
Phytochemical Analysis and Dermo-Cosmetic Evaluation of Cymbidium sp. (Orchidaceae) Cultivation By-Products

 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Plant Material
2.3. Plant Extraction
2.4. Chromatographic Fractionation of Ethyl Acetate Extracts from Pseudobulbs and Roots
2.5. TLC, HPLC, and NMR Analysis
2.6. UPLC-HRMS Analysis
2.7. DPPH
2.8. Tyrosinase Assay
2.9. Elastase Assay
2.10. Collagenase Assay
2.11. Statistical Analysis
3. Results
3.1. In Vitro Biological Evaluation of Cymbidium sp. By-Products’ Extracts
3.1.1. Antioxidant Potential of Cymbidium sp. Total Extracts
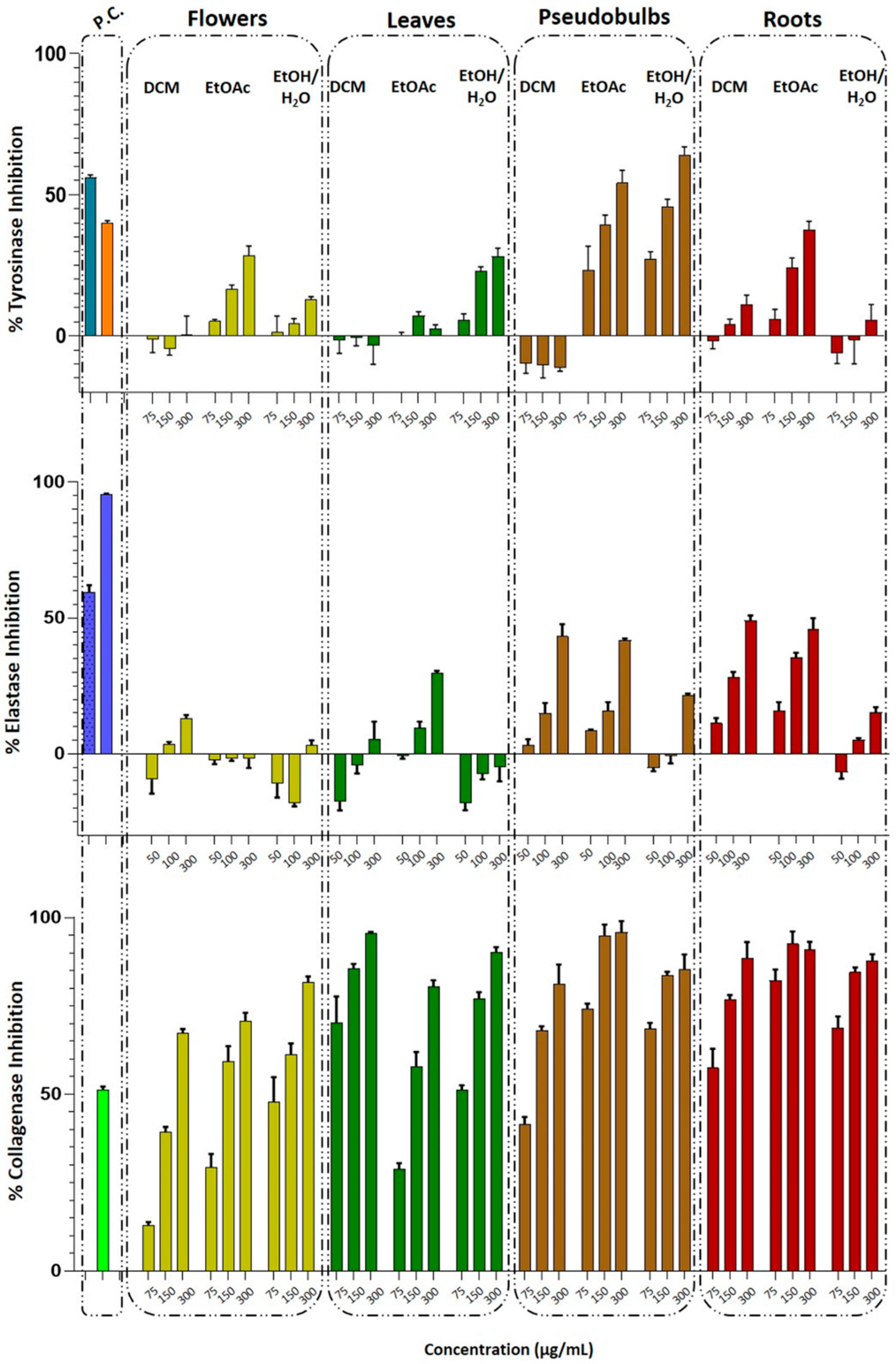
3.1.2. Determination of Anti-Tyrosinase, Anti-Elastase, and Anti-Collagenase Activity of Total Extracts
3.2. Phytochemical Analysis of Pseudobulb and Root EtOAc Extracts
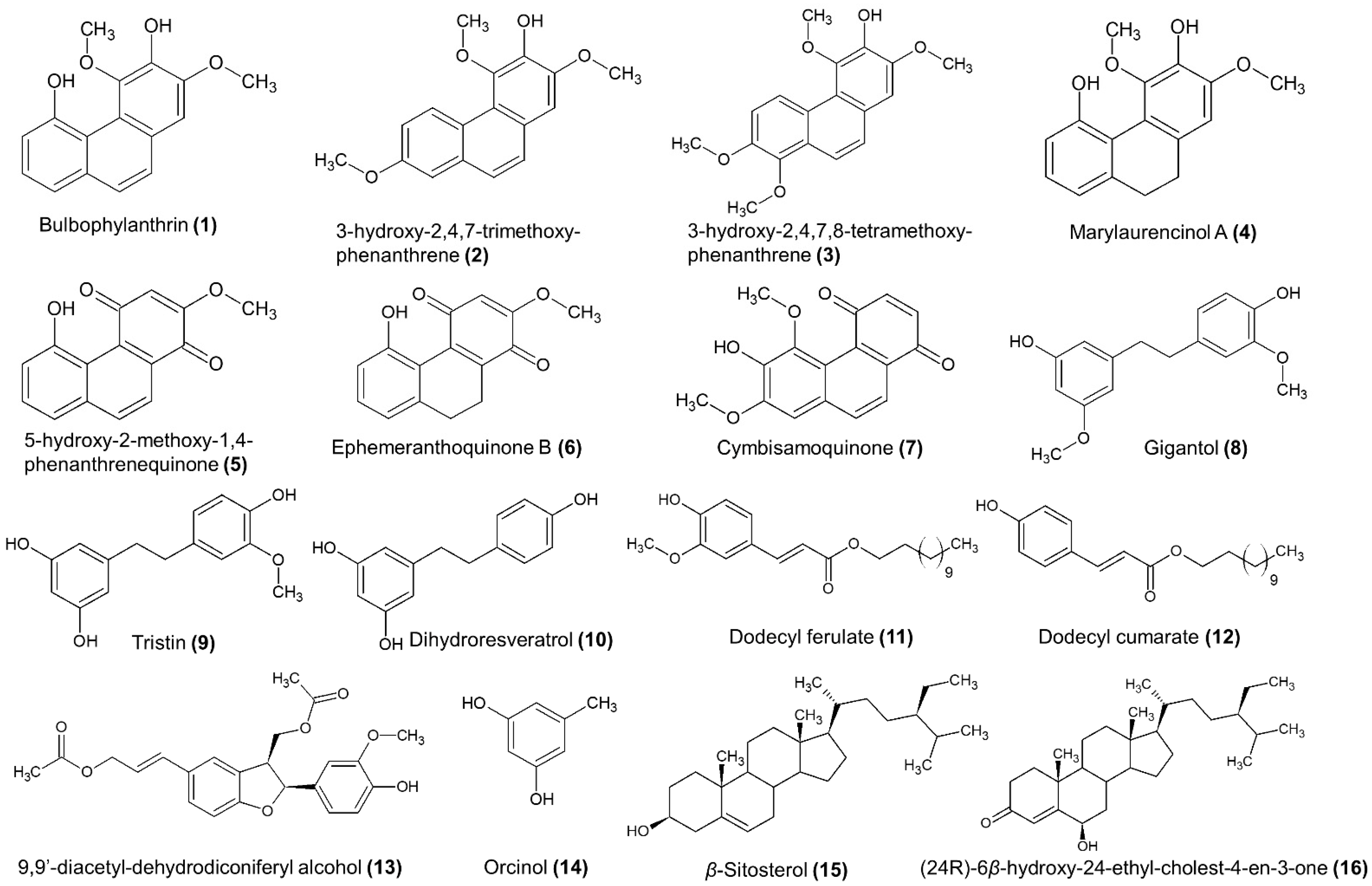
3.2.1. Isolation of the Main Secondary Metabolites
3.2.2. UPLC-HRMS/MS Analysis of Pseudobulb and Root Ethyl Acetate Extracts
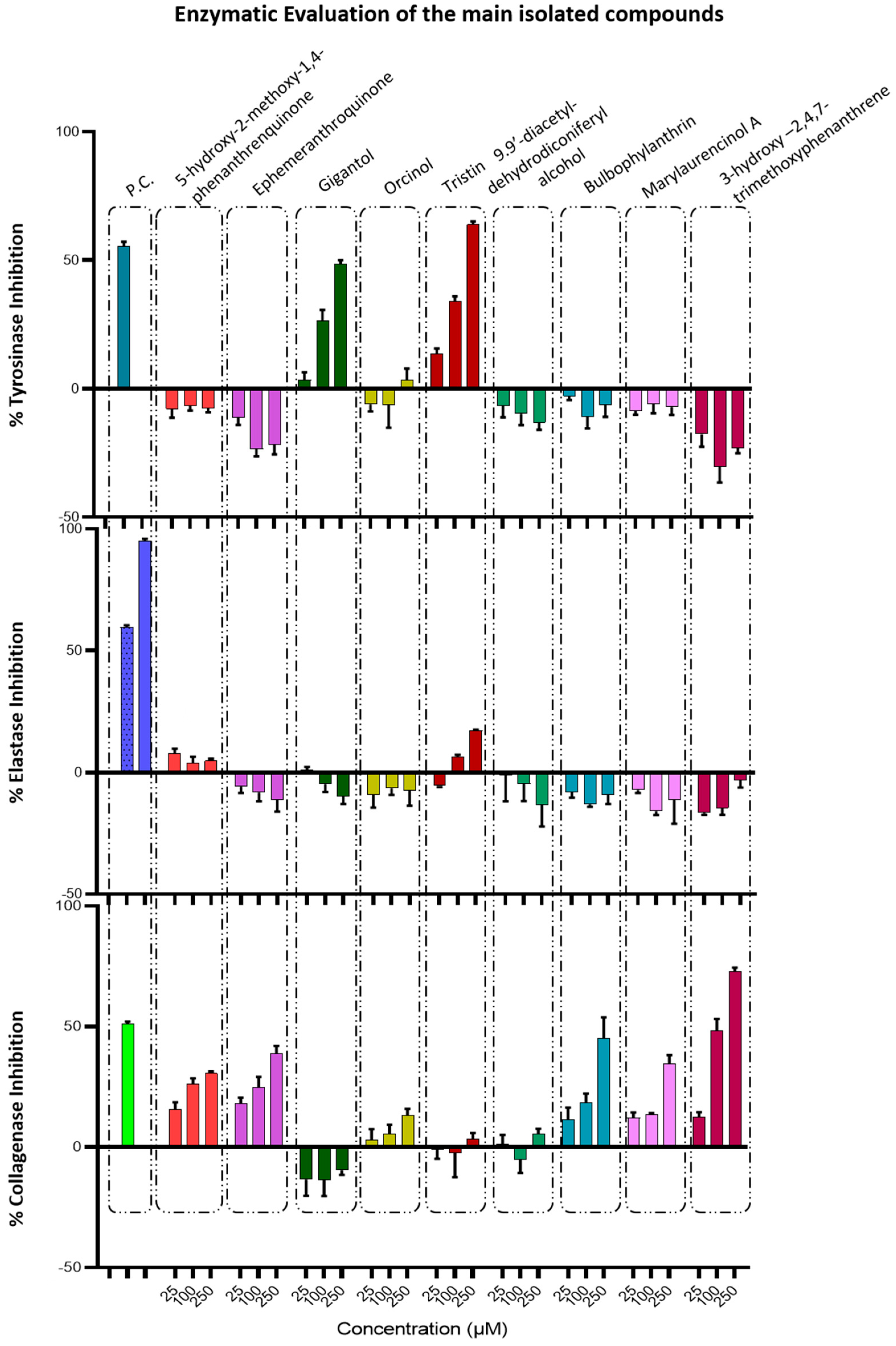
3.2.3. Determination of Anti-Tyrosinase, Anti-Elastase, and Anti-Collagenase Activity of Pure Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, R.M.P. Orchids: A review of uses in traditional medicine, its phytochemistry and pharmacology. J. Med. Plants Res. 2010, 4, 592–638. [Google Scholar]
- Hossain, M.M. Therapeutic orchids: Traditional uses and recent advances-An overview. Fitoterapia 2011, 82, 102–140. [Google Scholar] [CrossRef] [PubMed]
- The Plant List. 2020. Available online: http://www.theplantlist.org/ (accessed on 28 December 2021).
- Bulpitt, C.J.; Li, Y.; Bulpitt, P.F.; Wang, J. The use of orchids in Chinese medicine. J. R. Soc. Med. 2007, 100, 558–563. [Google Scholar] [CrossRef]
- Dioscorides, P. De Materia Medica; Frankfurt, Ed.; 1543 or De Materia Medica libra quinque 572 (In Imperial Library at Vienna); IBIDIS Press: Johannesburg, South Africa, 2000. [Google Scholar]
- Szlachetko, D.L. Genera et species Orchidalium.1. Pol. Bot. J. 2001, 46, 11–26. [Google Scholar]
- Barua, A.-K.; Ghosh, B.-B.; Ray, S.; Patra, A. Cymbinodin-A, a phenanthraquinone from Cymbidium Aloifolium. Phytochemistry 1990, 29, 3046–3047. [Google Scholar] [CrossRef]
- Blomqvist, L.; Leander, K.; Lüning, B.; Rosenblom, J. Studies on Orchidaceae Alkaloids XXIX. The absolute Configuration of Dendroprimine, an Alkaloid from Dendrobium primulinum Lindl. Acta Chem. Scand. 1972, 26, 3202–3326. [Google Scholar] [CrossRef]
- Chen, X.-J.; Mei, W.-L.; Cai, C.-H.; Guo, Z.-K.; Song, X.-Q.; Dai, H.-F. Four new bibenzyl derivatives from Dendrobium sinense. Phytochem. Lett. 2014, 9, 107–112. [Google Scholar] [CrossRef]
- Garcia-Gaytan, V.; Mendoza, M.D.L.N.R.; Hernández, M.S.; Trejo-Téllez, L.I.; Santos, M.E.P.; Ponce, G.V. Volatile Components in the Flower, Pedicellate Ovary and Aqueous Residue of Cymbidium sp. (ORCHIDACEAE). J. Anal. Sci. Methods Instrum. 2013, 3, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Majumder, P.; Sen, R. Pendulin, a polyoxygenated phenanthrene derivative from the orchid Cymbidium pendulum. Phytochemistry 1991, 30, 2432–2434. [Google Scholar] [CrossRef]
- Sut, S.; Maggi, F.; Dall’Acqua, S. Bioactive Secondary Metabolites from Orchids (Orchidaceae). Chem. Biodivers. 2017, 14, e1700172. [Google Scholar] [CrossRef]
- Hooker, J.D. Cymbidium and Cyperorchis. In Flora of British India; Reeve & Co: London, UK, 1890; Volume 6, pp. 8–15. [Google Scholar]
- Schlechter, R. Die Gattungen Cymbidium Sw. und Cyperorchis Bl. Repert. Nov. Specierum Regni Veg. 1924, 20, 96–110. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Saito, N.; Yokoi, M. Anthocyanins in the flowers of Cymbidium. Lindleyana 1996, 11, 214–219. [Google Scholar]
- Yoshikawa, K.; Ito, T.; Iseki, K.; Baba, C.; Imagawa, H.; Yagi, Y.; Morita, H.; Asakawa, Y.; Kawano, S.; Hashimoto, T. Phenanthrene Derivatives from Cymbidium Great Flower Marie Laurencin and Their Biological Activities. J. Nat. Prod. 2012, 75, 605–609. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Otsu, M.; Ito, T.; Asakawa, Y.; Kawano, S.; Hashimoto, T. Aromatic constituents of Cymbidium Great Flower Marie Laurencin and their antioxidative activity. J. Nat. Med. 2012, 67, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Baba, C.; Iseki, K.; Ito, T.; Asakawa, Y.; Kawano, S.; Hashimoto, T. Phenanthrene and phenylpropanoid constituents from the roots of Cymbidium Great Flower ‘Marylaurencin’ and their antimicrobial activity. J. Nat. Med. 2014, 68, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-J.; Cha, H.-S.; Lee, M.-H.; Kim, S.-Y.; Kim, S.H.; Kim, T.-J. Effects of Cymbidium Root Ethanol Extract on Atopic Dermatitis. Evid.-Based Complement. Altern. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Howlader, A.; Alam, M.; Ahmed, K.T.; Khatun, F.; Apu, A.S. Antinociceptive and Anti-inflammatory Activity of the Ethanolic Extract of Cymbidium aloifolium (L.). Pak. J. Biol. Sci. 2011, 14, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Lertnitikul, N.; Pattamadilok, C.; Chansriniyom, C.; Suttisri, R. A new dihydrophenanthrene from Cymbidium finlaysonianum and structure revision of cymbinodin-A. J. Asian Nat. Prod. Res. 2018, 22, 83–90. [Google Scholar] [CrossRef]
- Yoneyama, T.; Iseki, K.; Noji, M.; Imagawa, H.; Hashimoto, T.; Kawano, S.; Baba, M.; Kashiwada, Y.; Yahagi, T.; Matsuzaki, K.; et al. Marylosides A-G, Norcycloartane Glycosides from Leaves of Cymbidium Great Flower ‘Marylaurencin’. Molecules 2019, 24, 2504. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Wang, Z.; Su, Z.; Song, J.; Zhou, L.; Sun, Q.; Liu, S.; Li, S.; Li, Y.; Wang, M.; et al. Gigantol inhibits Wnt/β-catenin signaling and exhibits anticancer activity in breast cancer cells. BMC Complement. Altern. Med. 2018, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Axiotis, E.; Petrakis, E.A.; Halabalaki, M.; Mitakou, S. Phytochemical Profile and Biological Activity of Endemic Sideritis sipylea Boiss. in North Aegean Greek Islands. Molecules 2020, 25, 2022. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and Instrumental Approaches to Treat Hyperpigmentation. Pigment. Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Ishida, K. Biological Mechanisms Underlying the Ultraviolet Radiation-Induced Formation of Skin Wrinkling and Sagging I: Reduced Skin Elasticity, Highly Associated with Enhanced Dermal Elastase Activity, Triggers Wrinkling and Sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, J.; Angelis, A.; Aligiannis, N.; Rosalia, M.; Abedini, A.; Bakiri, A.; Reynaud, R.; Nuzillard, J.M.; Cangloff, S.-C.; Skaltsounis, A.L.; et al. In Vitro Dermo-Cosmetic Evaluation of Bark Extracts from Common Temperate Trees. Planta Med. 2016, 82, 1351–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidrianny, I.; Anggraeni, N.A.S.; Insanu, M. Antioxidant properties of peels extracts from three varieties of banana (Musa sp.) grown in West Java-Indonesia. Int. Food Res. J. 2018, 25, 57–64. [Google Scholar]
- Angelis, A.; Mavros, P.; Nikolaou, P.E.; Mitakou, S.; Halabalaki, M.; Skaltsounis, L. Phytochemical analysis of olive flowers’ hydroalcoholic extract and in vitro evaluation of tyrosinase, elastase and collagenase inhibition activity. Fitoterapia 2020, 143, 104602. [Google Scholar] [CrossRef]
- Majumder, P.; Kar, A.; Shoolery, J. Bulbophyllanthrin, a phenanthrene of the orchid Bulbophyllum leopardium. Phytochemistry 1985, 24, 2083–2087. [Google Scholar] [CrossRef]
- Liang, W.; Guo, X.; Nagle, D.G.; Zhang, W.-D.; Tian, X.-H. Genus Liparis: A review of its traditional uses in China, phytochemistry and pharmacology. J. Ethnopharmacol. 2019, 234, 154–171. [Google Scholar] [CrossRef]
- Zhou, D.; Chang, W.; Liu, B.; Chen, G.; Yang, Y.; Hao, Y.; Hou, Y.; Li, N. Stilbenes from the tubers of Bletilla striata with potential anti-neuroinflammatory activity. Bioorg. Chem. 2020, 97, 103715. [Google Scholar] [CrossRef]
- Langat, M.K.; Ndunda, B.M.; Salter, C.; Odusina, B.O.; Isyaka, S.M.; Mas-Claret, E.; Onocha, P.A.; Midiwo, J.O.; Nuzillard, J.-M.; Mulholland, D.A. Diterpenoids from the stem bark of Croton megalocarpoides Friis & M. G. Gilbert. Phytochem. Lett. 2020, 39, 1–7. [Google Scholar] [CrossRef]
- Kang, Y.-F.; Liu, C.-M.; Kao, C.-L.; Chen, C.-Y. Antioxidant and Anticancer Constituents from the Leaves of Liriodendron tulipifera. Molecules 2014, 19, 4235–4245. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhan, R.; Jiang, J.; Zhang, Y.; Dong, Y.; Chen, Y. A new ent-kaurane diterpenoid from Ixora amplexicaulis. Nat. Prod. Res. 2016, 30, 105–109. [Google Scholar] [CrossRef]
- Zhan, R.; Zhang, Y.; Chen, L.; Chen, Y. A new (propylphenyl)bibenzyl from Eria bambusifolia. Nat. Prod. Res. 2016, 30, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Dang, P.H.; Nguyen, H.X.; Le, T.H.; Van Do, T.N.; Nguyen, M.T.T. A new lignan from the stems of Buchanania lucida Blume (Anacardiaceae). Nat. Prod. Res. 2021, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lopes, S.P.; Yepes, L.M.; Pérez-Castillo, Y.; Robledo, S.M.; de Sousa, D. Alkyl and Aryl Derivatives Based on p-Coumaric Acid Modification and Inhibitory Action against Leishmania braziliensis and Plasmodium falciparum. Molecules 2020, 25, 3178. [Google Scholar] [CrossRef]
- Vansteelandt, M.; Blanchet, E.; Egorov, M.; Petit, F.; Toupet, L.; Bondon, A.; Monteau, F.; Le Bizec, B.; Thomas, O.P.; Pouchus, Y.F.; et al. Ligerin, an Antiproliferative Chlorinated Sesquiterpenoid from a Marine-Derived Penicillium Strain. J. Nat. Prod. 2013, 76, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sung, S.H.; Kim, Y.C. Antifibrotic Phenanthrenes of Dendrobium nobile Stems. J. Nat. Prod. 2007, 70, 1925–1929. [Google Scholar] [CrossRef]
- Won, J.-H.; Kim, J.-Y.; Yun, K.-J.; Lee, J.-H.; Back, N.-I.; Chung, H.-G.; Chung, S.A.; Jeong, T.-S.; Choi, M.-S.; Lee, K.-T. Gigantol Isolated from the Whole Plants of Cymbidium goeringii. Inhibits the LPS-Induced iNOS and COX-2 Expression via NF-κB Inactivation in RAW 264.7 Macrophages Cells. Planta Med. 2006, 72, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Deng, Q.; Zhang, Q.; Ma, Z.; Chen, B.; Yu, X.; Peng, H.; Yao, S.; Liu, J.; Ye, Y.; et al. Gigantol ameliorates CCl4-induced liver injury via preventing activation of JNK/cPLA2/12-LOX inflammatory pathway. Sci. Rep. 2020, 10, 22265. [Google Scholar] [CrossRef]
- Bi, Z.-M.; Wang, Z.-T.; Xu, L.-S.; Xu, G.-J. Study on phenolic constituents of Pholidota yunnanensis. J. Chin. Pharm. Sci. 2005, 40, 255–258. [Google Scholar]
- Ren, J.; Qian, X.-P.; Guo, Y.-G.; Li, T.; Yan, S.-K.; Jin, H.-Z.; Zhang, W.-D. Two new phenanthrene glycosides from Liparis regnieri Finet and their antibacterial activities. Phytochem. Lett. 2016, 18, 64–67. [Google Scholar] [CrossRef]
- Guo, X.-Y.; Wang, J.; Wang, N.-L.; Kitanaka, S.; Liu, H.-W.; Yao, X.-S. New Stilbenoids from Pholidota yunnanensis and Their Inhibitory Effects on Nitric Oxide Production. Chem. Pharm. Bull. 2006, 54, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Q.; Guo, Y.-Q.; Gao, W.-Y.; Zhang, T.-J.; Yan, L.-L. Two new phenanthrofurans from Pleione bulbocodioides. J. Asian Nat. Prod. Res. 2008, 10, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Duffhues, G.; Calzado, M.A.; de Vinuesa, A.G.; Caballero, F.J.; Ech-Chahad, A.; Appendino, G.B.; Krohn, K.; Fiebich, B.L.; Muñoz, E. Denbinobin, a naturally occurring 1,4-phenanthrenequinone, inhibits HIV-1 replication through an NF-κB-dependent pathway. Biochem. Pharmacol. 2008, 76, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.; Berthon, J.Y. Resveratrol: An original mechanism on tyrosinase inhibition. Int. J. Cosmet. Sci. 2000, 22, 219–226. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part of the Plant | Extraction Solvent | Extract Weight (g) | Yield (% w/w) |
|---|---|---|---|
| Flowers | DCM | 6.97 | 1.74 |
| EtOAc | 6.46 | 1.61 | |
| EtOH/H2O | 11.09 | 2.77 | |
| Leaves | DCM | 8.36 | 2.09 |
| EtOAc | 7.81 | 1.95 | |
| EtOH/H2O | 11.3 | 2.82 | |
| Pseudobulbs | DCM | 7.15 | 1.79 |
| EtOAc | 3.69 | 0.92 | |
| EtOH/H2O | 14.83 | 3.70 | |
| Roots | DCM | 2.06 | 0.65 |
| EtOAc | 3.68 | 0.92 | |
| EtOH/H2O | 9.54 | 2.39 |
| Part of Plant | Extraction Solvent | IC50 (μg/mL) |
|---|---|---|
| Flowers (Fl) | DCM | NA |
| EtOAc | NA | |
| EtOH/H2O | NA | |
| Leaves (L) | DCM | NA |
| EtOAc | NA | |
| EtOH/H2O | NA | |
| Pseudobulbs (Ps) | DCM | 165.78 |
| EtOAc | 114.18 | |
| EtOH/H2O | 233.32 | |
| Roots (R) | DCM | 174.61 |
| EtOAc | 127.17 | |
| EtOH/H2O | 304.02 |
| No. | Rt (min) | Experimental [M-H]− | Suggested EC | Delta (ppm) | RDB Eq. | Compound | EtOAc Extract | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| Ps | R | ||||||||
| Phenols, Phenolic acids, and derivatives | |||||||||
| 1 | 3.74 | 123.0453 | C7H7O2 | 1.44 | 4.5 | Orcinol (14) | + | - | * |
| 2 | 4.12 | 137.0245 | C7H5O3 | 0.97 | 5.5 | Salicylic acid | + | + | [1] |
| 3 | 4.14 | 137.0245 | C7H5O3 | 0.9 | 5.5 | p-hydroxybenzoic acid | + | + | [1] |
| 4 | 4.17 | 353.0883 | C16H17O9 | 1.46 | 8.5 | Chlorogenic acid | tr | tr | [24] |
| 5 | 5.18 | 193.0508 | C10H9O4 | 0.77 | 6.5 | Ferulic acid | - | tr | [1] |
| 6 | 5.37 | 163.0401 | C9H7O3 | 0.75 | 6.5 | p-coumaric acid | tr | tr | [1] |
| 7 | 5.55 | 151.0403 | C8H7O3 | 1.54 | 5.5 | Vanillin | tr | tr | [29] |
| 8 | 5.58 | 167.0352 | C8H7O4 | 1.49 | 5.5 | Vanillic acid | tr | tr | [44] |
| 9 | 7.85 | 153.0152 | C7H5O4 | 1.23 | 5.5 | Protocatechuic acid | + | + | [44] |
| 10 | 13.9 | 441.2648 | C27H37O5 | 0.821 | 9.5 | 9,9′-diacetyl-dehydrodiconiferyl alcohol (13) | tr | + | * |
| 11 | 14.06 | 361.2388 | C22H33O4 | 1.02 | 6.5 | Dodecyl ferulate (11) | + | + | * |
| 12 | 14.86 | 331.2283 | C21H31O3 | 1.21 | 6.5 | Dodecyl coumarate (12) | + | + | * |
| Dibenzyls and Stilbens | |||||||||
| 13 | 6.99 | 259.0976 | C15H15O4 | 0.07 | 8.5 | Tristin (9) | + | + | * |
| 14 | 7.11 | 229.0872 | C14H13O3 | 0.67 | 8.5 | Dihydroresveratrol (10) | + | tr | * |
| 15 | 7.4 | 259.0976 | C15H15O4 | 0.3 | 8.5 | Dendrosinen C | + | + | [36] |
| 16 | 8.66 | 273.1135 | C16H17O4 | 1.09 | 8.5 | Gigantol (8) | + | + | * |
| 17 | 8.7 | 289.1082 | C16H17O5 | 0.357 | 8.5 | Dendrosinen A | + | + | [36] |
| 18 | 8.72 | 243.1028 | C15H15O3 | 0.67 | 8.5 | Batatasin III | + | + | [12] |
| 19 | 8.85 | 227.0714 | C14H11O3 | 0.54 | 9.5 | Resveratrol | + | + | [12] |
| 20 | 9.77 | 301.1083 | C17H17O5 | 0.11 | 9.5 | Phoyunbene A or Phoyunbene B | + | + | [45] |
| 21 | 10.0 | 241.0871 | C15H13O3 | −0.24 | 9.5 | Thunalbene, Pholidotol D, Lusianthridin | tr | tr | [12] |
| Phenanthrenes and 1,4-phenanthrenquinones | |||||||||
| 22 | 5.85 | 243.0662 | C14H11O4 | 0.115 | 9.5 | Tetrahydroxy-9,10-dehydrophenanthrene | tr | + | |
| 23 | 7.43 | 283.0615 | C16H11O5 | 0.93 | 11.5 | Denbinobin, Isoxoflaccidin | + | + | [46] |
| 24 | 7.58 | 287.0925 | C16H15O5 | 1.26 | 9.5 | Coeloginanthridin, or 1,3,5-trihydroxy-2,4-dimethoxy-9,10-dihydrophenanthrene | tr | + | [2] |
| 25 | 7.77 | 253.0508 | C15H9O4 | 1.06 | 11.5 | 5-hydroxy-2-methoxy-1,4-phenanthrenequinone (5) | + | + | * |
| 26 | 8.16 | 283.0615 | C16H11O5 | 0.93 | 11.5 | Cymbisamoquinone (7) | + | + | * |
| 27 | 8.39 | 257.0817 | C15H13O4 | −0.90 | 9.5 | Trihydroxy-methoxy-9,10-dihydrophenanthrene | tr | + | [1] |
| 28 | 9.55 | 283.0977 | C17H15O4 | 0.38 | 10.5 | 3-hydroxy-2,4,7-trimethoxy-phenanthrene (2) | tr | + | * |
| 29 | 9.89 | 269.0821 | C16H13O4 | 0.81 | 10.5 | Bulbophyllanthrin (1) | tr | + | * |
| 30 | 10.01 | 271.0987 | C16H15O4 | 0.84 | 9.5 | Marylaurencinol A (4) | tr | + | * |
| 31 | 10.04 | 241.0871 | C15H13O3 | 0.25 | 9.5 | Hircinol or Coelonin | tr | + | [7] |
| 32 | 10.06 | 479.1506 | C30H23O6 | 1.228 | 19.5 | Blestriarene B | + | + | [2] |
| 33 | 10.4 | 255.0665 | C15H11O4 | 0.62 | 10.5 | Ephemeranthroquinone B (6) | + | + | * |
| 34 | 10.45 | 313.1084 | C17H15O4 | 0.71 | 10.5 | 3-hydroxy-2,4,7,8-tetramethoxy-phenanthrene (3) | tr | + | * |
| 35 | 11.41 | 461.1608 | C27H25O7 | 0.875 | 15.5 | Pleionesins C | + | + | [47] |
| Flavonoids | |||||||||
| 36 | 7.10 | 285.0407 | C15H9O6 | 0.87 | 11.5 | Kaempherol | + | tr | [2] |
| 37 | 7.10 | 285.0407 | C15H9O6 | 0.87 | 11.5 | Luteolin | + | tr | [29] |
| 38 | 7.18 | 301.0357 | C15H9O7 | 0.88 | 11.5 | Quercetin | + | tr | [2] |
| 39 | 7.29 | 269.0456 | C15H9O5 | 0.38 | 11.5 | Apigenin | + | tr | [26] |
| 40 | 7.39 | 299.0563 | C16H11O6 | 0.76 | 11.5 | Diosmetin | + | tr | [29] |
| 41 | 8.03 | 303.0513 | C15H11O7 | 1.49 | 10.5 | Taxifolin | + | tr | [29] |
| Other compounds | |||||||||
| 42 | 0.79 | 195.0512 | C6H11O7 | 0.9 | 1.5 | Gluconic acid or galactonic acid | + | + | [29] |
| 43 | 0.88 | 133.0143 | C4H5O5 | 0.85 | 2.5 | Malic acid | + | + | [29] |
| 44 | 1.10 | 191.0199 | C6H7O7 | 1.28 | 3.5 | Citric acid | tr | tr | [29] |
| 45 | 6.67 | 312.1243 | C18H18O4N | 0.636 | 10.5 | Moupinamide | + | + | [1] |
| 46 | 6.67 | 282.1138 | C17H16O3N | 1.04 | 10.5 | Paprazine | + | + | [1] |
| 47 | 7.03 | 342.1349 | C19H20O5N | 0.655 | 10.5 | n-trans-feruloyl 4′ O methyldopamine | + | + | [1] |
| 48 | 15.18 | 455.3532 | C30H47O3 | 0.44 | 7.5 | Oleanolic acid | + | tr | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Axiotis, E.; Angelis, A.; Antoniadi, L.; Petrakis, E.A.; Skaltsounis, L.A. Phytochemical Analysis and Dermo-Cosmetic Evaluation of Cymbidium sp. (Orchidaceae) Cultivation By-Products. Antioxidants 2022, 11, 101. https://doi.org/10.3390/antiox11010101
Axiotis E, Angelis A, Antoniadi L, Petrakis EA, Skaltsounis LA. Phytochemical Analysis and Dermo-Cosmetic Evaluation of Cymbidium sp. (Orchidaceae) Cultivation By-Products. Antioxidants. 2022; 11(1):101. https://doi.org/10.3390/antiox11010101
Chicago/Turabian StyleAxiotis, Evangelos, Apostolis Angelis, Lemonia Antoniadi, Eleftherios A. Petrakis, and Leandros A. Skaltsounis. 2022. "Phytochemical Analysis and Dermo-Cosmetic Evaluation of Cymbidium sp. (Orchidaceae) Cultivation By-Products" Antioxidants 11, no. 1: 101. https://doi.org/10.3390/antiox11010101