Gut Fungal Communities Are Influenced by Seasonality in Captive Baikal Teal (Sibirionetta formosa) and Common Teal (Anas crecca)

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Sample Collecting
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Bioinformatic Processing and Analysis of the Sequencing Data
2.5. Statistical Analysis
3. Results
3.1. General Characteristics of the Gut Fungal Sequences
3.2. Fungal Diversity and Community Composition
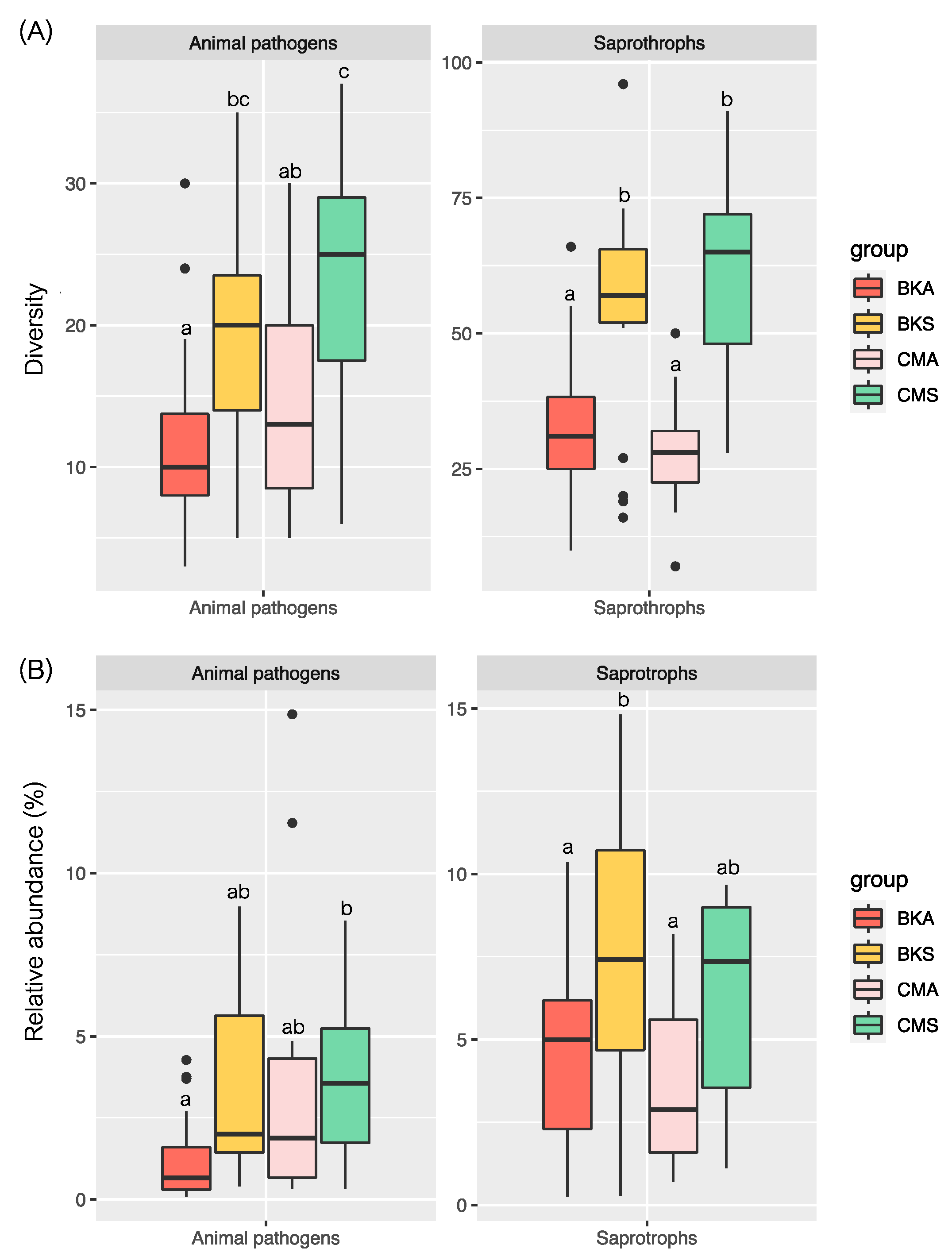
3.3. Potential Animal Pathogens and Saprotrophs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chin, V.K.; Yong, V.C.; Chong, P.P.; Amin Nordin, S.; Basir, R.; Abdullah, M. Mycobiome in the gut: A multiperspective review. Mediat. Inflamm. 2020, 2020, 9560684. [Google Scholar] [CrossRef] [PubMed]
- Kurilshikov, A.; Wijmenga, C.; Fu, J.; Zhernakova, A. Host genetics and gut microbiome: Challenges and perspectives. Trends. Immunol. 2017, 38, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Damare, J.M.; Hussong, D.; Weiner, R.M.; Colwell, R.R. Aerobic and facultatively anaerobic bacteria associated with the gut of Canada geese (Branta canadensis) and whistling swans (Cygnus columbianus columbianus). Appl. Environ. Microbiol. 1979, 38, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Xiang, X.; Dong, Y.; Cheng, L.; Zhou, L. Comparing the intestinal bacterial communities of sympatric wintering Hooded Crane (Grus monacha) and Domestic Goose (Anser anser domesticus). Avian Res. 2020, 11, 13. [Google Scholar] [CrossRef]
- Dong, Z.; Zhang, S.; Cheng, Y.; He, X.; Haase, I.; Liang, Y.; Jiang, Y.; Wu, Y. Comparative analysis of the intestinal tract microbiota and feeding habits of five sympatric flycatchers. Avian Res. 2022, 13, 100050. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, L.; Fu, R.; Cheng, L.; Yan, S.; Mahtab, N.; Song, Y. Effects of foraging site distances on the intestinal bacterial community compositions of the sympatric wintering Hooded Crane (Grus monacha) and Domestic Duck (Anas platyrhynchos domesticus). Avian Res. 2021, 12, 20. [Google Scholar] [CrossRef]
- Hill, S.C.; François, S.; Thézé, J.; Smith, A.L.; Simmonds, P.; Perrins, C.M.; Hoek, L.; Pybus, O.G. Impact of host age on viral and bacterial communities in a waterbird population. ISME J. 2022, 17, 215–226. [Google Scholar] [CrossRef]
- Mahtab, N.; Zhou, L.; Zhang, F.; Wang, W. Seasonal variations in the gut fungal communities of Hooded crane (Grus monacha) at wintering and stopover sites in China. Animals 2021, 11, 941. [Google Scholar] [CrossRef]
- Xiang, X.; Jin, L.; Yang, Z.; Zhang, N.; Zhang, F. Dramatic shifts in intestinal fungal community between wintering Hooded Crane and Domestic Goose. Avian Res. 2021, 12, 1. [Google Scholar] [CrossRef]
- Liu, G.; Xu, N.; Gong, Z.; Feng, J. Gut fungal communities of Anser erythropus wintering at Shengjin Lake and Caizi Lake in China. PJZ 2022, 55, 2163–2172. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Z.; Zhao, J.; Chen, Z.; Xiang, X. Significant differences in intestinal fungal community of hooded cranes along the wintering periods. Front. Microbiol. 2022, 13, 3205. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wu, F.T.; Zhou, Q.H.; Zhao, D.P. Comparative analysis of gut microbiota in captive and wild oriental white storks: Implications for conservation biology. Front. Microbiol. 2021, 12, 649466. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le, P.D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Li, J.; Chen, D.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; Tian, G.; Huang, Z.; et al. Fungi in gastrointestinal tracts of human and mice: From community to functions. Microb. Ecol. 2018, 75, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Su, M.; Duan, Y.; Huang, Y. Eurotium cristatum, a potential probiotic fungus from Fuzhuan brick tea, alleviated obesity in mice by modulating gut microbiota. Food Funct. 2019, 10, 5032–5045. [Google Scholar] [CrossRef] [PubMed]
- Pfavayi, L.T.; Sibanda, E.N.; Mutapi, F. The pathogenesis of fungal-related diseases and allergies in the African population: The state of the evidence and knowledge gaps. Int. Arch. Allergy Immunol. 2020, 181, 257–269. [Google Scholar] [CrossRef]
- Harrop, A.H.; McGowan, R.Y. 2009. Britain’s first Baikal Teal. British Birds 2020, 102, 691. [Google Scholar]
- Delany, S.; Scott, D.; Helmink, A.T.F. Waterbird Population Estimates; Wetlands International: Wageningen, the Netherlands, 2006. [Google Scholar]
- Guillemain, M.; Elmberg, J. The Teal; Bloomsbury Publishing: London, UK, 2014. [Google Scholar]
- Olney, P.J.S. The food and feeding habits of teal Anas crecca. Proc. Zool. Soc. Lond. 1963, 140, 169–210. [Google Scholar] [CrossRef]
- Euliss Jr, N.H.; Harris, S.W. Feeding ecology of northern pintails and green-winged teal wintering in California. J. Wildl. Manag. 1987, 51, 724–732. [Google Scholar] [CrossRef]
- Lebarbenchon, C.; Albespy, F.; Brochet, A.L.; Grandhomme, V.; Renaud, F.; Fritz, H.; Green, A.J.; Thomas, F.; van Der Werf, S.; Aubry, P.; et al. Spread of avian influenza viruses by common teal (Anas crecca) in Europe. PLoS ONE 2009, 4, e7289. [Google Scholar] [CrossRef]
- Hanson, B.A.; Swayne, D.E.; Senne, D.A.; Lobpries, D.S.; Hurst, J.; Stallknecht, D.E. Avian influenza viruses and paramyxoviruses in wintering and resident ducks in Texas. JWD 2005, 41, 624–628. [Google Scholar] [CrossRef]
- Starick, E.; Beer, M.; Hoffmann, B.; Staubach, C.; Werner, O.; Globig, A.; Strebelow, G.; Grund, C.; Durban, M.; Conraths, F.J.; et al. Phylogenetic analyses of highly pathogenic avian influenza virus isolates from Germany in 2006 and 2007 suggest at least three separate introductions of H5N1 virus. Vet. Microbiol. 2008, 128, 243–252. [Google Scholar] [CrossRef]
- Tanikawa, T.; Sakuma, S.; Yoshida, E.; Tsunekuni, R.; Nakayama, M.; Kobayashi, S. Comparative susceptibility of the common teal (Anas crecca) to infection with high pathogenic avian influenza virus strains isolated in Japan in 2004–2017. Vet. Microbiol. 2021, 263, 109266. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Bahl, M.I.; Baunwall, S.M.D.; Hvas, C.L.; Licht, T.R. Determining gut microbial dysbiosis: A review of applied indexes for assessment of intestinal microbiota imbalances. AEM 2021, 87, e00395-21. [Google Scholar] [CrossRef]
- Grond, K.; Sandercock, B.K.; Jumpponen, A.; Zeglin, L.H. The avian gut microbiota: Community, physiology and function in wild birds. JAB 2018, 49, e01788. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal. Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Oksanen, J. Vegan: An Introduction to Ordination. Available online: https://cran.r-project.org/web/packages/vegan/vignettes/intro-vegan.pdf (accessed on 23 April 2023).
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Q.; Doré, J.; Emmanuel, A.; Guarner, F.; Quigley, E.M.M. Gut microbiota and gastrointestinal health: Current concepts and future directions. Neurogastroenterol. Motil. 2013, 25, 4–15. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, V.J.; Song, S.J.; Delsuc, F.; Prest, T.L.; Oliverio, A.M.; Korpita, T.M.; Alexiev, A.; Amato, K.R.; Metcalf, J.L.; Kowalewski, M.; et al. The effects of captivity on the mammalian gut microbiome. ICB 2017, 57, 690–704. [Google Scholar] [CrossRef]
- Zhou, T.; Bodawatta, K.H.; Jiang, A. A network meta-analysis on comparison of invasive and non-invasive sampling methods to characterize intestinal microbiota of birds. Avian Res. 2023, 14, 100086. [Google Scholar] [CrossRef]
- Lewis, W.B.; Moore, F.R.; Wang, S. Characterization of the gut microbiota of migratory passerines during stopover along the northern coast of the Gulf of Mexico. J. Avian Biol. 2016, 47, 659–668. [Google Scholar] [CrossRef]
- Drovetski, S.V.; O’Mahoney, M.J.; Matterson, K.O.; Schmidt, B.K.; Graves, G.R. Distinct microbiotas of anatomical gut regions display idiosyncratic seasonal variation in an avian folivore. Animal Microbiome. 2019, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Maurice, C.F.; CL Knowles, S.; Ladau, J.; Pollard, K.S.; Fenton, A.; Pedersen, A.B.; Turnbaugh, P.J. Marked seasonal variation in the wild mouse gut microbiota. ISME J. 2015, 9, 2423–2434. [Google Scholar] [CrossRef]
- Yi, Y.; Lin, C.; Wang, W.; Song, J. Habitat and seasonal variations in bacterial community structure and diversity in sediments of a Shallow lake. Ecol. Indic. 2021, 120, 106959. [Google Scholar] [CrossRef]
- Wagner Mackenzie, B.; Chang, K.; Zoing, M.; Jain, R.; Hoggard, M.; Biswas, K.; Douglas, R.G.; Taylor, M.W. Longitudinal study of the bacterial and fungal microbiota in the human sinuses reveals seasonal and annual changes in diversity. Sci. Rep. 2019, 9, 17416. [Google Scholar] [CrossRef]
- Nino-Vega, G.A.; Munro, C.A.; San-Blas, G.; Gooday, G.W.; Gow, N.A.R. Differential expression of chitin synthase genes during temperature-induced dimorphic transitions in Paracoccidioides brasiliensis. Med. Mycol. 2000, 38, 31–39. [Google Scholar] [CrossRef]
- Rossi, V.; Ravanetti, A.; Pattori, E.; Giosue, S. Influence of temperature and humidity on the infection of wheat spikes by some fungi causing Fusarium head blight. J. Plant Pathol. 2021, 83, 189–198. [Google Scholar]
- Suradkar, K.P.; Hande, D.V.; Kadu, S.R. Seasonal diversity of endophytic fungi from ten medicinal plants. IJCMAS. 2014, 3, 260–265. [Google Scholar]
- Kakde, U.B.; Kakde, H.U.; Saoji, A.A. Seasonal variation of fungal propagules in a fruit market environment, Nagpur (India). Aerobiologia. 2001, 17, 177–182. [Google Scholar] [CrossRef]
- Selvanathan, S.; Indrakumar, I.; Johnpaul, M. Biodiversity of the endophytic fungi isolated from Calotropis gigantea (L.) R. Br. Recent. Res. Sci. Technol. 2011, 3, 94–100. [Google Scholar]
- Trevelline, B.K.; MacLeod, K.J.; Knutie, S.A.; Langkilde, T.; Kohl, K.D. In ovo microbial communities: A potential mechanism for the initial acquisition of gut microbiota among oviparous birds and lizards. Biol. Lett. 2018, 14, 20180225. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Ng, S.C. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef]
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. N. Biotechnol. 2013, 30, 685–694. [Google Scholar] [CrossRef]
- Siriyappagouder, P.; Kiron, V.; Lokesh, J.; Rajeish, M.; Kopp, M.; Fernandes, J. The intestinal mycobiota in wild zebrafish comprises mainly Dothideomycetes while Saccharomycetes predominate in their laboratory-reared counterparts. Front. Microbiol. 2018, 9, 387. [Google Scholar] [CrossRef]
- Oberwinkler, F. Evolutionary trends in Basidiomycota. Stapefia 2012, 96, 45–104. [Google Scholar]
- Hoppe, B.; Purahong, W.; Wubet, T.; Kahl, T.; Bauhus, J.; Arnstadt, T.; Hofrichter, M.; Buscot, F.; Krüger, D. Linking molecular deadwood-inhabiting fungal diversity and community dynamics to ecosystem functions and processes in Central European forests. Fungal Divers 2016, 77, 367–379. [Google Scholar] [CrossRef]
- de Abreu, D.P.B.; Machado, C.H.; Makita, M.T.; Botelho, C.F.M.; Oliveira, F.G.; da Veiga, C.C.P.; Martins, M.D.A.; Baroni, F.D.A. Intestinal lesion in a dog due to Cryptococcus gattii type VGII and review of published cases of canine gastrointestinal cryptococcosis. Mycopathologia. 2017, 182, 597–602. [Google Scholar] [CrossRef]
- Singh, K.; Ilkit, M.; Shokohi, T.; Tolooe, A.; Malik, R.; Seyedmousavi, S. Cryptococcosis: Emergence of Cryptococcus gattii in animals and zoonotic potential. In Emerging and Epizootic Fungal Infections in Animals; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 249–287. [Google Scholar]
- Navale, V.; Vamkudoth, K.R.; Ajmera, S.; Dhuri, V. Aspergillus derived mycotoxins in food and the environment: Prevalence, detection, and toxicity. Toxicol. Rep. 2021, 8, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Haesebrouck, F.; Martel, A. Aspergillus infections in birds: A review. Avian Pathol. 2010, 39, 325–331. [Google Scholar] [CrossRef]
- Desjardins, A.E. Gibberella from A (venaceae) to Z (eae). Annu. Rev. Phytopathol. 2003, 41, 177–198. [Google Scholar] [CrossRef]
- Vismer, H.F.; Shephard, G.S.; Van der Westhuizen, L.; Mngqawa, P.; Bushula-Njah, V.; Leslie, J.F. Mycotoxins produced by Fusarium proliferatum and F. pseudonygamai on maize, sorghum and pearl millet grains in vitro. Int. J. Food Microbiol. 2019, 296, 31–36. [Google Scholar] [CrossRef]
- Shaman, J.; Pitzer, V.E.; Viboud, C.; Grenfell, B.T.; Lipsitch, M. Absolute humidity and the seasonal onset of influenza in the continental United States. PLoS Biol. 2010, 8, e1000316. [Google Scholar] [CrossRef]
- Hillgarth, N.; Ramenofsky, M.; Winfield, J. Testosterone and sexual selection. Behav. Ecol. 1997, 8, 108–109. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- Mendes-Giannini, M.J.S.; Taylor, M.L.; Bouchara, J.B.; Burger, E.; Calich, V.L.G.; Escalante, E.D.; Hanna, S.A.; Lenzi, H.L.; Machado, M.P.; Miyaji, M.; et al. Pathogenesis II: Fungal responses to host responses: Interaction of host cells with fungi. Mycol. J. 2000, 38, 113–123. [Google Scholar] [CrossRef]
- Carpouron, J.E.; de Hoog, S.; Gentekaki, E.; Hyde, K.D. Emerging animal-associated fungal diseases. J. Fungi. 2022, 8, 611. [Google Scholar] [CrossRef]
- Brown, T.; Jordan, F.T.; Wood, A.M. Fungal diseases. In Poultry Diseases; Elsevier Saunders: Philadelphia, PA, USA, 2008; pp. 247–260. [Google Scholar]
- Odds, F.C. Candida and Candidosis: A Review and Bibliography; Bailliere Tindall: London, UK, 1988. [Google Scholar]
- Mancianti, F.; Nardoni, S.; Ceccherelli, R. Occurrence of yeasts in psittacines droppings from captive birds in Italy. Mycopathologia 2002, 153, 121–124. [Google Scholar] [CrossRef]
- Cafarchia, C.; Camarda, A.; Romito, D.; Campolo, M.; Quaglia, N.C.; Tullio, D.; Otranto, D. Occurrence of yeasts in cloacae of migratory birds. Mycopathologia 2006, 161, 229. [Google Scholar] [CrossRef]
- Soliman, E.S.; Sobeih, M.A.; Ahmad, Z.H.; Hussein, M.M.; Abdel-Latiff, H.; Moneim, A.A. Seasonal epidemiological surveillance on bacterial and fungal pathogens in broiler farms in Egypt. Int. J. Poult. Sci. 2009, 8, 720–727. [Google Scholar] [CrossRef]
- Dynowska, M.; Meissner, W.; Pacyńska, J. Mallard duck (Anas platyrhynchos) as a potential link in the epidemiological chain mycoses originating from water reservoirs. J. Vet. Res. 2013, 57, 323–328. [Google Scholar] [CrossRef]
- Medina, I.R.; Fuentes, L.R.; Arteaga, M.B.; Valcárcel, F.R.; Arbelo, F.A.; Del Castillo, D.P.; Suárez, S.D.; Quintana, O.F.; Gutiérrez, B.V.; Sergent, F.S.; et al. Pigeons and their droppings as reservoirs of Candida and other zoonotic yeasts. Rev. Iberoam. Micol. 2017, 34, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Gleason, F.H.; Marano, A.V.; Johnson, P.; Martin, W.W. Blastocladian parasites of invertebrates. Fungal Biol. Rev. 2010, 24, 56–67. [Google Scholar] [CrossRef]
- Benmayor, R.; Hodgson, D.J.; Perron, G.G.; Buckling, A. Host mixing and disease emergence. Curr. Biol. 2009, 19, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Browne, H.P.; Neville, B.A.; Forster, S.C.; Lawley, T.D. Transmission of the gut microbiota: Spreading of health. Nat. Rev. Microbiol. 2017, 15, 531–543. [Google Scholar] [CrossRef]
- De Vienne, D.M.; Hood, M.E.; Giraud, T. Phylogenetic determinants of potential host shifts in fungal pathogens. J. Evol. Biol. 2009, 22, 2532–2541. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakda, P.; Xiang, X.; Wu, Y.; Zhang, X.; Xu, W.; Zhou, L. Gut Fungal Communities Are Influenced by Seasonality in Captive Baikal Teal (Sibirionetta formosa) and Common Teal (Anas crecca). Animals 2023, 13, 2948. https://doi.org/10.3390/ani13182948
Sakda P, Xiang X, Wu Y, Zhang X, Xu W, Zhou L. Gut Fungal Communities Are Influenced by Seasonality in Captive Baikal Teal (Sibirionetta formosa) and Common Teal (Anas crecca). Animals. 2023; 13(18):2948. https://doi.org/10.3390/ani13182948
Chicago/Turabian StyleSakda, Patthanan, Xingjia Xiang, Yuannuo Wu, Xinying Zhang, Wenbin Xu, and Lizhi Zhou. 2023. "Gut Fungal Communities Are Influenced by Seasonality in Captive Baikal Teal (Sibirionetta formosa) and Common Teal (Anas crecca)" Animals 13, no. 18: 2948. https://doi.org/10.3390/ani13182948