
Micromorphological Traits of Balcanic Micromeria and Closely Related Clinopodium Species (Lamiaceae)

 , , , ,
, , , , 
Abstract
:
1. Introduction
2. Results and Discussion
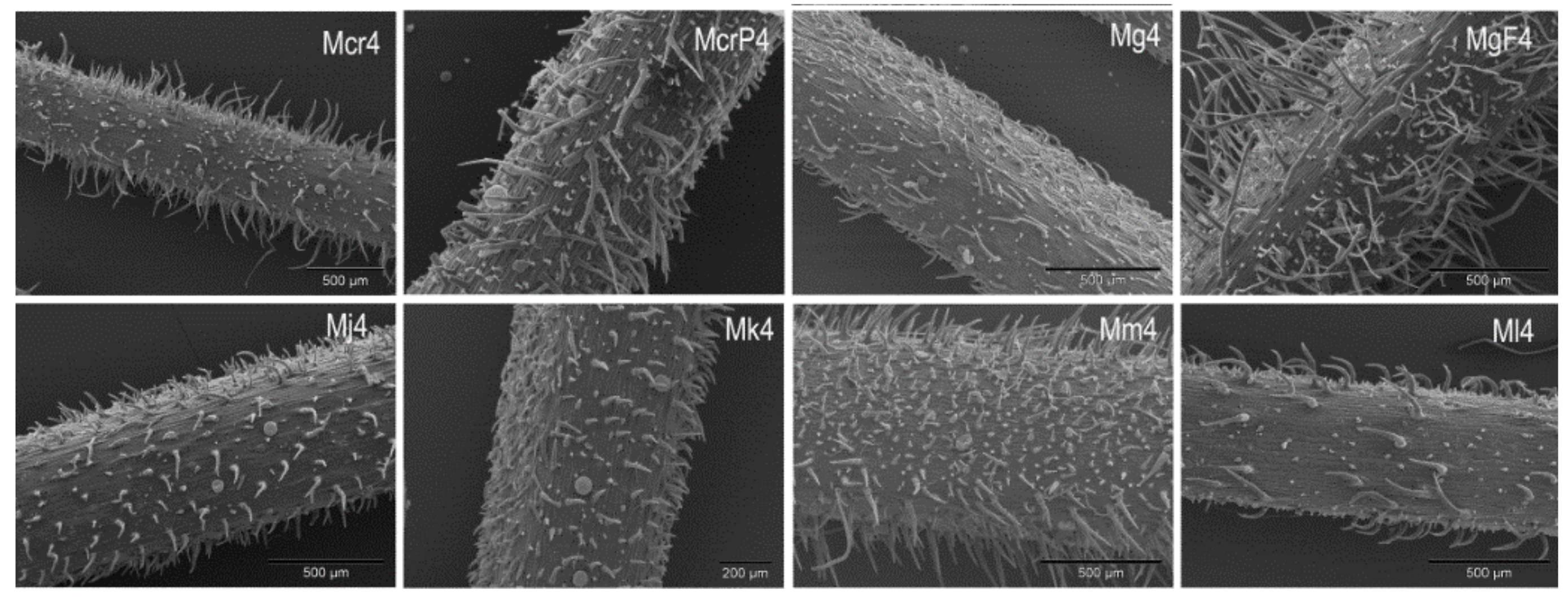
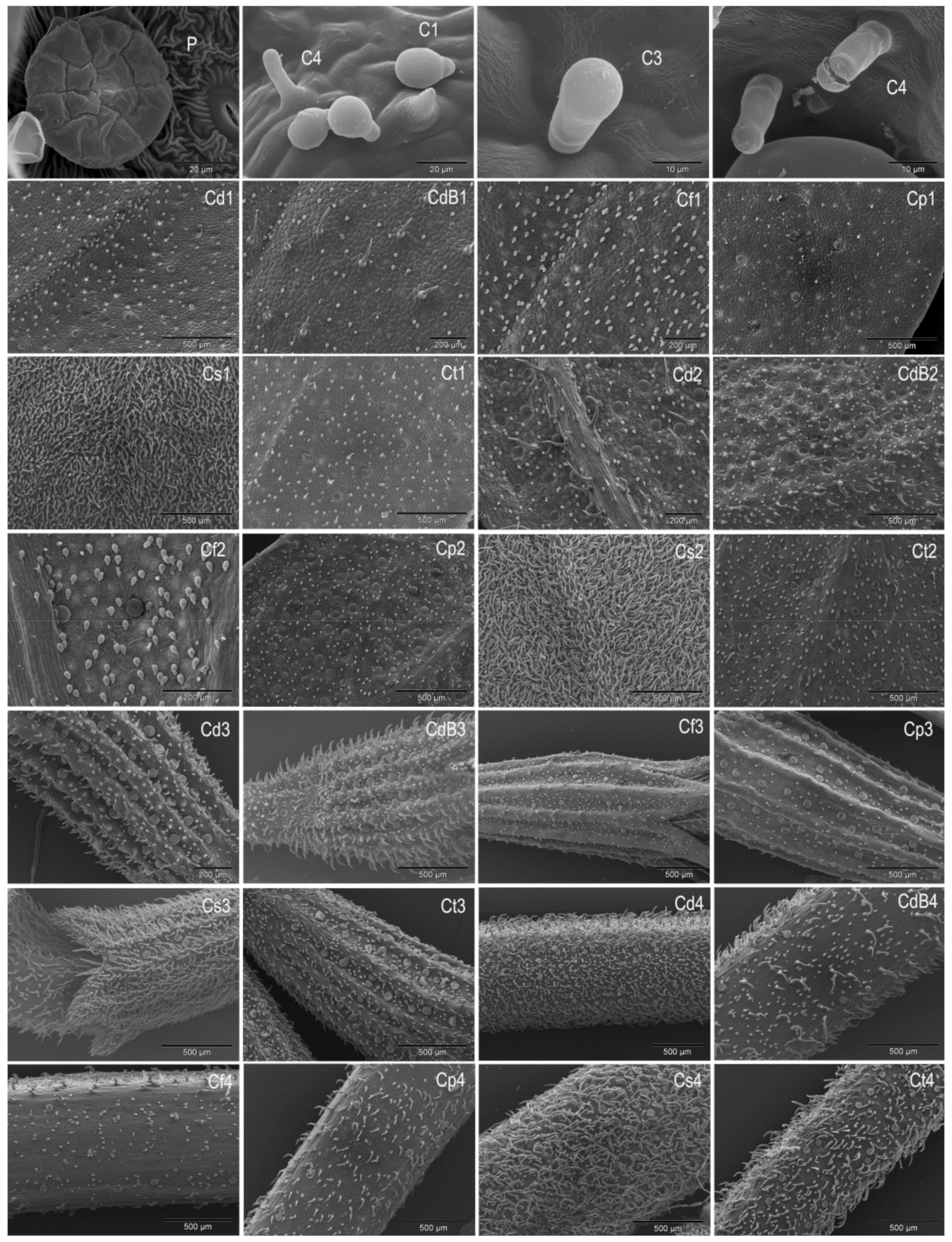
2.1. Trichomes
2.2. Pollen
3. Materials and Methods
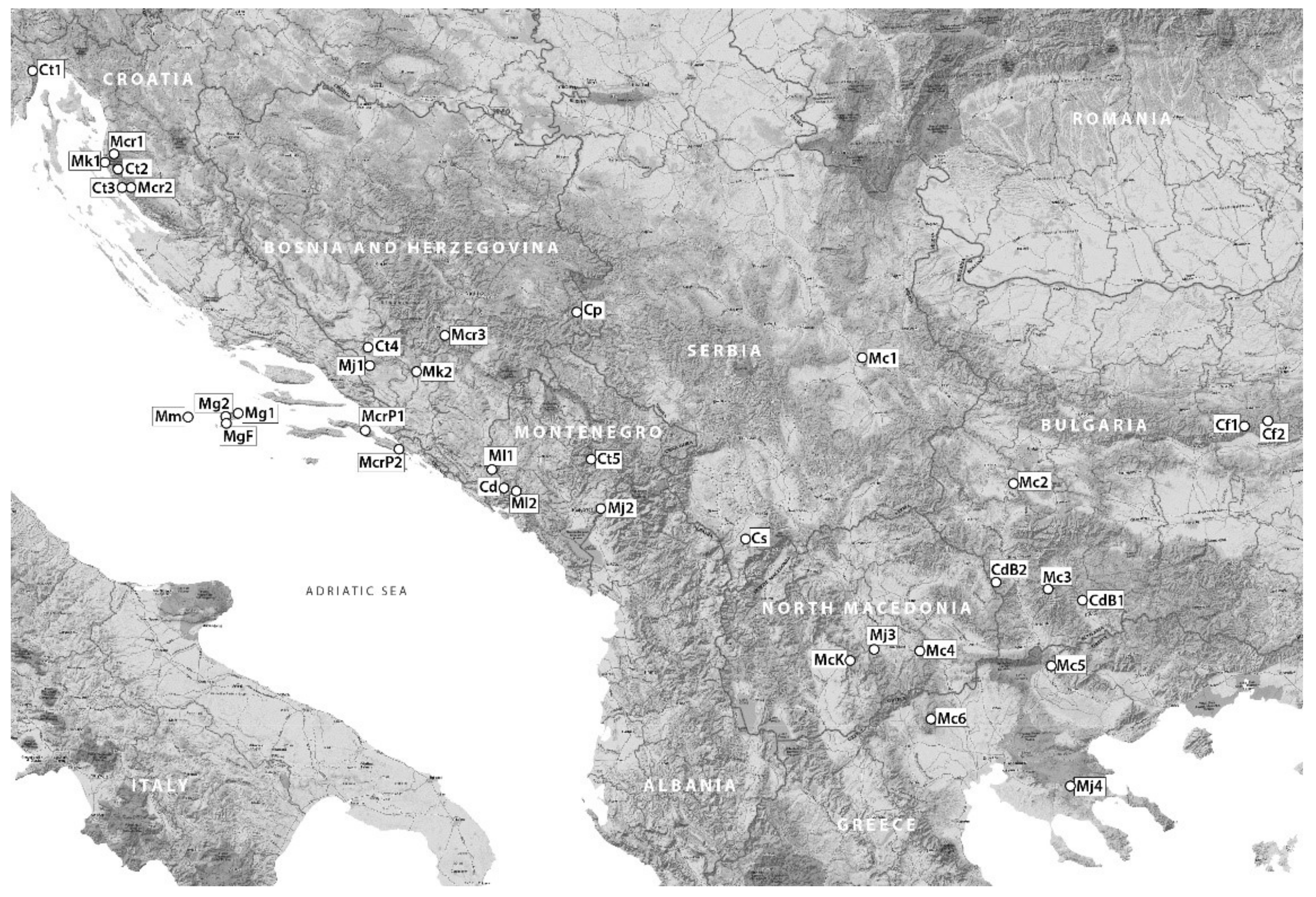
3.1. Plant Material
3.2. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Harley, R.M.; Atkins, S.; Budantsev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; De Kok, R.P.J.; Krestovskaja, T.; Morales, R. Labiatae. In The Families and Genera of Vascular Plants, 1st ed.; Kubitzki, K., Ed.; Springer: Berlin, Germany, 2004; Volume 4, pp. 167–282. [Google Scholar]
- Briquet, J. Labiatae. In Die Naturlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1895–1897; Volume 4, pp. 183–380. [Google Scholar]
- Killick, D.J.B. South African species of Satureia. Bothalia 1961, 7, 435–437. [Google Scholar] [CrossRef] [Green Version]
- Greuter, W.; Burdet, H.M.; Long, D. Satureja. In Med-Checklist; Conservatoire et Jardin Botaniques de la ville de Genève: Geneva, Switzerland, 1986; Volume 3, pp. 323–341. [Google Scholar]
- Bentham, G. Labiatae. In Prodromus Systematis Universalis Regni Vegetabilis; Candolle, A.P., Ed.; Treuttel et Würtz: Paris, France, 1848; Volume 12, pp. 212–226. [Google Scholar]
- Doroszenko, A. Taxonomic Studies on the Satureja Complex (Labiatae). Ph.D. Thesis, Edinburgh University, Edinburgh, UK, 1986. [Google Scholar]
- Valverde, R.M. Sinopsis y distribución del género Micromeria Bentham. Bot. Complut. 1993, 18, 157–168. [Google Scholar]
- Bräuchler, C.; Ryding, O.; Heubl, G. The genus Micromeria (Lamiaceae), a synoptical update. Willdenowia 2008, 38, 363–410. [Google Scholar] [CrossRef] [Green Version]
- Erhardt, W.; Götz, E.; Bödeker, N.; Seybold, S. Zander–Handwörterbuch der Pflanzennamen, 19th ed.; Eugen Ulmer GmbH und Co.: Stuttgart, Germany, 2014; p. 576. [Google Scholar]
- Bräuchler, C.; Meimberg, H.; Abele, T.; Heubl, G. Polyphyly of the genus Micromeria (Lamiaceae)—Evidence from cpDNA sequence data. Taxon 2005, 54, 639–650. [Google Scholar] [CrossRef]
- Bräuchler, C.; Meimberg, H.; Heubl, G. New names in old world Clinopodium—The transfer of the species of Micromeria sect. Pseudomelissa to Clinopodium. Taxon 2006, 55, 977–981. [Google Scholar] [CrossRef]
- Chater, A.O.; Guinea, E. Micromeria Benth. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 167–170. [Google Scholar]
- Werker, E.; Ravid, U.; Putievsky, E. Structure of glandular hairs and identification of the main components of their secreted material in some species of the Labiatae. Isr. J. Bot. 1985, 34, 31–45. [Google Scholar] [CrossRef]
- Husain, S.Z.; Marin, P.D.; Šilić, Č.; Qaser, M.; Petković, B. A micromorphological studies of some representative genera in the tribe Saturejeae. Bot. J. Linn. Soc. 1990, 103, 59–80. [Google Scholar] [CrossRef]
- Kremer, D.; Stabentheiner, E.; Dunkić, V.; Müller, I.D.; Vujić, L.; Kosalec, I.; Ballian, D.; Bogunić, F.; Bezić, N. Micromorphological and chemotaxonomical Traits of Micromeria croatica (Pers.) Schott. Chem. Biodivers. 2012, 9, 755–768. [Google Scholar] [CrossRef]
- Kremer, D.; Dunkić, V.; Ruščić, M.; Matevski, V.; Ballian, D.; Bogunić, F.; Eleftheriadou, E.; Stešević, D.; Kosalec, I.; Bezić, N.; et al. Micromorphological traits and essential oil contents of Micromeria kerneri Murb. and M. juliana (L.) Benth. (Lamiaceae). Phytochemistry 2014, 98, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Dunkić, V.; Kremer, D.; Grubešić, R.J.; Rodríguez, J.V.; Ballian, D.; Bogunić, F.; Stešević, D.; Kosalec, I.; Bezić, N.; Stabentheiner, E. Micromorphological and phytochemical traits of four Clinopodium L. species (Lamiaceae). S. Afr. J. Bot. 2017, 111, 232–241. [Google Scholar] [CrossRef]
- Kaya, A. Morphological characteristics of Clinopodium acinos and Clinopodium suaveolens (Lamiaceae) growing in Turkey. J. Res. Pharm. 2019, 23, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Mladenova, T.R.; Stoyanov, P.S.; Michova-Nankova, I.K.; Mladenov, R.D.; Boyadzhiev, D.T.; Bivolarska, A.V.; Todorov, K.T. Comparative leaf epidermis analyses of Micromeria frivaldszkyana (Degen) Velen. and Clinopodium vulgare L. (Lamiaceae) from Bulgarka Nature Park, Bulgaria. Ecol. Balk. 2019, 11, 133–140. [Google Scholar]
- Erdtman, G. Handbook of Palynology; Scandinavian Univ. Books: Munksgaard, Denmark, 1969. [Google Scholar]
- Moon, H.-K.; Vinckier, S.; Smets, E.; Huysmans, S. Palynological evolutionary trends within the tribe Menthae with special emphasis on subtribe Menthinae (Nepetoideae: Lamiaceae). Plant Syst. Evol. 2008, 275, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Firat, M.; Akçiçek, E.; Kaya, A. Clinopodium serpyllifolium subsp. sirnakense (Lamiaceae), a new taxon from south-eastern Anatolia, Turke. Phytotaxa 2015, 201, 131–139. [Google Scholar] [CrossRef]
- Doaigey, A.R.; El-Zaidy, M.; Alfarhan, A.; Milagy, A.E.-S.; Jacob, T. Pollen morphology of certain species of the family Lamiaceae in Saudi Arabia. Saudi J. Biol. Sci. 2018, 25, 354–360. [Google Scholar] [CrossRef]
- Padure, I.; Simic, S. Pollen and nutlet surface micromorphology of Clinopodium foliosum and C. menthifolium (Lamiaceae) from Istria. Joannea Botanik 2019, 16, 83–89. [Google Scholar]
- Payne, W.W. A glossary of plant hair terminology. Brittonia 1978, 30, 239–255. [Google Scholar] [CrossRef]
- Giuliani, C.; Bini, L.M. Insight into the structure and chemistry of glandular trichomes of Labiatae, with the emphasis on subfamily Lamioideae. Plant Syst. Evol. 2008, 276, 199–208. [Google Scholar] [CrossRef]
- Werker, E. Trichome Diversity and Development. In Advances in Botanical Research Incorporating Advances in Plant Pathology, 1st ed.; Hallahand, D.L., Gray, J., Callow, J.A., Eds.; Academic Press: London, UK, 2000; Volume 31, pp. 1–35. [Google Scholar]
- Corsi, G.; Bottega, S. Glandular hairs of Salvia officinalis: New data on morphology, localization and histochemistry in relation to function. Ann. Bot. 1999, 84, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Jia, P.; Liu, H.; Gao, T.; Xin, H. Glandular Trichomes and Essential Oil of Thymus quinquecostatus. Sci. World J. 2013, 3, 387952. [Google Scholar] [CrossRef] [Green Version]
- Kowalczuk, A.P.; Raman, V.; Galal, A.M.; Khan, I.A.; Siebert, D.J.; Zjawiony, J.K. Vegetative anatomy and micromorphology of Salvia divinorum (Lamiaceae). J. Nat. Med. 2014, 68, 63–73. [Google Scholar] [CrossRef]
- Stojičić, D.; Tošić, S.; Slavkovska, V.; Zlatković, B.; Budimir, S.; Janošević, D.; Uzelac, B. Glandular trichomes and essential oil characteristics of in vitro propagated Micromeria pulegium (Rochel) Benth. (Lamiaceae). Planta 2016, 244, 393–404. [Google Scholar] [CrossRef]
- Tozin, L.R.S.; Rodrigues, T.M. Morphology and histochemistry of glandular trichomes in Hyptis villosa Pohl ex Benth. (Lamiaceae) and differential labeling of cytoskeletal elements. Acta Bot. Brasilica 2017, 31, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Koca, F. Morphological and anatomical properties of Micromeria myrtifolia Boiss. et Hohen. Acta Pharm. Sci. 2002, 44, 235–242. [Google Scholar]
- Kremer, D.; Dunkić, V.; Stešević, D.; Kosalec, I.; Ballian, D.; Bogunić, F.; Bezić, N.; Stabentheiner, E. Micromorphological traits and essential oil of Micromeria longipedunculata Bräuchler (Lamiaceae). Cent. Eur. J. Biol. 2014, 9, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.; Jasnić, N.; Ascensão, L. Histochemical, micromorphology and ultrastructural investigation in glandular trichomes of Micromeria thymifolia. Bot. Serb. 2013, 37, 49–53. [Google Scholar]
- Hanlidou, E.; Kokkini, S.; Bosabalidis, A.M.; Bessiere, J.-M. Glandular trichomes and essential oil constituents of Calamintha menthifolia (Lamiaceae). Plant Syst. Evol. 1991, 177, 17–26. [Google Scholar] [CrossRef]
- Sonboli, A.; Fakhari, A.; Kanani, M.R.; Yousefzadi, M. Antimicrobial activity, essential oil composition and micromorphology of trichomes of Satureja laxiflora C. Koch from Iran. Z. Naturforsch C 2004, 59, 777–781. [Google Scholar] [CrossRef]
- Dunkić, V.; Kremer, D.; Müller, I.D.; Stabentheiner, E.; Kuzmić, S.; Grubešić, R.J.; Vujić, L.; Kosalec, I.; Randić, M.; Srečec, S.; et al. Chemotaxonomic and micromorphological traits of Satureja montana L. and S. subspicata Vis. (Lamiaceae). Chem. Biodivers. 2012, 9, 2825–2842. [Google Scholar] [CrossRef]
- Dodoš, T.; Janković, S.; Marin, P.D.; Rajčević, N. Essential oil composition and micromorphological traits of Satureja montana L., S. subspicata Bartel ex Vis., and S. kitaibelii Wierzb. ex Heuff. plant organs. Plants 2021, 10, 511. [Google Scholar] [CrossRef]
- Al-Zubaidy, A.M.A.; Hassan, K.I.; Jabbari, B.S. A comparative anatomical study of the genera Clinopodium L., Hymenocrater Fish. & Mey. and Melissa L. (Lamiaceae) in Kurdistan Region of Iraq. Diyala J. Pure Sci. 2015, 11, 30–43. [Google Scholar]
- Diklić, N. Micromeria Bentham. In Flora SR Srbije; Josifović, M., Ed.; Academie Serbe des Sciences et des Arts: Belgrade, Serbia, 1974; Volume 6, pp. 458–462. [Google Scholar]
- Šilić, Č. Monography of genuses Satureja L., Calamintha Miller, Micromeria Bentham, Acinos Miller and Clinopodium L. in flora of Yugoslavia; Zemaljski muzej BiH: Sarajevo, Bosnia and Herzegovina, 1979; pp. 172–262. [Google Scholar]
- Ančev, M. Micromeria Benth. In Flora Reipublicae Popularis Bulgaricae; Velčev, V., Ed.; Academiae Scientiarum Bulgaricae, Serdicae: Sofia, Bulgaria, 1989; Volume 9, pp. 356–362. [Google Scholar]
- Melnikov, D.G. New sections of the genus Clinopodium L. (Lamiaceae) and their synopsis. Turczaninowia 2015, 18, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, T. Flora Croatica; Alfa: Zagreb, Croatia, 2019; Volume 4, pp. 408–409. [Google Scholar]
- Matevski, V. Clinopodium albanicum (Griseb. ex K. Malý) Melnikov new species for the flora of the Republic of Macedonia. Contrib. Sect. Nat. Math. Biotech. Sci. 2018, 39, 123–128. [Google Scholar] [CrossRef]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Hill, T.; Lewicki, P. Statistics Methods and Applications; Stat Soft Inc.: Tulsa, OK, USA, 2006. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon/ | Trichomes | Leaf | Calyx | Stem | |
|---|---|---|---|---|---|
| Locality | Adaxial | Abaxial | |||
| M. cristata | NG | + | +/++ | ++ | + |
| ssp. cristata | peltate | + | + | ++ | + |
| Humsko | capitate C1 | +/++ | + | + | + |
| Brdo | capitate C2 | − | ±/+ | + | + |
| M. cristata | NG | + | +/++ | ++ | ++ |
| ssp. cristata | peltate | + | + | + | + |
| Vitosha Mt | capitate C1 | + | + | + | + |
| capitate C2 | − | ±/+ | + | ± | |
| M. cristata | NG | + | +/++ | ++ | +/++ |
| ssp. cristata | peltate | +/++ | + | +/++ | + |
| Pirin Mt | capitate C1 | + | + | + | + |
| capitate C2 | − | + | ± | ± | |
| M. cristata | NG | +/++ | +/++ | +/++ | + |
| ssp. cristata | peltate | + | + | + | + |
| Demir | capitate C1 | ±/+ | + | −/± | + |
| Kapija | capitate C2 | −/± | +/++ | +/++ | ±/+ |
| M. cristata | NG | +/++ | + | ++ | + |
| ssp. cristata | peltate | + | + | + | + |
| Nomos | capitate C1 | ±/+ | + | + | + |
| Serron | capitate C2 | − | + | + | +/++ |
| M. cristata | NG | + | +/++ | ++ | +/++ |
| ssp. cristata | peltate | + | + | + | ±/+ |
| Paikon Mt | capitate C1 | + | +/++ | + | + |
| capitate C2 | − | +/++ | + | +/++ | |
| M. cristata | NG | ++ | ++ | ++ | +/++ |
| ssp. kosaninii | peltate | +/++ | + | + | + |
| Pletvar | capitate C1 | ±/+ | ±/+ | + | + |
| capitate C2 | ±/+ | +/++ | ±/+/++ | + | |
| M. croatica | NG | + | + | + | + |
| Rossijev | peltate | − | +/++ | + | + |
| kuk | capitate C1 | − | ±/+ | ± | − |
| capitate C2 | ±/++ | ++ | ++ | − | |
| M. croatica | NG | + | + | ++ | + |
| Stupačinovo | peltate | − | + | −/± | + |
| capitate C1 | − | +/++ | + | ± | |
| capitate C2 | − | ±/++ | ± | +/++ | |
| M. croatica | NG | + | + | ++ | + |
| Dubočani | peltate | − | ++ | +/++ | + |
| capitate C1 | − | ±/++ | +/++ | ± | |
| capitate C2 | −/++ | ±/++ | ±/+ | +/++ | |
| M. croatica | NG | + | + | + | + |
| (syn. M. | peltate | − | ++ | + | + |
| pseudocroatica) | capitate C1 | ± | +/++ | + | + |
| Pijavičino | capitate C2 | ± | ++ | ++ | +/++ |
| M. croatica | NG | + | + | + | + |
| (syn. M. | peltate | − | ++ | + | + |
| pseudocroatica) | capitate C1 | ± | +/++ | + | + |
| Prapratno | capitate C2 | ± | ++ | ++ | +/++ |
| M. graeca | NG | ++ | ++ | +/++ | +/++ |
| ssp. graeca | peltate | + | + | + | + |
| Malo | capitate C1 | ± | ++ | ++ | + |
| zlo polje | capitate C2 | ± | + | ++ | + |
| M. graeca | NG | ++ | ++ | +/++ | +/++ |
| ssp. graeca | peltate | + | ++ | + | + |
| Komiža | capitate C1 | ± | ++ | + | + |
| capitate C2 | ± | + | ++ | ± | |
| M. graeca | NG | ++ | + | +/++ | ++ |
| ssp. fruticulosa | peltate | ±/++ | + | + | + |
| Sušac | capitate C1 | ± | + | + | + |
| capitate C2 | ± | + | + | + | |
| M. juliana | NG | + | + | ++ | +/++ |
| Grude | peltate | + | + | + | + |
| capitate C1 | ± | + | + | ±/+ | |
| capitate C2 | − | + | + | + | |
| M. juliana | NG | + | + | ++ | + |
| Cijevna | peltate | + | + | + | + |
| capitate C1 | + | ± | ++ | + | |
| capitate C2 | − | ++ | ++ | + | |
| M. juliana | NG | ++ | ++ | ++ | ++ |
| Rajec Reka | peltate | + | + | + | + |
| capitate C1 | ± | + | + | + | |
| capitate C2 | − | + | + | + | |
| M. juliana | NG | + | +/++ | +/++ | +/++ |
| Cholomon Mt | peltate | + | + | ±/+ | + |
| capitate C1 | + | + | + | + | |
| capitate C2 | − | ±/+ | ± | + | |
| M. kerneri | NG | +/++ | + | ++ | +/++ |
| Zavratnica | peltate | + | + | +/++ | + |
| bay | capitate C1 | ± | + | ± | + |
| capitate C2 | − | + | ++ | + | |
| M. kerneri | NG | +/++ | + | ++ | + |
| Mostar | peltate | ± | + | + | + |
| capitate C1 | ± | + | ±/+ | ± | |
| capitate C2 | − | ± | ++ | − | |
| M. | NG | + | + | + | +/++ |
| longipedunculata | peltate | − | + | + | ± |
| Jazina | capitate C1 | ± | +/++ | + | + |
| capitate C2 | − | + | + | ± | |
| M. | NG | + | + | + | + |
| longipedunculata | peltate | − | + | + | ± |
| Krivošije Mt | capitate C1 | ± | +/++ | + | + |
| capitate C2 | − | + | + | ±/+ | |
| M. microphylla | NG | + | + | ++ | + |
| Svetac Island | peltate | + | + | + | + |
| capitate C1 | −/±/+ | ±/+ | ± | ±/+ | |
| capitate C2 | − | + | +/++ | + | |
| Taxon/ | Trichomes | Leaf | Calyx | Stem | |
|---|---|---|---|---|---|
| Locality | Adaxial | Abaxial | |||
| C. dalmaticum | NG | + | +/++ | + | +/++ |
| Orjen | peltate | + | +/+ | + | + |
| capitate C1 | + | ±/+ | ±/+ | +/++ | |
| capitate C3 | ± | ++ | +/++ | − | |
| capitate C4 | − | ± | ± | ± | |
| C. dalmaticum | NG | + | +/++ | +/++ | + |
| (syn. M. bulgarica) | peltate | ± | + | +/++ | + |
| Mesta | capitate C1 | + | +/++ | ±/+/++ | ++ |
| River | capitate C3 | − | −/± | ±/+ | ± |
| Valley | capitate C4 | − | − | ±/+ | −/± |
| C. dalmaticum | NG | + | + | +/++ | + |
| (syn. M. bulgarica) | peltate | ± | ++ | +/++ | + |
| Vlahina Mt | capitate C1 | + | +/++ | + | ++ |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | − | −/± | −/± | |
| C. frivaldszkyanum | NG | ± | ± | + | −/+ |
| Malusha peak | peltate | ±/+ | + | +/++ | −/+ |
| capitate C1 | + | ++ | + | + | |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | −/± | + | − | |
| C. frivaldszkyanum | NG | ± | ±/+ | + | + |
| Vikanata Skala | peltate | ± | + | + | + |
| Nature | capitate C1 | + | + | + | + |
| Monument | capitate C3 | − | − | −/± | − |
| capitate C4 | − | − | + | − | |
| C. pulegium | NG | ± | + | + | + |
| Međeđa | peltate | + | ++ | ++ | + |
| capitate C1 | ± | +/++ | +/++ | + | |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | − | ± | − | |
| C. serpyllifolium | NG | +++ | +++ | ++ | ++ |
| Prizren | peltate | + | + | + | + |
| capitate C1 | + | ++ | ++ | +/++ | |
| capitate C3 | − | ± | ± | ± | |
| capitate C4 | − | ± | ± | − | |
| C. thymifolium | NG | ± | + | + | + |
| Učka Mt | peltate | + | +/++ | +/++ | + |
| capitate C1 | +/++ | +/++ | +/++ | +/++ | |
| capitate C3 | − | − | ± | − | |
| capitate C4 | − | − | ± | − | |
| C. thymifolium | NG | + | + | + | +/++ |
| Dokozina | peltate | + | +/++ | +/++ | + |
| plan | capitate C1 | + | ++ | ++ | + |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | − | − | − | |
| C. thymifolium | NG | + | + | + | + |
| Šušanj | peltate | + | ++ | +/++ | + |
| capitate C1 | + | ++ | +/++ | + | |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | − | ± | − | |
| C. thymifolium | NG | + | + | + | + |
| Blidinje | peltate | + | +/++ | +/++ | + |
| capitate C1 | + | ++ | +/++ | + | |
| capitate C3 | ± | − | − | − | |
| capitate C4 | − | − | − | − | |
| C. thymifolium | NG | ± | + | + | ±/+ |
| Manastir | peltate | + | +/++ | +/++ | + |
| Morače | capitate C1 | +/++ | ++ | ++ | +/++ |
| capitate C3 | − | − | − | − | |
| capitate C4 | − | − | ± | − | |
| Taxon | Polar Axis (P) | Equatorial Axis (E) | Pollen Size | P/E Ratio | |
|---|---|---|---|---|---|
| M. cristata ssp. cristata | Mean | 21.65 | 25.90 | Medium | 0.84 |
| Stdev | 1.10 | 2.28 | |||
| Min | 19.40 | 21.30 | |||
| Max | 23.60 | 28.60 | |||
| CV | 5.08 | 8.80 | |||
| M. cristata ssp. kosaninii | Mean | 21.00 | 22.94 | Small | 0.92 |
| Stdev | 0.91 | 1.10 | |||
| Min | 19.00 | 21.30 | |||
| Max | 22.70 | 25.00 | |||
| CV | 4.33 | 4.80 | |||
| M. croatica | Mean | 25.97 | 28.22 | Medium | 0.92 |
| Stdev | 1.86 | 1.93 | |||
| Min | 23.90 | 25.70 | |||
| Max | 30.50 | 32.40 | |||
| CV | 7.16 | 6.84 | |||
| M. croatica (syn. M. pseudocroatica) | Mean | 26.35 | 29.65 | Medium | 0.89 |
| Stdev | 2.38 | 2.80 | |||
| Min | 22.10 | 25.40 | |||
| Max | 30.00 | 23.90 | |||
| CV | 9.03 | 9.44 | |||
| M. graeca ssp. graeca | Mean | 28.65 | 34.65 | Medium | 0.83 |
| Stdev | 2.42 | 2.69 | |||
| Min | 24.60 | 31.20 | |||
| Max | 33.00 | 44.00 | |||
| CV | 8.45 | 7.76 | |||
| M. graeca ssp. fruticulosa | Mean | 26.73 | 33.86 | Medium | 0.79 |
| Stdev | 3.16 | 2.74 | |||
| Min | 22.40 | 28.10 | |||
| Max | 35.20 | 38.00 | |||
| CV | 11.82 | 8.09 | |||
| M. juliana | Mean | 23.37 | 27.10 | Medium | 0.86 |
| Stdev | 1.21 | 1.29 | |||
| Min | 21.30 | 25.10 | |||
| Max | 26.30 | 29.40 | |||
| CV | 5.18 | 4.76 | |||
| M. kerneri | Mean | 23.92 | 27.92 | Medium | 0.86 |
| Stdev | 1.40 | 1.02 | |||
| Min | 21.90 | 26.50 | |||
| Max | 26.20 | 29.90 | |||
| CV | 5.85 | 3.65 | |||
| M. longipedunculata | Mean | 18.18 | 21.10 | Small | 0.86 |
| Stdev | 0.74 | 1.67 | |||
| Min | 17.10 | 18.30 | |||
| Max | 19.40 | 25.50 | |||
| CV | 4.07 | 7.91 | |||
| M. microphylla | Mean | 18.65 | 21.80 | Small | 0.86 |
| Stdev | 1.18 | 0.90 | |||
| Min | 16.60 | 20.30 | |||
| Max | 20.80 | 23.60 | |||
| CV | 6.32 | 4.13 | |||
| C. dalmaticum | Mean | 26.67 | 32.47 | Medium | 0.82 |
| Stdev | 1.52 | 3.28 | |||
| Min | 24.40 | 27.50 | |||
| Max | 29.20 | 37.90 | |||
| CV | 5.70 | 10.10 | |||
| C. dalmaticum (syn. M. bulgarica) | Mean | 21.37 | 25.95 | Medium | 0.82 |
| Stdev | 2.82 | 1.35 | |||
| Min | 15.70 | 23.40 | |||
| Max | 24.10 | 27.30 | |||
| CV | 13.20 | 5.20 | |||
| C. frivaldszkyanum | Mean | 25.46 | 28.36 | Medium | 0.90 |
| Stdev | 1.23 | 0.99 | |||
| Min | 24.00 | 26.30 | |||
| Max | 28.50 | 29.40 | |||
| CV | 4.83 | 3.49 | |||
| C. pulegium | Mean | 26.87 | 28.90 | Medium | 0.93 |
| Stdev | 2.54 | 3.00 | |||
| Min | 25.00 | 26.10 | |||
| Max | 21.00 | 33.60 | |||
| CV | 9.45 | 10.38 | |||
| C. serpyllifolium | Mean | 22.27 | 26.21 | Medium | 0.85 |
| Stdev | 3.10 | 2.07 | |||
| Min | 12.60 | 22.60 | |||
| Max | 26.30 | 29.60 | |||
| CV | 13.92 | 7.89 | |||
| C. thymifolium | Mean | 22.15 | 23.52 | Small | 0.94 |
| Stdev | 1.21 | 1.17 | |||
| Min | 19.70 | 21.50 | |||
| Max | 24.60 | 25.90 | |||
| CV | 5.46 | 4.97 |
| Taxon | Mc4 | McK | Mcr2 | McrP1 | Mg1 | MgF |
|---|---|---|---|---|---|---|
| McK | 0.996467 | |||||
| Mcr2 | 0.000029 * | 0.000029 * | ||||
| McrP1 | 0.000029 * | 0.000029 * | 0.999995 | |||
| Mg1 | 0.000029 * | 0.000029 * | 0.000041 * | 0.000605 * | ||
| MgF | 0.000029 * | 0.000029 * | 0.981839 | 0.999995 | 0.013724 * | |
| Mj1 | 0.054533 | 0.000335 * | 0.000059 * | 0.000030 * | 0.000029 * | 0.000029 * |
| Mk1 | 0. 000813 * | 0.000030 * | 0.005354 * | 0.000199 * | 0.000029 * | 0.000032 * |
| Ml1 | 0.000029 * | 0.000032 * | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Mm | 0.000030 * | 0.000389 * | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Cd | 0.000029 * | 0.000029 * | 0.992281 | 1.000000 | 0.008540 * | 1.000000 |
| CdB1 | 1.000000 | 0.999997 | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Cf1 | 0.000029 * | 0.000029 * | 0.999806 * | 0.930688 | 0.000029 * | 0.464403 |
| Cp | 0.000029 * | 0.000029 * | 0.919882 * | 0.999711 | 0.037175 * | 1.000000 |
| Cs | 0.998023 | 0.469331 | 0.000029 | 0.000029 * | 0.000029 * | 0.000029 * |
| Ct4 | 0.999848 | 0.649443 | 0.000029 | 0.000029 * | 0.000029 * | 0.000029 * |
| Taxon | Mj1 | Mk | Ml1 | Mm | Cd | |
| Mk1 | 0.999474 | |||||
| Ml1 | 0.000029 * | 0.000029 * | ||||
| Mm | 0.000029 * | 0.000029 * | 0.999931 | |||
| Cd | 0.000029 * | 0.000035 * | 0.000029 * | 0.000029 * | ||
| CdB1 | 0.007514 * | 0.000077 * | 0.000029 * | 0.000037 * | 0.000029 * | |
| Cf1 | 0.003785 * | 0.151169 | 0.000029 * | 0.000029 * | 0.559547 | |
| Cp | 0.000029 * | 0.000030 * | 0.000029 * | 0.000029 * | 1.000000 | |
| Cs | 0.711451 | 0.081441 | 0.000029 * | 0.000029 * | 0.000029 * | |
| Ct4 | 0.534318 | 0.038831 * | 0.000029 * | 0.000029 * | 0.000029 * | |
| Taxon | Cd | CdB1 | Cf1 | Cp | Cs | |
| CdB1 | 0.000029 * | |||||
| Cf1 | 0.559547 | 0.000029 * | ||||
| Cp | 1.000000 | 0.000029 * | 0.272970 | |||
| Cs | 0.000029 * | 0.924334 | 0.000029 * | 0.000029 * | ||
| Ct4 | 0.000029 * | 0.977758 | 0.000029 * | 0.000029 * | 1.000000 | |
| Taxon | Mc4 | McK | Mcr2 | McrP1 | Mg1 | MgF |
|---|---|---|---|---|---|---|
| McK | 0.000032 * | |||||
| Mcr2 | 0.001274 * | 0.000029 | ||||
| McrP1 | 0.000029 * | 0.000029 * | 0.324744 | |||
| Mg1 | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * | ||
| MgF | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * | 0.983544 | |
| Mj1 | 0.638719 | 0.000029 * | 0.763081 | 0.000190 * | 0.000029 * | 0.000029 * |
| Mk1 | 0.012601 * | 0.000029 * | 1.000000 | 0.080933 | 0.000029 * | 0.000029 * |
| Ml1 | 0.000029 * | 0.044219 * | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Mm | 0.000029 * | 0.725134 | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Cd | 0.000029 * | 0.000029 * | 0.000029 * | 0.000039 * | 0.003949 | 0.379663 |
| CdB1 | 1.000000 | 0.000031 * | 0.001974 * | 0.000029 | 0.000029 * | 0.000029 * |
| Cf1 | 0.000381 * | 0.000029 * | 1.000000 | 0.518758 | 0.000029 * | 0.000029 * |
| Cp | 0.000031 * | 0.000029 * | 0.996257 | 0.990120 | 0.000029 * | 0.000029 * |
| Cs | 1.000000 | 0.000029 * | 0.014167 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Ct4 | 0.000793 * | 0.999419 | 0.000029 * | 0.000029 * | 0.000029 * | 0.000029 * |
| Taxon | Mj1 | Mk | Ml1 | Mm | Cd | |
| Mk1 | 0.977496 | |||||
| Ml1 | 0.000029 * | 0.000029 * | ||||
| Mm | 0.000029 * | 0.000029 * | 0.995845 | |||
| Cd | 0.000029 * | 0.000029 * | 0.000029* | 0.000029 * | ||
| CdB1 | 0.711996 | 0.018248 * | 0.000029 * | 0.000029 * | 0.000029 * | |
| Cf1 | 0.566977 | 0.999982 | 0.000029 * | 0.000029 * | 0.000029 * | |
| Cp | 0.055158 | 0.897260 | 0.000029 * | 0.000029 * | 0.000029 * | |
| Cs | 0.950327 | 0.090359 | 0.000029 * | 0.000029 * | 0.000029 * | |
| Ct4 | 0.000029 * | 0.000029 * | 0.000581 * | 0.083981 | 0.000029 * | |
| Taxon | Cd | CdB1 | Cf1 | Cp | Cs | |
| CdB1 | 0.000029 * | |||||
| Cf1 | 0.000029 * | 0.000598 * | ||||
| Cp | 0.000029 * | 0.000032 * | 0.999755 | |||
| Cs | 0.000029 * | 1.000000 | 0.004970 * | 0.000068 * | ||
| Ct4 | 0.000029 * | 0.000505 * | 0.000029 * | 0.000029 * | 0.000068 * | |
| Taxa—According to | Collecting Site | Voucher no. | |
|---|---|---|---|
| Bräuchler et al., 2008 (Code) | Balkan Literature | ||
| M. cristata ssp. cristata (Mc1) | M. cristata | Humsko brdo Mt (Serbia) | HFK-HR-51126 |
| M. cristata ssp. cristata (Mc2) | M. cristata | Vitosha Mt (Bulgaria) | HFK-HR-51132 |
| M. cristata ssp. cristata (Mc3) | M. cristata | Pirin Mt (Bulgaria) | HFK-HR-51133 |
| M. cristata ssp. cristata (Mc4) | M. cristata | Demir Kapija (N. Macedonia) * | HFK-HR-51141 |
| M. cristata ssp. cristata (Mc5) | M. cristata | Nomos Serron (Greece) | HFK-HR-51145 |
| M. cristata ssp. cristata (Mc6) | M. cristata | Paikon Mt (Greece) | HFK-HR-51146 |
| M. cristata ssp. kosaninii (McK) | M. kosaninii | Pletvar (N. Macedonia) * | HFK-HR-51142 |
| M. croatica (Mcr1) | M. croatica | Rossijev kuk (Croatia) | HFK-HR-51016 |
| M. croatica (Mcr2) | M. croatica | Stupačinovo (Croatia) * | HFK-HR-51017 |
| M. croatica (Mcr3) | M. croatica | Dubočani (BIH) | HFK-HR-51020 |
| M. croatica (McrP1) | M. pseudocroatica | Pijavičino (Croatia) * | HFK-HR-51032 |
| M. croatica (McrP2) | M. pseudocroatica | Prapratno (Croatia) | HFK-HR-51033 |
| M. graeca ssp. graeca (Mg1) | M. graeca | Malo zlo polje (Croatia) * | HFK-HR-51036 |
| M. graeca ssp. graeca (Mg2) | M. graeca | Komiža (Croatia) | HFK-HR-51037 |
| M. graeca ssp. fruticulosa (MgF) | M. fruticulosa | Sušac Island (Croatia) * | HFK-HR-51038 |
| M. juliana (Mj1) | M. juliana | Grude (BIH) * | HFK-HR-51043 |
| M. juliana (Mj2) | M. juliana | Cijevna Canyon (Montenegro) | HFK-HR-51045 |
| M. juliana (Mj3) | M. juliana | Rajec Reka (N. Macedonia) | HFK-HR-51168 |
| M. juliana (Mj4) | M. juliana | Cholomon Mt (Greece) | HFK-HR-51147 |
| M. kerneri (Mk1) | M. kerneri | Zavratnica (Croatia) * | HFK-HR-51018 |
| M. kerneri (Mk2) | M. kerneri | Mostar (BIH) | HFK-HR-51044 |
| M. longipedunculata (Ml1) | M. parviflora | Jazina (BIH) * | HFK-HR-51046 |
| M. longipedunculata (Ml2) | M. parviflora | Krivošije Mt (Montenegro) | HFK-HR-51047 |
| M. microphylla (Mm) | M. microphylla | Svetac Island (Croatia) * | HFK-HR-51039 |
| C. dalmaticum (Cd) | M. dalmatica | Orjen Mt (Montenegro) * | HFK-HR-51051 |
| C. dalmaticum (CdB1) | M. bulgarica | Mesta River Valley (Bulgaria) * | HFK-HR-51134 |
| C. dalmaticum (CdB2) | M. bulgarica | Vlahina Mt (Bulgaria) | HFK-HR-51135 |
| C. frivaldszkyanum (Cf1) | M. frivaldszkyana | Malusha peak (Bulgaria) * | HFK-HR-51136 |
| C. frivaldszkyanum (Cf2) | M. frivaldszkyana | Vikanata Skala Nature Monument (Bulgaria) | HFK-HR-51137 |
| C. pulegium (Cp) | M. pulegium | Međeđa (BIH) * | HFK-HR-51050 |
| C. serpyllifolium (Cs) | M. albanica | Prizren (Kosovo) * | HFK-HR-51074 |
| C. thymifolium (Ct1) | M. thymifolia | Učka Mt (Croatia) | HFK-HR-51077 |
| C. thymifolium (Ct2) | M. thymifolia | Dokozina plan (Croatia) | HFK-HR-51082 |
| C. thymifolium (Ct3) | M. thymifolia | Šušanj (Croatia) | HFK-HR-51081 |
| C. thymifolium (Ct4) | M. thymifolia | Blidinje (BIH) * | HFK-HR-51042 |
| C. thymifolium (Ct5) | M. thymifolia | Manastir Morača (Montenegro) | HFK-HR-51053 |
| Taxa–According to | Latitude | Longitude | Altitude a.s.l. (m) | |
|---|---|---|---|---|
| Bräuchler et al., 2008 (Code) | Balkan Literature | |||
| M. cristata (Mc1) | M. cristata | 43°22′45.0″ | 21°53′50.1″ | 387 |
| M. cristata (Mc2) | M. cristata | 42°29′33.4″ | 23°11′43.1″ | 980 |
| M. cristata (Mc3) | M. cristata | 41°46′11.4″ | 23°25′19.1″ | 1970 |
| M. cristata (Mc4) | M. cristata | 41°24′18.1″ | 22°15′47.0″ | 111 |
| M. cristata (Mc5) | M. cristata | 41°15′10.3″ | 23°24′49.8″ | 190 |
| M. cristata (Mc6) | M. cristata | 40°57′21.2″ | 22°20′02.0″ | 1650 |
| M. cristata ssp. kosaninii (McK) | M. kosaninii | 41°22′09.0″ | 21°39′06.1″ | 1020 |
| M. croatica (Mcr1) | M. croatica | 44°45′51.1″ | 4°59′17.1″ | 1641 |
| M. croatica (Mcr2) | M. croatica | 44°32′37.5″ | 15°10′04.7″ | 1058 |
| M. croatica (Mcr3) | M. croatica | 43°35′10.1″ | 18°04′44.0″ | 715 |
| M. croatica (McrP1) | M. pseudocroatica | 42°56′58.8″ | 17°22′02.1″ | 436 |
| M. croatica (McrP2) | M. pseudocroatica | 42°49′28.1″ | 17°39′57.9′ | 167 |
| M. graeca (Mg1) | M. graeca | 43°03′43.3′‘ | 16°12′55.9″ | 137 |
| M. graeca (Mg2) | M. graeca | 43°02′17.4″ | 16°05′49.5″ | 43 |
| M. graeca ssp. fruticulosa (MgF) | M. fruticulosa | 43°02′13.4″ | 16°05′50.5″ | 10 |
| M. juliana (Mj1) | M. juliana | 43°23′18.5″ | 17°23′27.8″ | 495 |
| M. juliana (Mj2) | M. juliana | 42°25′44.6″ | 19°28′53.5″ | 157 |
| M. juliana (Mj3) | M. juliana | 41°26′12.2″ | 21°52′06.4″ | 199 |
| M. juliana (Mj4) | M. juliana | 40°27′30.0″ | 23°31′04.6″ | 1100 |
| M. kerneri (Mk1) | M. kerneri | 44°42′02.2″ | 14°54′45.6″ | 161 |
| M. kerneri (Mk2) | M. kerneri | 43°20′27.7″ | 17°48′53.2″ | 61 |
| M. longipedunculata (Ml1) | M. parviflora | 42°42′11.7″ | 18°30′37.1″ | 347 |
| M. longipedunculata (Ml2) | M. parviflora | 42°32′51.8″ | 18°42′41.1″ | 624 |
| M. microphylla (Mm) | M. microphylla | 43°01′07.8″ | 15°45′08.2″ | 28 |
| C. dalmaticum (Cd) | M. dalmatica | 42°33′45.1″ | 18°37′36.6″ | 1074 |
| C. dalmaticum (CdB1) | M. bulgarica | 41°40′46.6″ | 23°43′29.7″ | 580 |
| C. dalmaticum (CdB2) | M. bulgarica | 41°50′30.6″ | 22°59′27.8″ | 1140 |
| C. frivaldszkyanum (Cf1) | M. frivaldszkyana | 42°45′01.0″ | 25°16′53.7′ | 1311 |
| C. frivaldszkyanum (Cf2) | M. frivaldszkyana | 42°45′57.6″ | 25°30′08.1″ | 1040 |
| C. pulegium (Cp) | M. pulegium | 43°44′01.2″ | 19°17′07.3″ | 563 |
| C. serpyllifolium (Cs) | M. albanica | 42°12′01.6″ | 20°45′57.7″ | 376 |
| C. thymifolium (Ct1) | M. thymifolia | 45°17′08.1″ | 14°12′02.4″ | 1189 |
| C. thymifolium (Ct2) | M. thymifolia | 44°39′04.3″ | 15°02′39.1″ | 1441 |
| C. thymifolium (Ct3) | M. thymifolia | 44°31′33.8″ | 15°06′45.1″ | 604 |
| C. thymifolium (Ct4) | M. thymifolia | 43°31′07.8″ | 17°23′09.8″ | 1195 |
| C. thymifolium (Ct5) | M. thymifolia | 42°45′50.9″ | 19°23′34.6″ | 301 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremer, D.; Stabentheiner, E.; Bogunić, F.; Ballian, D.; Eleftheriadou, E.; Stešević, D.; Matevski, V.; Ranđelović, V.; Ivanova, D.; Ruščić, M.; et al. Micromorphological Traits of Balcanic Micromeria and Closely Related Clinopodium Species (Lamiaceae). Plants 2021, 10, 1666. https://doi.org/10.3390/plants10081666
Kremer D, Stabentheiner E, Bogunić F, Ballian D, Eleftheriadou E, Stešević D, Matevski V, Ranđelović V, Ivanova D, Ruščić M, et al. Micromorphological Traits of Balcanic Micromeria and Closely Related Clinopodium Species (Lamiaceae). Plants. 2021; 10(8):1666. https://doi.org/10.3390/plants10081666
Chicago/Turabian StyleKremer, Dario, Edith Stabentheiner, Faruk Bogunić, Dalibor Ballian, Eleni Eleftheriadou, Danijela Stešević, Vlado Matevski, Vladimir Ranđelović, Daniella Ivanova, Mirko Ruščić, and et al. 2021. "Micromorphological Traits of Balcanic Micromeria and Closely Related Clinopodium Species (Lamiaceae)" Plants 10, no. 8: 1666. https://doi.org/10.3390/plants10081666