
Taxonomic Revisiting and Phylogenetic Placement of Two Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae)

 ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Scanning Electron Microscope (SEM)
2.1.1. Stem Micromorphology
2.1.2. Leaf Epidermal Cells
2.1.3. Stomatal Complex
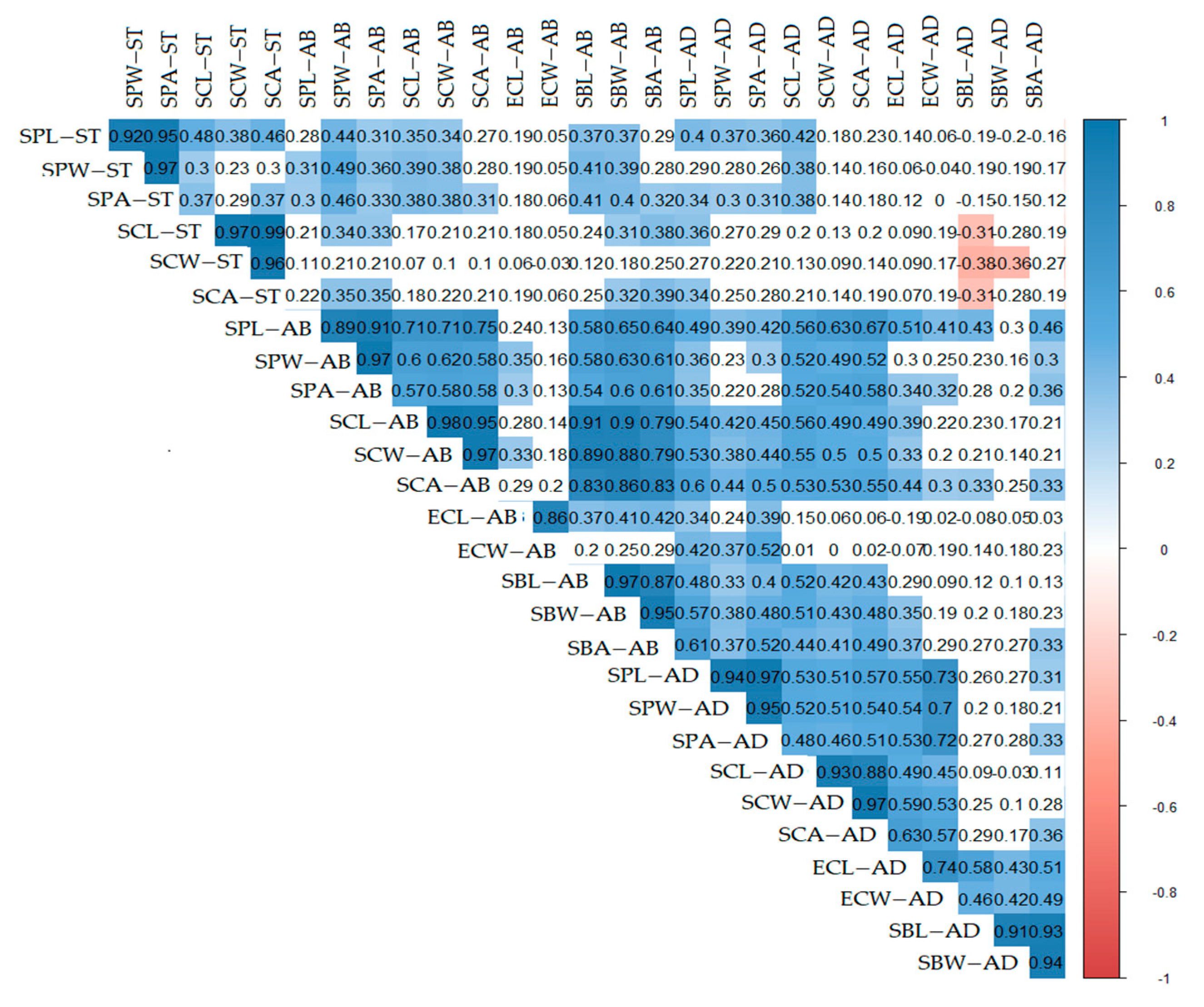
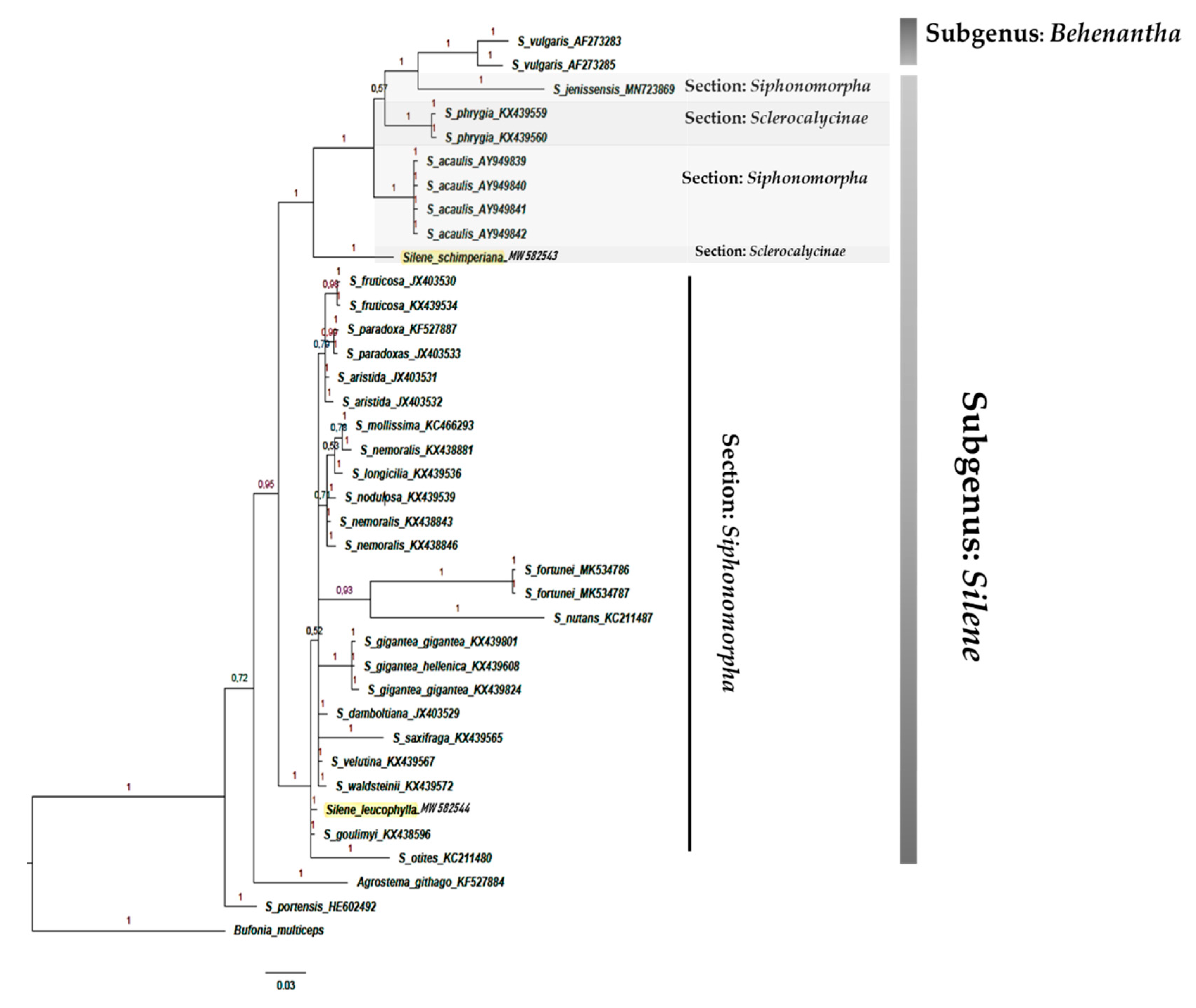
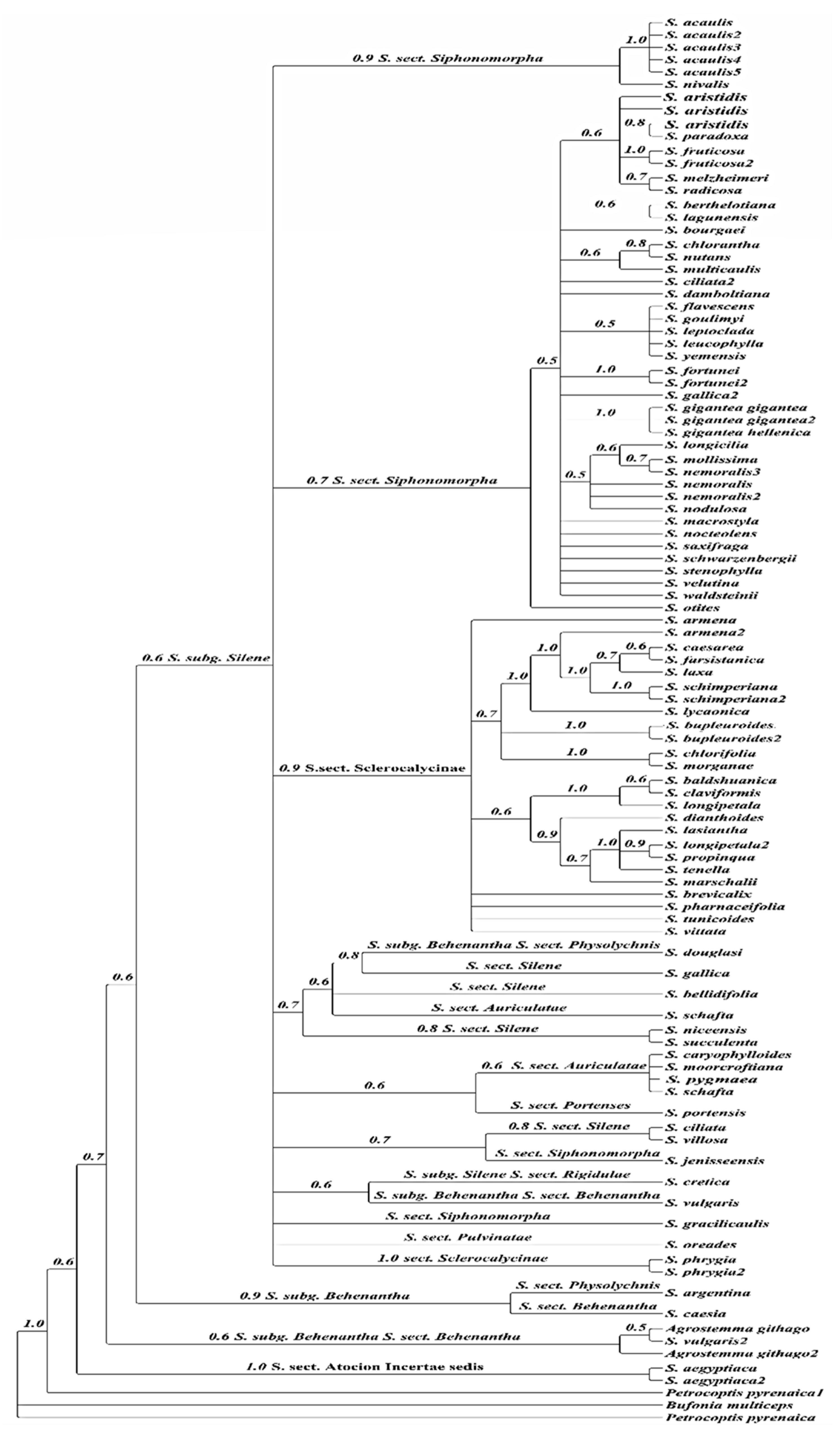
2.2. Phylogeny
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Specimens Examined
4.3. Scanning Electron Microscopy (SEM)
4.4. Statistical Analysis
4.5. DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Greenberg, A.K.; Donoghue, M.J. Molecular systematics and character evolution in Caryophyllaceae. Taxon 2011, 60, 1637–1652. [Google Scholar] [CrossRef]
- Rabeler, R.K.; Hartman, R.L. Caryophyllaceae in North America: Change over the last 110 years. J. Bot. Res. Inst. Tex. 2008, 2, 1285–1289. [Google Scholar]
- Madhani, H.; Rabeler, R.; Pirani, A.; Oxelman, B.; Heubl, G.; Zarre, S. Untangling phylogenetic patterns and taxonomic confusion in tribe Caryophylleae (Caryophyllaceae) with special focus on generic boundaries. Taxon 2018, 67, 83–112. [Google Scholar] [CrossRef] [Green Version]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabeler, R.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Hernández-Ledesma, P.; Berendsohn, W.G.; Borsch, T.; Von Mering, S.; Akhani, H.; Arias, S.; Castañeda-Noa, I.; Eggli, U.; Eriksson, R.; Flores-Olvera, H. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales. Willdenowia 2015, 45, 281–383. [Google Scholar] [CrossRef] [Green Version]
- Rohrbach, P. Conspectus systematicus specierum generis Silene Ann. Sci. Nat. Bot. 1868, 5, 369–382. [Google Scholar]
- Faried, A.; El-Banhawy, A.; Elqahtani, M. Taxonomic, DNA barcoding and phylogenetic reassessment of the Egyptian Ephedra L. (Ephedraceae). Catrina Int. J. Environ. Sci. 2018, 17, 1–13. [Google Scholar]
- El-Banhawy, A.; Al-Juhani, W. DNA barcoding and phylogeny of Phlomis aurea (Lamiaceae) endemic to Sinai Peninsula, Egypt. Pak. J. Bot. 2019, 51, 1263–1271. [Google Scholar] [CrossRef]
- Oxelman, B.; Rautenberg, A.; Thollesson, M.; Larsson, A.; Frajman, B.; Eggens, F.; Petri, A.; Aydin, Z.; Töpel, M.; Brandtberg-Falkman, A. Sileneae Taxonomy and Systematics. Available online: http://www.sileneae.info (accessed on 25 February 2021).
- Hosny, A.; El Hadidi, M.; Shamso, E. Taxonomic studies of Silenoideae (Cartophyllaceae) in Egypt. 1. Systematic revision of the genus Silene L. Taeckholmia 1992, 14, 1–36. [Google Scholar]
- Chowdhuri, P. Studies in the genus Silene. Notes R. Bot. Gard. Edinb. 1957, 22, 221–278. [Google Scholar]
- Royal Botanic Gardens Kew. World Checklist of Selected Plant Families (WCSP); Royal Botanic Gardens Kew: London, UK, 2018. [Google Scholar]
- Fakhry, A.M.; El-Keblawy, A.; Shabana, H.A.; Gamal, I.E.; Shalouf, A. Microhabitats Affect Population Size and Plant Vigor of Three Critically Endangered Endemic Plants in Southern Sinai Mountains, Egypt. Land 2019, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Omar, K. Community–Based Conservation of Threatened Plants Silene schimperiana and Polygala sinaica in South Sinai, Egypt. Available online: www.rufford.org (accessed on 15 February 2021).
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. J. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- El-Ghamery, A.; Morsy, M.; Abu-Shahba, M.J.T. Chromosome count and karyotype study on endemic species Silene schimperiana Boiss.(Caryophyllaceae), Sinai, Egypt. Taeckholmia 2016, 36, 102–114. [Google Scholar] [CrossRef]
- Zarrinkamar, F. Foliar anatomy of the Caryophyllaceae family in Arasbaran, NW Iran. Iran. J. Bot. 2001, 9, 93–102. [Google Scholar]
- Ahmad, K.; Khan, M.A.; Ahmad, M.; Zafar, M.; Arshad, M.; Ahmad, F. Taxonomic diversity of stomata in dicot flora of a district tank (N.W.F.P.) in Pakistan. Afr. J. Biotechnol. 2009, 8, 1052–1055. [Google Scholar]
- Hosseini, E.; Ghahremaninejad, F.; Assadi, M. The indumentum of vegetative and reproductive parts of annual species of Silene (Caryophyllaceae) in Iran. Prog. Biol. Sci. 2016, 6, 107–116. [Google Scholar]
- Sheibani, M.; Pirani, A. Trichome micromorphology of Silene (Caryophyllaceae) species in Iran. Iran. J. Bot. 2018, 24, 28–45. [Google Scholar]
- Chandra, S.; Rawat, D.; Verma, S.R.; Uniyal, P. Trichome diversity of the family Caryophyllaceae from Western Himalaya and their taxonomic implication. Trop. Plant Res. 2019, 6, 397–407. [Google Scholar] [CrossRef]
- Ullah, F.; Zafar, M.; Ahmad, M.; Shah, S.N.; Razzaq, A.; Sohail, A.; Zaman, W.; Çelik, A.; Ayaz, A.; Sultana, S. A systematic approach to the investigation of foliar epidermal anatomy of subfamily Caryophylloideae (Caryophyllaceae). Flora 2018, 246, 61–70. [Google Scholar] [CrossRef]
- Stace, C.A. Cuticular Studies as an Aid to Plant Taxonomy; British Museum (Natural History): London, UK, 1965. [Google Scholar]
- Casson, S.; Gray, J.E. Influence of environmental factors on stomatal development. New Phytol. 2008, 178, 9–23. [Google Scholar] [CrossRef]
- Dina, A.F.; Ibrahim, M.F.; Farag, R.; El-Gawad, H.G.A.; El-Banhawy, A.; Alaraidh, I.A.; Rashad, Y.M.; Lashin, I.; El-Yazied, A.A.; Elkelish, A.J.P. Influence of Polyethylene Glycol on Leaf Anatomy, Stomatal Behavior, Water Loss, and Some Physiological Traits of Date Palm Plantlets Grown In Vitro and Ex Vitro. Plants 2020, 9, 1440. [Google Scholar] [CrossRef] [PubMed]
- Hoveka, L.N.; van der Bank, M.; Bezeng, B.S.; Davies, T.J. Identifying biodiversity knowledge gaps for conserving South Africa’s endemic flora. Biodivers. Conserv. 2020, 29, 2803–2819. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant Endemism Centres and Biodiversity Hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Martín-Gómez, J.J.; Rewicz, A.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. Seed Morphology in Silene Based on Geometric Models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Parkhurst, D.F. The adaptive significance of stomatal occurrence on one or both surfaces of leaves. J. Ecol. 1978, 66, 367–383. [Google Scholar] [CrossRef]
- Takahashi, S.; Monda, K.; Negi, J.; Konishi, F.; Ishikawa, S.; Hashimoto-Sugimoto, M.; Goto, N.; Iba, K. Natural variation in stomatal responses to environmental changes among Arabidopsis thaliana ecotypes. PLoS ONE 2015, 10, e0117449. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, G.; Tsukaya, H. Organ size regulation in plants: Insights from compensation. Front. Plant Sci. 2011, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Gázquez, A.; Beemster, G.T. What determines organ size differences between species? A meta-analysis of the cellular basis. New Phytol. 2017, 215, 299–308. [Google Scholar] [CrossRef] [Green Version]
- El-Banhawy, A.; Acedo, C.; Qari, S.; Elkordy, A. Molecular Identification and Phylogenetic Placement of Rosa arabica Crép.(Rosaceae), a Critically Endangered Plant Species. Life 2020, 10, 335. [Google Scholar] [CrossRef]
- Rossatto, D.R.; Kolb, R.M. Gochnatia polymorpha (Less.) Cabrera (Asteraceae) changes in leaf structure due to differences in light and edaphic conditions. Acta Bot. Bras. 2010, 24, 605–612. [Google Scholar] [CrossRef]
- Schoch, P.-G.; Zinsou, C.; Sibi, M.J. Dependence of the stomatal index on environmental factors during stomatal differentiation in leaves of Vigna sinensis L. 1. Effect of light intensity. J. Exp. Bot. 1980, 31, 1211–1216. [Google Scholar] [CrossRef]
- Naciri, Y.; Du Pasquier, P.-E.; Lundberg, M.; Jeanmonod, D.; Oxelman, B. A phylogenetic circumscription of Silene sect. Siphonomorpha (Caryophyllaceae) in the Mediterranean Basin. Taxon 2017, 66, 91–108. [Google Scholar]
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene L. (Caryophyllaceae) en Méditerranée occidentale. IV: Species caeterae. Candollea 1985, 40, 5–34. [Google Scholar]
- Boissier, P.E. Flora Orientalis, Sive, Enumeratio Plantarum in Oriente a Graecia et Aegypto ad Indiae Fines Hucusque Observatarum; Georg, H., Ed.; Basileae: Genevae, Switzerland, 1875; Volume 1. [Google Scholar]
- Boulos, L. Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 1999. [Google Scholar]
- Coode, M.J.E.; Cullen, J. Silene L. Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, Scotland, 1967; Volume 2, pp. 179–242. [Google Scholar]
- Deflers, A. Voyage au Yémen, Journal D’une Excursion Botanique Faite en 1887 Dans les Montagnes de l’Arabie Heureuse; Kincksieck, P., Ed.; Wentworth Press: Paris, France, 1889; p. 306. [Google Scholar]
- Greuter, W. Studies in Greek Caryophylloideae: Agrostemma, Silene, and Vaccaria; Willdenowia: Berlin, Germany, 1995; pp. 105–142. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Ferreira, E.B.; Cavalcanti, P.P.; Nogueira, D.A. ExpDes: An R package for ANOVA and experimental designs. J. Appl. Math. 2014, 5, 2952. [Google Scholar] [CrossRef] [Green Version]
- Kolde, R. Pheatmap: Pretty Heatmaps; Version 1.0.8; Vienna, Austria, 2015. [Google Scholar]
- Viscosi, V.; Cardini, A. Leaf morphology, taxonomy and geometric morphometrics: A simplified protocol for beginners. PLoS ONE 2011, 6, e25630. [Google Scholar] [CrossRef] [Green Version]
- Soetewey, A. Correlation Coefficient and Correlation Test in R. 2020. Available online: https://www.statsandr.com/blog/correlation-coefficient-and-correlation-test-in-r/ (accessed on 20 February 2021).
- Alexander, P.J.; Rajanikanth, G.; Bacon, C.D.; Bailey, C.D. Recovery of plant DNA using a reciprocating saw and silica-based columns. Mol. Ecol. Notes 2007, 7, 5–9. [Google Scholar] [CrossRef]
- Doyle, J. DNA protocols for plants. In Molecular Techniques in Taxonomy; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.R.; Zhang, Y.H.; Nakamura, K.; Guan, B.C.; Qiu, Y.X. Developing DNA barcodes for species identification in Podophylloideae (Berberidaceae). J. Syst. Evol. 2014, 52, 487–499. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery, Salt Lake City, UT, USA, 18–21 July 2011; pp. 1–8. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Silene leucophylla Boiss. | Silene schimperiana Boiss. |
|---|---|---|
| Stem characters | ||
| Surface | Pubescent | Glabrous |
| Epicuticular Wax | Thin layer | Irregular crustose platelets |
| Trichomes | Present | Absent |
| Trichomes Type | Unicellular, non-glandular | Absent |
| Trichome Surface | Densely pustulate | Absent |
| Trichomes Length | 45–98 µm | Absent |
| Trichomes Width | 13–23 µm | Absent |
| Stomata Type | Anomocytic | Anomocytic |
| Abaxial leaf (AB) characters | ||
| Epidermal Cell Shape | Irregular oblong to bone-shape | Parallel tetra- to polygonal |
| Anticlinal Walls | Sunken channeled irregularly curved | Sunken straight |
| The curvature of Outer Periclinal Walls | Convex | Slightly Convex |
| Fine Relief of the Cell Wall | Highly ribbed | Irregular epicuticular crustose platelets |
| Trichomes Type | Unicellular, non-glandular | Absent |
| Trichome Surface | Densely irregular pustulate | Absent |
| Trichomes Length | 78–130 µm | Absent |
| Trichomes Width | 18–20 µm | Absent |
| Stomata Type | Diacytic | Diacytic |
| Stomata Level | Raised | Sunken |
| Guard Cell Surface | Smooth | Irregular epicuticular crustose platelets |
| Pore Shape | Elliptic slit | Linear slit |
| Adaxial leaf (AD) characters | ||
| Epidermal Cell Shape | Irregular oblong to bone-shape | Parallel penta- to polygonal |
| Anticlinal Walls | Sunken channeled irregularly curved | Sunken straight |
| The curvature of Outer Periclinal Walls | Convex | Slightly Convex |
| Fine Relief of the Cell Wall | Moderately ribbed | Irregular epicuticular crustose platelets |
| Trichomes Type | Unicellular, non-glandular | Absent |
| Trichome Surface | Densely irregular pustulate | Absent |
| Trichomes Length | 75–125 µm | Absent |
| Trichomes Width | 16–25 µm | Absent |
| Stomata Type | Diacytic | Diacytic |
| Stomata Level | Raised | Sunken |
| Guard Cell Surface | Smooth | Irregular epicuticular crustose platelets |
| Pore Shape | Elliptic slit | Elliptic + Linear slits |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
EL-Banhawy, A.; Nour, I.H.; Acedo, C.; ElKordy, A.; Faried, A.; AL-Juhani, W.; Gawhari, A.M.H.; Olwey, A.O.; Ellmouni, F.Y. Taxonomic Revisiting and Phylogenetic Placement of Two Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Plants 2021, 10, 740. https://doi.org/10.3390/plants10040740
EL-Banhawy A, Nour IH, Acedo C, ElKordy A, Faried A, AL-Juhani W, Gawhari AMH, Olwey AO, Ellmouni FY. Taxonomic Revisiting and Phylogenetic Placement of Two Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Plants. 2021; 10(4):740. https://doi.org/10.3390/plants10040740
Chicago/Turabian StyleEL-Banhawy, Ahmed, Iman H. Nour, Carmen Acedo, Ahmed ElKordy, Ahmed Faried, Widad AL-Juhani, Ahmed M. H. Gawhari, Asmaa O. Olwey, and Faten Y. Ellmouni. 2021. "Taxonomic Revisiting and Phylogenetic Placement of Two Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae)" Plants 10, no. 4: 740. https://doi.org/10.3390/plants10040740