
Proof-of-Concept Preclinical Use of Drosophila melanogaster in the Initial Screening of Immunomodulators

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Stocks and Maintenance
2.2. Bacterial Strain and Fly Infection Experiment
2.3. Survival Assay of Drosophila Melanogaster
2.4. Gene Expression Analysis
2.5. Data Analysis
3. Results
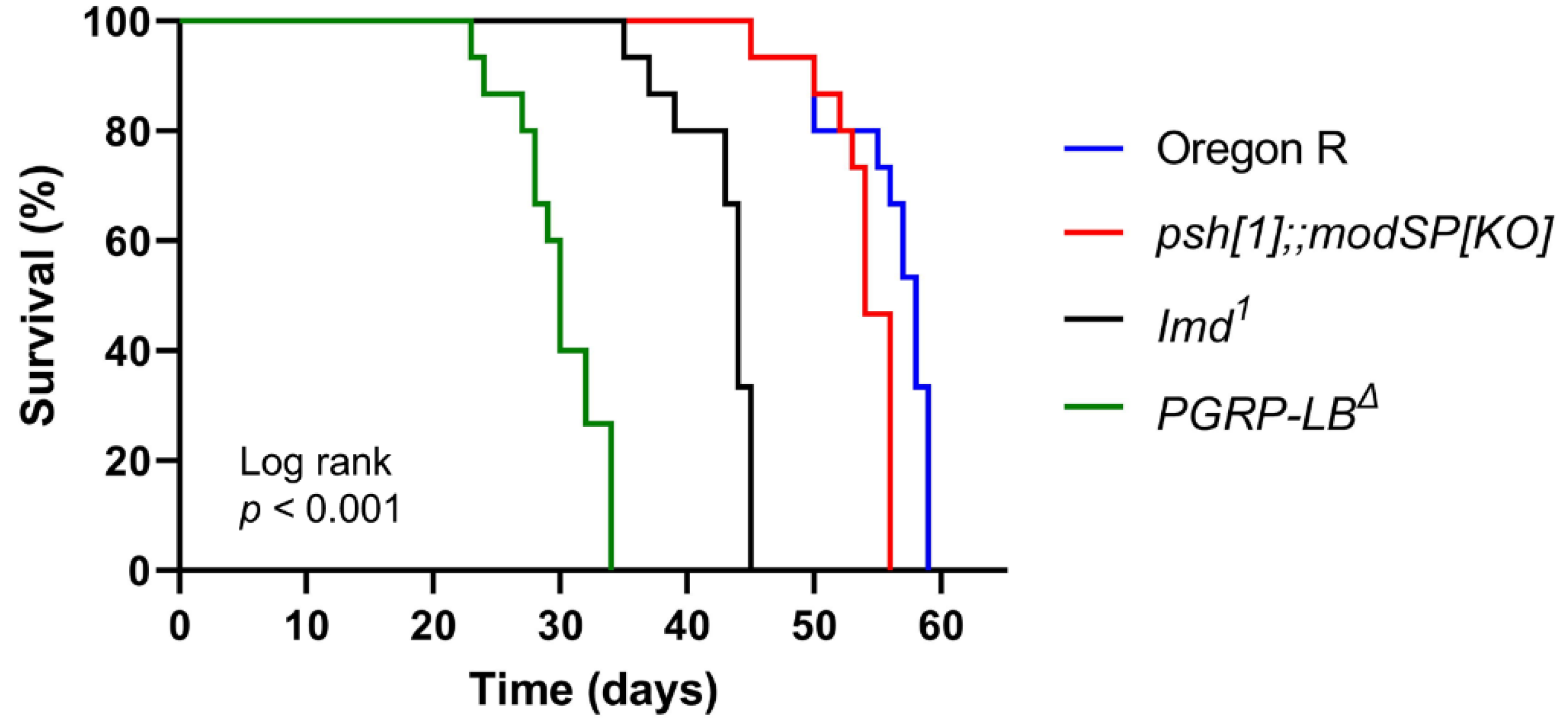
3.1. The Lifespan of Drosophila Melanogaster Is Associated with Its Genotypical Characteristics
3.2. Aspirin and Curcumin Improve the Lifespan of Wild-Type Drosophila melanogaster
3.3. Curcumin and EGCG Provide Antibacterial Protection against Staphylococcus aureus
3.4. Curcumin and EGCG Downregulate the NF-κB-Mediated Immune Response in D. melanogaster
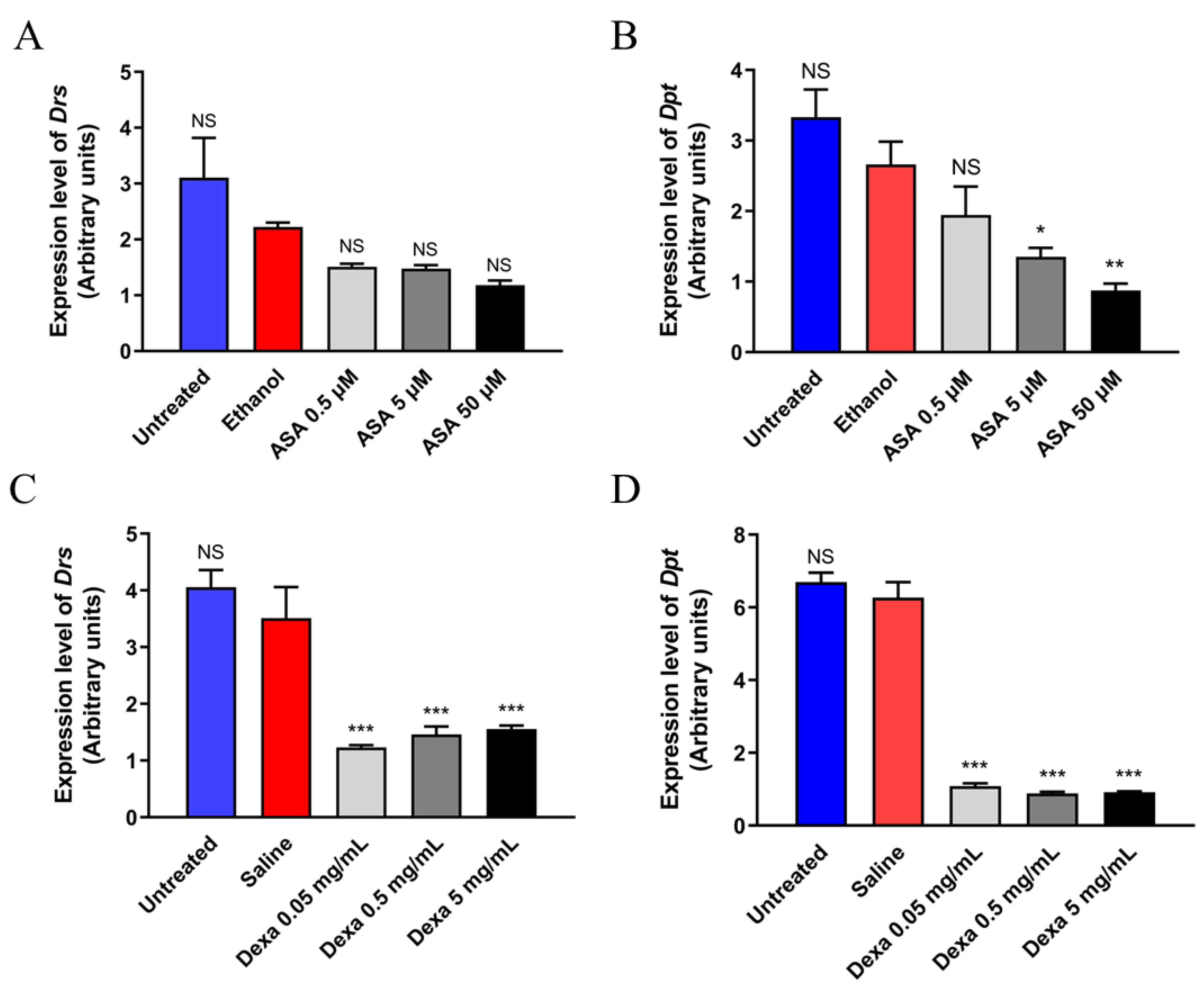
3.5. Aspirin and Dexamethasone Improve the Lifespan of Flies with Hyperactive Immune Phenotype
3.6. Downregulation of Innate Immune-Related Target Genes in D. melanogaster by Aspirin and Dexamethasone
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. Experimental animal models used for evaluation of potential immunomodulators: A mini review. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 211–216. [Google Scholar] [CrossRef]
- Shi, J.; Weng, J.-H.; Mitchison, T.J. Immunomodulatory drug discovery from herbal medicines: Insights from organ-specific activity and xenobiotic defenses. eLife 2021, 10, e73673. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches Towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [Green Version]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster—From microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Nainu, F.; Nakanishi, Y.; Shiratsuchi, A. Fruit fly as a model organism in the study of human diseases and drug discovery. J. Cent. Med. Educ. Sapporo Med. Univ. 2019, 10, 21–32. [Google Scholar]
- Fernández-Hernández, I.; Scheenaard, E.; Pollarolo, G.; Gonzalez, C. The translational relevance of Drosophila in drug discovery. EMBO Rep. 2016, 17, 471–472. [Google Scholar] [CrossRef] [Green Version]
- Morris, T.; Stables, M.; Hobbs, A.; de Souza, P.; Colville-Nash, P.; Warner, T.; Newson, J.; Bellingan, G.; Gilroy, D.W. Effects of Low-Dose Aspirin on Acute Inflammatory Responses in Humans. J. Immunol. 2009, 183, 2089–2096. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.M.; Burrell, C.J.; Douglas, R.M.; Debelle, P.; Davies, L. Adverse effects of aspirin, acetaminophen, and ibuprofen on immune function, viral shedding, and clinical status in rhinovirus-infected volunteers. J. Infect. Dis. 1990, 162, 1277–1282. [Google Scholar] [CrossRef]
- Brox, R.; Hackstein, H. Physiologically relevant aspirin concentrations trigger immunostimulatory cytokine production by human leukocytes. PLoS ONE 2021, 16, e0254606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chang, A.; Zou, Y.; Xu, H.; Cui, J.; Chen, Z.; Li, Y.; Du, Y.; Wu, J.; Yu, J.; et al. Aspirin Attenuates Cardiac Allograft Rejection by Inhibiting the Maturation of Dendritic Cells via the NF-κB Signaling Pathway. Front. Pharmacol. 2021, 12, 706748. [Google Scholar] [CrossRef] [PubMed]
- Bancos, S.; Bernard, M.P.; Topham, D.J.; Phipps, R.P. Ibuprofen and other widely used non-steroidal anti-inflammatory drugs inhibit antibody production in human cells. Cell. Immunol. 2009, 258, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joekel, D.E.; Deplazes, P. Optimized dexamethasone immunosuppression enables Echinococcus multilocularis liver establishment after oral egg inoculation in a rat model. Exp. Parasitol. 2017, 180, 27–32. [Google Scholar] [CrossRef]
- Thachil, A.J.; Shaw, D.P.; Nagaraja, K.V. Effects of Dexamethasone Immunosuppression on Turkey Clostridial Dermatitis. Avian Dis. 2014, 58, 433–436. [Google Scholar] [CrossRef]
- Hundakova, A.; Leva, L.; Toman, M.; Knotek, Z. A ferret model of immunosuppression induced with dexamethasone. Vet. Immunol. Immunopathol. 2022, 243, 110362. [Google Scholar] [CrossRef]
- Yuandani; Jantan, I.; Rohani, A.S.; Sumantri, I.B. Immunomodulatory Effects and Mechanisms of Curcuma Species and Their Bioactive Compounds: A Review. Front. Pharmacol. 2021, 12, 643119. [Google Scholar] [CrossRef]
- Catanzaro, M.; Corsini, E.; Rosini, M.; Racchi, M.; Lanni, C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules 2018, 23, 2778. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.L.; Chen, T.S.; Liou, S.Y.; Hsieh, C.C. Immunomodulatory effects of EGCG fraction of green tea extract in innate and adaptive immunity via T regulatory cells in murine model. Immunopharmacol. Immunotoxicol. 2014, 36, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.-C.; Cheng, H.-Y.; Lin, T.-S.; Chen, W.-H.; Lin, J.-H.; Lin, J.-J.; Lu, C.-C.; Chiang, J.-H.; Hsu, S.-C.; Wu, P.-P.; et al. Epigallocatechin Gallate (EGCG), Influences a Murine WEHI-3 Leukemia Model In Vivo Through Enhancing Phagocytosis of Macrophages and Populations of T- and B-Cells. In Vivo 2013, 27, 627–634. [Google Scholar]
- Saleh, F.; Raghupathy, R.; Asfar, S.; Oteifa, M.; Al-Saleh, N. Analysis of the effect of the active compound of green tea (EGCG) on the proliferation of peripheral blood mononuclear cells. BMC Complement. Altern. Med. 2014, 14, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nainu, F.; Asri, R.M.; Arsyad, A.; Manggau, M.A.; Amir, M.N. In vivo antibacterial activity of green algae Ulva reticulata against Staphylococcus aureus in Drosophila model of infection. Pharmacogn. J. 2018, 10, 993–997. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, M.; Gonsales, A.V.; Sartini, S.; Wahyudin, E.; Nainu, F. In vivo anti-staphylococcal activity of roselle (Hibiscus sabdariffa L.) calyx extract in Drosophila model of infection. J. Herbmed Pharmacol. 2019, 8, 41–46. [Google Scholar] [CrossRef]
- Nainu, F.; Djide, M.N.; Subehan, S.; Sartini, S.; Roska, T.P.; Salim, E.; Kuraishi, T. Protective Signatures of Roselle (Hibiscus sabdariffa L.) Calyx Fractions against Staphylococcus aureus in Drosophila Infection Model. HAYATI J. Biosci. 2020, 27, 306–313. [Google Scholar] [CrossRef]
- Nainu, F.; Asri, R.M.; Djide, M.N.; Ahsan, M.; Arfiansyah, R.; Sartini, S.; Alam, G. Protective effect of green algae Ulva reticulata against Pseudomonas aeruginosa in Drosophila infection model. HAYATI J. Biosci. 2019, 26, 163–171. [Google Scholar] [CrossRef]
- Paredes, J.C.; Welchman, D.P.; Poidevin, M.; Lemaitre, B. Negative Regulation by Amidase PGRPs Shapes the Drosophila Antibacterial Response and Protects the Fly from Innocuous Infection. Immunity 2011, 35, 770–779. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.Y.; Kim, K.H.; Lee, S.H.; Yoon, M.S.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Ahn, S.C.; Park, Y.C.; Park, Y.M. Curcumin inhibits immunostimulatory function of dendritic cells: MAPKs and translocation of NF-κB as potential targets. J. Immunol. 2005, 174, 8116–8124. [Google Scholar] [CrossRef] [Green Version]
- Tallei, T.E.; Fatimawali; Niode, N.J.; Idroes, R.; Zidan, B.M.R.M.; Mitra, S.; Celik, I.; Nainu, F.; Ağagündüz, D.; Bin Emran, T.; et al. A Comprehensive Review of the Potential Use of Green Tea Polyphenols in the Management of COVID-19. Evid.-Based Complement. Altern. Med. 2021, 2021, 7170736. [Google Scholar] [CrossRef]
- Steinmann, J.; Buer, J.; Pietschmann, T.; Steinmann, E. Anti-infective properties of epigallocatechin-3-gallate (EGCG), a component of green tea. Br. J. Pharmacol. 2013, 168, 1059–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, X.; Li, W.; Deng, X.; Deng, Y.; Niu, X. Curcumin protects mice from Staphylococcus aureus pneumonia by interfering with the self-assembly process of α-hemolysin. Sci. Rep. 2016, 6, 28254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisa, A.N.; Rahmasari, M.; Roska, T.P.; Arfiansyah, R.; Sartini, S.; Djide, M.N.; Harapan, H.; Nainu, F. Antimicrobial Effect of Roselle (Hibiscus sabdariffa L.) Water Fraction against Pseudomonas aeruginosa using Drosophila Infection Model. Biointerface Res. Appl. Chem. 2021, 11, 12877–12885. [Google Scholar]
- Sukarsih, Y.; Arfiansyah, R.; Roska, T.P.; Murdifin, M.; Kasim, S.; Nainu, F. Protective Effect of Ethanol Extract of Legundi (Vitex trifolia L.) Leaves against Staphylococcus aureus in Drosophila Infection Model. Biointerface Res. Appl. Chem. 2021, 11, 13989–13996. [Google Scholar]
- Østensen, M.; Khamashta, M.; Lockshin, M.; Parke, A.; Brucato, A.; Carp, H.; Doria, A.; Rai, R.; Meroni, P.; Cetin, I.; et al. Anti-inflammatory and immunosuppressive drugs and reproduction. Arthritis Res. Ther. 2006, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Giles, A.J.; Hutchinson, M.-K.N.D.; Sonnemann, H.M.; Jung, J.; Fecci, P.E.; Ratnam, N.M.; Zhang, W.; Song, H.; Bailey, R.; Davis, D.; et al. Dexamethasone-induced immunosuppression: Mechanisms and implications for immunotherapy. J. ImmunoTher. Cancer 2018, 6, 51. [Google Scholar] [CrossRef]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Hussain, M.; Javeed, A.; Ashraf, M.; Zhao, Y.; Mukhtar, M.M.; Rehman, M.U. Aspirin and immune system. Int. Immunopharmacol. 2012, 12, 10–20. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Aggarwal, B.B. “Spicing Up” of the Immune System by Curcumin. J. Clin. Immunol. 2007, 27, 19–35. [Google Scholar] [CrossRef]
- Pae, M.; Wu, D. Immunomodulating effects of epigallocatechin-3-gallate from green tea: Mechanisms and applications. Food Funct. 2013, 4, 1287–1303. [Google Scholar] [CrossRef] [PubMed]
- Gorzalczany, S.B.; Rodriguez Basso, A.G. Strategies to apply 3Rs in preclinical testing. Pharmacol. Res. Perspect. 2021, 9, e00863. [Google Scholar] [CrossRef] [PubMed]
- Nainu, F.; Rahmatika, D.; Emran, T.B.; Harapan, H. Potential Application of Drosophila melanogaster as a Model Organism in COVID-19-Related Research. Front. Pharmacol. 2020, 11, 588561. [Google Scholar] [CrossRef] [PubMed]
- van de Leemput, J.; Han, Z. Drosophila, a powerful model to study virus-host interactions and pathogenicity in the fight against SARS-CoV-2. Cell Biosci. 2021, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Herrera, P.; Cauchi, R.J. ACE and ACE2: Insights from Drosophila and implications for COVID-19. Heliyon 2021, 7, e08555. [Google Scholar] [CrossRef]
- Burden, N.; Chapman, K.; Sewell, F.; Robinson, V. Pioneering better science through the 3Rs: An introduction to the national centre for the replacement, refinement, and reduction of animals in research (NC3Rs). J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 198–208. [Google Scholar]
- Prescott, M.J.; Lidster, K. Improving quality of science through better animal welfare: The NC3Rs strategy. Lab Anim. 2017, 46, 152–156. [Google Scholar] [CrossRef]
- Nainu, F.; Salim, E.; Asri, R.M.; Hori, A.; Kuraishi, T. Neurodegenerative disorders and sterile inflammation: Lessons from a Drosophila model. J. Biochem. 2019, 166, 213–221. [Google Scholar] [CrossRef]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Models Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Willoughby, L.F.; Schlosser, T.; Manning, S.A.; Parisot, J.P.; Street, I.P.; Richardson, H.E.; Humbert, P.O.; Brumby, A.M. An in vivo large-scale chemical screening platform using Drosophila for anti-cancer drug discovery. Dis. Models Mech. 2013, 6, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Cagan, R. Drosophila as a novel therapeutic discovery tool for thyroid cancer. Thyroid 2010, 20, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Tzelepis, I.; Kapsetaki, S.E.; Panayidou, S.; Apidianakis, Y. Drosophila melanogaster: A first step and a stepping-stone to anti-infectives. Curr. Opin. Pharmacol. 2013, 13, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Maitra, U.; Ciesla, L. Using Drosophila as a platform for drug discovery from natural products in Parkinson’s disease. MedChemComm 2019, 10, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.; Sinadinos, C.; Johnston, A.; Sealey, M.; Mudher, A. Using Drosophila models of neurodegenerative diseases for drug discovery. Expert Opin. Drug Discov. 2011, 6, 129–140. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nainu, F.; Bahar, M.A.; Sartini, S.; Rosa, R.A.; Rahmah, N.; Kamri, R.A.; Rumata, N.R.; Yulianty, R.; Wahyudin, E. Proof-of-Concept Preclinical Use of Drosophila melanogaster in the Initial Screening of Immunomodulators. Sci. Pharm. 2022, 90, 11. https://doi.org/10.3390/scipharm90010011
Nainu F, Bahar MA, Sartini S, Rosa RA, Rahmah N, Kamri RA, Rumata NR, Yulianty R, Wahyudin E. Proof-of-Concept Preclinical Use of Drosophila melanogaster in the Initial Screening of Immunomodulators. Scientia Pharmaceutica. 2022; 90(1):11. https://doi.org/10.3390/scipharm90010011
Chicago/Turabian StyleNainu, Firzan, Muh. Akbar Bahar, Sartini Sartini, Reski Amalia Rosa, Nur Rahmah, Reski Amelia Kamri, Nur Rahma Rumata, Risfah Yulianty, and Elly Wahyudin. 2022. "Proof-of-Concept Preclinical Use of Drosophila melanogaster in the Initial Screening of Immunomodulators" Scientia Pharmaceutica 90, no. 1: 11. https://doi.org/10.3390/scipharm90010011