah, M.M.U.; Molla, A.H.; et al. Biofilm producing rhizobacteria with multiple plant growth-promoting traits promote growth of tomato under water-deficit stress. Front. Microbiol. 2020, 11, 542053. [Google Scholar] [CrossRef]
- Sansinenea, E. Bacillus spp.: As plant growth-promoting bacteria. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Singh, H., Keswani, C., Reddy, M., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019; pp. 225–237. [Google Scholar] [CrossRef]
- Su, Y.; Liu, C.; Fang, H.; Zhang, D. Bacillus subtilis: A universal cell factory for industry, agriculture, biomaterials and medicine. Microb. Cell Fact. 2020, 19, 173. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, Z.; Shao, J.; Xu, Z.; Liu, Y.; Xun, W.; Miao, Y.; Shen, Q.; Zhang, R. Biocontrol mechanisms of Bacillus: Improving the efficiency of green agriculture. Microb. Biotechnol. 2023, 16, 2250–2263. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting Rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Abd_Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Xu, M.; Sheng, J.; Chen, J.; Men, Y.; Gan, L.; Guo, S.; Shen, L. Bacterial community compositions of tomato (Lycopersicum esculentum Mill.) seeds and plant growth promoting activity of ACC deaminase producing Bacillus subtilis (HYT-12-1) on tomato seedlings. World J. Microbiol. Biotechnol. 2014, 30, 835–845. [Google Scholar] [CrossRef]
- Jain, S.; Varma, A.; Choudhary, D.K. Perspectives on nitrogen-fixing Bacillus species. In Soil Nitrogen Ecology; Cruz, C., Vishwakarma, K., Choudhary, D.K., Varma, A., Eds.; Springer: Cham, Switzerland, 2021; pp. 359–369. [Google Scholar] [CrossRef]
- Pattnaik, S.; Mohapatra, B.; Gupta, A. Plant growth-promoting microbe mediated uptake of essential nutrients (Fe, P, K) for crop stress management: Microbe–soil–plant continuum. Front. Agron. 2021, 3, 689972. [Google Scholar] [CrossRef]
- Berdi, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Epparti, P.; Eligar, S.M.; Sattur, A.; Kumar, B.S.G.; Halami, P.M. Characterization of dual bacteriocins producing Bacillus subtilis SC3.7 isolated from fermented food. Food Sci. Technol. 2022, 154, 112854. [Google Scholar] [CrossRef]
- Salazar, F.; Ortiz, A.; Sansinenea, E. Characterisation of two novel bacteriocin-like substances produced by Bacillus amyloliquefaciens ELI149 with broad-spectrum antimicrobial activity. J. Glob. Antimicrob. Resist. 2017, 11, 177–182. [Google Scholar] [CrossRef]
- Schneider, T.; Müller, A.; Miess, H.; Gross, H. Cyclic lipopeptides as antibacterial agents—Potent antibiotic activity mediated by intriguing mode of actions. Int. J. Med. Microbiol. 2014, 304, 37–43. [Google Scholar] [CrossRef]
- Helmy, N.M.; Parang, K. Cyclic peptides with antifungal properties derived from bacteria, fungi, plants, and synthetic sources. Pharmaceuticals 2023, 16, 892. [Google Scholar] [CrossRef]
- Gutiérrez-Chávez, C.; Benaud, N.; Ferrari, B.C. The ecological roles of microbial lipopeptides: Where are we going? Comput. Struct. Biotechnol. J. 2021, 19, 1400–1413. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.R.; Kanwar, S.S. Lipopeptides as the antifungal and antibacterial agents: Applications in food safety and therapeutics. BioMed Res. Int. 2015, 2015, 473050. [Google Scholar] [CrossRef] [PubMed]
- Bouchard-Rochette, M.; Machrafi, Y.; Cossus, L.; Nguyen, T.T.A.; Antoun, H.; Droit, A.; Tweddell, R.J. Bacillus pumilus PTB180 and Bacillus subtilis PTB185: Production of lipopeptides, antifungal activity, and biocontrol ability against Botrytis cinerea. Biol. Control 2022, 170, 104925. [Google Scholar] [CrossRef]
- Vignesh, M.; Shankar, S.R.M.; Subramani, N.; VedhaHari, B.N.; Ramzadevi, D. Study on spray-drying of Bacillus velezensis NKMV-3 strain, its formulation and bio efficacy against early blight of tomato. Biocatal. Agric. Biotechnol. 2022, 45, 102483. [Google Scholar] [CrossRef]
- Jia, Q.; Fan, Y.; Duan, S.; Qin, Q.; Ding, Y.; Yang, M.; Wang, Y.; Liu, F.; Wang, C. Effects of Bacillus amyloliquefaciens XJ-BV2007 on growth of Alternaria alternata and production of tenuazonic acid. Toxins 2023, 15, 53. [Google Scholar] [CrossRef]
- Kang, B.R.; Park, J.S.; Jung, W.J. Antifungal evaluation of fengycin isoforms isolated from Bacillus amyloliquefaciens PPL against Fusarium oxysporum f. sp. lycopersici. Microb. Pathog. 2020, 149, 104509. [Google Scholar] [CrossRef]
- Solanki, M.K.; Singh, R.K.; Srivastava, S.; Kumar, S.; Kashyup, P.L.; Srivastava, A.K. Characterization of antagonistic-potential of two Bacillus strains and their biocontrol activity against Rhizoctonia solani in tomato. J. Basic Microbiol. 2013, 53, 82–90. [Google Scholar] [CrossRef]
- Pei, D.; Zhang, Q.; Zhu, X.; Zhang, L. Biological control of Verticillium wilt and growth promotion in tomato by rhizospheric soil-derived Bacillus amyloliquefaciens Oj-2.16. Pathogens 2023, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Im, S.M.; Yu, N.H.; Joen, H.W.; Kim, S.O.; Park, H.W.; Park, A.R.; Kim, J.C. Biological control of tomato bacterial wilt by oxydifficidin and difficidin-producing Bacillus methylotrophicus DR-08. Pestic. Biochem. Physiol. 2020, 163, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Devappa, V.; Yadav, D.K. Suppression of tomato bacterial wilt incited by Ralstonia pseudosolanacearum using polyketide antibiotic-producing Bacillus spp. isolated from rhizospheric soil. Agriculture 2022, 12, 2009. [Google Scholar] [CrossRef]
- Chen, Q.; Qiu, Y.; Yuan, Y.; Wang, K.; Wang, H. Biocontrol activity and action mechanism of Bacillus velezensis strain SDTB038 against Fusarium crown and root rot of tomato. Front. Microbiol. 2022, 13, 994716. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Khalil, M.S.M.; Hassan, M.H.A.R.; Mahmoud, A.F.; Morsy, K.M.M. Involvement of secondary metabolites and extracellular lytic enzymes produced by plant growth promoting rhizobacteria in inhibiting the soilborne pathogens in faba bean plants. J. Trop. Plant Pests Dis. 2022, 22, 100–108. [Google Scholar] [CrossRef]
- Kumar, M.; Brar, A.; Yadav, M.; Chawade, A.; Vivekanand, V.; Pareek, N. Chitinases—Potential candidates for enhanced plant resistance towards fungal pathogens. Agriculture 2018, 8, 88. [Google Scholar] [CrossRef]
- Ghasemi, S.; Ahmadian, G.; Jelodar, N.B.; Rahimian, H.; Ghandili, S.; Dehestani, A.; Shariati, P. Antifungal chitinases from Bacillus pumilus SG2: Preliminary report. World J. Microbiol. Biotechnol. 2010, 26, 1437–1443. [Google Scholar] [CrossRef]
- Kilani-Feki, O.; Khedher, S.B.; Dammak, M.; Kamoun, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Tounsi, S. Improvement of antifungal metabolites production by Bacillus subtilis V26 for biocontrol of tomato postharvest disease. Biol. Control 2016, 95, 73–82. [Google Scholar] [CrossRef]
- Diabankana, R.G.C.; Shulga, E.U.; Validov, S.Z.; Afordoanyi, D.M. Genetic characteristics and enzymatic activities of Bacillus velezensis KS04AU as a stable biocontrol agent against phytopathogens. Int. J. Plant Biol. 2022, 13, 201–222. [Google Scholar] [CrossRef]
- Rocha, F.Y.O.; de Oliveira, C.M.; da Silva, P.R.A.; de Melo, L.H.V.; do Carmo, M.G.F.; Baldani, J.I. Taxonomical and functional characterization of Bacillus strains isolated from tomato plants and their biocontrol activity against races 1, 2 and 3 of Fusarium oxysporum f. sp. lycopersici. Appl. Soil Ecol. 2017, 120, 8–19. [Google Scholar] [CrossRef]
- Dubinkina, V.; Fridman, Y.; Pandey, P.P.; Maslov, S. Multistability and regime shifts in microbial communities explained by competition for essential nutrients. eLife 2019, 8, e49720. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Mates, A.P.K.; Pontes, N.C.; Halfeld-Vieira, B.A. Bacillus velezensis GF267 as a multi-site antagonist for the control of tomato bacterial spot. Biol. Control 2019, 137, 104013. [Google Scholar] [CrossRef]
- Tan, S.; Jiang, Y.; Song, S.; Huang, J.; Ling, N.; Xu, Y.; Shen, Q. Two Bacillus amyloliquefaciens strains isolated using the competitive tomato root enrichment method and their effects on suppressing Ralstonia solanacearum and promoting tomato plant growth. Crop Prot. 2013, 43, 134–140. [Google Scholar] [CrossRef]
- Wu, K.; Su, L.; Fang, Z.; Yuan, S.; Wang, L.; Shen, B.; Shen, Q. Competitive use of root exudates by Bacillus amyloliquefaciens with Ralstonia solanacearum decreases the pathogenic population density and effectively controls tomato bacterial wilt. Sci. Hortic. 2017, 218, 132–138. [Google Scholar] [CrossRef]
- Khan, A.; Doshi, H.V.; Thakur, M.C. Bacillus spp.: A prolific siderophore producer. In Bacilli and Agrobiotechnology; Islam, M., Rahman, M., Pandey, P., Jha, C., Aeron, A., Eds.; Springer: Cham, Switzerland, 2016; pp. 309–323. [Google Scholar] [CrossRef]
- Saha, R.; Saha, N.; Donofrio, R.S.; Bestervelt, L.L. Microbial siderophores: A mini review. J. Basic Microbiol. 2012, 53, 303–317. [Google Scholar] [CrossRef]
- Roskova, Z.; Skarohlid, R.; McGachy, L. Siderophores: An alternative bioremediation strategy? Sci. Total Environ. 2022, 819, 153144. [Google Scholar] [CrossRef] [PubMed]
- Kalam, S.; Basu, A.; Podile, A.R. Functional and molecular characterization of plant growth promoting Bacillus isolates from tomato rhizosphere. Hellyon 2020, 6, e04734. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Park, D.H.; Kim, J.Y.; Kim, B.S. Biological control of grey mold and growth promotion of tomato using Bacillus spp. isolated from soil. Trop. Plant Pathol. 2016, 41, 169–176. [Google Scholar] [CrossRef]
- Dong, H.; Gao, R.; Dong, Y.; Yao, Q.; Zhu, H. Bacillus velezensis RC116 inhibits the pathogens of bacterial wilt and fusarium wilt in tomato with multiple biocontrol traits. Int. J. Mol. Sci. 2023, 24, 8527. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Chauhan, A.; Sharma, R.; Sehgal, R.; Shirkot, C.K. Potential of Bacillus amyloliquefaciens for biocontrol of bacterial canker of tomato incited by Clavibacter michiganensis ssp. michiganensis. Microb. Pathog. 2019, 130, 196–203. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Gámez-Arcas, S.; Baroja-Fernández, E.; García-Gómez, P.; Muñoz, F.J.; Almagro, G.; Bahaji, A.; Sánchez-López, Á.M.; Pozueta-Romero, J. Action mechanisms of small microbial volatile compounds in plants. J. Exp. Bot. 2022, 73, 498–510. [Google Scholar] [CrossRef]
- Peñuelas, J.; Asensio, D.; Tholl, D.; Wenke, K.; Rosenkranz, M.; Piechulla, B.; Schnitzler, J.P. Biogenic volatile emissions from the soil. Plant Cell Environ. 2014, 37, 1866–1891. [Google Scholar] [CrossRef]
- Ramyabharathi, S.A.; Raguchander, T. Mode of action of Bacillus subtilis EPCO16 against tomato Fusarium wilt. Biochem. Cell. Arch. 2014, 14, 47–50. [Google Scholar]
- Sachdev, S.; Singh, R.P. Isolation, characterisation and screening of native microbial isolates for biocontrol of fungal pathogens of tomato. Clim. Chang. Environ. Sustain. 2018, 6, 46–58. [Google Scholar] [CrossRef]
- Wang, S.; Hu, T.; Jiao, Y.; Wei, J.; Cao, K. Isolation and characterization of Bacillus subtilis EB-28, an endophytic bacterium strain displaying biocontrol activity against Botrytis cinerea Pers. Front. Agric. China 2009, 3, 247–252. [Google Scholar] [CrossRef]
- Raza, W.; Wang, J.; Wu, Y.; Ling, N.; Wei, Z.; Huang, Q.; Shen, Q. Effects of volatile organic compounds produced by Bacillus amyloliquefaciens on the growth and virulence traits of tomato bacterial wilt pathogen Ralstonia solanacearum. Appl. Microbiol. Biotechnol. 2016, 100, 7639–7650. [Google Scholar] [CrossRef] [PubMed]
- Awan, Z.A.; Shoaib, A.; Schenk, P.M.; Ahmad, A.; Alansi, S.; Paray, B.A. Antifungal potential of volatiles produced by Bacillus subtilis BS-01 against Alternaria solani in Solanum lycopersicum. Front. Plant Sci. 2023, 13, 1089562. [Google Scholar] [CrossRef]
- Guo, J.; Xu, Y.; Liang, S.; Zhou, Z.; Zhang, C.; Li, K.; Peng, X.; Qin, S.; Xing, K. Antifungal activity of volatile compounds from Bacillus tequilensis XK29 against Botrytis cinerea causing gray mold on cherry tomatoes. Postharvest Biol. Technol. 2023, 198, 112239. [Google Scholar] [CrossRef]
- Lee, S.W.; Lee, S.H.; Balaraju, K.; Park, K.S.; Nam, K.W.; Park, J.W.; Park, K. Growth promotion and induced disease suppression of four vegetable crops by a selected plant growth-promoting rhizobacteria (PGPR) strain Bacillus subtilis 21-1 under two different soil conditions. Acta Physiol. Plant. 2014, 36, 1353–1362. [Google Scholar] [CrossRef]
- Chowdappa, P.; Kumar, S.P.M.; Lakshmi, M.J.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Farzand, A.; Moosa, A.; Zubair, M.; Khan, A.R.; Massawe, V.C.; Tahir, H.A.S.; Sheikh, T.M.M.; Ayaz, M.; Gao, X. Suppression of Sclerotinia sclerotiorum by the induction of systemic resistance and regulation of antioxidant pathways in tomato using fengycin produced by Bacillus amyloliquefaciens FZB42. Biomolecules 2019, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Song, C.; Muñoz, C.; Kuipers, O. Bacillus cabrialesii BH5 protects tomato plants against Botrytis cinerea by production of specific antifungal compounds. Front. Microbiol. 2021, 12, 707609. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Xu, W.; Hu, Y.; Tian, R.; Wang, Z. Bacillus velezensis YYC promotes tomato growth and induces resistance against bacterial wilt. Biol. Control 2022, 172, 104977. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Belachew, S.T.; Yoon, E.; Chun, S.C. Expression of β-1,3-glucanase (GLU) and phenylalanine ammonia-lyase (PAL) genes and their enzymes in tomato plants induced after treatment with Bacillus subtilis CBR05 against Xanthomonas campestris pv. vesicatoria. J. Gen. Plant Pathol. 2017, 83, 7–13. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Abdalla, S.A.; Sleem, M.M. Endophytic Bacillus subtilis SR22 triggers defense responses in tomato against rhizoctonia root rot. Plants 2022, 11, 2051. [Google Scholar] [CrossRef]
- Van Loon, L.C. Plant responses to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Liebmann, B.; Kalantidis, K.; Vassilakos, N.; Skandalis, N. Bacillus amyloliquefaciens MBI600 differentially induces tomato defense signaling pathways depending on plant part and dose of application. Sci. Rep. 2019, 9, 19120. [Google Scholar] [CrossRef]
- Appu, M.; Ramalingam, P.; Sathiyanarayanan, A.; Huang, J. An overview of plant defense-related enzymes responses to biotic stresses. Plant Gene 2021, 27, 100302. [Google Scholar] [CrossRef]
- Ongena, M.; Jourdan, E.; Adam, A.; Michel, P.; Brans, A.; Joris, B.; Arpigny, J.L.; Thonart, P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis mbi600 promotes growth of tomato plants and induces systemic resistance contributing to the control of soilborne pathogens. Plants 2021, 10, 1113. [Google Scholar] [CrossRef]
- Awan, Z.A.; Shaaib, A. Combating early blight infection by employing Bacillus subtilis in combination with plant fertilizers. Curr. Plant Biol. 2019, 20, 100125. [Google Scholar] [CrossRef]
- Kipngeno, P.; Losenge, T.; Maina, N.; Kahangi, E.; Juma, P. Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biol. Control 2015, 90, 92–95. [Google Scholar] [CrossRef]
- Wu, K.; Fang, Z.; Wang, L.; Yuan, S.; Guo, R.; Shen, B.; Shen, Q. Biological potential of bioorganic fertilizer fortified with bacterial antagonist for the control of tomato bacterial wilt and the promotion of crop yields. J. Microbiol. Biotechnol. 2016, 26, 1755–1764. [Google Scholar] [CrossRef]
- Bahramisharif, A.; Rose, L.E. Efficacy of biological agents and compost on growth and resistance of tomatoes to late blight. Planta 2019, 249, 790–813. [Google Scholar] [CrossRef]
- Ji, X.; Li, J.; Meng, Z.; Zhang, S.; Dong, B.; Qiao, K. Synergistic effect of combined application of a new fungicide fluopimomide with a biocontrol agent Bacillus methylotrophicus TA-1 for management of gray mold in tomato. Plant Dis. 2019, 103, 1991–1997. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Luo, K.; Jiang, H.; Deng, Y.; Bai, L.; Zhou, X. Combined use of Bacillus subtilis strain B-001 and bactericide for the control of tomato bacterial wilt. Pest Manag. Sci. 2017, 73, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Myresiotis, C.K.; Karaoglanidis, G.S.; Vryzas, Z.; Papadopoulou-Mourkidou, E. Evaluation of plant-growth-promoting rhizobacteria, acibenzolar-S-methyl and hymexazol for integrated control of Fusarium crown and root rot on tomato. Pest Manag. Sci. 2012, 68, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Esquivel-Cervantes, L.F.; Tlapal-Bolaños, B.; Tovar-Pedraza, J.M.; Pérez-Hernández, O.; Leyva-Mir, S.G.; Camacho-Tapia, M. Efficacy of biorational products for managing diseases of tomato in greenhouse production. Plants 2022, 11, 1638. [Google Scholar] [CrossRef] [PubMed]
- Mousa, M.A.A.; Abo-Elyousr, K.A.M.; Abdel Alal, A.M.K.; Alshareef, N.O. Management Fusarium wilt disease in tomato by combinations of Bacillus amyloliquefaciens and peppermint oil. Agronomy 2021, 11, 2536. [Google Scholar] [CrossRef]
- Abdeljalil, N.O.B.; Vallance, J.; Gerbore, J.; Yacoub, A.; Daami-Remadi, M.; Rey, P. Combining potential oomycete and bacterial biocontrol agents as a tool to fight tomato Rhizoctonia root rot. Biol. Control 2021, 155, 104521. [Google Scholar] [CrossRef]
- Zhao, X.; Hendriks, M.; Deleu, E.; Spanoghe, P.; Höfte, M.; van Overbeek, L.; Uyttendaele, M. Prevalence, attachment ability and strength of the biological control agent Bacillus thuringiensis on tomato. Food Microbiol. 2023, 112, 104235. [Google Scholar] [CrossRef]
- Chien, Y.C.; Huang, C.H. Biocontrol of bacterial spot on tomato by foliar spray and growth medium application of Bacillus amyloliquefaciens and Trichoderma asperellum. Eur. J. Plant Pathol. 2020, 156, 995–1003. [Google Scholar] [CrossRef]
- Kulimushi, S.M.; Muiru, W.M.; Mutitu, E.W. Potential of Trichoderma spp., Bacillus subtilis and Pseudomonas fluorescens in the management of early blight in tomato. Biocontrol Sci. Technol. 2021, 31, 912–923. [Google Scholar] [CrossRef]
- Devi, N.O.; Tombisana Devi, R.K.; Debbarma, M.; Hajong, M.; Thokchom, S. Effect of endophytic Bacillus and arbuscular mycorrhiza fungi (AMF) against Fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. Egypt. J. Biol. Pest Control 2022, 32, 1. [Google Scholar] [CrossRef]
- Sultana, F.; Hossain, M.M. Assessing the potentials of bacterial antagonists for plant growth promotion, nutrient acquisition, and biological control of Southern blight disease in tomato. PLoS ONE 2022, 17, e0267253. [Google Scholar] [CrossRef]
- Elsayed, T.R.; Jacquiod, S.; Nour, E.H.; Sørensen, S.J.; Smalla, K. Biocontrol of bacterial wilt disease through complex interaction between tomato plant, antagonists, the indigenous rhizosphere microbiota, and Ralstonia solanacearum. Front. Microbiol. 2020, 10, 2835. [Google Scholar] [CrossRef]
- Solanki, M.K.; Solanki, A.C.; Rai, S.; Srivastava, S.; Kashyap, B.K.; Divvela, P.K.; Kumar, S.; Yandigeri, M.S.; Kashyap, P.L.; Shrivastava, A.K.; et al. Functional interplay between antagonistic bacteria and Rhizoctonia solani in the tomato plant rhizosphere. Front. Microbiol. 2022, 13, 990850. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.E. Efficiency of Trichoderma viride and Bacillus subtilis as biocontrol agents against root rot caused by Fusarium solani in tomato. Egypt. J. Agric. Res. 2019, 97, 507–516. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karačić, V.; Miljaković, D.; Marinković, J.; Ignjatov, M.; Milošević, D.; Tamindžić, G.; Ivanović, M. Bacillus Species: Excellent Biocontrol Agents against Tomato Diseases. Microorganisms 2024, 12, 457. https://doi.org/10.3390/microorganisms12030457
Karačić V, Miljaković D, Marinković J, Ignjatov M, Milošević D, Tamindžić G, Ivanović M. Bacillus Species: Excellent Biocontrol Agents against Tomato Diseases. Microorganisms. 2024; 12(3):457. https://doi.org/10.3390/microorganisms12030457
Chicago/Turabian StyleKaračić, Vasiljka, Dragana Miljaković, Jelena Marinković, Maja Ignjatov, Dragana Milošević, Gordana Tamindžić, and Milan Ivanović. 2024. "Bacillus Species: Excellent Biocontrol Agents against Tomato Diseases" Microorganisms 12, no. 3: 457. https://doi.org/10.3390/microorganisms12030457