Diet Composition of the Wild Stump-Tailed Macaque (Macaca arctoides) in Perlis State Park, Peninsular Malaysia, Using a Chloroplast tRNL DNA Metabarcoding Approach: A Preliminary Study

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
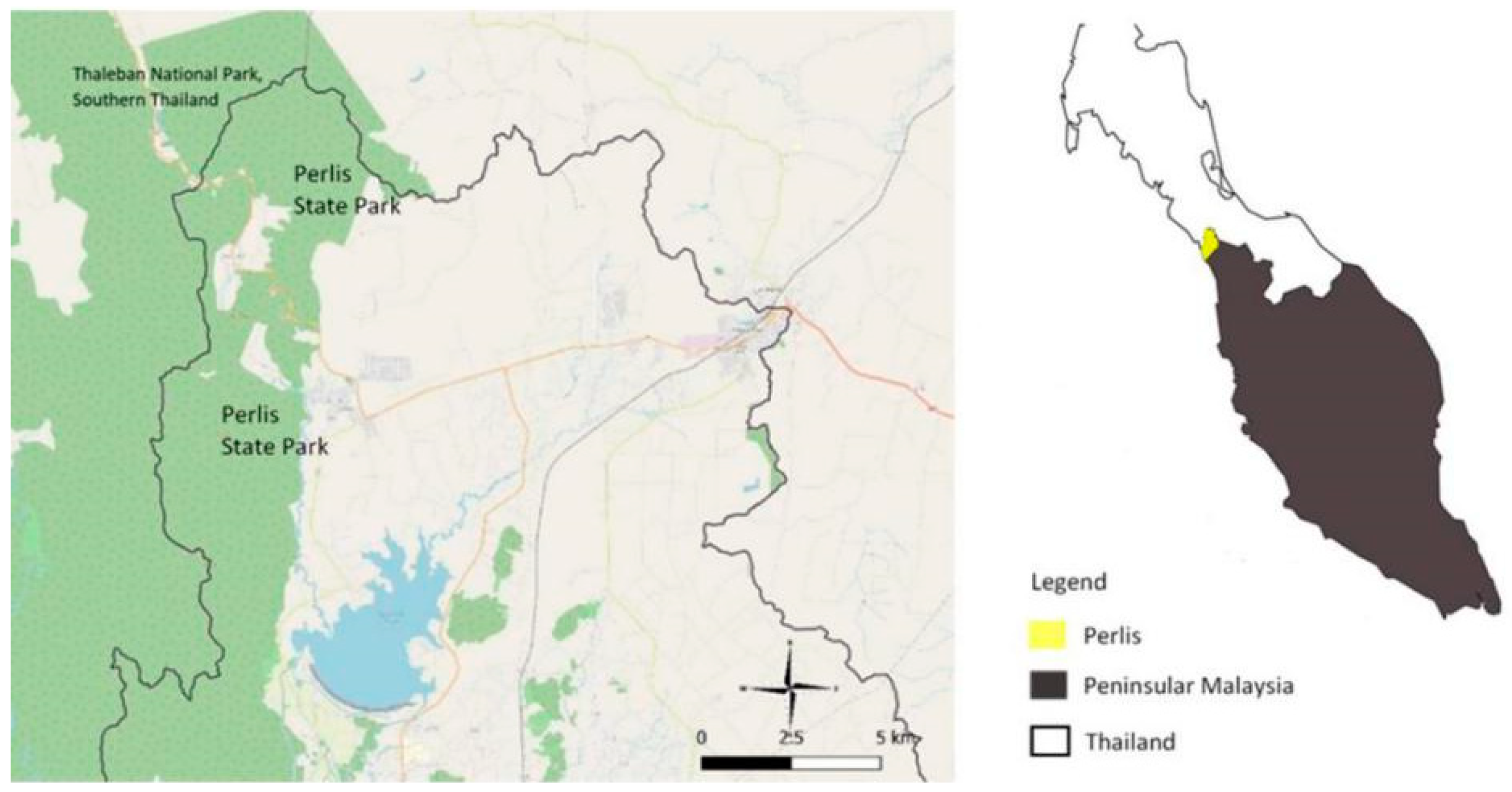
2.1. Fecal Sample Collection
2.2. DNA Extraction
2.3. PCR Amplification
2.4. Illumina MiniSeq-DNA Sequencing of tRNL Gene
2.5. Statistical Analysis
3. Results
3.1. NGS Data Analysis and Sequence Filtering
3.2. Diet Richness and Composition of the Wild M. Arctoides in PSP
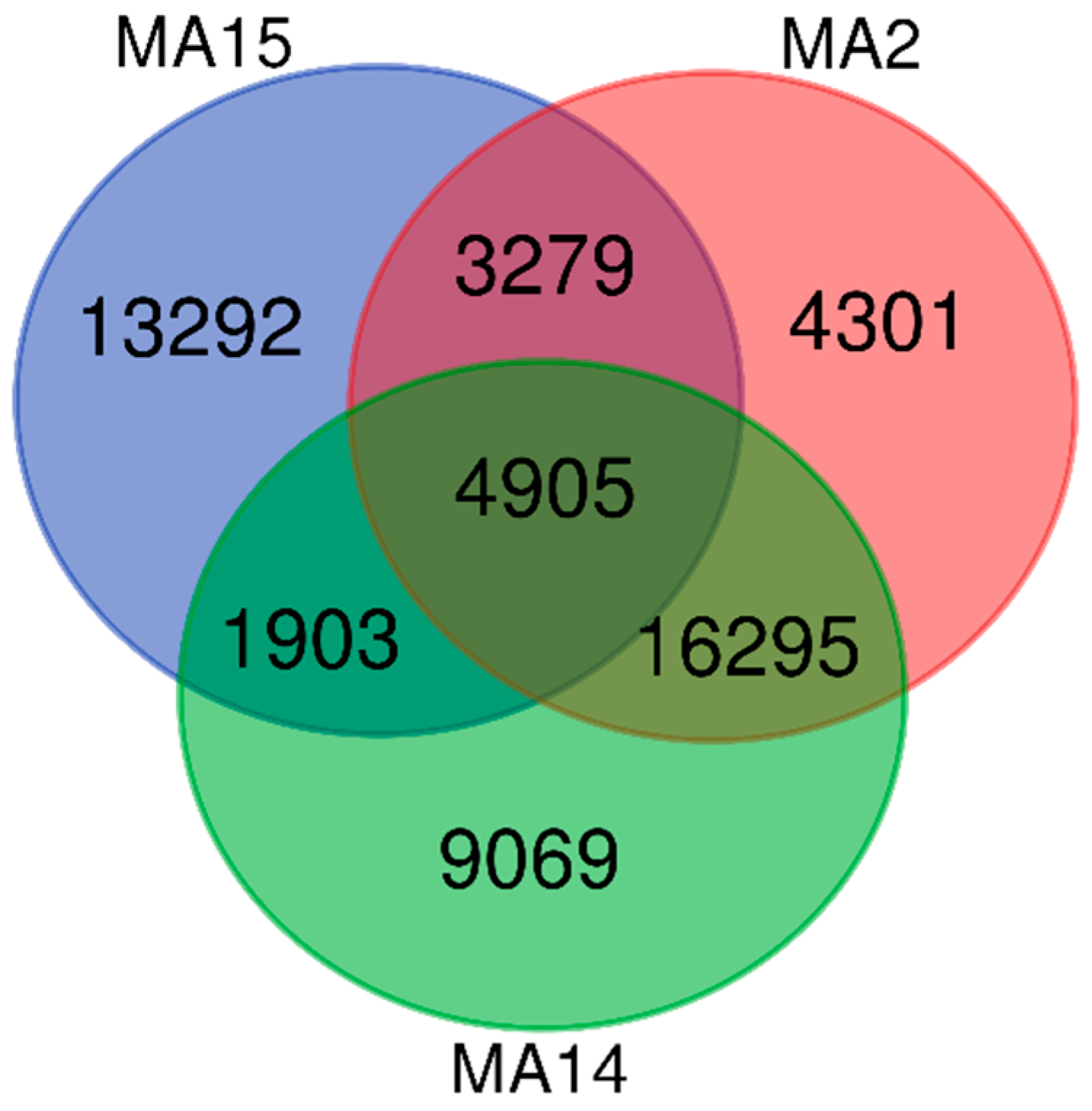
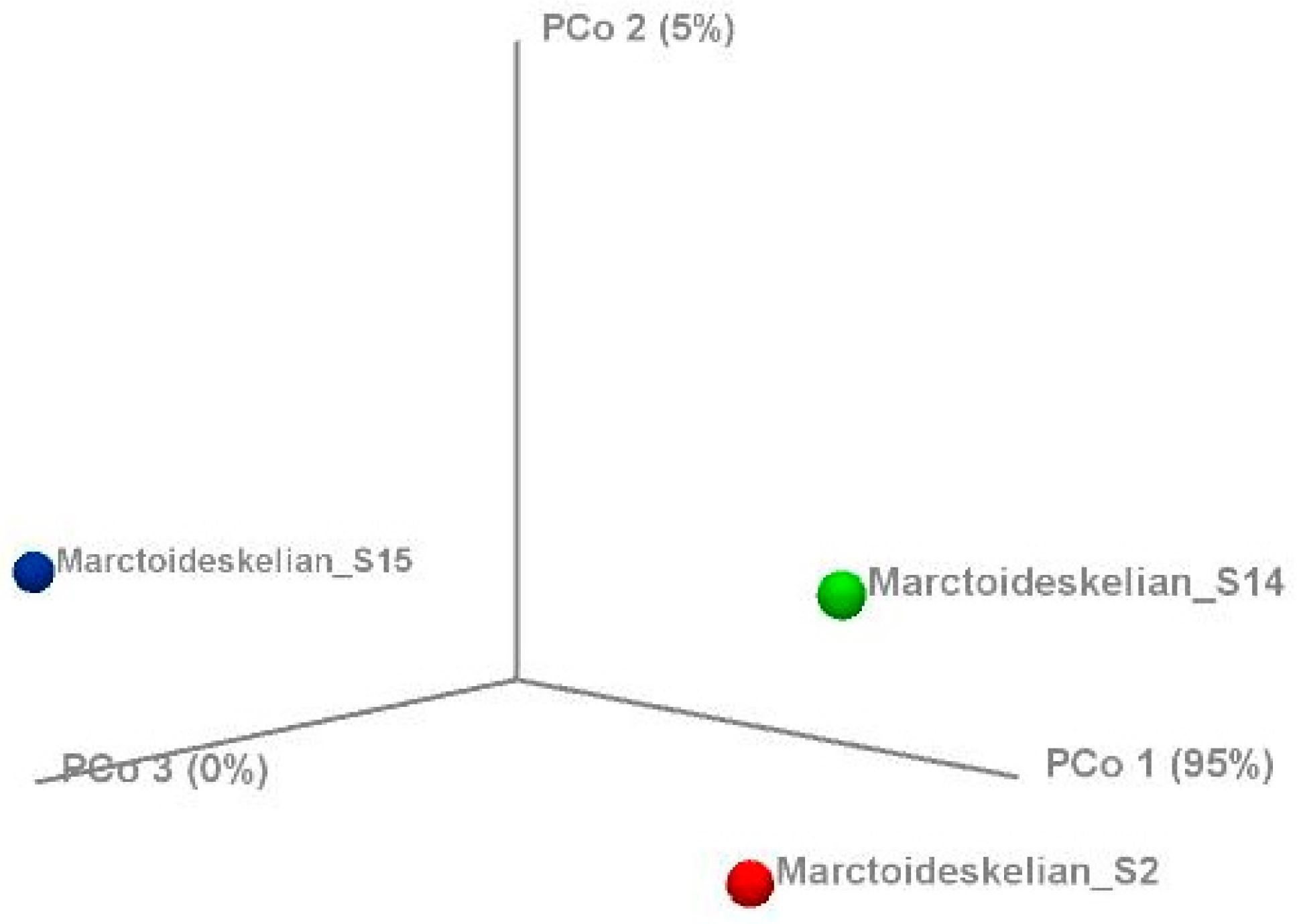
3.3. Heatmap, PCoA, Diversity Indices, and Venn Diagram
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roos, C.; Boonratana, R.; Supriatna, J.; Fellowes, J.R.; Groves, C.P.; Nash, S.D.; Rylands, A.B.; Mittermeier, R.A. An updated taxonomy and conservation status review of Asian primates. Asian Primates J. 2014, 4, 2–38. [Google Scholar]
- Jayaraj, V.K.; Daud, S.H.M.; Azhar, M.-I.; Mohd, S.A.; Mokhtar, S.I.; Abdullah, M.T. Diversity and conservation status of mammals in Wang Kelian State Park, Perlis, Malaysia. J. Species List Distrib. 2013, 9, 1439–1448. [Google Scholar] [CrossRef] [Green Version]
- Syamil, A.R.; Mohd-Ridwan, A.R.; Amsah, M.A.; Abdul-Latiff, M.A.B.; Md-Zain, B.M. Population census and age category character of stump-tailed macaque, Macaca arctoides, in Northern Peninsular Malaysia. Biodiversitas 2019, 20, 2446–2452. [Google Scholar]
- Rahimatsah, A.; Kasim, O. The Administration and Management of Perlis State Park; Faridah-Hanum, L., Osman, K., Yussof, A.R., Latiff, A., Eds.; Forest Department of Perlis: Kangar, Perlis, Malaysia, 2002.
- Chan, K.O.; Grismer, L.L.; Sharma, D.S.; Daicus, B.; Norhayati, A. New herpetofaunal records for Perlis State Park and adjacent areas. Malay. Nat. J. 2009, 61, 277–284. [Google Scholar]
- Syamsul Herman, M.A.; Ahmad, S.; Ramachandran, S.; Rusli, M.Y. Length of stay of discrete outdoor recreational experiences of Perlis State Park. Malays For. 2013, 76, 133–139. [Google Scholar]
- Ampeng, A.; Md-Zain, B.M. Ranging patterns of critically endangered colobine, Presbytis chrysomelas chrysomelas. Sci. World J. 2012, 594382, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Md-Zain, B.M.; Mohamad, M.; Ernie-Muneerah, M.A.; Ampeng, A.; Jasmi, A.; Lakim, M.; Mahani, M.C. Phylogenetic relationships of Malaysian monkeys, Cercopithecidae, based on mitochondrial cytochrome c sequences. Genet. Mol. Res. 2010, 9, 1987–1996. [Google Scholar] [CrossRef]
- Nayak, J.K.; Singh, P. Primatology. In Biological Anthropology Series—1; SSDN Publishers & Distributors: New Delhi, India, 2017. [Google Scholar]
- Sha, J.C.M.; Hanya, G. Diet, activity, habitat use, and ranging of two neighboring groups of food-enhanced long-tailed macaques (Macaca fascicularis). Am. J. Primatol. 2013, 75, 581–592. [Google Scholar] [CrossRef]
- Nila, S.; Suryobroto, B.; Widayati, K.A. Dietary variation of long tailed macaques (Macaca fascicularis) in Telaga Warna, Bogor, West Java. Hayati 2014, 21, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Ruslin, F.; Matsuda, I.; Md-Zain, B.M. The feeding ecology and dietary overlap in two sympatric primate species, the long-tailed macaque (Macaca fascicularis) and dusky langur (Trachypithecus obscurus obscurus), in Malaysia. Primates 2018, 60, 41–50. [Google Scholar] [CrossRef]
- Ruppert, N.; Holzner, A.; See, K.W.; Gisbrecht, A.; Beck, A. Activity budgets and habitat use of wild southern pig-tailed macaques (Macaca nemestrina) in oil palm plantation and forest. Int. J. Primatol. 2018, 39, 237–251. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant. Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Gielly, L.; Miquel, C.; Valentini, A.; Vermat, T.; Corthier, G.; Brochmann, C.; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 2007, 35, e14. [Google Scholar] [CrossRef] [Green Version]
- Srivathsan, A.; Sha, J.C.M.; Vogler, A.P.; Meier, R. Comparing the effectiveness of metagenomics and metabarcoding for diet analysis of a leaf-feeding monkey (Pygathrix nemaeus). Mol. Ecol. Resour. 2015, 15, 250–261. [Google Scholar] [CrossRef]
- Srivathsan, A.; Ang, A.; Vogler, A.P.; Meier, R. Fecal metagenomics for the simultaneous assessment of diet, parasites, and population genetics of an understudied primate. Front. Zool. 2016, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mallott, E.K.; Garber, P.A.; Malhi, R.S. trnL outperforms rbcL as a DNA metabarcoding marker when compared with the observed plant component of the diet of wild white-faced capuchins (Cebus capucinus, Primates). PLoS ONE 2018, 13, e0199556. [Google Scholar] [CrossRef]
- Song, S.J.; Amir, A.; Metcalf, J.L.; Amato, K.R.; Xu, Z.Z.; Humphrey, G.; Knight, R. Preservation methods differ in fecal microbiome stability. Msystems 2016, 1, e00021-16. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Latiff, M.A.B.; Aifat, N.R.; Yaakop, S.; Md-Zain, B.M. A noninvasive molecular approach: Exploiting species-locus-specific pcr primers in defeating numts and dna cross-contamination of cercopithecidae. J. Anim. Plant Sci. 2017, 27, 1015–1023. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Srivastava, A. Conservation of Threatened Primates of Northeast India. Primate Conserv. 2006, 20, 107–113. [Google Scholar] [CrossRef]
- Fooden, J. The bear macaque, Macaca arctoides: A systematic review. J. Hum. Evol. 1990, 19, 607–686. [Google Scholar] [CrossRef]
- Bradley, B.J.; Stiller, M.; Doran-Sheehy, D.M.; Harris, T.; Chapman, C.A.; Vigilant, L.; Poinar, H. Plant DNA sequences from feces: Potential means for assessing diets of wild primates. Am. J. Primatol. 2007, 69, 699–705. [Google Scholar] [CrossRef]
- Da Silva, M.J.F.; Minhos, T.; Sa, R.M.; Bruford, M. Using genetics as a tool in primate conservation. Nat. Educ. Knowl. 2012, 3, 10. [Google Scholar]
- Austerlitz, F.; David, O.; Schaeffer, B.; Bleakley, K.; Olteanu, M.; Leblois, R.; Veuille, M.; Laredo, C. DNA barcode analysis: A comparison of phylogenetic and statistical classification methods. BMC Bioinf. 2009, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.B.; He, L.J.; Crozier, R.H.; Muster, C.; Zhu, C.D. Estimating sample sizes for DNA barcoding. Mol. Phylogenet. Evol. 2010, 54, 1035–1039. [Google Scholar] [CrossRef]
- Liu, J.; Provan, J.; Gao, L.M.; Li, D.Z. Sampling strategy and potential utility of indels for DNA barcoding of closely related plant species: A case study in Taxus. Int. J. Mol. Sci. 2012, 13, 8740–8751. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Huang, C.; Tang, C.; Huang, L.; Tang, H.; Ma, G.; Zhou, Q. Dietary adaptations of Assamese macaques (Macaca assamensis) in limestone forests in Southwest China. Am. J. Primatol. 2014, 77, 171–185. [Google Scholar] [CrossRef]
- Herman, S.; Shuib, A.; Ramachandran, S.; Mohd Rusli, Y.; Richards, A. The need for recreational economic valuation at Perlis State Park. Malays. For. 2012, 75, 73–80. [Google Scholar]
- Zakaria, R.; Said, M.N.M.; Ibrahim, F.H. Association of tree communities with soil properties in a semi deciduous forest of Perlis, Peninsular Malaysia. Biotropia 2019, 27, 69–79. [Google Scholar]
- Galetti, M.; Pedroni, F. Seasonal diet of capuchin monkeys (Cebus apella) in a semideciduous forest in south-east Brazil. J. Trop. Ecol. 1994, 10, 27–39. [Google Scholar] [CrossRef]
- Estrada, A.; Estrada, R. Patterns of predation in a free-ranging troop of stumptail macaques (Macaca arctoides): Relations to the ecology II. Primates 1977, 18, 633–646. [Google Scholar] [CrossRef]
- Estrada, A.; Sandoval, J.M.; Manzolillo, D. Further data on predation by free-ranging stumptail macaques (Macaca arctoides). Primates 1978, 19, 401–407. [Google Scholar] [CrossRef]
- Quéméré, E.; Hibert, F.; Miquel, C.; Lhuillier, E.; Rasolondraibe, E.; Champeau, J.; Rabarivola, C.; Nusbaumer, L.; Chatelain, C.; Gautier, L.; et al. A DNA metabarcoding study of a primate dietary diversity and plasticity across its entire fragmented range. PLoS ONE 2013, 8, e58971. [Google Scholar] [CrossRef] [Green Version]
- Okuda, T.; Manokaran, N.; Matsumoto, Y.; Niiyama, K.; Thomas, S.C.; Ashton, P.S. Pasoh: Ecology of a Lowland Rain Forest in Southeast. Asia; Springer: Tokyo, Japan; Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Badron, U.H.; Talip, N.; Mohamad, A.L.; Affenddi, A.E.A.; Juhari, A.A.A. Studies on leaf venation in selected taxa of the genus Ficus L. (Moraceae) in Peninsular Malaysia. Trop. Life Sci. Res. 2014, 25, 111–125. [Google Scholar]
- Mars, M. Fig (Ficus carica L.) genetic resources and breeding. Acta Hortic. 2003, 605, 19–27. [Google Scholar] [CrossRef]
- Shanahan, M.; Samson, S.O.; Compton, S.G.; Corlett, R. Fig-eating by vertebrate frugivores: A global review. Biol. Rev. Camb. Philos. Soc. 2001, 76, 529–572. [Google Scholar] [CrossRef]
- Hambali, K.; Ismail, A.; Md-Zain, B.M.; Aainaa, A.; Abdul Karim, F. Diet of long-tailed macaques (Macaca fascicularis) at the entrance of Kuala Selangor Nature Park (anthropogenic habitat): Food selection that leads to human-macaque conflict. Acta Biol. Malays. 2014, 3, 58–68. [Google Scholar]
- Kaewpanus, K.; Aggimarangsee, N.; Sitasuwan, N.; Wangpakapattanawong, P. Diet and feeding behavior of Assamese macaque (Macaca assamensis) at Tham Pla temple, Chiang Rai Province, Northern Thailand. J. Wildl. Thail. 2015, 22, 2558. [Google Scholar]
- Riley, E.P.; Tolbert, B.; Farida, W.R. Nutritional content explains the attractiveness of cacao to crop raiding Tonkean macaques. Curr. Zool. 2013, 59, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Wei, H.; Huang, Z.; Huang, C. Diet of the Assamese macaque Macaca assamensis in limestone habitats of Nonggang, China. Curr. Zool. 2011, 57, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Lopes, J.C.; Chatrou, L.W.; Mello-Silva, R.; Rudall, P.J.; Sajo, M.G. Phylogenomics and evolution of floral traits in the neotropical tribe Malmeeae (Annonaceae). Mol. Phylogenet. Evol. 2017, 118, 379–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennington, R.T.; Lavin, M.; Ireland, H.; Klitgaard, B.; Preston, J.; Hu, J.M. Phylogenetic relationships of basal Papilionoid legumes based upon sequences of the chloroplast trnL intron. Syst. Bot. 2001, 26, 537–556. [Google Scholar]
- Cardoso, D.; Pennington, R.T.; de Queiroz, L.P.; Boatwright, J.S.; Van Wyk, B.E.; Wojciechowski, M.F.; Lavin, M. Reconstructing the deep-branching relationships of the papilionoid legumes. S. Afr. J. Bot. 2013, 89, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Pennington, R.T.; Stirton, C.H.; Schrire, B.D. Tribe Sophoreae. In Legumes of the World; Lewis, G., Schrire, B., Mackinder, B., Lock, M., Eds.; Royal Botanic Gardens: Kew, UK, 2005; pp. 227–249. [Google Scholar]
- Rafidah, A.R.; Chew, M.Y.; Ummul-Nazrah, A.R.; Kamarudin, S. The flora of Tasik Bera, Pahang, Malaysia. Malay. Nat. J. 2010, 62, 249–306. [Google Scholar]
- Salleh, N.; Azeman, S.; Kiew, R.; Kamin, I.; Cheng-Kong, R.C. Plant checklist of the Bukit Nanas forest reserve, Kuala Lumpur, Malaysia. One Ecosyst. 2017, 2, e13708. [Google Scholar] [CrossRef] [Green Version]
- Chatrou, L.W.; Pirie, M.D.; Erkens, R.H.J.; Couvreur, T.L.P.; Neubig, K.M.; Abbott, R.; Mols, J.B.; Maas, J.W.; Saunders, R.M.K.; Chase, M.W. A new subfamilial and tribal classification of the pantropical flowering plant family Annonaceae informed by molecular phylogenetics. Bot. J. Linn. Soc. 2012, 169, 5–40. [Google Scholar] [CrossRef] [Green Version]
- Chatrou, L.W.; Escribano, M.P.; Viruel, M.A.; Maas, J.W.; Richardson, J.E.; Hormaza, J.I. Flanking regions of monomorphic microsatellite loci provide a new source of data for plant species-level phylogenetics. Mol. Phylogenet. Evol. 2009, 53, 726–733. [Google Scholar] [CrossRef]
- Mols, J.B.; Keßler, P.J.A.; Rogstad, S.H.; Saunders, R.M.K. Reassignment of six Polyalthia species to the New Genus Maasia (Annonaceae): Molecular and morphological congruence. Syst. Bot. 2008, 33, 490–494. [Google Scholar] [CrossRef]
- Valentini, A.; Miquel, C.; Nawaz, M.A.; Bellemain, E.; Coissac, E.; Pompanon, F.; Gielly, L.; Cruaud, C.; Nascetti, G.; Wincker, P.; et al. New perspectives in diet analysis based on DNA barcoding and parallel pyrosequencing: The trnL approach. Mol. Ecol. Resour. 2008, 9, 51–60. [Google Scholar] [CrossRef]
- Stata, M.; Sage, T.L.; Hoffmann, N.; Covshoff, S.; Wong, G.K.; Sage, R.F. Mesophyll chloroplast investment in C3, C4 and C2 species of the genus Flaveria. Plant. Cell Physiol. 2016, 57, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Aziz, S.A.; Clements, G.R.; Peng, L.Y.; Campos-Arceiz, A.; McConkey, K.R.; Forget, P.-M.; Gan, H.M. Elucidating the diet of the island flying fox (Pteropus hypomelanus) in Peninsular Malaysia through Illumina next-generation sequencing. PeerJ 2017, e3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Level | M. arctoides tRNL |
|---|---|
| Order | 29 |
| Family | 46 |
| Genus | 124 |
| Species | 145 |
| Sample | MA15 | MA2 | MA14 |
|---|---|---|---|
| Simpson_1-D | 0.9937 | 0.9711 | 0.9644 |
| Shannon_H | 8.106 | 7.387 | 7.288 |
| Evenness_e^H/S | 0.1417 | 0.05611 | 0.04545 |
| Chao-1 | 25,210 | 40,080 | 38,610 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, N.A.; Abdul-Latiff, M.A.B.; Mohd-Ridwan, A.R.; Yaakop, S.; Nor, S.M.; Md-Zain, B.M. Diet Composition of the Wild Stump-Tailed Macaque (Macaca arctoides) in Perlis State Park, Peninsular Malaysia, Using a Chloroplast tRNL DNA Metabarcoding Approach: A Preliminary Study. Animals 2020, 10, 2215. https://doi.org/10.3390/ani10122215
Osman NA, Abdul-Latiff MAB, Mohd-Ridwan AR, Yaakop S, Nor SM, Md-Zain BM. Diet Composition of the Wild Stump-Tailed Macaque (Macaca arctoides) in Perlis State Park, Peninsular Malaysia, Using a Chloroplast tRNL DNA Metabarcoding Approach: A Preliminary Study. Animals. 2020; 10(12):2215. https://doi.org/10.3390/ani10122215
Chicago/Turabian StyleOsman, Nur Azimah, Muhammad Abu Bakar Abdul-Latiff, Abd Rahman Mohd-Ridwan, Salmah Yaakop, Shukor Md Nor, and Badrul Munir Md-Zain. 2020. "Diet Composition of the Wild Stump-Tailed Macaque (Macaca arctoides) in Perlis State Park, Peninsular Malaysia, Using a Chloroplast tRNL DNA Metabarcoding Approach: A Preliminary Study" Animals 10, no. 12: 2215. https://doi.org/10.3390/ani10122215