
Molecular Phylogeny of Trifolium L. Section Trifolium with Reference to Chromosome Number and Subsections Delimitation

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. DNA Isolation and DNA Sequence
4.3. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellison, N.W.; Liston, A.; Steiner, J.J.; Williams, W.M.; Taylor, N.L. Molecular phylogenetics of the clover genus (Trifolium—Leguminosae). Mol. Phylogenet. Evol. 2006, 39, 688–705. [Google Scholar] [CrossRef]
- Gillett, J.M.; Taylor, N.L.; Collins, M. World of Clovers; Collins, M., Ed.; Iowa State, University Press: Ames, IA, USA, 2001. [Google Scholar]
- Scoppola, A.; Tirado, J.L.; Gutiérrez, F.M.; Magrini, S. The genus Trifolium (Fabaceae) in south Europe: A critical review on species richness and distribution. Nord. J. Bot. 2018, 36, njb-01723. [Google Scholar] [CrossRef]
- Sprent, J.I.; Platzmann, J. Nodulation in Legumes; Royal Botanic Gardens Kew: London, UK, 2001. [Google Scholar]
- Taylor, N.; Smith, R. Red clover breeding and genetics. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1980; Volume 31, pp. 125–154. [Google Scholar]
- Katznelson, J.; Putiyevsky, E. Cytogenetic studies in trifolium spp. related to berseem II. Relationships within the echinata group. Theor. Appl. Genet. 1974, 44, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Putiyevsky, E.; Katznelson, J. Cytogenetic studies in Trifolium spp. related to berseem I. Intra- and interspecific hybrid seed formation. Theor. Appl. Genet. 1973, 43, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Putiyevsky, E.; Katznelson, J.; Zohary, D. Cytogenetic studies in Trifolium spp related to Berseem. IV. The relationships in the Alexandrinum and Vavilovi crossability groups and the origin of cultivated Berseem. Theor. Appl. Genet. 1975, 45, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Quesenberry, K.H. Red Clover Science. In Current Plant Sciences and Biology in Agriculture; Kluwer Academic Publisher: Dordrecht, The Netherlands, 1996; Volume 28, pp. 141–160. [Google Scholar]
- Williams, W.M.; Ellison, N.W.; Ansari, H.A.; Verry, I.M.; Hussain, S.W. Experimental evidence for the ancestry of allotetraploid Trifolium repens and creation of synthetic forms with value for plant breeding. BMC Plant Biol. 2012, 12, 1–10. [Google Scholar] [CrossRef]
- Morris, J.; Greene, S. Defining a Multiple-Use Germplasm Collection for the Genus Trifolium. Crop Sci. 2001, 41, 893–901. [Google Scholar] [CrossRef]
- Boissier, E. Trifolium. In Flora Orientalis, Sive, Enumeratio Plantarum in Oriente a Graecia et Aegypto ad Indiae Fines Hucusque Observatarum; Georg, H., Ed.; Basileae: Genevae, Switzerland, 1872; Volume 2. [Google Scholar]
- Zohary, M. Origins and evolution in the genus. Trifolium. Bot. Notiser. 1972, 125, 501–511. [Google Scholar]
- Zohary, M.; Heller, D. The Genus Trifolium; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1984. [Google Scholar]
- Goldblatt, P.; Johnson, D.E. Tropicos Database: Index to Plant Chromosome Numbers. 2001–2003; Missouri Botanical Garden: St Louis, MO, USA, 2006. [Google Scholar]
- Watson, L.; Sayed-Ahmed, H.; Badr, A. Molecular phylogeny of Old World Trifolium (Fabaceae), based on plastid and nuclear markers. Plant Syst. Evol. 2000, 224, 153–171. [Google Scholar] [CrossRef]
- Badr, A. Electrophoretic studies of seed proteins in relation to chromosomal criteria and the relationships of some taxa of Trifolium. Taxon 1995, 44, 183–191. [Google Scholar] [CrossRef]
- Badr, A.; El-Shazly, H.H.; Watson, L.E. Origin and ancestry of Egyptian clover (Trifolium alexandrinum L.) as revealed by AFLP markers. Genet. Resour. Crop Evol. 2008, 55, 21–31. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Zoric, L.; Merkulov, L.; Lukovic, J.; Boza, P. Comparative analysis of qualitative anatomical characters of Trifolium L.(Fabaceae) and their taxonomic implications: Preliminary results. Plant Syst. Evol. 2012, 298, 205–219. [Google Scholar] [CrossRef]
- Tripp, E.A.; Lendemer, J.C. Sleepless nights: When you can’t find anything to use but molecules to describe new taxa. Taxon 2014, 63, 969–971. [Google Scholar] [CrossRef]
- Senn, H.A. Chromosome number relationship Leguminosae. Bibliogr. Genet. 1938, 12, 175–336. [Google Scholar]
- Britten, E.J. Chromosome numbers in the genus Trifolium. Cytologia 1963, 28, 428–449. [Google Scholar] [CrossRef]
- Pritchard, R.D. Equity theory: A review and critique. Organ. Behav. Hum. Decis. Process. 1969, 4, 176–211. [Google Scholar] [CrossRef]
- Goldblatt, P. Cytology and Phylogeny of Leguminosae; Polhill, R.M., Raven, P.H., Eds.; Royal Botanic Gardens: Kew, UK, 1981; Volume 2, pp. 427–463. [Google Scholar]
- El-Kholy, M. Cytology of Some Trifolium Species; Al-Azhar University: Cairo, Egypt, 1990. [Google Scholar]
- Bermúdez-Torres, K.; Ferval, M.; Hernández-Sánchez, A.M.; Tei, A.; Gers, C.; Wink, M.; Legal, L. Molecular and Chemical Markers to Illustrate the Complex Diversity of the Genus Lupinus (Fabaceae). Diversity 2021, 13, 263. [Google Scholar] [CrossRef]
- Khalili, Z.; Ghalenoyi, S.; Maassoumi, A.A.; Kazempour-Osaloo, S. Phylogenetic relationships, biogeography and taxonomic delimitation of Astragalus sect. Acanthophace (Fabaceae) using cpDNA and nrDNA ITS sequences analyses. Plant Biosyst. 2021, 155, 291–301. [Google Scholar]
- Ashkenazy, H.; Cohen, O.; Pupko, T.; Huchon, D. Indel reliability in indel-based phylogenetic inference. Genome Biol. Evol. 2014, 6, 3199–3209. [Google Scholar] [CrossRef]
- Kalinkina, V.A.; Mikhaylova, Y.V.; Kislov, D.E. Diversity and taxonomy of the Trifolium lupinaster polymorphic complex in Eastern Europe and Asia. Flora 2020, 267, 151597. [Google Scholar] [CrossRef]
- Jreisat, C.S.; Laten, H.M. Ribosomal RNA Internal Transcribed Regions Identify Possible Misidentification or Mislabeling among Trifolium (Clover) Specimens from Germplasm Collections. Crop Sci. 2017, 57, 322–326. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 1–9. [Google Scholar] [CrossRef]
- Aaronsohn, A. Agricultural Explorations in Palestine; USDA: Washington, DC, USA, 1910; 80.
- Bobrov, E. Vidi Kleverov URSS (in Russian). Acta Inst. Bot. N. Acad. Sci. USSR Ser. 1 1947, 6, 164–336. [Google Scholar]
- Trabut, M. L’indigénat de la Fève en Algérie. Bull. Soc. Bot. Fr. Rev. Bibl. 1910, 57, 424–429. [Google Scholar] [CrossRef]
- Eig, A. A historical–phytosociological essay on Palestine forests of Quercus aegilops L. ssp.ithaburensis (Des) in past and present. Beih. Bot. Cent. 1934, 51B, 225–272. [Google Scholar]
- Snell, R.; Aarssen, L.W. Life history traits in selfing versus outcrossing annuals: Exploring the ‘time-limitation’ hypothesis for the fitness benefit of self-pollination. BMC Ecol. 2005, 5, 1–14. [Google Scholar] [CrossRef]
- Aarssen, L. Why are most selfers annuals? A new hypothesis for the fitness benefit of selfing. Oikos 2000, 89, 606–612. [Google Scholar] [CrossRef]
- Nosrati, H.; Feizi, M.H.; Razban-Haghighi, A.; Seyed-Tarrah, S. Impact of life history on genetic variation in Trifolium (Fabaceae) estimated by ISSR. Environ. Exp. Biol. 2015, 13, 83–88. [Google Scholar]
- Malaviya, D.R.; Roy, A.K.; Kaushal, P.; Kumar, B. Affinity between Trifolium alexandrinum and T. apertum—cytological investigation in embryo rescued hybrid. Cytologia 2004, 69, 425–429. [Google Scholar] [CrossRef]
- Riasat, M.; Sadeghian, S. Karyology Study of Ten Trifolium Species in Fars Province. J. Genet. Res. 2019, 5, 136–142. [Google Scholar]
- Kazimierski, T.; Kazimierska, E. Structure of anther heads in some Trifolium L. species. Acta Soc. Bot. Pol. 1974, 43, 321–329. [Google Scholar] [CrossRef]
- Kiran, Y.; Şahİn, A.; Türkoğlu, İ.; Emre, İ.; Kurşat, M. Karyological notes on seven Trifolium L. taxa from Turkey. Turk. J. Sci. Technol. 2015, 10, 11–16. [Google Scholar]
- Kataoka, R.; Hara, M.; Kato, S.; Isobe, S.; Sato, S.; Tabata, S.; Ohmido, N. Integration of linkage and chromosome maps of red clover (Trifolium pratense L.). Cytogenet. Genome Res. 2012, 137, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Higgins, D.G.; Bleasby, A.J.; Fuchs, R. CLUSTAL V: Improved software for multiple sequence alignment. Bioinformatics 1992, 8, 189–191. [Google Scholar] [CrossRef]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence-based phylogenetic analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef]
- Young, N.D.; Healy, J. GapCoder automates the use of indel characters in phylogenetic analysis. BMC Bioinform. 2003, 4, 1–6. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]



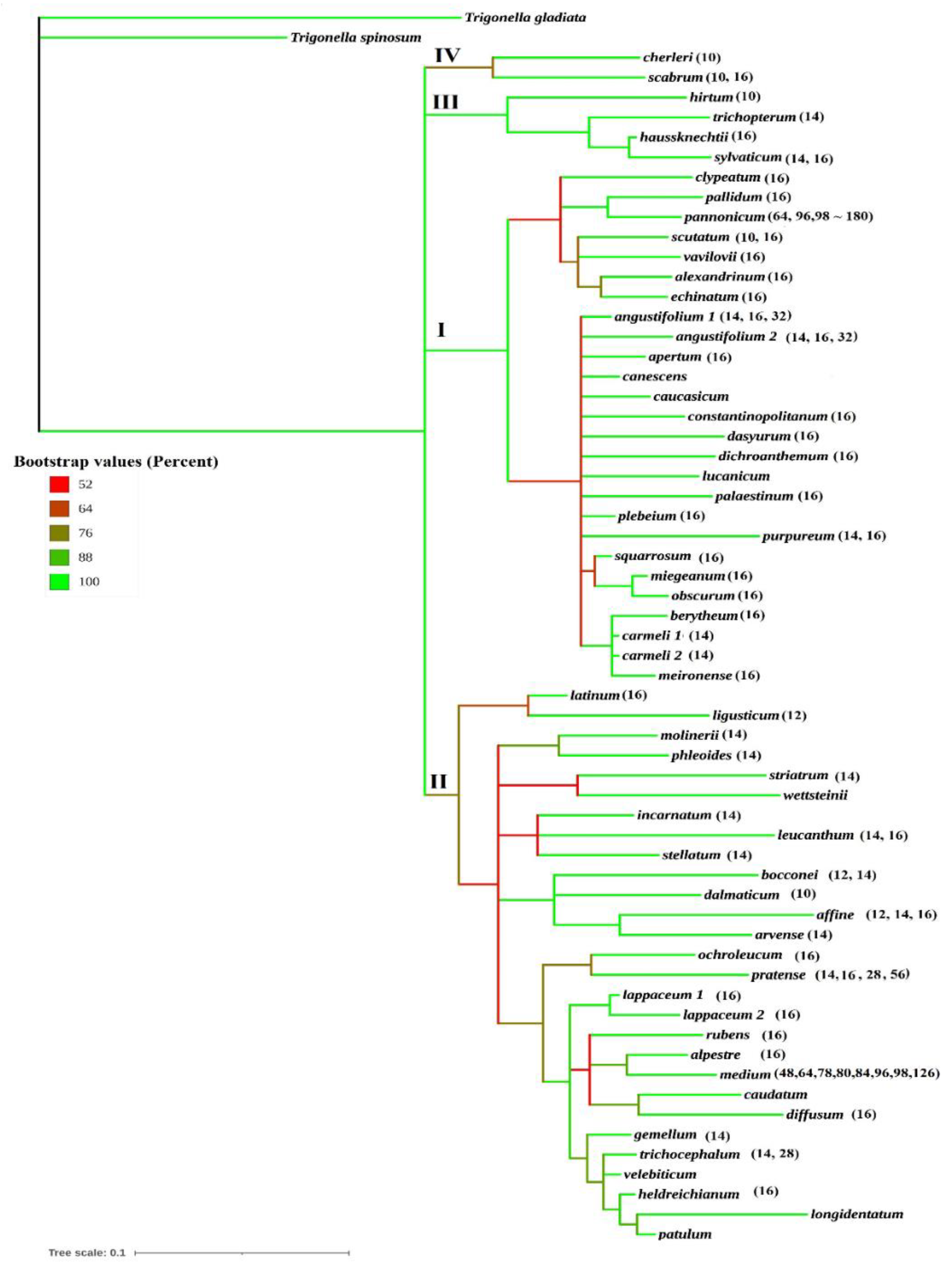{kind=link}
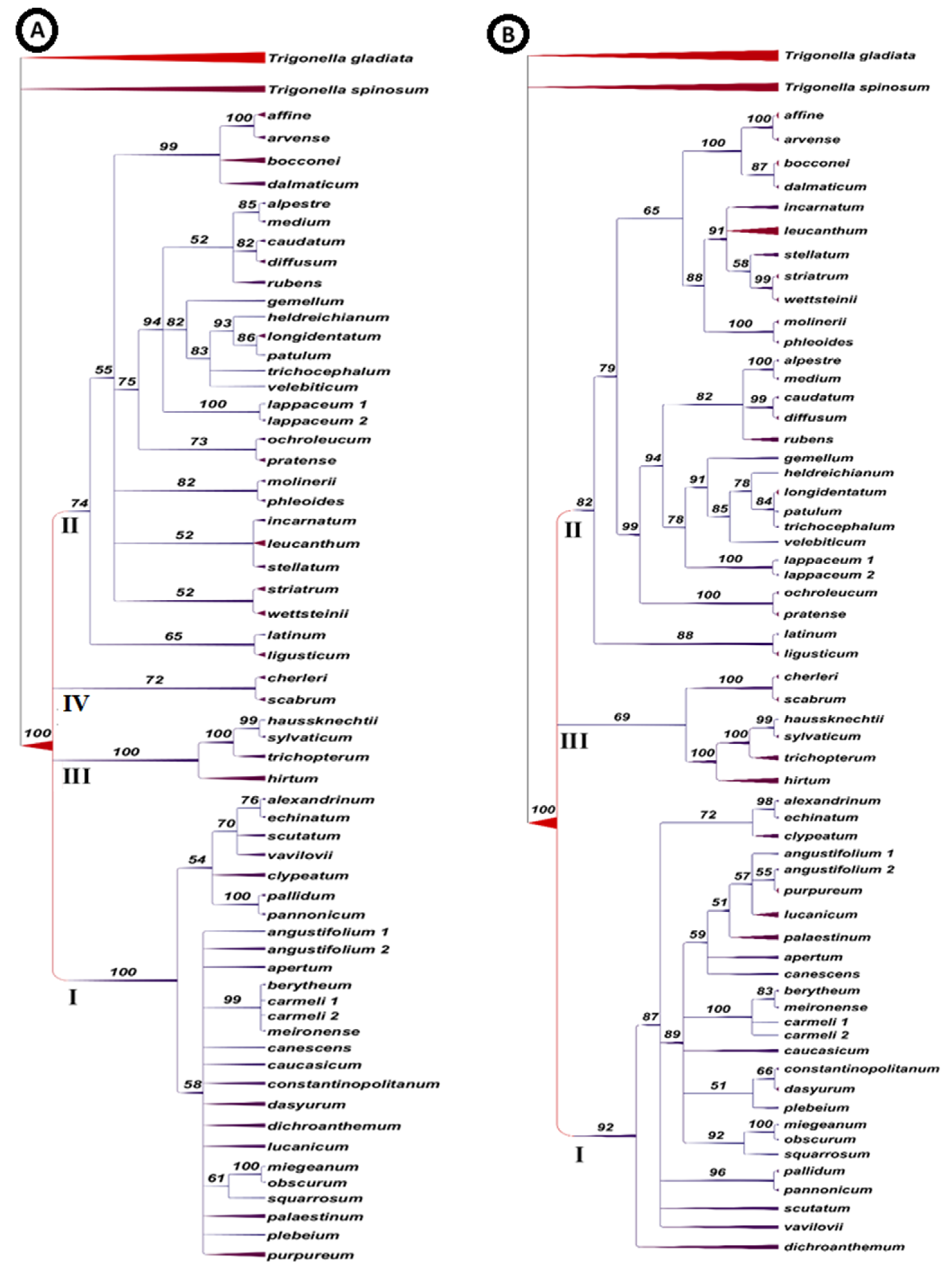{kind=link}
| Trifolium Species | 2n Chromosome Number | Subsection | New Clade |
|---|---|---|---|
| T. alexandrinum | 16 | Alexandrina | Clade I |
| T. vavilovii | 16 | ||
| T. apertum | 16 | ||
| T. berytheum | 16 | ||
| T. meironense | 16 | ||
| T. scutatum | 10, 16 | Clypeata | |
| T. clypeatum | 16 | ||
| T. plebeium | 16 | ||
| T. pallidum | 16 | Trifolium | |
| T. lucanicum | - | Scabroidea | |
| T. dasyurum | 16 | Angustifolia | |
| T. dichroanthum | 16 | ||
| T. palaestinum | 16 | ||
| T. angustifolium | 14, 16, 32 | ||
| T. purpureum | 14, 16 | ||
| T. carmeli | 14 | Echinata | |
| T. echinatum | 16 | ||
| T. canescens | - | Ochroleuca | |
| T. caucasicum | - | ||
| T. pannonicum | 64, 96, 98~180 | ||
| T. miegeanum | 16 | Urceolata | |
| T. obscurum | 16 | ||
| T. squarrosum | 16 | ||
| T. constantinopolitanum | 16 | ||
| T. striatum | 14 | Stenosemium | Clad II |
| T. latinum | 16 | Echinata | |
| T. alpestre | 16 | Alpesteria | |
| T. rubens | 16 | ||
| T. medium | 48, 64, 78, 80, 84, 96, 98, 126 | Intermedia | |
| T. heldreichianum | 16 | ||
| T. patulum | - | ||
| T. velebiticum | - | ||
| T. wettsteinii | - | ||
| T. caudatum | - | Ochroleuca | |
| T. longidentatum | - | ||
| T. trichocephalum | 14, 28 | ||
| T. ochroleucum | 16 | ||
| T. gemellum | 14 | Pheloidea | |
| T. phleoides | 14 | ||
| T. ligusticum | 12 | ||
| T. leucanthum | 14, 16 | Urceolata | |
| T. diffusum | 16 | Trifolium | |
| T. pratense | 14, 16, 28, 56 | ||
| T. molinerii | 14 | Stellata | |
| T. incarnatum | 14 | ||
| T. stellatum | 14 | ||
| T. lappaceum | 16 | Lappacea | |
| T. affine | 12, 14, 16 | Arvensia | |
| T. arvense | 14 | ||
| T. bocconei | 12, 14 | Trichoptera | |
| T. dalmaticum | 10 | Scabroidea | |
| T. sylvaticum | 14, 16 | Stellata | Clade III |
| T. hirtum | 10 | Lappacea | |
| T. cherleri | 10 | ||
| T. haussknechtii | 16 | Angustifolia | |
| T. scabrum | 10, 16 | Scabroidea | |
| T. trichopterum | 14 | Trichoptera |
| No | Trifolium Species | Source and Accession Number * | Country of Origin | Gene Bank ITS1/ITS2 |
|---|---|---|---|---|
| 1 | T. affine Presl. | SRPIS 369014 | Turkey | MW683982/MW684022 |
| 2 | T. alexandrinum L. | ARC, EG 10 | Egypt | AF154381/AF154605 |
| 3 | T. alpestre L. | Kew 11097 | Switzerland | AF154400/AF154624 |
| 4 | T. angustifolium L. | MU 9491 | USA | MW683983/MW684023 |
| 5 | T. angustifolium L. | BGUZ 8023 | Switzerland | AF154388/AF154612 |
| 6 | T. apertum Bobrov | SRPIS 314117 | Russia | MW683984/MW684024 |
| 7 | T. arvense L. | SRPIS 120077 | Turkey | MW683985/MW684025 |
| 8 | T. berytheum Bois. and T. Blanche | SRPIS 369019 | Turkey | MW683986/MW684026 |
| 9 | T. bocconei Savi | SRPIS 224643 | Morocco | MW683987/MW684027 |
| 10 | T. canescens Willd. | WRPIS 418866 | Italy | MW683988/MW684028 |
| 11 | T. carmeli Boiss. | IPK TRIF 100/75 | USA | MW683989/MW684029 |
| 12 | T. carmeli | SRPIS 353422 | Israel | MW683990/MW684030 |
| 13 | T. caucasicum Tausch* | WRPIS 597496 | Russia | MW683991/MW684031 |
| 14 | T. caudatum Boiss. | WRPIS 251858 | Italy | MW683992/MW684032 |
| 15 | T. cherleri L. | Italy 589 | Italy | AF154357/AF154582 |
| 16 | T. clypeatum L. | ICARDA 1149 | Syria | AF154383/AF154607 |
| 17 | T. constantinopolitanum Sert. | ICARDA 1114 | Syria | AF154386/AF154610 |
| 18 | T. dalmaticum Vis. | WRPIS 516292 | Serbia | MW683993/MW684033 |
| 19 | T. dasyurum C. Presl. | SRPIS 369029 | Australia | AF154387/AF154611 |
| 20 | T. dichroanthum Boiss. | SRPIS 292474 | Israel | MW683994/MW684034 |
| 21 | T. diffusum Ehrh. | Kew 1094 | Serbia | AF154401/AF154625 |
| 22 | T. echinatum N.B. | ICARDA 1758 | Syria | AF154382/AF154606 |
| 23 | T. gemellum Willd. | SRPIS 302967 | Spain | MW683995/MW684035 |
| 24 | T. haussknechtii Boiss. | WRPIS G31176-98i | Bulgaria | MW683996/MW684036 |
| 25 | T. heldreichianum Hausskn. | WRPIS 419289 | Greece | MW683997/MW684037 |
| 26 | T. hirtum All. | Kew 25085 | Greece | AF154359/AF154932 |
| 27 | T. incarnatum L. | Kew 55767 | England | AF154392/AF154932 |
| 28 | T. lappaceum L. | ICARDA 108405 | Syria | MW683998/MW684038 |
| 29 | T. lappaceum L.. | Italy 529 | Italy | AF154395/AF154619 |
| 30 | T. latinum Sebast. | IOWA State | USA | MW683999/MW684039 |
| 31 | T. leucanthum M.B. | ICARDA 1111 | Syria | AF154394/AF154618 |
| 32 | T. ligusticum Lois. | SRPIS 419415 | Greece | MW684000/MW684040 |
| 33 | T. longidentatum Nabelek | WRPIS 542831 | Bosnia, Herzegovina | MW684001/MW684041 |
| 34 | T. lucanicum Gasp. Ex Guss. | Kew 13921 | Greece | AF154390/AF154614 |
| 35 | T. medium. L. | WRPIS 325481 | Russia | MW684002/MW684042 |
| 36 | T. meironense Zoh. and Lern. | ICARDA 69062 | Algeria | MW684003/MW684043 |
| 37 | T. miegeanum Maire | SRPIS 258406 | Portugal | MW684004/MW684044 |
| 38 | T. molinerii (incarnatum L.) | SRPIS 591667 | Bulgaria | MW684005/MW684045 |
| 39 | T. obscurum Savi | SRPIS 369057 | Italy | MW684006/MW684046 |
| 40 | T. ochroleucum Huds. | Kew 7559 | Greece | AF154397/AF154621 |
| 41 | T. palaestinum Boiss. | SRPIS 369060 | Palestine | MW684007/MW684047 |
| 42 | T. pallidum Waldst. and Kit. | Kew 31516 | Greece | AF154385/AF154609 |
| 43 | T. pannonicum Jacq. | BGUZ s.n. | Switzerland | AF154384/AF154608 |
| 44 | T. patulum Tausch | WRPIS 604678-97i | Serbia | MW684008/MW684048 |
| 45 | T. phleoides Willd. | SRPIS 298420 | Turkey | MW684009/MW684049 |
| 46 | T. plebeium Boiss. | ICARDA 67905 | Syria | MW684010/MW684050 |
| 47 | T. pratense L. | JBVU 1044 | France | AF154396/AF154620 |
| 48 | T. purpureum Loisel | ICARDA 938 | Syria | AF154389/AF154613 |
| 49 | T. rubens L. | IPP 135 | France | AF154398/AF154622 |
| 50 | T. scabrum L. | ICARDA 939 | Syria | AF154358/AF154583 |
| 51 | T. scutatum Boiss. | ICARDA 67459 | Syria | MW684011/MW684051 |
| 52 | T. squarrosum L. | SRPIS 233720 | Italy | MW684012/MW684052 |
| 53 | T. stellatum L. | ICARDA 947 | Syria | AF154393/AF154617 |
| 54 | T. striatum L. | Kew 5337 | England | AF154391/AF154615 |
| 55 | T. sylvaticum Gerard | SRPIS 120194 | Turkey | MW684013/MW684053 |
| 56 | T. trichocephalum Bieb. | WRPIS 251210-64 | North Macedonia | MW684014/MW684054 |
| 57 | T. trichopterum Pancic | SRPIS 583430 | Bulgaria | MW684015/MW684055 |
| 58 | T. vavilovii Eig | Vencent 11160 | USA | MW684016/MW684056/ |
| 59 | T. velebiticum Degen | WRPIS G30679-97i | USA | MW684017/MW684057 |
| 60 | T. wettsteinii Dorfl. and Hay | WRPIS G31121 | USA | MW684018/MW684058 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, H.I.S.; Badr, A.; El-Shazly, H.H.; Watson, L.; Fouad, A.S.; Ellmouni, F.Y. Molecular Phylogeny of Trifolium L. Section Trifolium with Reference to Chromosome Number and Subsections Delimitation. Plants 2021, 10, 1985. https://doi.org/10.3390/plants10101985
Ahmed HIS, Badr A, El-Shazly HH, Watson L, Fouad AS, Ellmouni FY. Molecular Phylogeny of Trifolium L. Section Trifolium with Reference to Chromosome Number and Subsections Delimitation. Plants. 2021; 10(10):1985. https://doi.org/10.3390/plants10101985
Chicago/Turabian StyleAhmed, Hanan I. Sayed, Abdelfattah Badr, Hanaa H. El-Shazly, Linda Watson, Ahmed S. Fouad, and Faten Y. Ellmouni. 2021. "Molecular Phylogeny of Trifolium L. Section Trifolium with Reference to Chromosome Number and Subsections Delimitation" Plants 10, no. 10: 1985. https://doi.org/10.3390/plants10101985