
Bioactive Secondary Metabolites of the Genus Diaporthe and Anamorph Phomopsis from Terrestrial and Marine Habitats and Endophytes: 2010–2019


Abstract
:1. Introduction
2. Bioactive Secondary Metabolites from Phomopsis
2.1. Polyketides
2.1.1. Xanthones
2.1.2. Chromones
2.1.3. Chromanones
2.1.4. Benzofuranones
2.1.5. Pyrones
2.1.6. Quinones
2.1.7. Phenols
2.1.8. Oblongolides
2.1.9. Unclassified Polyketides
2.2. Terpenoids
2.2.1. Monoterpenoids
2.2.2. Sesquiterpenoids
2.2.3. Diterpenoids
2.2.4. Triterpenoids
2.3. Steroids
2.4. Macrolides
2.5. Alkaloids
2.6. Flavonoids
3. Bioactive Secondary Metabolites from Diaporthe spp.
3.1. Polyketides
3.1.1. Xanthones
3.1.2. Chromones
3.1.3. Chromanones
3.1.4. Furanones
3.1.5. Pyrones
3.1.6. Quinones
3.1.7. Phenols
3.1.8. Oblongolides
3.1.9. Unclassified Polyketides
3.2. Terpenoids
3.3. Steriods
3.4. Ten-Membered Lactones
3.5. Alkaloids
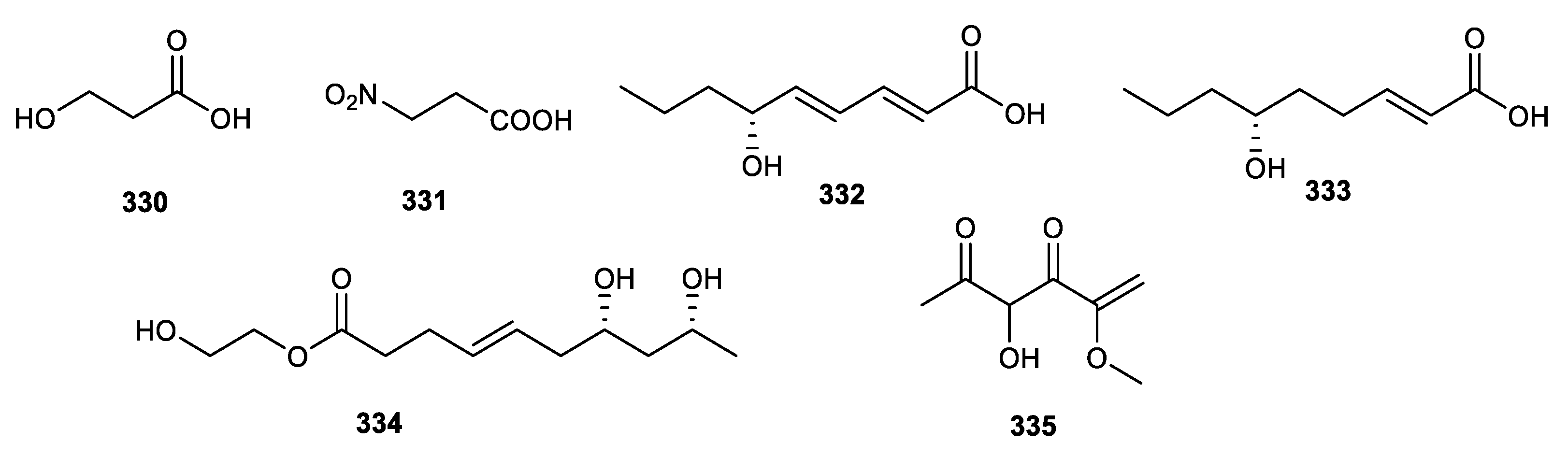
3.6. Fatty Acids
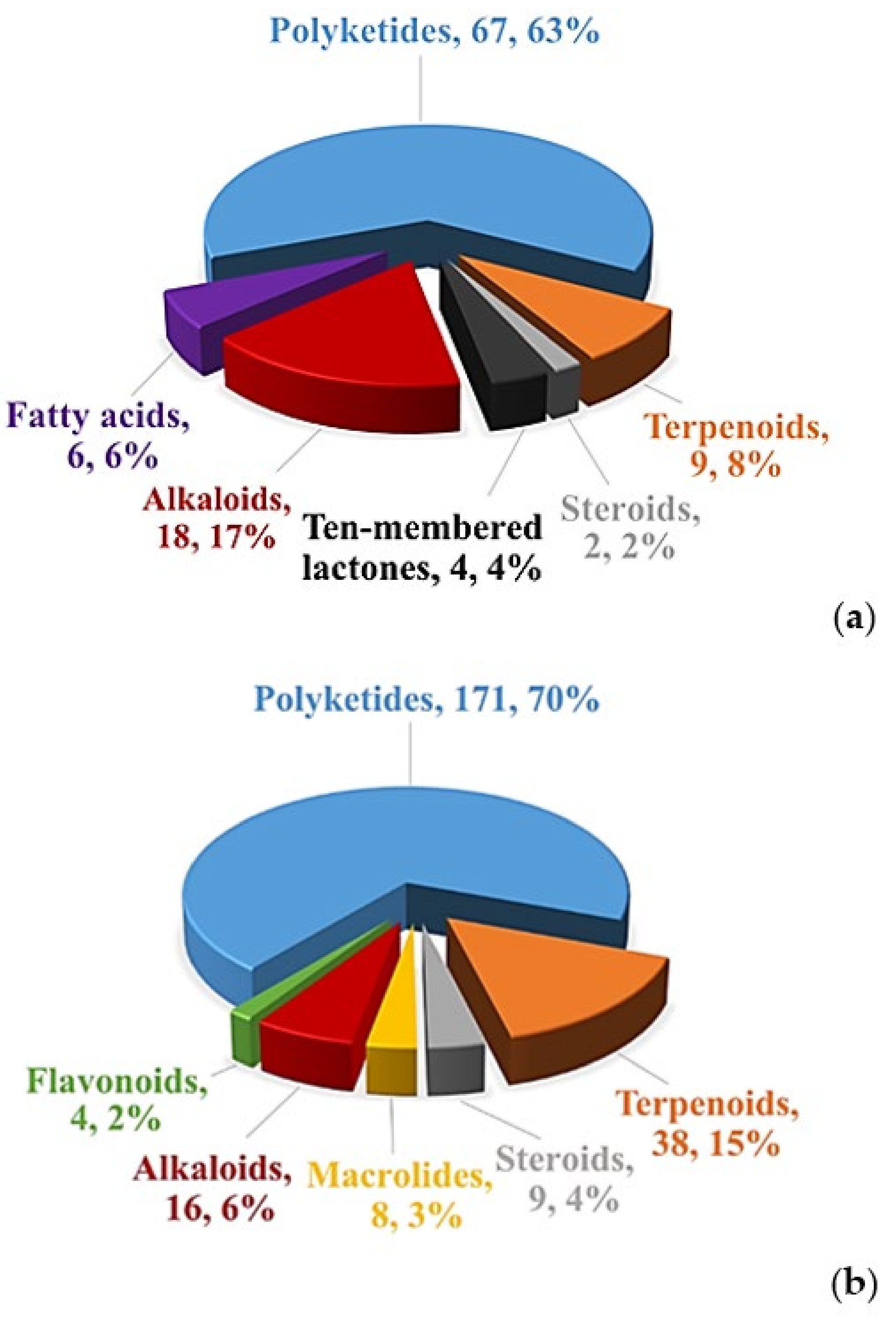
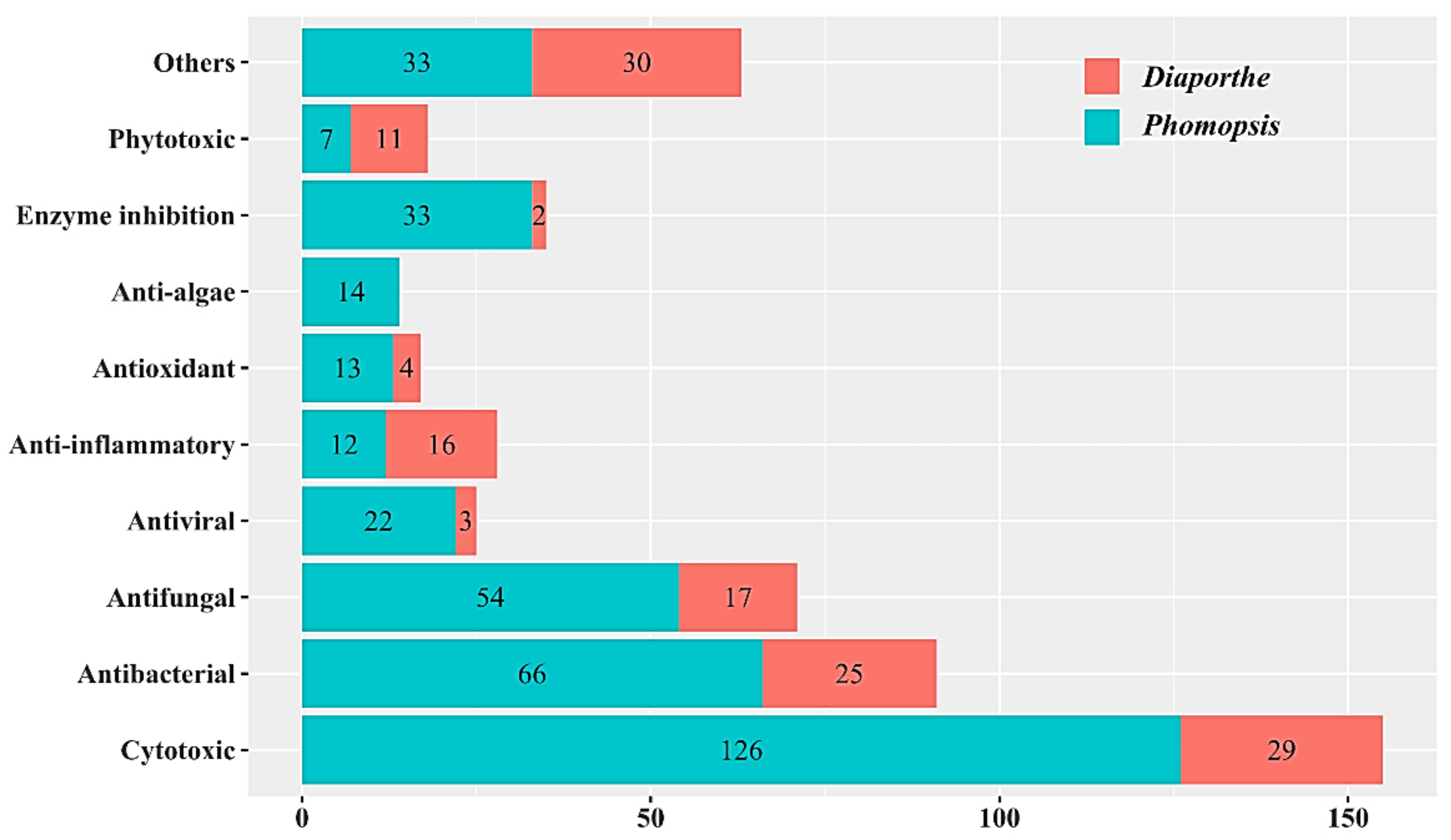
4. Characteristics of Bioactive Secondary Metabolites from the Genus Diaporthe and Anamorph Phomopsis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Chen, Y.Y.; Liu, J.K.J. Unravelling Diaporthe species associated with woody hosts from karst formations (Guizhou) in China. J. Fungi 2020, 6, 251. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, F.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 153–187. [Google Scholar] [CrossRef]
- Santos, J.M.; Correia, V.G.; Phillips, A.J.L. Primers for mating-type diagnosis in Diaporthe and Phomopsis: Their use in teleomorph induction in vitro and biological species definition. Fungal Biol. 2010, 114, 255–270. [Google Scholar] [CrossRef]
- Gong, J.L.; Lu, Y.; Wu, W.H.; He, C.P.; Liang, Y.Q.; Huang, X.; Zheng, J.L.; Xi, J.G.; Tang, S.B.; Yi, K.X. First report of Phomopsis heveicola (anamorph of Diaporthe tulliensis) causing leaf blight of Coffee (Coffea arabica) in China. Plant Dis. 2020, 104, 570–571. [Google Scholar] [CrossRef]
- Rehner, S.A.; Uecker, F.A. Nuclear ribosomal internal transcribed spacer phylogeny and host diversity in the coelomycete Phomopsis. Can. J. Bot. 1994, 72, 1666–1674. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandecic, K.; Cosic, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Hou, C.L. Three new species of Diaporthe from China based on morphological characters and DNA sequence data analyses. Phytotaxa 2019, 422, 157–174. [Google Scholar] [CrossRef]
- Leon, M.; Berbegal, M.; Rodriguez-Reina, J.M.; Elena, G.; Abad-Campos, P.; Ramon-Albalat, A.; Olmo, D.; Vicent, A.; Luque, J.; Miarnau, X.; et al. Identification and characterization of Diaporthe spp. associated with twig cankers and shoot blight of almonds in Spain. Agronomy 2020, 10, 1062. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D.; et al. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Santos, L.; Alves, A.; Alves, R. Evaluating multi-locus phylogenies for species boundaries determination in the genus Diaporthe. PeerJ 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Huang, Y.J.; Fang, M.J.; Wang, J.F.; Zheng, Z.H.; Su, W.J. Cytotoxic and antimicrobial metabolites from marine lignicolous fungi, Diaporthe sp. FEMS Microbiol. Lett. 2005, 251, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.H.; Teles, H.L.; Zanardi, L.M.; Marx Young, M.C.; Eberlin, M.N.; Hadad, R.; Pfenning, L.H.; Costa-Neto, C.M.; Castro-Gamboa, I.; Bolzani, V.d.S.; et al. Cadinane sesquiterpenoids of Phomopsis cassiae, an endophytic fungus associated with Cassia spectabilis (Leguminosae). Phytochemistry 2006, 67, 1964–1969. [Google Scholar] [CrossRef]
- Niaz, S.I.; Khan, D.; Naz, R.; Safdar, K.; Ul Abidin, S.Z.; Khan, I.U.; Gul, R.; Khan, W.U.; Khan, M.A.U.; Lan, L. Antimicrobial and antioxidant chlorinated azaphilones from mangrove Diaporthe perseae sp. isolated from the stem of Chinese mangrove Pongamia pinnata. J. Asian Nat. Prod. Res. 2020. [Google Scholar] [CrossRef]
- Yang, Z.J.; Zhang, Y.F.; Wu, K.; Xu, Y.X.; Meng, X.G.; Jiang, Z.T.; Ge, M.; Shao, L. New azaphilones, phomopsones A-C with biological activities from an endophytic fungus Phomopsis sp. CGMCC No.5416. Fitoterapia 2020, 145. [Google Scholar] [CrossRef]
- Da Rosa, B.V.; Kuhn, K.R.; Ugalde, G.A.; Zabot, G.L.; Kuhn, R.C. Antioxidant compounds extracted from Diaporthe schini using supercritical CO2 plus cosolvent. Bioprocess Biosyst. Eng. 2020, 43, 133–141. [Google Scholar] [CrossRef]
- Fan, M.M.; Xiang, G.; Chen, J.W.; Gao, J.; Xue, W.W.; Wang, Y.X.; Li, W.H.; Zhou, L.; Jiao, R.H.; Shen, Y.; et al. Libertellenone M, a diterpene derived from an endophytic fungus Phomopsis sp. S12, protects against DSS-induced colitis via inhibiting both nuclear translocation of NF-κB and NLRP3 inflammasome activation. Int. Immunopharmacol. 2020, 80. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Ogilvie, K.K.; Watson, A.K. Phytotoxic metabolites of Phomopsis convolvulus, a host-specific pathogen of field bindweed. Can. J. Chem. 1992, 70, 2276–2284. [Google Scholar] [CrossRef]
- Zhang, C.W.; Ondeyka, J.G.; Herath, K.B.; Guan, Z.Q.; Collado, J.; Platas, G.; Pelaez, F.; Leavitt, P.S.; Gurnett, A.; Nare, B.; et al. Tenellones A and B from a Diaporthe sp.: Two highly substituted benzophenone inhibitors of parasite cGMP-dependent protein kinase activity. J. Nat. Prod. 2005, 68, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Gao, Y.H.; Niu, D.Y.; Yang, L.Y.; Gao, X.M.; Du, G.; Hu, Q.F. Xanthone derivatives from the fermentation products of an endophytic fungus Phomopsis sp. Fitoterapia 2013, 91, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Qiu, S.X.; She, Z.G.; Lin, Y.C. A new xanthone derivative from the marine fungus Phomopsis sp. (No. SK7RN3G1). Chem. Nat. Compd. 2013, 49, 31–33. [Google Scholar] [CrossRef]
- Yuan, L.; Huang, W.; Du, G.; Gao, X.; Yang, H.; Hu, Q.; Ma, Y. Isolation of xanthones from the fermentation products of the endophytic fungus of Phomopsis amygdali. Chem. Nat. Compd. 2015, 51, 460–463. [Google Scholar] [CrossRef]
- Huang, R.; Ma, K.X.; Xie, X.S.; Wang, T.; Wu, S.H. Secondary metabolites of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2015, 51, 392–394. [Google Scholar] [CrossRef]
- Yuan, L.; Huang, W.Z.; Zhou, K.; Wang, Y.D.; Dong, W.; Lou, J.; Li, L.M.; Du, G.; Yang, H.Y.; Ma, Y.H.; et al. Xanthones from the fermentation products of an endophytic fungus Phomopsis sp. Heterocycles 2015, 91, 381–387. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, H.; Li, Y.; Ye, Y.; Hu, Q.; Gao, X.; Du, G. A new xanthone from the fermentation products of endophytic fungus of Phomopsis species. Asian J. Chem. 2014, 26, 4591–4593. [Google Scholar] [CrossRef]
- Hu, Q.; Yang, Y.; Yang, S.; Cao, H.; Chunyang, M.; Yang, H.; Gao, X.; Du, G. Xanthones from the fermentation products of the endophytic fungus of Phomopsis amygdali. Chem. Nat. Compd. 2015, 51, 456–459. [Google Scholar] [CrossRef]
- Huang, Z.; Yang, J.; Lei, F.; She, Z.; Lin, Y. A new xanthone O-glycoside from the mangrove endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2013, 49, 27–30. [Google Scholar] [CrossRef]
- Roensberg, D.; Debbab, A.; Mandi, A.; Vasylyeva, V.; Boehler, P.; Stork, B.; Engelke, L.; Hamacher, A.; Sawadogo, R.; Diederich, M.; et al. Pro-apoptotic and immunostimulatory tetrahydroxanthone dimers from the endophytic fungus Phomopsis longicolla. J. Org. Chem. 2013, 78, 12409–12425. [Google Scholar] [CrossRef] [PubMed]
- Shiono, Y.; Sasaki, T.; Shibuya, F.; Yasuda, Y.; Koseki, T.; Supratman, U. Isolation of a phomoxanthone A derivative, a new metabolite of tetrahydroxanthone, from a Phomopsis sp. isolated from the mangrove, Rhizhopora mucronata. Nat. Prod. Commun. 2013, 8, 1735–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meixiang, H.; Jing, L.; Lan, L.; Sheng, Y.; Jun, W.; Yongcheng, L.J.M.D. Phomopsichin A–D; four new chromone derivatives from mangrove endophytic fungus Phomopsis sp. 33#. Mar. Drugs 2016, 14, 215. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.; Kim, J.; Choi, J.N.; Ponnusamy, K.; Jeon, Y.; Kim, S.U.; Kim, J.G.; Lee, C.H. Identification, fermentation, and bioactivity against xanthomonas oryzae of antimicrobial metabolites isolated from Phomopsis longicolla S1B4. J. Microbiol. Biotechnol. 2010, 20, 494–500. [Google Scholar] [CrossRef]
- Ding, B.; Yuan, J.; Huang, X.; Wen, W.; Zhu, X.; Liu, Y.; Li, H.; Lu, Y.; He, L.; Tan, H.J.M.D. New dimeric members of the phomoxanthone family: Phomolactonexanthones A, B and deacetylphomoxanthone C isolated from the fungus Phomopsis sp. Mar. Drugs 2013, 11, 4961–4972. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Wang, Z.; Xia, G.; Huang, X.; Xu, F.; Chen, W.; She, Z. Three new chromone derivatives produced by Phomopsis sp. HNY29-2B from Acanthus ilicifolius linn. Chin. J. Chem. 2017, 35, 1889–1893. [Google Scholar] [CrossRef]
- Wu, Q.; Guo, Y.; Guo, Z.K.; Chu, Y.L.; Wang, T.; Tan, R.X. Two new cytosporones from the culture of endophytic Phomopsis sp. Chem. Nat. Compd. 2013, 48, 938–941. [Google Scholar] [CrossRef]
- Hu, H.B.; Luo, Y.F.; Wang, P.; Wang, W.J.; Wu, J. Xanthone-derived polyketides from the Thai mangrove endophytic fungus Phomopsis sp. xy21. Fitoterapia 2018, 131, 265–271. [Google Scholar] [CrossRef]
- Yang, J.X.; Qiu, S.; She, Z.; Lin, Y. A new isochroman derivative from the marine fungus Phomopsis sp. (No. Gx-4). Chem. Nat. Compd. 2014, 50, 424–426. [Google Scholar] [CrossRef]
- Ahmed, I.; Hussain, H.; Schulz, B.; Draeger, S.; Padula, D.; Pescitelli, G.; van Ree, T.; Krohn, K. Three new antimicrobial metabolites from the endophytic fungus Phomopsis sp. Eur. J. Org. Chem. 2011, 2011, 2867–2873. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, K.; Xu, Y.; Xia, X.; Wang, X.; Ge, M.; Shao, L. Three novel chromanones with biological activities from the endophytic fungus Phomopsis CGMCC No. 5416. J. Antibiot. 2020, 73, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, R.; Guo, Z.; She, Z.; Lin, Y. A new naphtho-γ-pyrone from mangrove endophytic fungus ZSU-H26. Chem. Nat. Compd. 2010, 46, 15–18. [Google Scholar] [CrossRef]
- Yang, J.; Xu, F.; Huang, C.; Li, J.; She, Z.; Pei, Z.; Lin, Y. Metabolites from the mangrove endophytic fungus Phomopsis sp. (#zsu-H76). Eur. J. Org. Chem. 2010, 2010, 3692–3695. [Google Scholar] [CrossRef]
- Yang, J.X.; Chen, Y.; Huang, C.; She, Z.; Lin, Y. A new isochroman derivative from the marine fungus Phomopsis sp. (No. ZH-111). Chem. Nat. Compd. 2011, 47, 13–16. [Google Scholar] [CrossRef]
- Tang, J.W.; Wang, W.G.; Li, A.; Yan, B.C.; Chen, R.; Li, X.N.; Du, X.; Sun, H.D.; Pu, J.X. Polyketides from the endophytic fungus Phomopsis sp. sh917 by using the one strain/many compounds strategy. Tetrahedron 2017, 73, 3577–3584. [Google Scholar] [CrossRef]
- Adelin, E.; Martin, M.T.; Cortial, S.; Retailleau, P.; Lumyong, S.; Ouazzani, J. Bioactive polyketides isolated from agar-supported fermentation of Phomopsis sp. CMU-LMA, taking advantage of the scale-up device, Platotex. Phytochemistry 2013, 93, 170–175. [Google Scholar] [CrossRef]
- Tao, M.H.; Chen, Y.C.; Wei, X.Y.; Tan, J.W.; Zhang, W.M. Chemical constituents of the endophytic fungus Phomopsis sp. A240 isolated from Taxus chinensis var. mairei. Helv. Chim. Acta 2014, 97, 426–430. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Islam, M.T.; Facey, P.; Douanla-Meli, C.; von Tiedemann, A.; Laatsch, H. Depsidones and other constituents from Phomopsis sp. CAFT69 and its host plant Endodesmia calophylloides with potent inhibitory effect on motility of zoospores of grapevine pathogen Plasmopara viticola. Phytochem. Lett. 2012, 5, 657–664. [Google Scholar] [CrossRef]
- Chapla, V.M.; Zeraik, M.L.; Ximenes, V.F.; Zanardi, L.M.; Lopes, M.N.; Cavalheiro, A.J.; Silva, D.H.S.; Young, M.C.M.; da Fonseca, L.M.; Bolzani, V.S.; et al. Bioactive secondary metabolites from Phomopsis sp., an endophytic fungus from Senna spectabilis. Molecules 2014, 19, 6597–6608. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xu, L.; Yang, L.; Huang, Y.; Li, S.; Shen, Y. Phomopsidone A, a novel depsidone metabolite from the mangrove endophytic fungus Phomopsis sp. A123. Fitoterapia 2014, 96, 146–151. [Google Scholar] [CrossRef]
- Kornsakulkarn, J.; Somyong, W.; Supothina, S.; Boonyuen, N.; Thongpanchang, C. Bioactive oxygen-bridged cyclooctadienes from endophytic fungus Phomopsis sp. BCC 45011. Tetrahedron 2015, 71, 9112–9116. [Google Scholar] [CrossRef]
- Xu, J.L.; Liu, Z.M.; Chen, Y.C.; Tan, H.B.; Li, H.H.; Li, S.N.; Guo, H.; Huang, Z.L.; Gao, X.X.; Liu, H.X.; et al. Lithocarols A-F, six tenellone derivatives from the deep-sea derived fungus Phomopsis lithocarpus FS508. Bioorg. Chem. 2019, 87, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Wang, Z.C.; Hu, W.Y.; Yan, K.L.; Wang, X.L.; Yang, H.M.; Yang, H.Y.; Gao, Y.H.; Liu, Q.; Hu, Q.F. Three new 3-methyl-2-arylbenzofurans from the fermentation products of an endophytic fungus Phomopsis sp. and their anti-TMV activity. Phytochem. Lett. 2017, 21, 287–290. [Google Scholar] [CrossRef]
- Song, H.C.; Qin, D.; Han, M.J.; Wang, L.; Zhang, K.; Dong, J.Y. Bioactive 2-pyrone metabolites from an endophytic Phomopsis asparagi SWUKJ5.2020 of Kadsura angustifolia. Phytochem. Lett. 2017, 22, 235–240. [Google Scholar] [CrossRef]
- Hussain, H.; Ahmed, I.; Schulz, B.; Draeger, S.; Krohn, K. Pyrenocines J-M: Four new pyrenocines from the endophytic fungus, Phomopsis sp. Fitoterapia 2012, 83, 523–526. [Google Scholar] [CrossRef]
- Yang, Z.; Ding, J.; Ding, K.; Chen, D.; Cen, S.; Ge, M. Phomonaphthalenone A: A novel dihydronaphthalenone with anti-HIV activity from Phomopsis sp. HCCB04730. Phytochem. Lett. 2013, 6, 257–260. [Google Scholar] [CrossRef]
- Li, X.B.; Chen, G.Y.; Liu, R.J.; Zheng, C.J.; Song, X.M.; Han, C.R. A new biphenyl derivative from the mangrove endophytic fungus Phomopsis longicolla HL-2232. Nat. Prod. Res. 2017, 31, 2264–2267. [Google Scholar] [CrossRef]
- Mishra, P.D.; Verekar, S.A.; Deshmukh, S.K.; Joshi, K.S.; Fiebig, H.H.; Kelter, G. Altersolanol A: A selective cytotoxic anthraquinone from a Phomopsis sp. Lett. Appl. Microbiol. 2015, 60, 387–391. [Google Scholar] [CrossRef]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
- Klaiklay, S.; Rukachaisirikul, V.; Phongpaichit, S.; Pakawatchai, C.; Saithong, S.; Buatong, J.; Preedanon, S.; Sakayaroj, J. Anthraquinone derivatives from the mangrove-derived fungus Phomopsis sp. PSU-MA214. Phytochem. Lett. 2012, 5, 738–742. [Google Scholar] [CrossRef]
- Hussain, H.; Tchimene, M.K.; Ahmed, I.; Meier, K.; Steinert, M.; Draeger, S.; Schulz, B.; Krohn, K. Antimicrobial chemical constituents from the endophytic fungus Phomopsis sp. from Notobasis syriaca. Nat. Prod. Commun. 2011, 6, 1905–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, R.; Chen, S.; Liu, Z.; Tan, C.; Huang, X.; She, Z. A new α-pyrone from the mangrove endophytic fungus Phomopsis sp. HNY29-2B. Nat. Prod. Res. 2017, 31, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Farooq, U.; Hussain, H.; Ahmed, I.; Rheinheimer, J.; Draeger, S.; Schulz, B.; van Ree, T. Phomosines H-J, novel highly substituted biaryl ethers, isolated from the endophytic fungus Phomopsis sp. from Ligustrum vulgare. Nat. Prod. Commun. 2011, 6, 1907–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.S.; Liang, M.J.; Mi, Q.L.; Chen, W.; Ling, J.; Chen, X.; Li, J.; Yang, G.Y.; Hu, Q.F.; Wang, W.G.; et al. Two new diphenyl ether derivatives from the fermentation products of the endophytic fungus Phomopsis asparagi. Chem. Nat. Compd. 2019, 55, 843–846. [Google Scholar] [CrossRef]
- Gao, Y.H.; Zheng, R.; Li, J.; Kong, W.S.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; et al. Three new diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. J. Asian Nat. Prod. Res. 2019, 21, 316–322. [Google Scholar] [CrossRef]
- Li, Z.J.; Yang, H.Y.; Li, J.; Liu, X.; Ye, L.; Kong, W.S.; Tang, S.Y.; Du, G.; Liu, Z.H.; Zhou, M.; et al. Isopentylated diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. J. Antibiot. 2018, 71, 359–362. [Google Scholar] [CrossRef]
- Yang, H.Y.; Duan, Y.Q.; Yang, Y.K.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; Hu, Q.F.; et al. Two new diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. Chem. Nat. Compd. 2019, 55, 428–431. [Google Scholar] [CrossRef]
- Yang, H.Y.; Duan, Y.Q.; Yang, Y.K.; Li, J.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; et al. Three new napthalene derivatives from the endophytic fungus Phomopsis fukushii. Phytochem. Lett. 2017, 22, 266–269. [Google Scholar] [CrossRef]
- Li, X.M.; Zeng, Y.C.; Chen, J.H.; Yang, Y.K.; Li, J.; Ye, L.; Du, G.; Zhou, M.; Hu, Q.F.; Guangyu, Y.; et al. Two new naphthalene derivatives from the fermentation products of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2019, 55, 618–621. [Google Scholar] [CrossRef]
- Xu, J.L.; Liu, H.X.; Chen, Y.C.; Tan, H.B.; Guo, H.; Xu, L.Q.; Li, S.N.; Huang, Z.L.; Li, H.H.; Gao, X.X.; et al. Highly substituted benzophenone aldehydes and eremophilane derivatives from the deep-Sea derived fungus Phomopsis lithocarpus FS508. Mar. Drugs 2018, 16, 329. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.X.; Shen, X.T.; Huang, R.; Wang, T.; Xie, X.S.; Liu, S.W.; Wu, S.H.; He, J. Bioactiye metabolites produced by the endophytic fungus Phomopsis sp. YM355364. Nat. Prod. Commun. 2014, 9, 669–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, S.L.; Li, Y.P.; Xiang, H.Y.; Liu, Y.; Wang, Y.D.; Kong, L.P.; Du, G.; Hu, Q.F.; Chen, Y.J.; Wang, W.G. Histone deacetylase inhibitor induced lipase Inhibitors from endophytic Phomopsis sp. 0391. Rec. Nat. Prod. 2020, 14, 42–47. [Google Scholar] [CrossRef]
- Kongprapan, T.; Xu, X.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J.; Chen, J.; Shen, X. Cytosporone derivatives from the endophytic fungus Phomopsis sp. PSU-H188. Phytochem. Lett. 2017, 22, 219–223. [Google Scholar] [CrossRef]
- Xu, J.; Tan, H.; Chen, Y.; Li, S.; Guo, H.; Huang, Z.; Li, H.; Gao, X.; Liu, H.; Zhang, W. Lithocarpinols A and B, a pair of diastereomeric antineoplastic tenellone derivatives from the deep-sea derived fungus Phomopsis lithocarpus FS508. Chin. Chem. Lett. 2019, 30, 439–442. [Google Scholar] [CrossRef]
- Chen, Y.G.; Pan, J.H.; Xu, F.; Liu, F.; Yang, J.X.; Huang, C.H.; Xu, C.L.; Lu, Y.J.; Cai, X.L.; She, Z.G.; et al. A new indene derivative from the marine fungus Phomopsis sp. (No. GX7-4A). Chem. Nat. Compd. 2010, 46, 230–232. [Google Scholar] [CrossRef]
- Tan, Q.W.; Fang, P.H.; Ni, J.C.; Gao, F.; Chen, Q.J. Metabolites Produced by an Endophytic Phomopsis sp. and Their Anti-TMV Activity. Molecules 2017, 22, 2073. [Google Scholar] [CrossRef] [Green Version]
- Shiono, Y.; Muslihah, N.I.; Suzuki, T.; Ariefta, N.R.; Anwar, C.; Nurjanto, H.H.; Aboshi, T.; Murayama, T.; Tawaraya, K.; Koseki, T.; et al. New eremophilane and dichlororesorcinol derivatives produced by endophytes isolated from Ficus ampelas. J. Antibiot. 2017, 70, 1133–1137. [Google Scholar] [CrossRef]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Srikitikulchai, P.; Srichomthong, K.; Lumyong, S. Oblongolides from the endophytic fungus Phomopsis sp. BCC 9789. J. Nat. Prod. 2010, 73, 55–59. [Google Scholar] [CrossRef]
- Lin, T.; Wang, G.H.; Lin, X.; Hu, Z.Y.; Chen, Q.C.; Xu, Y.; Zhang, X.K.; Chen, H.F. Three new oblongolides from Phomopsis sp. XZ-01, an endophytic fungus from Camptotheca acuminate. Molecules 2011, 16, 3351–3359. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Jiang, B.G.; Li, X.N.; Wang, Y.T.; Liu, S.S.; Zheng, K.X.; He, J.; Wu, S.H. Polyoxygenated cyclohexenoids with promising α-glycosidase inhibitory activity produced by Phomopsis sp. YE3250, an endophytic fungus derived from Paeonia delavayi. J. Agric. Food Chem. 2018, 66, 1140–1146. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Troise, C.; Santini, A.; Tuzi, A.; Vurro, M.; Ash, G.; Evidente, A. Phomentrioloxin: A phytotoxic pentasubstituted geranylcyclohexentriol produced by Phomopsis sp., a potential mycoherbicide for Carthamus lanatus Biocontrol. J. Nat. Prod. 2012, 75, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.L.; Mottier, N.; Jeanneret-Gris, J.; Christen, D.; Tabacchi, R.; Abou-Mansour, E. Differential production of phytotoxins from Phomopsis sp. from grapevine plants showing esca symptoms. J. Agric. Food Chem. 2014, 62, 8602–8607. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.S.; Wu, Y.; Qiao, Y.B.; Guo, Y.; Wang, J.P.; Hu, Z.X.; Zhang, Q.; Li, X.N.; Huang, J.F.; Zhou, Q.; et al. Protoilludane, illudalane, and botryane sesquiterpenoids from the endophytic fungus Phomopsis sp. TJ507A. J. Nat. Prod. 2018, 81, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Zanardi, L.M.; Bolzani, V.d.S.; Cavalheiro, A.J.; Siqueira Silva, D.H.; Trevisan, H.C.; Araujo, A.R.; Silva, G.H.; Teles, H.L.; Young, M.C.M. Sesquiterpenes produced by endophytic fungus Phomopsis cassiae with antifungal and acetylcholinesterase inhibition activities. Quim. Nova 2012, 35, 2233–2236. [Google Scholar] [CrossRef] [Green Version]
- Hemtasin, C.; Kanokmedhakul, S.; Kanokmedhakul, K.; Hahnvajanawong, C.; Soytong, K.; Prabpai, S.; Kongsaeree, P. Cytotoxic pentacyclic and tetracyclic aromatic sesquiterpenes from Phomopsis archeri. J. Nat. Prod. 2011, 74, 609–613. [Google Scholar] [CrossRef]
- Ma, X.; Wang, W.; Li, E.; Gao, F.; Guo, L.; Pei, Y. A new sesquiterpene from the entomogenous fungus Phomopsis amygdali. Nat. Prod. Res. 2016, 30, 276–280. [Google Scholar] [CrossRef]
- Qian, Y.X.; Kang, J.C.; Luo, Y.K.; He, J.; Wang, L.; Li, Q.R. Secondary metabolites of an endophytic fungus Phomopsis castaneae-mollissimae. Chem. Nat. Compd. 2018, 54, 346–347. [Google Scholar] [CrossRef]
- Wei, W.; Gao, J.; Shen, Y.; Chu, Y.L.; Xu, Q.; Tan, R.X. Immunosuppressive diterpenes from Phomopsis sp. S12. Eur. J. Org. Chem. 2014, 2014, 5728–5734. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, X.; Chen, J.W.; Shen, Y.; Jiang, N.; Tan, R.X.; Jiao, R.H.; Ge, H.M. Anti-inflammatory diterpenoids from an endophytic fungus Phomopsis sp. S12. Tetrahedron Lett. 2019, 60. [Google Scholar] [CrossRef]
- Zhang, Y.; Hao, F.; Liu, N.; Xu, Y.; Jia, A.; Yang, Z.; Xia, X.; Liu, C. Stereochemical determination of a new and cytotoxic euphane triterpenoid from the plant endophytic fungus Phomopsis chimonanthi. J. Antibiot. 2013, 66, 679–682. [Google Scholar] [CrossRef]
- Peyrat, L.A.; Eparvier, V.; Eydoux, C.; Guillemot, J.C.; Litaudon, M.; Stien, D. Betulinic acid, the first lupane-type triterpenoid isolated from both a Phomopsis sp. and its host plant Diospyros carbonaria benoist. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Huang, R.; Miao, C.P.; Chen, Y.W. Two new steroids from an endophytic fungus Phomopsis sp. Chem. Biodivers. 2013, 10, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.X.; Wu, Y.; Xie, S.S.; Sun, W.G.; Guo, Y.; Li, X.N.; Liu, J.J.; Li, H.; Wang, J.P.; Luo, Z.W.; et al. Phomopsterones A and B, two functionalized ergostane-type steroids from the endophytic fungus Phomopsis sp. TJ507A. Org. Lett. 2017, 19, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Adelin, E.; Servy, C.; Cortial, S.; Levaique, H.; Martin, M.T.; Retailleau, P.; Le Goff, G.; Bussaban, B.; Lumyong, S.; Ouazzani, J. Isolation, structure elucidation and biological activity of metabolites from Sch-642305-producing endophytic fungus Phomopsis sp. CMU-LMA. Phytochemistry 2011, 72, 2406–2412. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, H.; Chen, Y.; Li, S.; Huang, Z.; Guo, H.; Li, H.; Gao, X.; Liu, H.; Zhang, W. Lithocarpins A-D: Four tenellone-macrolide conjugated [4 + 2] hetero-adducts from the deep-sea derived fungus Phomopsis lithocarpus FS508. Org. Chem. Front. 2018, 5, 1792–1797. [Google Scholar] [CrossRef]
- Yan, B.C.; Wang, W.G.; Hu, D.B.; Sun, X.; Kong, L.M.; Li, X.N.; Du, X.; Luo, S.H.; Liu, Y.; Li, Y.; et al. Phomopchalasins A and B, two cytochalasans with polycyclic-fused skeletons from the endophytic fungus Phomopsis sp. shj2. Org. Lett. 2016, 18, 1108–1111. [Google Scholar] [CrossRef]
- Luo, Y.F.; Zhang, M.; Dai, J.G.; Pedpradab, P.; Wang, W.J.; Wu, J. Cytochalasins from mangrove endophytic fungi Phomopsis spp. xy21 and xy22. Phytochem. Lett. 2016, 17, 162–166. [Google Scholar] [CrossRef]
- Jouda, J.B.; Tamokou, J.D.D.; Mbazoa, C.D.; Douala-Meli, C.; Sarkar, P.; Bag, P.K.; Wandji, J. Antibacterial and cytotoxic cytochalasins from the endophytic fungus Phomopsis sp. harbored in Garcinia kola (Heckel) nut. BMC Complement. Altern. Med. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Zhou, Y.; Li, H.F.; Ye, Y.H.; Guo, J.H. Antifungal metabolites from Phomopsis sp. By254, an endophytic fungus in Gossypium hirsutum. Afr. J. Microbiol. Res. 2011, 5, 1231–1236. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.S.; Peng, C.J.; Cheng, M.J.; Wu, H.C.; Chan, H.Y.; Hsieh, S.Y.; Yuan, G.F.; Chen, I.S. Chemical constituents of the endophytic fungus Phomopsis asparagi isolated from the plant Peperomia sui. Chem. Nat. Compd. 2018, 54, 504–508. [Google Scholar] [CrossRef]
- Chen, S.C.; Liu, Z.M.; Tan, H.B.; Chen, Y.C.; Li, S.N.; Li, H.H.; Guo, H.; Zhu, S.; Liu, H.X.; Zhang, W.M. Tersone A-G, new pyridone alkaloids from the deep-sea fungus Phomopsis tersa. Mar. Drugs 2019, 17, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Huang, M.; Li, X.; Liu, L.; Chen, B.; Wang, J.; Lin, Y. Phochrodines A-D, first naturally occurring new chromenopyridines from mangrove entophytic fungus Phomopsis sp. 33#. Fitoterapia 2018, 124, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Verekar, S.A.; Mishra, P.D.; Sreekumar, E.S.; Deshmukh, S.K.; Fiebig, H.H.; Kelter, G.; Maier, A. Anticancer activity of new depsipeptide compound isolated from an endophytic fungus. J. Antibiot. 2014, 67, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Katz, L. Manipulation of modular polyketide syntheses. Chem. Rev. 1997, 97, 2557–2575. [Google Scholar] [CrossRef]
- Le Pogam, P.; Boustie, J. Xanthones of lichen source: A 2016 update. Molecules 2016, 21, 294. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.D.; Jiang, Y.Y.; Guo, F.X.; Chen, L.X.; Xu, L.L.; Zhang, W.; Liu, B. The antitumor activity of naturally occurring chromones: A review. Fitoterapia 2019, 135, 114–129. [Google Scholar] [CrossRef]
- Mayuri, B.; Kavitha, P.; Basavoju, S.; Bhargavi, G.; Reddy, K.L. Synthesis, structural characterisation and biological evolution of chromanones. J. Mol. Struct. 2017, 1145, 1–9. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Cheng, J.P. Catalytic asymmetric synthesis of chiral benzofuranones. Adv. Synth. Catal. 2014, 356, 1172–1198. [Google Scholar] [CrossRef]
- McGlacken, G.P.; Fairlamb, I.J.S. 2-Pyrone natural products and mimetics: Isolation, characterisation and biological activity. Nat. Prod. Rep. 2005, 22, 369–385. [Google Scholar] [CrossRef]
- Bolton, J.L.; Dunlap, T. Formation and biological targets of quinones: Cytotoxic versus cytoprotective effects. Chem. Res. Toxicol. 2017, 30, 13–37. [Google Scholar] [CrossRef]
- Jones, T.J.M.; Douglas, C.J. The metabolism and toxicity of quinones, quinonimines, quinone methides, and quinone-thioethers. Curr. Drug Metab. 2002, 3, 425–438. [Google Scholar] [CrossRef]
- Gajera, H.P.; Gevariya, S.N.; Hirpara, D.G.; Patel, S.V.; Golakiya, B.A. Antidiabetic and antioxidant functionality associated with phenolic constituents from fruit parts of indigenous black jamun (Syzygium cumini L.) landraces. J. Food Sci. Technol. 2017, 54, 3180–3191. [Google Scholar] [CrossRef] [PubMed]
- Shing, T.K.M.; Yang, J. A short synthesis of natural (−)-oblongolide via an intramolecular or a transannular diels-alder reaction. J. Org. Chem. 1995, 60, 5785–5789. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.J.; Huang, M.Q.; Bao, J.L.; Chen, X.P.; Wang, Y.T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investig. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef] [PubMed]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol. 2011, 3, 228–249. [Google Scholar] [CrossRef]
- Zielinska-Blajet, M.; Feder-Kubis, J. Monoterpenes and their derivatives-recent development in biological and medical applications. Int. J. Mol. Sci. 2020, 21, 7078. [Google Scholar] [CrossRef]
- Chen, D.L.; Wang, B.W.; Sun, Z.C.; Yang, J.S.; Xu, X.D.; Ma, G.X. Natural nitrogenous sesquiterpenoids and their bioactivity: A review. Molecules 2020, 25, 2485. [Google Scholar] [CrossRef]
- Chen, L.; Lu, X.; El-Seedi, H.; Teng, H. Recent advances in the development of sesquiterpenoids in the treatment of type 2 diabetes. Trends Food Sci. Technol. 2019, 88, 46–56. [Google Scholar] [CrossRef]
- Su, Y.D.; Su, J.H.; Hwang, T.L.; Wen, Z.H.; Sheu, J.H.; Wu, Y.C.; Sung, P.J. Briarane diterpenoids isolated from octocorals between 2014 and 2016. Mar. Drugs 2017, 15, 44. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Kinghorn, A.D. Natural product triterpenoids and their semi-synthetic derivatives with potential anticancer activity. Planta Med. 2019, 85, 802–814. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.U.; Ismail, M.; Khurram, M.; Ullah, I.; Rabbi, F.; Iriti, M. Bioactive steroids and saponins of the genus Trillium. Molecules 2017, 22, 2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, J.; He, S.; Yan, X. A review study on macrolides isolated from cyanobacteria. Mar. Drugs 2017, 15, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.K.; Tripathi, G.; Kishore, N.; Singh, R.K.; Singh, A.; Tiwari, V.K. Drug development against tuberculosis: Impact of alkaloids. Eur. J. Med. Chem. 2017, 137, 504–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Luo, X.; Yang, J.; Chen, F.; Lin, X.; Chen, C.; Zhou, X.; Liu, S.; Liu, Y. Structurally diverse polyketides from the mangrove-derived fungus Diaporthe sp. SCSIO 41011 with their anti-influenza A virus activities. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ruan, Q.; Jiang, S.; Qu, Y.; Chen, J.; Zhao, M.; Yang, B.; Liu, Y.; Zhao, Z.; Cui, H. Cytochalasins and polyketides from the fungus Diaporthe sp. GZU-1021 and their anti-inflammatory activity. Fitoterapia 2019, 137. [Google Scholar] [CrossRef]
- Niu, Z.; Chen, Y.; Guo, H.; Li, S.N.; Li, H.H.; Liu, H.X.; Liu, Z.; Zhang, W. Cytotoxic polyketides from a deep-sea sediment derived fungus Diaporthe phaseolorum FS431. Molecules 2019, 24, 3062. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Ding, M.; Huang, D.; Zhang, Z.; Liu, H.; Huang, H.; She, Z. Chroman-4-one and pyrano[4,3-b]chromenone derivatives from the mangrove endophytic fungus Diaporthe phaseolorum SKS019. RSC Adv. 2017, 7, 20128–20134. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhao, J.; Liang, X.; Lv, X.; Li, Y.; Qu, J.; Liu, Y. Dothiorelone derivatives from an endophyte Diaporthe pseudomangiferaea inhibit the activation of human lung fibroblasts MRC-5 cells. Fitoterapia 2018, 127, 7–14. [Google Scholar] [CrossRef]
- Bungihan, M.E.; Tan, M.A.; Kitajima, M.; Kogure, N.; Franzblau, S.G.; dela Cruz, T.E.E.; Takayama, H.; Nonato, M.G. Bioactive metabolites of Diaporthe sp. P133, an endophytic fungus isolated from Pandanus amaryllifolius. J. Nat. Med. 2011, 65, 606–609. [Google Scholar] [CrossRef]
- De Medeiros, A.G.; Savi, D.C.; Mitra, P.; Shaaban, K.A.; Jha, A.K.; Thorson, J.S.; Rohr, J.; Glienke, C. Bioprospecting of Diaporthe terebinthifolii LGMF907 for antimicrobial compounds. Folia Microbiol. 2018, 63, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Meepagala, K.M.; Briscoe, W.E.; Techen, N.; Johnson, R.D.; Clausen, B.M.; Duke, S.O. Isolation of a phytotoxic isocoumarin from Diaporthe eres-infected Hedera helix (English ivy) and synthesis of its phytotoxic analogs. Pest Manag. Sci. 2018, 74, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Singamaneni, V.; Sharma, N.; Kumar, A.; Arora, D.; Kushwaha, M.; Bhushan, S.; Jaglan, S.; Gupta, P. Valproic acid induces three novel cytotoxic secondary metabolites in Diaporthe sp., an endophytic fungus from Datura inoxia Mill. Bioorganic Med. Chem. Lett. 2018, 28, 2217–2221. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, Z.; Lin, X.; Lu, C.; Shen, Y. A new polyketide from Diaporthe sp. SXZ-19, an endophytic fungal strain of Camptotheca acuminate. Nat. Prod. Res. 2013, 27, 2100–2104. [Google Scholar] [CrossRef] [PubMed]
- Evidente, M.; Boari, A.; Vergura, S.; Cimmino, A.; Vurro, M.; Ash, G.; Superchi, S.; Evidente, A. Structure and absolute configuration of kongiidiazadione, a new phytotoxic 3-substituted-5-diazenylcyclopentendione produced by Diaporthe Kongii. Chirality 2015, 27, 557–562. [Google Scholar] [CrossRef]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef]
- Riga, R.; Happyana, N.; Quentmeier, A.; Zammarelli, C.; Kayser, O.; Hakim, E.H. Secondary metabolites from Diaporthe lithocarpus isolated from Artocarpus heterophyllus. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef]
- Ratnaweera, P.B.; Jayasundara, J.M.N.M.; Herath, H.H.M.S.D.; Williams, D.E.; Rajapaksha, S.U.; Nishantha, K.M.D.W.P.; de Silva, E.D.; Andersen, R.J. Antifeedant, contact toxicity and oviposition deterrent effects of phyllostine acetate and phyllostine isolated from the endophytic fungus Diaporthe miriciae against Plutella xylostella larvae. Pest Manag. Sci. 2020, 76, 1541–1548. [Google Scholar] [CrossRef]
- Wulansari, D.; Julistiono, H.; Nurkanto, A.; Agusta, A. Antifungal activity of (+)-2,2′-epicytoskyrin A and its membrane-disruptive action. Makara J. Sci. 2016, 20, 160–166. [Google Scholar] [CrossRef]
- Tian, W.; Liao, Z.; Zhou, M.; Wang, G.; Wu, Y.; Gao, S.; Qiu, D.; Liu, X.; Lin, T.; Chen, H. Cytoskyrin C, an unusual asymmetric bisanthraquinone with cage-like skeleton from the endophytic fungus Diaporthe sp. Fitoterapia 2018, 128, 253–257. [Google Scholar] [CrossRef]
- Specian, V.; Sarragiotto, M.H.; Pamphile, J.A.; Clemente, E. Chemical characterization of bioactive compounds from the endophytic fungus Diaporthe helianthi isolated from Luehea divaricata. Braz. J. Microbiol. 2012, 43, 1174–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reveglia, P.; Pacetti, A.; Masi, M.; Cimmino, A.; Carella, G.; Marchi, G.; Mugnai, L.; Evidente, A. Phytotoxic metabolites produced by Diaporthe eres involved in cane blight of grapevine in Italy. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Noriler, S.A.; Savi, D.C.; Ponomareva, L.V.; Rodrigues, R.; Rohr, J.; Thorson, J.S.; Glienke, C.; Shaaban, K.A. Vochysiamides A and B: Two new bioactive carboxamides produced by the new species Diaporthe vochysiae. Fitoterapia 2019, 138. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.P.B.; Aguilar-Perez, M.M.; Arnold, A.E.; Rios, N.; Coley, P.D.; Kursar, T.A.; Cubilla-Rios, L. Chemical constituents and their antibacterial activity from the tropical endophytic fungus Diaporthe sp. F2934. J. Appl. Microbiol. 2016, 120, 1501–1508. [Google Scholar] [CrossRef] [Green Version]
- Ola, A.R.B.; Debbab, A.; Kurtan, T.; Broetz-Oesterhelt, H.; Aly, A.H.; Proksch, P. Dihydroanthracenone metabolites from the endophytic fungus Diaporthe melonis isolated from Annona squamosa. Tetrahedron Lett. 2014, 55, 3147–3150. [Google Scholar] [CrossRef]
- Nakashima, K.I.; Tomida, J.; Kamiya, T.; Hirai, T.; Morita, Y.; Hara, H.; Kawamura, Y.; Adachi, T.; Inoue, M. Diaporthols A and B: Bioactive diphenyl ether derivatives from an endophytic fungus Diaporthe sp. Tetrahedron Lett. 2018, 59, 1212–1215. [Google Scholar] [CrossRef]
- Cui, H.; Lin, Y.; Luo, M.; Lu, Y.; Huang, X.; She, Z. Diaporisoindoles A-C: Three isoprenylisoindole alkaloid derivatives from the mangrove endophytic fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef]
- Cui, H.; Liu, Y.; Li, J.; Huang, X.; Yan, T.; Cao, W.; Liu, H.; Long, Y.; She, Z. Diaporindenes A-D: Four unusual 2,3-dihydro-1H-indene analogues with anti-inflammatory activities from the mangrove endophytic fungus Diaporthe sp. SYSU-HQ3. J. Org. Chem. 2018, 83, 11804–11813. [Google Scholar] [CrossRef]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and phomentrioloxins B and C Produced by Diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef]
- Luo, X.; Lin, X.; Tao, H.; Wang, J.; Li, J.; Yang, B.; Zhou, X.; Liu, Y. Isochromophilones A-F, cytotoxic chloroazaphilones from the marine mangrove endophytic fungus Diaporthe sp. SCSIO 41011. J. Nat. Prod. 2018, 81, 934–941. [Google Scholar] [CrossRef]
- Chen, C.J.; Liu, X.X.; Zhang, W.J.; Zang, L.Y.; Wang, G.; Ng, S.W.; Tan, R.X.; Ge, H.M. Sesquiterpenoids isolated from an endophyte fungus Diaporthe sp. RSC Adv. 2015, 5, 17559–17565. [Google Scholar] [CrossRef]
- Mandavid, H.; Rodrigues, A.M.S.; Espindola, L.S.; Eparvier, V.; Stien, D. Secondary metabolites isolated from the amazonian endophytic fungus Diaporthe sp. SNB-GSS10. J. Nat. Prod. 2015, 78, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Y.; Li, H.; Li, S.; Tan, H.; Liu, Z.; Li, D.; Liu, H.; Zhang, W. Four new metabolites from the endophytic fungus Diaporthe lithocarpus A740. Fitoterapia 2019, 137. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Kusari, P.; Kayser, O.; Spiteller, M. Endophytic Diaporthe sp. LG23 produces a potent antibacterial tetracyclic triterpenoid. J. Nat. Prod. 2015, 78, 2128–2132. [Google Scholar] [CrossRef]
- Ito, A.; Maeda, H.; Tonouchi, A.; Hashimoto, M. Relative and absolute structure of phomolide C. Biosci. Biotechnol. Biochem. 2015, 79, 1067–1069. [Google Scholar] [CrossRef] [Green Version]
- Yedukondalu, N.; Arora, P.; Wadhwa, B.; Malik, F.A.; Vishwakarma, R.A.; Gupta, V.K.; Riyaz-Ul-Hassan, S.; Ali, A. Diapolic acid A-B from an endophytic fungus, Diaporthe terebinthifolii depicting antimicrobial and cytotoxic activity. J. Antibiot. 2017, 70, 212–215. [Google Scholar] [CrossRef]
- Brissow, E.R.; da Silva, I.P.; de Siqueira, K.A.; Senabio, J.A.; Pimenta, L.P.; Januario, A.H.; Magalhaes, L.G.; Furtado, R.A.; Tavares, D.C.; Sales Junior, P.A.; et al. 18-Des-hydroxy cytochalasin: An antiparasitic compound of Diaporthe phaseolorum-92C, an endophytic fungus isolated from Combretum lanceolatum Pohl ex Eichler. Parasitol. Res. 2017, 116, 1823–1830. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, D.; Liang, Y.; Liu, X.; Cao, F.; Qin, Y.; Mo, T.; Xu, Z.; Li, J.; Yang, R. Cytochalasins from endophytic Diaporthe sp. GDG-118. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef]
- Chang, F.R.; Wang, S.W.; Li, C.Y.; Lu, Y.Y.; Liu, S.Y.V.; Chen, C.Y.; Wu, Y.C.; Cheng, Y.B. Natural products from Diaporthe arecae with anti-angiogenic activity. Isr. J. Chem. 2019, 59, 439–445. [Google Scholar] [CrossRef]
- Cui, H.; Yu, J.; Chen, S.; Ding, M.; Huang, X.; Yuan, J.; She, Z. Alkaloids from the mangrove endophytic fungus Diaporthe phaseolorum SKS019. Bioorganic Med. Chem. Lett. 2017, 27, 803–807. [Google Scholar] [CrossRef]
- Schloss, S.; Hackl, T.; Herz, C.; Lamy, E.; Koch, M.; Rohn, S.; Maul, R. Detection of a toxic methylated derivative of phomopsin A produced by the legume-infesting fungus Diaporthe toxica. J. Nat. Prod. 2017, 80, 1930–1934. [Google Scholar] [CrossRef] [PubMed]
- Sebastianes, F.L.S.; Cabedo, N.; El Aouad, N.; Valente, A.M.M.P.; Lacava, P.T.; Azevedo, J.L.; Pizzirani-Kleiner, A.A.; Cortes, D. 3-Hydroxypropionic acid as an antibacterial agent from endophytic fungi Diaporthe phaseolorum. Curr. Microbiol. 2012, 65, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Yang, X.Q.; Wan, C.P.; Wang, B.Y.; Yin, H.Y.; Shi, L.J.; Wu, Y.M.; Yang, Y.B.; Zhou, H.; Ding, Z.T. Potential antihyperlipidemic polyketones from endophytic Diaporthe sp. JC-J7 in Dendrobium nobile. RSC Adv. 2018, 8, 41810–41817. [Google Scholar] [CrossRef] [Green Version]
- Yenn, T.W.; Ring, L.C.; Nee, T.W.; Khairuddean, M.; Zakaria, L.; Ibrahim, D. Endophytic Diaporthe sp. ED2 produces a novel anti-candidal ketone derivative. J. Microbiol. Biotechnol. 2017, 27, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- El-Helw, E.A.E.; Hashem, A.I. Synthesis and antitumor activity evaluation of some pyrrolone and pyridazinone heterocycles derived from 3-((2-oxo-5-(p-tolyl)furan-3(2H)-ylidene)methyl)quinolin-2(1H)-one. Synth. Commun. 2020, 50, 1046–1055. [Google Scholar] [CrossRef]
- Zheng, C.J.; Shao, C.L.; Chen, M.; Niu, Z.G.; Zhao, D.L.; Wang, C.Y. Merosesquiterpenoids and ten-membered macrolides from a soft coral-derived Lophiostoma sp. fungus. Chem. Biodivers. 2015, 12, 1407–1414. [Google Scholar] [CrossRef]
- Jozwiak, M.; Filipowska, A.; Fiorino, F.; Struga, M. Anticancer activities of fatty acids and their heterocyclic derivatives. Eur. J. Pharmacol. 2020, 871. [Google Scholar] [CrossRef]






















{kind=link}
{kind=link}
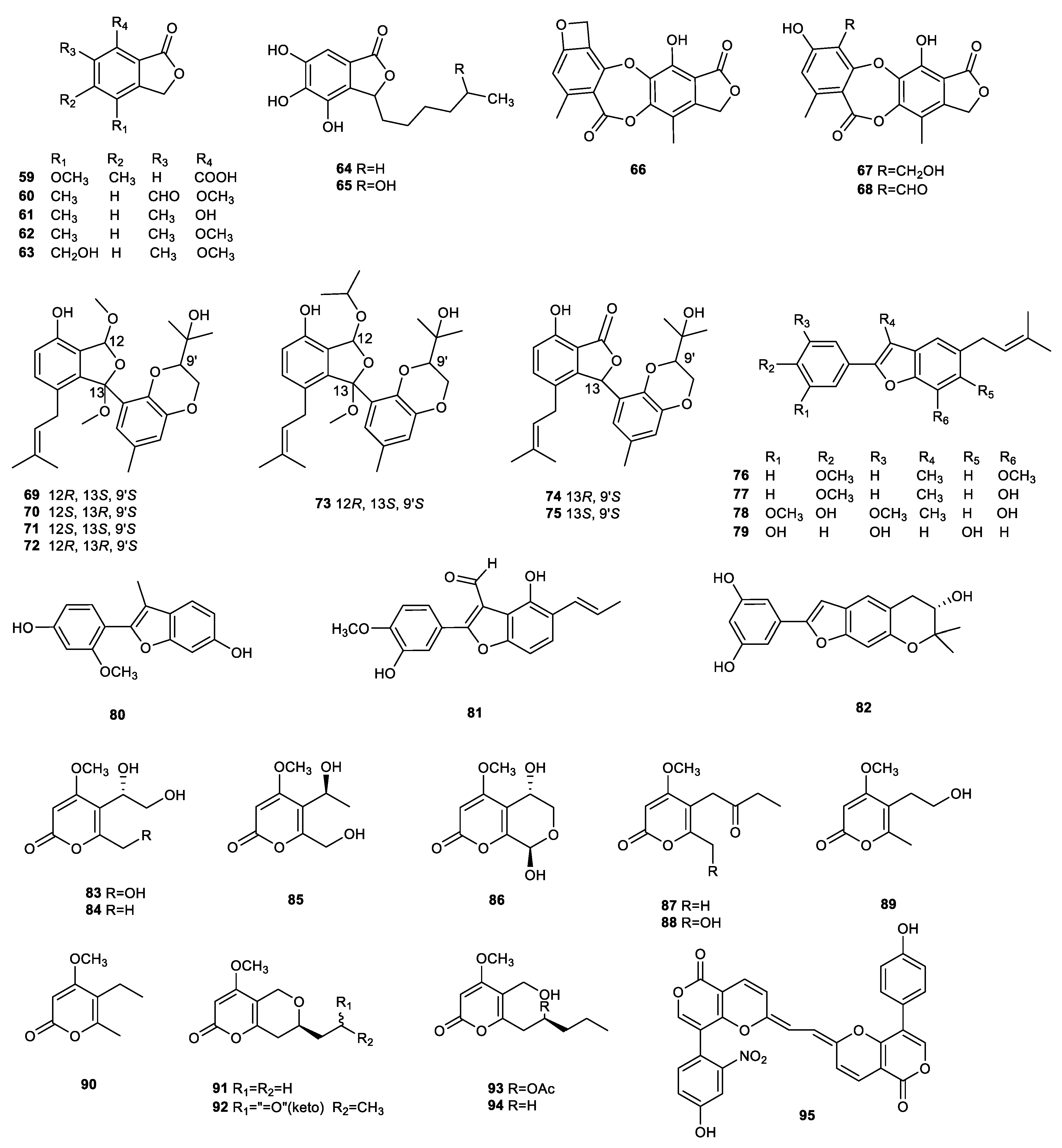{kind=link}
{kind=link}
{kind=link}
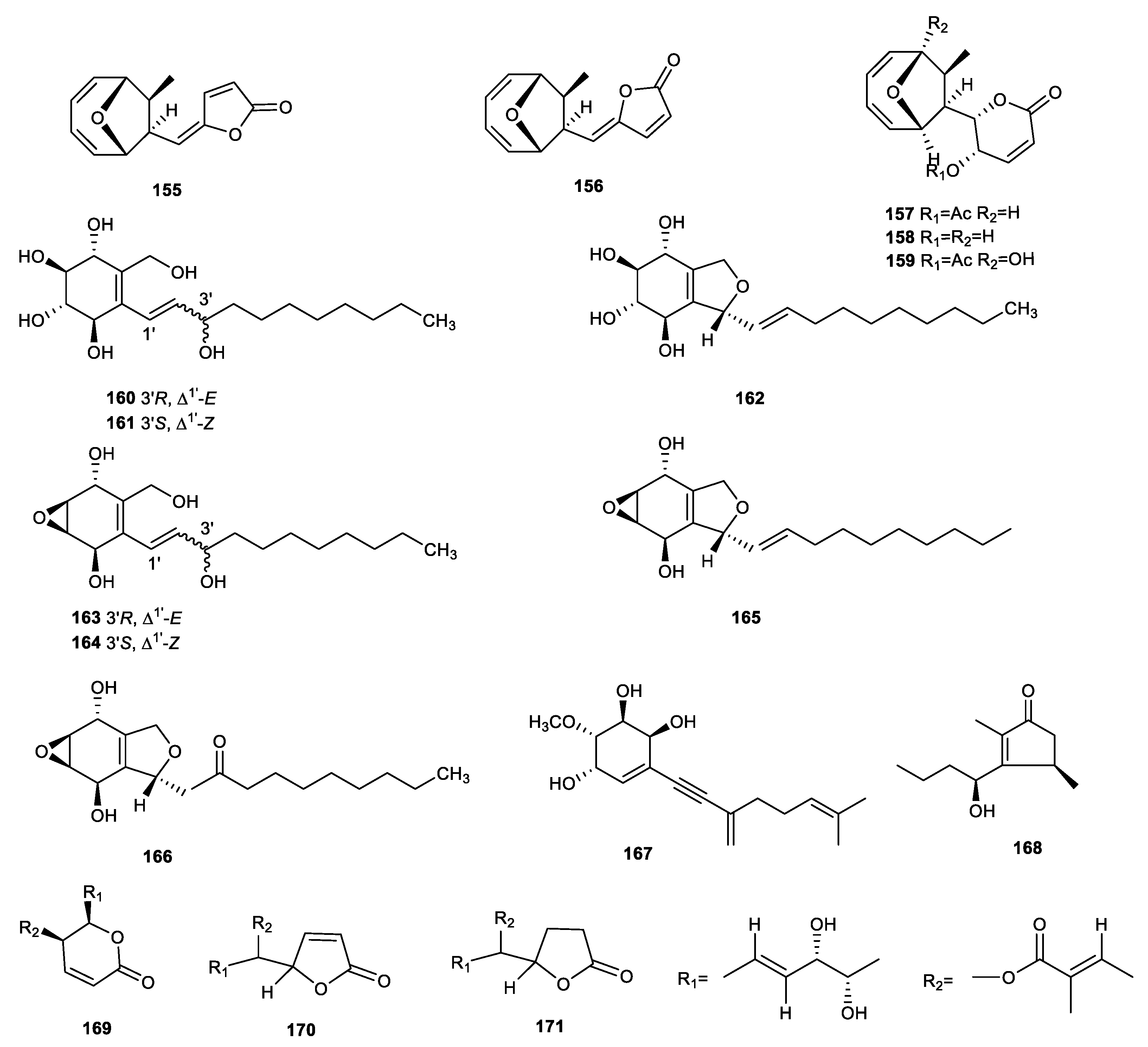{kind=link}
{kind=link}
{kind=link}
{kind=link}
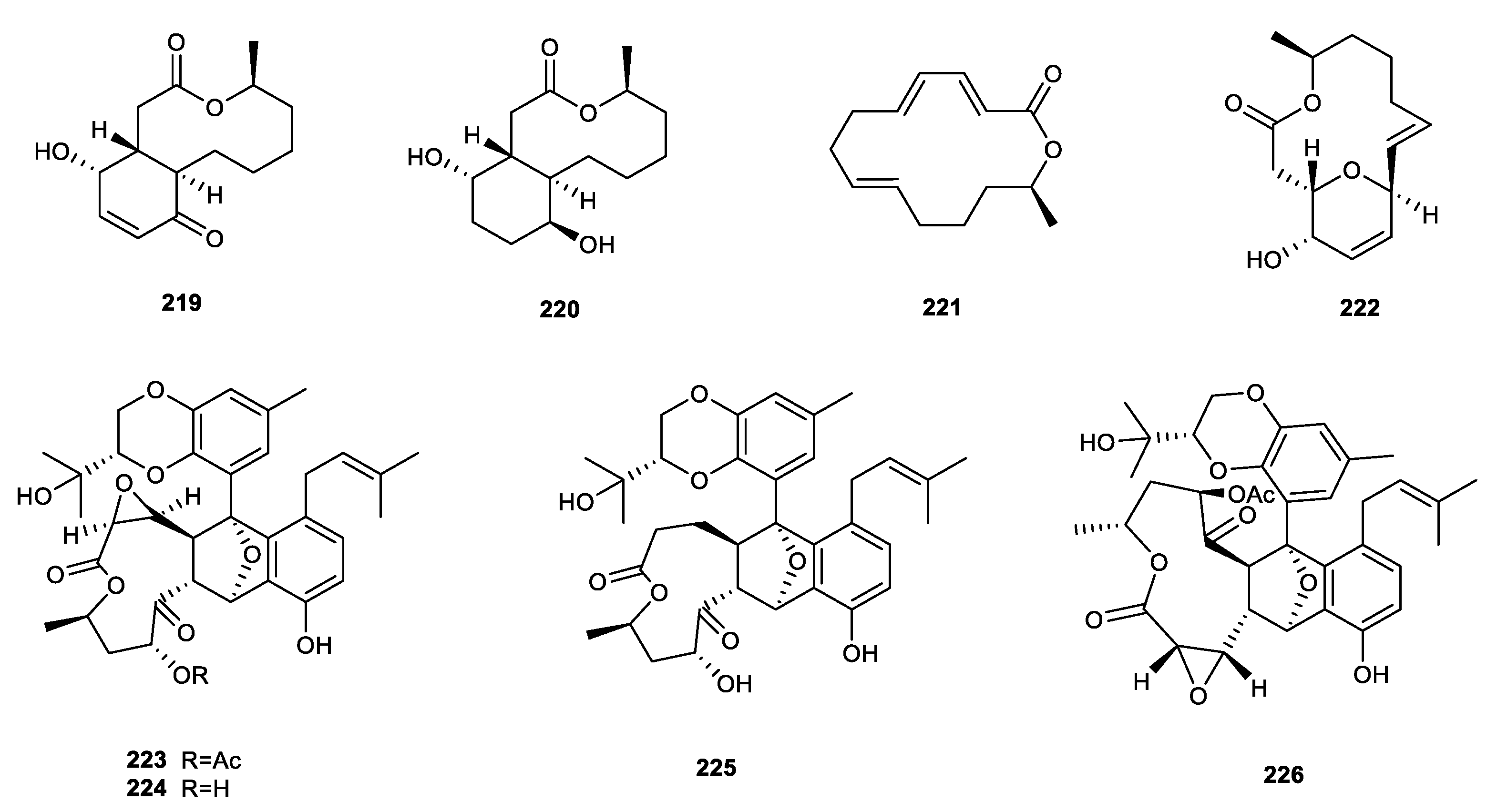{kind=link}
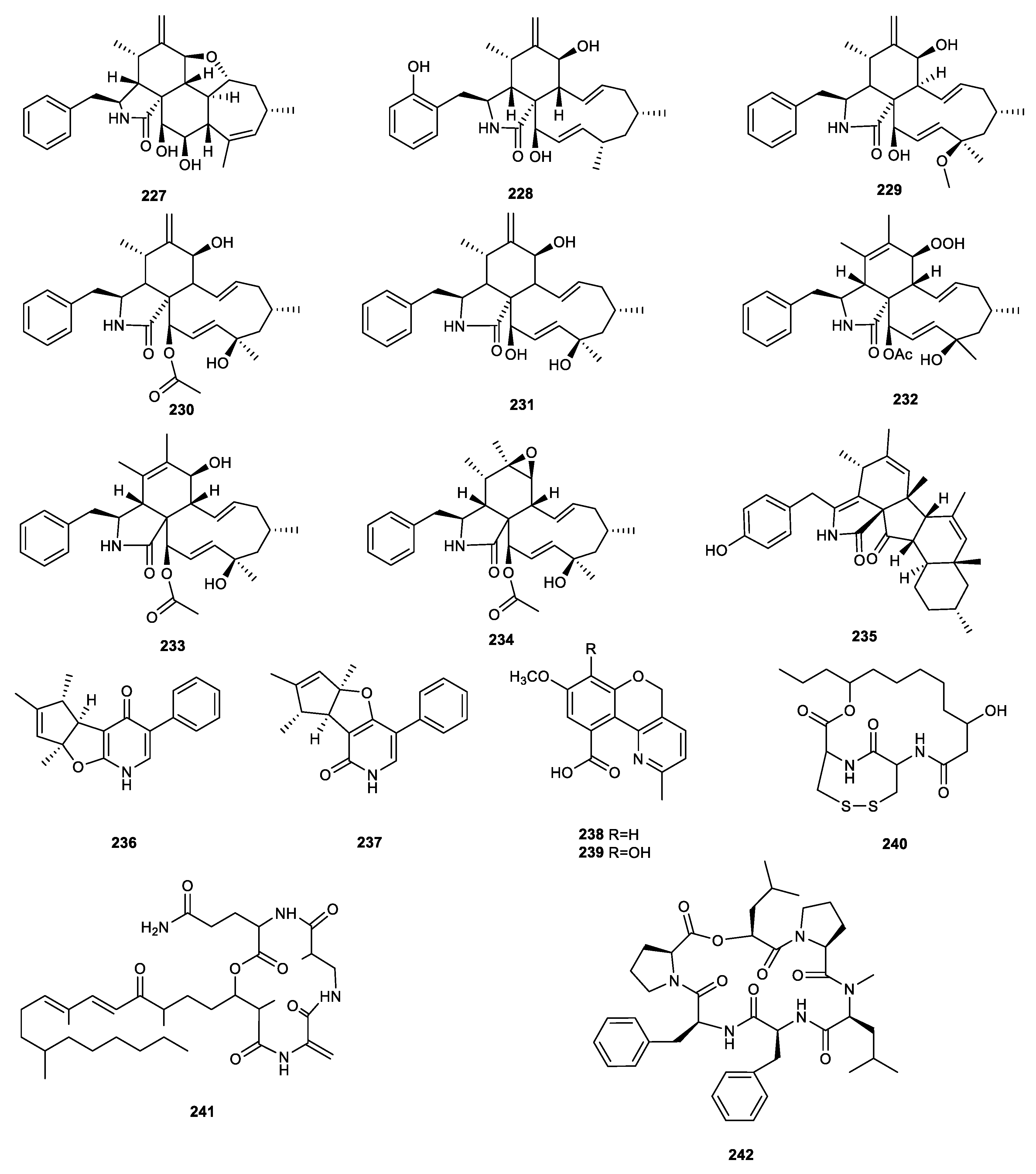{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Structural Types | Compounds | Strains | Habitats (T/M a) | Activities | Refs. |
|---|---|---|---|---|---|---|
| 1 | Xanthones | 1,5-Dihydroxy-3-hydroxyethyl-6-methoxy-carbonylxanthone | Phomopsis sp. | Paris polyphylla var. yunnanensis (T) | Cytotoxic | [23] |
| 2 | 1-Hydroxy-5-methoxy-3-hydroxyethyl-6-methoxycarbonylxanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 3 | 1-Hydroxy-3-hydroxyethyl-8-ethoxycarbonyl-xanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 4 | Pinselin | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 5 | 1-Hydroxy-8-(hydroxymethyl)-3-methoxy-6-methylxanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 6 | 2,6-Dihydroxy-3-methyl-9-oxoxanthene-8-carboxylic acid methyl ester | Phomopsis sp. (No. SK7RN3G1) | Sediment (M) | Cytotoxic | [24] | |
| 7 | 4,5-Dihydroxy-3-(2-hydroxyethyl)-1-methoxy-8-methoxy- carbonylxanthone | P. amygdali | Paris axialis (T) | Cytotoxic | [25] | |
| 8 | 1,8-Dihydroxy-4-(2-hydroxyethyl)-3-methoxyxanthone | P. amygdali | P. axialis (T) | Cytotoxic | [25] | |
| 9 | Hydroxyvertixanthone | Phomopsis sp. YM 355364 | Aconitum carmichaelii (T) | Antimicrobial | [26] | |
| 10 | Dalienxanthone A | Phomopsis sp. | Paris daliensis (T) | Cytotoxic | [27] | |
| 11 | Dalienxanthone B | Phomopsis sp. | P. daliensis (T) | Cytotoxic | [27] | |
| 12 | Dalienxanthone C | Phomopsis sp. | P. daliensis (T) | Cytotoxic | [27] | |
| 13 | Paucinervin E | P. amygdali | P. axialis (T) | Cytotoxic | [25] | |
| 14 | 1,3-Dihydroxy-4-(1,3,4-trihydroxybutan-2-yl)-8-methoxy-9H-xanthen-9-one | P. amygdali | P. polyphylla var. yunnanensis (T) | Cytotoxic | [28] | |
| 15 | 3-Methoxy-1,4,8-trihydroxy-5-(1ʹ,3ʹ,4ʹ-trihydroxybutan-2ʹ-yl)-xanthone | P. amygdali | P. axialis (T) | Cytotoxic | [29] | |
| 16 | 8-Methoxy-1,3,4-trihydroxy-5-(1ʹ,3ʹ,4ʹ-trihydroxybutan-2ʹ-yl)-xanthone | P. amygdali | P. axialis (T) | Cytotoxic | [29] | |
| 17 | Secosterigmatocystin | Phomopsis sp. P. amygdali | P. polyphylla var. yunnanensis (T) P. axialis (T) | Cytotoxic Cytotoxic | [23] [29] | |
| 18 | 3,8-Dihydroxy-4-(2,3-dihydroxy-1-hydroxymethylpropyl)-1-methoxyxanthone | Phomopsis sp. | P. daliensis (T) | Cytotoxic | [27] | |
| 19 | Oliganthins E | Phomopsis sp. | P. daliensis (T) | Cytotoxic | [27] | |
| 20 | Dihydrosterigmatocystin | P. amygdali | P. axialis(T) | Cytotoxic | [29] | |
| 21 | Vieillardixanthone | P. amygdali | P. axialis (T) | Cytotoxic | [29] | |
| 22 | 1,7-Dihydroxy-2-methoxy-3-(3-methylbut-2-enyl)xanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 23 | 1-Hydroxy-4,7-dimethoxy-6-(3-oxobutyl)-xanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 24 | Asperxanthone | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 25 | 6-O-Methyl-2-deprenylrheediaxanthone B | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Cytotoxic | [23] | |
| 26 | Cratoxylumxanthone D | Phomopsis sp. | P. daliensis (T) | Cytotoxic | [27] | |
| 27 | 3-O-(6-O-α-L-Arabinopyranosyl)-β-D-glucopyranosyl-1,4-dimethoxyxanthone | Phomopsis sp. (ZH76) | Excoecaria agallocha (M) | Cytotoxic | [30] | |
| 28 | Phomoxanthone A | P. longicolla Phomopsis sp. IM 41-1 Phomopsis sp. 33# | Sonneratia caseolaris (M) Rhizhopora mucronata (M) Rhizophora stylosa (M) | Pro-apoptotic Antimicrobial Inhibiting acetylcholinesterase and α-glucosidase, Antioxidant | [31] [32] [33] | |
| 29 | 12-O-Deacetyl-phomoxanthone A | Phomopsis sp. IM 41-1 | R. mucronata (M) | Antimicrobial | [32] | |
| 30 | Dicerandrol A | P. longicolla S1B4 Phomopsis sp. HNY29-2B | - b Acanthus ilicifolius (M) | Antimicrobial Cytotoxic | [34] [35] | |
| 31 | Dicerandrol B | P. longicolla S1B4 Phomopsis sp. HNY29-2B | - b A. ilicifolius (M) | Antibacterial Cytotoxic | [34] [35] | |
| 32 | Dicerandrol C | P. longicolla S1B4 | - b | Antibacterial | [34] | |
| 33 | Deacetylphomoxanthone B | P. longicolla S1B4 Phomopsis sp. HNY29-2B | - b A. ilicifolius(M) | Antibacterial Cytotoxic | [34] [35] | |
| 34 | Penexanthone A | Phomopsis sp. HNY29-2B | A. ilicifolius (M) | Cytotoxic | [35] | |
| 35 | Chromones | (+)-Phomopsichin A | Phomopsis sp. 33# | R. stylosa (M) | Antimicrobial, Antioxidant, Inhibiting acetylcholinesterase and α-glucosidase | [33] |
| 36 | (−)-Phomopsichin B | Phomopsis sp. 33# | R. stylosa (M) | Antimicrobial, Antioxidant, Inhibiting acetylcholinesterase and α-glucosidase | [33] | |
| 37 | Phomopsichin C | Phomopsis sp. 33# | R. stylosa (M) | Antimicrobial, Antioxidant, Inhibiting acetylcholinesterase and α-glucosidase | [33] | |
| 38 | Phomopsichin D | Phomopsis sp. 33# | R. stylosa (M) | Antimicrobial, Antioxidant, Inhibiting acetylcholinesterase and α-glucosidase | [33] | |
| 39 | Chaetocyclinone B | Phomopsis sp. HNY29-2B | A. ilicifolius (M) | Cytotoxic | [36] | |
| 40 | Pestalotiopsone F | Phomopsis sp. IFB-ZS1-S4 | Scaevola hainanensis (M) | Inhibiting neuraminidase | [37] | |
| 41 | Phomoxanthone F | Phomopsis sp. xy21 | Xylocarpus granatum (M) | Anti-HIV | [38] | |
| 42 | 5-Hydroxy-3-hydroxymethyl-2-methyl-7-methoxychromone | Phomopsis sp. (No. Gx-4) | Sediment (M) | Cytotoxic, Inhibiting the growth of SIV branch | [39] | |
| 43 | Phomochromone A | Phomopsis sp. | Cistus monspeliensis (T) | Antimicrobial, Antialgal | [40] | |
| 44 | Phomochromone B | Phomopsis sp. | C. monspeliensis (T) | Antimicrobial, Antialgal | [40] | |
| 45 | Phomochromanone A | Phomopsis sp. CGMCC No. 5416 | Achyranthes bidentata (T) | Cytotoxic, Anti-HIV | [41] | |
| 46 | Phomochromanone B | Phomopsis sp. CGMCC No. 5416 | A. bidentata (T) | Cytotoxic, Anti-HIV | [41] | |
| 47 | 5-Hydroxy-6,8-dimethoxy-2-benzyl-4H-naphtho[2,3-b]-pyran-4-one | Phomopsis sp. ZSU-H26 | E. agallocha (M) | Cytotoxic | [42] | |
| 48 | Phomopsis-H76 A | Phomopsis sp. (#zsu-H76) | E. agallocha (M) | Accelerating the growth of SIV branch | [43] | |
| 49 | Chromanones | (3R,4S)-3,4-Dihydro-4,5,8-trihydroxy-3-methylisocoumarin | Phomopsis sp. (No. ZH-111) | Sediment (M) | Accelerating the growth of SIV branch, Cytotoxic | [44] |
| 50 | (3R,4S)-3,4-Dihydro-8-hydroxy-4-methoxy-3-methylisocoumarin | Phomopsis sp. (No. Gx-4) | Sediment (M) | Cytotoxic, Accelerating the growth of SIV branch | [39] | |
| 51 | 3,4-Dihydro-8-hydroxy-3-methyl-1H-2-benzopyran-1-one-5-carboxylic acid | Phomopsis sp. (No. Gx-4) | Sediment (M) | Cytotoxic, Accelerating the growth of SIV branch | [39] | |
| 52 | 5,8-Dihydroxy-4-methylcoumarin | Phomopsis sp. (No. Gx-4) | Sediment (M) | Cytotoxic, Inhibiting the growth of SIV branch | [39] | |
| 53 | (10S)-Diaporthin | Phomopsis sp. sh917 | Isodon eriocalyx var. laxiflora (T) | Antiangiogenic | [45] | |
| 54 | Cytosporone D | Phomopsis sp. CMU-LMA | Alpinia malacensis (T) | Antimicrobial, Inibiting DnaG primase | [46] | |
| 55 | Alternariol | Phomopsis sp. A240 Phomopsis sp. CAFT69 Phomopsis sp. | Taxus chinensis var. mairei (T) Endodesmia calophylloides (T) Senna spectabilis (T) | Cytotoxic Motility inhibitory and zoosporicidal potential Anti-inflammatory | [47] [48] [49] | |
| 56 | Alternariol-5-O-methyl ether | Phomopsis sp. CAFT69 | E. calophylloides (T) | Motility inhibitory and zoosporicidal potential | [48] | |
| 57 | 5ʹ-Hydroxyalternariol | Phomopsis sp. A240 Phomopsis sp. CAFT69 | T. chinensis var. mairei (T) E. calophylloides (T) | Antioxidant Motility inhibitory and zoosporicidal potential | [47] [48] | |
| 58 | Phomochromanone C | Phomopsis sp. CGMCC No. 5416 | A. bidentata (T) | Cytotoxic, Pro-apoptotic | [41] | |
| 59 | Benzofuranones | 7-Methoxy-6-methyl-3-oxo-1,3-dihydroisobenzofuran-4-carboxylic acid | Phomopsis sp. A123 | Kandelia candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] |
| 60 | Diaporthelactone | Phomopsis sp. A123 | K. candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] | |
| 61 | 7-Hydroxy-4,6-dimethy-3H-isobenzofuran-1-one | Phomopsis sp. A123 | K. candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] | |
| 62 | 7-Methoxy-4,6-dimethyl-3H-isobenzofuran-1-one | Phomopsis sp. A123 | K. candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] | |
| 63 | 4-(Hydroxymethyl)-7-methoxy-6-methyl-1(3H)-isobenzofuranone | Phomopsis sp. (No. ZH-111) | Sediment (M) | Inhibiting the growth of SIV branch, Cytotoxic | [44] | |
| 64 | Cytosporone E | Phomopsis sp. BCC 45011 | X. granatum(M) | Cytotoxic, Antimalarial | [51] | |
| 65 | Cytosporone P | Phomopsis sp. BCC 45011 | X. granatum (M) | Antimalarial | [51] | |
| 66 | Phomopsidone A | Phomopsis sp. A123 | K. candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] | |
| 67 | Excelsione | Phomopsis sp. A123 | K. candel (M) | Cytotoxic, Antifungal, Antioxidant | [50] | |
| 68 | Excelsional | Phomopsis sp. CAFT69 | E. calophylloides (T) | Motility inhibitory and zoosporicidal potential | [48] | |
| 69 | Lithocarol A | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 70 | Lithocarol B | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 71 | Lithocarol C | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 72 | Lithocarol D | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 73 | Lithocarol E | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 74 | Lithocarol F | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 75 | Isoprenylisobenzofuran A | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [52] | |
| 76 | 7-Methoxy-2-(4-methoxyphenyl)-3-methyl-5-(3-prenyl)-benzofuran | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 77 | 2-(4-Methoxyphenyl)-3-methyl-5-(3-prenyl)-benzofuran-7-ol | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 78 | 2-(4-Hydroxy-3,5-dimethoxyphenyl)-3-methyl-5-(3-prenyl) benzofuran-7-ol | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 79 | Moracin N | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 80 | 2-(2′-Methoxy-4′-hydroxy)-aryl-3-methy-6-hydroxybenzofuran | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 81 | Iteafuranal B | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 82 | Moracin P | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-TMV | [53] | |
| 83 | Pyrones | Phomaspyrone A | P. asparagi SWUKJ5.2020 | Kadsura angustifolia (T) | Cytotoxic | [54] |
| 84 | Macommelin-8,9-diol | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 85 | Phomaspyrone B | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 86 | Phomaspyrone C | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 87 | Phomaspyrone D | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 88 | Phomaspyrone E | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 89 | Macommelin-9-ol | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 90 | Macommelin | P. asparagi SWUKJ5.2020 | K. angustifolia (T) | Cytotoxic | [54] | |
| 91 | Pyrenocine J | Phomopsis sp. | Cistus salvifolius (T) | Antifungal, Antibacterial, Algicidal | [55] | |
| 92 | Pyrenocine K | Phomopsis sp. | C. salvifolius (T) | Antifungal, Antibacterial, Algicidal | [55] | |
| 93 | Pyrenocine L | Phomopsis sp. | C. salvifolius (T) | Antibacterial, Algicidal | [55] | |
| 94 | Pyrenocine M | Phomopsis sp. | C. salvifolius (T) | Antifungal, Antibacterial, Algicidal | [55] | |
| 95 | Phomopsis-H76 C | Phomopsis sp. (#zsu-H76) | E. agallocha (M) | Inhibiting the growth of SIV branch | [43] | |
| 96 | Quinones | Anhydrojavanicin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] |
| 97 | Dihydroanhydrojavanicin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 98 | Fusarubin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 99 | Javanicin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 100 | 2-Acetonyl-3methyl-5-hydroxy-7-methoxy-naphthazarin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 101 | Bostrycoidin | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 102 | Altersolanol B | P. longicolla HL-2232 | Bruguiera sexangula var. rhynchopetala (M) | Antibacterial | [57] | |
| 103 | Altersolanol A | Phomopsis sp. (PM0409092) P. foeniculi | Nyctanthes arbor-tristis (T) Foeniculum vulgare (T) | Cytotoxic Phytotoxic | [58] [59] | |
| 104 | (2R,3S)-7-Ethyl-1,2,3,4-tetrahydro-2,3,8-trihdroxy-6-methoxy-3-methyl-9,10-anthracenedione | Phomopsis sp. PSU-MA214 | Rhizophora apiculata (M) | Cytotoxic, Antibacterial | [60] | |
| 105 | Altersolanol J | P. foeniculi | F. vulgare (T) | Phytotoxic | [59] | |
| 106 | 2-Hydroxymethyl-4β,5α,6β-trihydroxycyclohex-2-en | Phomopsis sp. | Notobasis syriaca (T) | Antibacterial, Algicidal | [61] | |
| 107 | (−)-Phyllostine | Phomopsis sp. | N. syriaca (T) | Antifungal, Antibacterial, Algicidal | [61] | |
| 108 | (+)-Epiepoxydon | Phomopsis sp. | N. syriaca (T) | Antibacterial, Algicidal | [61] | |
| 109 | (+)-Epoxydon monoacetate | Phomopsis sp. | N. syriaca (T) | Antifungal, Antibacterial, Algicidal | [61] | |
| 110 | Phomonaphthalenone A | Phomopsis sp. HCCB04730 | Radix Stephaniae Japonicae (T) | Cytotoxic, Anti-HIV | [56] | |
| 111 | Ampelanol | Phomopsis sp. HNY29-2B | A. ilicifolius (M) | Antibacterial | [62] | |
| 112 | Phenols | Phomosine K | Phomopsis sp. | N. syriaca (T) | Antibacterial | [61] |
| 113 | Phomosine A | Phomopsis sp. | Ligustrum vulgare (T) | Antifungal, Antibacterial, Inhibiting algae | [63] | |
| 114 | Phomosine B | Phomopsis sp. | L. vulgare (T) | Antifungal, Antibacterial | [63] | |
| 115 | Phomosine C | Phomopsis sp. | L. vulgare (T) | Antifungal, Antibacterial | [63] | |
| 116 | Phomosine D | Phomopsis sp. | L. vulgare (T) | Antifungal, Inhibiting algae | [63] | |
| 117 | Phomosine I | Phomopsis sp. | L. vulgare (T) | Antifungal, Antibacterial | [63] | |
| 118 | 4-(3-Methoxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-(hydroxymethyl)phenol | P. asparagi | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [64] | |
| 119 | 4-(3-Hydroxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-(hydroxymethyl)phenol | P. asparagi | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [64] | |
| 120 | 4-(3-Methoxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-methylphenol | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [65] | |
| 121 | 4-(3-Hydroxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-methylphenol | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [65] | |
| 122 | 4-(3-Methoxy-5-methylphenoxy)-2-(3-hydroxypropyl)-6-methylphenol | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [65] | |
| 123 | 1-(4-(3-Methoxy-5-methylphenoxy)-2-methoxy-6-methylphenyl)-3-methylbut-3-en-2-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [66] | |
| 124 | 1-(4-(3-(Hydroxymethyl)-5methoxyphenoxy)-2-methoxy-6-methylphenyl)-3-methylbut-3-en-2-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [66] | |
| 125 | 1-(4-(3-Hydroxy-5(hydroxymethyl)phenoxy)-2-methoxy-6-methylphenyl)-3-methylbut-3-en-2-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [66] | |
| 126 | 1-[2-Methoxy-4-(3-methoxy-5-methylphenoxy)-6-methylphenyl]-ethanone | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [67] | |
| 127 | 1-[4-(3-(Hydroxymethyl)-5-methoxyphenoxy)-2-methoxy-6-methylphenyl]-ethanone | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [67] | |
| 128 | 3-Hydroxy-1-(1,8-dihydroxy-3,6-dimethoxynaphthalen-2-yl)propan-1-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [68] | |
| 129 | 3-Hydroxy-1-(1,3,8-trihydroxy-6-methoxynaphthalen-2-yl)propan-1-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [68] | |
| 130 | 3-Hydroxy-1-(1,8-dihydroxy-3,5-dimethoxynaphthalen-2-yl)propan-1-one | P. fukushii | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [68] | |
| 131 | 5-Methoxy-2-methyl-7-(3-methyl-2-oxobut-3-enyl)-1-naphthaldehyde | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [69] | |
| 132 | 2-(Hydroxymethyl)-5-methoxy-7-(3-methyl-2-oxobut-3-enyl)-1-naphthaldehyde | Phomopsis sp. | P. polyphylla var. yunnanensis (T) | Anti-MRSA | [69] | |
| 133 | Tenellone H | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [70] | |
| 134 | 16-Acetoxycytosporone B | Phomopsis sp. YM 355364 | A. carmichaeli (T) | Antifungal | [71] | |
| 135 | Cytosporone B | Phomopsis sp. 0391 Phomopsis sp. PSU-H188 | P. polyphylla var. yunnanensis (T) Hevea brasiliensis (T) | Inhibiting lipase Protecting pancreatic β-cells | [72] [73] | |
| 136 | Dothiorelone A | Phomopsis sp. 0391 | P. polyphylla var. yunnanensis (T) | Inhibiting lipase | [72] | |
| 137 | Lithocarpinol A | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [74] | |
| 138 | Lithocarpinol B | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [74] | |
| 139 | Phomoindene A | Phomopsis sp. (No. GX7-4A) | Sediment (M) | Cytotoxic | [75] | |
| 140 | 4-Hydroxybenzaldehyde | Phomopsis sp. YM 355364 | A. carmichaelii (T) | Antimicrobial | [26] | |
| 141 | 5,5′-Dimethoxybiphenyl-2,2′-diol | P. longicolla HL-2232 | B. sexangula var. rhynchopetala (M) | Antibacterial | [57] | |
| 142 | Phomonitroester | Phomopsis sp. PSU-MA214 | R. apiculate (M) | Cytotoxic | [60] | |
| 143 | Cytosporone U | Phomopsis sp. FJBR-11 | Brucea javanica (T) | Anti-TMV | [76] | |
| 144 | Altenusin | Phomopsis sp. CAFT69 | E. calophylloides (T) | Motility inhibitory and zoosporicidal potential | [48] | |
| 145 | Cosmochlorin D | Phomopsis sp. N-125 | Ficus ampelas (T) | Cytotoxic, Growth-inhibition activity | [77] | |
| 146 | Cosmochlorin E | Phomopsis sp. N-125 | F. ampelas (T) | Cytotoxic, Growth-inhibition activity | [77] | |
| 147 | Oblongolides | Oblongolide Z | Phomopsis sp. BCC 9789 | Musa acuminate (T) | Cytotoxic, Anti-HSV-1 | [78] |
| 148 | Oblongolide Y | Phomopsis sp. BCC 9789 | M. acuminate (T) | Cytotoxic | [78] | |
| 149 | Oblongolide C1 | Phomopsis sp. XZ-01 | Camptotheca acuminate (T) | Cytotoxic | [79] | |
| 150 | Oblongolide P1 | Phomopsis sp. XZ-01 | C. acuminate (T) | Cytotoxic | [79] | |
| 151 | Oblongolide X1 | Phomopsis sp. XZ-01 | C. acuminate (T) | Cytotoxic | [79] | |
| 152 | 6-Hydroxyphomodiol | Phomopsis sp. XZ-01 | C. acuminate (T) | Cytotoxic | [79] | |
| 153 | Oblongolide C | Phomopsis sp. XZ-01 | C. acuminate (T) | Cytotoxic | [79] | |
| 154 | 2-Deoxy-4α-hydroxyoblongolide X | Phomopsis sp. BCC 9789 | M. acuminate (T) | Anti-HSV-1 | [78] | |
| 155 | Unclassified polyketides | Phomoxydiene C | Phomopsis sp. BCC 45011 | X. granatum (M) | Cytotoxic, Antimalarial | [51] |
| 156 | 1893 A | Phomopsis sp. BCC 45011 | X. granatum (M) | Cytotoxic | [51] | |
| 157 | Mycoepoxydiene | Phomopsis sp. BCC 45011 | X. granatum (M) | Cytotoxic, Antimalarial | [51] | |
| 158 | Deacetylmycoepoxydiene | Phomopsis sp. BCC 45011 | X. granatum (M) | Cytotoxic, Antimalarial | [51] | |
| 159 | Phomoxydiene A | Phomopsis sp. BCC 45011 | X. granatum (M) | Cytotoxic, Antimalarial | [51] | |
| 160 | Phomopoxide A | Phomopsis sp. YE3250 | Paeonia delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 161 | Phomopoxide B | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 162 | Phomopoxide C | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 163 | Phomopoxide D | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 164 | Phomopoxide E | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 165 | Phomopoxide F | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 166 | Phomopoxide G | Phomopsis sp. YE3250 | P. delavayi (T) | Cytotoxic, Antifungal, Inhibiting α-glycosidase | [80] | |
| 167 | Phomentrioloxin | Phomopsis sp. | Carthamus lanatus (T) | Phytotoxic | [81] | |
| 168 | Phomotenone | Phomopsis sp. | C. monspeliensis (T) | Antifungal, Antibacterial, Antialgal | [40] | |
| 169 | Phomopsolide B | Phomopsis sp. DC275 | Vitis vinifera (T) | Antibacterial, Phytotoxic | [82] | |
| 170 | Phomopsolidone A | Phomopsis sp. DC275 | V. vinifera (T) | Antibacterial, Phytotoxic | [82] | |
| 171 | Phomopsolidone B | Phomopsis sp. DC275 | V. vinifera (T) | Antibacterial, Phytotoxic | [82] | |
| 172 | Monoterpenoids | Acropyrone | Phomopsis sp. HNY29-2B | A. ilicifolius (M) | Antibacterial | [62] |
| 173 | Nectriapyrone | P. foeniculi | F. vulgare (T) | Phytotoxic | [59] | |
| 174 | (1S,2S,4S)-Trihydroxy-p-menthane | Phomopsis sp. | C. monspeliensis (T) | Antibacterial, Antialgal | [40] | |
| 175 | Sesquiterpenoids | Phomophyllin A | Phomopsis sp. TJ507A | Phyllanthus glaucus (T) | Inhibiting BACE1 | [83] |
| 176 | Phomophyllin B | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 177 | Phomophyllin C | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 178 | Phomophyllin D | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 179 | Phomophyllin E | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 180 | Phomophyllin F | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 181 | Phomophyllin G | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 182 | Radulone B | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 183 | Phomophyllin I | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 184 | Onitin | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 185 | (7R,9S,10R)-3,9-Di-hidroxicalamenene | P. cassiae | Cassia spectabilis (T) | Inhibiting acetylcholinesterase, Antifungal | [84] | |
| 186 | (7R,9R,10R)-3,9-Di-hidroxicalamenene | P. cassiae | C. spectabilis (T) | Inhibiting acetylcholinesterase, Antifungal | [84] | |
| 187 | (7S,10R)-3-Hidroxicalamen-8-one | P. cassiae | C. spectabilis (T) | Inhibiting acetylcholinesterase, Antifungal | [84] | |
| 188 | Aristelegone-A | P. cassiae | C. spectabilis (T) | Inhibiting acetylcholinesterase, Antifungal | [84] | |
| 189 | Phomoarcherin A | P. archeri | Vanilla albidia (T) | Cytotoxic | [85] | |
| 190 | Phomoarcherin B | P. archeri | V. albidia (T) | Cytotoxic, Antimalarial | [85] | |
| 191 | Phomoarcherin C | P. archeri | V. albidia (T) | Cytotoxic | [85] | |
| 192 | Kampanol A | P. archeri | V. albidia (T) | Cytotoxic | [85] | |
| 193 | (+)-S-1-Methyl-abscisic-6-acid | P. amygdali | Call midge (T) | Antibacterial | [86] | |
| 194 | (+)-S-Abscisic acid | P. amygdali | C. midge (T) | Antibacterial | [86] | |
| 195 | 7-Hydroxy-10-oxodehydrodihydrobotrydial | Phomopsis sp. TJ507A | P. glaucus (T) | Inhibiting BACE1 | [83] | |
| 196 | Curcumol | P. castaneae-mollissimae GQH87 | Artemisia annua (T) | Cytotoxic | [87] | |
| 197 | 9-Hydroxyphomopsidin | Phomopsis sp. CAFT69 | E. calophylloides (T) | Motility inhibitory and zoosporicidal potential | [48] | |
| 198 | Phomopsidin | Phomopsis sp. CAFT69 | E. calophylloides (T) | Motility inhibitory and zoosporicidal potential | [48] | |
| 199 | AA03390 | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [70] | |
| 200 | Diterpenoids | Libertellenone J | Phomopsis sp. S12 | Illigera rhodantha (T) | Anti-inflammatory | [88] |
| 201 | Libertellenone C | Phomopsis sp. S12 | - b | Anti-inflammatory | [89] | |
| 202 | Libertellenone T | Phomopsis sp. S12 | - b | Anti-inflammatory | [89] | |
| 203 | Pedinophyllol K | Phomopsis sp. S12 | - b | Anti-inflammatory | [89] | |
| 204 | Pedinophyllol L | Phomopsis sp. S12 | - b | Anti-inflammatory | [89] | |
| 205 | Fusicoccin J | P. amygdali | C. midge (T) | Antibacterial | [86] | |
| 206 | 3α-Hydroxyfusicoccin J | P. amygdali | C. midge (T) | Antibacterial | [86] | |
| 207 | Triterpenoids | 3S,22R,26-Trihydroxy-8,24E-euphadien-11-one | P. chimonanthi | Tamarix chinensis (T) | Cytotoxic | [90] |
| 208 | Betulinic acid | Phomopsis sp. SNB-LAP1-7-32 | Diospyros carbonaria (T) | Antiviral, Cytotoxic | [91] | |
| 209 | Oleanolic acid | P. castaneae-mollissi mae GQH87 | A. annua (T) | Cytotoxic | [87] | |
| 210 | Steroids | (14β,22E)-9,14-Dihydroxyergosta-4,7,22-triene-3,6-dione | Phomopsis sp. | A. carmichaeli (T) | Antifungal | [92] |
| 211 | (5α,6β,15β,22E)-6-Ethoxy-5,15-dihydroxyergosta-7,22-dien-3-one | Phomopsis sp. | A. carmichaeli (T) | Antifungal | [92] | |
| 212 | Calvasterol A | Phomopsis sp. | A. carmichaeli (T) | Antifungal | [92] | |
| 213 | Calvasterol B | Phomopsis sp. | A. carmichaeli (T) | Antifungal | [92] | |
| 214 | Ganodermaside D | Phomopsis sp. | A. carmichaeli (T) | Antifungal | [92] | |
| 215 | Dankasterone A | Phomopsis sp. YM 355364 | A. carmichaeli (T) | Antifungal, Anti-influenza | [71] | |
| 216 | 3β,5α,9α-Trihydroxy-(22E,24R)-ergosta-7,22-dien-6-one | Phomopsis sp. YM 355364 | A. carmichaeli (T) | Antifungal | [71] | |
| 217 | Phomopsterone B | Phomopsis sp. TJ507A | P. glaucus (T) | Anti-inflammatory | [93] | |
| 218 | Cyathisterol | Phomopsis sp. YM 355364 | A. carmichaelii (T) | Antifungal | [26] | |
| 219 | Macrolides | Sch-642305 | Phomopsis sp. CMU-LMA | Alpinia malaccensis (T) | Cytotoxic, Antimicrobial | [94] |
| 220 | LMA-P1 | Phomopsis sp. CMU-LMA | A. malaccensis (T) | Cytotoxic | [94] | |
| 221 | Benquoine | Phomopsis sp. CMU-LMA | A. malaccensis (T) | Cytotoxic, Antimicrobial | [94] | |
| 222 | Aspergillide C | Phomopsis sp. IFB-ZS1-S4 | S. hainanensis (M) | Inhibiting neuraminidase | [37] | |
| 223 | Lithocarpin A | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [95] | |
| 224 | Lithocarpin B | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [95] | |
| 225 | Lithocarpin C | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [95] | |
| 226 | Lithocarpin D | P. lithocarpus FS508 | Sediment (M) | Cytotoxic | [95] | |
| 227 | Alkaloids | Phomopchalasin B | Phomopsis sp. shj2 | I. eriocalyx var. laxiflora (T) | Antimigratory | [96] |
| 228 | Phomopsichalasin G | P. spp. xy21 and xy22 | X. granatum (M) | Cytotoxic | [97] | |
| 229 | 18-Metoxycytochalasin J | Phomopsis sp. | Garcinia kola (T) | Cytotoxic, Antibacterial | [98] | |
| 230 | Cytochalasin H | Phomopsis sp. Phomopsis sp. By254 Phomopsis sp. | G. kola (T) Gossypium hirsutum (T) S. spectabilis (T) | Cytotoxic, Antibacterial Antifungal Inhibiting acetylcholinesterase, Anti-inflammatory | [98] [99] [49] | |
| 231 | Cytochalasin J | Phomopsis sp. Phomopsis sp. P. asparagi | G. kola (T) S. spectabilis (T) Peperomia sui (T) | Cytotoxic, Antibacterial Anti-inflammatory Antiandrogen | [98] [49] [100] | |
| 232 | Phomopchalasin C | Phomopsis sp. shj2 | I. eriocalyx var. laxiflora (T) | Cytotoxic, Anti-inflammatory, Antimigratory | [96] | |
| 233 | Cytochalasin N | Phomopsis sp. By254 | G. hirsutum (T) | Antifungal | [99] | |
| 234 | Epoxycytochalasin H | Phomopsis sp. By254 | G. hirsutum (T) | Antifungal | [99] | |
| 235 | Diaporthalasin | Phomopsis sp. PSU-H188 | H. brasiliensis (T) | Anti-MRSA | [73] | |
| 236 | (+)-Tersone E | P. tersa FS441 | Sediment (M) | Antibacterial, Cytotoxic | [101] | |
| 237 | ent-Citridone A | P. tersa FS441 | Sediment (M) | Antibacterial | [101] | |
| 238 | Phochrodine C | Phomopsis sp. 33# | R. stylosa (M) | Anti-inflammatory | [102] | |
| 239 | Phochrodine D | Phomopsis sp. 33# | R. stylosa (M) | Anti-inflammatory, Antioxidant | [102] | |
| 240 | PM181110 | P. glabrae | Pongamia pinnata (T) | Anticancer | [103] | |
| 241 | Fusaristatin A | P. longicolla S1B4 | - b | Antibacterial | [34] | |
| 242 | Exumolide A | Phomopsis sp. (No. ZH-111) | Sediment (M) | Accelerating the growth of SIV branch, Cytotoxic | [44] | |
| 243 | Flavonoids | Quercetin | P. castaneae-mollissimae GQH87 | A. annua (T) | Cytotoxic | [87] |
| 244 | Luteolin | P. castaneae-mollissimae GQH87 | A. annua (T) | Cytotoxic | [87] | |
| 245 | Naringenin | P. castaneae-mollissimae GQH87 | A. annua (T) | Cytotoxic | [87] | |
| 246 | Luteolin-7-O-glucoside | P. castaneae-mollissimae GQH87 | A. annua (T) | Cytotoxic | [87] |
| Number | Structural Types | Compounds | Strains | Habitats (T/M a) | Activities | Refs. |
|---|---|---|---|---|---|---|
| 247 | Xanthones | 3,8-Dihydroxy-6-methyl-9-oxo-9H-xanthene-1-carboxylate | Diaporthe sp. SCSIO 41011 | Rhizophora stylosa (M) | Anti-IAV | [125] |
| 28 | Phomoxanthone A | Diaporthe sp. GZU-1021 D. phaseolorum FS431 | Chiromanteshae- matochir (M)Sediment (M) | Anti-inflammatory Cytotoxic | [126] [127] | |
| 248 | Chromones | Penialidin A | Diaporthe sp. GZU-1021 | Chiromanteshae matochir (M) | Anti-inflammatory | [126] |
| 35 | (+)-Phomopsichin A | D. phaseolorum SKS019 | Acanthus ilicifolius (M) | Inhibitory effects on osteoclastogenesis | [128] | |
| 249 | (−)-Phomopsichin A | D. phaseolorum SKS019 | A. ilicifolius (M) | Inhibitory effects on osteoclastogenesis | [128] | |
| 250 | (+)-Phomopsichin B | D. phaseolorum SKS019 | A. ilicifolius (M) | Inhibitory effects on osteoclastogenesis | [128] | |
| 36 | (−)-Phomopsichin B | D. phaseolorum SKS019Diaporthe sp. GZU-1021 | A. ilicifolius (M) Chiromateshaem atochir (M) | Inhibitory effects on osteoclastogenesis Anti-inflammatory | [128] [126] | |
| 251 | Diaporchromanone C | D. phaseolorum SKS019 | A. ilicifolius (M) | Inhibitory effects on osteoclastogenesis | [128] | |
| 252 | Diaporchromanone D | D. phaseolorum SKS019 | A. ilicifolius (M) | Inhibitory effects on osteoclastogenesis | [128] | |
| 40 | Pestalotiopsone F | Diaporthe sp. SCSIO 41011 | R. stylosa (M) | Anti-IAV | [125] | |
| 253 | Pestalotiopsone B | Diaporthe sp. SCSIO 41011 D. pseudomangiferaea | R. stylosa (M) Tylophora ouata (T) | Anti-IAV Antifibrotic | [125] [129] | |
| 254 | Diaportheone A | Diaporthe sp. P133 | Pandanus amaryllifolius (T) | Antitubercular | [130] | |
| 255 | Diaportheone B | Diaporthe sp. P133 | P. amaryllifolius (T) | Antitubercular | [130] | |
| 53 | Chromanones | (10S)-Diaporthin | D. terebinthifolii LGMF907 | Schinus terebinthifolius (T) | Antibacterial | [131] |
| 256 | Orthosporin | D. terebinthifolii LGMF907 | S. terebinthifolius (T) | Antibacterial | [131] | |
| 54 | Cytosporone D | D. pseudomangiferaea | T. ouata (T) | Cytotoxic, Antioxidant Antidiabetic | [129] | |
| 257 | Mucorisocoumarin A | D. pseudomangiferaea | T. ouata (T) | Antifibrotic | [129] | |
| 258 | 3,4-Dihydro-8-hydroxy-3,5-dimethyl-isocoumarin | D. eres | Hedera helix (T) | Phytotoxic | [132] | |
| 259 | Diportharine A | Diaporthe sp. | Datura inoxia (T) | Antioxidant | [133] | |
| 260 | Furanones | (1R,2E,4S,5R)-1-[(2R)-5-Oxotetrahydrofuran-2-yl]-4,5-dihydroxy-hex-2-en-1-yl(2E)-2-methylbut-2-enoate | Diaporthe sp. SXZ-19 | Camptotheca acuminate (T) | Cytotoxic | [134] |
| 261 | Butyl 5-[(1R)-1-hydroxyethyl]-γ-oxofuran-2-butanoate | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 262 | 3,4-Dihydro-5ʹ-[(1R)-1-hydroxyethyl] [2,2ʹ-bifuran]-5(2H)-one | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 263 | 3,4-Dihydro-5ʹ-[(1R)-1-hydroxymethylethyl][2,2ʹ-bifuran]-5(2H)-one | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 264 | Kongiidiazadione | D. Kongii | Carthamus lanatus (T) | Phytotoxic, Antibacterial | [135] | |
| 265 | Pyrones | Phomopsolide A | D. maritima | Picea mariana(T) Picea rubens (T) | Antifungal, Antibiotic | [136] |
| 169 | Phomopsolide B | D. maritima | P. mariana (T) P. rubens (T) | Antifungal, Antibiotic | [136] | |
| 266 | Phomopsolide C | D. maritima | P. mariana (T)P. rubens (T) | Antifungal, Antibiotic | [136] | |
| 267 | (S,E)-6-(4-Hydroxy-3-oxopent-1-en-1-yl)-2H-pyran-2-one | D. maritima | P. mariana (T) P. rubens (T) | Antifungal, Antibiotic | [136] | |
| 268 | 7-Hydroxy-6-metoxycoumarin | D. lithocarpus | Artocarpus heterophyllus (T) | Antifungal | [137] | |
| 269 | Coumarin | D. lithocarpus | A. heterophyllus (T) | Antibacterial | [137] | |
| 270 | Quinones | Phyllostine acetate | D. miriciae | Cyperus iria (T) | Antifeedant, Contact toxicity, Oviposition deterrent activities | [138] |
| 107 | (−)-Phyllostine | D. miriciae | C. iria (T) | Antifeedant, Contact toxicity, Oviposition deterrent activities | [138] | |
| 271 | Biatriosporin N | Diaporthe sp. GZU-1021 | Chiromanteshae- matochir (M) | Anti-inflammatory | [126] | |
| 272 | Emodin | D. lithocarpus | A. heterophyllus (T) | Cytotoxic, Antibacterial | [137] | |
| 273 | 1,2,8-Trihydroxyanthraquinone | D. lithocarpus | A. heterophyllus (T) | Antibacterial | [137] | |
| 274 | (+)-2,2′-Epicytoskyrin A | Diaporthe sp. GNBP-10 | Uncaria gambir Roxb (T) | Antifungal | [139] | |
| 275 | Cytoskyrin C | Diaporthe sp. | Anoectochilus roxburghii (T) | Cytotoxic, Activating the NF-κB pathway | [140] | |
| 276 | (+)-Epicytoskyrin | Diaporthe sp. | A. roxburghii (T) | Cytotoxic, Activating the NF-κB pathway | [140] | |
| 277 | Phenols | Tyrosol | D. helianthin D. eres | Luehea divaricate (T) Vitis vinifera (T) | Antagonistic Phytotoxic | [141] [142] |
| 278 | 2,5-Dihydroxybenzyl alcohol | D. vochysiae LGMF1583 | Vochysia divergens (T) | Cytotoxic | [143] | |
| 140 | 4-Hydroxybenzaldehyde | D. eres | V. vinifera (T) | Phytotoxic | [142] | |
| 279 | p-Cresol | D. eres | V. vinifera (T) | Phytotoxic | [142] | |
| 280 | 4-Hydroxybenzoic acid | D. eres | V. vinifera (T) | Phytotoxic | [142] | |
| 281 | Arbutin | D. lithocarpus | A. heterophyllus (T) | Cytotoxic | [137] | |
| 113 | Phomosine A | Diaporthe sp. F2934 | Siparuna gesnerioides (T) | Antibacterial | [144] | |
| 115 | Phomosine C | Diaporthe sp. F2934 | S. gesnerioides (T) | Antibacterial | [144] | |
| 282 | Flavomannin-6,6′-di-O-methyl ether | D. melonis | Annona squamosal (T) | Antimicrobial | [145] | |
| 283 | Acetoxydothiorelone B | D. pseudomangiferaea | T. ouata (T) | Antifibrotic | [129] | |
| 284 | Dothiorelone B | D. pseudomangiferaea | T. ouata (T) | Antifibrotic | [129] | |
| 285 | Dothiorelone L | D. pseudomangiferaea | T. ouata (T) | Antifibrotic | [129] | |
| 286 | Dothiorelone G | D. pseudomangiferaea | T. ouata (T) | Antifibrotic | [129] | |
| 287 | Diaporthol A | Diaporthe sp. ECN-137 | Phellodendron amurense (T) | Anti-migration | [146] | |
| 288 | Diaporthol B | Diaporthe sp. ECN-137 | P. amurense (T) | Anti-migration | [146] | |
| 289 | Tenellone C | Diaporthe sp. SYSU-HQ3 | Excoecaria agallocha (M) | MptpB inhibitory | [147] | |
| 290 | Tenellone D | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 291 | Diaporindene A | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 292 | Diaporindene B | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 293 | Diaporindene C | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 294 | Diaporindene D | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 75 | Isoprenylisobenzofuran A | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 295 | Oblongolides | Oblongolide D | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] |
| 296 | Oblongolide H | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 297 | Oblongolide P | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 298 | Oblongolide V | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] | |
| 299 | Unclassified polyketides | Phomentrioloxin B | D. gulyae | C. lanatus (T) | Phytotoxic | [149] |
| 300 | epi-Isochromophilone II | Diaporthe sp. SCSIO 41011 | R. stylosa (M) | Cytotoxic | [150] | |
| 301 | Isochromophilone D | Diaporthe sp. SCSIO 41011 | R. stylosa (M) | Cytotoxic | [150] | |
| 302 | Monoterpenoids | (1R,2R,4R)-Trihydroxy-p-menthane | Diaporthe sp. SXZ-19 | C. acuminate (T) | Cytotoxic | [134] |
| 303 | Gulypyrone A | D. gulyae | C. lanatus (T) | Phytotoxic | [149] | |
| 304 | Gulypyrone B | D. gulyae | C. lanatus (T) | Phytotoxic | [149] | |
| 173 | Nectriapyrone | D. Kongii | C. lanatus (T) | Phytotoxic | [135] | |
| 305 | Sesquiterpenoids | Diaporol R | Diaporthe sp. | R. stylosa (M) | Cytotoxic | [151] |
| 306 | Eremofortin F | Diaporthe sp. SNB-GSS10 | Sabicea cinerea (T) | Cytotoxic | [152] | |
| 307 | Lithocarin B | D. lithocarpus A740 | Morinda officinalis (T) | Cytotoxic | [153] | |
| 308 | Lithocarin C | D. lithocarpus A740 | M. officinalis (T) | Cytotoxic | [153] | |
| 309 | Triterpenoids | 19-Nor-lanosta-5(10),6,8,24-tetraene-1α,3β,12β,22S-tetraol | Diaporthe sp. LG23 | Mahonia fortunei (T) | Antibacterial | [154] |
| 216 | Steriods | 3β,5α,9α-Trihydroxy-(22E,24R)-ergosta-7,22-dien-6-one | Diaporthe sp. LG23 | M. fortunei (T) | Antibacterial | [154] |
| 310 | Chaxine C | Diaporthe sp. LG23 | M. fortunei (T) | Antibacterial | [154] | |
| 311 | Ten-membered lactones | Phomolide C | Diaporthe sp. | Aucuba japonica var. borealis (T) | Inhibitory of proliferation of human colon adenocarcinoma cells | [155] |
| 312 | Xylarolide | D. terebinthifolii | Glycyrrhiza glabra (T) | Antimicrobial, Cytotoxic | [156] | |
| 313 | Phomolide G | D. terebinthifolii | G. glabra (T) | Antibacterial | [156] | |
| 314 | Xylarolide A | Diaporthe sp. | D. inoxia (T) | Cytotoxic, Antioxidant | [133] | |
| 315 | Alkaloids | 18-Des-hydroxy cytochalasin H | D. phaseolorum-92C | Combretum lanceolatum (T) | Inhibiting leishmanicidal, Antioxidant, Cytotoxic | [157] |
| 316 | 21-Acetoxycytochalasin J2 | Diaporthe sp. GDG-118 | Sophora tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 317 | 21-Acetoxycytochalasin J3 | Diaporthe sp. GDG-118 | S. tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 318 | Cytochalasin J3 | Diaporthe sp. GDG-118 | S. tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 230 | Cytochalasin H | Diaporthe sp. GDG-118 Diaporthe sp. GZU-1021 | S. tonkinensis (T) Chiromanteshae matochir (M) | Antifungal, AntibacterialAnti-inflammatory | [158] [126] | |
| 319 | 7-Acetoxycytochalasin H | Diaporthe sp. GDG-118 | S. tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 231 | Cytochalasin J | Diaporthe sp. GDG-118 | S. tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 320 | Cytochalasin E | Diaporthe sp. GDG-118 | S. tonkinensis (T) | Antifungal, Antibacterial | [158] | |
| 321 | 21-O-Deacetyl-L-696,474 | Diaporthe sp. GZU-1021 | Chiromanteshae matochir (M) | Anti-inflammatory | [126] | |
| 322 | Cordysinin A | D. arecae | Kandelia obovate (M) | Anti-angiogenic | [159] | |
| 323 | 5-Deoxybostrycoidin | D. phaseolorum SKS019 | A. ilicifolius (M) | Cytotoxic | [160] | |
| 241 | Fusaristatin A | D. phaseolorum SKS019 | A. ilicifolius (M) | Cytotoxic | [160] | |
| 324 | Vochysiamide B | D. vochysiae LGMF1583 | V. divergens (T) | Antibacterial, Cytotoxic | [143] | |
| 325 | Diaporisoindole A | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 326 | Diaporisoindole B | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 327 | Diaporisoindole D | Diaporthe sp. SYSU-HQ3 Diaporthe sp. SYSU-HQ3 | E. agallocha (M) E. agallocha (M) | Anti-inflammatory MptpB inhibitory | [148] [147] | |
| 328 | Diaporisoindole E | Diaporthe sp. SYSU-HQ3 | E. agallocha (M) | Anti-inflammatory | [148] | |
| 329 | Phomopsin F | D. toxica | -b | Cytotoxic | [161] | |
| 330 | Fatty acids | 3-Hydroxypropionic acid | D. phaseolorum | Laguncularia racemose (M) | Antimicrobial | [162] |
| 331 | 3-Nitropropionic acid | D. gulyae | C. lanatus (T) | Phytotoxic | [149] | |
| 332 | Diapolic acid A | D. terebinthifolii | G. glabra (T) | Antibacterial | [156] | |
| 333 | Diapolic acid B | D. terebinthifolii | G. glabra (T) | Antibacterial | [156] | |
| 334 | Diaporthsin E | Diaporthe sp. JC-J7 | Dendrobium nobile (T) | Antihyperlipidemic | [163] | |
| 335 | 3-Hydroxy-5-methoxyhex-5-ene-2,4-dione | Diaporthe sp. ED2 | Orthosiphon stamieus (T) | Antifungal | [164] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, T.-C.; Lu, Y.-H.; Wang, J.-F.; Song, Z.-Q.; Hou, Y.-G.; Liu, S.-S.; Liu, C.-S.; Wu, S.-H. Bioactive Secondary Metabolites of the Genus Diaporthe and Anamorph Phomopsis from Terrestrial and Marine Habitats and Endophytes: 2010–2019. Microorganisms 2021, 9, 217. https://doi.org/10.3390/microorganisms9020217
Xu T-C, Lu Y-H, Wang J-F, Song Z-Q, Hou Y-G, Liu S-S, Liu C-S, Wu S-H. Bioactive Secondary Metabolites of the Genus Diaporthe and Anamorph Phomopsis from Terrestrial and Marine Habitats and Endophytes: 2010–2019. Microorganisms. 2021; 9(2):217. https://doi.org/10.3390/microorganisms9020217
Chicago/Turabian StyleXu, Tang-Chang, Yi-Han Lu, Jun-Fei Wang, Zhi-Qiang Song, Ya-Ge Hou, Si-Si Liu, Chuan-Sheng Liu, and Shao-Hua Wu. 2021. "Bioactive Secondary Metabolites of the Genus Diaporthe and Anamorph Phomopsis from Terrestrial and Marine Habitats and Endophytes: 2010–2019" Microorganisms 9, no. 2: 217. https://doi.org/10.3390/microorganisms9020217