
Additions to the Knowledge of the Genus Pezicula (Dermateaceae, Helotiales, Ascomycota) in China

State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
*
Authors to whom correspondence should be addressed.
Biology 2022, 11(10), 1386; https://doi.org/10.3390/biology11101386
Submission received: 3 August 2022
/
Revised: 19 September 2022
/
Accepted: 20 September 2022
/
Published: 23 September 2022
(This article belongs to the Special Issue Studies on Species Diversity and Phylogeny of Ascomycota (Fungi))
Abstract
:Simple Summary
Species of the genus Pezicula are endophytes, phytopathogens, or saprobes, and some of them have biocontrol potential, promoting plant growth and resistance to environmental stress. The species diversity of Pezicula in China was reviewed. Two new species (P. ellipsoids and P. fusispora) and two new Chinese records (P. acericola and P. carpinea) were discovered using morphological and molecular approaches. Descriptions and illustrations of macroscopic and microscopic features were provided for the new and newly found taxa. Pezicula aurantiaca was excluded from the fungal catalogue of China. Together with the previously reported taxa (P. cinnamomea, P. ericae, P. heterochroma, P. magnispora, P. melanigena, P. neosporulosa, P. ocellata, P. rhizophila, P. cf. rubi, and P. subcarnea), 14 Pezicula species are currently known from China. The results provide updated information and improve our understanding of the genus.
Abstract
We describe two new species of Pezicula (Dermateaceae, Ascomycota), P. ellipsoides and P. fusispora, which are discovered in China. Pezicula ellipsoides sp. nov. is distinct in producing 2–3 stipitate apothecia on a basal stroma with a light yellow hymenium, broadly ellipsoid 0–1(–2)-septate ascospores, and divergent DNA sequence data. Pezicula fusispora sp. nov. is characterized by sessile apothecia, 0.3–0.8 mm in diam, a yellowish hymenium, J+ asci 135–170 × 15–21 μm, multiseptate ascospores, 33–48 × 7–10.3 μm, and growing on rotten bamboo. In addition, Pezicula acericola and P. carpinea are reported for the first time from China, and 11 Pezicula species previously reported from China are reviewed and briefly noted, of which one was a misidentified species. Phylogenetic analyses inferred from ITS nrDNA sequences confirm the placement of P. ellipsoides and P. cinnamomea in the genus Pezicula.
1. Introduction
Pezicula Tul. & C. Tul. belongs to the family Dermateaceae Fr. (Helotiales, Leotiomycetes), which was established in 1869 and typified by P. carpinea (Pers.) Tul. ex Fuckel [1]. The sexual stages of the genus are often found as bright-colored, fleshy, pruinose or pulverulent, sessile to short-stipitate apothecia on woody substrates [2]. The asexual stages of Pezicula were originally placed in Cryptosporiopsis Bubák & Kabát, which is treated as a later synonym according to the ‘one fungus–one name’ rule of the International Code of Nomenclature for algae, fungi, and plants [3]. Pezicula species are endophytes, phytopathogens, or saprobes, and some of them have the abilities of antagonizing plant pathogens, producing compounds with antimicrobial activities, promoting plant growth, and increasing plant resistance to environmental stress [2,4,5,6,7]. Several regional investigations of the genus have been carried out [8,9,10,11]. A monographic treatment of Pezicula was published by Verkley in 1999, in which 26 species were accepted [2]. Eleven taxa were further introduced, ten species were transferred to Pezicula from other genera, and one name was treated as synonym thereafter [3,5,12,13,14,15,16,17,18]. One hundred and sixty-two epithets are listed under Pezicula in the Index Fungorum online database [1]. In recent years, morphological characters combined with PCR-based restriction fragment length polymorphism (RFLP) patterns of ribosomal DNA as well as DNA sequence analyses were used for reliable species identification. Forty-six species are currently recognized in the genus.
Phylogenetically, Pezicula is closely related to Rhizodermea Verkley & J.D. Zijlstra [5] and Dermea Fr. [19], and also associated with Coleophoma Höhn., Davidhawksworthia Crous, Neofabraea H.S. Jacks., Parafabraea Chen Chen, Verkley & Crous, Phlyctema Desm., and Pseudofabraea Chen Chen, Verkley & Crous based on sequence analyses of the internal transcribed spacer region (ITS), partial large subunit nrDNA (LSU), partial β-tubulin region (tub2), and partial RNA polymerase II second largest subunit (rpb2) regions [5,19,20,21,22]. The above-mentioned genera can be separated from each other by the morphology of sexual and/or asexual stages as well as DNA sequences.
In China, the first report of Pezicula species was attributed to Teng (1963) based on specimens of P. ocellata (Pers.) Seaver collected from Ningxia and Qinghai provinces [23]. Additional species have been recorded from different regions [7,9,12,13,24,25,26,27,28,29,30]. During our studies on the species diversity of Dermateaceae in China, herbarium specimens were re-examined, and recent collections of the genus were identified on the basis of morphological features and molecular data. The occurrence of 11 Pezicula species in China are summarized and reviewed based on the literature. In addition, two new species are described, and two new records are added to the fungal catalogue of China.
2. Materials and Methods
2.1. Morphological Observations
The study was based on newly collected materials and specimens preserved in the Herbarium Mycologicum Academiae Sinicae (HMAS). Macroscopic characters were recorded according to the field notes or observed under a SZX7 stereomicroscope (Olympus Corporation, Tokyo, Japan). Dried apothecia were rehydrated with distilled water and sectioned at a thickness of 15−20 μm with a YD-1508A freezing microtome (Yidi Medical Equipment Co., Ltd., Jinhua, Jiangsu, China). Iodine reactions of ascus apical apparatus were tested in Melzer’s reagent and Lugol’s solution with or without 3% KOH pretreatment [31]. Microscopic examinations and measurements were taken from dried apothecia under a BH-2 microscope (Olympus Corporation, Tokyo, Japan). Microscopic photographs were taken using an Axiocam 305 color microscope camera (Zeiss Corporation, Göttingen, Germany) attached to an Axioskop 2 Plus microscope (Zeiss Corporation, Göttingen, Germany).
2.2. DNA Extraction, PCR, and Sequencing
Genomic DNA was extracted from dried apothecia using a Plant Genomic DNA Kit (TIANGEN Biotech. Co., Beijing, China). PCR reaction mixtures had a final volume of 30 μL, containing 15 μL 2 × Taq MasterMix (CWBiotech. Co., Beijing, China), 1.5 μL of each primer (10 mM), 2 μL DNA, and 10 μL deionized water. The nuclear ribosomal internal transcribed spacer (ITS) region was amplified using the primers ITS1 and ITS4 [32]. PCR reactions were carried out in an Applied Biosystems 2720 thermocycler (Foster City, CA, USA) under the following conditions: 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 53 °C for 30 s and 30 s at 72 °C, and a final extension of 72 °C for 10 min. PCR products were purified and sequenced at Tianyi Huiyuan Bioscience and Technology Co. Ltd. (Beijing, China).
2.3. Sequence Assembly, Alignment, and Phylogenetic Analyses
Newly generated sequences were assembled using BioEdit 7.0.9.0 [33] and submitted to GenBank. ITS sequences of representative Pezicula species and two outgroup taxa were retrieved from GenBank (Table 1). The sequence dataset was aligned with MUSCLE [34] and manually adjusted where necessary in BioEdit 7.0.9.0 [33].
The ITS matrix was analyzed using Maximum Likelihood (ML) and Bayesian inference (BI) methods. ML analysis was conducted in RAxML 8.0 [35] using a GTRGAMMA model and tested with 1000 bootstrap replicates. BI analysis was performed using MrBayes 3.1.2 [36]. The best-fit nucleotide substitution model was estimated by MrModeltest 2.3 [37] with the Akaike information criterion. Two parallel runs of four simultaneous chains of Markov Chain Monte Carlo were performed for 2,000,000 generations with trees sampled every 100 generations. The first quarter of the sampled trees were discarded as the burn-in phase, and the remaining trees were used for calculating posterior probabilities (PP) in the majority rule consensus tree. The phylogenetic trees were visualized in FigTree v1.4.4 [38].
3. Results
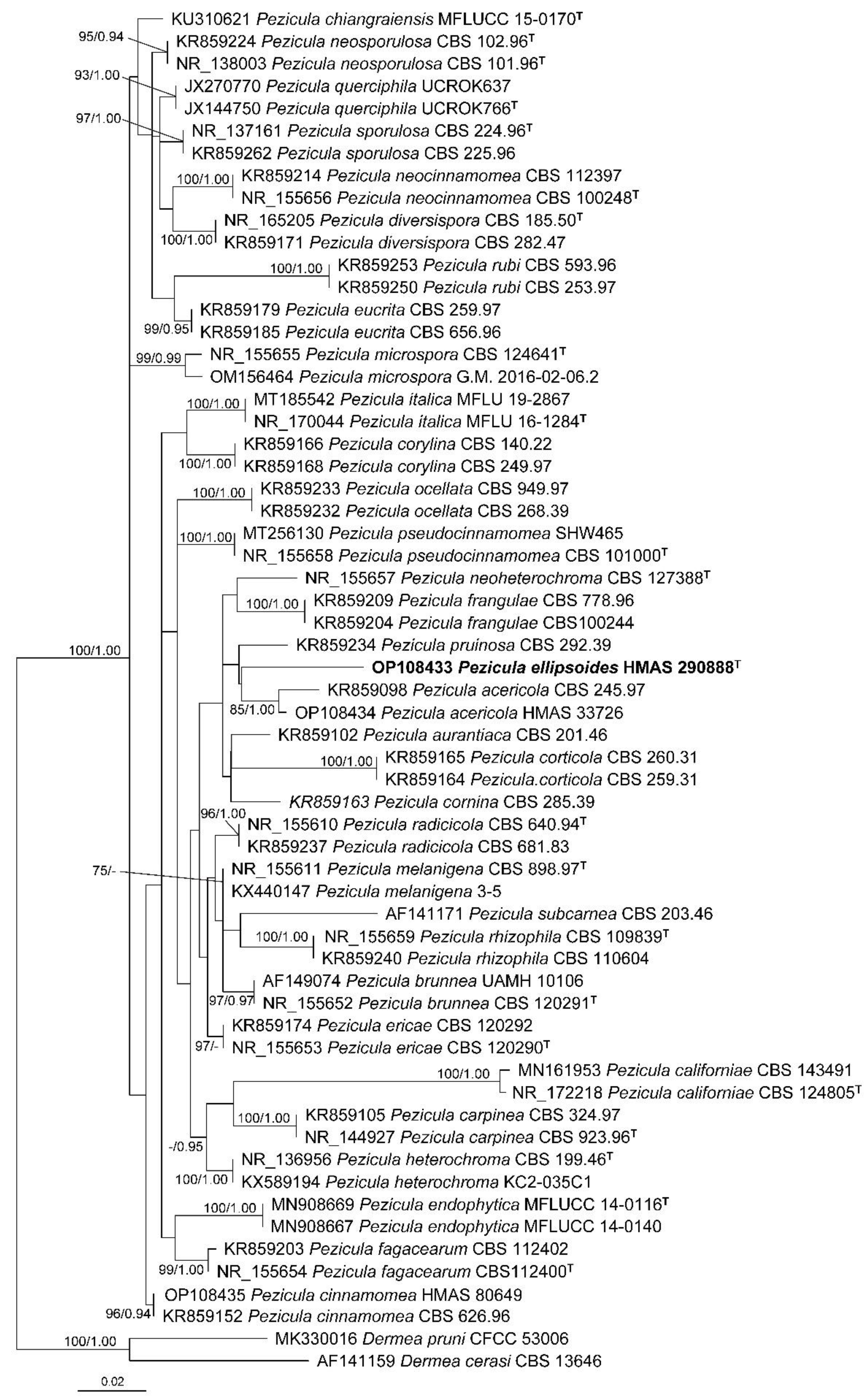
3.1. Phylogeny
A total of 61 ITS sequences representing 33 Pezicula species and two outgroup taxa were included in the phylogenetic analyses (Table 1). The ITS alignment consisted of 542 characters including gaps (TreeBASE accession number: 29639). GTR+I+G was selected as the best-fit model. The resulting maximum likelihood phylogenetic tree is shown in Figure 1 with bootstrap support values of ML (≥50%) and Bayesian posterior probabilities (≥0.90) shown at the nodes from left to right. All the samples of Pezicula formed a well-supported monophyletic group, and all the investigated species were recognized.
3.2. Taxonomy
3.2.1. New Species
Fungal Names: FN571258.
Etymology: The specific epithet refers to the ascospore shape of the species.
Holotype: CHINA, Hubei, Shennongjia, alt. 2500 m, on rotten bark, 14 September 2014, H.D. Zheng et al. 9542, HMAS 290888.
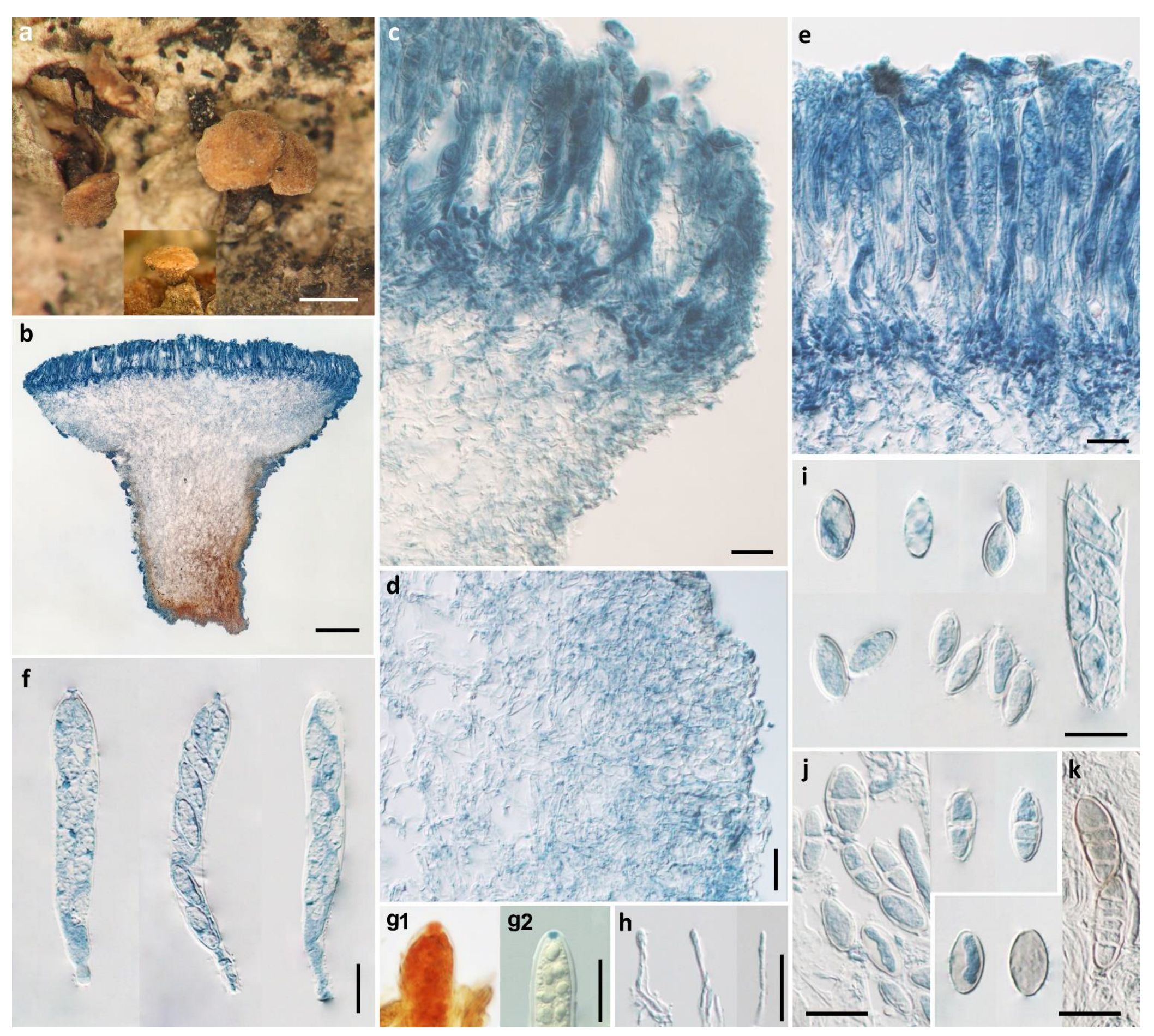
Apothecia erumpent, solitary or 2–3 on a basal stroma, discoid with surface convex, margin entire, short stipitate, 1–2 mm in diam; hymenium light yellow when fresh, pale brownish-yellow when dry; receptacle surface concolorous. Ectal excipulum of textura angularis, 50–80 μm thick, cells hyaline to light brown, isodiametric or nearly so, 4–16 μm in diam. Medullary excipulum of textura intricata, 50–300 μm thick, hyphae hyaline, 3.5–6 μm wide, becoming light brown in stipe and some swollen up to 20 μm in diam. Subhymenium hyaline, composed of interwoven hyphae, about 40 μm thick. Hymenium 140–150 μm thick. Asci arising from simple septa, 8-spored, cylindrical-clavate, with apex rounded to conical-rounded, narrowed gradually or abruptly into a short stalk, apical rings J+, becoming blue only after KOH pretreatment in Melzer’s reagent and not changing color without KOH pretreatment, and becoming red without KOH pretreatment and blue after KOH pretreatment in Lugol’s solution, 110–128 × 15–18 μm. Ascospores ellipsoid, with ends rounded or somewhat pointed, 0–1(–2)-septate, tardily becoming 3–5-septate, thin-walled, hyaline to light brown, multiguttulate, irregularly uniseriate in asci, 15.5–22.5 × 7.5–12 μm. Paraphyses filiform, septate, simple, hyaline, 2–2.5 μm wide, slightly enlarged at apex, equaling to or slightly exceeding the asci.
Notes: In the BLASTN search of the ITS region of P. ellipsoides in NCBI, P. cinnamomea (CBS 242.60, MH857970) shares the highest sequence similarity of 96.55% (503/521), followed by P. brunnea (CBS 120291, NR_155652) and P. acericola (CBS 248.97, KR859101), both with 96.35% (502/521) similarity. The phylogenetic analyses confirmed the placement of P. ellipsoides in Pezicula and its distinction from any other species of the genus.
Pezicula puberula (E.J. Durand) Verkley is similar to P. ellipsoides in the morphology of apothecia, asci, and ascospores, but differs in the presence of its whitish pruinate hymenium surface, up to 15 apothecia attached to a single basal stroma, cells of ectal excipulum 10–25 × 5–8 μm, and receptacle surface covered by amorphous crystals [2]. The ascospores of P. ellipsoides resemble those of P. sepium (Desm.) Dennis; however, the latter possesses smaller (less than 1.4 mm in diam), sessile to subsessile apothecia, thick-walled medullary excipular cells, 7–12 × 5–8 μm, and larger asci, 130–165 × 15–23 μm [2].
Pezicula fusispora W.Y. Zhuang & H.D. Zheng, sp. nov. Figure 3.
Fungal Names: FN571257.
Etymology: The specific epithet refers to the fusoid ascospores of the fungus.
Holotype: CHINA, Hunan, Yizhang, Mangshan, alt. 1200 m, on rotten bamboo, 11 April 2002, W.Y. Zhuang, B. Liu & X.Z. Liu 4162, HMAS 290892.
Apothecia slightly erumpent, solitary or 2–5 on a basal stroma, discoid, sessile; disc circular, elliptical to irregular, slightly convex, concave when dry, hymenium is bright yellow to yellowish-brown, dull yellow with olivaceous tint when dry, 0.3–0.8 mm in diam; receptacle near and at the margin concolorous with the hymenium, brownish at flanks and towards the base; margin entire or slightly torn. Ectal excipulum of textura angularis, 20–30 μm thick, cells brown, isodiametric or nearly so, 5–12 × 4–10 μm. Medullary excipulum of textura intricata, poorly developed, 30–40 μm thick, hyphae hyaline, 2–3 μm in diam. Subhymenium indistinguishable. Hymenium 155–175 μm thick. Asci arising from simple septa, 8-spored, cylindrical-clavate, with apex conical-rounded, narrowed gradually into a short, thin stalk, apical rings J+, blue in Melzer’s reagent and Lugol’s solution without KOH pre-treatment, 135–170 × 15–21 μm. Ascospores fusoid, with ends pointed, inequilateral, 3–7-septate, thin-walled, hyaline to light brown, multiguttulate, irregularly biseriate in asci, 33–48 × 7–10.3 μm. Paraphyses filiform, septate, simple, hyaline, 2.5–3 μm wide, slightly enlarged at apex, equaling to or slightly exceeding the asci.
Other specimen examined: CHINA, Hunan, Nanyue, Zhurongfeng, alt. 700 m, on rotten bamboo, 11 April 2002, W.Y. Zhuang 4136, HMAS 290893.
Notes: Pezicula fusispora can be easily distinguished by the combination of sessile apothecia with bright yellow to yellowish-brown hymenium, J+ asci in iodine solutions, and multiseptate ascospores which are fusoid and 33–48 × 7–10.3 μm in size. Pezicula aurantiaca and P. ocellata also have sessile apothecia but are different in hymenium color, J− ascus apical apparatus, and ascospores that are ellipsoid, smaller, and with fewer septa [2]. Unfortunately, no DNA sequence was retrieved from the voucher specimens, and the phylogenetic position of the fungus within the genus awaits future investigation.
3.2.2. New Records for China
Pezicula acericola (Peck) Peck ex Sacc. & Berl., Atti Inst. Veneto Sci. Lett., ed Arti (Sér. 6) 3: 725, 1885. Figure 4.
≡Nodularia acericola Peck, Ann. Rep. N.Y. St. Mus. 24: 98, 1872.
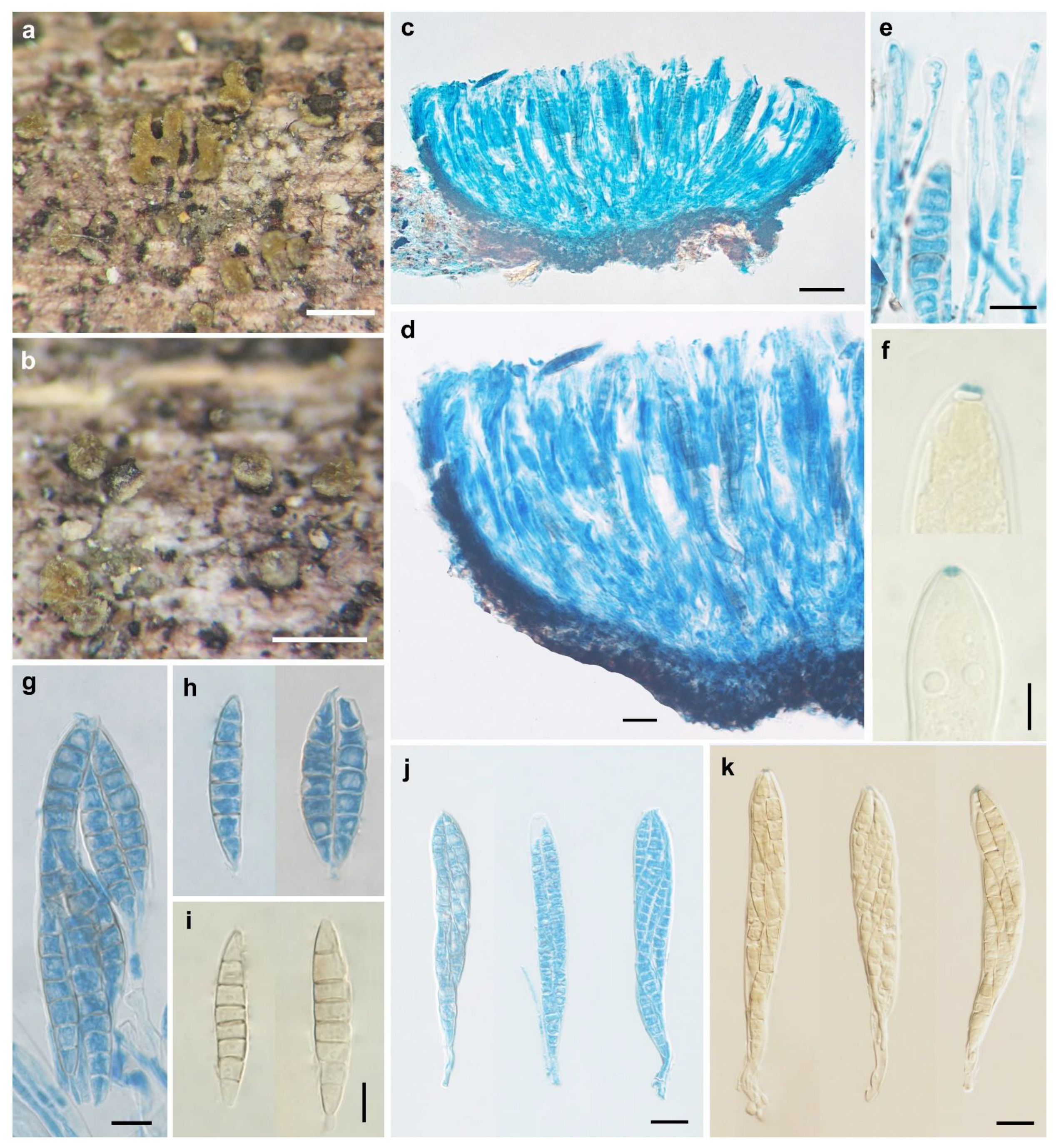
Apothecia erumpent, a few to a large number in circular to linear clusters on a basal stroma, discoid with surface plane to slightly convex, margin entire or irregularly torn, short stipitate; hymenium yellow when fresh, light cinnamon when dry, 0.3–3 mm in diam; receptacle concolorous to paler than hymenium. Ectal excipulum of textura angularis, 25–180 μm thick, cells subhyaline to light brown, isodiametric or nearly so, 4–16.5 μm in diam. Medullary excipulum of textura intricata, 20–190 μm thick, hyphae hyaline to light brown, 4–8.5 μm wide. Subhymenium indistinguishable. Hymenium 125–150 μm thick. Asci arising from croziers, clavate, apex conical-rounded, narrowed gradually or abruptly into a short stalk, 8-spored, apical rings J+, becoming blue only after KOH pretreatment in Melzer’s reagent, and becoming red without KOH pretreatment and blue after KOH pretreatment in Lugol’s solution, 98.5–126 × 16.5–22 μm. Ascospores ellipsoid with ends narrower, slightly inequilateral, 0(–3)-septate, thin-walled, hyaline, multiguttulate, irregularly biseriate in asci, 24.5–33 × 7.5–11 μm. Paraphyses filiform, septate, hyaline, slightly swollen to 3–5 μm wide at apex and 1.5–3 μm wide below, equaling to or slightly exceeding the asci.
Specimens examined: China, Jilin, Antu County, Mt. Changbai, alt. 1600 m, on rotten twig, 11 August 1960, Y.C. Yang, J.R. Li & F.S. Yuan 673 (HMAS 33726); Jilin, Antu County, Mt. Changbai Forestry Station, alt. 1750 m, on rotten bark, 25 July 1960, Y.C. Yang, J.R. Li & F.S. Yuan 420 (HMAS 33749).
Known distribution: Asia (China, Japan), Europe (Austria, Czech Republic, France, Italy, the Netherlands, Sweden, Switzerland, United Kingdom), and North America (Canada, USA#) (# indicating country of type locality).
Notes: Pezicula acericola is widely distributed in Europe and North America [2]. The macroscopic and microscopic features of the Chinese materials coincide with those from other countries. The ITS sequence of the Chinese collection (HMAS 33726) has a 98.89% similarity with those of the Canadian materials (KR859094-KR859101). This is a new record for China. The two voucher specimens (HMAS 33726 and HMAS 33749) were originally misidentified as P. aurantiaca, which is clearly different in possessing sessile apothecia that are mostly solitary or 2–3(–4) on a basal stroma, asci J− in Melzer’s reagent, and ascospores smaller in size (14.5–23 × 6–10 μm) [2].
Pezicula carpinea (Pers.) Tul. ex Fuckel, Jb. Nassau. Ver. Naturk. 23–24: 279, 1870. Figure 5.
≡Peziza carpinea Pers., Syn. Meth. Fung. (Göttingen) 2: 673, 1801.
Apothecia erumpent, mostly 2–15 on a basal stroma, discoid, margin slightly raised, drying concave, substipitate to short-stipitate, 0.5–2.0 mm in diam; hymenium pruinose, ochraceous when dry; receptacle concolorous with hymenium. Ectal excipulum of textura angularis to textura globulosa, 25–40 μm thick, cells hyaline to brown, isodiametric or nearly so, axis of cells arranged at a high to right angle to the outer surface, 5–20 μm in diam. Medullary excipulum of textura intricata, 110–220 μm thick, hyphae hyaline to light brown, 4–5.5 μm wide. Subhymenium composed of interwoven hyphae, 40–55 μm thick, cells hyaline. Hymenium 150–165 μm thick. Asci arising from simple septa, cylindrical-clavate, apex rounded to truncate-rounded, narrowed gradually into a stalk of varying length, 8-spored, apical rings J+, becoming blue in Melzer’s reagent and Lugol’s solution without KOH pre-treatment, 129–165 × 21–25 μm. Ascospores ellipsoid to broadly ellipsoid, slightly inequilateral, with ends rounded, nonseptate, thin-walled, hyaline, multiguttulate, uniseriate to biseriate in asci, 19.2–24.6 × 10.5–11.5 μm. Paraphyses filiform, septate, simple or branched at upper part, hyaline to very pale brown, swollen to 3.5–5 μm at apex and 2–3 μm wide below, exceeding the asci by 15–25 μm.
Specimen examined: China, Tibet, Longzhan, Gibala, on rotten wood, 16 July, 1975, M. Zang 295 (HKAS 5295).
Known distribution: Asia (China), Europe (Austria, Czech Republic, France, Germany#, Hungary, the Netherlands, Slovenia, Sweden, United Kingdom), North America (Canada, USA).
Notes: Pezicula carpinea is the type species of the genus, which was previously reported from Europe and North America [2,5]. The specific epithet of the species refers to its common occurrence on Carpinus sp. The sexual and asexual characteristics of the species have been well-documented and illustrated in previous studies [2,5]. The features of the Chinese collection are concordant with those from other regions of the world. The overmatured ascospores were recorded as yellow-brown and muriform [2], but such ascospores were not found in the Chinese material due to the different stage of development.
3.2.3. Species Previously Recorded from China
Pezicula cinnamomea (DC.) Sacc., Syll. Fung. 8: 311, 1889.
≡Peziza cinnamomea DC., Fl. franç., Edn 3 (Paris) 5/6: 23, 1815.
Specimen examined: China, Guangxi, Wuming, Mt. Daming, alt. 1200 m, on rotten twig of a conifer tree, 22 December 1997, W.P. Wu & W.Y. Zhuang 1902 (HMAS 74848).
Known distribution: Asia (China, Japan,), Europe (Austria, Belgium, Czech Republic, Denmark, France#, Germany, Netherlands, Norway, Russia, Spain, Sweden, Switzerland, United Kingdom), North America (Canada, USA), Central America (Costa Rica), Oceania (New Zealand).
Notes: The species was reported from Guangxi Province in China [26]. It occurs on broad-leaved trees as well as conifers [2].
Pezicula ericae (Sigler) P.R. Johnst., in Johnston, Seifert, Stone, Rossman & Marvanová, IMA Fungus 5(1): 104, 2014.
≡Cryptosporiopsis ericae Sigler, in Sigler, Allan, Lim, Berch & Berbee, Stud. Mycol. 53: 57, 2005.
Known distribution: Asia (China), Europe (Norway), North America (USA#).
Notes: Pezicula ericae was originally described in the USA as an endophytic fungus from roots of ericaceous plants [10]. Subsequently, strains of the species were isolated from orchids [39,40], aspen [11], pine [41], spruce [42], fir [43], and other hosts [6,44], in addition to ericaceous plants [45,46,47]. The species is able to promote plant growth [48], improve the drought resistance ability of plants [49], and synthesize compounds with antimicrobial activities against phytopathogenic bacteria and fungi [6,44].
Pezicula heterochroma Verkley, Stud. Mycol. 44: 94, 1999.
Known distribution: Asia (China), North America (Canada#).
Notes: The type of the species grew on recently dead bark of Alnus mollis in Canada [2]. The Chinese materials were recorded as an endophyte of Vaccinium dunalianum, with strong antagonistic effects against a wood-decaying fungus, Poria placenta (Fr.) Cooke (≡ Rhodonia placenta (Fr.) Niemelä, K.H. Larss. & Schigel) [30].
Pezicula magnispora Z.H. Zhong & Zheng Wang, in Zhong, Wang & Pfister, Mycotaxon 78: 161, 2001.
Specimen examined: China, Sichuan, Luding, Mt. Gongga, alt. 1200 m, on herbaceous stem, 19 August 1997, Wang Z. 2097 (holotype, HMAS 75908).
Known distribution: Asia (China#).
Notes: The species was originally described from Sichuan Province and is distinctive in producing large, muriform ascospores (46–58 × 14.5–20 μm) and growing on herbaceous stems [12]. Further distribution was not reported. The type specimen of P. magnispora is too scanty to extract DNA, and sequence data are not available.
Pezicula melanigena (T. Kowalski & Halmschl.) P.R. Johnst., in Johnston, Seifert, Stone, Rossman & Marvanová, IMA Fungus 5(1): 104, 2014.
≡Cryptosporiopsis melanigena T. Kowalski & Halmschl. (as ‘Cryptosporioposis’), in Kowalski, Halmschlager & Schrader, Mycol. Res. 102(3): 348, 1998.
Known distribution: Asia (China), Europe (Austria#).
Notes: Pezicula melanigena was originally isolated from roots of healthy and declining 80–120-yr-old oak trees (Quercus robur and Q. petraea) in Austria and is only known from its asexual stage. It was also stated that the intensity of culture pigmentation, growth rate, size of macroconidia, color of chlamydospores, and shape of setae are variable among isolates of the species [50]. The fungus was detected in forest soil samples from northeastern China via a high-throughput sequencing method [28].
Pezicula neosporulosa Z.L. Yuan & Verkley, Mycoscience 56: 206, 2014.
Known distribution: Asia (China, Korea), Europe (the Netherlands#), Oceania (New Zealand).
Notes: The species was originally introduced to science based on the materials from the Netherlands and China [13], and the Chinese samples were all endophytes of Abies beshanzuensis from Zhejiang Province. Subsequently, a strain was isolated from soil in Korea [51]. It is also known as an endophytic fungus of Pseudowintera colorata, an endemic medicinal plant of New Zealand [52]. The strain VDBF-2 isolated from Vaccinium dunalianum in Yunnan Province showed strong antagonistic effects against plant pathogens and wood-decaying fungi [30]. Based on comparative and pan-genomic analyses of the endophytic P. neosporulosa, an example of convergent genomic adaptation to endophytism was revealed, mostly determined by enzymes involved in carbohydrate metabolism and secondary metabolite biosynthesis [53].
Pezicula ocellata (Pers.) Seaver, North American Cup-fungi, (Inoperculates) (New York): 345, 1951.
≡Peziza ocellata Pers., Syn. Meth. Fung. (Göttingen) 2: 667, 1801.
Specimens examined: China, Ningxia, Mt. Helan, alt. 2600 m, on rotten twig, 23 June 1961, S.J. Han, Q.M. Ma & Y. Liu 2200 (HMAS 32060); Qinghai, Qilian, Ladonggou, alt. 2800 m, on living bark of ?Salix pseudotangii C. Wang & C.Y. Yu, 18 Aug. 1958, Q.M. Ma 521 (HMAS 33643).
Known distribution: Asia (China). Europe (Czech Republic, Denmark, France#, Germany, Luxembourg, Norway, Poland, Sweden, UK), North America (Canada).
Notes: Pezicula ocellata is prominent in its sessile and orange to orange-brown apothecia with a white margin [2], and the color illustrations of the sexual and asexual stages were provided by Chen et al. (2016) [5].
Pezicula rhizophila (Verkley & J.D. Zijlstra) P.R. Johnst., in Johnston, Seifert, Stone, Rossman & Marvanová, IMA Fungus 5(1): 104, 2014.
≡Cryptosporiopsis rhizophila Verkley & J.D. Zijlstra, in Verkley, Zijlstra, Summerbell & Berendse, Mycol. Res. 107(6): 694, 2003.
Known distribution: Asia (China), Europe (the Netherlands#).
Notes: Pezicula rhizophila is known as an endophytic fungus of several ericaceous plants [54]. An endophytic strain FLR13 of the species was isolated from roots of cultivated blueberry in China, which showed a strong antagonistic ability against five plant pathogens, the ability of plant growth enhancement, and its application potential as a biocontrol agent in agriculture [7]. The genome data of the strain were available in the NCBI database (Accession: JAJONI000000000), for which no relevant paper has been published.
Pezicula cf. rubi (Lib.) Niessl, in Rabenhorst, Fungi Europ. Exsicc. no. 2122. 1876.
≡Patellaria rubi Lib., Pl. crypt. Arduenna, fasc. (Liège) 3(nos 201–300): no. 231, 1834.
Specimen examined: China, Yunnan, Xishuangbanna, on bark of Tetrastigma sp., 22 October 1988, Korf, Zang, Chen & W.Y. Zhuang 229 (HMAS 72156).
Known distribution: Asia (China), Europe (Austria, Belgium#, Czech Republic, Denmark, France, Germany, Italy, Netherlands, Sweden, UK), North America (Canada).
Notes: Pezicula rubi normally grows on Rubus or Rosa, while the natural substrate of the Chinese collection was recorded as Tetrastigma sp. [25] which is not a common substrate for the fungus.
Pezicula subcarnea J.W. Groves, Mycologia 33: 517. 1941.
Known distribution: Asia (China), North America (Canada#, USA).
3.2.4. Species Excluded from the Fungal Catalogue of China
Pezicula aurantiaca (Rehm) Rehm, Ber. Bayer. Bot. Ges. 13: 198, 1912.
≡Habrostictis aurantiaca Rehm, Ber. naturhist. Augsburg 26: 67, 1881.
Notes: Pezicula aurantiaca was originally described from Austria and also known from Canada [2]. The species was reported from China based on two specimens [21], which were reidentified as P. acericola based on morphological features as well as DNA sequence data. Thus, P. aurantiaca should be excluded from the fungal catalogue of China [29].
4. Discussion
In this study, two novel species and two new Chinese records of Pezicula were discovered based on morphological features, and the positions of two of them, P. ellipsoides and P. acericola, were further confirmed by phylogenetic inference. Eleven species of the genus previously recorded from China were reviewed, of which one misidentified name should be excluded. Up to now, 14 Pezicula species are known from the country, among which nine were known as saprobes [12,23,24,25,26], four were plant endophytes [7,13,30,45], and one was even reported in soil [28] but might also be plant-associated.
As far as the lifestyles of Pezicula species are concerned, they are supposed to be initially endophytic in living plant tissues but switch to saprophytic when their hosts die, and those which are weakly phytopathogenic may cause diseases when their hosts are under stress [2,5]. Some of the endophytic species show antifungal effects against plant pathogens, some may produce antibiotics and fungicidal or herbicidal compounds [2,4,6,7,30,56,57,58,59], and others possess the ability to promote plant growth [7,48]. In view of the antimicrobial activity and plant growth promotion of Pezicula, they might have application potential in plant disease control and increasing crop yield.
In most cases, morphological characteristics are sufficient to distinguish Pezicula species, especially those that have both sexual and asexual stages. DNA sequence data are very helpful in discovering new species and understanding relationships among taxa within the group. ITS, LSU, rpb2, tub2, and translation elongation factor-1 alpha (tef-1α) gene sequences have been used in the taxonomic and phylogenetic studies of the genus [5,13,14,18]. Based on the results of the previous studies and our analyses, the variations of the ITS region appear to be able to distinguish Pezicula species correctly. Additional research should be carried out regarding species diversity, the connection of sexual and asexual stages, the selection of supplementary DNA barcodes, and the exploration of potential uses of the members of the genus. Knowing the species diversity is the first step in gaining this knowledge. We expect to find additional Pezicula species in our future surveys in different regions of the country.
5. Conclusions
The species diversity of the genus Pezicula in China was explored. Two new species and two new Chinese records of the genus were discovered, described, and illustrated. The previously reported taxa were reviewed with one excluded from the fungal catalogue of China, resulting in 14 species that are currently known from the country.
Author Contributions
H.-D.Z. conducted morphological observations, molecular experiments, and phylogenetic analyses, and wrote the manuscript draft; W.-Y.Z. revised and polished the draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the National Natural Science Foundation of China (31770019).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Names of the new species were formally registered in the database Fungal Names (https://nmdc.cn/fungalnames, accessed on 1 August 2022). Specimens were deposited in the Herbarium Mycologicum Academiae Sinicae (HMAS). The newly generated sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank, accessed on 1 August 2022). Alignment and trees were submitted to TreeBase (https://www.treebase.org, accessed on 6 September 2022).
Acknowledgments
The authors would like to thank the curators of HMAS and HKAS for their help in sending the specimens on loan. We are also grateful to the reviewers for their valuable suggestions and helpful comments, and to all co-collectors of the specimens for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Index Fungorum. Available online: http://www.indexfungorum.org/names/Names.asp (accessed on 18 June 2022).
- Verkley, G.J.M. A monograph of the genus Pezicula and its anamorphs. Stud. Mycol. 1999, 44, 1–180. [Google Scholar]
- Johnston, P.R.; Seifert, K.A.; Stone, J.K.; Rossman, A.Y.; Marvanova, L. Recommendations on generic names competing for use in Leotiomycetes (Ascomycota). IMA Fungus 2014, 5, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Sucker, J.; Aust, H.J.; Krohn, K.; Ludewig, K.; Jones, P.G.; Döring, D. Biologically active secondary metabolites of endophytic Pezicula species. Mycol. Res. 1995, 99, 1007–1015. [Google Scholar] [CrossRef]
- Chen, C.; Verkley, G.J.M.; Sun, G.; Groenewald, J.Z.; Crous, P.W. Redefining common endophytes and plant pathogens in Neofabraea, Pezicula, and related genera. Fungal Biol. 2016, 120, 1291–1322. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Zhang, D.-J.; Liu, J.-L.; Pan, H.-Y.; Qin, J.-C.; Zhang, Y.-H. Antifungal effects of volatile organic compounds from the endophytic fungus Cryptosporiopsis ericae Cc-HG-7 isolated from Coptis chinensis Franch. Biocontrol. Sci. Tech. 2018, 28, 496–508. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Hu, Y.; Guo, W.; Wu, Z.; Zhang, Y.; Cao, Z. Endophytes from blueberry roots and their antifungal activity and plant growth enhancement effects. Rhizosphere 2021, 20, 100454. [Google Scholar] [CrossRef]
- Dennis, R.W.G. New or interesting British microfungi, II. Kew Bull. 1974, 29, 157–179. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; p. 1527. [Google Scholar]
- Sigler, L.; Allan, T.; Lim, S.R.; Berch, S.; Berbee, M. Two new Cryptosporiopsis species from roots of ericaceous hosts in western North America. Stud. Mycol. 2005, 53, 53–62. [Google Scholar] [CrossRef]
- Wang, W.; Tsuneda, A.; Gibas, C.F.; Currah, R.S. Cryptosporiopsis species isolated from the roots of aspen in central Alberta: Identification, morphology, and interactions with the host, in vitro. Can. J. Bot. 2007, 85, 1214–1226. [Google Scholar] [CrossRef]
- Zhong, Z.; Wang, Z.; Pfister, D.H. Pezicula magnispora, a new species on an herbaceous plant. Mycotaxon 2001, 78, 161–165. [Google Scholar]
- Yuan, Z.; Verkley, G.J.M. Pezicula neosporulosa sp. nov. (Helotiales, Ascomycota), an endophytic fungus associated with Abies spp. in China and Europe. Mycoscience 2015, 56, 205–213. [Google Scholar] [CrossRef]
- Ekanayaka, A.H.; Daranagama, D.A.; Ariyawansa, H.A.; Jones, E.B.G.; Bakhali, A.H.; Hyde, K.D. Pezicula chiangraiensis sp. nov. from Thailand. Mycotaxon 2016, 131, 739–748. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-J.; McKenzie, E.H.C.; Liu, J.-K.; Bhat, D.J.; Dai, D.-Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.-L.; Shang, Q.-J.; et al. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Braun, U.; Bensch, K. Annotated list of taxonomic novelties published in “Fungi Europaei Exsiccati, Klotzschii Herbarium Vivum Mycologicum Continuato, Editio Nova, Series Secunda” Cent. 1 to 26 issued by G.L. Rabenhorst between 1859 and 1881 (secondpart—Cent. 11 to 20). Schlechtendalia 2021, 38, 191–262. [Google Scholar]
- Ma, X.-Y.; Kang, J.-C.; Hyde, K.D.; Doilom, M.; Chomnunti, P. Pezicula endophytica sp. nov., endophytic in Dendrobium in Thailand. Mycotaxon 2021, 136, 563–577. [Google Scholar] [CrossRef]
- Mehrabi, M.; Asgari, B.; Wijayawardene, N.N.; Hyde, K.D. Description of Dermea persica (Dermateaceae, Helotiales), a new asexual Ascomycete from Iran, and an updated key to Dermea species. Phytotaxa 2018, 367, 25–37. [Google Scholar] [CrossRef]
- Abeln, E.C.A.; de Pagter, M.A.; Verkley, G.J.M. Phylogeny of Pezicula, Dermea and Neofabraea inferred from partial sequences of the nuclear ribosomal RNA gene cluster. Mycologia 2000, 92, 685–693. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. They seldom occur alone. Fungal Biol. 2016, 120, 1392–1415. [Google Scholar] [CrossRef]
- Jiang, N.; Tian, C.M. Re-collection of Dermea prunus in China, with a description of D. chinensis sp. nov. MycoKeys 2019, 50, 79–91. [Google Scholar] [CrossRef]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; p. 808. [Google Scholar]
- Liou, S.-C.; Chen, Z.-C. Notes on Taiwan Discomycetes I. (Pezizales and Helotales). Taiwania 1977, 22, 29–43. [Google Scholar] [CrossRef]
- Zhuang, W.-Y.; Wang, Z. Discomycetes of tropical China. II. Collections from Yunnan. Mycotaxon 1998, 69, 339–358. [Google Scholar]
- Zhuang, W.-Y. Fungal flora of tropical Guagnxi, China: Discomycetes of tropica China. IV. More fungi from Guangxi. Mycotaxon 1999, 72, 325–337. [Google Scholar]
- Zhang, C.Y. Diversity of Ericoid Mycorrhiza and the Related Fungi Isolated from Hair Roots of Thododendron fortunei L. Ph.D. Thesis, Beijing Forest University, Beijing, China, 2008. [Google Scholar]
- Zhuang, W.-Y.; Zheng, H.-D.; Zeng, Z.-Q. Species Catalogue of China, Volumen 3 Fungi, Cup-Fungi; Science Press: Beijing, China, 2018; p. 142. [Google Scholar]
- Luo, Q.S. Fungi Diversity in the Soil of Forest in Tangwanghe National Park. Master’s Thesis, Northeast Agricultural University, Haerbin, China, 2018. [Google Scholar]
- Yan, D.; Zeng, W.L.; Chen, X.X.; Liu, Y.; Wang, L.C.; Liu, H.M.; Zhao, P. Phytopathogenic antagonistic activities of the endophytic fungi from the twigs of Vaccinium dunalianum. J. Cent. South Univ. Tech. 2021, 41, 64–71. [Google Scholar]
- Baral, H.-O. Iodine Reaction in Ascomycetes: Why Is Lugol’s Solution Superior to Melzer’s Reagent? Available online: http://www.gbif-mycology.de/HostedSites/Baral/IodineReaction.htm (accessed on 18 March 2011).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A. FigTree v1.4.4, Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, Scotland, 2008. Available online: http://tree.bio.ed.ac.uk/software/figtree/(accessed on 22 May 2019).
- Chen, J.; Dong, H.-L.; Meng, Z.-X.; Guo, S.-X. Cadophora malorum and Cryptosporiopsis ericae isolated from medicinal plants of the Orchidaceae in China. Mycotaxon 2010, 112, 457–461. [Google Scholar] [CrossRef]
- Lee, M.S.; Yang, Y.L.; Wu, C.Y.; Chen, Y.L.; Lee, C.K.; Tzean, S.S.; Lee, T.H. Efficient identification of fungal antimicrobial principles by tandem MS and NMR database. J. Food Drug Anal. 2019, 27, 860–868. [Google Scholar] [CrossRef]
- Xu, F.; Wang, C.; Chu, H.; Tang, M. Research on root fungal community of Pinus tabuliformis in pine wilt disease damaged and undamaged areas. Acta Bot. Boreali-Occident. Sin. 2014, 34, 1627–1634. [Google Scholar]
- Terhonen, E.; Keriö, S.; Sun, H.; Asiegbu, F.O. Endophytic fungi of Norway spruce roots in boreal pristine mire, drained peatland and mineral soil and their inhibitory effect on Heterobasidion parviporum in vitro. Fungal Ecol. 2014, 9, 17–26. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Rao, L.B.; Chen, Y.C.; Zhang, C.L.; Wu, Y.G. From pattern to process: Species and functional diversity in fungal endophytes of Abies beshanzuensis. Fungal Biol. 2011, 115, 197–213. [Google Scholar] [CrossRef]
- Devi, L.S.; Joshi, S.R. Evaluation of the antimicrobial potency of silver nanoparticles biosynthesized by using an endophytic fungus, Cryptosporiopsis ericae PS4. J. Microbiol. 2014, 52, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Zhao, X.Y.; Liu, C.L.; Bai, L.; Zhao, M.; Li, L.L. Diversity and characteristics of colonization of root-associated fungi of Vaccinium uliginosum. Sci. Rep. 2018, 8, 15283. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-C.; Ye, Y.-S.; Lin, W.-R. Characteristics of root-cultivable endophytic fungi from Rhododendron ovatum Planch. Braz. J. Microbiol. 2019, 50, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, M.; Ma, H.-C.; Wu, J.-R. Mycorrhizal diversity of Rhododendron racemosum Franch. in Yunnan Jiaozi Mountain. J. Yunnan Univ. Nat. Sci. 2019, 41, 1062–1072. [Google Scholar]
- Yao, Q.; Chen, M.; Zhou, Y.; Zhu, H.-H. New Pezicula ericae (3-41) Strain Useful in Cultivating Blueberry, Promoting Growth of Blueberry Shoots, and Increasing Amount of Growth. CN107988087A, 4 May 2018. [Google Scholar]
- Mu, D.; Du, N.; Zwiazek, J.J. Inoculation with ericoid mycorrhizal associations alleviates drought stress in lowland and upland velvetleaf blueberry (Vaccinium myrtilloides) seedlings. Plants 2021, 10, 2786. [Google Scholar] [CrossRef]
- Kowalski, T.; Halmschlager, E.; Schrader, K. Cryptosporiopsis melanigena sp. nov., a root-inhabiting fungus of Quercus robur and Q. petraea. Mycol. Res. 1998, 102, 347–354. [Google Scholar] [CrossRef]
- Das, K.; Lee, S.-Y.; Choi, H.-W.; Eom, A.-H.; Cho, Y.-J.; Jung, H.-Y. Taxonomy of Arthrinium minutisporum sp. nov., Pezicula neosporulosa, and Acrocalymma pterocarpi: New records from soil in Korea. Mycobiology 2020, 48, 450–463. [Google Scholar] [CrossRef]
- Purushotham, N.; Jones, E.; Monk, J.; Ridgway, H. Fungal communities in the native New Zealand medicinal plant Pseudowintera colorata (Horopito) are determined by plant organ type and host maturity with key members promoting plant growth. Microorganisms 2021, 9, 2576. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.-L.; Wu, Q.; Xu, L.-X.; Druzhinina, I.S.; Stukenbrock, E.H.; Nieuwenhuis, B.P.S.; Zhong, Z.; Liu, Z.-J.; Wang, X.; Cai, F.; et al. Genomic landscape of a relict fir-associated fungus reveals rapid convergent adaptation towards endophytism. ISME J. 2022, 16, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Verkley, G.J.M.; Zijlstra, J.D.; Summerbell, R.C.; Berendse, F. Phylogeny and taxonomy of root-inhabiting Cryptosporiopsis species, and C. rhizophila sp. nov., a fungus inhabiting roots of several Ericaceae. Mycol. Res. 2003, 107, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Groves, J.W. Pezicula carnea and Pezicula subcarnea. Mycologia 1941, 33, 510–522. [Google Scholar] [CrossRef]
- Noble, H.M.; Langley, D.; Sidebottom, P.J.; Lane, S.J.; Fisher, P.J. An echinocandin from an endophytic Cryptosporiopsis sp. and Pezicula sp. in Pinus sylvestris and Fagus sylvatica. Mycol. Res. 1991, 95, 1439–1440. [Google Scholar] [CrossRef]
- Strobel, G.A.; Miller, R.V.; Martinez-Miller, C.; Condron, M.M.; Teplow, D.B.; Hess, W.M. Cryptocandin, a potent antimycotic from the endophytic fungus Cryptosporiopsis cf. quercina. Microbiology 1999, 145, 1919–1926. [Google Scholar] [CrossRef]
- Sugie, Y.; Dekker, K.A.; Inagaki, T.; Kim, Y.J.; Sakakibara, T.; Sakemi, S.; Sugiura, A.; Brennan, L.; Duignan, J.; Sutcliffe, J.A.; et al. A novel antibiotic CJ-17,572 from a fungus, Pezicula sp. J. Antibiot. 2002, 55, 19–24. [Google Scholar] [CrossRef]
- Wang, J.; Wang, G.; Zhang, Y.; Zheng, B.; Zhang, C.; Wang, L. Isolation and identification of an endophytic fungus Pezicula sp. in Forsythia viridissima and its secondary metabolites. World J. Microbiol. Biotech. 2014, 30, 2639–2644. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood phylogenetic tree inferred from ITS sequences of Pezicula species. Dermea cerasi and D. pruni were used as outgroup taxa. Types are indicated with the superscript letter “T”. The new species proposed in the present study is in bold. Bootstrap support values ≥50% and Bayesian posterior probabilities ≥0.90 are shown at the nodes. The scale bar represents the number of nucleotide substitutions per site.
Figure 1.
Maximum likelihood phylogenetic tree inferred from ITS sequences of Pezicula species. Dermea cerasi and D. pruni were used as outgroup taxa. Types are indicated with the superscript letter “T”. The new species proposed in the present study is in bold. Bootstrap support values ≥50% and Bayesian posterior probabilities ≥0.90 are shown at the nodes. The scale bar represents the number of nucleotide substitutions per site.

Figure 2.
Pezicula ellipsoides H.D. Zheng & W.Y. Zhuang (HMAS 290888, holotype). (a) Dry apothecia on natural substrate; (b) longitudinal section of apothecium; (c) longitudinal section of apothecium (partial) showing excipular and hymenium structure at margin and upper flank; (d) excipular structure of stipe; (e) hymenium; (f) asci; (g1) IKI reaction of ascus apical ring without KOH pretreatment; (g2) IKI reaction of ascus apical ring after KOH pretreatment; (h) paraphyses; (i) ascospores; (j) ascospores with 0–1 septa; (k) light brown ascospores. Scale bars: (a) = 1 mm; (b) = 200 μm; (c–k) = 20 μm. Mounting media: (b–f, h–k) lactophenol cotton blue; (g1, g2) Lugol’s solution.
Figure 2.
Pezicula ellipsoides H.D. Zheng & W.Y. Zhuang (HMAS 290888, holotype). (a) Dry apothecia on natural substrate; (b) longitudinal section of apothecium; (c) longitudinal section of apothecium (partial) showing excipular and hymenium structure at margin and upper flank; (d) excipular structure of stipe; (e) hymenium; (f) asci; (g1) IKI reaction of ascus apical ring without KOH pretreatment; (g2) IKI reaction of ascus apical ring after KOH pretreatment; (h) paraphyses; (i) ascospores; (j) ascospores with 0–1 septa; (k) light brown ascospores. Scale bars: (a) = 1 mm; (b) = 200 μm; (c–k) = 20 μm. Mounting media: (b–f, h–k) lactophenol cotton blue; (g1, g2) Lugol’s solution.

Figure 3.
Pezicula fusispora W.Y. Zhuang & H.D. Zheng (HMAS 290892, holotype). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecium; (d) longitudinal section of apothecium (partial) showing excipular and hymenium structure; (e) paraphyses; (f) IKI reaction of ascus apical ring without KOH pretreatment; (g–i) ascospores; (j,k) asci. Scale bars: (a,b) = 0.5 mm; (c) = 50 μm; (d,j,k) = 20 μm; (e–i) = 10 μm. Mounting media: (c–e,g,h,j) lactophenol cotton blue; (f,i,k) Lugol’s solution.
Figure 3.
Pezicula fusispora W.Y. Zhuang & H.D. Zheng (HMAS 290892, holotype). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecium; (d) longitudinal section of apothecium (partial) showing excipular and hymenium structure; (e) paraphyses; (f) IKI reaction of ascus apical ring without KOH pretreatment; (g–i) ascospores; (j,k) asci. Scale bars: (a,b) = 0.5 mm; (c) = 50 μm; (d,j,k) = 20 μm; (e–i) = 10 μm. Mounting media: (c–e,g,h,j) lactophenol cotton blue; (f,i,k) Lugol’s solution.

Figure 4.
Pezicula acericola (Peck) Peck ex Sacc. & Berl. (HMAS 33726). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecia; (d) longitudinal section of apothecium (half) showing hymenium and excipular structure; (e) hymenium; (f) excipular structure at flank; (g) asci; (h1) IKI reaction of ascus apical ring without KOH pretreatment; (h2) IKI reaction of ascus apical ring after KOH pretreatment; (i) paraphyses; (j) ascospores. Scale bars: (a) = 2 cm; (b) = 2 mm; (c) = 100 μm; (d–j) = 20 μm. Mounting media: (c–g,i,j) lactophenol cotton blue; (h1,h2) Lugol’s solution.
Figure 4.
Pezicula acericola (Peck) Peck ex Sacc. & Berl. (HMAS 33726). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecia; (d) longitudinal section of apothecium (half) showing hymenium and excipular structure; (e) hymenium; (f) excipular structure at flank; (g) asci; (h1) IKI reaction of ascus apical ring without KOH pretreatment; (h2) IKI reaction of ascus apical ring after KOH pretreatment; (i) paraphyses; (j) ascospores. Scale bars: (a) = 2 cm; (b) = 2 mm; (c) = 100 μm; (d–j) = 20 μm. Mounting media: (c–g,i,j) lactophenol cotton blue; (h1,h2) Lugol’s solution.

Figure 5.
Pezicula carpinea (Pers.) Tul. ex Fuckel (HKAS 5295). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecium; (d) excipular structure of margin; (e) excipular structure of flank; (f) excipular structure adjacent to flank and stipe; (g) apices of asci and paraphyses; (h) asci; (i) IKI reaction of apical ring; (j,k) ascospores. Scale bars: (a,b) = 1 mm; (c) = 200 μm; (d,h) = 20 μm; (e–g,i–k) = 10 μm. Mounting media: (c–h,j) lactophenol cotton blue; (i) Lugol’s solution; (k) 3% KOH.
Figure 5.
Pezicula carpinea (Pers.) Tul. ex Fuckel (HKAS 5295). (a,b) Dry apothecia on natural substrate; (c) longitudinal section of apothecium; (d) excipular structure of margin; (e) excipular structure of flank; (f) excipular structure adjacent to flank and stipe; (g) apices of asci and paraphyses; (h) asci; (i) IKI reaction of apical ring; (j,k) ascospores. Scale bars: (a,b) = 1 mm; (c) = 200 μm; (d,h) = 20 μm; (e–g,i–k) = 10 μm. Mounting media: (c–h,j) lactophenol cotton blue; (i) Lugol’s solution; (k) 3% KOH.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of taxa and GenBank accession numbers of ITS sequences used in the study.
| Species | Strain/Voucher Specimen | Locality | ITS |
|---|---|---|---|
| Dermea cerasi (Pers.) Fr. | CBS 136.46 | USA | AF141159 |
| D. pruni (Teng) J.W. Groves | CFCC 53006 | China | MK330016 |
| Pezicula acericola (Peck) Peck ex Sacc. & Berl. | CBS 245.97 | Canada | KR859098 |
| P. acericola | HMAS 33726 | China | OP108434 ** |
| P. aurantiaca (Rehm) Rehm | CBS 201.46 | Canada | KR859102 |
| P. brunnea (Sigler) P.R. Johnst. | CBS 120291T | Canada | NR_155652 |
| P. brunnea | UAMH 10106 | AF149074 | |
| P. californiae (Cheew., Denman & Crous) P.R. Johnst. | CBS 124805T | USA | NR_172218 |
| P. californiae | CBS 143491 | MN161953 | |
| P. carpinea (Pers.) Tul. ex Fuckel | CBS 923.96T | Germany | NR_144927 |
| P. carpinea | CBS 324.97 | The Netherlands | KR859105 |
| P. chiangraiensis Ekanayaka & K.D. Hyde | MFLUCC 15-0170T | Thailand | KU310621 |
| P. cinnamomea (DC.) Sacc. | CBS 239.96 | France | KR859124 |
| P. cinnamomea | HMAS 80649 | The Netherlands | OP108435 ** |
| P. cornina (Peck) P.R. Johnst. | CBS 285.39 | Canada | KR859163 |
| P. corticola (C.A. Jørg.) Nannf. | CBS 259.31 | Denmark | KR859164 |
| P. corticola | CBS 260.31 | Denmark | KR859165 |
| P. corylina J.W. Groves | CBS 140.22 | UK | KR859166 |
| P. corylina | CBS 249.97 | Canada | KR859168 |
| P. diversispora (Robak) P.R. Johnst. | CBS 185.50T | Norway | NR_165205 |
| P. diversispora | CBS 282.47 | KR859171 | |
| P. ellipsoides H.D. Zheng & W.Y. Zhuang | HMAS 290888T | China | OP108433 ** |
| P. endophytica X.Y. Ma, K.D. Hyde & J.C. Kang | MFLUCC 14-0116T | MN908669 | |
| P. endophytica | MFLUCC 14-0140 | MN908667 | |
| P. ericae (Sigler) P.R. Johnst. | CBS 120290T | USA | NR_155653 |
| P. ericae | CBS 120292 | Canada | KR859174 |
| P. eucrita (P. Karst.) P. Karst. | CBS 259.97 | USA | KR859179 |
| P. eucrita | CBS 656.96 | Germany | KR859185 |
| P. fagacearum Chen Chen, Verkley & Crous | CBS 112400T | Italy | NR_155654 |
| P. fagacearum | CBS 112402 | Italy | KR859203 |
| P. frangulae (Fr.) Fuckel | CBS 778.96 | The Netherlands | KR859209 |
| P. frangulae | CBS 100244 | Denmark | KR859204 |
| P. heterochroma Verkley | CBS 199.46T | Canada | NR_136956 |
| P. heterochroma | KC2-035C1 | KX589194 | |
| P. italica W.J. Li, Camporesi & K.D. Hyde | MFLU 16-1284T | Italy | NR_170044 |
| P. italica | MFLU 19-2867 | Italy | MT185542 |
| P. melanigena (T. Kowalski & Halmschl.) P.R. Johnst. | CBS 898.97T | Austria | NR_155611 |
| P. melanigena | 3–5 | KX440147 | |
| P. microspora Chen Chen, Verkley & Crous | CBS 124641T | Italy | NR_155655 |
| P. microspora | G.M. 2016-02-06.2 | OM156464 | |
| P. neocinnamomea Chen Chen, Verkley & Crous | CBS 100248T | Denmark | NR_155656 |
| P. neocinnamomea | CBS 112397 | KR859214 | |
| P. neoheterochroma Chen Chen, Verkley & Crous | CBS 127388T | Austria | NR_155657 |
| P. neosporulosa Z.L. Yuan & Verkley | CBS 101.96T | The Netherlands | NR_138003 |
| P. neosporulosa | CBS 102.96T | The Netherlands | KR859224 |
| P. ocellata (Pers.) Seaver | CBS 949.97 | Luxembourg | KR859233 |
| P. ocellata | CBS 268.39 | Germany | KR859232 |
| P. pruinosa Farl. | CBS 292.39 | Canada | KR859234 |
| P. pseudocinnamomea Chen Chen, Verkley & Crous | CBS 101000T | The Netherlands | NR_155658 |
| P. pseudocinnamomea | SHW465 | MT256130 | |
| P. querciphila (S.C. Lynch, Eskalen & Zambino) P.R. Johnst. | UCROK766T | USA | JX144750 |
| P. querciphila | UCROK637 | USA | JX270770 |
| P. radicicola (Kowalski & Bartnik) P.R. Johnst. | CBS 640.94T | Poland | NR_155610 |
| P. radicicola | CBS 681.83 | Austria | KR859237 |
| P. rhizophila (Verkley & J.D. Zijlstra) P.R. Johnst. | CBS 109839T | The Netherlands | NR_155659 |
| P. rhizophila | CBS 110604 | The Netherlands | KR859240 |
| P. rubi (Lib.) Niessl | CBS 253.97 | USA | KR859250 |
| P. rubi | CBS 593.96 | The Netherlands | KR859253 |
| P. sporulosa Verkley | CBS 224.96T | The Netherlands | NR_137161 |
| P. sporulosa | CBS 225.96T | The Netherlands | KR859262 |
| P. subcarnea J.W. Groves | CBS 203.46 | Canada | AF141171 |
Newly generated sequence data are marked with **. Types are indicated with the superscript “T”.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zheng, H.-D.; Zhuang, W.-Y. Additions to the Knowledge of the Genus Pezicula (Dermateaceae, Helotiales, Ascomycota) in China. Biology 2022, 11, 1386. https://doi.org/10.3390/biology11101386
AMA Style
Zheng H-D, Zhuang W-Y. Additions to the Knowledge of the Genus Pezicula (Dermateaceae, Helotiales, Ascomycota) in China. Biology. 2022; 11(10):1386. https://doi.org/10.3390/biology11101386
Chicago/Turabian StyleZheng, Huan-Di, and Wen-Ying Zhuang. 2022. "Additions to the Knowledge of the Genus Pezicula (Dermateaceae, Helotiales, Ascomycota) in China" Biology 11, no. 10: 1386. https://doi.org/10.3390/biology11101386
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.