
Diversity and Pathogenicity of Botryosphaeriaceae and Phytophthora Species Associated with Emerging Olive Diseases in Italy

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Surveys and Sampling
2.2. Isolation of Fruit Rot Agents
2.3. Isolation of Branch Canker Agents
2.4. Isolation of Collar and Root Rot Agents
2.5. Identification of the Isolates
2.6. Pathogenicity Assays
2.7. Data Analysis
3. Results
3.1. Field Surveys
3.2. Fungal and Fungal-Like Species Associated with Symptomatic Samples
3.3. Pathogenicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tous, J.; Ferguson, L. Mediterranean fruits. Prog. New Crops 1996, 416, 430. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World, 2nd ed.; Claredon Press: Oxford, UK, 1994; pp. 137–143. [Google Scholar]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Terral, J.F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigg, D. Olive oil, the Mediterranean and the world. GeoJournal 2001, 53, 163–172. [Google Scholar] [CrossRef]
- Vossen, P. Olive oil: History, production, and characteristics of the world’s classic oils. HortScience 2007, 42, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.D. Olive entrepreneurship in the southern hemisphere: The case of New Zealand. Int. J. Rural Manag. 2008, 4, 67–85. [Google Scholar] [CrossRef]
- Torres, M.; Pierantozzi, P.; Searles, P.; Rousseaux, M.C.; García-Inza, G.; Miserere, A.; Bodoira, R.; Contreras, C.; Maestri, D. Olive Cultivation in the Southern Hemisphere: Flowering, Water Requirements and Oil Quality Responses to New Crop Environments. Front. Plant Sci. 2017, 8, 1830. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, D.; Farooqi, T.J.A.; Ma, L.; Deng, Y.; Jia, Z. The olive (Olea europaea L.) industry in China: Its status, opportunities and challenges. Agrofor. Syst. 2019, 93, 395–417. [Google Scholar] [CrossRef]
- The Food and Agriculture Organization. Crops. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 18 July 2023).
- ISTAT, Istituto Nazionale di Statistica. Available online: https://www.istat.it/en/ (accessed on 18 July 2023).
- Daane, K.M.; Johnson, M.W. Olive fruit fly: Managing an ancient pest in modern times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Nigro, F.; Boscia, D.; Antelmi, I.; Ippolito, A. Fungal species associated with severe decline of olive in southern Italy. J. Plant Pathol. 2013, 95, 668. [Google Scholar]
- Úrbez-Torres, J.R.; Peduto, F.; Vossen, P.M.; Krueger, W.H.; Gubler, W.D. Olive twig and branch dieback: Etiology, incidence, and distribution in California. Plant Dis. 2013, 97, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Diaz, R.M.; Cirulli, M.; Bubici, G.; Del Mar Jimenez-Gasco, M.; Antoniou, P.P.; Tjamos, E.C. Verticillium wilt, a major threat to olive production: Current status and future prospects for its management. Plant Dis. 2012, 96, 304–329. [Google Scholar] [CrossRef] [Green Version]
- Talhinhas, P.; Loureiro, A.; Oliveira, H. Olive anthracnose: A yield-and oil quality-degrading disease caused by several species of Colletotrichum that differ in virulence, host preference and geographical distribution. Mol. Plant Pathol. 2018, 19, 1797–1807. [Google Scholar] [CrossRef] [Green Version]
- Moral, J.; Agustí-Brisach, C.; Raya, M.C.; Jurado-Bello, J.; López-Moral, A.; Roca, L.F.; Chattaoui, M.; Rhouma, A.; Nigro, F.; Sergeeva, V.; et al. Diversity of Colletotrichum species associated with olive anthracnose worldwide. J. Fungi 2021, 7, 741. [Google Scholar] [CrossRef] [PubMed]
- Buonaurio, R.; Almadi, L.; Famiani, F.; Moretti, C.; Agosteo, G.E.; Schena, L. Olive leaf spot caused by Venturia oleaginea: An updated review. Front. Plant Sci. 2023, 13, 1061136. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Favaron, F.; Sella, L. Characterization of Phytophthora and Botryosphaeriaceae species associated with emerging olive diseases in north-east Italy. In Abstracts of Presentations at the XXV Congress of the Italian Phytopathological Society (SIPaV). J. Plant Pathol. 2019, 101, 811–848. [Google Scholar]
- Sánchez-Hernández, E.; Muñoz-García, M.; Brasier, C.M.; Trapero-Casas, A. Identity and pathogenicity of two Phytophthora taxa associated with a new root disease of olive trees. Plant Dis. 2001, 85, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Raimondo, M.L.; Cibelli, F.; Phillips, A.J.L.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum associated with a decline of olives in southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Pathol. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- Phillips, A.J.; Rumbos, I.C.; Alves, A.; Correia, A. Morphology and phylogeny of Botryosphaeria dothidea causing fruit rot of olives. Mycopathologia 2005, 159, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Sergeeva, V.; Alves, A.; Phillips, A.J. Neofusicoccum luteum associated with leaf necrosis and fruit rot of olives in New South Wales, Australia. Phytopathol. Mediterr. 2009, 48, 294–298. [Google Scholar]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, T.J. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathology 2010, 100, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Rodríguez, L.; Mondino-Hintz, P.; Alaniz-Ferro, S. Diversity of Botryosphaeriaceae species causing stem canker and fruit rot in olive trees in Uruguay. J. Phytopathol. 2022, 170, 264–277. [Google Scholar] [CrossRef]
- Brunetti, A.; Matere, A.; Lumia, V.; Pasciuta, V.; Fusco, V.; Sansone, D.; Marangi, P.; Cristella, N.; Faggioli, F.; Scortichini, M.; et al. Neofusicoccum mediterraneum is involved in a twig and branch dieback of olive trees observed in Salento (Apulia, Italy). Pathogens 2022, 11, 53. [Google Scholar] [CrossRef]
- Ivić, D.; Petrović, E.; Godena, S. Fungi associated with canker diseases on olive in Istria (Croatia). J. Cent. Eur. Agric. 2023, 24, 470–475. [Google Scholar] [CrossRef]
- González, M.; Pérez-Sierra, A.; Serrano, M.S.; Sanchez, M.E. Two Phytophthora species causing decline of wild olive (Olea europaea subsp. europaea var. sylvestris). Plant Pathol. 2017, 66, 941–948. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Favaron, F.; Sella, L. First report of Phytophthora acerina, P. pini, and P. plurivora causing root rot and sudden death of olive trees in Italy. Plant Dis. 2020, 104, 996. [Google Scholar] [CrossRef]
- Scanu, B.; Jung, T.; Masigol, H.; Linaldeddu, B.T.; Jung, M.H.; Brandano, A.; Mostowfizadeh-Ghalamfarsa, R.; Janoušek, J.; Riolo, M.; Cacciola, S.O. Phytophthora heterospora sp. nov., a new pseudoconidia-producing sister species of P. palmivora. J. Fungi 2021, 7, 870. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.H.; Chiang, K.S.; Del Ponte, E.M. Plant disease severity estimated visually: A century of research, best practices, and opportunities for improving methods and practices to maximize accuracy. Trop. Plant Pathol. 2022, 47, 25–42. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Ruzzon, B.; Montecchio, L. Coniella granati and Phytophthora palmivora the main pathogens involved in pomegranate dieback and mortality in north-eastern Italy. Ital. J. Mycol. 2020, 49, 92–100. [Google Scholar]
- Linaldeddu, B.T.; Mulas, A.A.; Bregant, C.; Piras, G.; Montecchio, L. First Report of Phytophthora pistaciae causing root and collar rot on nursery plants of Pistacia lentiscus in Italy. Plant Dis. 2020, 104, 1564. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lamari, L. Assess: Image Analysis Software for Plant Disease Quantification; APS Press: St. Paul, MN, USA, 2002. [Google Scholar]
- Taylor, R.K.; Hale, C.N.; Hartill, W.F.T. A stem canker disease of olive (Olea europaea) in New Zealand. N. Z. J. Crop Hortic. Sci. 2001, 29, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Triki, M.A.; HadjTaieb, S.K.; Cheffi, M.; Gharbi, Y.; Rhouma, A. First report of dieback of olive trees caused by Neofusicoccum australe in Tunisia. J. Plant Pathol. 2015, 97, 212. [Google Scholar]
- Manca, D.; Bregant, C.; Maddau, L.; Pinna, C.; Montecchio, L.; Linaldeddu, B.T. First report of canker and dieback caused by Neofusicoccum parvum and Diplodia olivarum on oleaster in Italy. Ital. J. Mycol. 2020, 49, 85–91. [Google Scholar]
- Batista, E.; Lopes, A.; Alves, A. What do we know about Botryosphaeriaceae? An overview of a worldwide cured dataset. Forests 2021, 12, 313. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Maddau, L.; Franceschini, A.; Alves, A.; Phillips, A.J.L. Botryosphaeriaceae species associated with lentisk dieback in Italy and description of Diplodia insularis sp. nov. Mycosphere 2016, 7, 962–977. [Google Scholar] [CrossRef]
- Smahi, H.; Belhoucine-Guezouli, L.; Berraf-Tebbal, A.; Chouih, S.; Arkam, M.; Franceschini, A.; Linaldeddu, B.T.; Phillips, A.J.L. Molecular characterization and pathogenicity of Diplodia corticola and other Botryosphaeriaceae species associated with canker and dieback of Quercus suber in Algeria. Mycosphere 2017, 8, 1261–1272. [Google Scholar] [CrossRef]
- Urbez-Torres, J. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, 5–45. [Google Scholar]
- Fiorenza, A.; Gusella, G.; Vecchio, L.; Aiello, D.; Polizzi, G. Diversity of Botryosphaeriaceae species associated with canker and dieback of avocado (Persea americana) in Italy. Phytopathol. Mediterr. 2023, 62, 47–63. [Google Scholar]
- Moral, J.; Morgan, D.; Trapero, A.; Michailides, T.J. Ecology and epidemiology of diseases of nut crops and olives caused by Botryosphaeriaceae fungi in California and Spain. Plant Dis. 2019, 103, 1809–1827. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The main species associated with cankers and dieback of Fraxinus excelsior in north-eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Calice, U.; Linaldeddu, B.T.; Maddau, L.; Superchi, S.; Evidente, A. Diplofuranoxin, a disubstituted dihydrofuranone, was produced together with sphaeropsidin A and epi-sphaeropsidone by Diplodia subglobosa, an emerging ash (Fraxinus excelsior L.) pathogen in Europe. Phytochemistry 2022, 202, 113302. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Maddau, L.; Masi, M.; Linaldeddu, B.T.; Evidente, A. Secondary metabolites produced by Sardiniella urbana, a new emerging pathogen on European hackberry. Nat. Prod. Res 2019, 33, 1862–1869. [Google Scholar] [CrossRef]
- Saccardo, P.A. Conspectus generum fungorum Italiae inferorium. Michelia 1880, 2, 1–38. [Google Scholar]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfi, A.; Maddau, L.; Cimmino, A.; Linaldeddu, B.T.; Franceschini, A.; Serra, S.; Basso, S.; Melck, D.; Evidente, A. Cyclobotryoxide, a phytotoxic metabolite produced by the plurivorous pathogen Neofusicoccum australe. J. Nat. Prod. 2012, 75, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Tang, J.J.; Zhang, A.L.; Gao, J.M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef]
- Cimmino, A.; Maddau, L.; Masi, M.; Linaldeddu, B.T.; Pescitelli, G.; Evidente, A. Fraxitoxin, a new isochromanone isolated from Diplodia fraxini. Chem. Biodivers. 2017, 14, e1700325. [Google Scholar] [CrossRef]
- Di Lecce, R.; Marco, M.A.S.I.; Linaldeddu, B.T.; Pescitelli, G.; Maddau, L.; Evidente, A. Bioactive secondary metabolites produced by the emerging pathogen Diplodia olivarum. Phytopathol. Mediterr. 2021, 60, 129–138. [Google Scholar] [CrossRef]
- Rumbos, I.C. Dieback symptoms on olive trees caused by the fungus Eutypa lata 1. EPPO Bull. 1993, 23, 441–445. [Google Scholar] [CrossRef]
- Carlucci, A.; Lops, F.; Cibelli, F.; Raimondo, M.L. Phaeoacremonium species associated with olive wilt and decline in southern Italy. Eur. J. Plant Pathol. 2015, 141, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Trouillas, F.P.; Gubler, W.D. Identification and characterization of Eutypa leptoplaca, a new pathogen of grapevine in Northern California. Mycol. Res. 2004, 108, 1195–1204. [Google Scholar] [CrossRef]
- Bregant, C.; Rossetto, G.; Meli, L.; Sasso, N.; Montecchio, L.; Brglez, A.; Piškur, B.; Ogris, N.; Maddau, L.; Linaldeddu, B.T. Diversity of Phytophthora species involved in new diseases of mountain vegetations in Europe with the description of Phytophthora pseudogregata sp. nov. Forests 2023, 14, 1515. [Google Scholar] [CrossRef]
- Bregant, C.; Mulas, A.A.; Rossetto, G.; Deidda, A.; Maddau, L.; Piras, G.; Linaldeddu, B.T. Phytophthora mediterranea sp. nov., a new species closely related to Phytophthora cinnamomi from nursery plants of Myrtus communis in Italy. Forests 2021, 12, 682. [Google Scholar] [CrossRef]
- Bregant, C.; Batista, E.; Hilário, S.; Linaldeddu, B.T.; Alves, A. Phytophthora species involved in Alnus glutinosa decline in Portugal. Pathogens 2023, 12, 276. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Nowakowska, J.A.; Oszako, T. Phytophthora species isolated from ash stands in Białowieża Forest nature reserve. For. Pathol. 2016, 46, 660–662. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Sikora, K.; Galko, J.; Kunca, A. Occurrence of Phytophthora species in riparian stands of black alder (Alnus glutinosa) in Slovakia. For. Pathol. 2023, 53, e12800. [Google Scholar] [CrossRef]
- Seddaiu, S.; Linaldeddu, B.T. First Report of Phytophthora acerina, P. plurivora, and P. pseudocryptogea associated with declining common alder trees in Italy. Plant Dis. 2020, 104, 1874. [Google Scholar] [CrossRef]
- Hong, C.; Gallegly, M.E.; Richardson, P.A.; Kong, P. Phytophthora pini Leonian resurrected to distinct species status. Mycologia 2011, 103, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, C.; Biscontri, M.; Tabet, D.; Vettraino, A.M. The never-ending presence of Phytophthora species in Italian nurseries. Pathogens 2022, 12, 15. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of Phytophthora species from declining Mediterranean maquis vegetation, including two new species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef] [Green Version]
- Sims, L.L.; Chee, C.; Bourret, T.; Hunter, S.; Garbelotto, M. Genetic and phenotypic variation of Phytophthora crassamura isolates from California nurseries and restoration sites. Fungal Biol. 2019, 123, 159–169. [Google Scholar] [CrossRef]
- Bregant, C.; Rossetto, G.; Deidda, A.; Maddau, L.; Franceschini, A.; Ionta, G.; Raiola, A.; Montecchio, L.; Linaldeddu, B.T. Phylogeny and pathogenicity of Phytophthora species associated with artichoke crown and root rot and description of Phytophthora marrasii sp. nov. Agriculture 2021, 11, 873. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Schena, L.; Agosteo, G.E.; Magnano di San Lio, G.; Cacciola, S.O. Phytophthora oleae sp. nov. causing fruit rot of olive in southern Italy. Plant Pathol. 2018, 67, 1362–1373. [Google Scholar] [CrossRef]
- González, M.; Pérez-Sierra, A.; Sánchez, M.E. Phytophthora oleae, a new root pathogen of wild olives. Plant Pathol. 2019, 68, 901–907. [Google Scholar] [CrossRef]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Cooke, D.E.; Duncan, J.M.; Hansen, E.M. Multiple new phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides–P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef]
- Hansen, E.; Delatour, C. Phytophthora species in oak forests of north-east France. Ann. For. Sci. 1999, 56, 539–547. [Google Scholar] [CrossRef]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First report of root rot caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef]
- Rodriguez-Padron, C.; Siverio, F.; Perez-Sierra, A.; Rodriguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythium vexans recovered from avocado orchards in the Canary Islands, including Phytophthora niederhauserii as a new pathogen of avocado. Phytopathol. Mediterr. 2018, 57, 89–106. [Google Scholar]
- Bourret, T.B.; Fajardo, S.N.; Frankel, S.J.; Rizzo, D.M. Cataloging Phytophthora Species of Agriculture, Forests, Horticulture, and Restoration Outplantings in California, USA: A Sequence-Based Meta-Analysis. Plant Dis. 2023, 107, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Sanchez-Hernandez, E.; Kirk, S.A. Phytophthora inundata sp. nov., a part heterothallic pathogen of trees and shrubs in wet or flooded soils. Mycol. Res. 2003, 107, 477–484. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
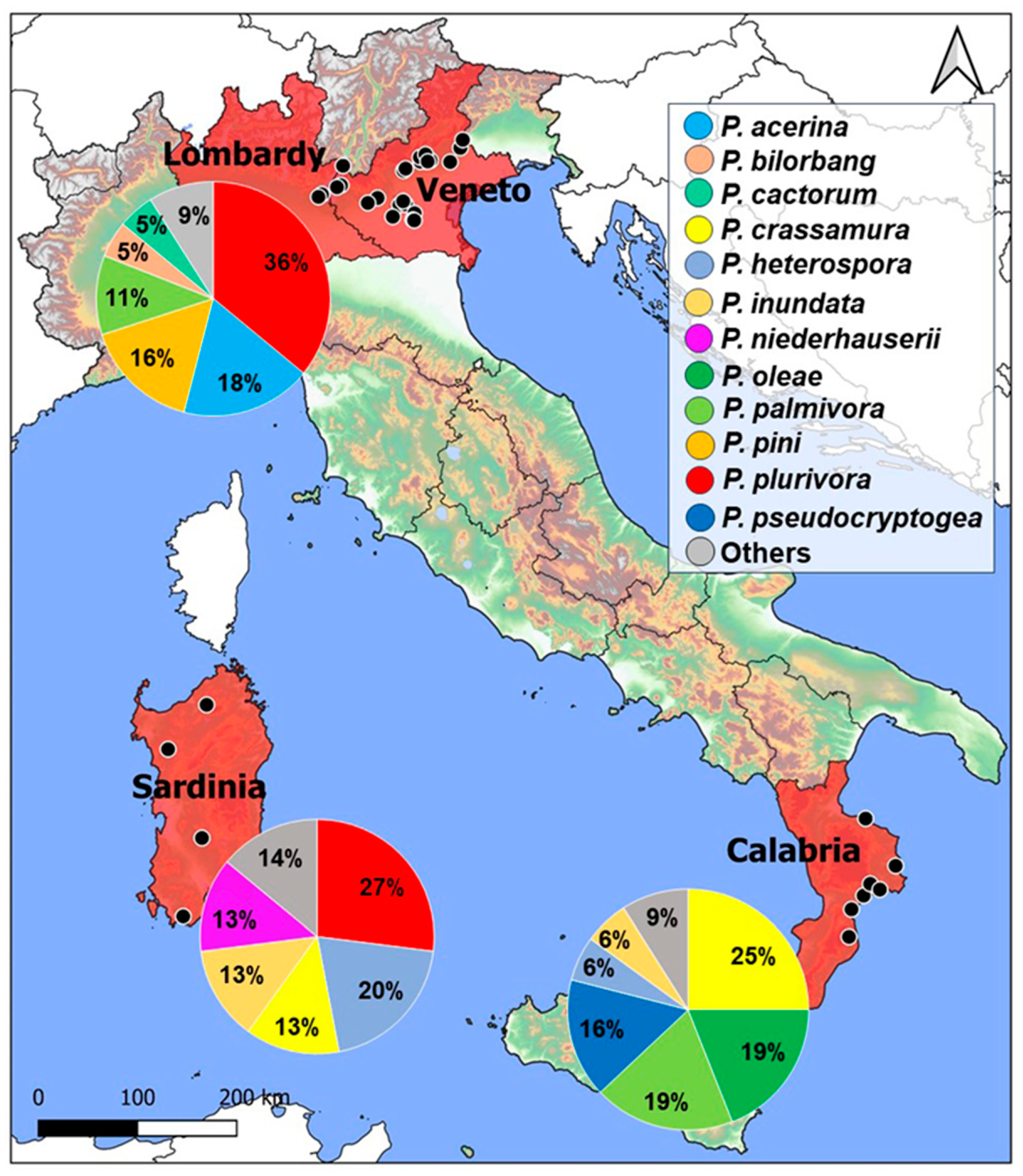{kind=link}
| Italian Region | Sites | Cultivar | Type of Samples | |||
|---|---|---|---|---|---|---|
| Unripe Drupes | Ripe Drupes | Stem/Branches | Collar/Roots | |||
| Veneto | 1 | Casaliva | 6 | - | 10 | 12 |
| 2 | Grignano | - | - | 15 | 3 | |
| 3 | Grignano | 3 | - | 15 | - | |
| 4 | Grignano | 2 | - | 13 | - | |
| 5 | Grignano | - | - | 19 | - | |
| 6 | Grignano | - | - | 51 | - | |
| 7 | Grignano | - | - | 4 | 2 | |
| 8 | Grignano | - | 4 | 13 | 2 | |
| 9 | Leccino, frantoio, Trepp, favarol | - | - | 21 | - | |
| 10 | Leccino, grignano, trepp | 16 | 35 | 17 | 20 | |
| 11 | Grignano | - | - | 14 | 2 | |
| 12 | Grignano | - | - | 8 | - | |
| 13 | Grignano | 12 | - | 7 | 5 | |
| 14 | Grignano | - | - | 6 | - | |
| 15 | Arbequina, arbosana | 10 | - | 3 | 16 | |
| 16 | Rasara, leccino, matosso, itrana | - | 15 | 7 | 2 | |
| 17 | Rasara | 50 | 50 | 75 | 7 | |
| 18 | Frantoio, leccino | - | 5 | 4 | 2 | |
| 19 | Frantoio, leccino, Grignano | - | 8 | 6 | 2 | |
| 20 | Frantoio, leccino | - | 10 | 5 | - | |
| 21 | Leccino, grignano, Frantoio | - | 3 | 3 | 2 | |
| 22 | Frantoio, leccino, pendolino | - | 10 | 5 | - | |
| 23 | Leccino, frantoio, Grignano | - | 10 | 7 | 3 | |
| 24 | Frantoio, leccino | - | 5 | 5 | 2 | |
| Lombardy | 25 | Frantoio | - | 6 | 5 | 3 |
| 26 | Frantoio | - | 5 | 3 | 2 | |
| Sardinia | 27 | Frantoio | 100 | 100 | 10 | 10 |
| 28 | Bosana | - | - | 10 | - | |
| 29 | Semidana | - | - | 5 | 7 | |
| 30 | Pizz’e carroga | 100 | - | - | - | |
| Calabria | 31 | Carolea | - | - | - | 5 |
| 32 | Biancolilla | - | - | - | 5 | |
| 33 | Nocellara del Belice | - | - | - | 5 | |
| 34 | Frantoio | - | - | - | 3 | |
| 35 | Carolea | - | - | - | 3 | |
| 36 | Carolea | - | - | - | 3 | |
| 37 | Carolea | - | - | - | 3 | |
| 38 | Carolea | - | - | - | 2 | |
| Fungal Species | Accession Numbers | Type of Samples | Sites | |||
|---|---|---|---|---|---|---|
| UD | RD | SB | R | |||
| Apiospora arundinis | OR284845 | 2 | - | 2 | - | 1, 9 |
| Botryosphaeria dothidea | OR284846 | 208 | 73 | 16 | - | 6, 10, 15–23, 25–27, 30 |
| Diplodia africana | OR284847 | - | 1 | - | - | 27 |
| Diplodia fraxini | OR284848 | 2 | - | 15 | - | 10, 17 |
| Diplodia mutila | OR284849 | 32 | 38 | 12 | - | 6, 9, 10, 12, 16–21, 27 |
| Diplodia olivarum | OR284850 | 9 | 24 | 15 | - | 1, 6, 7, 9, 11, 12, 21, 24, 27, 30 |
| Diplodia sapinea | OR284851 | - | 1 | - | - | 27 |
| Diplodia seriata | OR284852 | 31 | 8 | 47 | - | 1–4, 6–8, 10, 11,13, 15–17, 27, 30 |
| Diplodia subglobosa | OR284853 | - | 4 | 14 | - | 1–3, 9, 15, 17, 20 |
| Dothiorella iberica | OR284854 | 1 | - | 2 | - | 10, 16 |
| Dothiorella omnivora | OR284855 | - | - | 1 | - | 16 |
| Dothiorella sarmentorum | OR284856 | - | 3 | 11 | - | 1–6, 17 |
| Dothiorella sempervirentis | OR284857 | - | - | 1 | - | 6 |
| Eutypa lata | OR284858 | - | - | 3 | - | 9, 16 |
| Eutypa leptoplaca | OR284859 | - | - | 2 | - | 1, 9 |
| Neofusicoccum australe | OR284860 | - | 29 | 1 | - | 27 |
| Neofusicoccum cryptoaustrale | OR284861 | - | 11 | 4 | - | 27, 29 |
| Neofusicoccum luteum | OR284862 | 2 | 5 | - | - | 27 |
| Neofusicoccum mediterraneum | OR284863 | 3 | 5 | 7 | - | 27, 28 |
| Neofusicoccum parvum | OR284864 | 7 | 18 | 90 | - | 1, 2, 4, 5, 6, 8–10, 15–28 |
| Phaeoacremonium iranianum | OR284865 | - | - | 2 | - | 14 |
| Phaeoacremonium scolyti | OR284866 | - | - | 2 | - | 9, 14 |
| Sardiniella urbana | OR284867 | - | 1 | - | - | 17 |
| Phytophthora acerina | OR284868 | - | - | - | 14 | 8, 10, 24, 27 |
| Phytophthora bilorbang | OR284869 | - | - | - | 6 | 10, 15, 38 |
| Phytophthora cactorum | OR284870 | - | - | - | 4 | 11, 17 |
| Phytophthora cinnamomi | OR284871 | - | - | - | 2 | 19 |
| Phytophthora citricola | OR284872 | - | - | - | 1 | 31 |
| Phytophthora crassamura | OR284873 | - | - | - | 10 | 29, 31–33, 36 |
| Phytophthora heterospora | OR284874 | - | - | - | 5 | 27, 29, 35 |
| Phytophthora inundata | OR284875 | - | - | - | 4 | 29, 31, 32 |
| Phytophthora nicotianae | OR284876 | - | - | - | 2 | 15 |
| Phytophthora niederhauserii | OR284877 | - | - | - | 4 | 10, 27 |
| Phytophthora oleae | OR284878 | - | - | - | 6 | 31, 32, 34 |
| Phytophthora palmivora | OR284879 | - | - | - | 15 | 7, 25–27, 33, 34, 37 |
| Phytophthora pini | OR284880 | - | - | - | 12 | 11, 15, 18 |
| Phytophthora plurivora | OR284881 | - | - | - | 31 | 1, 6, 8, 12, 27 |
| Phytophthora pseudocryptogea | OR284882 | - | - | - | 5 | 31, 33, 36, 37, 38 |
| Fungal Species | Drupe Lesion Size 1 | Positive Re-Isolation (%) |
|---|---|---|
| Botryosphaeria dothidea | 2.4 ± 0.8 b | 100 |
| Diplodia mutila | 2.3 ± 0.9 b | 100 |
| Diplodia olivarum | 2.7 ± 2.1 b | 100 |
| Neofusicoccum parvum | 6.3 ± 2.6 a | 100 |
| Control | 0.0 | - |
| Fungal Species | Length Necrotic Lesion 1 | Wilting Symptoms | Positive Re-Isolation (%) |
|---|---|---|---|
| Apiospora arundinis | 1.2 ± 0.4 ef | no | 20 |
| Botryosphaeria dothidea | 6.1 ± 1.2 de | no | 100 |
| Diplodia fraxini | 33.5 ± 6.5 b | yes | 100 |
| Diplodia olivarum | 14.6 ± 2.6 c | no | 100 |
| Diplodia seriata | 6.2 ± 1.3 d | no | 100 |
| Diplodia subglobosa | 13.6 ± 2.6 c | no | 100 |
| Eutypa lata | 6.1 ± 1.2 de | no | 80 |
| Eutypa leptoplaca | 5.8 ± 1.2 de | no | 80 |
| Neofusicoccum parvum | 58.2 ± 10.1 a | yes | 100 |
| Phaeoacremonium iranianum | 5.2 ± 1.2 de | no | 60 |
| Control | 0.2 ± 0.4 f | no | - |
| Phytophthora Species | Length Necrotic Lesion 1 | Positive Re-Isolation (%) |
|---|---|---|
| Phytophthora acerina | 27.7 ± 4.9 a | 100 |
| Phytophthora bilorbang | 13.8 ± 5.9 c | 100 |
| Phytophthora cactorum | 20.1 ± 5.8 b | 100 |
| Phytophthora cinnamomi | 12.6 ± 2.1 c | 100 |
| Phytophthora citricola | 7.3 ± 3.8 d | 100 |
| Phytophthora crassamura | 4.8 ± 2.3 de | 100 |
| Phytophthora heterospora | 11.3 ± 4.2 c | 100 |
| Phytophthora inundata | 2.4 ± 0.5 e | 80 |
| Phytophthora oleae | 7.2 ± 2.5 d | 100 |
| Phytophthora palmivora | 19.1 ± 4.1 b | 100 |
| Phytophthora pini | 29.5 ± 5.4 a | 100 |
| Phytophthora plurivora | 28.7 ± 3.5 a | 100 |
| Phytophthora pseudocryptogea | 13.1 ± 3.8 c | 100 |
| Control | 1.6 ± 1.2 e | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linaldeddu, B.T.; Rossetto, G.; Maddau, L.; Vatrano, T.; Bregant, C. Diversity and Pathogenicity of Botryosphaeriaceae and Phytophthora Species Associated with Emerging Olive Diseases in Italy. Agriculture 2023, 13, 1575. https://doi.org/10.3390/agriculture13081575
Linaldeddu BT, Rossetto G, Maddau L, Vatrano T, Bregant C. Diversity and Pathogenicity of Botryosphaeriaceae and Phytophthora Species Associated with Emerging Olive Diseases in Italy. Agriculture. 2023; 13(8):1575. https://doi.org/10.3390/agriculture13081575
Chicago/Turabian StyleLinaldeddu, Benedetto T., Giovanni Rossetto, Lucia Maddau, Thomas Vatrano, and Carlo Bregant. 2023. "Diversity and Pathogenicity of Botryosphaeriaceae and Phytophthora Species Associated with Emerging Olive Diseases in Italy" Agriculture 13, no. 8: 1575. https://doi.org/10.3390/agriculture13081575