New Taxa of the Family Amniculicolaceae (Pleosporales, Dothideomycetes, Ascomycota) from Freshwater Habitats in Spain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Specimen Examination
2.2. Phenotypic Study
2.3. DNA Extraction PCR Amplification and Sequencing
2.4. Phylogenetic Analysis
3. Results
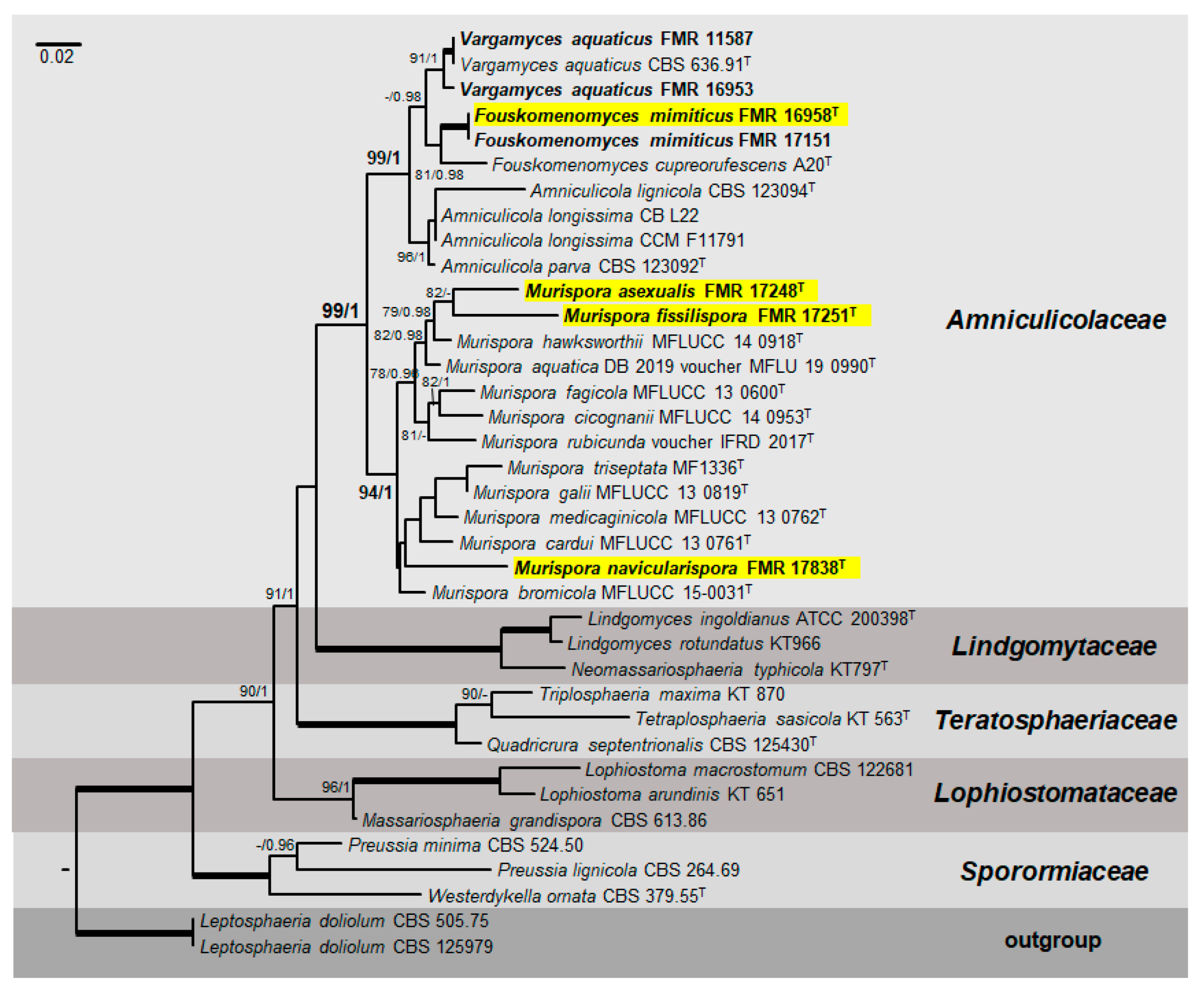
3.1. Phylogenetic Analyses
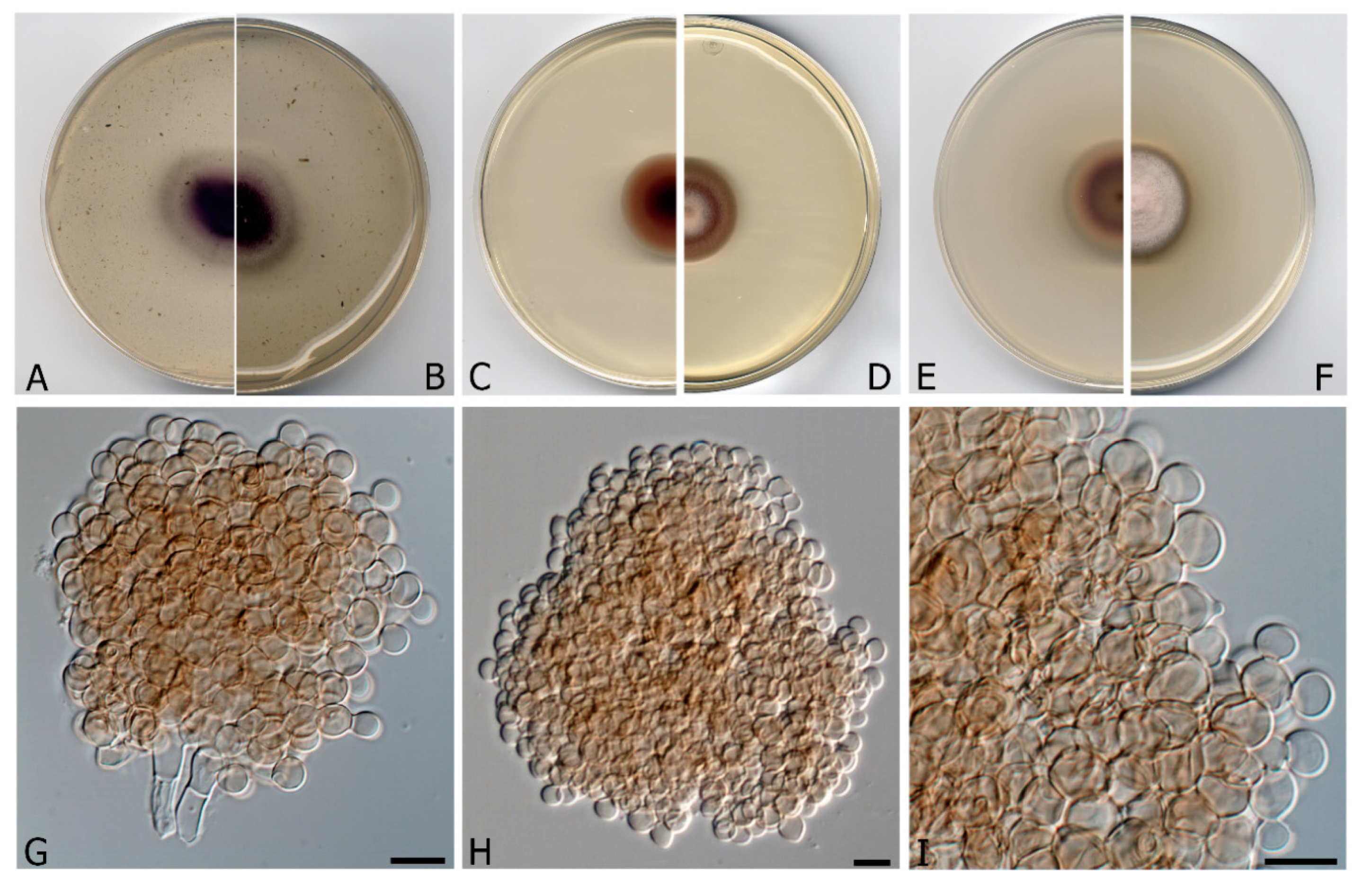
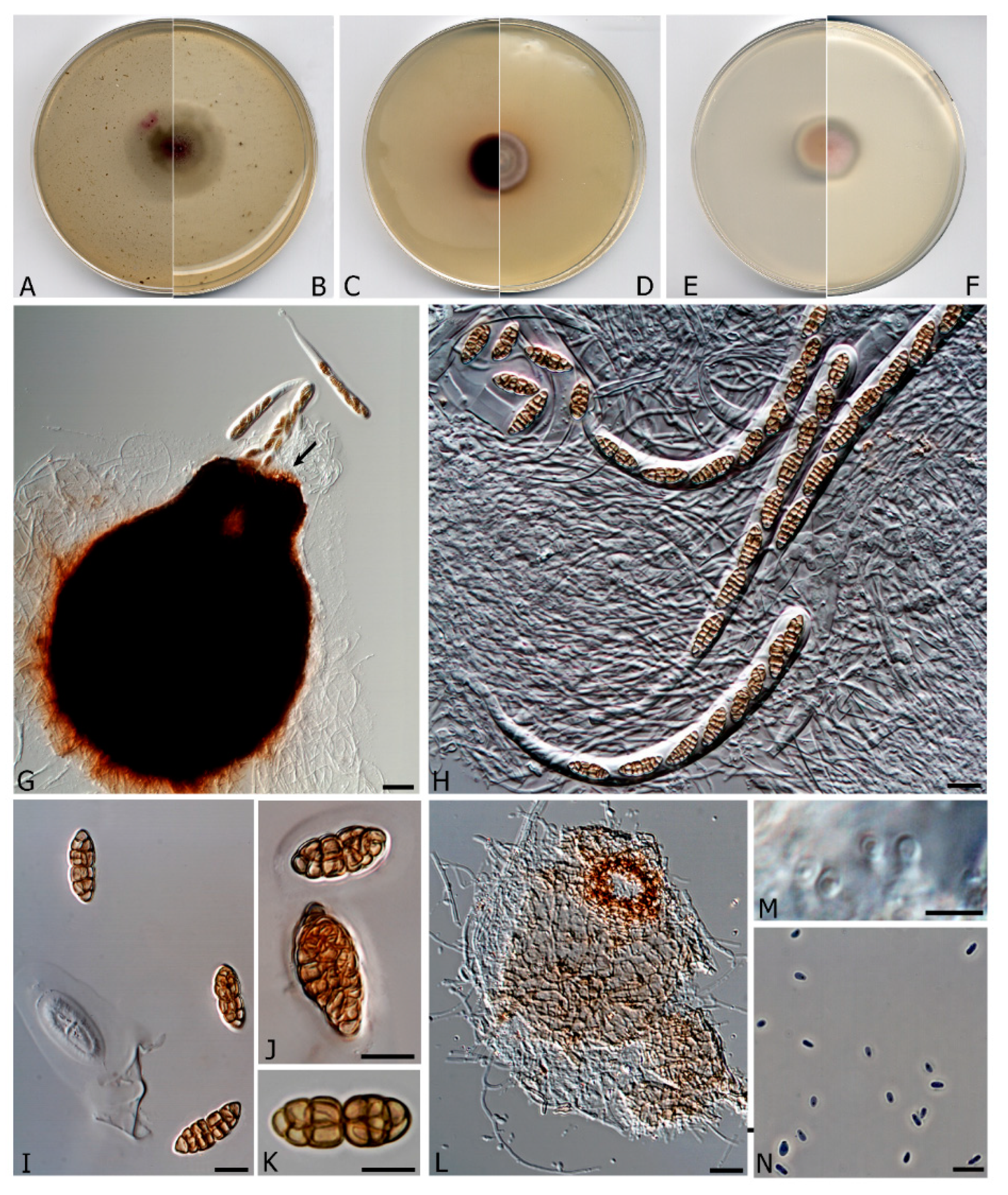
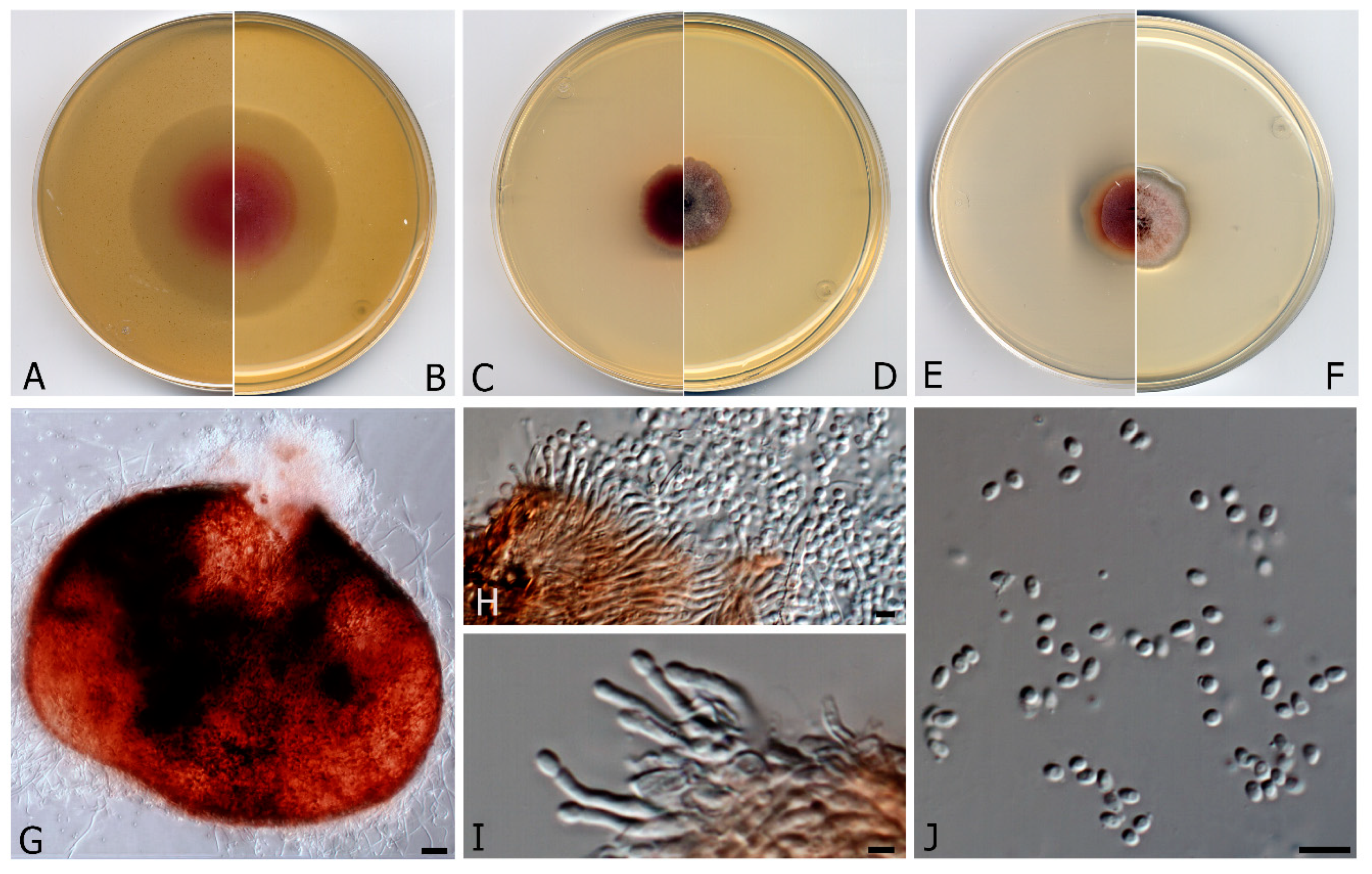
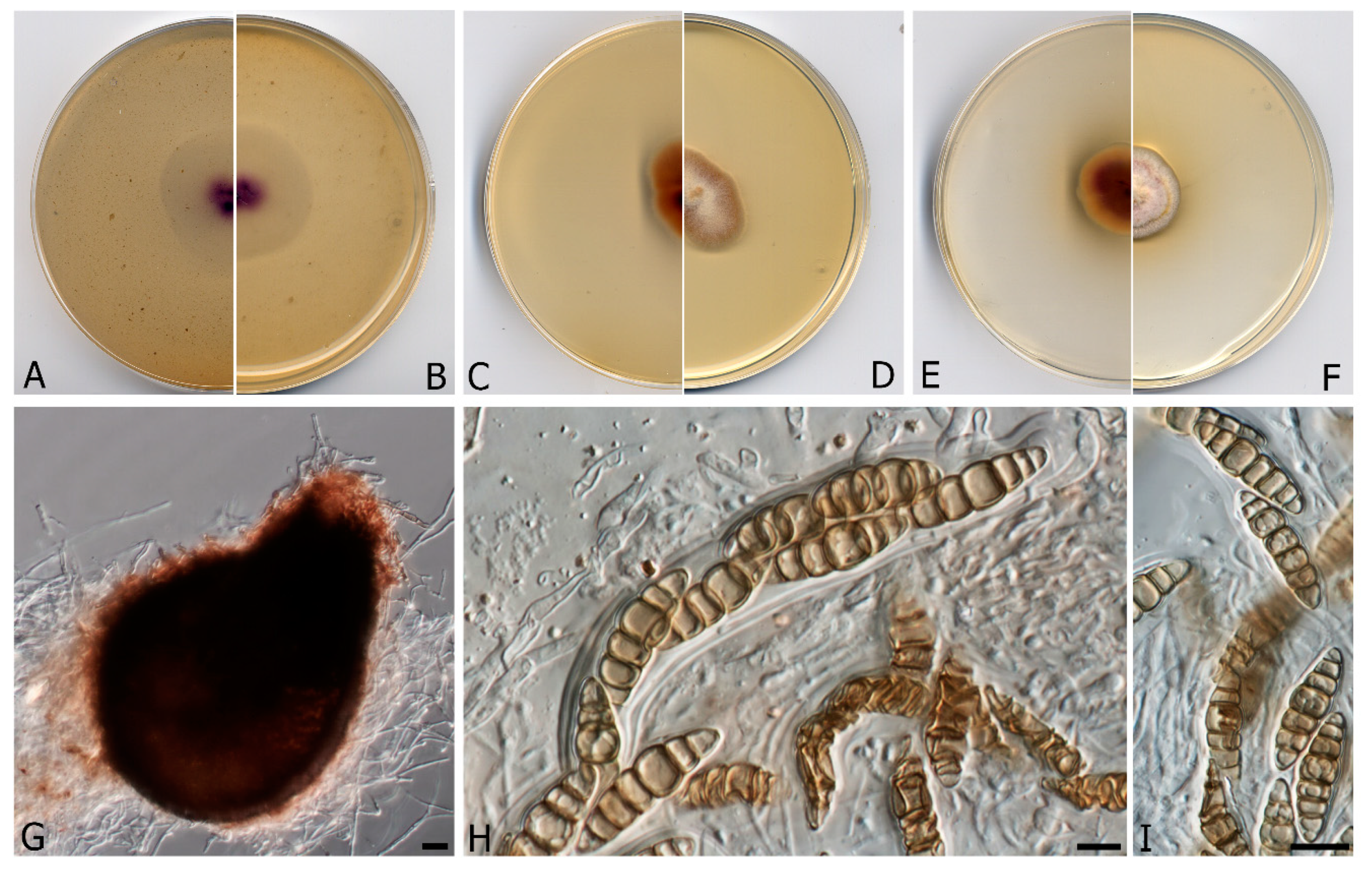
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grossart, H.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 17, 339–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic Habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Gessner, M.O.; Gulis, V.; Kuehn, K.A.; Chauvet, E.; Suberkropp, K. Fungal decomposers of plant litter in aquatic ecosystems. In Environmental and Microbial Relationships. The Mycota, 2nd ed.; Kubicek, C.P., Druzhinina, I.S., Eds.; Springer: Heidelberg/Berlin, Germany, 2007; Volume 4, pp. 301–324. [Google Scholar]
- Webster, J. Experiment with spores of aquatic hyphomycetes: I Sedimentation, and impaction on smooth surfaces. Ann. Bot. 1959, 23, 595–611. [Google Scholar] [CrossRef]
- Wurzbacher, C.M.; Bärlocher, F.; Grossart, H.P. Fungi in lake ecosistems. Aquat. Microb. Ecol. 2010, 59, 125–149. [Google Scholar] [CrossRef] [Green Version]
- Dhanasekaran, V.; Jeewon, R.; Hyde, K.D. Molecular Taxonomy, origins and evolution of freshwater ascomycetes. Fungal. Divers. 2006, 23, 351–390. [Google Scholar]
- Gareth Jones, E.B. Form and function of fungal spore appendages. Mycoscience 2006, 47, 167–183. [Google Scholar] [CrossRef]
- Shearer, C.A.; Raja, H.A.; Miller, A.N.; Nelson, P.; Tanaka, K.; Hirayama, K.; Marvanová, L.; Hyde, K.D.; Zhang, Y. The molecular phylogeny of freshwater Dothideomycetes. Stud. Mycol. 2009, 64, 145–153. [Google Scholar] [CrossRef]
- Pang, K.L.; Abdel-Wahab, M.A.; Sivichai, S.; El-Sharouney, H.M.; Gareth Jones, E.B. Jahnulales (Dothideomycetes, Ascomycota): A new order of lignicolous freshwater ascomycetes. Mycol. Res. 2002, 106, 31–42. [Google Scholar] [CrossRef]
- Raja, H.A.; Tanaka, K.; Hirayama, K.; Miller, A.N.; Shearer, C.A. Freshwater ascomycetes: Two new species of Lindgomyces (Lindgomycetaceae, Pleosporales, Dothideomycetes) from Japan and USA. Mycologia 2011, 103, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; Paguigan, N.D.; Fournier, J.; Oberlies, N.H. Additions to Lindgomyces (Lindgomycetaceae, Pleosporales, Dothideomycetes), Including two new species occurring on submerged wood from North Carolina, USA, with notes on secondary metabolite profiles. Mycol. Prog. 2017, 16, 535–552. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Shearer, C.A. Freshwater ascomycetes: Natipusillaceae, a new family of tropical fungi, including Natipusilla bellaspora sp. nov. from the peruvian Amazon. Mycologia. 2012, 104, 569–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jeewon, R.; Fournier, J.; Hyde, K.D. Multi-Gene phylogeny and morphotaxonomy of Amniculicola lignicola: A Novel Freshwater Fungus from France and Its Relationships to the Pleosporales. Mycol. Res. 2008, 112, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fournier, J.; Crous, P.W.; Pointing, S.B.; Hyde, K.D. Phylogenetic and morphological assessment of two new species of Amniculicola and their allies (Pleosporales). Pers. Mol. Phylogeny Evol. Fungi 2009, 23, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Schoch, C.L.; Fournier, J.; Crous, P.W.; de Gruyter, J.; Woudenberg, J.H.C.; Hirayama, K.; Tanaka, K.; Pointing, S.B.; Spatafora, J.W.; et al. Multi-locus phylogeny of Pleosporales: A taxonomic, ecological and evolutionary Re-evaluation. Stud. Mycol. 2009, 64, 85–102. [Google Scholar] [CrossRef]
- Wanasinghe; Dhanushka, N.; Gareth Jones, E.B.; Camporesi, E.; Mortimer, P.E.; Xu, J.; Bahkali, A.H.; Hyde, K.D. The Genus Murispora. Cryptogam. Mycol. 2015, 36, 419–448. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Luecking, R.; et al. Fungal Diversity Notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Bao, D.; Wanasinghe, D.; Luo, Z.; Mortimer, P.E.; Kumar, V.; Su, H.; Hyde, K.D. Murispora aquatica sp. nov. and Murispora fagicola, a new record from freshwater habitat in China. Pytotaxa 2019, 416, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; CBS Laboratory Manual Series; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010; p. 390. [Google Scholar]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth & Bisby’s Dictionary of the Fungi, 8th ed.; CAB International: Oxon, UK, 1995; p. 616. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; p. 256. [Google Scholar]
- Chupp, C. Further notes on double cover-glass mounts. Mycologia 1940, 32, 269–270. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and Phylogeny of Gliocladium Analysed from Nuclear Large Subunit Ribosomal DNA Sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S. Primers for Elongation Factor 1-α (EF1-α). 2001. Available online: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf. (accessed on 2 February 2020).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0. for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic. Acids. Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic. Acids. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling High-Impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. JModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Voglmayr, H. Spirosphaera cupreorufescens sp. nov., a rare aeroaquatic fungus. Stud. Mycol. 2004, 50, 221–228. [Google Scholar]
- Gareth Jones, E.B.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.N.; Hyde, K.D.; Liu, J.K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of New Marine Dothideomycetes and Sordariomycetes from Mangroves and Deep-Sea Sediments. Bot. Mar. 2020, 63, 155–181. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.K.; Aptroot, A.; Crous, P.W.; Hyde, K.D.; Jeewon, R. The Polyphyletic Nature of Pleosporales: An Example from Massariosphaeria Based on RDNA and RBP2 Gene Phylogenies. Mycol. Res. 2007, 111, 1268–1276. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Gené, J.; Castañeda-Ruiz, R.F.; Mena-Portales, J.; Crous, P.W.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain | GenBank Accession No. | ||
|---|---|---|---|---|
| LSU | ITS | tef1 | ||
| Amniculicola lignicola | CBS 123094T | MH874798 | ----- | GU456278 |
| Amniculicola longissima | CB L22 | GU266240 | AY204596 | ----- |
| Amniculicola lonsissima | CCM-F10304 | JN673029 | AY204594 | ----- |
| Amniculicola parva | CBS 123092T | FJ795497 | MH863272 | GU349065 |
| Fouskomenomyces cupreorufescens | A20T | AY616236 | AY616232 | ----- |
| Fouskomenomyces mimiticus | FMR 16,958T | LR824585 | LR824586 | LR824584 |
| Fouskomenomyces mimiticus | FMR 17,151 = CBS 146935 | LR824588 | LR824587 | LR824589 |
| Leptosphaeria dolium | CBS 125979 | JF740283 | JF740208 | ----- |
| Leptosphaeria dolium | CBS 505.75 | GQ387576 | JF740205 | GU349069 |
| Lindgomyces ingoldianus | ATCC 200398T | AB521736 | JF419898 | ----- |
| Lindgomyces rotundatus | KT966 | AB521739 | JF419901 | ----- |
| Lophiostoma macrostomum | CBS 122681 | EU552141 | EU552141 | LC001753 |
| Lophiostoma arundinis | KT 651 | AB618999 | JN942965 | LC001738 |
| Massariosphaeria grandispora | CBS 613.86 | FJ795507 | ----- | GU349036 |
| Murispora aquatica | MFLU 19-0990T | MN325075 | MN325085 | MN337969 |
| Murispora asexualis | FMR 17,248T = CBS 146937 | LR824596 | LR824593 | LR824590 |
| Murispora bromicola | MFLUCC 15-0031T | NG_059595 | NR_164235 | KT305999 |
| Murispora cardui | MFLUCC 13-0761T | NG_059607 | KT736082 | KT709190 |
| Murispora cicognanii | MFLUCC 14-0953T | NG_059609 | NR_155381 | MK109804 |
| Murispora fagicola | MFLUCC 13-0600T | NG_060797 | NR_155379 | KT709188 |
| Murispora fissilispora | FMR 17,251T = CBS 146936 | LR824597 | LR824594 | LR824591 |
| Murispora galii | MFLUCC 13-0819T | KT709175 | NR_154629 | KT709189 |
| Murispora haswksworthii | MFLUCC 14-091T | KT709180 | NR_138414 | KT709192 |
| Murispora medicaginicola | MFLUCC 13-0762T | NG_059609 | NR_155380 | KT709191 |
| Murispora navicularispora | FMR 17,838T | LR824598 | LR824595 | LR824592 |
| Murispora rubicunda | IFRD 2017T | FJ795507 | ----- | GU456289 |
| Murispora triseptata | MF1336T | MK411002 | ----- | ----- |
| Neomassariosphaeria typhicola | KT797T | AB521747 | JF419906 | ----- |
| Preussia lignícola | CBS 264.69 | MH878448 | ----- | GU349027 |
| Preussia minima | CBS 524.50 | MH868263 | MH856741 | DQ677897 |
| Quadricrura septentrionalis | CBS 125430 | MH875152 | NR_119402 | ----- |
| Triplosphaeria máxima | KT 870 | AB524637 | NR_119407 | ----- |
| Tetraplosphaeria sasicola | KT 563T | AB524631 | AB524807 | ----- |
| Vargamyces aquaticus | CBS 636.9T | KY853539 | NR_154471 | ----- |
| Vargamyces aquaticus | FMR 11587 | KY853538 | KY853475 | ----- |
| Vargamyces aquaticus | FMR 16,953 | LR812096 | LR812095 | ----- |
| Westerdykella ornata | CBS 379.55T | NG_057861 | NR_103587 | GU349021 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magaña-Dueñas, V.; Stchigel, A.M.; Cano-Lira, J.F. New Taxa of the Family Amniculicolaceae (Pleosporales, Dothideomycetes, Ascomycota) from Freshwater Habitats in Spain. Microorganisms 2020, 8, 1355. https://doi.org/10.3390/microorganisms8091355
Magaña-Dueñas V, Stchigel AM, Cano-Lira JF. New Taxa of the Family Amniculicolaceae (Pleosporales, Dothideomycetes, Ascomycota) from Freshwater Habitats in Spain. Microorganisms. 2020; 8(9):1355. https://doi.org/10.3390/microorganisms8091355
Chicago/Turabian StyleMagaña-Dueñas, Viridiana, Alberto M. Stchigel, and José F. Cano-Lira. 2020. "New Taxa of the Family Amniculicolaceae (Pleosporales, Dothideomycetes, Ascomycota) from Freshwater Habitats in Spain" Microorganisms 8, no. 9: 1355. https://doi.org/10.3390/microorganisms8091355