
Fantastic Downy Mildew Pathogens and How to Find Them: Advances in Detection and Diagnostics

 ,
,  , , and
, , and 
Abstract
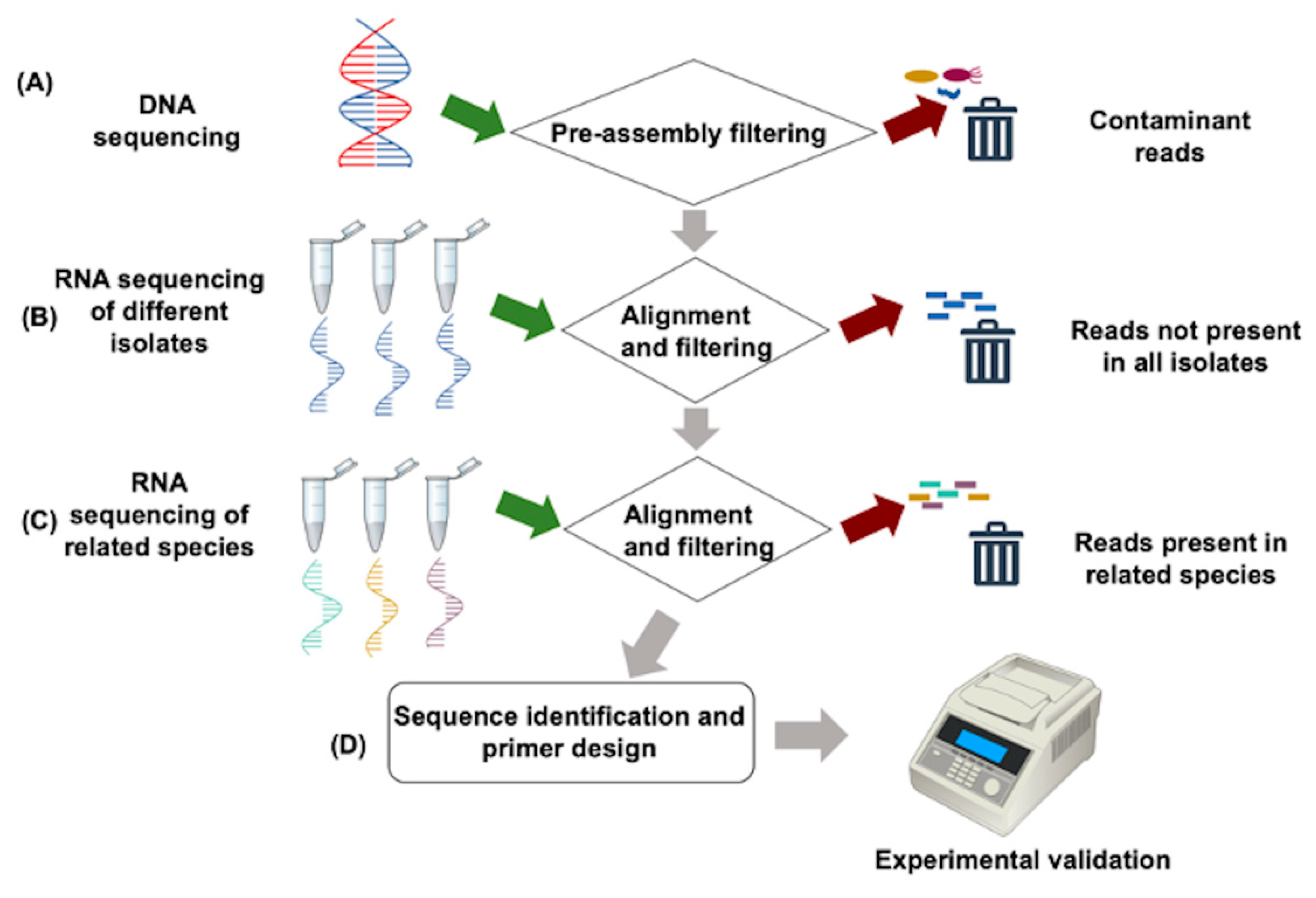
:1. Downy Mildew Pathogens and How to Find Them
2. Advantages and Disadvantages of Different Marker Types

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Loci/Type | Advantages | Disadvantages | Examples |
|---|---|---|---|---|
| Nuclear | Ribosomal and Internal Transcribed Spacer (ITS) | -High reproducibility -Abundant copies -Common primers | -Copy heterogeneity -Low resolution for cryptic species -Species cross-reactivity | Pe destructor [70] Pe arborescens [71] Ps belbahrii [72] Plasmopara spp. [64] B.lactucae [64] Ps humuli [73] |
| Housekeeping | -Common primers -known genes | -Low polymorphisms and reproducibility -Limited for phylogenetic analysis | Ps belbahrii [26] Ps cubensis [74] | |
| Species-specific | -species-specific primers | -Low polymorphisms and reproducibility -Limited for phylogenetic analysis | Ps humuli [23] Ps cubensis [32,33] | |
| Multilocus | -Improves phylogenetic interpretation -Infraspecific resolution -High variability | -Low reproducibility -More labor | Ps cubensis [66] Ps. humuli [66]. | |
| Mitochondrial | Single locus | -Improves phylogenetic interpretation -Infraspecific resolution -High variability | -Uniparental heritance -Limited in detecting hybrid species | B. lactucae [75] Ps cubensis [76] |
| Antigen | ELISA | -Speed and simplicity -High-throughput | -Requires monoclonal antibodies -Species cross-reactivity -Limited use for biosurveillance | Pe destructor [54] |
| Immunostrips | -Speed and simplicity -Cost-effective -Portability | -Requires monoclonal antibody -Species cross-reactivity | Still not available for downy mildew pathogens | |
| Enzymatic profile | Isozymes | -Codominant markers -Complement to phenotypic data | -Large amount of tissue is required -Low polymorphisms -Influenced by the environment | Pernosclerospora spp. [58] P. halstedii [59] |
3. Advances in Genomics Provide New Opportunities and Challenges for Diagnostic Marker Development
4. Beyond Pathogen Detection: Diagnostics for Pathogen-Informed Management
| Active Ingredient | FRAC | Target Site | Resistance in Oomycetes | Molecular Mechanisms Known? | Downy Mildews Where Molecular Mechanisms Have Been Identified |
|---|---|---|---|---|---|
| Mefenoxam | 4 | RNA polymerase I | Yes [127] | In Phytophthora infestans but not in a downy mildew pathogen [128,129,130]. | N/A |
| Famoxadone and fenamidone | 11 | Complex III: cytochrome bc1 (ubiquinol oxidase) at Qo site (cyt b gene) | Yes, [33,120] | Yes, G143A amino acid change in cyt b gene | Pl. viticola Ps. cubensis |
| Cyazofamid | 21 | Complex III: cytochrome bc1 (ubiquinone reductase) at Qi site | Yes [126] | Yes, in Pl. viticola an insertion in cyt B gene (i.e., E203-DE-V204) | Pl. viticola |
| Ethaboxam and zoxamide | 22 | β-tubulin | Yes [123] | Yes, C239S amino acid change in β-tubulin gene | Pl. viticola |
| Cymoxanil | 27 | Unknown | Yes [132] | Unknown | N/A |
| Propamocarb | 28 | Cell membrane permeability, fatty acids (proposed) | Yes [131] | Unknown | N/A |
| Fluazinam | 29 | Uncouplers of oxidative phosphorylation | Yes [133] | Unknown | N/A |
| Dimethomorph and mandipropamid | 40 | Cellulose synthase | Yes [33,124] | Yes, G1105W/V/S amino acid change in cellulose synthase gene (cesA3) | Pl. viticola Ps. cubensis |
| Fluopicolide | 43 | Delocalization of spectrin-like proteins | Yes [131] | Unknown | N/A |
| Ametoctradin | 45 | Complex III: cytochrome bc1 (ubiquinone reductase) at Qo site, stigmatellin-binding subsite | Yes [134] | Yes, S34L amino acid change in Pl. viticola in the cyt B gene | Pl. viticola |
| Oxathiapiprolin and fluoxapiprolin | 49 | Lipid homeostasis and transfer/storage | Yes [125] | Yes, G769W amino acid change in PcORP1 gene in Phytophthora capsici | N/A |
| Picarbutrazox | U17 | Unknown | Not officially reported | Not found | N/A |
5. Lab and Field-Deployable Platforms for Downy Mildew Diagnostics and Early Detection
6. Future Prospects: Portable Sequencing for Diagnostics and Biosurveillance
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gascuel, Q.; Martinez, Y.; Boniface, M.-C.; Vear, F.; Pichon, M.; Godiard, L. The sunflower downy. mildew pathogen Plasmopara halstedii: Plasmopara halstedii, sunflower downy mildew. Mol. Plant Pathol. 2015, 16, 109–122. [Google Scholar] [CrossRef]
- Salgado-Salazar, C.; Shiskoff, N.; Daughtrey, M.; Palmer, C.L.; Crouch, J.A. Downy Mildew: A serious disease threat to rose health worldwide. Plant Dis. 2018, 102, 1873–1882. [Google Scholar] [CrossRef] [Green Version]
- Wallace, E.C.; Quesada-Ocampo, L.M. First report of Downy Mildew on Buffalo Gourd (Cucurbita foetidissima) caused by Pseudoperonospora cubensis in North Carolina. Plant Dis. 2015, 99, 1861. [Google Scholar] [CrossRef]
- Wallace, E.C.; Adams, M.; Ivors, K.; Ojiambo, P.S.; Quesada-Ocampo, L.M. First report of Pseudoperonospora cubensis causing Downy Mildew on Momordica balsamina and M. charantia in North Carolina. Plant Dis. 2014, 98, 1279. [Google Scholar] [CrossRef]
- Wallace, E.; Choi, Y.J.; Thines, M.; Quesada-Ocampo, L.M. First Report of Plasmopara aff. australis on Luffa cylindrica in the United States. Plant Dis. 2015, 100, 537. [Google Scholar] [CrossRef]
- Dussert, Y.; Mazet, I.D.; Couture, C.; Gouzy, J.; Piron, M.-C.; Kuchly, C.; Bouchez, O.; Rispe, C.; Mestre, P.; Delmotte, F. A High-Quality Grapevine Downy Mildew genome assembly reveals rapidly evolving and lineage-specific putative host adaptation genes. GBE 2019, 11, 954–969. [Google Scholar] [CrossRef]
- Savory, E.A.; Granke, L.L.; Quesada-Ocampo, L.M.; Varbanova, M.; Hausbeck, M.K.; Day, B. The cucurbit downy mildew pathogen Pseudoperonospora cubensis. Mol. Plant Pathol. 2011, 12, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Purayannur, S.; Miles, T.D.; Gent, D.H.; Pigg, S.; Quesada-Ocampo, L.M. Hop Downy Mildew Caused by Pseudoperonospora humuli: A diagnostic guide. Plant Health Prog. 2020, 19, 173–179. [Google Scholar] [CrossRef]
- Coley-Smith, J.R. Persistence and identification of downy mildew Pseudoperonospora humuli (Miy. and Tak.) Wilson in hop rootstocks. Ann. Appl. Biol. 1964, 53, 129–132. [Google Scholar] [CrossRef]
- Wyenandt, C.A.; Simon, J.E.; Pyne, R.M.; Homa, K.; McGrath, M.T.; Zhang, S.; Raid, R.N.; Ma, L.-J.; Wick, R.; Guo, L.; et al. Basil Downy Mildew (Peronospora belbahrii): Discoveries and challenges relative to its control. Phytopathology 2015, 105, 885–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado-Salazar, C.; Rivera, Y.; Veltri, D.; Crouch, J.A. Polymorphic SSR markers for Plasmopara obducens (Peronosporaceae), the newly emergent downy mildew pathogen of Impatiens (Balsaminaceae). APPS 2015, 3, 1500073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derevnina, L.; Chin-Wo-Reyes, S.; Martin, F.; Wood, K.; Froenicke, L.; Spring, O.; Michelmore, R. Genome sequence and architecture of the tobacco downy mildew pathogen Peronospora tabacina. MPMI 2015, 28, 1198–1215. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-J.; Hong, S.-B.; Shin, H.-D. Re-consideration of Peronospora farinosa infecting Spinacia oleracea as distinct species, Peronospora effusa. Mycolo. Res. 2007, 111, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.C.; De Leon, C.; Payak, M.M. Diseases of maize in South and South-East Asia: Problems and progress. Crop. Prot. 1993, 12, 414–422. [Google Scholar] [CrossRef]
- Beans, C. Scientists Are Fighting for the Stricken Pickle against This Tricky Disease [Radio Broadcast]. NPR Radio. 14 December 2018. Available online: www.npr.org/sections/thesalt/2018/12/14/672229450/672229450/scientists-are-fighting-for-the-stricken-against-this-tricky-disease (accessed on 18 February 2021).
- Edwardson, J.R. Hops—Their botany, history, production and utilization. Econ. Bot. 1952, 6, 160–175. [Google Scholar] [CrossRef]
- Holmes, G.J.; Ojiambo, P.S.; Hausbeck, M.K.; Quesada-Ocampo, L.M.; Keinath, A.P. Resurgence of cucurbit downy mildew in the United States: A watershed event for research and extension. Plant Dis. 2015, 99, 428–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frederiksen, R.A.; Renfro, B.L. Global status of maize downy mildew. Annu. Rev. Phytopathol. 1977, 15, 249–271. [Google Scholar] [CrossRef]
- Chen, W.-J.; Delmotte, F.; Cervera, S.R.; Douence, L.; Greif, C.; Corio-Costet, M.-F. At least two origins of fungicide resistance in grapevine downy mildew populations. Appl. Environ. Microbiol. 2007, 73, 5162–5172. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.E.; Eastwell, K.C.; Grove, G.G.; Barbour, J.D.; Ocamb, C.M.; Alldredge, J.R. Sensitivity of Pseudoperonospora humuli (the causal agent of hop downy mildew) from Oregon, Idaho, and Washington to fosetyl-Al (Aliette). Plant Health Prog. 2004, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Pintore, I.; Gilardi, G.; Gullino, M.L.; Garibaldi, A. Detection of mefenoxam-resistant strains of Peronospora belbahrii, the causal agent of basil downy mildew, transmitted through infected seeds. Phytoparasitica 2016, 44, 563–569. [Google Scholar] [CrossRef]
- Keinath, A.P.; Miller, S.A.; Smart, C.D. Response of Pseudoperonospora cubensis to preventative fungicide applications varies by state and year. Plant Health Prog. 2019, 20, 142–146. [Google Scholar] [CrossRef]
- Rahman, A.; Góngora-Castillo, E.; Bowman, M.J.; Childs, K.L.; Gent, D.H.; Martin, F.N.; Quesada-Ocampo, L.M. Genome sequencing and transcriptome analysis of the hop downy mildew pathogen Pseudoperonospora humuli reveal species-specific genes for molecular detection. Phytopathology 2019, 109, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Withers, S.; Gongora-Castillo, E.; Gent, D.; Thomas, A.; Ojiambo, P.S.; Quesada-Ocampo, L.M. Using next-generation sequencing to develop molecular diagnostics for Pseudoperonospora cubensis, the cucurbit downy mildew pathogen. Phytopathology 2016, 106, 1105–1116. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Meneses, M.; Ristaino, J.B. Detection and quantification of Peronospora tabacina using a real-time polymerase chain reaction assay. Plant Dis. 2011, 95, 673–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, D.; Tian, M. A qPCR approach to quantify the growth of basil downy mildew pathogen Peronospora belbahrii during infection. Curr. Plant Biol. 2018, 15, 2–7. [Google Scholar] [CrossRef]
- Salcedo, A.; Hausbeck, M.; Pigg, S.; Quesada-Ocampo, L.M. Diagnostic guide for cucurbit downy mildew. Plant Health Prog. 2020, 19, 166–172. [Google Scholar] [CrossRef]
- Kandel, S.L.; Mou, B.; Shishkoff, N.; Shi, A.; Subbarao, K.V.; Klosterman, S.J. Spinach downy mildew: Advances in our understanding of the disease cycle and prospects for disease management. Plant Dis. 2019, 103, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Y.; Van den Langenberg, K.M.; Wehner, T.C.; Ojiambo, P.S.; Hausbeck, M.K.; Quesada-Ocampo, L.M.; Lebeda, A.; Sierotzki, H.; Gisi, U. Resurgence of Pseudoperonospora cubensis: The causal agent of cucurbit downy mildew. Phytopathology 2015, 105, 998–1012. [Google Scholar] [CrossRef] [Green Version]
- Standish, J.R.; Raid, R.N.; Pigg, S.; Quesada-Ocampo, L.M. A Diagnostic guide for basil downy mildew. Plant Health Prog. 2020, 21, 77–81. [Google Scholar] [CrossRef]
- Wallace, E.C.; D’Arcangelo, K.N.; Quesada-Ocampo, L.M. Population analyses reveal two host-adapted clades of Pseudoperonospora cubensis, the causal agent of cucurbit downy mildew, on commercial and wild cucurbits. Phytopathology 2020, 110, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Standish, J.; D’Arcangelo, K.N.; Quesada-Ocampo, L.M. Clade-specific biosurveillance of Pseudoperonospora cubensis using spore traps for precision disease management of cucurbit downy mildew. Phytopathology 2020, 6, 231. [Google Scholar] [CrossRef]
- Rahman, A.; Miles, T.D.; Martin, F.N.; Quesada-Ocampo, L.M. Molecular approaches for biosurveillance of the cucurbit downy mildew pathogen, Pseudoperonospora cubensis. Can. J. Plant Pathol. 2017, 39, 282–296. [Google Scholar] [CrossRef]
- Crandall, S.G.; Rahman, A.; Quesada-Ocampo, L.M.; Martin, F.N.; Bilodeau, G.J.; Miles, T.D. Advances in diagnostics of downy mildews: Lessons learned from other oomycetes and future challenges. Plant Dis. 2018, 102, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, F.; Scalenghe, R.; Davino, S.; Panno, S.; Scuderi, G.; Ruisi, P.; Villa, P.; Stroppiana, D.; Boschetti, M.; Goulart, L.R.; et al. Advanced methods of plant disease detection. A review. Agron. Sustain. Dev. 2015, 35, 1–25. [Google Scholar] [CrossRef] [Green Version]
- The Royal Society. Reaping the Benefits Science and the Sustainable Intensification of Global Agriculture; The Royal Society: London, UK, 2009. [Google Scholar]
- Klein, J. Genome reconstruction of the non-culturable spinach downy mildew Peronospora effusa by metagenome filtering. PLoS ONE 2020, 15, e0225808. [Google Scholar] [CrossRef]
- Aegerter, B.J.; Nuñez, J.J.; Davis, R.M. Environmental factors affecting rose downy mildew and development of a forecasting model for a nursery production system. Plant Dis. 2003, 87, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Granke, L.L.; Morrice, J.J.; Hausbeck, M.K. Relationships between airborne Pseudoperonospora cubensis sporangia, environmental conditions, and cucumber downy mildew severity. Plant Dis. 2014, 98, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, R.A.; Koike, S.T.; Fox, A.D.; Anchieta, A.; Subbarao, K.V.; Klosterman, S.J.; McRoberts, N. Season-long dynamics of spinach downy mildew determined by spore trapping and disease incidence. Phytopathology 2016, 106, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.A.; Beed, F.D.; Harmon, C.L. Plant disease diagnostic capabilities and networks. Annu. Rev. Phytopathol. 2009, 47, 15–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Paul, R.; Ba Tis, T.; Saville, A.C.; Hansel, J.C.; Yu, T.; Ristaino, J.B.; Wei, Q. Non-invasive plant disease diagnostics enabled by smartphone-based fingerprinting of leaf volatiles. Nat. Plants 2019, 5, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. PCR heads into the field. Nat. Methods 2015, 12, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Qin, W.; Huang, X.; Kong, F.; Schoen, C.D.; Feng, J.; Wang, Z.; Zhang, H. Development and application of loop-mediated isothermal amplification (LAMP) for detection of Plasmopara viticola. Sci. Rep. 2016, 6, 28935. [Google Scholar] [CrossRef] [PubMed]
- Riethmuller, A.; Voglmayr, H.; Goker, M.; Weiss, M.; Oberwinkler, F. Phylogenetic relationships of the downy mildews (Peronosporales) and related groups based on nuclear large subunit ribosomal DNA sequences. Mycologia 2002, 94, 834. [Google Scholar] [CrossRef] [PubMed]
- Goulter, K.; Randles, J. Serological and molecular techniques to detect and identify plant pathogens. In Plant Pathogens and Plant Diseases; Brown, J.F., Ogle, H.J., Eds.; Rockvale Publications: Armidale, Australia, 1997; pp. 172–191. [Google Scholar]
- Aslam, S.; Tahir, A.; Aslam, M.F.; Alam, M.W.; Shedayi, A.A.; Sadia, S. Recent advances in molecular techniques for the identification of phytopathogenic fungi–a mini review. J. Plant Interact. 2017, 12, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Torrance, L. Use of monoclonal antibodies in plant pathology. Eur. J. Plant Pathol. 1995, 101, 351–363. [Google Scholar] [CrossRef]
- Peruski, A.H.; Peruski, L.F. Immunological methods for detection and identification of infectious disease and biological warfare agents. CVI 2003, 10, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Narayanasamy, P. Microbial Plant Pathogens-Detection and Disease Diagnosis; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-90-481-9734-7. [Google Scholar]
- Martin, F.N.; Abad, Z.G.; Balci, Y.; Ivors, K. Identification and detection of Phytophthora: Reviewing our progress, identifying our needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, K.; Kobayashi, M.; Tomita, M.; Kubota, N.; Suga, H.; Hyakumachi, M. Production and evaluation of monoclonal antibodies for the detection of Pythium sulcatum in Soil. J. Phytopathol. 2002, 150, 97–104. [Google Scholar] [CrossRef]
- Rittenburg, J.; Petersen, F.; Miller, S.; Grothaus, G. Development of a rapid, field-usable immunoassay format, for detection and quantitation of Pythium, Rhizoctonia and Sclerotinia spp. in plant tissue. Phytopathology 1988, 78, 1519. [Google Scholar]
- Kennedy, R.; Wakeham, A.J. Development of detection systems for the sporangia of Peronospora destructor. In The Downy Mildews-Genetics, Molecular Biology and Control; Lebeda, A., Spencer-Phillips, P.T.N., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 147–155. [Google Scholar]
- Mahaffee, W.; Grove, G.; Stoll, R. Inoculum detection for managing grape powdery mildew. In Practical Winery and Vineyards; Springer: Dordrecht, The Netherlands, 2011; pp. 26–32. [Google Scholar]
- Estrada-Garcia, M.T.; Callow, J.A.; Green, J.R. Monoclonal antibodies to the adhesive cell coat secreted by Pythium aphanidermatum zoospores recognise 200 × 103 Mr glycoproteins stored within large peripheral vesicles. J. Cell Sci. 1990, 95, 199. [Google Scholar]
- Hardham, A.R.; Suzaki, E.; Perkin, J.L. Monoclonal antibodies to isolate-, species-, and genus-specific components on the surface of zoospores and cysts of the fungus Phytophthora cinnamomi. Can. J. Bot. 1986, 64, 311–321. [Google Scholar] [CrossRef]
- Bonde, M.R. Isozyme Analysis to differentiate species of Peronosclerospora causing downy mildews of maize. Phytopathology 1984, 74, 1278. [Google Scholar] [CrossRef]
- Komjáti, H.; Bakonyi, J.; Spring, O.; Virányi, F. Isozyme analysis of Plasmopara halstedii using cellulose-acetate gel electrophoresis. Plant Pathol. 2007, 57, 57–63. [Google Scholar] [CrossRef]
- Blair, J.E.; Coffey, M.D.; Park, S.-Y.; Geiser, D.M.; Kang, S. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genet. Biol. 2008, 45, 266–277. [Google Scholar] [CrossRef]
- Schroeder, K.L.; Martin, F.N.; de Cock, A.W.A.M.; Lévesque, C.A.; Spies, C.F.J.; Okubara, P.A.; Paulitz, T.C. Molecular detection and quantification of Pythium species: Evolving taxonomy, new tools, and challenges. Plant Dis. 2013, 97, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Wallace, E.C.; Quesada-Ocampo, L.M. Analysis of microsatellites from the transcriptome of downy mildew pathogens and their application for characterization of Pseudoperonospora populations. Peer J. 2017, 5, e3266. [Google Scholar] [CrossRef] [Green Version]
- Perumal, R.; Nimmakayala, P.; Erattaimuthu, S.R.; No, E.-G.; Reddy, U.K.; Prom, L.K.; Odvody, G.N.; Luster, D.G.; Magill, C.W. Simple sequence repeat markers useful for sorghum downy mildew (Peronosclerospora sorghi) and related species. BMC Genet. 2008, 9, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voglmayr, H.; Riethmüller, A.; Göker, M.; Weiss, M.; Oberwinkler, F. Phylogenetic relationships of Plasmopara, Bremia and other genera of downy mildew pathogens with pyriform haustoria based on Bayesian analysis of partial LSU rDNA sequence data. Mycol. Res. 2004, 108, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Thines, M.; Voglmayr, H.; Gker, M. Taxonomy and phylogeny of the downy mildews (Peronosporaceae). In Oomycete Genetics and Genomics; Lamour, K., Kamoun, S., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 47–75. [Google Scholar]
- Runge, F.; Choi, Y.-J.; Thines, M. Phylogenetic investigations in the genus Pseudoperonospora reveal overlooked species and cryptic diversity in the P. cubensis species cluster. Eur. J. Plant Pathol. 2011, 129, 135–146. [Google Scholar] [CrossRef]
- Yang, X.; Hong, C. Differential usefulness of nine commonly used genetic markers for identifying Phytophthora species. Front. Microbiol. 2018, 9, 2334. [Google Scholar] [CrossRef] [Green Version]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in-silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Malapi-Wight, M.; Salgado-Salazar, C.; Demers, J.E.; Clement, D.L.; Rane, K.K.; Crouch, J.A. Sarcococca blight: Use of whole-genome sequencing for fungal plant disease diagnosis. Plant Dis. 2016, 100, 1093–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heyden, H.; Bilodeau, G.; Carisse, O.; Charron, J.-B. Monitoring of Peronospora destructor primary and secondary inoculum by real-time qPCR. Plant Dis. 2020, 104, 3183–3191. [Google Scholar] [CrossRef]
- Montes-Borrego, M.; Muñoz-Ledesma, F.J.; Jiménez-Díaz, R.M.; Landa, B.B. Real-time PCR quantification of Peronospora arborescens, the opium poppy downy mildew pathogen, in seed stocks and symptomless infected plants. Plant Dis. 2011, 95, 143–152. [Google Scholar] [CrossRef]
- Farahani-Kofoet, R.D.; Römer, P.; Grosch, R. Systemic spread of downy mildew in basil plants and detection of the pathogen in seed and plant samples. Mycol. Prog. 2012, 11, 961–966. [Google Scholar] [CrossRef]
- Gent, D.H.; Nelson, M.E.; Farnsworth, J.L.; Grove, G.G. PCR detection of Pseudoperonospora humuli in air samples from hop yards. Plant Pathol. 2009, 58, 1081–1091. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, M.H.; Lee, S. Identification of Pseudoperonospora cubensis using real-time PCR and high- resolution melting (HRM) analysis. J. Gen. Plant Pathol. 2016, 82, 110–115. [Google Scholar] [CrossRef]
- Kunjeti, S.G.; Anchieta, A.; Martin, F.N.; Choi, Y.-J.; Thines, M.; Michelmore, R.W.; Koike, S.T.; Tsuchida, C.; Mahaffee, W.; Subbarao, K.V.; et al. Detection and quantification of Bremia lactucae by spore trapping and quantitative PCR. Phytopathology 2016, 106, 1426–1437. [Google Scholar] [CrossRef] [Green Version]
- Summers, C.F.; Adair, N.L.; Gent, D.H.; McGrath, M.T.; Smart, C.D. Pseudoperonospora cubensis and P. humuli detection using species-specific probes and high-definition melt curve analysis. Can. J. Plant Pathol. 2015, 37, 315–330. [Google Scholar] [CrossRef]
- Lindner, D.L.; Banik, M.T. Intragenomic variation in the ITS rDNA region obscures phylogenetic relationships and inflates estimates of operational taxonomic units in genus Laetiporus. Mycologia 2011, 103, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Thines, M.; Choi, Y.-J. Evolution, Diversity, and taxonomy of the Peronosporaceae, with focus on the Genus Peronospora. Phytopathology 2016, 106, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A Molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Anchieta, A.; McRoberts, N.; Koike, S.T.; Subbarao, K.V.; Voglmayr, H.; Choi, Y.-J.; Thines, M.; Martin, F.N. Coupling spore traps and quantitative PCR assays for detection of the downy mildew pathogens of spinach (Peronospora effusa) and Beet (P. schachtii). Phytopathology 2014, 104, 1349–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, M.N.; Ocamb, C.M.; Grünwald, N.J.; Mancino, L.E.; Gent, D.H. Genetic and pathogenic relatedness of Pseudoperonospora cubensis and P. humuli. Phytopathology 2011, 101, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, G.J.; Martin, F.N.; Coffey, M.D.; Blomquist, C.L. Development of a multiplex assay for genus- and species-specific detection of Phytophthora based on differences in mitochondrial gene order. Phytopathology 2014, 104, 733–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, M.B.; Walsh, K.; Boonham, N.; O’Neill, T.; Pearson, S.; Barker, I. Development of real-time PCR (TaqMan®) assays for the detection and quantification of Botrytis cinerea in planta. Plant Physiol. Biochem. 2005, 43, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Badotti, F.; de Oliveira, F.S.; Garcia, C.F.; Vaz, A.B.M.; Fonseca, P.L.C.; Nahum, L.A.; Oliveira, G.; Góes-Neto, A. Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol. 2017, 17, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosdidier, M.; Aguayo, J.; Marçais, B.; Ioos, R. Detection of plant pathogens using real-time PCR: How reliable are late Ct values? Plant Pathol. 2017, 66, 359–367. [Google Scholar] [CrossRef]
- Kulik, T.; Bilska, K.; Żelechowski, M. Promising perspectives for detection, identification, and quantification of plant pathogenic fungi and oomycetes through targeting mitochondrial DNA. IJMS 2020, 21, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-J.; Beakes, G.; Glockling, S.; Kruse, J.; Nam, B.; Nigrelli, L.; Ploch, S.; Shin, H.-D.; Shivas, R.G.; Telle, S.; et al. Towards a universal barcode of oomycetes-a comparison of the cox 1 and cox 2 loci. Mol. Ecol. Resour. 2015, 15, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Mansfield, M.A.; Park, B.; Geiser, D.M.; Ivors, K.L.; Coffey, M.D.; Grünwald, N.J.; Martin, F.N.; Lévesque, C.A.; Blair, J.E. The Promise and pitfalls of sequence-based identification of plant-pathogenic fungi and oomycetes. Phytopathology 2010, 100, 732–737. [Google Scholar] [CrossRef]
- Galtier, N.; Nabholz, B.; Glémin, S.; Hurst, G.D.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Nuclear DNA analyses in genetic studies of populations: Practice, problems and prospects. Mol. Ecol. 2003, 12, 563–584. [Google Scholar] [CrossRef]
- Robideau, G.P.; De Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef]
- Thines, M.; Sharma, R.; Rodenburg, S.Y.A.; Gogleva, A.; Judelson, H.S.; Xia, X.; van den Hoogen, J.; Kitner, M.; Klein, J.; Neilen, M.; et al. The genome of Peronospora belbahrii reveals high heterozygosity, a low number of canonical effectors, and TC-rich promoters. MPMI 2020, 33, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Straub, S.C.; Fishbein, M.; Livshultz, T.; Foster, Z.; Parks, M.; Weitemier, K.; Cronn, R.C.; Liston, A. Building a model: Developing genomic resources for common milkweed (Asclepias syriaca) with low coverage genome sequencing. BMC Genom. 2011, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Kroon, L.P.N.M.; Bakker, F.T.; van den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Villa, N.O.; Kageyama, K.; Asano, T.; Suga, H. Phylogenetic relationships of Pythium and Phytophthora species based on ITS rDNA, cytochrome oxidase II and β-tubulin gene sequences. Mycologia 2006, 98, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, K.; Klosterman, S.J.; Derevnina, L.; Martin, F.; Bertier, L.D.; Koike, S.; Reyes-Chin-Wo, S.; Mou, B.; Michelmore, R. Comparative genomics of downy mildews reveals potential adaptations to biotrophy. BMC Genom. 2018, 19, 851. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Xia, X.; Cano, L.M.; Evangelisti, E.; Kemen, E.; Judelson, H.; Oome, S.; Sambles, C.; van den Hoogen, D.J.; Kitner, M.; et al. Genome analyses of the sunflower pathogen Plasmopara halstedii provide insights into effector evolution in downy mildews and Phytophthora. BMC Genom. 2015, 16, 741. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, K.; Gil, J.; Bertier, L.D.; Kenefick, A.; Wood, K.J.; Zhang, L.; Reyes-Chin-Wo, S.; Cavanaugh, K.; Tsuchida, C.; Wong, J.; et al. Genomic signatures of heterokaryosis in the oomycete pathogen Bremia Lact. Nat. Commun. 2019, 10, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouxel, M.; Papura, D.; Nogueira, M.; Machefer, V.; Dezette, D.; Richard-Cervera, S.; Carrere, S.; Mestre, P.; Delmotte, F. Microsatellite markers for characterization of native and introduced populations of Plasmopara viticola, the causal agent of grapevine downy mildew. Appl. Environ. Microbiol. 2012, 78, 6337–6340. [Google Scholar] [CrossRef] [Green Version]
- Thilliez, G.J.A.; Armstrong, M.R.; Lim, T.-Y.; Baker, K.; Jouet, A.; Ward, B.; Oosterhout, C.; van Jones, J.D.G.; Huitema, E.; Birch, P.R.J.; et al. Pathogen enrichment sequencing (PenSeq) enables population genomic studies in oomycetes. New Phytol. 2019, 221, 1634–1648. [Google Scholar] [CrossRef] [Green Version]
- Jouet, A.; Saunders, D.G.O.; McMullan, M.; Ward, B.; Furzer, O.; Jupe, F.; Cevik, V.; Hein, I.; Thilliez, G.J.A.; Holub, E.; et al. Albugo candida race diversity, ploidy and host-associated microbes revealed using DNA sequence capture on diseased plants in the field. New Phytol. 2019, 221, 1529–1543. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Jones, M.; Koutsovoulos, G.; Clarke, M.; Blaxter, M. Blobology: Exploring raw genome data for contaminants, symbionts and parasites using taxon-annotated GC-coverage plots. Front. Genet. 2013, 4, 237. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S. Metabolic Diversity and novelties in the oomycetes. Annu. Rev. Microbiol. 2017, 71, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Ah-Fong, A.M.V.; Kagda, M.S.; Abrahamian, M.; Judelson, H.S. Niche-specific metabolic adaptation in biotrophic and necrotrophic oomycetes is manifested in differential use of nutrients, variation in gene content, and enzyme evolution. PLoS Pathog. 2019, 15, e1007729. [Google Scholar] [CrossRef]
- Feng, C.; Correll, J.C.; Kammeijer, K.E.; Koike, S.T. Identification of new races and deviating strains of the spinach downy mildew pathogen Peronospora farinosa f. sp. spinaciae. Plant Dis. 2014, 98, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.E.; Inaba, T.; Cohen, Y. Physiological specialization in Pseudoperonospora cubensis. Phytopathology 1987, 77, 1621–1624. [Google Scholar] [CrossRef]
- Mohammed, A.E.; You, M.P.; Al-lami, H.F.D.; Barbetti, M.J. Pathotypes and phylogenetic variation determine downy mildew epidemics in Brassica spp. in Australia. Plant Pathol. 2018, 67, 1514–1527. [Google Scholar] [CrossRef]
- Radwan, G.L.; Perumal, R.; Isakeit, T.; Magill, C.W.; Prom, L.K.; Little, C.R. Screening exotic sorghum germplasm, hybrids, and elite lines for resistance to a new virulent pathotype (P6) of Peronosclerospora sorghi causing downy mildew. Plant Health Prog. 2011, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Thakur, R.P.; Rai, K.N.; Rao, V.P.; Rao, A.S. Genetic resistance of pearl millet male-sterile lines to diverse Indian pathotypes of Sclerospora graminicola. Plant Dis. 2001, 85, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Jeuken, M.; Lindhout, P. Lactuca saligna, a non-host for lettuce downy mildew (Bremia lactucae), harbors a new race-specific Dm gene and three QTLs for resistance. Theor. Appl. Genet. 2002, 105, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Call, A.D.; Criswell, A.D.; Wehner, T.C.; Ando, K.; Grumet, R. Resistance of cucumber cultivars to a new strain of cucurbit downy mildew. Horts 2012, 47, 171–178. [Google Scholar] [CrossRef]
- Gascuel, Q.; Bordat, A.; Sallet, E.; Pouilly, N.; Carrere, S.; Roux, F.; Vincourt, P.; Godiard, L. Effector polymorphisms of the sunflower downy mildew pathogen Plasmopara halstedii and their use to identify pathotypes from field isolates. PLoS ONE 2016, 11, e0148513. [Google Scholar] [CrossRef] [PubMed]
- Pecrix, Y.; Buendia, L.; Penouilh-Suzette, C.; Maréchaux, M.; Legrand, L.; Bouchez, O.; Rengel, D.; Gouzy, J.; Cottret, L.; Vear, F.; et al. Sunflower resistance to multiple downy mildew pathotypes revealed by recognition of conserved effectors of the oomycete Plasmopara halstedii. Plant J. 2019, 97, 730–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purayannur, S.; Cano, L.M.; Bowman, M.J.; Childs, K.L.; Gent, D.H.; Quesada-Ocampo, L.M. The effector repertoire of the hop downy mildew pathogen Pseudoperonospora humuli. Front. Genet. 2020, 11, 910. [Google Scholar] [CrossRef] [PubMed]
- Purayannur, S.; Cano, L.M.; Bowman, M.J.; Childs, K.L.; Quesada-Ocampo, L.M. Host-specific effectors of the cucurbit downy mildew pathogen Pseudoperonospora cubensis. Phytopathology 2020, 110, S1.1–S1.26. [Google Scholar] [CrossRef]
- Jupe, F.; Witek, K.; Verweij, W.; Śliwka, J.; Pritchard, L.; Etherington, G.J.; Maclean, D.; Cock, P.J.; Leggett, R.M.; Bryan, G.J.; et al. Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 2013, 76, 530–544. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Cano, L.M.; Raffaele, S.; Bain, R.A.; Cooke, L.R.; Etherington, G.J.; Deahl, K.L.; Farrer, R.A.; Gilroy, E.M.; Goss, E.M.; et al. Genome analyses of an aggressive and invasive lineage of the irish potato famine pathogen. PLoS Pathog. 2012, 8, e1002940. [Google Scholar] [CrossRef]
- Goldenhar, K.E.; Hausbeck, M.K. Fungicides for control of downy mildew on pickling cucumber in Michigan. Plant Health Prog. 2019, 20, 165–169. [Google Scholar] [CrossRef]
- D’Arcangelo, K.N.; Adams, M.L.; Kerns, J.P.; Quesada-Ocampo, L.M. Assessment of fungicide product applications and program approaches for control of downy mildew on pickling cucumber in North Carolina. Crop. Protect. 2021, 140, 105412. [Google Scholar] [CrossRef]
- Ishii, H.; Fraaije, B.A.; Sugiyama, T.; Noguchi, K.; Nishimura, K.; Takeda, T.; Amano, T.; Hollomon, D.W. Occurrence and Molecular Characterization of Strobilurin Resistance in cucumber powdery mildew and downy mildew. Phytopathology 2001, 91, 1166–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, M.; Waldner, M.; Olaya, G.; Cohen, Y.; Gisi, U.; Sierotzki, H. Resistance mechanism to carboxylic acid amide fungicides in the cucurbit downy mildew pathogen Pseudoperonospora cubensis: Resistance mechanism to CAA fungicides in Pseudoperonospora cubensis. Pest. Manag. Sci. 2011, 67, 1211–1214. [Google Scholar] [CrossRef]
- Feng, X.; Baudoin, A. First report of carboxylic acid amide fungicide resistance in Plasmopara viticola (Grapevine Downy Mildew) in North America. Plant Health Prog. 2018, 19, 139. [Google Scholar] [CrossRef]
- Noel, Z.A.; Sang, H.; Roth, M.G.; Chilvers, M.I. Convergent Evolution of C239S Mutation in Pythium spp. β-tubulin coincides with inherent insensitivity to ethaboxam and implications for other Peronosporalean oomycetes. Phytopathology 2019, 109, 2087–2095. [Google Scholar] [CrossRef]
- Blum, M.; Gamper, H.A.; Waldner, M.; Sierotzki, H.; Gisi, U. The cellulose synthase 3 (CesA3) gene of oomycetes: Structure, phylogeny and influence on sensitivity to carboxylic acid amide (CAA) fungicides. Fungal Biol. 2012, 116, 529–542. [Google Scholar] [CrossRef]
- Miao, J.; Cai, M.; Dong, X.; Liu, L.; Lin, D.; Zhang, C.; Pang, Z.; Liu, X. Resistance assessment for oxathiapiprolin in Phytophthora capsici and the detection of a point mutation (G769W) in PcORP1 that confers resistance. Front. Microbiol. 2016, 7, 615. [Google Scholar] [CrossRef] [Green Version]
- Cherrad, S.; Hernandes, C.; Steva, H.; Vacher, S. Resistance de Plasmopara viticola aux inhibiteurs du complexe III: Un point sur la caracterisation phenotypique et genotypique des souches. Plant Dis. 2018, 7, 11–12. [Google Scholar]
- Reuveni, M. Development of Resistance to metalaxyl in Pseudoperonospora cubensis. Plant Dis. 1980, 64, 1108. [Google Scholar] [CrossRef]
- Matson, M.E.H.; Small, I.M.; Fry, W.E.; Judelson, H.S. Metalaxyl resistance in Phytophthora infestans: Assessing role of RPA190 gene and diversity within clonal lineages. Phytopathology 2015, 105, 1594–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, E.; Young, V.; Sierotzki, H.; Scalliet, G.; Birch, P.R.J.; Cooke, D.E.L.; Csukai, M.; Whisson, S.C. Sequence diversity in the large subunit of RNA polymerase I contributes to Mefenoxam insensitivity in Phytophthora infestans: Basis of Mefenoxam insensitivity in P. infestans. Mol. Plant Pathol. 2014, 15, 664–676. [Google Scholar] [CrossRef]
- Ayala-Usma, D.A.; Danies, G.; Myers, K.; Bond, M.O.; Romero-Navarro, J.A.; Judelson, H.S.; Restrepo, S.; Fry, W.E. Genome-wide association study identifies single nucleotide polymorphism markers associated with mycelial growth (at 15, 20, and 25 °C), Mefenoxam resistance, and mating type in Phytophthora infestans. Phytopathology 2020, 110, 822–833. [Google Scholar] [CrossRef]
- Thomas, A.; Neufeld, K.N.; Seebold, K.W.; Braun, C.A.; Schwarz, M.R.; Ojiambo, P.S. Resistance to Fluopicolide and Propamocarb and baseline sensitivity to Ethaboxam among isolates of Pseudoperonospora cubensis from the Eastern United States. Plant Dis. 2018, 102, 1619–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toffolatti, S.L.; Venturini, G.; Campia, P.; Cirio, L.; Bellotto, D.; Vercesi, A. Sensitivity to cymoxanil in Italian populations of Plasmopara viticola oospores: Sensitivity to cymoxanil in Italian populations of Plasmopara viticola oospores. Pest. Manag. Sci. 2015, 71, 1182–1188. [Google Scholar] [CrossRef]
- Schepers, H.T.A.M.; Kessel, G.J.T.; Lucca, F.; Förch, M.G.; van den Bosch, G.B.M.; Topper, C.G.; Evenhuis, A. Reduced efficacy of fluazinam against Phytophthora infestans in the Netherlands. Eur. J. Plant Pathol. 2018, 151, 947–960. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, S.; Remuson, F.; Caddoux, L.; Barrès, B. Investigation of the sensitivity of Plasmopara viticola to amisulbrom and ametoctradin in French vineyards using bioassays and molecular tools. Pest Manag. Sci. 2019, 75, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Miles, T.D.; Neill, T.; Colle, M.; Warneke, B.; Robinson, G.; Stergiopoulos, I.; Mahaffee, W.F. Allele-specific detection methods for QoI fungicide resistant Erysiphe necator in vineyards. Plant Dis. 2020, 105, 175–182. [Google Scholar] [CrossRef]
- Broccanello, C.; Chiodi, C.; Funk, A.; McGrath, J.M.; Panella, L.; Stevanato, P. Comparison of three PCR-based assays for SNP genotyping in plants. Plant Methods 2018, 14, 28. [Google Scholar] [CrossRef] [Green Version]
- West, J.S.; Atkins, S.D.; Emberlin, J.; Fitt, B.D.L. PCR to predict risk of airborne disease. Trends Microbiol. 2008, 16, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Leach, C.M.; Hildebrand, P.D.; Sutton, J.C. Sporangium discharge by Peronospora destructor: Influence of humidity, red-infrared radiation, and vibration. Phytopathology 1982, 72, 1052–1056. [Google Scholar] [CrossRef]
- Aylor, D.E.; Taylor, G.S. Escape of Peronospora tabacina spores from a field of diseased tobacco plants. Phytopathology 1983, 73, 525–529. [Google Scholar] [CrossRef]
- Granke, L.L.; Hausbeck, M.K. Dynamics of Pseudoperonospora cubensis Sporangia in commercial cucurbit fields in Michigan. Plant Dis. 2011, 95, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, K.N.; Isard, S.A.; Ojiambo, P.S. Relationship between disease severity and escape of Pseudoperonospora cubensis sporangia from a cucumber canopy during downy mildew epidemics. Plant Pathol. 2013, 62, 1366–1377. [Google Scholar] [CrossRef]
- Thiessen, L.D.; Keune, J.A.; Neill, T.M.; Turechek, W.W.; Grove, G.G.; Mahaffee, W.F. Development of a grower-conducted inoculum detection assay for management of grape powdery mildew. Plant Pathol. 2016, 65, 238–249. [Google Scholar] [CrossRef]
- Dhar, N.; Mamo, B.E.; Subbarao, K.V.; Koike, S.T.; Fox, A.; Anchieta, A.; Klosterman, S.J. Measurements of aerial spore load by qPCR facilitates lettuce downy mildew risk advisement. Plant Dis. 2020, 104, 82–93. [Google Scholar] [CrossRef]
- Belbahri, L.; Calmin, G.; Pawlowski, J.; Lefort, F. Phylogenetic analysis and real-time PCR detection of a presumably undescribed Peronospora species on sweet basil and sage. Mycolog. Res. 2005, 109, 1276–1287. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, A.; Minuto, G.; Bertetti, D.; Gullino, M.L. Seed transmission of Peronospora sp. of Basil/Samenübertragung von Peronospora sp. an Basilicum. J. Plant Dis. Prot. 2004, 111, 465–469. [Google Scholar]
- Landa, B.B.; Montes-Borrego, M.; Muñoz-Ledesma, F.J.; Jiménez-Díaz, R.M. Phylogenetic analysis of downy mildew pathogens of opium poppy and PCR-Based in planta and seed detection of Peronospora arborescens. Phytopathology 2007, 97, 1380–1390. [Google Scholar] [CrossRef] [Green Version]
- Villari, C.; Mahaffee, W.F.; Mitchell, T.K.; Pedley, K.F.; Pieck, M.L.; Hand, F.P. Early detection of airborne inoculum of Magnaporthe oryzae in turfgrass fields using a quantitative LAMP assay. Plant Dis. 2017, 101, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Si Ammour, M.; Bilodeau, G.J.; Tremblay, D.M.; Van der Heyden, H.; Yaseen, T.; Varvaro, L.; Carisse, O. Development of real-time isothermal amplification assays for on-site detection of Phytophthora infestans in potato leaves. Plant Dis. 2017, 101, 1269–1277. [Google Scholar] [CrossRef] [Green Version]
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase polymerase amplification for diagnostic applications. Clin. Chem. 2016, 62, 947–958. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Kubota, R.; Alvarez, A.; Su, W.; Jenkins, D. FRET-based assimilating probe for sequence-specific real-time monitoring of loop-mediated isothermal amplification (LAMP). Biol. Eng. Trans. 2011, 4, 81–100. [Google Scholar] [CrossRef]
- Oscorbin, I.P.; Belousova, E.A.; Zakabunin, A.I.; Boyarskikh, U.A.; Filipenko, M.L. Comparison of fluorescent intercalating dyes for quantitative loop-mediated isothermal amplification (qLAMP). Biotechniques 2016, 61, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Mohd-Naim, N.F.; Safavieh, M.; Ahmed, M.U. Colorimetric nucleic acid detection on paper microchip using loop mediated isothermal amplification and crystal violet dye. ACS Sens. 2017, 2, 1713–1720. [Google Scholar] [CrossRef]
- Ma, X.; Shu, Y.; Nie, K.; Qin, M.; Wang, D.; Gao, R.; Wang, M.; Wen, L.; Han, F.; Zhou, S. Visual detection of pandemic influenza A H1N1 Virus 2009 by reverse-transcription loop-mediated isothermal amplification with hydroxynaphthol blue dye. J. Virol. Methods 2010, 167, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Corless, C.E.; Guiver, M.; Borrow, R.; Edwards-Jones, V.; Kaczmarski, E.B.; Fox, A.J. Contamination and sensitivity issues with a real-time universal 16S rRNA PCR. J. Clin. Microbiol. 2000, 38, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Thiessen, L.D.; Neill, T.M.; Mahaffee, W.F. Development of a quantitative loop-mediated isothermal amplification assay for the field detection of Erysiphe necator. Peer J. 2018, 6, e4639. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, H.; Kawana, T.; Fukushima, E.; Suzutania, T. Tolerance of loop-mediated isothermal amplification to a culture medium and biological substances. J. Biochem. Biophys. Methods 2007, 70, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Francois, P.; Tangomo, M.; Hibbs, J.; Bonetti, E.-J.; Boehme, C.C.; Notomi, T.; Perkins, M.D.; Schrenzel, J. Robustness of a loop-mediated isothermal amplification reaction for diagnostic applications. FEMS Immunol. Med. Microbiol. 2011, 62, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Nixon, G.; Garson, J.A.; Grant, P.; Nastouli, E.; Foy, C.A.; Huggett, J.F. Comparative study of sensitivity, linearity, and resistance to inhibition of digital and nondigital polymerase chain reaction and loop mediated isothermal amplification assays for quantification of human cytomegalovirus. Anal. Chem. 2014, 86, 4387–4394. [Google Scholar] [CrossRef]
- Nguyen, P.L.; Sudheesh, P.S.; Thomas, A.C.; Sinnesael, M.; Haman, K.; Cain, K.D. Rapid detection and monitoring of Flavobacterium psychrophilum in water by using a handheld, field-portable quantitative PCR system. J. Aquat. Anim. Health 2018, 30, 302–311. [Google Scholar] [CrossRef]
- Stagg, H.E.B.; Merino, R.; Oladimeji, P.; Steiropoulos, N.; Munro, E.S. Preliminary validation of a portable real-time RT-PCR assay for the detection of salmonid alphavirus. Bull. Eur. Assoc. Fish. Pathol. 2018, 38, 121–128. [Google Scholar]
- DeShields, J.B.; Bomberger, R.A.; Woodhall, J.W.; Wheeler, D.L.; Moroz, N.; Johnson, D.A.; Tanaka, K. On-site molecular detection of soil-borne phytopathogens using a portable real-time PCR system. J. Vis. Exp. 2018, 132, e56891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiessen, L.D.; Neill, T.M.; Mahaffee, W.F. Assessment of Erysiphe necator ascospore release models for use in the mediterranean climate of Western Oregon. Plant Dis. 2018, 102, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Miles, T.D.; Martin, F.N.; Coffey, M.D. Development of rapid isothermal amplification assays for detection of Phytophthora spp. in Plant Tissue. Phytopathology 2015, 105, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Rojas, J.A.; Miles, T.D.; Coffey, M.D.; Martin, F.N.; Chilvers, M.I. Development and application of qPCR and RPA genus- and species-specific detection of Phytophthora sojae and P. sansomeana root rot pathogens of soybean. Plant Dis. 2017, 101, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.S.; Zaikova, E.; Goerlitz, D.S.; Bai, Y.; Tighe, S.W. Real-time DNA sequencing in the antarctic dry valleys using the Oxford nanopore sequencer. J. Biomol. Tech. 2017, 28, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Castro-Wallace, S.L.; Chiu, C.Y.; John, K.K.; Stahl, S.E.; Rubins, K.H.; McIntyre, A.B.R.; Dworkin, J.P.; Lupisella, M.L.; Smith, D.J.; Botkin, D.J.; et al. Nanopore DNA sequencing and genome assembly on the international space station. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalupowicz, L.; Dombrovsky, A.; Gaba, V.; Luria, N.; Reuven, M.; Beerman, A.; Lachman, O.; Dror, O.; Nissan, G.; Manulis-Sasson, S. Diagnosis of plant diseases using the Nanopore sequencing platform. Plant Pathol. 2019, 68, 229–238. [Google Scholar] [CrossRef]
- Badial, A.B.; Sherman, D.; Stone, A.; Gopakumar, A.; Wilson, V.; Schneider, W.; King, J. Nanopore sequencing as a surveillance tool for plant pathogens in plant and insect tissues. Plant Dis. 2018, 102, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, G.V.; Cook, N.M.; Bueno-Sancho, V.; Lewis, C.M.; Persoons, A.; Mitiku, A.D.; Heaton, M.; Davey, P.E.; Abeyo, B.; Alemayehu, Y.; et al. MARPLE, a point-of-care, strain-level disease diagnostics and surveillance tool for complex fungal pathogens. BMC Biol. 2019, 17, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boykin, L.M.; Sseruwagi, P.; Alicai, T.; Ateka, E.; Mohammed, I.U.; Stanton, J.-A.L.; Kayuki, C.; Mark, D.; Fute, T.; Erasto, J.; et al. Tree Lab: Portable genomics for early detection of plant viruses and pests in Sub-Saharan Africa. Genes 2019, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Vangah, S.J.; Katalani, C.; Booneh, H.A.; Hajizade, A.; Sijercic, A.; Ahmadian, G. CRISPR-based diagnosis of infectious and noninfectious diseases. Biol. Proced. Online 2020, 22, 22. [Google Scholar] [CrossRef]
- Wang, X.; Shang, X.; Xingxu, H.X. Next-generation pathogen diagnosis with CRISPR/Cas-based detection methods. Emergy Microbes Infect. 2020, 9, 1682–1691. [Google Scholar] [CrossRef]
- Dai, T.; Yang, X.; Hu, T.; Jiao, B.; Xu, Y.; Zheng, X.; Shen, D. Comparative evaluation of a novel recombinase polymerase amplification-lateral flow dipstick (RPA-LFD) assay, LAMP, conventional PCR, and leaf-disc baiting methods for detection of Phytophthora sojae. Front. Microbiol. 2019, 10, 1884. [Google Scholar] [CrossRef] [Green Version]
- Crannell, Z.A.; Rohrman, B.; Richards-Kortum, R. Equipment-free incubation of recombinase polymerase amplification reactions using body heat. PLoS ONE 2014, 9, e112146. [Google Scholar] [CrossRef]
- Xiang, X.; Qian, K.; Zhang, Z.; Lin, F.; Xie, Y.; Liu, Y.; Yang, Z. CRISPR-cas systems based molecular diagnostic tool for infectious diseases and emerging 2019 novel coronavirus (COVID-19) pneumonia. J. Drug Target. 2020, 28, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.; Collins, J.J.; Zhang, F. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salcedo, A.F.; Purayannur, S.; Standish, J.R.; Miles, T.; Thiessen, L.; Quesada-Ocampo, L.M. Fantastic Downy Mildew Pathogens and How to Find Them: Advances in Detection and Diagnostics. Plants 2021, 10, 435. https://doi.org/10.3390/plants10030435
Salcedo AF, Purayannur S, Standish JR, Miles T, Thiessen L, Quesada-Ocampo LM. Fantastic Downy Mildew Pathogens and How to Find Them: Advances in Detection and Diagnostics. Plants. 2021; 10(3):435. https://doi.org/10.3390/plants10030435
Chicago/Turabian StyleSalcedo, Andres F., Savithri Purayannur, Jeffrey R. Standish, Timothy Miles, Lindsey Thiessen, and Lina M. Quesada-Ocampo. 2021. "Fantastic Downy Mildew Pathogens and How to Find Them: Advances in Detection and Diagnostics" Plants 10, no. 3: 435. https://doi.org/10.3390/plants10030435