
Fungi Affecting Wall Paintings of Historical Value: A Worldwide Meta-Analysis of Their Detected Diversity



1
Department of Ecological and Biological Sciences (DEB), University of Tuscia, 01100 Viterbo, Italy
2
Department of Sciences, Roma Tre University, 00146 Rome, Italy
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Appl. Sci. 2022, 12(6), 2988; https://doi.org/10.3390/app12062988
Submission received: 4 February 2022
/
Revised: 9 March 2022
/
Accepted: 10 March 2022
/
Published: 15 March 2022
(This article belongs to the Special Issue The Role of Fungi in Biodeterioration of Cultural Heritage: New Insights for Their Control)
Abstract
:Wall paintings have been a cultural expression of human creativity throughout history. Their degradation or destruction represents a loss to the world’s cultural heritage, and fungi have been identified as a major contributor to their decay. We provide a critical review of fungi isolated from worldwide wall paintings between 1961–2021. One-hundred three scientific papers were reviewed focusing on fungal diversity, isolation protocols, and spatial distribution of data. The study sites were grouped into five environmental categories on the basis of the expected major microclimatic conditions (temperature, relative humidity, ventilation), and the possible relationship with the species found was investigated. The highest number of records were localized in Europe, with 38 sites on a total of 74, 20 of which were from Italy. A total of 378 fungal entries were obtained, consisting of 1209 records, belonging to 260 different species and 173 genera. The accuracy level in taxa determination was highly variable among different papers analyzed. Data showed a dominance of Ascomycota, mainly of orders Eurotiales and Hypocreales probably due to their wide distribution and easily air dispersed spores and due to the possible pitfalls linked to the isolation methods, favoring rapidly growing taxa. Statistical analyses revealed that fungal communities were not strictly linked to environmental categories with different ventilation, temperature, and humidity. Such findings may be due to the wide geographical area, the wide heterogeneity of the data, and/or the absence of standardized sampling and analyses protocols. They could also be the result of the dominance of some prevailing factors in the various sites that mask the influence one of each other.
1. Introduction
Wall paintings are among the most representative elements of figurative artworks and have been developed by human creativity since prehistoric times [1]. Their technique of execution requires a layered structure consisting of a support, a ground, and a paint layer, which changed over time across different cultures (secco, such as tempera, or frescoes) [2]. In secco technique, which is the earliest, the preparation layers are applied, but the colors remain on the surface, whereas in the frescoes the colors are applied before the mortar dries, allowing their in-depth penetration [2]. The employed colors usually have a mineral origin, but some pigments can also be derived from plants. Organic compounds can later be added during restoration or because of other human activities (e.g., firing candles in the churches) [3].
The observed deterioration phenomena of mural paintings depend largely on the materials used and the environmental conditions [4]. Indeed, mural paintings are subject to a variety of biodeterioration phenomena, which varies depending on the humidity, lighting, temperature, ventilation, and nutrients, which also select the occurring biological agents [1,5]. Furthermore, many environmental factors may synergistically or antagonistically contribute to the deteriorating actions of microorganisms [6]. Organisms belonging to all domains (bacteria, algae, fungi, animals and sometimes also lichens, mosses, ferns, and higher plants) have been isolated from the surfaces of mural paintings [4,7]. Indeed, given the indoor conditions of most mural paintings, photoautotrophs are highly limited, while fungi and bacteria are more frequent [8]. Bacteria with reduced nutritional needs have been often suggested to be the first colonizers. With their death and lyses, they release organic matter that promotes the growth of secondary colonizers, such as fungi [9,10,11]. Fungi, instead, can produce a large assortment of enzymes and have the remarkable ability to grow and thrive in a wide variety of environmental conditions, including low water activity [12]. Fungi have been rightly recognized as the most common cause of biodeterioration of painted surfaces and other artworks, causing both physical and chemical deterioration phenomena, with aesthetic and structural consequences [1,13,14].
Generally, damage is due to the mycelial growth on the substrate, hyphal penetration, and fruiting bodies production onto and into the substrate, all of which increases the volume and number of cracks, causing the rupture of the pigment layer and leading to surface fragments detachments [1,15]. Fungal colonization generally starts on the surface and then moves in-depth, up to decreasing painted layer cohesion and cause exfoliations and loss of the paint [9,16]. A study carried out by Dornieden and colleagues demonstrated that some fungi, as the so called microcolonial black fungi, are among the most dangerous for cultural heritages and can influence the resistance to shear and torsion stress of mortar and marble, contributing to the separation of different layers of material in mural paintings [17]. Aesthetic damages are also frequent, due to pigment discolorations, mycelial pigmentation, and/or the release of organic pigments of different colors, depending on the species involved. Moreover, secondary compounds such as extracellular enzymes and/or organic acids are generally released in the substrate from fungal hyphae, and this may cause chemical alterations of the mineral constituents of the surfaces as well as the original pigments [9,16]. The secretion of organic acids (e.g., oxalic, citric, succinic, formic, malic, acetic, fumaric, glyoxylic, gluconic, and tartaric acids) also plays a significant role in chemical attack, causing acidification of the substrate [18,19]. They can cause dissolution of cations and chelation of metal ions from mortar and mineral pigments, leading to the formation of stable metal complexes whose crystallization causes an increase of internal pressure resulting in cracking, peeling, and the eventual loss of mural fragments [20].
Awareness of the considerable role played by microorganisms in the preservation of art objects and historical buildings dates back to the 1950s [21]. Ionita and colleagues provided one of the first detailed descriptions of the mycoflora involved in the deterioration of mural paintings of monasteries in Moldavia, noting that it was favored by the various nutritional sources present in the materials used for the realization of the frescoes and by local environmental parameters [21]. This was perhaps one of the first statements of the importance of interdisciplinary studies to prevent and control deterioration processes and define restoration and preservation strategies. Two interesting mini-reviews were later published by Garg and Ciferri teams [1,16]. Many papers have been published after that, showing a growing awareness of the degradative role of fungi as well as the importance of mycological analyses as an integral part of the state-of-the-art system of wall painting safeguards [22].
Despite the fact that the fungal role in the deterioration of frescoes has been documented by a huge number of papers, a global inventory of fungal diversity and their optimal settlement conditions is not yet available. These paintings are mainly present in confined and semi-confined environments, both hypogean and non-hypogean. A fungal alteration pattern dependent on the environmental conditions of these different sites was expected. Those present in hypogean environments are often subjected to a constant extreme humidity, promoting fungal spores germination and mycelial growth. The amount and type of available nutrients also affects the fungal growth rate and the type of fungal taxa. Nutrients may arrive from the external environment as airborne particles, and the more confined are the environments, the lower are the air spores dispersion phenomena. With this contribution, we aimed to describe the diversity of fungal colonizers involved in the deterioration of wall paintings, as well as their distribution under different environmental conditions. Additionally, we aimed to determine if a correlation among the different species recorded and the different types of environments-hypogean, non-hypogean, confined, non-confined, and open-exists and to speculate on their preferential habitat and their possible origin. A dataset of all the fungal taxa occurring on wall paintings based on bibliographic references was created for these purposes.
2. Materials and Methods
2.1. The Bibliographic Search
An extensive search was made among peer reviewed literature, proceedings to conferences, and books. The literature was identified using international databases, such as Scopus (https://www.scopus.com, 29 December 2021), Science Direct (https://www.sciencedirect.com, 29 December 2021), Web of Science (http://www.webofknowledge.com, 29 December 2021) and Google Scholar (https://scholar.google.com, 29 December 2021), that were consulted by using keywords such as ‘wall paintings’, ‘mural paintings’, ‘frescoes’, ‘fungi’, ‘biodeterioration’, ‘microbial deterioration’, and ‘biodeteriogenic agents’. The thematic databases of ICCROM (International Centre for the Study of the Preservation and Restoration of Cultural Property) and the Italian ISCR (Istituto Superiore per la Conservazione e il Restauro) were also consulted, being important reference institutions in the field. Such sources were fundamental in the search of literature related to congresses and reports, that are not found by the most common scientific reference tools. The search covered more than 50 years, dating back to the first papers published in the 1960s (Figure 1), even if mostly of the papers containing useful taxonomic information were published after the 1980s.
2.2. The Database
A list of all the entries retrieved, corresponding to taxa identified at both species and genus level, was compiled. Few were the entries referring to taxa above the rank of genus and they are listed at the end of the database. Current names of the taxa were reported according to the Index Fungorum (www.indexfungorum.org, 20 January 2022). The synonyms under which the different species were eventually reported in the analyzed papers were also indicated. Those entries recorded on paintings that have been the subject of multiple studies over the years, such as Takamatsuzuka and Kitora tumuli in Japan [23,24,25], were reported once accompanied by all bibliographic references.
2.2.1. The Geographic Localization of the Study Sites
The geographical locations and coordinates of the studied monuments were retrieved using Google Maps. The graphical representation on a map of sites distribution was performed using the 3D Map function of the Excel package. Some papers dealt with an unidentified number of monuments, as in the case of ‘Monasteries in Romania’ [21], ‘several churches’ in Northern Portugal [26], or ‘Ajanta caves’ in India [27,28,29,30,31,32]. In these cases, one or more sites were counted, depending on the details provided by the authors within the studied area and on their geographical distribution. Instead, some papers dealt with different monuments from the same area (e.g., different buildings in the historical site of Herculanum, Italy), that have been considered as a single site for the purpose of this study. This is why the total number of monuments is higher than the total number of geographical sites assessed in the statistical analyses.
2.2.2. Isolation and Identification Methods Used
The following information has been recorded: the type of culture media used, the growth temperature and incubation time, and the methods used for isolates identification.
2.2.3. The Types of Environments
The environments that housed the wall paintings object of the studies considered were grouped into five categories, based on whether they were hypogean or not, as well as the expected ventilation, confinement, and moisture conditions. The following are the definitions of the categories:
- C-HE: Confined Hypogean Environment (isolated, without air circulation, with generally high humidity levels and relatively stable and low temperatures) as tumuli, close tombs, or prehistoric caves. They are often characterized by not negligible organic matter inputs from dripping waters, animals, and their fecal pellets and may be influenced by the presence of visitors, promoting the introduction and movement of airborne particulate.
- NC-HE: Non-Confined Hypogean Environment (hypogean environments with reduced air circulation), such as underground crypts, catacombs, rupestrian churches, or Roman houses. These sub-aerial environments offer semi- or non-confined situations that are partially isolated from the external environment, with humidity and temperature comparatively more stable than the outdoor conditions but influenced by external day-night cycles and seasonal fluctuations.
- C-NHE: Confined Non-Hypogean Environment: the sub-aerial environment of churches, monasteries, temples, refectories, and castle chapels represents a confined condition in which the microbial community is enclosed in a mesocosm. They are partially isolated from the external environment and have relatively more stable humidity and temperature than outdoor conditions, but they are influenced by external day-night cycles, seasonal variations in temperature, and relative humidity values. They are more prone to microbial attacks since they include more visited sites.
- SC-NHE: Semi Confined Non-Hypogean Environment (open towards the outdoor environment but protected by wide changes in environmental parameters), such as ancient archaeological buildings and private homes. They are open to the outdoors but sheltered from rain and ventilation; they all experience seasonal and daily relative humidity and temperature fluctuations.
- O-SPE: Open and Semi-Protected or Protected Environment; they include buildings’ walls or collapsed caves, which are among the most exposed sites to biodeterioration risks.
2.3. Statistical Analyses
A data dissimilarity matrix was inferred using the Jaccard’s dissimilarity index [33], and a hierarchical cluster analysis was performed on this matrix using the UPGMA method. Two dendrograms relating dissimilarities between sites and entries were plotted. Entries identified at the genus level were considered as diverse elements. The Silhouette index was used to resolve the optimal number of clusters [34]. A contingency table between the obtained clusters and the environmental categories to which they belonged was also constructed to assess the relationship between their data. An indicator species analysis of the individual clusters was performed, which identifies associations between entries or combinations of entries and clusters, using the Indval index [35,36].
All analyses were performed with the R Software with the packages ade4, vegan, gclus, cluster, vegclust, and indicspecies.
3. Results
3.1. The Fungal Data Set
A total of 103 papers dealing with the fungal deterioration of wall paintings were collected, regarding 107 different monuments grouped in 74 sites. A total of 378 fungal entries were obtained, consisting of 1209 records belonging to 173 genera and 260 species (Table 1).
The taxonomic distribution of the total fungal diversity and within different types of environments, at phylum and order level, is reported in Figure 2. Ascomycota was the dominant phylum, ranging from 89 to 97% (except for the environment O-SPE where a great proportion of undetermined taxa was retrieved) and accounting for 100% of the fungal entries in 40 out of 74 sites. The other two phyla identified were Basidiomycota (1–6%), more abundant in C-NHE, and Mucoromycota, reaching a maximum value (2.35%) in C-NHE (Figure 2A).
The entries were distributed in 39 orders. Eurotiales and Hypocreales were the most represented ones, together accounting for more than half of total fungal diversity. Pleosporales, Capnodiales, Sordariales, and Chaetothyriales were also well represented orders within Ascomycota. Basidiomycota accounted for a maximum of around 6% of all taxa, with Agaricales, Helotiales, and Saccharomycetales being the most abundant ones accordingly. Mucorales was the most abundant order within the phylum Mucoromycota.
The accuracy in taxa determination varied among the different papers analyzed, with many entries identified only at genus level. In fact, a total of 107 out of 378 entries (28.3%) referred to genera not determined at the species level, and it was not possible to quantify the number of possible different species belonging to these genera within different papers. A total of 61 genera (35.2%) were represented by a single species. Aspergillus and Penicillium (order Eurotiales) were the most frequently recorded genera and were represented by a greater number of species, 40 and 46, respectively, present in 44 (59.4%) and 32 (43.2%) sites, which increased to 54 (73%) and 51 (68.9%) when those sites where the genera were reported as undetermined at the species level were also considered. Their contribution to the total number of records was significant, with 219 (18.1%) and 154 (12.7%) records, respectively. Other genera frequently recorded were Alternaria and Fusarium with seven species each; Acremonium, Cladosporium, and Trichoderma with six species; Curvularia and Talaromyces with five species; and Chaetomium with four species.
3.2. The Geographic Distribution of the Study Sites
The data came from 107 monuments, grouped in 74 sites and distributed among 19 different countries. The countries where they were reported as well as the different number of sites were graphically represented in Figure 3.
3.3. Isolation and Identification Methods
A dominance of culture-based methods associated with morphological identification (58%) and target regions sequencing (31%) was recorded, the latter has become progressively dominant since the 2000s. The culture media used varied depending on the research purposes and included Czapek Dox agar (CZ), malt extract agar (MEA), malt agar (MA), potato dextrose agar (PDA), Sabouraud dextrose agar (SAB), and oatmeal agar (OA) among the most frequently used. The use of mycological agar (MYC), Cook’s Rose Bengal (CRB), and yeast peptone dextrose (YPD) was sporadic. Data on growth temperature and incubation time were frequently missing, accounting for 31.25% (Figure 4A, green) and 54.12% (Figure 4B, green) of all papers, respectively. This trend was particularly evident in the earliest papers where more attention was paid to fungal species than to the conditions used to isolate them (e.g., [40,47,84,88]). The most frequent temperature settings were 25 °C (35%) and within the range 27–32 °C (21.25%). When reported, the incubation frequently corresponded to 7 days (31.76%) (Figure 4B). When isolated strains were identified by molecular approaches, the identification was performed targeting different barcoding regions, such as the complete internal transcribed spacer (ITS), a part of it (ITS1), portions of 18S (SSU), 26S (LSU), and β-tubulin.
3.4. Distribution of Fungal Entries in Different Environments
The distribution of the sites among the environmental categories revealed a predominance of wall painting recorded in C-NHE, which alone accounted for 54% of the sites. The hypogean environments accounted for 36.5% of the sites, distributed between C-HE (18.9%) and NC-HE (17.6%). The remaining two categories, SC-NHE and O-SPE, were less represented, comprising 4.1% and 5.4% of the sites, respectively.
The cluster analysis at the level of different sites resulted in a general dispersion, with no distinct clusters retrieved. Several clusters consisted of one or few sites highly different one to each other (13-11-14-3-15-10-9-7-12). Other clusters (4-5-6) were slightly more similar to each other and contained many entries (Figure 5).
The contingency table highlighted that the obtained clusters did not have a strong correlation with the suggested environmental categories (Table 2). However, some clusters showed some affinities: cluster 15 with cat. 4 (SC-NHE) (aff. index 17); cluster 6 with cat. 5 (O-SPE) (aff. index 12.8); clusters 2,4,5 with cat. 3 (C-NHE) (aff. indexes 8.1, 9, and 6.2, respectively); cluster 4 with cat. 2 (NC-HE) (aff. index 5.9); cluster 1 with cat. 1 (C-HE) (aff. index 8.2).
When the correlation among different entries was analyzed, very scattered results were retrieved (Figure S1). However, some entries or combinations of them showed some affinities (specificity and fidelity above 0.5) with certain clusters such as Alternaria alternata with cluster 2 (Indval Index = 0.75); Acrothecium sp. and Penicillium sp. with cluster 4 (Indval Index = 0.70 and 0.66, respectively); Parengyodontium album and Rhinocladiella-like with cluster 5 (Indval Index = 0.70 and 0. 64); Actinomucor elegans, Penicillium pancosmium, Acremoniella atra, Alternaria angustiovoidea, Scolecobasidium anomalum, Scolecobasidium lascauxensis, when considered in combination, the entries Trichoderma sp. and Verticillium sp. with cluster 1 (Indval Index = 0.70, 0.70, 0.70, 0.70, 0.70, and 0.70, respectively); and Akanthomyces lecanii with cluster 9 (Indval Index = 0.93).
4. Discussion
4.1. The Fungal Data Set
Even if fungi have been suggested as secondary colonizers of painted mural substrates, they are among the most common microbial life-forms present in these environments and the primary cause of their biodeterioration [1,13,14]. The wide biodiversity observed confirms the potential key role of fungi in such colonization process and suggests a combination of causes that can favor their growth. Their broad enzymatic activities allow them to grow on every type of material, or wherever they find organic matter. Fungi recorded belonged to species generally reported from natural environments like soils, plants, and air where they live as saprotrophs, as well as plant and animal parasites and pathogens. A detailed survey of their diversity and distribution should become a prerequisite before any restoration measures in order to prevent further damages [39]. Most records belonged to Ascomycota, with Eurotiales being the most common order, due to the prevalence of Aspergillus and Penicillium genera. The former was one of the most frequently isolated genera, with A. flavus and A. niger among the more frequently recorded species. As reported in the literature, even from the first older papers in this field, these two genera, along with Alternaria, Fusarium, Cladosporium, Mortierella, Chaetomium, and Acremonium, are among the most common deteriogens of such paintings [1,11,15,31,39,46,62,78,87,91]. These taxa are ubiquitous, and their frequent occurrence is due to the production of numerous conidia, which are widespread in the environment because they are easily dispersed by air. A diversity of filamentous fungi, with the most predominant genera Penicillium, Cladosporium, Aspergillus, and Trichoderma, were also isolated from mural paintings of the Parish Church of Santo Aleixo (Portugal). Their dehydrogenase activity was determined, as an indicator of the presence of metabolic active cells to allow a deeper insight on the deteriogenic role of the isolates [19].
Species of these genera were recorded on indoor frescoes in numerous monasteries in Romania, possibly favored by the organic components and vegetal pigments used, as well as high moisture levels caused by frequent rainwater penetration, which also resulted in the formation of efflorescences [55]. Cladosporium species can cope in a variety of harsh environmental conditions thanks to their low nutritional requirements (i.e., in oligotrophic conditions). Otherwise, Chaetomium species are proteolytic and cellulolytic ascomycetes, favored by nutrient-rich substrates [22,65,117]. They were reported as the most frequent microfungi on the frescoes of the St. Damian Monastery in Assisi (Italy) [46] and on frescoes in a Serbian church [22]. Furthermore, a community of Aspergillus, Penicillium, Cladosporium, and Chaetomium species was recorded from Medieval wall paintings in Styria (Austria), forming spots of different colors [39]. This group of genera was dominant on two deteriorating frescos in St Clare’s Refectory of the Monastery of St Damian in Assisi [46].
Hypocreales was the second most abundant order, accounting for 18% of total fungal diversity, within which Acremonium, Trichoderma, and Fusarium were among the most common genera. Hypocreales is one of the largest orders of filamentous ascomycetes and exhibits a broad range of ecologies, ranging from plant-associated nutritional modes to animal pathogens (e.g., insect pathogens) and mycoparasites [118]. Neocosmospora solani, recorded in Thailand, India, Japan, and France; Simplicillium lamellicola, recorded in Russia; and Clonostachys rosea [48,58], recorded in Japan, are examples of mycoparasitic species, while Parengiodontium album is an insect parasite and was recorded in several countries (Germany, Russia, Romania, Austria, Italy, and England) [109]. The recurrent presence of mites and insects pointed out their possible role in spreading fungi on painted surfaces [46,119].
Finally, the plant pathogen species Fusarium oxysporum has been shown to produce an extracellular pinkish pigment that disfigures and aesthetically damages colonized mural paintings and stone surfaces with permanent stains [78].
Phylum Basidiomycota was present with several occasional species, mostly represented by one or two records, and comprises litter, soil, and wood-saprotrophs, ectomycorrhizal, epiphyte, and plant-pathogen species. Their occurrence must be regarded as sporadic, potentially aided by root penetration. The possible role of roots as a carrier for rhizosphere microorganisms, like a dripping line for water condensation, and as an organic carbon source by root exudates has been hypothesized [57,120]. In any case, a Basidiomycete was also recorded at the entrance of Roman catacombs [121], possibly due to spores carried by water infiltrations and germinating using organic nutrients from the soil and/or the phototrophic biofilm.
Mucoromycota was present with few species and records, and black meristematic fungi were rarely recorded as well. These latter may grow on a wide range of substrates and are resistant to a variety of environmental stresses, as well as being widely distributed epi- and endolithically on monuments [122,123]. Although the biodiversity of black fungi on historical monuments is not fully elucidated, recent samplings indicate that they are also present on wall paintings and that their rare finding could be linked to the isolation protocols used, generally favoring fast-growing species [124]. Two new species of the genus Neodevrisia have been found in the restricted sampling area of the Vallerano cave and another, still undescribed, from Maijishan grottoes [45,99,108]. Scolecobasidium lascauxensis and S. anomalus were isolated and described from black stains in Lascaux Cave, France [85,115], while the chaetothyrialean black fungi Cladophialophora, Exophiala, and Phialophora have been reported from different sites [23,24,42,98].
Yeasts have been rarely reported, such as Saccharomycetales (Ascomycota) that usually grow by individual yeast cells or Rhodotorula spp. (Basidiomycota) often linked to pink/orange stains due to the release of carotenoids [19,93].
Among those more commonly reported, some species such as those belonging to the genera Alternaria, Fusarium, Aspergillus, Penicillium, and Cladosporium may be responsible for annoying allergic and toxic reactions suffered by conservationists and visitors [81,125,126]. Alternaria alternata is a very common fungal species, frequently recorded on frescoes. Its spores are recognized as common powerful aeroallergens, and indoor environments offer higher levels of exposure to this risk than open-air [59,127]. Records of Fusarium species have also been reported, such as F. solani in the Lascaux caves [97] and F. oxysporum in many sites. They are mainly plant pathogens, but they can also be the causal agents of human mycoses [97,128,129]. Some Aspergillus species are pathogenic to humans and animals and are responsible for clinical manifestations (https://www.aspergillus.org.uk/species-archive/, 29 December 2021). Among these, A. fumigatus is a human pathogenic fungus recorded on frescoes within different environments (3,11,22,29,42,49,70,71), causing infections in humans which can be fatal in immunocompromised patients (61). Aspergillus flavus has been frequently recorded in monasteries, churches, temples, caves, and tombs, mostly due to their numerous aerosolized spores. It mainly grows in the soil, but it is also a facultative and opportunistic pathogen of both animals and plants, producing mycotoxins that are highly harmful to humans [130].
In light of the above, the identification of the species deteriorating wall painting is needed for the protection of restorers and visitors. However, the temperature values characterizing confined and semi-confined sites are generally too low for potential pathogenic fungi. In fact, truly thermophilic fungi which cannot grow at temperatures below 20 °C are not active in these environments, at least during winter in temperate regions.
4.2. Geographic Distribution
Considering the geographic distribution of the data, just one site among the studied paintings comes from the Americas (the Cathedral of Havana at Cuba) [82]. The highest number of records was from Europe, with 70 monuments, mostly representative of hypogean environments and of churches and historical buildings, with a considerable prevalence of Italian monuments (39). A total of 26 monuments were from Asia, while the 13 African ones were all from Egypt.
This distribution arises from the old tradition of people of the Euro-Mediterranean area of using such artistic expression, starting from the old prehistoric caves to the Etruscan and Greek-Roman traditions until the consolidated use both in the decoration of Christian churches and historical buildings [2]. In the case of the Egyptian area, the recorded taxa derived from the old tombs of the Pharaohs [54,72,73,77,78], and similarly in East Asia, the tradition is mainly found in kings’ and Emperors’ tombs [23,24,25,38]. Most of the ancient paintings in buildings do not exist anymore, due to frequent rebuilt or remake of the materials [131].
Our results suggest that the monuments studied were often confined to restricted geographic areas. In any case a wider geographic distribution than that recorded may be possible, as a number of sites may have escaped the search. In fact, even if formally available on the web and on the major repositories, because of the language barriers, some studies could not be taken into account.
4.3. Isolation and Identification Methods
Culture-based methods favor the growth of microorganisms best fitting with the laboratory conditions used (namely, culture media, temperatures, and incubation times). In this study, we found that the most frequent experimental settings were favorable to fast-growing, highly-sporulating fungi, with the use of culture media rich in easily accessible carbon sources, alongside short incubation times and optimal growth temperatures favoring their sporulation. Otherwise, lower growth temperatures (≤20 °C), wide temperature ranges, different isolation media, and a longer incubation time could enlarge the detectable culturable fraction.
Since the early 2000s, molecular phylogenetic methods have highlighted the limitations of morphological identification, allowing us to gain a better understanding of the kingdom of Fungi [132]. Nowadays, the identification by barcode regions sequencing is a common practice. Even if the nuclear ITS region has been recognized as a fungal barcode, its discriminating power changes within the taxonomical groups, and other/more barcode regions are often necessary to have a reliable identification [133]. This is the case of the identification of species within large groups, as Fusarium, Penicillium, Aspergillus, and Cladosporium genera, where cryptic species can be detected only by sequencing multiple molecular markers [134].
In detail, Fusarium species determination has been best made with the combined phylogeny of protein coding genes such as elongation factor (TEF1), RNA polymerase (RPB2) and the partial β-tubulin (BT2) gene [134]. To discriminate between Penicillium and Aspergillus species, β-tubulin (BT2) and calmodulin (cmdA) genes have been proposed as secondary barcodes, respectively [135,136]. While the most phylogenetic informative markers for Cladosporium were TEF1 and actin gene (actA), ITS sequences being identical for species of the same complex [137,138].
The correct identification of strains is required in order to provide restorers more information about strains’ ecology and degradative potential. In this light, standardized identification protocols should be implemented.
High throughput sequencing methods have recently been applied to cultural heritage purposes. These methods represent a powerful tool to define the whole fungal diversity present but not necessarily to deepen the mechanisms and the main actors of the deterioration phenomena [139]. The combination of culture-based and molecular methods should be used for a better understanding of deterioration processes. Indeed, pure cultured microorganisms represent the key to uncover settlers’ physiological and ecological traits, as well as representing a resource for many in silico applications and barcoded identifications [123,124].
4.4. Distribution of Taxa in the Different Environments
The most prevalent sites were confined non-hypogean environments, which are characterized by varied thermo-hygrometric temperatures and air movement. Hypogean (both confined and non-confined), where nutrients and humidity can favor fungal growth were represented as well.
Temperature and relative humidity are among the environmental parameters most important to microbial colonization capability, and in the case of heterotrophs, a certain amount of nutrients is also needed [4,140]. It is well known that fungi rapidly grow when relative humidity is higher than 65% and when a certain quantity of nutrients is available. The low values of temperatures, even if are not favorable for microbial growth by themselves, have a positive effect in contributing to increase in humidity, favoring water condensation on surfaces. Walls, especially in hypogean environments, generally provide these requirements [1]. Temperatures in confined environments are generally more stable than in non-confined environments, where daily and seasonal changes may occur, with ranges that have effects on microbial settlement. Elevated moisture values and stable temperatures have been reported as ideally suited to promote microbial growth on surfaces in catacombs sites [7,94,141]. Indeed, the highest risk occurs when high humidity is coupled with high temperature values, and negative effects of rising temperatures arise only if their highest values can strongly influence the humidity values [142]. In the case of hypogea, the underground conditions favor the maintaining of humidity.
Air movement differences between confined, semi-confined, and non-confined environments were expected to alter the number and type of fungal species recorded as well as incoming nutrients from the outside environment. A great proportion of entries in the database belonged to soil and litter dwellers such as saprotrophs, producing numerous spores that are well adapted to air-borne dispersal, and therefore, air ventilation may have a significant impact on the risk of contamination [143]. The more limited air volume movement of confined mural paintings compared to semi-confined ones was suggested to decrease the number of air-borne dust particles, with biofilm communities relying more on internal interactions between different microorganisms than on the external organic inputs [17]. Among the first aerobiological studies, Savulescu and Ionita reported a greater number of isolates inside the studied monasteries than outside of them, probably due to a more favorable microclimate inside the church, which favors the development of microorganisms [55]. Pangallo and colleagues proposed for the first time a comparative analysis of the microbial component of paintings and the surrounding air to gather information on the origin of fungal contamination [70]. Aside from the importance of aerobiological studies for the conservation and prevention of microbial attacks on indoor painted surfaces [144], these studies have received little attention. In light of the large number of fungal species potentially harmful for restorers and visitors, constant monitoring of air spore quality and concentration, as well as the use of air filters to reduce fungal spores concentrations, would be required for site conservation [62,101,144,145].
Significant correlations between the different taxa and the various environmental categories have not been recorded. Indeed, such data is not the result of the absence of a correlation between fungal growths and environmental conditions but can be consequence of several other influencing conditions that hide it. In fact, many are the ecological requirements that shape the ecological niches of the different species (i.e., the limiting factors), but the most conditioning factors are those that result in a quantity proximal to the upper or lower tolerance limit of an organism [146]. Then, for the various sites examined, some factors may become more relevant if their values are closer to the tolerance limit of certain organism, but this does not mean that other parameters do not play a role [147].
Indeed, our results may be influenced by the wide number of taxa in the wide geographic distribution of sites and by the different methodologies used to characterize the fungal diversity. In fact, different sampling techniques and isolation conditions were used within the studies we analyzed. Other factors that allow fungi to thrive and/or survive in a variety of conditions are their wide nutritional versatility and range of adaptations. The presence of numerous genera that are widespread and highly sporulating and hence present in all the environmental categories must also be considered. The absence of evident correlations could have been determined by the absence of distinct boundaries between the categories identified, with overlapping microclimatic conditions which could have resulted in overlaps within their respective microbial communities. Finally, the heterogeneity of the data, with taxa identified at the genus or species level, may have resulted in dispersed clusters and hampered the ability to demonstrate any relationship.
This result seems to be in line with other studies. The influence of environmental factors such as temperature, relative humidity, and the opening or closure of the temples was not evident for fungal growths on wall paintings of 12 archaeological sites in the central and western parts of Thailand [76]. Furthermore, a stronger relationship with the age of five caves in China than with the environmental conditions, such as temperature and relative humidity, was proposed to explain the observed differences in fungal communities [64]. In two distinct mural paintings, instead, the differences recorded in the microbial communities were associated to the different organic input origin (i.e., wine cellar evaporation, and insect exuvia/excrements) and the microclimatic conditions. The more humid conditions favored the growth of actinomycetes, bacteria, and dark-pigmented fungi, while the other showed a biofilm, mainly dominated by xerotolerant and patchy growing sporulating fungi [17]. Differences in fungal communities were also recorded on mural paintings of two subterranean ancient Chinese tombs dating back over 1700 years, mostly due to variations in interior temperature and relative humidity as well as to their history and drawing techniques used [51].
Other significant concerns could be related to the identification of the isolated species, which was initially based solely on morphological observation. Indeed, phylogenetic molecular approaches are nowadays routinely applied, providing a universal tool for accurately identifying fungal species.
New methodologies such as omics techniques are now available; however, they rarely provide information at species or genus level, and there is no guarantee that the recorded taxa are actively growing. Moreover, culture-dependent approaches may not provide a real picture of the microbial diversity actively growing at the sampling time. This is because not all fungi actively growing on the deteriorated substrates can grow under laboratory conditions, and fungi growing under laboratory conditions may not actively grow on the sampled surfaces. Therefore, a combination of culture-based and molecular approaches may be needed to gain a clear picture of the actual biodiversity present on the painted surfaces as well as to have strains to investigate their potential degradative roles.
5. Conclusions
This study contributes to illustrate the high fungal diversity on wall paintings and raises awareness about the fungal threat on the deterioration of such artworks. Ascomycota was the most common phylum, with Eurotiales and Hypocreales as the most common orders. Statistical analyses did not enhance core communities that can be considered characteristics of different environmental categories of sites hosting wall paintings. Our results were likely due to the heterogeneity and fragmentation of the data in the databases, the dispersed geographical area considered, and the complexity of factors which can condition the biological growths. It is therefore crucial to cover the knowledge gaps through (i) international collaborations, (ii) enlarging the isolation and cultivation protocols as to easily detect also strains different from fast growing ones, and (iii) standardizing the identification protocols. Standardizing and improving the site descriptions (e.g., repeated microclimatic data) could allow for possible relations between site and their settlers and for further comparisons among different environmental conditions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app12062988/s1, Figure S1: Cluster analysis on the Jaccard distance of the distribution of the different entries retrieved from the papers analyzed; Table S1: List of all the references associated to each site defined in the analyses.
Author Contributions
Conceptualization, L.Z. and G.C.; methodology, L.Z., F.C., D.I. and G.C.; data curation, L.Z., F.C., D.I. and G.C.; writing—original draft preparation, L.Z. and F.C.; writing—review and editing, L.Z., F.C., D.I. and G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors wish to acknowledge Giulio Zangari (Department of Sciences, University of ‘Roma Tre’) for his support in some statistical elaborations. Authors wish to acknowledge Giulia Corsetti Antonini and Matt Hudson for the revision of the English language.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Garg, K.L.; Jain, K.K.; Mishra, A.K. Role of fungi in the deterioration of wall paintings. Sci. Total Environ. 1995, 167, 255–271. [Google Scholar] [CrossRef]
- Mora, P.; Mora, L.; Philippot, P. The Conservation of Wall Paintings; Butterworths: London, UK, 1984; 576p. [Google Scholar]
- Giannini, C.; Tapete, D. Materiali e procedimenti esecutivi della pittura murale. In Il Laboratorio dell’Arte, Fonti e Ricerche per la Storia delle Tecniche Artistiche; Il Prato: Saonara, Italy, 2009; Volume 2, 160p. [Google Scholar]
- Caneva, G.; Nugari, M.P.; Nugari, M.P.; Salvadori, O. Plant Biology for Cultural Heritage: Biodeterioration and Conservation; Getty Publications: Los Angeles, CA, USA, 2008. [Google Scholar]
- Caneva, G.; Bartoli, F.; Fontani, M.; Mazzeschi, D.; Visca, P. Changes in biodeterioration patterns of mural paintings: Multi-temporal mapping for a preventive conservation strategy in the Crypt of the Original Sin (Matera, Italy). J. Cult. Herit. 2019, 40, 59–68. [Google Scholar] [CrossRef]
- Scheerer, S.; Ortega Morales, O.; Gaylarde, C.; Allen, I.; Laskin, S.; Geoffrey, G. Microbial deterioration of stone monuments—An updated overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef]
- Albertano, P.; Urzì, C. Structural interactions among epilithic cyanobacteria and heterotrophic microorganisms in Roman hypogea. Microb. Ecol. 1999, 38, 244–252. [Google Scholar] [CrossRef]
- Ranalli, G.; Zanardini, E.; Andreotti, A.; Colombini, M.P.; Corti, C.; Bosch-Roig, P.; De Nuntiis, P.; Lustrato, G.; Mandrioli, P.; Rampazzi, L.; et al. Hi-tech restoration by two-steps biocleaning process of Triumph of Death fresco at the Camposanto Monumental Cemetery (Pisa, Italy). J. Appl. Microbiol. 2018, 125, 800–812. [Google Scholar] [CrossRef]
- Sáiz-Jiménez, C.; Samson, R.A. Biodegradacion de obras de arte. Hongos implicados en la degradacion de los frescos del monasterio de la Rabida (Huelva). Bot. Macaronesica 1981, 8–9, 255–264. [Google Scholar]
- Pepe, O.; Sannino, L.; Palomba, S.; Anastasio, M.; Blaiotta, G.; Villani, F.; Moschetti, G. Heterotrophic microorganisms in deteriorated medieval wall paintings in Southern Italian churches. Microbiol. Res. 2010, 165, 21–32. [Google Scholar] [CrossRef]
- Gomoiu, I.; Cojoc, R.L.; Enache, M.I.; Neagu, S.E.; Mohanu, D.; Mohanu, I. Microbial ability to colonize mural painting and its substrate. Acta Phys. Polo. A 2018, 134, 383–386. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biot. 2013, 97, 9637–9646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosado, T.; Gil, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Oxalate biofilm formation in mural paintings due to microorganisms—A comprehensive study. Int. Biodeter. Biodegr. 2013, 85, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ciferri, O. Microbial degradation of paintings. Appl. Environ. Microbiol. 1999, 65, 879–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dornieden, T.; Gorbushina, A.A.; Krumbein, W.E. Biodecay of cultural heritage as a space/time-related ecological situation—An evaluation of a series of studies. Int. Biodeter. Biodegr. 2000, 46, 261–270. [Google Scholar] [CrossRef]
- Rosado, T.; Martins, M.R.; Pires, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Enzymatic monitorization of mural paintings biodegradation and biodeterioration. Int. J. Conserv. Sci. 2013, 4, 603–612. [Google Scholar]
- Rosado, T.; Gil, M.; Caldeira, A.T.; Martins, M.D.R.; Dias, C.B.; Carvalho, L.; Mirão, J.; Candeias, A.E. Material characterization and biodegradation assessment of mural paintings: Renaissance frescoes from Santo Aleixo Church, Southern Portugal. Int. J. Architect. Herit. 2015, 8, 835–852. [Google Scholar] [CrossRef]
- Unković, N.; Dimkić, I.; Stupar, M.; Stanković, S.; Vukojević, J.; Ljaljević Grbić, M. Biodegradative potential of fungal isolates from sacral ambient: In vitro study as risk assessment implication for the conservation of wall paintings. PLoS ONE 2018, 13, e0190922. [Google Scholar] [CrossRef]
- Ionita, I. Contributions to the study of the biodeterioration of the work of art and of historic monuments. II. Species of fungi involved in the deterioration of mural paintings from the monasteries of Moldavia. Rev. Roum. Biol. Série Bot. 1973, 18, 179–189. [Google Scholar]
- Unković, N.; Grbić, M.L.; Stupar, M.; Savković, Ž.; Jelikić, A.; Stanojević, D.; Vukojević, J. Fungal-induced deterioration of mural paintings: In situ and mock-model microscopy analyses. Microsc. Microanal. 2016, 22, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, J.; Kiyuna, T.; An, K.D.; Nagatsuka, Y.; Handa, Y.; Tazato, N.; Hata-Tomita, J.; Nishijima, M.; Koide, T.; Yaguchi, Y.; et al. Microbiological survey of the stone chambers of Takamatsuzuka and Kitora tumuli, Nara Prefecture, Japan: A milestone in elucidating the cause of biodeterioration of mural paintings. In Proceedings of the 31st International Symposium on the Conservation and Restoration of Cultural Property—Study of Environmental Conditions Surrounding Cultural Properties and Their Protective Measures, Tokyo, Japan, 5–7 February 2008; pp. 51–73. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.619.5561&rep=rep1&type=pdf (accessed on 31 January 2022).
- Sugiyama, J.; Kiyuna, T.; Nishijima, M.; An, K.D.; Nagatsuka, Y.; Tazato, N.; Handa, Y.; Handa-Tomita, J.; Sato, Y.; Kigawa, R.; et al. Polyphasic insights into the microbiomes of the Takamatsuzuka tumulus and Kitora tumulus. J. Gen. Appl. Microbiol. 2017, 63, 63–113. [Google Scholar] [CrossRef] [Green Version]
- Kiyuna, T.; An, K.D.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. Molecular assessment of fungi in “black spots” that deface murals in the Takamatsuzuka and Kitora Tumuli in Japan: Acremonium sect. Gliomastix including Acremonium tumulicola sp. nov. and Acremonium felinum comb. nov. Mycoscience 2011, 52, 1–17. [Google Scholar] [CrossRef]
- Marco, A.; Santos, S.; Caetano, J.; Pintado, M.; Vieira, E.; Moreira, P.R. Basil essential oil as an alternative to commercial biocides against fungi associated with black stains in mural painting. Build. Environ. 2020, 167, 106459. [Google Scholar] [CrossRef]
- Tilak, S.T. Biodeterioration of paintings in Ajanta. In Biodeterioration of Cultural Property; Agrawal, O.P., Dhawan, S., Eds.; Macmillan: New Delhi, India, 1991; pp. 204–212. [Google Scholar]
- Agrawal, O.P.; Dhawan, S.; Garg, K.L.; Shaheen, F.; Pathak, N.; Misra, A. Study of biodeterioration of the Ajanta wall paintings. Int. Biodeterior. 1988, 24, 121–129. [Google Scholar] [CrossRef]
- Garg, K.L.; Dhawan, S.; Bhatnagar, I.K. Microbicides for preservation of wall paintings. In Biodeterioration and Biodegradation; Rossmoore, H.W., Ed.; Elsevier Applied Science: Barking, UK, 1991; Volume 8, pp. 505–507. [Google Scholar]
- Dhawan, S.; Misra, A.; Garg, K.L.; Pathak, N. Laboratory evaluation of orto-phenyl-phenol and p-chloro-m-cresol for the control of some fungal forms of Ajanta wall paintings. In Biodeterioration of Cultural Property; Agrawal, O.P., Dhawan, S., Eds.; Macmillan: New Delhi, India, 1991; pp. 313–338. [Google Scholar]
- Dhawan, S. Microbial deterioration of mural paintings. Biodeterior. Mater. 2002, 2, 95–105. [Google Scholar]
- Dhawan, S.; Garg, K.L.; Pathak, N. Microbial analysis of Ajanta wall paintings & their possible control in situ. In Biodeterioration of Cultural Property, Proceedings of the 2nd International Conference, Yokohama, Japan, 5–8 October 1992; International Communications Specialists: Tokyo, Japan, 1993; Volume 2, pp. 245–262. [Google Scholar]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef] [Green Version]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Sun, J.Z.; Ge, Q.Y.; Zhu, Z.B.; Zhang, X.L.; Liu, X.Z. Three dominating hypocrealean fungi of the ‘white mold spots’ on acrylic varnish coatings of the murals in a Koguryo tomb in China. Phytotaxa 2019, 397, 225–236. [Google Scholar] [CrossRef]
- Jeong, S.H.; Lee, H.J.; Lee, M.Y.; Chung, Y.J. Conservation environment for mural tomb in Goa-ri, Goryeong. J. Cons. Sci. 2017, 33, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Berner, M.; Wanner, G.; Lubitz, W. A comparative study of the fungal flora present in medieval wall paintings in the chapel of the castle Herberstein and in the parish church of St Georgen in Styria, Austria. Int. Biodeter. Biodegr. 1997, 40, 53–61. [Google Scholar] [CrossRef]
- Sáiz-Jiménez, C.; Samson, R.A. Microorganisms and environmental pollution as deteriorating agents of the frescoes of the Monastery of “Santa María de la Rábida”, Huelva, Spain. In Proceedings of the ICOM, Committee for Conservation, 6th Triennial Meeting, Ottawa, ON, Canada, 21–25 September 1981. [Google Scholar]
- Rebricova, N.L. Some ecological aspects of protection of old Russian wall paintings from microbiological deterioration. In Biodeterioration of Cultural Property; Agrawal, O.P., Dhawan, S., Eds.; Macmillan: New Delhi, India, 1991; pp. 294–306. [Google Scholar]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-pigmented biodeteriogenic fungi in Etruscan tombs: New data on their culture dependent diversity and favouring conditions. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef]
- Rebricova, N.L. Micromycetes taking part in deterioration of old Russian wall paintings. In Recent Advances in Biodeterioration and Biodegradation. Biodeterioration of Cultural Heritage; Naya Prokash: Calcutta, India, 1993; Volume 1, pp. 205–232. [Google Scholar]
- Martins, R.; Fialho, S.; Lima, M.; Tavares, D.; Mirão, J.; Valadas, S.; Candeias, A.E. Biodegradation assessment of a 16th century fresco from Southern Portugal. Microsc. Microanal. 2009, 15, 65–66. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Wu, F.; Ma, W.; Zhang, Y.; Gu, J.-D.; Duan, Y.; Xu, R.; Feng, H.; Wang, W.; Li, S.-W. Insights into the bacterial and fungal communities and microbiome that causes a microbe outbreak on ancient wall paintings in the Maijishan Grottoes. Int. Biodeter. Biodegr. 2021, 163, 105250. [Google Scholar] [CrossRef]
- Guglielminetti, M.; De Giuli Morghen, C.; Radaelli, A.; Bistoni, F.; Carruba, G.; Spera, G.; Caretta, G. Mycological and Ultrastructural studies to evaluate biodeterioration of mural paintings. Detection of fungi and mites in frescos of the Monastery of St Damian in Assisi. Int. Biodeter. Biodegr. 1994, 33, 269–283. [Google Scholar] [CrossRef]
- Agarossi, G.; Ferrari, R.; Monte, M. Biocides in the control of biodeterioration. In The Conservation of Monuments in the Mediterranean Basin, Proceedings of the First International Symposium, 7–10 June Bari, Italy, 1989; Grafo Edizioni: Brescia, Italy, 1990; pp. 511–517. [Google Scholar]
- Karpovich-Tate, N.; Rebrikova, N.L. Microbial communities on damaged frescoes and building materials in the cathedral of the Nativity of the Virgin in the Pafnutii-Borovskii monastery, Russia. Int. Biodeter. Biodegr. 1991, 27, 281–296. [Google Scholar] [CrossRef]
- Crippa, A. Funghi isolati da affreschi murali in antiche chiese di Pavia. In Atti Società Italiana Scienze Naturali Museo Civico Storia Naturale Milano; 1983; Volume 124, pp. 3–10. Available online: https://www.biodiversitylibrary.org/item/267082#page/10/mode/1up (accessed on 31 January 2022).
- Sorlini, C.; Sacchi, M.; Ferrari, A. Microbiological deterioration of Gambara’s frescoes exposed to open air in Brescia, Italy. Int. Biodeter. 1987, 23, 167–179. [Google Scholar] [CrossRef]
- Ma, W.; Wu, F.; Tian, T.; He, D.; Zhang, Q.; Gu, J.-D.; Duand, Y.; Mae, D.; Wang, W.; Feng, H. Fungal diversity and its contribution to the biodeterioration of mural paintings in two 1700-year-old tombs of China. Int. Biodeter. Biodegr. 2020, 152, 104972. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Petersen, K. Distribution of microorganisms on ancient wall paintings as related to associated faunal elements. Int. Biodeter. Biodegr. 2000, 46, 277–284. [Google Scholar] [CrossRef]
- Veneranda, M.; Prieto-Taboada, N.; de Vallejuelo, S.F.O.; Maguregui, M.; Morillas, H.; Marcaida, I.; Castro, K.; Madariaga, J.M.; Osanna, M. Biodeterioration of Pompeian mural paintings: Fungal colonization favoured by the presence of volcanic material residues. Environ. Sci. Poll. Res. 2017, 24, 19599–19608. [Google Scholar] [CrossRef]
- Gambino, M.; Ahmed, M.A.A.A.; Villa, F.; Cappitelli, F. Zinc oxide nanoparticles hinder fungal biofilm development in an ancient Egyptian tomb. Int. Biodeter. Biodegr. 2017, 122, 92–99. [Google Scholar] [CrossRef]
- Savulescu, A.; Ionita, I. Contributions to the study of the biodeterioration of the works of art and historic monuments, I. Species of fungi isolated from frescoes. Rev. Roum. Biol. 1971, 16, 201–206. [Google Scholar]
- Unković, N.; Erić, S.; Šarić, K.; Stupar, M.; Savković, Ž.; Stanković, S.; Stanojević, O.; Dimkić, I.; Vukojević, J.; Ljaljević Grbić, M. Biogenesis of secondary mycogenic minerals related to wall paintings deterioration process. Micron 2017, 100, 1–9. [Google Scholar] [CrossRef]
- Mishra, A.K.; Garg, K.L. Microbial deterioration of wall paintings. In Biodeterioration of Cultural Property 3, Proceedings of the 3rd International Conference on Biodeterioration of Cultural Property, Bangkok, Thailand, 4–7 July 1995; Conservation Science Division, Office of Archaeology and National Museums: Bangkok, Thailand, 1995; pp. 630–642. [Google Scholar]
- Emoto, Y. Microbiological investigation of ancient tombs with paintings: Ozuka tomb in Fukuoka and Chibusan tomb in Kumamoto. Sci. Conserv. 1974, 12, 95–102. [Google Scholar]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. [Google Scholar] [CrossRef]
- Stupar, M.; Grbić, M.L.; Simić, G.S.; Jelikić, A.; Vukojević, J.; Sabovljević, M. A sub-aerial biofilms investigation and new approach in biocide application in cultural heritage conservation: Holy Virgin Church (Gradac Monastery, Serbia). Indoor Built Environ. 2012, 23, 584–593. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Heyrman, J.; Dornieden, T.; Gonzalez-Delvalle, M.; Krumbein, W.E.; Laiz, L.; Petersen, L.; Saiz-Jimenez, C.; Swings, J. Bacterial and fungal diversity and biodeterioration problems in mural painting environments of St. Martins church (Greene–Kreiensen, Germany). Int. Biodeter. Biodegr. 2004, 53, 13–24. [Google Scholar] [CrossRef]
- Ripka, K. Identification of Microorganisms on Stone and Mural Paintings Using Molecular Methods. Ph.D. Thesis, University of Wien, Vienna, Austria, 2005; p. 148. Available online: http://othes.univie.ac.at/13628/1/endvers.2.pdf (accessed on 2 December 2021).
- Ma, Y.; Zhang, H.; Du, Y.; Tian, T.; Xiang, T.; Liu, X.; Wu, F.; An, L.; Wang, W.; Gu, J.-D.; et al. The community distribution of bacteria and fungi on ancient wall paintings of the Mogao Grottoes. Sci. Rep. 2015, 5, 7752. [Google Scholar] [CrossRef]
- Unković, N.; Ljaljević Grbić, M.; Subakov-Simić, G.; Stupar, M.; Vukojević, J.; Jelikić, A.; Stanojević, D. Biodeteriogenic and toxigenic agents on 17th century mural paintings and facade of the old church of the Holy Ascension (Veliki Krčimir, Serbia). Indoor Built Environ. 2015, 25, 826–837. [Google Scholar] [CrossRef]
- Rosado, T.; Mirão, J.; Candeias, A.; Caldeira, A.T. Microbial communities analysis assessed by pyrosequencing—A new approach applied to conservation state studies of mural paintings. Anal. Bioanal. Chem. 2014, 406, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepe, O.; Palomba, S.; Sannino, L.; Blaiotta, G.; Ventorino, V.; Moschetti, G.; Villani, F. Characterization in the archaeological excavation site of heterotrophic bacteria and fungi of deteriorated wall painting of Herculaneum in Italy. J. Environ. Biol. 2011, 32, 241–250. [Google Scholar] [PubMed]
- Sprocati, A.R.; Alisi, C.; Tasso, F.; Vedovato, E.; Barbabietola, N.; Cremisini, C. A microbiological survey of the Etruscan Mercareccia tomb (Italy): Contribution of microorganisms to deterioration and restoration. In Art 2008, Proceedings of the 9th International Conference on NDT of Art, Jerusalem, Israel, 25–30 May 2008; NDT of Art: Jerusalem, Israel, 2008; 9p. [Google Scholar]
- Sampǒ, S.; Luppi Mosca, A.M. A study of the fungi occurring on 15th century frescoes in Florence, Italy. Int. biodeterior. 1989, 25, 343–353. [Google Scholar] [CrossRef]
- Pangallo, D.; Kraková, L.; Chovanová, K.; Šimonovičová, A.; De Leo, F.; Urzì, C. Analysis and comparison of the microflora isolated from fresco surface and from surrounding air environment through molecular and biodegradative assays. World J. Microb. Biot. 2012, 28, 2015–2027. [Google Scholar] [CrossRef]
- Pangallo, D.; Chovanová, K.; Šimonovicová, A.; De Leo, F.; Urzì, C. Assessment of the biodeterioration risk of the Ladislav lagend fresco in Velka Lomnica (SK) through non-invasive methods. In Proceedings of the 11th International Congress on Deterioration and Conservation of Stone, Torun, Poland, 15–20 September 2008; Volume 1, pp. 457–464. [Google Scholar]
- Helmi, F.M.; Elmitwalli, H.R.; Rizk, M.A.; Hagrassy, A.F. Antibiotic extraction as a recent biocontrol method for Aspergillus niger and Aspergillus flavus fungi in ancient Egyptian mural paintings. Mediterr. Archaeol. Arc. 2011, 11, 1–7. [Google Scholar]
- Khalaphallah, R.; El-Derby, A.A. The effect of nano-TiO2 and plant extracts on microbial strains isolated from Theban ancient Egyptian royal tomb painting. Afr. J. Microbiol. Res. 2015, 9, 1424–1430. [Google Scholar] [CrossRef]
- Elhagrassy, A.F. Isolation and characterization of actinomycetes from mural paintings of Snu-Sert-Ankh tomb, their antimicrobial activity, and their biodeterioration. Microbiol. Res. 2018, 216, 47–55. [Google Scholar] [CrossRef]
- Chaisrisook, C.; Suwanarit, P.; Aranyanak, C. Fungal deterioration of mural paintings in the royal temple. In Biodeterioration of Cultural Property 3, Proceedings of the 3rd International Conference on Biodeterioration of Cultural Property, Bangkok, Thailand, 4–7 July 1995; Conservation Science Division, Office of Archaeology and National Museums: Bangkok, Thailand, 1995; pp. 116–137. [Google Scholar]
- Senbua, W.; Wichitwechkarn, J. Molecular identification of fungi colonizing art objects in Thailand and their growth inhibition by local plant extracts. 3 Biotech 2019, 9, 356. [Google Scholar] [CrossRef]
- Stoyancheva, G.; Krumova, E.; Kostadinova, N.; Miteva-Staleva, J.; Grozdanov, P.; Ghaly, M.F.; Sakr, A.A.; Angelova, M. Biodiversity of contaminant fungi at different coloured materials in ancient Egypt Tombs and Mosques. Cr. Acad. Bul. Sci. 2018, 71, 907–915. [Google Scholar] [CrossRef]
- Sakr, A.; Ghaly, M.; Helal, G.; Abdel Haliem, M. Effect of thymol against fungi deteriorating mural paintings at Tell Basta tombs, Lower Egypt. Int. J. Res. Stud. Biosci. 2012, 6, 8–23. [Google Scholar] [CrossRef]
- Raschle, P. Experience of combating moulds during restoration of ceiling paintings in a Swiss baroque monastery church. Biodeterioration 1983, 5, 427–433. [Google Scholar]
- Rosado, T.; Falé, A.; Gil, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Understanding the influence of microbial contamination on colour alteration of pigments used in wall paintings—The case of red and yellow ochres and ultramarine blue. Color Res. Appl. 2019, 44, 783–789. [Google Scholar] [CrossRef]
- Di Carlo, E.; Chisesi, R.; Barresi, G.; Barbaro, S.; Lombardo, G.; Rotolo, V.; Palla, F. Fungi and bacteria in indoor Cultural Heritage environments: Microbial-related risks for artworks and human health. Environ. Ecol. Res. 2016, 4, 257–264. [Google Scholar] [CrossRef]
- Cepero, A.; Martinez, P.; Castro, J.; Sanchez, A.; Machado, J. The biodeterioration of cultural property in the republic of Cuba: A review of some experiences. In Biodeterioration of Cultural Property, Proceedings of the 2nd International Conference, Yokohama, Japan, 5–8 October 1992; International Communications Specialists: Tokyo, Japan, 1993; pp. 479–487. [Google Scholar]
- Arai, H. Relationship between fungi and brown spots found in various materials. In Biodeterioration of Cultural Property, Proceedings of the 2nd International Conference, Yokohama, Japan, 5–8 October 1992; International Communications Specialists: Tokyo, Japan, 1993; pp. 320–336. [Google Scholar]
- Barbieri, N.; Bassi, M.; Dassù, G.; Rossi, F. Gli affreschi del tempio repubblicano di Brescia: Condizioni ambientali ed inquinamento microbiologico. Arte Lomb. 1986, 76/77, 113–117. [Google Scholar]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Envir. Sci. Tech. 2012, 46, 3762–3770. [Google Scholar] [CrossRef]
- Bartolini, M.; Nugari, M.P.; Pietrini, A.M.; Ricci, S.; Roccardi, A.; Filetici, M.G. Gli ambienti ipogei delle domus romane al Celio: Indagini biologiche per il controllo e la prevenzione del biodeterioramento. Kermes La Riv. Del Restauro 2010, 23, 45–54. [Google Scholar]
- Vasanthakumar, A.; DeAraujo, A.; Mazurek, J.; Schilling, M.; Mitchell, R. Microbiological survey for analysis of the brown spots on the walls of the tomb of King Tutankhamun. Int. Biodeter. Biodegr. 2013, 79, 56–63. [Google Scholar] [CrossRef]
- Tonolo, A.; Giacobini, C. Microbiological changes of frescoes. In Recent Advances in Conservation; Thomson, G., Ed.; Butterworths: London, UK, 1961; pp. 62–64. [Google Scholar]
- Arai, H. The environmental analysis of archaeological sites. TrAC Trends Anal. Chem. 1990, 9, 213–216. [Google Scholar] [CrossRef]
- Sorlini, C.; Allievi, L.; Sacchi, M.; Ferrari, A. Microorganisms present in deteriorated materials of the Palazzo della Ragione in Milan. Int. Biodeterior. Bull. 1982, 18, 105–110. [Google Scholar]
- Rosado, T.; Mirão, J.; Candeias, A.; Caldeira, A.T. Characterizing microbial diversity and damage in mural paintings. Microsc. Microanal. 2015, 21, 78. [Google Scholar] [CrossRef]
- Popescu, A.; Arai, H.; Minatoya, T. Biodeterioration aspects of the Probota Monastery and possibilities for its restoration. In Biodeterioration of Cultural Property 3, Proceedings of the 3rd International Conference on Biodeterioration of Cultural Property, Bangkok, Thailand, 4–7 July 1995; Conservation Science Division, Office of Archaeology and National Museums: Bangkok, Thailand, 1995; pp. 255–271. [Google Scholar]
- Gargani, G. Fungus contamination of Florence art masterpieces before and after the 1966 disaster. In Biodeterioration of Materials, Microbiological and Allied Aspects; Elsevier: Amsterdam, The Netherlands, 1968; pp. 252–257. [Google Scholar]
- Saarela, M.; Alakomi, H.L.; Suihko, M.L.; Maunuksela, L.; Raaska, L.; Mattila-Sandholm, T. Heterotrophic microorganisms in air and biofilm samples from Roman catacombs, with special emphasis on actinobacteria and fungi. Int. Biodeter. Biodegr. 2004, 54, 27–37. [Google Scholar] [CrossRef]
- Palla, F.; Billeci, N.; Mancuso, F.P.; Pellegrino, L.; Lorusso, L.C. Microscopy and molecular biology techniques for the study of biocenosis diversity in semi-confined environments. Conserv. Sci. Cult. Herit. 2010, 10, 185–194. [Google Scholar] [CrossRef]
- Nagatsuka, Y.; Kiyuna, T.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. Candida tumulicola sp. nov. and Candida takamatsuzukensis sp. nov., novel yeast species assignable to the Candida membranifaciens clade, isolated from the stone chamber of the Takamatsuzuka tumulus. Int. J. Syst. Evol. Microbiol. 2009, 59, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Jacquet, C.; Dennetiere, B.; Lacoste, S.; Bousta, F.; Orial, G.; Cruaud, C.; Couloux, A.; Roquebert, M.F. Invasion of the French Paleolithic painted cave of Lascaux by members of the Fusarium solani species complex. Mycologia 2007, 99, 526–533. [Google Scholar] [CrossRef]
- Kiyuna, T.; An, K.D.; Kigawa, R.; Sano, C.; Sugiyama, J. Two new Cladophialophora species, C. tumbae sp. nov. and C. tumulicola sp. nov., and chaetothyrialean fungi from biodeteriorated samples in the Takamatsuzuka and Kitora Tumuli. Mycoscience 2018, 59, 75–84. [Google Scholar] [CrossRef]
- Zucconi, L.; Gagliardi, M.; Isola, D.; Onofri, S.; Andaloro, M.C.; Pelosi, C.; Pogliani, C.; Selbmann, L. Biodeterioration agents dwelling in or on the wall paintings of the Holy Saviour’s cave (Vallerano, Italy). Int. Biodeter. Biodegr. 2012, 70, 40–46. [Google Scholar] [CrossRef]
- Bianchi, A.; Favali, M.A.; Barbieri, N.; Bassi, M. The use of fungicides on mold-covered frescoes in S. Eusebio in Pavia. Int. Biodeterior. Bull. 1980, 16, 45–51. [Google Scholar]
- Pitzurra, L.; Bellezza, T.; Giammarioli, M.; Giraldi, M.; Sbaraglia, G.; Spera, G.; Bistoni, F. Microbial environmental monitoring of the refectory in the monastery of St. Anna in Foligno, Italy. Aerobiologia 1999, 15, 203–209. [Google Scholar] [CrossRef]
- Bassi, M.; Giacobini, C. Scanning electron microscopy: A new technique in the study of the microbiology of works of art. Int. Biodeter. Biodegr. 2001, 48, 55–66. [Google Scholar] [CrossRef]
- Jeffries, P. Biodeterioration of wall paintings in Canterbury Cathedral. In Biodeterioration of Cultural Property; Agrawal, O.P., Dhawan, S., Eds.; Macmillan: Delhi, India, 1991; pp. 287–293. [Google Scholar]
- Fiorillo, F.; Fiorentino, S.; Montanari, M.; Monaco, C.R.; Del Bianco, A.; Vandini, M. Learning from the past, intervening in the present: The role of conservation science in the challenging restoration of the wall painting Marriage at Cana by Luca Longhi (Ravenna, Italy). Herit. Sci. 2020, 8, 10. [Google Scholar] [CrossRef]
- Kiyuna, T.; An, K.D.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. Mycobiota of the Takamatsuzuka and Kitora Tumuli in Japan, focusing on the molecular phylogenetic diversity of Fusarium and Trichoderma. Mycoscience 2008, 49, 298–311. [Google Scholar] [CrossRef]
- Stomeo, F.; Portillo, M.C.; Gonzalez, J.M. Assessment of bacterial and fungal growth on natural substrates: Consequences for preserving caves with prehistoric paintings. Curr. Microbiol. 2009, 59, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Kiyuna, T.; An, K.D.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. Bristle-like fungal colonizers on the stone walls of the Kitora and Takamatsuzuka Tumuli are identified as Kendrickiella phycomyces. Mycoscience 2012, 53, 446–459. [Google Scholar] [CrossRef]
- Egidi, E.; de Hoog, G.S.; Isola, D.; Onofri, S.; Quaedvlieg, W.; de Vries, M.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and taxonomy of meristematic rock-inhabiting black fungi in the Dothideomycetes based on multi-locus phylogenies. Fungal Divers. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Nugari, M.P.; Pietrini, A.M.; Caneva, G.; Imperi, F.; Visca, P. Biodeterioration of mural paintings in a rocky habitat: The Crypt of the Original Sin (Matera, Italy). Int. Biodeter. Biodegr. 2009, 63, 705–711. [Google Scholar] [CrossRef]
- Jeffries, P. Growth of Beauveria alba on mural paintings in Canterbury Cathedral. Int. Biodeter. Biodegr. 1986, 22, 11–13. [Google Scholar]
- Leplat, J.; Francois, A.; Bousta, F. White fungal covering on the wall paintings of the Saint-Savin-sur-Gartempe Abbey church crypt: A case study. Int. Biodeter. Biodegr. 2017, 122, 29–37. [Google Scholar] [CrossRef]
- Milanesi, C.; Baldi, F.; Vignani, R.; Ciampolini, F.; Faleri, C.; Cresti, M. Fungal deterioration of medieval wall fresco determined by analysing small fragments containing copper. Int. Biodeter. Biodegr. 2006, 57, 7–13. [Google Scholar] [CrossRef]
- Moza, M.I.; Mironescu, M.; Georgescu, C.; Florea, A.; Bucşa, L. Isolation and characterisation of moulds degrading mural paintings. Ann. RSCB 2012, 17, 136–142. [Google Scholar]
- An, K.D.; Kiyuna, T.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. The identity of Penicillium sp. 1, a major contaminant of the stone chambers in the Takamatsuzuka and Kitora Tumuli in Japan, is Penicillium paneum. Anton. van Leeuw. 2009, 96, 579. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Two new species of the genus Ochroconis, O. lascauxensis and O. anomala isolated from black stains in Lascaux Cave, France. Fungal Biol. 2012, 116, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Caneva, G.; Tescari, M.; Bartoli, F.; Nugari, M.P.; Pietrini, A.M.; Salvadori, O. Ecological mapping for the preventive conservation of prehistoric mural paintings in rock habitats: The site of Filiano (Basilicata, Italy). Conserv. Sci. Cult. Herit. 2015, 15, 53–59. [Google Scholar] [CrossRef]
- Unković, N.; Ljaljević Grbić, M.; Stupar, M.; Vukojević, J.; Subakov-Simić, G.; Jelikić, A.; Stanojević, D. ATP bioluminescence method: Tool for rapid screening of organic and microbial contaminants on deteriorated mural paintings. Nat. Prod. Res. 2015, 33, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Aime, M.C.; Grigoriev, I.V.; Martin, F.; Stajich, J.E.; Blackwell, M. The fungal tree of life: From molecular systematics to genome-scale phylogenies. In The Fungal Kingdom; Heitman, J., Howlett, B.J., Crous, P.W., Stukenbrock, E.H., James, T.Y., Gow, N.A.R., Eds.; ASM press: Washington, DC, USA, 2007; pp. 1–34. [Google Scholar] [CrossRef] [Green Version]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeterior. Biodegrad. 2008, 62, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Isola, D.; Bartoli, F.; Langone, S.; Ceschin, S.; Zucconi, L.; Caneva, G. Plant DNA barcode as a tool for root identification in hypogea: The Case of the Etruscan Tombs of Tarquinia (Central Italy). Plants 2021, 10, 1138. [Google Scholar] [CrossRef]
- Cuzman, O.A.; Tapete, D.; Fratini, F.; Mazzei, B.; Riminesi, C.; Tiano, P. Assessing and facing the biodeteriogenic presence developed in the Roman Catacombs of Santi Marco, Marcelliano e Damaso, Italy. Eur. J. Sci. Theol. 2014, 10, 185–197. [Google Scholar]
- Isola, D.; Selbmann, L.; Meloni, P.; Maracci, E.; Onofri, S.; Zucconi, L. Detrimental rock black fungi and biocides: A study on the Monumental Cemetery of Cagliari. In Science and Technology for the Conservation of Cultural Heritage; Rogerio-Candelera, M.A., Lazzari, M., Cano, E., Eds.; CRC Press: London, UK, 2013; pp. 83–86. [Google Scholar]
- Isola, D.; Bartoli, F.; Meloni, P.; Caneva, G.; Zucconi, L. Black fungi and stone heritage conservation: Ecological and metabolic assays for evaluating colonization potential and responses to traditional biocides. Appl. Sci. 2022, 12, 2038. [Google Scholar] [CrossRef]
- Isola, D.; Scano, A.; Orrù, G.; Prenafeta-Boldú, F.X.; Zucconi, L. Hydrocarbon-contaminated sites: Is there something more than Exophiala xenobiotica? New insights into black fungal diversity using the long cold incubation method. J. Fungi 2021, 7, 817. [Google Scholar] [CrossRef]
- Sterflinger, K.; Pinzari, F. The revenge of time: Fungal deterioration of cultural heritage with particular reference to books, paper and parchment. Environ. Microbiol. 2012, 14, 559–566. [Google Scholar] [CrossRef]
- Pyzik, A.; Ciuchcinski, K.; Dziurzynski, M.; Dziewit, L. The Bad and the Good—Microorganisms in Cultural Heritage Environments—An Update on Biodeterioration and Biotreatment Approaches. Materials 2021, 14, 177. [Google Scholar] [CrossRef]
- Salo, P.M.; Arbes, S.J.; Sever, M.; Jaramillo, R.; Cohn, R.D.; London, S.J.; Zeldin, D.C. Exposure to Alternaria alternata in US homes is associated with asthma symptoms. J. Aller. Cl. Imm. 2006, 118, 892–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Magnon, K.C.; Cox, P.A.; Revankar, S.G.; Sanche, S.; Geiser, D.M.; Juba, J.H.; van Burik, J.-A.H.; et al. Genetic diversity of human pathogenic members of the Fusarium oxysporum complex inferred from multilocus DNA sequence data and amplified fragment length polymorphism analyses: Evidence for the recent dispersion of a geographically widespread clonal lineage and nosocomial origin. J. Clin. Microbiol. 2004, 42, 5109–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domsch, K.H. Compendium of Soil Fungi; Taxonomically Revised by Walter Gams; IHW: Eching, Germany, 2007; 672p. [Google Scholar]
- Hyde, K.D.; Al-Hatmi, A.M.S.; Andersen, B.; Boekhout, T.; Buzina, W.; Dawson, T.L.; Eastwood, D.C.; Gareth Jones, E.B.; de Hoog, G.S.; Kang, Y.; et al. The world’s ten most feared fungi. Fungal Divers. 2018, 93, 161–194. [Google Scholar] [CrossRef]
- Forster, A.M.; Thomson, D.; Richards, K.; Pilcher, N.; Vettese, S. Western and Eastern building conservation philosophies: Perspectives on permanence and impermanence. Int. J. Architect. Her. 2019, 13, 870–885. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lu Cking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Tekpinar, A.D.; Kalmer, A. Utility of various molecular markers in fungal identification and phylogeny. Nova Hedwig. 2019, 109, 187–224. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Van Den Ende, A.H.G.; Stielow, J.B.; Van Diepeningen, A.D.; Seifert, K.A.; McCormick, W.; Assabgui, R.; Gräfenhan, T.; De Hoog, S.G.; Levesque, C.A. Evaluation of two novel barcodes for species recognition of opportunistic pathogens in Fusarium. Fungal Biol. 2016, 120, 231–245. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, T.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 7, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Visagie, C.M.; Frisvad, J.C. Recommendations to prevent taxonomic misidentification of genome-sequenced fungal strains. Microbiol. Resour. Announc. 2021, 10, e01074-20. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Gene, J.; Sutton, D.A.; Wiederhold, N.P.; Cano-Lira, J.F.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia 2016, 36, 281–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterflinger, K.; Little, B.; Pinar, G.; Pinzari, F.; de los Rios, A.; Gu, J.D. Future directions and challenges in biodeterioration research on historic materials and cultural properties. Int. Biodeterior. Biodegrad. 2018, 129, 10–12. [Google Scholar] [CrossRef]
- Caneva, G.; Isola, D.; Lee, H.J.; Chung, Y.J. Biological risk for hypogea: Shared data from Etruscan tombs in Italy and ancient tombs of the Baekje dynasty in Republic of Korea. Appl. Sci. 2020, 10, 6104. [Google Scholar] [CrossRef]
- Sanchez-Moral, S.; Canaveras, J.C.; Laiz, L.; Saiz-Jimenez, C.; Bedoya, J.; Luque, L. Biomediated precipitation of calcium carbonate metastable phases in hypogean environments: A short review. Geomicrobiol. J. 2003, 20, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Caneva, G.; Langone, S.; Bartoli, F.; Cecchini, A.; Meneghini, C. Vegetation cover and tumuli’s shape as affecting factors of microclimate and biodeterioration risk for the conservation of Etruscan tombs (Tarquinia, Italy). Sustainability 2021, 13, 3393. [Google Scholar] [CrossRef]
- Savković, Ž.; Stupar, M.; Unković, N.; Knežević, A.; Vukojević, J.; Ljaljević Grbić, M. Fungal Deterioration of Cultural Heritage Objects. In Biodegradation; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Nugari, M.P.; Realini, M.; Roccardi, A. Contamination of mural paintings by indoor airborne fungal spores. Aerobiologia 1993, 9, 131–139. [Google Scholar] [CrossRef]
- Mandrioli, P.; Caneva, G.; Sabbioni, C. Cultural Heritage and Aerobiology. Methods and Measurement Techniques for Biodeterioration Monitoring; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; 260p. [Google Scholar] [CrossRef]
- Odum, E.P.; Odum, H.T.; Andrews, J. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1971; Volume 3. [Google Scholar]
- Caneva, G.; Bartoli, F.; Savo, V.; Futagami, Y.; Strona, G. Combining statistical tools and ecological assessments in the study of biodeterioration patterns of stone temples in Angkor (Cambodia). Sci. Rep. 2016, 6, 32601. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
PRISMA flow diagram of the search process.

Figure 2.
Taxonomic composition of the fungal diversity recorded on wall paintings on the total dataset and in relation to the different environments, at (A) phylum and (B) order level.
Figure 2.
Taxonomic composition of the fungal diversity recorded on wall paintings on the total dataset and in relation to the different environments, at (A) phylum and (B) order level.

Figure 3.
Geographic distribution of the reviewed study’s 107 monuments. Nations are indicated with the international alpha-3 code: ITA: Italy, EGY: Egypt, THA: Thailand, PRT: Portugal, CHN: China, ROU: Romania, AUT: Austria, GER: Germany, JAP: Japan, IND: India, RUS: Russian Federation, ESP: Spain, SRB: Republic of Serbia, CUB: Cuba, FRA: France, KOR: Republic of Korea, SVK: Slovak Republic, CHE: Swiss, GBR: Great Britain. In the blue rectangle Cuba.
Figure 3.
Geographic distribution of the reviewed study’s 107 monuments. Nations are indicated with the international alpha-3 code: ITA: Italy, EGY: Egypt, THA: Thailand, PRT: Portugal, CHN: China, ROU: Romania, AUT: Austria, GER: Germany, JAP: Japan, IND: India, RUS: Russian Federation, ESP: Spain, SRB: Republic of Serbia, CUB: Cuba, FRA: France, KOR: Republic of Korea, SVK: Slovak Republic, CHE: Swiss, GBR: Great Britain. In the blue rectangle Cuba.

Figure 4.
Values/ranges of (A) growth temperature and (B) incubation time recorded in culture-based protocols.
Figure 4.
Values/ranges of (A) growth temperature and (B) incubation time recorded in culture-based protocols.

Figure 5.
Cluster dendrogram. Cluster analysis on the Jaccard distance of the composition at the level of the different sites. Sites belonged to the following categories: Site1–Site14 C-HE; Site15–Site27 NC-HE; Site 28–Site67 C-NHE; Site 68–Site70 SC-NHE; Site 71–Site74 O-SPE. The references associated to each site are reported in Supplementary Table S1.
Figure 5.
Cluster dendrogram. Cluster analysis on the Jaccard distance of the composition at the level of the different sites. Sites belonged to the following categories: Site1–Site14 C-HE; Site15–Site27 NC-HE; Site 28–Site67 C-NHE; Site 68–Site70 SC-NHE; Site 71–Site74 O-SPE. The references associated to each site are reported in Supplementary Table S1.


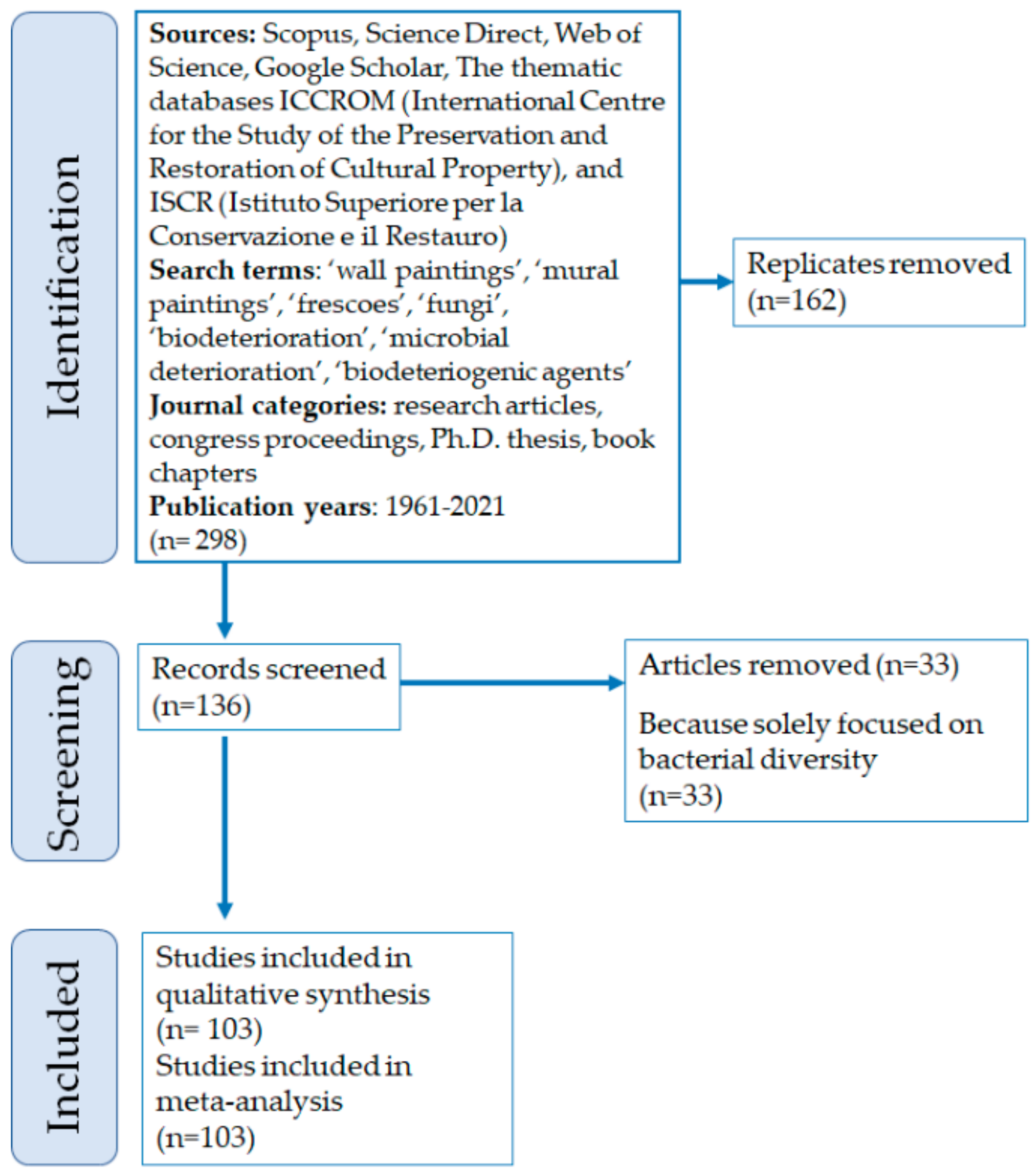{kind=link}
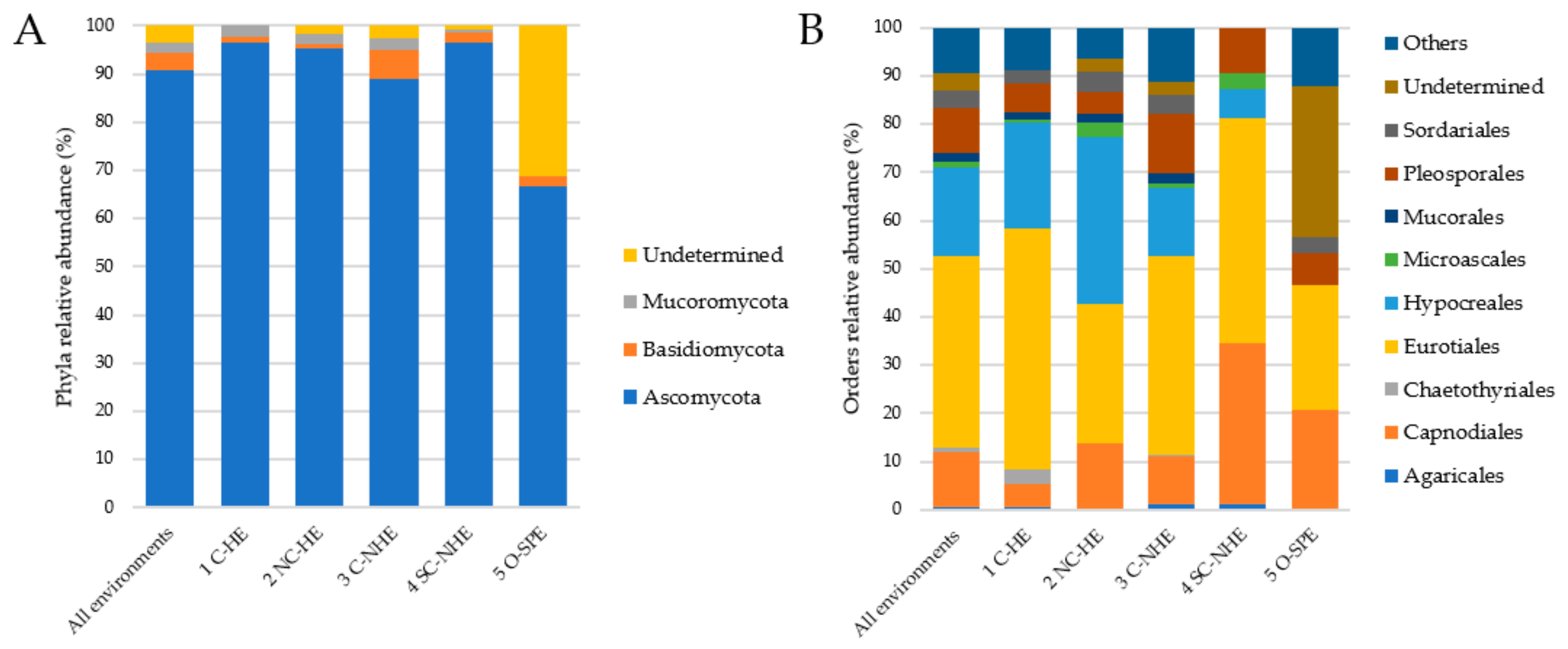{kind=link}
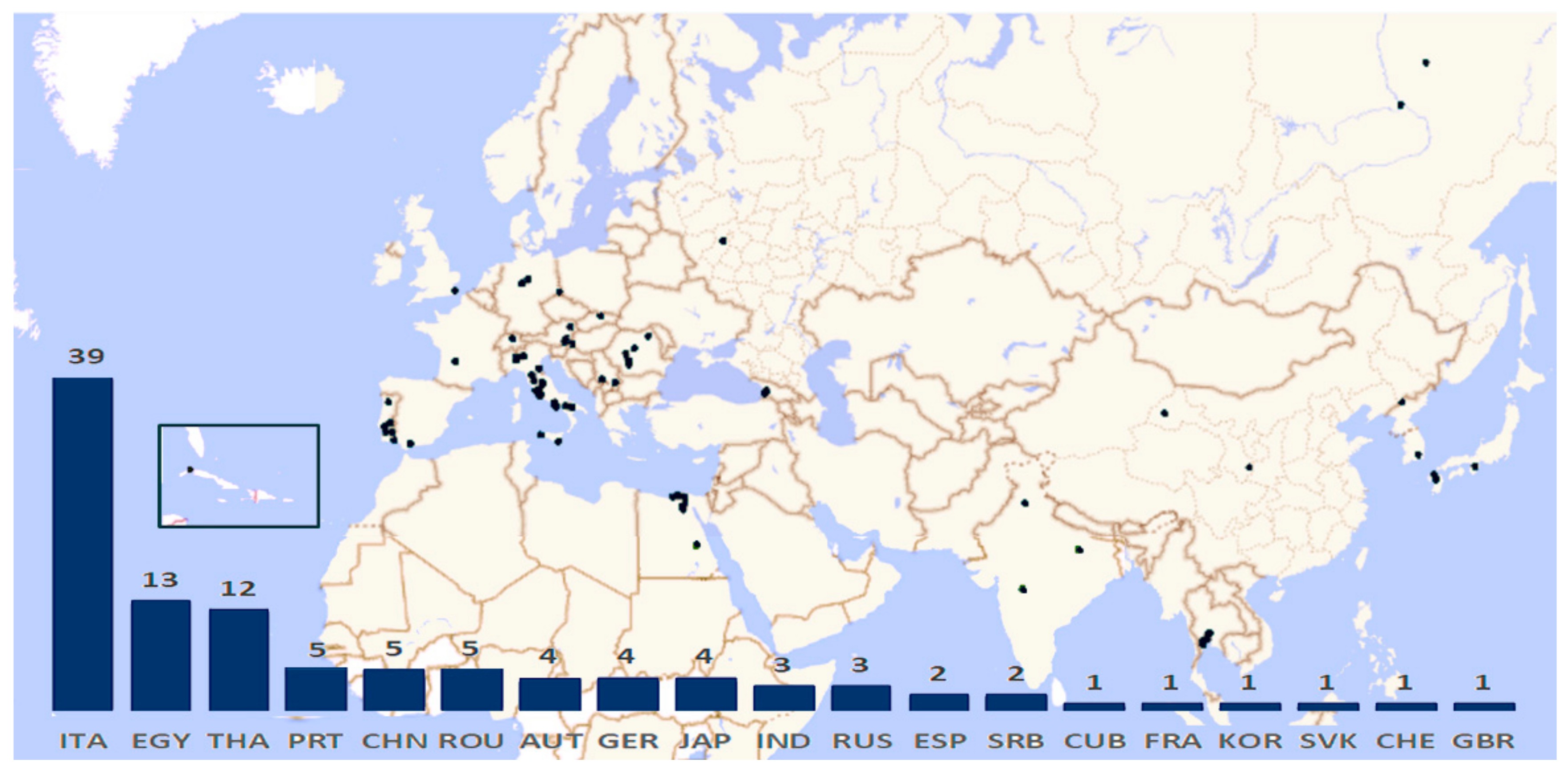{kind=link}
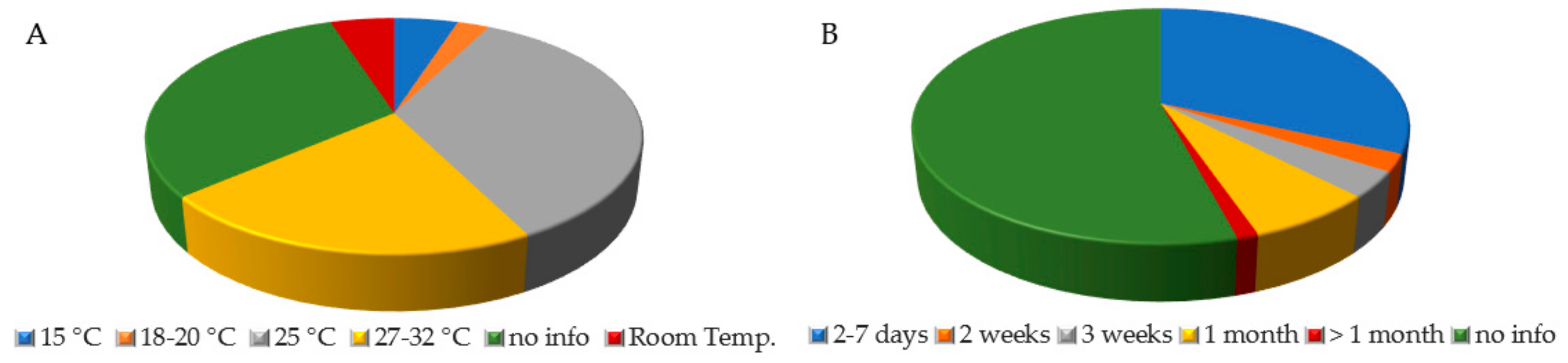{kind=link}
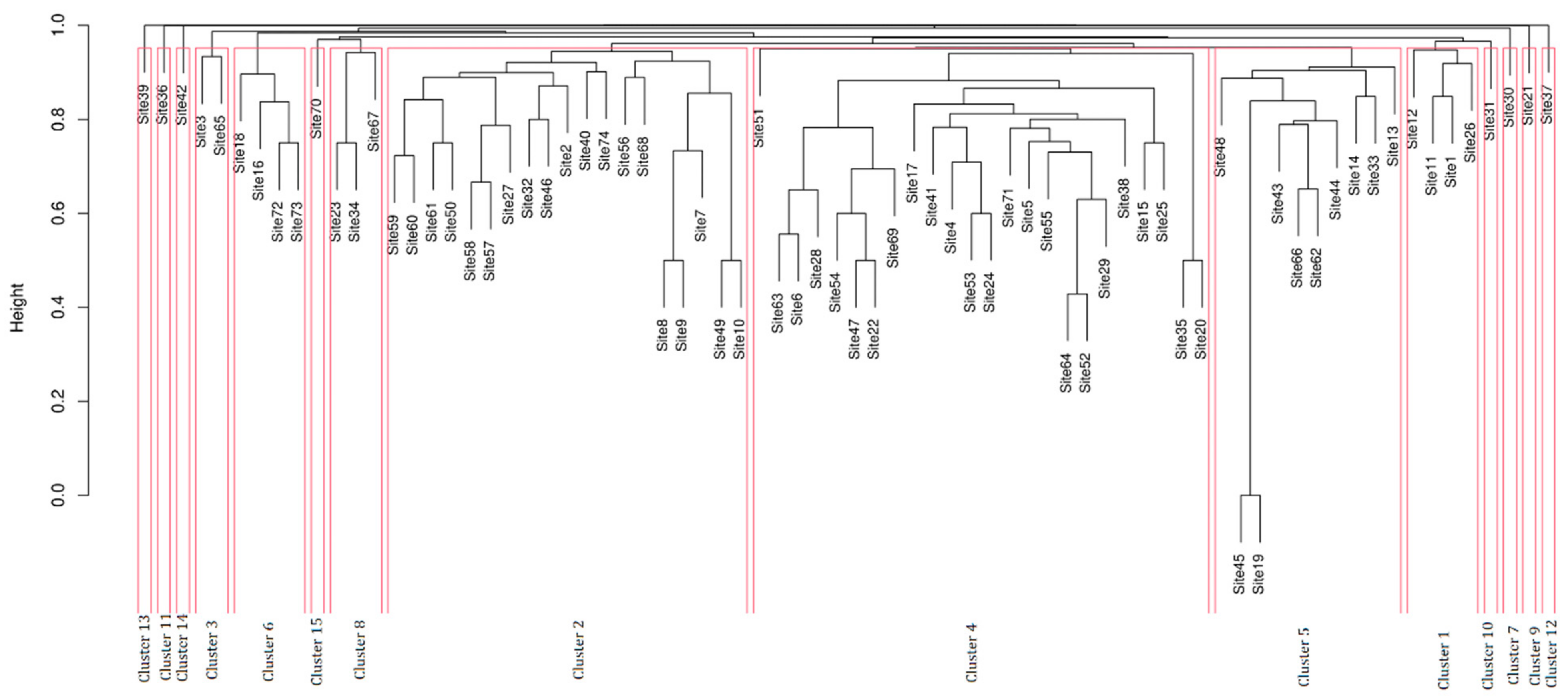{kind=link}
Table 1.
List of the fungal entries retrieved from the different papers grouped by genera, in association with the corresponding references and the environmental categories where they have been registered.
Table 1.
List of the fungal entries retrieved from the different papers grouped by genera, in association with the corresponding references and the environmental categories where they have been registered.
| Genus | Fungal Name | References | Environment |
|---|---|---|---|
| Acremoniella | Acremoniella atra | [21] | C-NHE |
| Acremonium | Acremonium camptosporum | [37] | NC-HE |
| Acremonium charticola | [38,39,40,41] | NC-HE, C-NHE | |
| Acremonium masseei | [23,25] | C-HE | |
| Acremonium murorum (syn. Gliomastix murorum) | [23,24,25,28,32] | C-HE, NC-HE | |
| Acremonium rutilum (syn. A. roseum) | [21] | C-NHE | |
| Acremonium cf. rutilum | [39] | C-NHE | |
| Acremonium sp. | [23,38,39,40,41,42,43,44,45,46,47] | C-HE, NC-HE, C-NHE | |
| Acrodontium | Acrodontium crateriforme | [48] | C-NHE |
| Acrophialophora | Acrophialophora fusispora (syn. A. nainiana) | [28,32] | NC-HE |
| Acrostalagmus | Acrostalagmus luteoalbus (syn. Verticillium lateritium) | [49] | C-NHE |
| Acrothecium | Acrothecium sp. | [50] | O-SPE |
| Actinomucor | Actinomucor elegans | [51] | C-HE |
| Akanthomyces | Akanthomyces lecanii (syn. Verticillium lecanii) | [39,43,52] | NC-HE, C-NHE |
| Allophoma | Allophoma labilis (syn. Phoma labilis) | [53] | SC-NHE |
| Alternaria | Alternaria alternata (syn. A. tenuis and Ulocladium alternariae) | [21,26,28,29,31,32,40,46,49,51,53,54,55,56,57,58,59] | All environments |
| Alternaria angustiovoidea | [60] | C-NHE | |
| Alternaria chartarum (syn. Ulocladium chartarum) | [21,61] | C-NHE | |
| Alternaria dianthi | [31] | C-NHE | |
| Alternaria longipes | [31] | C-NHE | |
| Alternaria longissima | [28,31,32] | NC-HE, C-NHE | |
| Alternaria oudemansii (syn. Ulocladium oudemansii) | [62] | NC-HE, C-NHE | |
| Alternaria tenuissima | [21,22,24,31,46,57,63] | NC-HE, C-NHE, O-SPE | |
| Alternaria sp. (syn. Ulocladium sp.) | [10,11,21,23,26,38,41,46,64,65] | C-HE, NC-HE, C-NHE, O-SPE | |
| Amphinema | Amphinema sp. | [66] | C-NHE |
| Amyloporia | Amyloporia sinuosa (syn. Antrodia sinuosa) | [11] | C-NHE |
| Antrodia | Antrodia sp. | [66] | C-NHE |
| Apiotrichum | Apiotrichum sp. (syn. Hyalodendron sp.) | [43] | C-NHE |
| Arachnomyces | Arachnomyces sp. | [45] | NC-HE |
| Armillaria | Armillaria sp. | [66] | C-NHE |
| Arthrinium | Arthrinium arundinis | [40] | C-NHE |
| Arthrinium phaeospermum (syn. Papularia sphaerosperma) | [28,32] | NC-HE | |
| Arthrinium sp. | [46,65] | C-NHE; O-SPE | |
| Arthrobotrys | Arthrobotrys sp. | [23] | C-HE |
| Ascochyta | Ascochyta medicaginicola (syn. Phoma medicaginis) | [20,56] | O-SPE |
| Ascochyta sp. | [63] | C-NHE | |
| Ascotricha | Ascotricha guamensis | [32] | NC-HE |
| Aspergillus | Aspergillus aeneus | [67] | SC-NHE |
| Aspergillus amstelodami (syn. Eurotium amstelodami) | [46,55,63] | C-NHE | |
| Aspergillus aureolatus | [20,56] | O-SPE | |
| Aspergillus auricomus | [22] | O-SPE | |
| Aspergillus candidus | [28,32,39,68] | C-HE, NC-HE, C-NHE | |
| Aspergillus clavatus | [37] | C-HE | |
| Aspergillus creber | [20,56] | O-SPE | |
| Aspergillus echinulatus | [21] | C-NHE | |
| Aspergillus europaeus | [20,56] | O-SPE | |
| Aspergillus fischeri (syn. Neosartorya fischeri) | [62] | NC-HE; C-NHE | |
| Aspergillus flavipes | [20,56] | O-SPE | |
| Aspergillus flavus (syn. A. oryzae) | [10,17,20,28,30,31,32,56,57,68,69,70,71,72,73,74,75] | C-HE, NC-HE, C-NHE, O-SPE | |
| Aspergillus fumigatus | [30,31,38,46,61,76,77,78,79] | C-HE, NC-HE, C-NHE | |
| Aspergillus glaucus group | [79] | C-NHE | |
| Aspergillus ivoriensis | [67] | SC-NHE | |
| Aspergillus japonicus | [78] | C-HE | |
| Aspergillus melleus | [67] | SC-NHE | |
| Aspergillus multicolor | [67] | SC-NHE | |
| Aspergillus nidulans (syn. Emericella nidulans) | [28,30,31,32,46,51,61,70,71,78] | C-HE, NC-HE, C-NHE | |
| Aspergillus niger | [10,17,19,20,28,30,31,32,49,50,53,54,55,56,72,74,75,77,80,81,82] | All environments | |
| Aspergillus niger group | [46] | C-NHE | |
| Aspergillus ochraceus | [38,49,67] | C-HE, C-NHE, SC-NHE | |
| Aspergillus ostianus | [20,56,67] | SC-NHE, O-SPE | |
| Aspergillus pallidofulvus | [20,56] | O-SPE | |
| Aspergillus parasiticus | [20,56] | O-SPE | |
| Aspergillus penicilloides | [83] | C-HE | |
| Aspergillus petrakii | [67] | SC-NHE | |
| Aspergillus proliferans | [28,32] | NC-HE | |
| Aspergillus protuberus | [67] | SC-NHE | |
| Aspergillus puniceus | [67] | SC-NHE | |
| Aspergillus repens | [21,41] | C-NHE | |
| Aspergillus restrictus | [63,84] | C-HE, C-NHE | |
| Aspergillus sclerotiorum | [76] | C-NHE | |
| Aspergillus spectabilis (syn. Emericella spectabilis) | [67] | SC-NHE | |
| Aspergillus stellatus (syn. Emericella variecolor) | [67] | SC-NHE | |
| Aspergillus sydowii | [28,32,39,51,52,62,69] | C-HE, NC-HE, C-NHE | |
| Aspergillus terreus | [29,30,31,32,78] | C-HE, NC-HE, C-NHE | |
| Aspergillus unguis | [76] | C-NHE | |
| Aspergillus ustus | [46,67] | C-NHE, SC-NHE | |
| Aspergillus versicolor | [17,28,31,32,39,41,43,46,49,51,55,61,67,68,69,70,71,78,82,84,85,86] | C-HE, NC-HE, C-NHE, SC-NHE | |
| Aspergillus wentii | [28,31,32] | NC-HE, C-NHE | |
| Aspergillus sp. | [11,18,19,22,23,24,27,44,45,46,50,64,66,69,75,77,82,87,88,89,90,91,92,93,94,95] | C-HE, NC-HE, C-NHE, O-SPE | |
| Aureobasidium | Aureobasidium pullulans | [31,41,43,45,49,52,57] | NC-HE, C-NHE |
| Beauveria | Beauveria bassiana | [48] | C-NHE |
| Beauveria sp. | [45,52] | NC-HE, C-NHE | |
| Bispora | Bispora sp. | [65] | O-SPE |
| Bjerkandera | Bjerkandera adusta | [77] | C-NHE |
| Blastobotrys | Blastobotrys aristatus | [39] | C-NHE |
| Blastomyces | Blastomyces sp. | [84] | C-HE |
| Botryotrichum | Botryotrichum atrogriseum | [17,55,59] | NC-HE, C-NHE |
| Botryotricum domesticum | [60] | C-NHE | |
| Botryotrichum murorum (syn. Chaetomium murorum) | [20,21,46,55,56] | C-NHE, O-SPE | |
| Botrytis | Botrytis cinerea | [40,41,43,46,69] | C-NHE |
| Brunneochlamydosporium | Brunneochlamydosporium nepalense (syn. Acremonium nepalense) | [38,85] | C-HE |
| Burgoa | Burgoa sp. | [23] | C-HE |
| Candida | Candida takamatsuzukensis | [23,96] | C-HE |
| Candida tumulicola | [23,96] | C-HE | |
| Candida sp. | [23,24,65] | C-HE, NC-HE | |
| Capronia | Capronia coronata | [85] | C-HE |
| Cephalotrichum | Cephalotrichum verrucisporum (syn. Doratomyces verrucisporus) | [23,24] | C-HE |
| Cephalotrichum sp. (syn. Doratomyces sp.) | [23,24] | C-HE | |
| Cephalosporium | Cephalosporium sp. | [88,90] | C-NHE |
| Chaetomium | Chaetomium ancistrocladum | [20,56] | O-SPE |
| Chaetomium elatum | [40] | C-NHE | |
| Chaetomium globosum | [21,22,26,31,32,38,41,46,51,55] | C-HE, NC-HE, C-NHE, O-SPE | |
| Chaetomium piluliferum (syn. Botryotrichum piluliferum) | [21,55] | C-NHE | |
| Chaetomium sp. | [9,22,27,31,39,40,43,46,65,70,71,72] | NC-HE, C-NHE, O-SPE | |
| Chondrostereum | Chondrostereum sp. | [66] | C-NHE |
| Chrysosporium | Chrysosporium pseudomerdarium | [85] | C-HE |
| Chrysosporium sp. | [47,52,62,97] | C-HE, NC-HE, C-NHE, O-SPE | |
| Circinella | Circinella muscae (syn. Circinella sydowii) | [55] | C-NHE |
| Cladophialophora | Cladophialophora tumulicola | [24,98] | C-HE |
| Cladosporium | Cladosporium cladosporioides | [22,28,29,30,31,32,38,41,46,49,51,57,59,68,69,85,99,100] | C-HE, NC-HE, C-NHE, O-SPE |
| Cladosporium cucumerinum | [46,49,51] | C-HE, C-NHE | |
| Cladosporium herbarum | [21,28,31,32,39,46,51,55,58,63,82] | C-HE, NC-HE, C-NHE | |
| Cladosporium macrocarpum | [60] | NC-HE | |
| Cladosporium sphaerospermum | [5,7,9,22,28,31,32,39,40,41,42,43,46,48,51,52,62,63,69] | C-HE, NC-HE, C-NHE, O-SPE | |
| Cladosporium uredinicola | [20,56] | O-SPE | |
| Cladosporium xylophilum | [60] | C-NHE | |
| Cladosporium sp. | [15,18,19,22,23,27,42,43,44,45,52,58,62,64,65,66,71,75,77,80,82,86,87,90,91,94,101,102,103] | All environments | |
| Clonostachys | Clonostachys rosea (syn. Gliocladium roseum) | [58] | C-HE |
| Cochliobolus | Cochliobolus geniculatus (syn. Curvulata geniculata) | [32,76] | NC-HE, C-NHE |
| Collariella | Collariella bostrychodes (syn. Chaetomium bostrychodes) | [28,32] | NC-HE |
| Coltricia | Coltricia sp. | [66] | C-NHE |
| Coprinellus | Coprinellus aokii (syn. Coprinus aokii) | [67] | SC-NHE |
| Coprinopsis | Coprinopsis atramentaria | [38] | C-HE |
| Coprinopsis cothurnata (syn. Coprinus cothurnatus) | [63] | C-NHE | |
| Cordyceps | Cordyceps farinosa (syn. Isaria farinosa) | [68] | C-HE |
| Coriolopsis | Coriolopsis sp. | [66] | C-NHE |
| Cunninghamella | Cunninghamella echinulata | [9,28,32,40,55] | NC-HE, C-NHE |
| Cunninghamella elegans | [38] | C-HE | |
| Cunninghamella sp. | [23,24] | C-HE | |
| Curvularia | Curvularia australiensis (syn. Drechslera australiensis) | [28,29,30,31,32] | NC-HE, C-NHE |
| Curvularia hawaiiensis (syn. Drechslera hawaiiensis) | [28,29,30,31,32] | NC-HE, C-NHE | |
| Curvularia lunata | [28,30,31,32,76,83] | NC-HE, C-NHE | |
| Curvularia pallescens | [29,30,31,32] | NC-HE, C-NHE | |
| Curvularia spicifera (syn. Drechslera spicifera) | [46] | C-NHE | |
| Curvularia sp. | [75] | C-NHE | |
| Cutaneotrichosporon | Cutaneotrichosporon mucoides (syn. Trichosporon mucoides) | [63] | C-NHE |
| Cylindrocarpon | Cylindrocarpon sp. | [23,24] | C-HE |
| Cyphellophora | Cyphellophora olivacea | [42] | C-HE |
| Cyphellophora sp. | [42] | C-HE | |
| Cyphellostereum | Cyphellostereum sp. | [66] | C-NHE |
| Cystoderma | Cystoderma sp. | [66] | C-NHE |
| Devriesia | Devriesia sp. | [45] | NC-HE |
| Dichotomophilus | Dichotomophilus indicus (syn. Chaetomium indicum) | [55] | C-NHE |
| Didymella | Didymella glomerata (syn. Phoma glomerata) | [21,40] | C-NHE |
| Dipodascus | Dipodascus geotrichum | [55] | C-NHE |
| Dipodascus sp. (syn. Geotrichum sp.) | [47,75] | NC-HE, C-NHE | |
| Discostroma | Discostroma corticola (syn. Seimatosporium lichenicola) | [20,56] | O-SPE |
| Drechslera | Drechslera sp. | [65] | O-SPE |
| Emericella | Emericella ruber | [31] | C-NHE |
| Emericella sp. | [49,75] | C-HE, C-NHE | |
| Engyodontium | Engyodontium sp. | [45,69] | C-HE, NC-HE, C-NHE |
| Epicoccum | Epicoccum nigrum (syn. Epicoccum purpurascens) | [20,28,31,32,46,49,56,58] | C-HE, NC-HE, C-NHE, O-SPE |
| Epicoccum sp. | [27,65,87,90] | C-HE, NC-HE, C-NHE, O-SPE | |
| Eurotium | Eurotium halophilicum | [104] | C-NHE |
| Eurotium herbariorum | [68] | C-HE | |
| Eurotium sp. | [47,65,89,92] | C-HE, NC-HE, C-NHE | |
| Exophiala | Exophiala angulospora | [42,98] | C-HE |
| Exophiala moniliae | [85] | C-HE | |
| Exophiala sp. | [23,42] | C-HE | |
| Fomitopsis | Fomitopsis vinosa | [63] | C-NHE |
| Fusarium | Fusarium chlamydosporum | [51] | C-HE |
| Fusarium culmorum | [31] | C-NHE | |
| Fusarium equiseti | [53] | SC-NHE | |
| Fusarium fujikuroi (syn. F. moniliforme) | [29,30,32,74] | C-HE, NC-HE | |
| Fusarium oxysporum | [23,28,31,32,38,46,49,58,67,73,105] | C-HE, NC-HE, C-NHE, SC-NHE | |
| Fusarium proliferatum | [77] | C-NHE | |
| Fusarium sporotrichioides | [38] | C-HE | |
| Fusarium sp. | [10,23,24,27,31,39,43,47,62,64,82,91,94,106] | C-HE, NC-HE, C-NHE | |
| Fuscoporia | Fuscoporia sp. | [66] | C-NHE |
| Fusidium | Fusidium viride | [48] | C-NHE |
| Ganoderma | Ganoderma sp. | [66] | C-NHE |
| Gliomastix | Gliomastix tumulicola (syn. Acremonium tumulicola) | [23,25] | C-HE |
| Gliomastix sp. | [58,97] | C-HE | |
| Gloiothele | Gloiothele sp. | [66] | C-NHE |
| Helminthosporium | Helminthosporium sp. | [65,75,82] | C-NHE, O-SPE |
| Humicola | Humicola fuscoatra | [49] | C-NHE |
| Humicola udagawae | [38] | C-HE | |
| Humicola sp. | [31] | C-NHE | |
| Hyphodontia | Hyphodontia alutaria | [66] | C-NHE |
| Hyphodontia sp. | [66] | C-NHE | |
| Hyphodontiella | Hyphodontiella sp. | [66] | C-NHE |
| Hypholoma | Hypholoma sp. | [66] | C-NHE |
| Idriella | Idriella sp. | [58] | C-HE |
| Kendrickiella | Kendrickiella phycomyces | [23,24,107] | C-HE |
| Kernia | Kernia geniculotricha | [67] | SC-NHE |
| Kernia hippocrepida | [67] | SC-NHE | |
| Lactarius | Lactarius sp. | [66] | C-NHE |
| Lecanicillium | Lecanicillium psalliotae | [38,59] | C-HE, NC-HE |
| Lecanicillium sp. | [68] | C-HE | |
| Leptobacillium | Leptobacillium muralicola | [37] | NC-HE |
| Leptosphaeria | Leptosphaeria sp. | [65] | O-SPE |
| Leptosphaerulina | Leptosphaerulina sp. | [64] | NC-HE |
| Macrophomina | Macrophomina phaseolina | [28,31,32] | NC-HE |
| Malbranchea | Malbranchea sp. | [46] | C-NHE |
| Mammaria | Mammaria echinobotryoides | [83] | C-HE |
| Memnoniella | Memnoniella sp. | [31] | C-NHE |
| Metapochonia | Metapochonia bulbillosa (syn. Verticillium bulbillosum) | [47] | C-HE, NC-HE |
| Metapochonia suchlasporia (syn. Verticillium suchlasporium) | [52] | NC-HE, C-NHE | |
| Meyerozyma | Meyerozyma guilliermondii | [60] | NC-HE |
| Microascus | Microascus brevicaulis (syn. Scopulariopsis brevicaulis) | [21,52,68] | C-HE, NC-HE, C-NHE |
| Microascus chartarum (syn. Scopulariopsis chartarum) | [62] | NC-HE, C-NHE | |
| Microascus cirrosus | [67] | SC-NHE | |
| Microascus sp. | [68] | C-HE | |
| Microdochium | Microdochium lycopodinum | [38] | C-HE |
| Monilinia | Monilinia sp. (syn. Monilia sp.) | [82] | C-NHE |
| Monocillium | Monocillium-like | [23] | C-HE |
| Monodictys | Monodictys castaneae (syn. Stemphylium macrosporoideum) | [21] | C-NHE |
| Monodictys sp. | [31,58] | C-HE, C-NHE | |
| Mortierell | Mortierella alpina | [47] | NC-HE |
| Mortierella ambigua | [38] | C-HE | |
| Mortierella parvispora (syn. M. gracilis) | [31] | C-NHE | |
| Mortierella sp. | [47,58] | C-HE, NC-HE | |
| Mucor | Mucor plumbeus (syn. M. spinosus) | [21] | C-NHE |
| Mucor racemosus (syn. M. globosus) | [28,32,47,60] | NC-HE | |
| Mucor silvaticus | [28,32] | NC-HE | |
| Mucor sp. | [23,46,58,64,82,93] | C-HE, NC-HE, C-NHE | |
| Myxotrichum | Myxotrichum stipitatum | [46] | C-NHE |
| Myxotrichum sp. | [46] | C-NHE | |
| Nectria | Nectria sp. | [15] | C-NHE |
| Neocosmospora | Neocosmospora solani (syn. Fusarium solani) | [23,28,31,76,97,105] | C-HE, NC-HE, C-NHE |
| Neodevriesia | Neodevriesia modesta (syn. Devriesia modesta) | [99,108] | O-SPE |
| Neodevriesia simplex (syn. Devriesia simplex) | [99,108] | O-SPE | |
| Neodevriesia sp. | [45] | NC-HE | |
| Neosartorya | Neosartorya sp. | [62] | NC-HE, C-NHE |
| Neosetophoma | Neosetophoma cerealis (syn. Coniothyrium cerealis) | [69] | C-NHE |
| Neurospora | Neurospora intermedia | [76] | C-NHE |
| Neurospora sp. | [82] | C-NHE | |
| Nigrospora | Nigrospora oryzae (syn. N. sphaerica) | [28,32,82] | C-HE, NC-HE, C-NHE |
| Nigrospora sp. | [28,82] | NC-HE, C-NHE | |
| Oidiodendron | Oidiodendron cereale | [69] | C-NHE |
| Oidiodendron tenuissimum | [49] | C-NHE | |
| Ophiostoma | Ophiostoma sp. | [23] | C-HE |
| Paecilomyces | Paecilomyces variotii | [28,29,30,31,32,61] | NC-HE, C-NHE |
| Paecilomyces sp. | [27,46,68,75,82,97] | C-HE, NC-HE, C-NHE | |
| Parengyodontium | Parengyodontium album (syn. Beauveria alba, Tritirachium album, and Engyodontium album) | [9,21,39,40,41,42,43,48,52,59,68,94,103,109,110,111] | C-HE, NC-HE, C-NHE |
| Penicillium | Penicillium aethiopicum | [67] | SC-NHE |
| Penicillium albicans | [93] | C-NHE | |
| Penicillium aurantiogriseum (syn. P. verrucosum var. cyclopium) | [9,41,43,51] | C-HE, C-NHE | |
| Penicillium brevicompactum | [39,40,46,68,69,94] | C-HE, NC-HE, C-NHE | |
| Penicillium camemberti | [93] | C-NHE | |
| Penicillium canescens (syn. P. raciborski) | [40,93] | C-NHE | |
| Penicillium carneum | [67] | SC-NHE | |
| Penicillium chrysogenum (syn. P. notatum) | [9,21,40,41,46,51,54,60,67,69,81,87,93,112,113] | C-HE, NC-HE, C-NHE, SC-NHE | |
| Penicillium citreonigrum (syn. P. citreoviride) | [22,58] | C-HE, O-SPE | |
| Penicillium citrinum | [31,32,40,58,76,93] | C-HE, NC-HE, C-NHE | |
| Penicillium commune | [51,53,68,93] | C-HE, C-NHE, SC-NHE | |
| Penicillium concentricum | [67] | SC-NHE | |
| Penicillium coprobium | [67] | SC-NHE | |
| Penicillium corylophilum | [44,91] | C-NHE | |
| Penicillium daleae | [91] | C-NHE | |
| Penicillium decumbens | [9,40,93] | C-NHE | |
| Penicillium dierckxii (syn. P. fellutanum) | [46] | C-NHE | |
| Penicillium digitatum | [66] | C-NHE | |
| Penicillium dipodomyicola | [67] | SC-NHE | |
| Penicillium expansum | [46,49] | C-NHE | |
| Penicillium fuscoglaucum | [60] | C-NHE | |
| Penicillium glabrum (syn. P. frequentans) | [9,39,46,49,66,93,94] | NC-HE, C-NHE | |
| Penicillium granulatum | [31] | C-NHE | |
| Penicillium griseofulvum | [46,56,57,67] | C-NHE, SC-NHE | |
| Penicillium herquei | [46] | C-NHE | |
| Penicillium italicum | [53] | SC-NHE | |
| Penicillium javanicum (syn. Eupenicillium javanicum) | [62] | NC-HE, C-NHE | |
| Penicillium lanosum | [20,56,100] | NC-HE, O-SPE | |
| Penicillium lilacinum | [21,55] | C-NHE | |
| Penicillium meleagrinum | [38,69] | C-HE, C-NHE | |
| Penicillium miczynskii | [47] | SC-NHE | |
| Penicillium olsonii | [51] | C-HE | |
| Penicillium oxalicum | [49,58] | C-HE, C-NHE | |
| Penicillium pancosmium | [38] | C-HE | |
| Penicillium paneum | [23,67,114] | C-HE, SC-NHE | |
| Penicillium polonicum | [51] | C-HE | |
| Penicillium purpurescens | [31] | C-NHE | |
| Penicillium purpurogenum | [69,93] | C-NHE | |
| Penicillium restrictum | [93] | C-NHE | |
| Penicillium simplicissimum (syn. P. janthinellum) | [58,93] | C-HE, C-NHE | |
| Penicillium spinulosum (syn. P. nigricans) | [9,40,84] | C-HE, C-NHE | |
| Penicillium thomii | [93] | C-NHE | |
| Penicillium turbatum | [53] | SC-NHE | |
| Penicillium verrucosum | [39,46] | C-NHE | |
| Penicillium vulpinum | [67] | SC-NHE | |
| Penicillium sp. | [11,15,18,19,23,24,26,28,31,43,44,45,46,47,48,58,61,64,66,70,75,77,78,80,82,86,88,89,90,91,94,101] | C-HE, NC-HE, C-NHE | |
| Pestalotia | Pestalotia sp. | [26] | C-NHE |
| Phialophora | Phialophora sp. | [23,24,43,47] | C-HE, C-NHE |
| Phlebia | Phlebia sp. | [66] | C-NHE |
| Pholiota | Pholiota sp. | [66] | C-NHE |
| Phoma | Phoma sp. | [23,31] | C-HE, C-NHE |
| Physalacria | Physalacria sp. | [66] | C-NHE |
| Pleospora | Pleospora sp. | [65] | O-SPE |
| Postia | Postia sp. | [66] | C-NHE |
| Preussia | Preussia terricola | [68] | C-HE |
| Preussia sp. | [68] | C-HE | |
| Pseudogymnoascus | Pseudogymnoascus pannorum (syn. Geomyces pannorum and Chrysosporium pannorum) | [39,43,48] | C-NHE |
| Pseudozyma | Pseudozyma prolifica | [51] | C-HE |
| Purpureocillium | Purpureocillium lilacinus (syn. Paecilomyces lilacinus) | [19,38,49] | C-HE, C-NHE |
| Pyrenophora | Pyrenophora biseptata (syn. Drechslera biseptata) | [28,32] | NC-HE |
| Radulomyces | Radulomyces sp. | [66] | C-NHE |
| Rhinocladiella | Rhinocladiella-like | [23] | C-HE |
| Rhizoctonia | Rhizoctonia solani (syn. Thanatephorus cucumeris) | [20,28,32,56] | NC-HE, O-SPE |
| Rhizopus | Rhizopus stolonifer (syn. R. nigricans) | [28,31,32,53,57,73,74] | C-HE, NC-HE, C-NHE |
| Rhizopus sp. | [27,101] | NC-HE, C-NHE | |
| Rhodotorula | Rhodotorula glutinis | [49] | C-NHE |
| Rhodotorula mucilaginosa | [66] | C-NHE | |
| Rhodotorula sp. | [15,18,66,69,80,91] | C-NHE | |
| Russula | Russula sp. | [66] | C-NHE |
| Sagenomella | Sagenomella griseoviridis | [24] | C-HE |
| Sagenomella striatispora | [24] | C-HE | |
| Sagenomella sp. | [44] | NC-HE | |
| Sarocladium | Sarocladium bacillisporum (syn. Acremonium bacillisporum) | [94] | NC-HE |
| Sarocladium kiliense (syn. Acremonium kiliense) | [21,52] | NC-HE, C-NHE | |
| Sarocladium strictum (syn. Acremonium cfr. strictum) | [23,52] | C-HE, NC-HE, C-NHE | |
| Schizophyllum | Schizophyllum commune | [64] | C-NHE |
| Schizophyllum sp. | [67] | C-NHE | |
| Schizopora | Schizopora paradoxa (syn. Hyphodontia paradoxa) | [64] | C-NHE |
| Scolecobasidium | Scolecobasidium anomalum (syn. Ochroconis anomala) | [115] | C-HE |
| Scolecobasidium lascauxensis | [85,115] | C-HE | |
| Scolecobasidium tshawytschae (syn. Ochroconis tshawytschae | [46] | C-NHE | |
| Scopulariopsis | Scopulariopsis brevicaulis | [21,55] | C-NHE |
| Scopulariopsis fusca | [39] | C-NHE | |
| Scopulariopsis sp. | [46,47,81,90] | NC-HE, C-NHE | |
| Scytalidium | Scytalidium sp. | [70,71] | C-NHE |
| Simplicillium | Simplicillium lamellicola (syn. Verticillium lamellicola) | [48] | C-NHE |
| Skeletocutis | Skeletocutis sp. | [67] | C-NHE |
| Sordaria | Sordaria humana | [28,32] | NC-HE |
| Sphaerostilbella | Sphaerostilbella sp. (syn. Gliocladium sp.) | [23,93,97] | C-HE, C-NHE |
| Sporothrix | Sporothrix sp. | [44,91] | NC-HE, C-NHE |
| Sporotrichum | Sporotrichum sp. | [41,43] | C-NHE |
| Stachybotrys | Stachybotrys chartarum (syn. S. atra) | [21,31,48,54,84] | NC-HE, C-NHE |
| Stachybotrys cylindrosporus | [21] | C-NHE | |
| Stachybotrys echinatus (syn. Memnoniella echinata) | [28,31,32] | NC-HE | |
| Stachybotrys sp. | [9,27,31] | NC-HE, C-NHE | |
| Stagonosporopsis | Stagonosporopsis lupini | [60] | C-NHE |
| Stemphylium | Stemphylium botryosum | [41] | C-NHE |
| Stemphylium pyriforme | [55] | C-NHE | |
| Stemphylium sp. | [93] | C-NHE | |
| Stereum | Stereum sp. | [66] | C-NHE |
| Syncephalastrum | Syncephalastrum sp. | [75] | C-NHE |
| Talaromyces | Talaromyces aculeatus | [51] | C-HE |
| Talaromyces flavus | [38,69] | C-HE | |
| Talaromyces pinophilus (syn. Penicillium pinophilum) | [54,94] | C-HE, NC-HE | |
| Talaromyces rugulosus (syn. Penicillium rugulosum) | [68,69,113] | C-HE, C-NHE | |
| Talaromyces variabilis (syn. Penicillium variabile) | [69] | C-NHE | |
| Thysanorea | Thysanorea papuana | [85] | C-HE |
| Tilletiopsis | Tilletiopsis sp. | [69] | C-NHE |
| Torrubiella | Torrubiella alba (syn. Lecanicillium aranearum) | [94] | NC-HE |
| Torrubiella sp. | [68,94] | C-HE, NC-HE | |
| Torula | Torula herbarum | [55] | C-NHE |
| Torula sp. | [46] | C-NHE | |
| Tricharina | Tricharina sp. | [64] | NC-HE |
| Trichocladium | Trichocladium asperum | [68] | C-HE |
| Trichoderma | Trichoderma harzianum | [29,30,31,32,69] | NC-HE, C-NHE |
| Trichoderma sect. Longibrachiatum | [23,105] | C-HE | |
| Trichoderma virens (syn. Gliocladium virens) | [58] | C-HE | |
| Trichoderma viride | [23,55,58] | C-HE, C-NHE | |
| Trichoderma sp. | [19,23,24,31,44,47,75,82,85,95,97] | C-HE, NC-HE, C-NHE | |
| Trichothecium | Trichothecium indicum (syn. Acremonium indicum) | [28,31,32] | NC-HE |
| Trichothecium roseum | [21,92] | C-NHE | |
| Tritirachium | Tritirachium sp. | [67,82] | C-HE, C-NHE |
| Tubaria | Tubaria sp. | [66] | C-NHE |
| Tyromyces | Tyromyces sp. | [66] | C-NHE |
| Umbelopsis | Umbelopsis ramanniana (syn. Mortierella ramanniana) | [39,47] | NC-HE, C-NHE |
| Venturia | Venturia carpophila (syn. Cladosporium carpophilum) | [32] | NC-HE |
| Verticillium | Verticillium alboatrum | [32] | NC-HE |
| Verticillium sp. | [23,48,52,58,97] | C-HE, NC-HE, C-NHE | |
| Wallemia | Wallemia sebi | [63] | C-NHE |
| Wallemia sp. | [92] | C-NHE | |
| Westerdykella | Westerdykella sp. | [64] | NC-HE |
| Xylodon | Xylodon nespoli | [66] | C-NHE |
| Xylodon nothofagi | [66] | C-NHE | |
| Xylodon raduloides | [66] | C-NHE | |
| Zygosporium | Zygosporium masoni | [23] | C-HE |
| Basidiomycota (Phylum) | [64] | NC-HE | |
| Black meristematic fungi | [5,99,116] | NC-HE, O-SPE | |
| Chaetomiaceae (Family) | [64] | NC-HE | |
| Filobasidiales | [64] | NC-HE | |
| Hyaline sterile mycelia | [62] | NC-HE, C-NHE | |
| Melanized sterile mycelia | [62] | NC-HE, C-NHE | |
| Pezizomycotina (Subphylum) | [64] | NC-HE | |
| Undetermined dark pigmented fungi | [17] | O-SPE | |
| Undetermined yeasts | [46] | C-NHE | |
| Uredinales (Order) | [65] | O-SPE | |
| Ustilaginales (Order) | [65] | O-SPE |
Table 2.
Contingency table and affinity indices. (A) Contingency table between the obtained clusters and the environmental categories to which they are correlated and (B) their relative affinity indices.
Table 2.
Contingency table and affinity indices. (A) Contingency table between the obtained clusters and the environmental categories to which they are correlated and (B) their relative affinity indices.
| (A) | Contingency Table | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clusters | ||||||||||||||||
| 13 | 11 | 14 | 3 | 6 | 15 | 8 | 2 | 4 | 5 | 1 | 10 | 9 | 7 | 12 | ||
| Environmental categories | C-HE | 1 | 5 | 3 | 2 | 3 | ||||||||||
| NC-HE | 2 | 1 | 1 | 6 | 1 | 1 | 1 | |||||||||
| C-NHE | 1 | 1 | 1 | 1 | 2 | 11 | 13 | 7 | 1 | 1 | 1 | |||||
| SC-NHE | 1 | 1 | 1 | |||||||||||||
| O-SPE | 2 | 1 | 1 | |||||||||||||
| (B) | Affinity Indices | |||||||||||||||
| Clusters | ||||||||||||||||
| 13 | 11 | 14 | 3 | 6 | 15 | 8 | 2 | 4 | 5 | 1 | 10 | 9 | 7 | 12 | ||
| Environmental categories | C-HE | 1.82 | 4.8 | 1.37 | 1.46 | 8.2 | ||||||||||
| NC-HE | 3.93 | 1.31 | 0.21 | 5.89 | 0.39 | 0.98 | 3.93 | |||||||||
| C-NHE | 1.28 | 1.28 | 1.28 | 0.64 | 1.7 | 11 | 8.98 | 6.25 | 1.28 | 1.28 | 1.28 | |||||
| SC-NHE | 17 | 1 | 0.71 | |||||||||||||
| O-SPE | 12.8 | 0.67 | 0.53 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zucconi, L.; Canini, F.; Isola, D.; Caneva, G. Fungi Affecting Wall Paintings of Historical Value: A Worldwide Meta-Analysis of Their Detected Diversity. Appl. Sci. 2022, 12, 2988. https://doi.org/10.3390/app12062988
AMA Style
Zucconi L, Canini F, Isola D, Caneva G. Fungi Affecting Wall Paintings of Historical Value: A Worldwide Meta-Analysis of Their Detected Diversity. Applied Sciences. 2022; 12(6):2988. https://doi.org/10.3390/app12062988
Chicago/Turabian StyleZucconi, Laura, Fabiana Canini, Daniela Isola, and Giulia Caneva. 2022. "Fungi Affecting Wall Paintings of Historical Value: A Worldwide Meta-Analysis of Their Detected Diversity" Applied Sciences 12, no. 6: 2988. https://doi.org/10.3390/app12062988
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.