
Investigations on Fungi Isolated from Apple Trees with Die-Back Symptoms from Basilicata Region (Southern Italy)




1
School of Agricultural, Forestry, Food and Environmental Sciences (SAFE), University of Basilicata, Viale dell’Ateneo Lucano 10, 85100 Potenza, Italy
2
Department of Pharmacy, University of Salerno, Via Giovanni Paolo II 132, 84084 Salerno, Italy
3
Department of Engineering and Environmental Protection, Faculty of Agriculture, University of Agricultural Sciences and Veterinary Medicine, No. 3-5, Calea Manastur Street, 400372 Cluj-Napoca, Romania
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(10), 1374; https://doi.org/10.3390/plants11101374
Submission received: 2 March 2022
/
Revised: 17 May 2022
/
Accepted: 18 May 2022
/
Published: 21 May 2022
(This article belongs to the Special Issue Decline of Mediterranean Fruit Crops and Forests Associated with Fungal Trunk Pathogens)
Abstract
:Val d’Agri is an important orchard area located in the Basilicata Region (Southern Italy). A phenomenon affecting cv. “Golden Delicious” apples which lead to tree death has been observed in the past several years in this area. This phenomenon has already been detected in about 20 hectares and is rapidly expanding. The symptoms observed were “scaly bark” and extensive cankers, mainly located in the lower part of the trunk, associated with wood decay. Dead plants ranged from 20% to 80% and, in many cases, trees were removed by farmers. In order to identify the causes of this phenomenon, investigations were started in autumn/winter 2019. In order to determine the possible causal agents, fungal and bacterial isolations, from symptomatic tissues, were performed in laboratory. Bacterial isolations gave negative results, whereas pure fungal cultures (PFCs) were obtained after 3–4 passages on potato dextrose agar (PDA) media. Genetic material was extracted from each PFC and amplified by PCR using three pairs of primers: ITS5/4, Bt2a/Bt2b and ACT-512F/ACT-783R. The amplicons were directly sequenced, and nucleotide sequences were compared with those already present in the NCBI GenBank nucleotide database. All isolated fungi were identified based on morphological features and multilocus molecular analyses. Neofusicoccum parvum, Diaporthe eres and Trametes versicolor were most frequently isolated, while Pestalotiopsis funerea, Phomopsis spp. and Diaporthe foeniculina were less frequently isolated. All nucleotide sequences obtained in this study have been deposited into the EMBL database. Pathogenicity tests showed that N. parvum was the most pathogenic and aggressive fungus, while Phomopsis sp. was demonstrated to be the less virulent one. All the investigated fungi were repeatedly reisolated from artificially inoculated twigs of 2-year-old apple trees, cv. “Golden Delicious”, and subsequently morphologically and molecularly identified. The role played by the above-mentioned fungi in the alterations observed in field is also discussed.
1. Introduction
Apple die-back syndrome is a complex disease initially characterized by a stunted appearance of the plants and the presence of chlorosis symptoms on leaves. As the disease develops, cracks and necrotic lesions of the cortex appear mostly at the base of the stem and at the grafting point. Subsequently, “scaly bark” and extensive cankers, generally located in the lower part of the trunk develop, which are also associated with wood decay, and the progressive death of the trees is registered. Many authors worldwide investigated the apple die-back syndrome, attributing it to different causal agents, such as various phytopathogens or other possible physiological causes [1,2,3,4,5]. Furthermore, in 2011, Cloete et al. [6] reported the presence of the die-back syndrome and cankers on apples and pears in South Africa, from which they isolated several fungal pathogens belonging to the Diplodia, Phaeoacremonium, Phomopsis, Neofusicoccum and Eutypa genera. A very recent study by Di Francesco et al. [5], which characterized, in Brazil, apple cultivars’ susceptibility to Neofusicoccum parvum (Pennycook & Samuels) P.W. Crous, Slippers & A.J.L. Phillips, stated that due to climate change, this fungus is emerging as a new pathogen on species of the Rosaceae plant family. An apple tree die-back syndrome causing severe tree losses was also observed in the main apple producing regions in Tunisia, as described by Souli et al. [7]. The authors identified, both morphologically and molecularly, Phytophthora and Pythium species as being the causal agents and the factors that promoted apple tree die-back. They also showed that soil salinity contributed to increase the disease severity [7].
In Italy, apple cultivation is mainly concentrated in the northeast part of the country, specifically in Trentino Alto Adige/Südtirol region. This area comprises about half of the harvested hectares and has an intensive growing system, producing almost 70% of the Italian apples. However, some other Italian regions such as Veneto, Emilia-Romagna, Piedmont, Campania and Basilicata also successfully cultivate apple trees with quite relevant productions. Overall, in 2019 in Italy, the surface cultivated with apple was about 58,000 hectares and the apple production reported in the same year was about 23 million quintals [8].
Due to the economic importance of this crop in Italy, many studies regarding its cultivation and phytosanitary status have been performed. The phytosanitary surveys on apples cultivated in Bolzano area, was investigated by Lindner [9]. The author reported the cortical damage of apple cvs. “Gala”, “Golden Delicious” and “Red Delicious”, in spring 2007, and alterations very much resembling the “blister bark” and “paper bark” symptoms often associated the with withering and drying of the branches were also described. Nevertheless, the author reported that they did not isolate any bacterial or fungal pathogen from the plants, concluding that the cause of the observed symptoms was probably to be found among particularly unfavorable climatic conditions registered during the winter and spring periods. The same author also observed damages at the grafting point level and identified two fungi, known as canker agents, namely Phomopsis mali and Diplodia malorum.
The apple tree die-back syndrome was reported during 2008–2009 in many apple orchards in the north of Italy and since that period many other trees have become infected, especially young plants.
The phytosanitary status of the apples from Trentino region (Northern Italy) was investigated by Prodorutti et al. [10]. The authors reported an increase in the die-back symptoms on apple trees, showing that plants were usually stunted with cracking and necrosis in the lower part of the trunk and on the graft union site. The trees died during the growing season. Furthermore, the same authors reported that the incidence of the disease was, in some cases, very high (reaching almost 80%), and that the most affected trees were the youngest ones of about 2–5 years old that had been subjected to various types of stress. One bacterium, Pseudomonas syringae pv. syringae, and a few fungi, such as Phomopsis spp., Neonectria spp. And some Botryosphaeriaceae, were isolated from trunk tissues, taken from symptomatic trees. Their role in the syndrome expression was also demonstrated, even if they appeared not to cause the death of the artificially inoculated branches [10].
The Val d’Agri area, located in the Basilicata Region (Southern Italy), has geographical and climatic conditions that favor apple cultivation. In particular, “Fuji” and “Golden Delicious” apple cultivars are mostly grown in the area. Apple orchards belong to private farmers and are kept as small-scale cultivation systems. During the autumn/winter 2019, in the Val d’Agri area, on about 20 hectares cultivated with apples, symptoms characterized by “scaly bark” and extensive cankers, mainly located in the lower part of the trunk and associated with wood decay, were observed. Additionally, dead plants ranged from 20 to 80%, and in many cases, trees were removed by farmers. In order to identify the causes of this phenomenon, investigations were started in 2019. It is to be mentioned that despite good prevention and control measures employed so far against the key diseases on fruit trees in the south of Italy, including the Basilicata region, knowledge about the distribution and the pathogens involved in apple die-back syndrome is still missing. These data are very important since the presence of the die-back syndrome could economically affect the growers in the region. The identification and characterization of various fungal and bacterial pathogens attacking fruit trees, including apples, were initially based on only the morphological features of the pure cultures obtained in vitro [11,12]. However, over time, despite the ease of application, the morphological features proved to be inefficient to further classify fungal and bacterial pathogens. Therefore, other solutions, including molecular approaches, were investigated in order to identify and characterize the phytopathogens associated with the die-back symptoms [6,7,13,14]. Nowadays, several gene regions or genes, such as the Internal Transcribed Spacer (ITS) of the ribosomal DNA (rDNA), β-tubulin (TUB-2) and actin (ACT) protein-coding genes, are extensively utilized to identify and characterize phytopathogens [15,16,17,18,19,20,21,22,23].
The aim of the present study was to investigate the die-back syndrome on apple orchards from the Val d’Agri area (Basilicata region, Southern Italy). More precisely, the main objectives of the present study were to: (1) identify fungi or bacteria eventually associated with the die-back symptoms observed on apple trees; and (2) perform pathogenicity tests on apple trees in order to verify the involvement of the identified pathogens in the apple die-back disease observed in the Val d’Agri area.
2. Results
2.1. Pathogens Isolations
Pure culture fungal isolates on PDA media from die-back symptomatic material obtained in this study were selected for further characterization through morphological and cultural characteristics, DNA sequencing and phylogenetic analysis (Table 1).
From apple die-back symptomatic samples, the above-described fungi were isolated with different frequencies. Among the most frequently isolated fungi were the N. parvum, with a 55% isolation frequency (IF%), followed by D. eres, with a 15% IF, and T. versicolor, with a 14% IF. All the other fungi were less frequently isolated with an IF ranging from 10–12%, except for Phomopssis spp., which was very rarely isolated (<5% IF).
Despite repeated trials to isolate bacteria from symptomatic apple wood, no bacterial colonies were ever obtained. During the investigation for the identification of the apple die-back disease cause no symptom or damage of the root system were noticed. Furthermore, all isolation attempts, performed from roots taken from the symptomatic apple trees, gave negative results.
2.2. Morphological Identification
Based on their cultural and morphological features, pure fungal isolates were classified in five distinct genera: Neofusicoccum [24], Diaporthe [25,26,27,28], Trametes [29,30,31,32,33], Pestalotiopsis [34,35,36] and Phomopsis [6,37,38,39,40] (Table 1 and Figure 1). In particular, in the case of Diaporthe grayish or white colonies on PDA and alpha and beta conidia were observed; in the case of Neofusicoccum, grey-black colonies and fusiform conidia, nonseptate when young and biseptate ellipsoidal (partially light brown with a darker middle center) when old, were detected. In the case of Phomopsis, white colonies and alpha and beta conidia were observed. For Pestalotiopsis, reddish colonies and 4-septate conidia, fusiform to ellipsoid and straight to slightly curved, were noticed. Trametes genus was identified based on white colonies and the presence of clavate basidia with an inflated epibasidial segment, 4-spored, clamped at the base and basidiospore cylindrical in large spores with slightly inflated top, ellipsoid to ovoid.
2.3. Molecular Characterization
The PCR amplifications for each gene investigated yielded amplicons of expected sizes: ITS5/ITS4 (~700 bp), tub-2 (~500 bp) and ACT (~300 bp), which, after direct sequencing in both directions, using the same primers as for the amplification, led to 26 nucleotide sequences (Table 1). A megablast search, excluding “uncultured/environmental sample sequences”, performed in the NCBI’s nucleotide database (www.ncbi.org, accessed on 12 January 2022) for all nucleotide sequences obtained in this study, identified at Genus level all fungal isolates (Table 1).
2.4. Phylogenetic Analysis
Single locus analysis gave consistent results for all three loci (ITS, tub-2 and ACT), and the topology of trees was congruent in terms of species grouping. All sequences obtained in this study have been deposited in the European Molecular Biology Laboratory (EMBL-EBI) nucleotide database (www.ebi.ac.uk, accessed on 12 January 2022) and their GenBank accession numbers are presented in Table 1.
The final alignment dataset, for the ITS region, was composed of a total number of 658 characters. It contained 58 nucleotide sequences, including five outgroup species, namely: Diaporthella corylina (acc. no. KC343004) utilized for Diaporthe fungi; Valsa japonica (acc. no AF191185) for Phomopsis sp. fungi; Sordaria alcina (acc. no. AY681198) for the Pestaliopsis sp. Fungal group; Grifola frondosa (acc. No. AY049140) utilized for Trametes sp. Fungi; and for the Neofusicoccum parvum group, Diplodia seriata (acc. no. MH221102). Phylogenetic analysis based on the ITS region variation showed that fungal isolates from the same species clustered together into the same clade and, as expected, outgroup species were placed separately from the other groups (Figure 2).
Overall, the 53 nucleotide sequences obtained in this study, based on the ITS region sequence analysis, clustered in two clades which contained all six fungal species. Two of these species belonged to Diaporthe genus, namely, Diaporthe eres Nitschke and D. foeniculina (Sacc.) Udayanga & Castl., and others have been identified as N. parvum, Pestalotiopsis funerea (Desm.) Steyaert, Phomopsis sp. Sacc. & Roum. and T. versicolor (L.) Lloyd (Figure 2). Within the phylogenetic tree, the first clade grouped together five of the species previously mentioned, while the second clade contained only one species, Trametes versicolor, clearly separated from the others. Additionally, the fungal species isolated and identified in this study were positioned close to similar reference species downloaded from the GenBank for each fungus, and their location was well supported by very high (97–100%) bootstrap values (Figure 2). The ITS data confirmed the previous preliminary fungal identification based on morphological features.
Since ITS alone does not provide sufficient resolution to exactly classify fungi at species level, other loci were considered for the phylogeny-based identification of the taxa investigated in this study. In particular, the β-tubulin (tub-2) gene, a very well-known molecular locus extensively used in phylogenetic studies of phytopathogenic fungi [35,38,39,40], was examined. A total number of 39 nucleotide sequences of the tub-2 partial gene were obtained and employed along with the species D. corylina, B. dothidea and S. alcina, used as outgroups in phylogenetic analysis, which was carried out using the NJ method, as performed for the ITS (Figure 3).
Results from the phylogenetic analysis of the fungal species from this study, based on the tub-2 gene, have shown that they grouped together with similar species from the GenBank database. A better separation within the clades and subclades compared to what obtained from the ITS was also observed (Figure 3). Two separate clades were obtained from the nucleotide sequences investigated in this study. In one, Neofusicoccum isolates grouped together into the same subgroup, which was very well supported (98% bootstrap support value), and were separated from the Pestalotiopsis subclade, which was also highly supported (99% bootstrap support value), whereas the Diaporthe isolates were all placed in a separate clade and were clearly distinguished in two subclades as species, e.g., D. eres and D. foeniculina, both sharing their vicinity with the same outgroup, D. corylina (Figure 3). All fungal species based the tub-2 gene variation were grouped together with the same species from the GenBank with an elevated bootstrap support (98–99%). The separation of the fungal species within each clade or subclade, strongly supported by high bootstrap values, was 99% for D. eres, D. feoniculina and N. parvum and 98% in the case of P. funerea (Figure 3).
Regarding the third gene, namely actin, despite our repeated PCR trials, amplicons could not be obtained for all fungal species (Figure 4). Therefore, the alignment for the ACT gene used in phylogenetic analysis contained 298 characters and involved only 24 nucleotide sequences. Moreover, ACT gene analysis showed that the two Diaporthe species identified in this study along with their reference species from the GenBank were well separated from the Neofusicoccum sp. isolates and, thus, clustered in two different clades also supported by very high bootstrap values of 94% and 99%, respectively. In addition, Neofusicoccum isolates were grouped together with their reference species with a 99% bootstrap support value (Figure 4). The phylogenetic reconstruction based on the ACT gene reconfirmed the molecular identification based on other loci at species level for each fungal species analyzed and also was in concordance with the preliminary morphological characterization.
Multilocus phylogenetic analyses for Diaporthe, Neofusicoccum and Pestalotiopsis spp. isolates showed that the topology of the trees was congruent in terms of grouping for all fungal species investigated, supporting the single locus phylogenetic outcomes (Figures S1–S3 in Supplementary Material).
2.5. Pathogenicity Trial
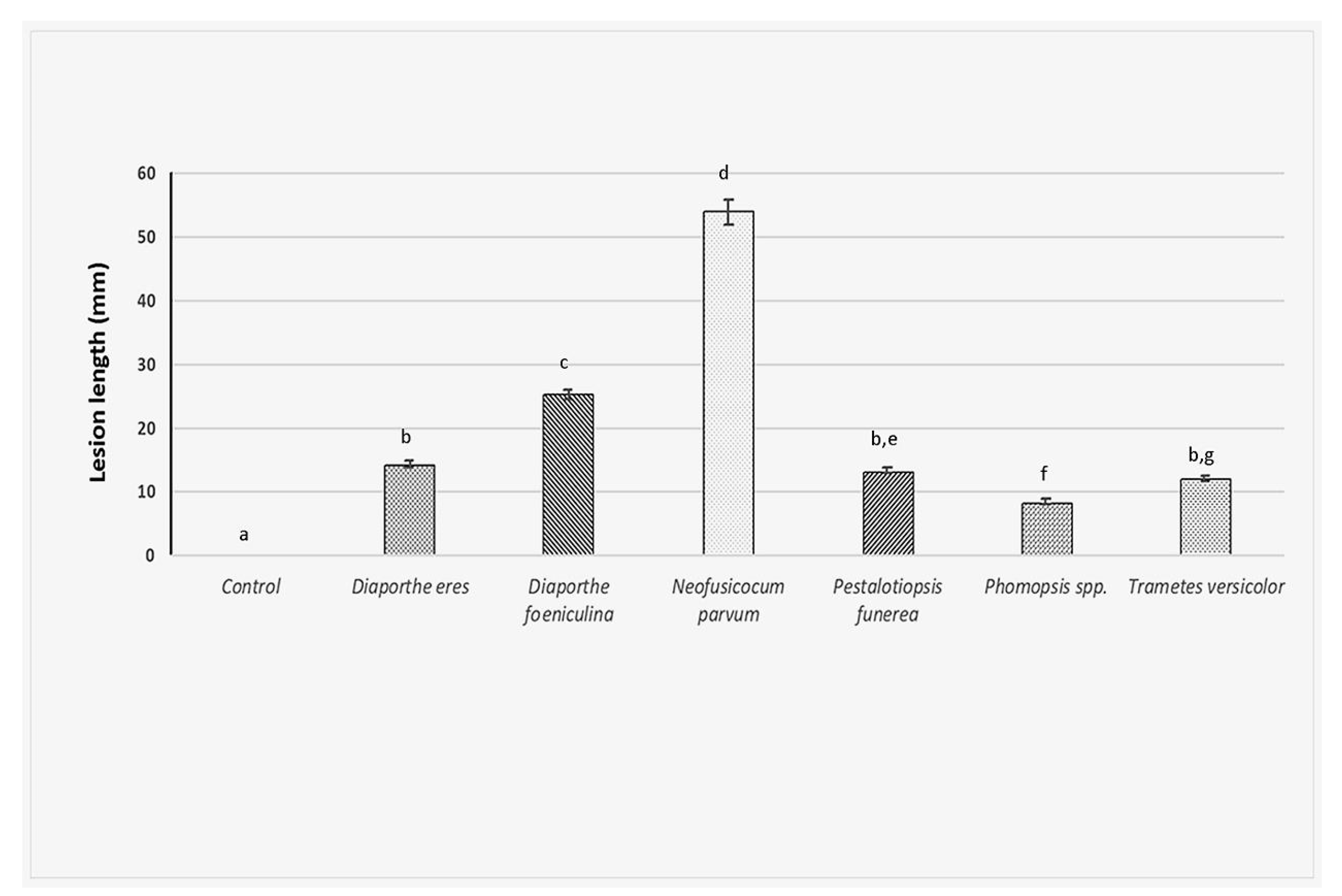
In the artificial inoculations test, on twigs of 2-year-old apple trees (cv. “Golden Delicious”), using the six fungi investigated in this study, the size of the observed lesions greatly varied among isolates (F6,77 = 390, p < 0.001). Furthermore, the tested fungal isolates produced lesions in the host that were always larger than those observed from the control (Tukey t-tests, p < 0.001 in all cases). Most of the fungal isolates (D. eres, P. funerea, Phomopsis spp. and T. versicolor) developed smaller lesions than D. foeniculina and N. parvum. No lesion developed after control treatment (Figure 5).
Among all fungi investigated, N. parvum produced the longest lesions (53.98 mm). It proved to be also the most pathogenic since the inoculated tree showed very strong die-back symptoms, such as reddish-brown cankers on the twigs, associated with internally brown necrosis. Finally, the death of all twigs and whole branches was observed at one month after artificial infection. D. feoniculina produced 25 mm length cankers on twigs with internally brown necrosis and death of some twigs and branches was also noticed. D. eres, P. funerea and Phomopsis formed similar lesions as described above but of a shorter length, which ranged between 8–14 mm. In the case of T. versicolor, symptoms of wood caries were also seen (Figure 5).
All the inoculated fungi were always reisolated from the lesions and based on molecular methods were identical to the cultures used for inoculation.
3. Discussion
This study is the first to address the presence of the die-back syndrome on apple orchards in the Val d’Agri region and to isolate and further characterize fungal species which could be involved in the observed disease through morphology, DNA sequencing and phylogenetic analysis. The search for the causal agent of the apple die-back syndrome contributed to in vitro isolation of six fungi, already known to be involved in different diseases in apple and other plant species. Both cultural and morphological features of the five phytopathogenic genera, namely Neofusicoccum, Diaporthe, Trametes, Pestalotiposis and Phomopsis, identified in the present study were consistent and resembled the above-mentioned ones.
The morphological classification of fungi is an inexpensive and rapid tool but has also many limitations. As a consequence, current mycotaxonomy has changed a lot, now employing other methodological approaches, such as phylogeny, chemotaxonomy, genetics, ecology or molecular biology [35,41,42,43,44,45,46]. The preliminary identification of the fungi isolated from the apple plant, in the present study based on morphological features, was confirmed by molecular outcomes gained from the sequencing of the ITS region, (tub-2 and actin (ACT) genes. A lesser variation was noticed over the ITS region, for all fungal species investigated, and only in the case of the reference sequences downloaded from the database, which was probably due to sequencing errors. It is widely accepted that sometimes the sequences deposited in the GenBank are of poor quality and around 30% of the ITS sequences deposited may be associated with the wrong taxon [47].
ITS locus alone, despite its advantages and official recognition as a DNA barcoding marker [48,49,50,51,52], can be limited in providing enough resolution in the case of closely-related fungal species [53]. Considering all these limitations, for an accurate fungal species identification other loci like tub-2 and ACT were explored, showing that nucleotide variation was higher in ACT gene, followed by the tub-2 gene for all fungal species investigated.
Typically, different wood-rotting fungi have been associated with the die-back syndrome in apple over the years as Coriolus spp., Stereum spp., Schizophyllum commune [1]. Apart from these, Sphaeropsis pyriputrescens Xiao & J.D Dogers fungus was reported to cause cankers and twig die-back on apple and crabapple trees in the USA [54]. Cloete et al. [6] found that apple and pear trees in South Africa are the hosts of many fungi associated with the die-back symptoms, such as Diplodia spp., Neofusicoccum spp., Phaeoacremonium spp. and Phomopsis sp. Very recent studies of Jabiri et al. [55] reported symptoms of dieback disease, such as root rot, yellow leaves and wilting, caused by Phytopytium vexans on young apple trees (6–10 years old) of cv. “Golden Delicious” in Morocco.
The Diaporthe genus has also been associated to the shoot canker or fruit rot in pear [56,57]. Dissanayake et al. [28,58], based on molecular phylogenetic analysis, revealed seven new species, within the above-mentioned genus, in Italy. Among the Diaporthe species, D. eres has recently been reported to be linked to necrosis and stem cankers and caused the death of young apple rootstocks in Canada [59]. Moreover, D. eres, is among the most serious phytopathogenic fungi affecting many plant species all over the world [26,28,56,57,58,59,60,61,62,63]. The outcomes from this study, showing the frequent isolation from apple with die-back symptoms of D. eres, agree with the previous studies by Sessa et al. [64] who reported the D. eres isolation from peach and apple with wood disease symptoms, such as wedge-shaped necrosis and canker. Additionally, the identification of more than one species of Diaporthe, namely D. eres and D. neotheicola, on the same plant species in our study is in agreement with the earlier works of van Niekerk et al. [40] and of Thompson et al. [65], who reported that the same host plant may be colonized by different Diaporthe spp. at the same time.
Neofusicoccum parvum, which recently emerged as a phytopathogen, was also among the most frequently isolated fungi in the present study. The Neofusicoccum Crous, Slippers and A.J. L. Phillips Genus was described by Crous et al. [24], aiming to include Botryosphaeriaceae with Fusicoccum-like anamorphs [66]. This fungus has been already reported to cause cankers on many cultivable plant species [67], including apple and pear [68], but it seems that it is expanding its host range, as demonstrated by the very recent study by Choi et al., in Korea [69]. Moreover, N. parvum was the most aggressive fungus in our study, as demonstrated by both natural and artificial inoculations on apple from Val d’Agri, also causing the death of the entire tree.
Trametes versicolor was among the wood fungi frequently isolated in the present investigation. Fungi from the Genus Trametes are white rot polypores. Nearly 60 species are known worldwide on many hosts, and some are used for medicinal purposes [70,71,72], The taxonomy situation within T. versicolor is still complex, since unresolved phylogenies and unclear species boundaries exist [31,33]. The study of Kile [73], who examined host-pathogen relationships between the apple tree, T. versicolor and factors affecting host susceptibility, showed that the fungus was a facultative parasite which caused the white rot of the sapwood and the susceptibility of living wood to fungal decay increased with the age of the tree, due to a natural decline in host plant resistance. The frequent isolation of T. versicolor from apple trees showing die-back symptoms agree with the study of Darbyshire et al. [74], which associated the die-back of apple trees in Australia to the wood-rotting fungus T. versicolor and also showed that it is a low-sugar disease. An association between Coriolus versicolor (syn. T. versicolor) and the die-back disease of apples in Washington state, described by Dilley and Covey, supported the present study outcomes [75]. We can assume that frequent in vitro isolation of T. versicolor from apple trees obtained in our study and the die-back symptoms observed after natural and/or artificial inoculation to this host can be associated with this disease.
Many studies reported that the Diaporthe (Phomopsis) fungi has been associated with shoot blight and canker, decay, wilting, necrosis of bark and fruit rot in several fruit tree species worldwide [27,37,76,77,78]. The isolation of Phomopsis sp. in this study agrees with the earlier studies by Pertot and Vindimian [79], who reported the diffusion of P. mali, causing the dieback of young apple trees in Trentino (Northern Italy). Cloete et al. [6] also found three Phomopsis sp. isolates from pear and apple exhibiting die-back symptoms and considered them as a possible inoculum source for grapevine trunk disease pathogens. Our outcomes concord with the results of Bai et al. [56], who identified the presence of these fungi on pear in China, and of Kanematsu et al. [37], who, in Japan, showed that they were responsible for shoot cankers.
Regarding Diaporthe species, our results from pathogenicity tests are similar to those reported by Sessa et al. [64], who investigated the diversity and the virulence of the Diaporthe species associated with wood disease symptoms in deciduous fruit trees in Uruguay. The same authors recognized them to be the causal agents of twig and branch cankers, showing that D. eres and D. foeniculina produced necrosis. Furthermore, another study by Abramczyk et al. [62] characterized isolates of D. eres based on morphological and pathological characteristics, which were isolated from fruit plants and genetically identified as D. eres species complex [78]. Additionally, they demonstrated that in pathogenicity tests D. eres produced small necrosis of about 12–17 mm in diameter, occurring at the site of inoculation. D. eres colonies were obtained from the artificially inoculated tissue, again confirming the results obtained in our study on this fungus, showing its pathogenic abilities towards apple trees.
Pathogenicity tests results showing that N. parvum was the most virulent among all fungal species isolated and identified on apple trees from the Val d’Agri region match those by Cloete et al. [6]. The authors analyzed fungi associated with die-back symptoms of apple and pear trees cultivated in proximity of grapevine in Western Cape, South Africa, and found that a species of Neofusicoccum (N. australe) was among the most virulent species towards apple, with mean necrotic lesions of about 40.2 mm length. In the same study, Phomospsis sp. was observed to be less virulent (necrotic lesions of about 11.8 mm in length), and this was similar to the results obtained in the present study (necrotic lesions of about 8 mm in length). Another study by Espinoza et al. [80] found that Neofusicoccum spp. was associated with the stem canker and dieback of blueberry in Chile and reported, for the first time, N. parvum as a canker-causing agent on blueberry. In their study, the same authors performed pathogenicity tests on kiwi, blueberry and apple and found that N. parvum was the most aggressive fungus, in all hosts, and this is also in accordance with our results.
4. Materials and Methods
4.1. Biologic Material
Pieces of symptomatic trunks from the apple cv. “Golden Delicious”, showing die-back symptoms, were collected in autumn/winter 2019. During this period, apple orchards located in the Val d’Agri area were surveyed for the presence of apple die-back symptoms. A total number of 50 samples, made of pieces of living material (bark and cankered trunks) showing die-back symptoms, more specifically, “scaly bark” and extensive cankers, mainly located in the lower part of the trunk, and wood decay were obtained from trees 3–12 years old (Figure 6). They were brought to the Plant Pathology Laboratory at the University of Basilicata and stored in fridges, at 4 °C, until used.
4.2. Pathogen Isolation
Symptomatic wood pieces were cut under laminar flow sterile conditions into small parts, surface-sterilized by soaking in a 70% ethanol solution for 1 min, in a 1% NaOCl solution for 1 min, in 70% ethanol solution for another 30 sec and finally rinsed in sterile water for 2 min. After sterilization, the trunk pieces were dried on a sterile paper and cut into small parts. Small parts of about 2 × 2 mm taken from the margins between necrotic and healthy tissue were placed on petri plates containing potato dextrose agar (PDA, Oxoid Ltd., Hants, UK), amended with streptomycin sulphate (40 mg L−1, MerckKGaA, Darmstadt, Germany) and were incubated at 25 °C in the dark until growth could be detected. Subcultures were performed from the growing hyphae onto PDA and incubated under the same conditions. Pure cultures were created for all obtained PDA plates.
To isolate and identify bacterial pathogens probably linked to the die-back syndrome symptomatic wood trunk, samples were first surface sterilized and prepared, as reported by Schaad et al. [81].
4.3. Morphological Identification
All fungal isolates obtained in this study were stored, as pure cultures (PFC), in the culture collection of the Plant Pathology Laboratory of the School of Agriculture, Forestry, Food and Environmental Sciences (SAFE) at the University of Basilicata on PDA slants and maintained at 4 °C in fridge.
Fungal isolates were examined using a Axioscope microscope (Zeiss, Jena, Germany) and preliminary identified by morphological characteristics.
4.4. Molecular Characterization
For molecular characterization, genomic DNA was extracted from fresh PFC mycelia of each isolate, 7–10 days old, through an extraction protocol described by Mang et al. [43]. Genomic DNA quality and quantity were checked using a Nanodrop ND-1000 spectrophotometer (Thermo Scientific Inc., Willmington, DE, USA) and the material was stored at −20 °C in 1.5 mL Eppendorf tubes until further use. In order to determine the fungal species, three different genes/regions were amplified. Namely, the internal transcribed spacers (ITS1 and ITS2) of the ribosomal RNA (ITS); β-tubulin (tub-2) and actin (ACT). The oligonucleotides used for PCR amplifications were: ITS5/ITS4 [82], Bt2a/Bt2b [83] and ACT512F/ACT783R [84] (Table 2).
PCR amplifications were performed under the conditions explained in Mang et al. [43,85] for ITS only. For the other two genes the Phire Direct PCR Master mix (Thermo Scientific Inc., USA) was used, following manufacturer’s instructions with some modifications. PCR mixtures were composed of 10 µL of 2X Phire Plant PCR Buffer (including 1.5 mM MgCl2 and 20 µM of dNTPs), Primers 0.5 µM each; 0.4 µL of Phire Hot Start II DNA polymerase enzyme, 5 µL of template DNA (20 ng/µL) and double distilled water up to 20 µL. The PCR cycling protocol consisted of: an initial denaturation at 98 °C for 5 min for 1 cycle; then 40 cycles of denaturation at 98 °C for 5 s; annealing at 60 °C for ITS and at 62 °C for tub-2 and ACT genes for 5 s; extension at 72 °C for 20 s, followed by a final extension at 72 °C for 1 min for 1 cycle. All PCR products were separated in 1.5% agarose gels in Tris-Acetic acid-EDTA (TAE) buffer and visualized under the UV after staining with SYBR Safe DNA Gel Stain (ThermoFisher Scientific™, Carlsbad, CA, USA). A 100-bp GeneRuler Express DNA Ladder (ThermoFisher Scientific™ Baltics UAB, Vilnius, Lithuania) was used as a molecular weight marker. Direct sequencing of all PCR products was performed by BMR Genomics [Padua, Italy], using a 3130xl automatic sequencer in both directions and using the same primers as for the PCR. Subsequently, the sequence information was analyzed by the local alignment search tool using BLASTn [86,87] in the National Center for Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov/BLAST, accessed on 12 January 2022). Annotations were based on BLAST searches with a minimum of 99–100% identity over at least 80% of the length of the nucleotide sequence, which are the commonly used thresholds for reliable sequence annotation [88]. Nucleotide sequences primary identification was carried out through the BLASTn search tool program [86,87] of the NCBI by comparing all sequences obtained in this study with those already present in the database.
4.5. Sequences Alignments and Phylogenetic Analysis
All nucleotide sequences produced by this study and identified based on high sequence identity (>99–100%) to similar species already present in nucleotide databases, along with few additional reference sequences downloaded from GenBank (http://www.ncbi.nlm.nih.gov/GenBank, accessed on 12 January, 2022), were used for the phylogenetic analysis. Subsequently, they were manually edited and aligned with the ClustalX version 2.0 [88] program, using the MEGA X (Molecular Evolutionary Genetic Analysis) [89] phylogeny package to build representative alignments (Table 3). As reported in previous studies by Slippers et al. [66] and by Crous et al. [24], N. parvum and N. ribis are closely related cryptic species within the recently described Genus Neofusicoccum (Botryosphaeriaceae, Ascomycetes). Therefore, in case of the ACT gene, when no other reference species were available in the GenBank nucleotide database, this fungal species was also used, allowing us to perform the phylogenetic investigation (Table 3).
Phylogeny reconstructions were performed with MEGAX [89] for each gene using the neighbor-joining (NJ) statistical method [90] with an interior branch test and 1000 bootstrap replications [91,92], nucleotide substitution type and the Tajima–Nei substitution model [93] with uniform rates among sites. A deletion was used as treatment for gaps and missing data and the codon positions included were 1st, 2nd, 3rd and noncoding sites. The evolutionary distances computed using the Tajima–Nei method [93] are in the units of the number of base substitutions/site. The same procedure described previously was used for the tub-2 and ACT genes. An unequal number of nucleotide sequences were involved in the phylogenetic analyses for each gene investigated, which was caused by the lack of positive PCR and sequencing results for some of the genes and also the nonexistence of nucleotide sequences in the GenBank database. Therefore, only single gene phylogenies could be performed, each one containing all nucleotide sequences obtained in this study for the examined gene plus reference species downloaded from the GenBank.
In particular, the reference sequences representing the relevant species used to build alignments for species identification were: for D. eres and D. foeniculina (D. corylina strain CBS121124 and only for ACT gene D. helianthi strain AR4131), N. parvum (Diplodia seriata strain ASJ297; or Botryosphaeria dothidea strains HL1 and HPLW1 for the tub-2 and ACT genes, respectively), Pestalotiopsis sp. (Sordaria alcina strain CBS 109460), Phomopsis sp. (Valsa japonica isolate CBS375.29) and Trametes versicolor (Grifola frondosa isolate WC835) (Table 3).
A different number of nucleotide sequences were obtained for each gene and fungal species in this study; therefore, a multilocus phylogeny with three genes (ITS + tub-2 + ACT) was possible only for Diaporthe and Neofusicoccum spp., while for Pestalotiopsis spp. a two-gene phylogeny (ITS + tub-2) was performed using the Seaview5 program, as presented in Table S1 and in Figures S1–S3. (Supplementary material).
4.6. Pathogenicity Trials
A trial was conducted under field conditions to examine the formation of lesions on twigs of 2-year-old apple trees (cv. “Golden Delicious”), using a common protocol. In particular, the pathogenicity tests were performed using 4 mm diameter mycelial plugs taken from the margins of 7-day-old cultures on PDA amended with antibiotic streptomycin sulphate (40 mg L–1, MerckKGaA, Darmstadt, Germany). An equal number of young apple shoots were equally treated but using only sterile agar plugs, which were left as controls. One fungal isolate was used for each apple tree, according to the fungal species identified and characterized in this study, and each treatment was replicated four times. A wound of the about the size of the agar plug was made on each woody shoot, in the phloem and cortex tissue, with a sterile scalpel. Immediately after wounding, the plug was positioned in the center of the wound and covered by a sterile water wetted cotton piece. In order to avoid a rapid dehydration, lesion sites were wrapped with parafilm (Pechiney Plastic Packaging, Menasha, WI, USA). For each fungal pathogen the trial layout was a randomized block design with four repetitions using twigs as experimental units. The whole pathogenicity trial consisted of six fungal pathogens, isolated in this study from apple trees in Val d’Agri, and an agar plug only. Following inoculation, all young apple trees were placed in a greenhouse, where they were kept under natural light conditions at 22 °C and at about 70% relative humidity. After 30 days of inoculation, apple twigs were inspected for lesion development and after 45 days post-inoculation, when their necrosis was evident, the twigs were removed and brought to the laboratory for immediate analysis. The number of twigs with necrosis was recorded and, after the removal of the bark, the length of the developed canker lesions was measured. In order to reisolate the causal agent, small pieces (approx. 5 mm length) of diseased wooden tissue were cut from the edge of the necrotic lesions from the inoculated twigs and, after surface disinfection, were placed in petri dishes containing PDA and antibiotic streptomycin sulphate. Plates were incubated for 7 days at 20 °C in an incubator, under dark conditions, until growth was detected. Subsequently, the identification of the reisolated fungi was carried out by both morphological features and molecular analysis, using the protocols for morphological identification, DNA extraction and PCR conditions described above.
4.7. Statistical Analysis
Since the data obtained from the lesion measurements were normally distributed (Shapiro–Wilk tests [94], followed by a Holm–Bonfferoni [95] correction), a one-way ANOVA was used to test for mean differences among the investigated fungal isolates. Tukey post hoc tests for multiple comparisons of means were also performed to detect significant differences among the treatments. The statistical analyses performed in this study were performed using the R version 3.6.2 software (R Core Team, Vienna, Austria) [96].
5. Conclusions
Fungal species investigated in this study are well known to be involved in the fruit tree trunk diseases. The present study demonstrated that among all fungi investigated, N. parvum was the most aggressive and may be involved in the heavy decline of apple trees in the Val D’Agri area. In addition, other fungi, such as D. eres, D. foeniculina, P. funerea, T. versicolor and Phomopsis spp., could have contributed to the aggravation of the existing symptoms. Our field observations allowed us to assume that fungi, and in particular N. parvum, could penetrate the trees through wounds created by cuttings. Therefore, to avoid this, it is necessary to protect the wounds, in particular after cuttings.
Given the economic importance of apples worldwide, more investigations related to the role played by the phytopathogens discovered in this study, which are involved in die-back disease on apple trees, seem necessary. Future outcomes will be expected to add beneficial knowledge to better understand this complex disease in order to establish appropriate strategies to protect this regionally relevant and worldwide nutritionally important crop.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11101374/s1, Figure S1: PhyML tree (ITS + tub-2 + ACT) for Diaporthe spp.; Figure S2: PhyML tree (ITS + tub-2 + ACT) for Neofusicoccum spp.; Figure S3: PhyML tree (ITS + tub-2) for Pestalotiopsis spp.; Table S1: Multilocus phylogeny parameters used in the study.
Author Contributions
Conceptualization, S.M.M. and I.C.; methodology, S.M.M.; software, S.M.M. and A.M.; validation, S.M.M., C.M., A.M. and I.C.; formal analysis, S.M.M.; investigation, S.M.M.; resources, I.C.; data curation, S.M.M., I.C., C.M. and A.M.; writing—original draft preparation, S.M.M.; writing—review and editing, S.M.M., I.C., C.M. and A.M.; visualization, S.M.M., I.C., C.M. and A.M.; supervision, I.C. and S.M.M.; project administration, I.C. and S.M.M.; funding acquisition, I.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Basilicata Region-Phytosanitary Office (Matera, Italy), under the project entitled: “Epidemiological studies regarding the presence and spread in Basilicata of pathogens of agricultural and forestry plants with particular concern on those of quarantine. Molecular characterization of the pathogens and possibilities of fighting against them”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors declare that the data supporting the findings of this study are available within the article and in the supplementary materials (Figures S1–S3 and Table S1).
Acknowledgments
The authors thank Giuseppe Malvasi and Vincenzo Pucciariello (Phytosanitary Office of Matera, Department of Agricultural and Forestry Politics, Basilicata Region, Italy) for furnishing the information regarding the presence of the disease investigated in this study and for their valuable help with the field investigations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Doepel, R.F. Wood-rotting fungi associated with dieback in apple trees: Progress report: Orchard surveys and laboratory studies. J. Dep. Agric. West. Aust. 1962, 3, 668–678. Available online: https://researchlibrary.agric.wa.gov.au/journal_agriculture4/vol3/iss9/2 (accessed on 15 January 2022).
- Jeffers, S.N.; Aldwinckle, H.S.; Burr, T.J.; Arnerson, P.A. Phytophthora and Phytium species associated with crown rot in New York apple orchards. Phytopathology 1982, 72, 533–538. [Google Scholar] [CrossRef]
- Latorre, B.A.; Rioja, M.E.; Wilcox, W.F. Phytophthora species associated with crown rot of apple in Chile. Plant Dis. 2001, 85, 603–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, M.; Andrews, P.K.; Reganold, J.P.; Lévesque, A. Frequency, virulence, and metalaxyl sensitivity of Phytium spp. isolated from apple roots under conventional and organic production systems. Plant Dis. 2002, 86, 669–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Rusin, C.; Di Foggia, M.; Marceddu, S.; Rombolà, A.; Botelho, R.V.; Baraldi, E. Characterization of apples cultivars susceptibility to Neofusicoccum parvum Brazilian strains. Eur. J. Plant Pathol. 2020, 156, 939–951. [Google Scholar] [CrossRef]
- Cloete, M.; Fourie, P.H.; Damm, U.; Crous, P.W.; Moster, L. Fungi associated with die-back symptoms of apple and pear trees, a possible inoculum source of grapevine trunk disease pathogens. Phytopathol. Mediterr. 2011, 50, S176–S190. [Google Scholar] [CrossRef]
- Souli, M.; Paloma, A.-C.; Prez-Sierra, A.; Fattouch, S.; Armengol, J.; Boughalleb, N. Etiology of apple tree dieback in Tunisia and abiotic factors associated with the disease. Afric. J. Microbiol. Res. 2014, 8, 2272–2281. [Google Scholar] [CrossRef] [Green Version]
- ISTAT 2019. Coltivazioni Legnose Fruttifere. Available online: http://dati.istat.it (accessed on 15 January 2022).
- Lindner, L. Analisi diagnostiche. In Diagnostica Fitopatologica. Relazioni D’attività anni 2014–2006. Centro di Sperimentazione Laimburg, Bolzano (BZ), 77 Pag. (Diagnostic Analyses. Phytopathology Diagnostic. Activity Reports Years 2012–2006; Experimental Center Laimburg: Bolzano, Italy, 2014; 77p. [Google Scholar]
- Prodorutti, D.; Cainelli, C.; Gualandri, V.; Profaizer, D.; Dallago, G.; Branz, A.; Deliati, I.; Pertot, I.; Angeli, G. Moria e deperimento del melo in Trentino. Atti Giornate Fitopatol. 2012, 2, 619–621. [Google Scholar]
- Kienholz, J.R. Comparative study of the apple anthracnose and perennial canker fungi. J. Agric. Res. 1939, 59, 635–665. [Google Scholar]
- Pennycook, S.R.; Samuels, G.J. Botryosphaeria and Fusicoccum species associated with ripe fruit rot of Actinidia deliciosa (Kiwifruit) in New Zealand. Mycotaxon 1985, 24, 445–458. [Google Scholar]
- Hantula, J.; Lilja, A.; Nuorteva, H.; Parikka, P.; Werres, S. Pathogenicity, morphology and genetic variation of Phytophthora cactorum from strawberry, apple, rhododendron, and silver birch. Mycol. Res. 2000, 104, 1062–1068. [Google Scholar] [CrossRef]
- Petróczy, M.; Palkovics, L. First report of Monilia polystroma on apple in Hungary. Eur. J. Plant Pathol. 2009, 125, 343–347. [Google Scholar] [CrossRef]
- Hibett, D.S. Ribosomal RNA and fungal systematics. Trans. Mycol. Soc. Jpn. 1992, 33, 533–556. [Google Scholar]
- Arzanlou, M.; Bakhshi, M. ITS-rDNA sequences differentiate a new lineage of Diplodia associated with canker disease of apple in Iran. Plant Pathol. Quar. 2012, 2, 132–141. [Google Scholar] [CrossRef]
- Diaz, G.A.; Mostert, L.; Halleen, F.; Lolas, M.; Gutierrez, M.; Ferrada, E.; Latorre, B.A. Diplodia seriata Associated with Botrypshaeria Canker and Dieback in Apples Trees in Chile. Plant Dis. 2019, 103, 1025. [Google Scholar] [CrossRef]
- McKean, P.G.; Vaughan, S.; Gull, K. The extended tubulin superfamily. J. Cell Sci. 2001, 114, 2723–2733. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Phillips, A.J.L.; Crous, P.; Alves, A. Diaporthe species on Rosaceae with descriptions of D. pyracanthae sp. nov. and D. malorum sp. nov. Mycosphere 2017, 8, 485–511. [Google Scholar] [CrossRef]
- Pan, T.-T.; Xue, D.-S.; Li, B.-H.; Lian, S.; Wang, C.-X. Characterization of the pathogen causing shoot dieback on apple. Acta Phytopathol. Sinica 2020, 50, 107–111. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z.; Qin, G.; He, C.; Li, B.; Tian, S. Actin is required for cellular development and virulence of Botrytis cinerea via the mediation of secretory proteins. Msystems 2020, 5, e00732-19. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ding, Z.; Zhou, Z.Q.; Wang, Y.Z.; Guo, L.Y. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is Botryosphaeria dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Wennecker, M. Fungal Pathogens in Pome Fruit Orchards and Causal Agents of Postharvest Decay. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 25 February 2019; p. 200. [Google Scholar]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Phillips, A.; Alves, A.; Burgess, T.I.; Barber, P.A.; Groenewald, J.Z. Resolving phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Udayanga, D.; Liu, X.Z.; Crous, P.W.; McKenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, W. Diaporthe: A genus of endophitic, saprofitic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Yan, J.Y.; Li, X.H. The current status of species in Diaporthe. Mycosphere 2017, 8, 1106–1156. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Ryvarden, L. North American Polypores Vol. 2: Megasporoporia–Wrightoporia; Fungiflora: Oslo, Norway, 1987; pp. 437–885. [Google Scholar]
- Ryvarden, L.; Gilbertson, R.L. European Polypores: Part 2: Meripilus–Tyromyces; Fungiflora: Oslo, Norway, 1994; pp. 394–743. [Google Scholar]
- Justo, A.; Hibbett, D.S. Phylogenetic classification of Trametes (Basidiomycota, Polyporales) based on a five-marker dataset. Taxon 2011, 60, 1567–1583. [Google Scholar] [CrossRef]
- Welti, S.; Moreau, P.-A.; Favel, A.; Courtecuisse, R.; Haon, M.; Navarro, D.; Taussac, S.; Lesage-Meessen, L. Molecular phylogeny of Trametes and related genera, and description of a new genus Leiotrametes. Fungal Divers. 2012, 55, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Carlson, A.; Justo, A.; Hibbett, D.S. Species delimitation in Trametes: A comparison of ITS, RPB1, RPB2 and TEF1 Gene Phylogenies. Mycologia 2014, 106, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Summerell, B.A.; Swart, L.; Denman, S.; Taylor, J.E.; Beziudenhout, C.M.; Palm, M.E.; Mariconowitz, S.; Groenewald, J.Z. Fungal pathogens of Proteaceae. Persoonia 2011, 27, 20–45. [Google Scholar] [CrossRef] [Green Version]
- Maharachchikumbura, S.S.N.; Guo, L.-D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Kanematsu, S.; Kobayashi, T.; Kudo, A.; Ohtsu, Y. Conidial morphology, pathogenicity and culture characteristics of Phomopsis isolates from peach, Japanese pear and apple in Japan. Jpn. J. Phytopathol. 1999, 65, 264–273. [Google Scholar] [CrossRef]
- Santos, J.M.; Phillips, A.J.L. Resolving the complex of Diaporthe (Phomopsis) species occurring on Feoniculum vulgare in Portugal. Fungal Divers. 2009, 34, 111–125. [Google Scholar]
- Van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, J.M.; Groenewald, J.Z.; Farr, D.F.; Fourie, P.H.; Halleen, F.; Crous, P.W. Reassessment of Phomopsis species on grapevine. Australas. Plant Pathol. 2005, 34, 27–39. [Google Scholar] [CrossRef]
- Koenraadt, S.; Somerville, S.C.; Jones, A.L. Characterization of mutations in the beta-tubulin gene of benomyl-resistant field strains of Venturia inequalis and other plant pathogenic fungi. Phytopathology 1992, 82, 1348–1354. [Google Scholar] [CrossRef]
- Begerow, D.; Beate, J.; Oberwinkler, F. Evolutionary relationships among β-tubulin gene sequences of basidiomycetous fungi. Mycol. Res. 2004, 108, 1257–1263. [Google Scholar] [CrossRef]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Diver. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Mang, S.M.; Raciopi, R.; Camele, I.; Rana, G.L.; D’Auria, M. Use of metabolic profiles to distinguish three Monilinia species. Plant Ptahol. 2015, 97, 55–59. [Google Scholar] [CrossRef]
- Sofo, A.; Elshafie, H.S.; Scopa, A.; Mang, S.M.; Camele, I. Impact of airborne zinc polution on the antimicrobial activity of olive oil and teh microbial metabolic profiles of Z-contaminated soils in an Italian olive orchard. J. Trace Elem. Med. Biol. 2018, 49, 276–284. [Google Scholar] [CrossRef]
- Hofstetter, V.; Buyck, B.; Eyssartier, G.; Schnee, S.; Gindro, K. The unabearable lightness of sequence-based identification. Fungal Diver. 2019, 96, 243–284. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Jeewon, R.; Chomnunti, P.; Wanasinghe, D.N.; Norphanphoun, C.; Karunarathna, A.; Pem, D.; Perera, R.H.; Camporesi, E.; McKenzie, E.H.; et al. Taxonomic circumscription of Diaporthales based on multigene phylogeny and morphology. Fungal Divers. 2018, 93, 241–443. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Wibberg, D.; Stadler, M.; Lambert, C.; Bunk, B.; Spröer, C.; Rückert, C.; Kalinowski, J.; Cox, R.J.; Kuhnert, E. High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers. 2021, 106, 7–28. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium; Fungal Barcoding Consortium Author List; Bolchacova, E. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Kiss, L. Limits of nuclear ribosomal DNA internal transcribed spacer (ITS) sequences as species barcode for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, E1811. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.L.; Boal, R.J. A New Canker and Twig Dieback Disease of Apple and Crabapple Trees Caused by Sphaeropsis pyriputrescens in Washington State. Plant Dis. 2005, 89, 10. [Google Scholar] [CrossRef]
- Jabiri, S.; Lahlali, R.; Bahra, C.; Amraoui, M.B.; Tahiri, A.; Amiri, S. First report of Phytopythium vexans associated with dieback disease of apple trees in Morocco. J. Plant Pathol. 2020, 102, 1319. [Google Scholar] [CrossRef]
- Bai, Q.; Zhai, L.F.; Chen, X.R.; Hong, N.X.; Xu, W.X.; Wang, G.P. Biological and molecular characterization of five Phomopsis species associated with pear shoot canker in China. Plant Dis. 2015, 99, 1704–1712. Available online: http://apsjournals.apsnet.org/loi/pdis (accessed on 18 January 2021). [CrossRef] [Green Version]
- Bertetti, D.; Guarnaccia, V.; Spadaro, D.; Gullino, M. First report of fruit rot caused by Diaporthe eres Nitschkle on European pear cultivated in Italy. (Diaporthe eres Nitschkle, agente di marciume su frutti di pero in Italia). Prot. Delle Colt. 2018, 5, 10–13. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Zhang, W.; Yan, J.Y.; Li, X.H. Molecular phylogenetic analysis reveals seven new Diaporthe species from Italy. Mycosphere 2017, 8, 853–877. [Google Scholar] [CrossRef]
- Ali, S.; Renderos, W.; Bevis, E.; Hebb, J.; Abassi, P.A. Diaporthe eres causes stem cankers and death of young apple rootstocks in Canada. Can. J. Plant Pathol. 2020, 42, 217–227. [Google Scholar] [CrossRef]
- Florian, V.C.; Puia, C.; Groza, R.; Suciu, L.A.; Florian, T. Study of the major pathogens that lead to apple fruit decay during storage. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 538–545. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Huang, S.; Xia, J.; Zhang, X.; Li, Z. Morphological and molecular identification of Diaporthe species in south-western China, with description of eight new species. MycoKeys 2021, 77, 65–95. [Google Scholar] [CrossRef]
- Abramczyk, B.; Król, E.D.; Zalewska, E.D.; Zimowska, B. Morphological characteristics and pathogenicity of Diaporthe eres isolates to the fruit tree shoots. Acta Sci. Pol. -Hortorum Cultus 2018, 17, 125–133. [Google Scholar] [CrossRef]
- Chaisiri, C.; Liu, X.; Lin, Y.; Fu, Y.; Zhu, F.; Luo, C. Phylogenetic and Haplotype Network Analyses of Diaporthe eres Species in China Based on Sequences of Multiple Loci. Biology 2021, 10, 179. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Bettucci, L.; Lupo, S. Diversity and virulence of Diaporthe species associated with wood diseases symptoms in deciduous fruit trees in Uruguay. Phytopathol. Mediterr. 2017, 56, 431–444. [Google Scholar] [CrossRef]
- Thompson, S.M.; Tan, Y.P.; Young, A.J.; Neate, S.M.; Aitken, E.A.B.; Shivas, R.G. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Pers. Molec. Phylogen. Evol. Fungi 2011, 27, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Smit, W.A.; Crous, P.W.; Countinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Taxonomy, phylogeny and identification of Botryosphaeriaceace associated with pome and stone fruit trees in South Africa and other regions of the world. Plant Pathol. 2007, 56, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.D.; Tang, G.H.; Peng, S.B.; Chen, H.; Zhai, M.Z. Neofusicoccum parvum causing canker of seedlings of Juglans regia in China. J. For. Res. 2015, 26, 1019–1024. [Google Scholar] [CrossRef]
- Shen, Y.-M.; Chao, C.-H.; Liu, H.L. First Report of Neofusicoccum parvum Associated with Stem Canker and Dieback of Asian Pear Trees in Taiwan. Plant Dis. 2010, 94, 1062. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Paul, N.C.; Lee, K.-H.; Kim, H.-J.; Sang, H. Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii). Forests 2021, 12, 440. [Google Scholar] [CrossRef]
- Zmitrovich, I.V.; Ezhov, O.N.; Wasser, S.P. A survey of species of genus Trametes Fr. (higher basidiomycetes) with estimation of their medicinal source potential. Int. J. Med. Mushrooms 2012, 14, 307–319. [Google Scholar] [CrossRef]
- Habtermariam, S. Trametes versicolor (Synn. Coriolus versicolor) Polysaccharides in Cancer Therapy: Targets and Efficiency. Biomedicines 2020, 8, 135. [Google Scholar] [CrossRef]
- Pop, R.M.; Puia, I.C.; Puia, A.; Chedea, V.S.; Leopold, N.; Bocsan, I.C.; Buzoianu, A.D. Characterization of Trametes versicolor: Medicinal Mushroom with Important Health Benefits. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Kile, G. The Host-Pathogen Relationship between Apple and Trametes versicolor and Factors Affecting Host Susceptibility. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 1971. [Google Scholar]
- Darbyshire, B. Factors Influencing the Susceptibility of Apple Trees to Trametes (Polystictus) versicolor. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 1968. [Google Scholar]
- Dilley, M.A.; Covey, R.P., Jr. Association of Coriolus versicolor with a dieback disease of apples in Washington State. Plant Dis. 1981, 85, 77–78. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.Z.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, L.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Karaoglanidi, G.; Bardas, G. First Report of Phomopsis Fruit Decay on Apple Caused by Phomopsis mali in Greece. Plant Dis. 2006, 90, 375. [Google Scholar] [CrossRef]
- Abramczyk, B.; Król, E. Use of RAPD-PCR and its markers for identification of Diaporthe/Phomopsis from fruit trees in south-eastern Poland. Acta Sci. Pol. -Hortorum Cultus 2016, 15, 161–175. Available online: http://ww.acta.media.pl/pl/action/getfullphp?id=5213 (accessed on 8 January 2020).
- Pertot, I.; Vindimian, M.E. Diffusione di Phomopsis mali, agente di moria in giovani impianti di melo in Trentino. Inform. Fitopatol. 1998, 48, 45–49. Available online: http://hdl.handle.net/10449/17658 (accessed on 8 January 2020).
- Espinoza, J.G.; Briceño, E.X.; Chávez, E.R.; Úrbez-Torrez, J.R.; Latorre, B.A. Neofusicoccum spp. Associated with Stem Canker and Dieback of Blueberry in Chile. Plant Dis. 2009, 93, 1187–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaad, N.W.; Jones, J.B.; Chun, W. Laboratory Guide for the Identification of Plant Pathogenic Bacteria, 3rd ed.; The American Phytopathological Society: St. Paul, MN, USA, 2001; 373p. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplified and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Mang, S.M.; Bonomo, M.G.; Camele, I.; Saturnino, C.; Capasso, A.; Salzano, G. Preliminary Investigation of Allergenic Fungi in an Indoor Environment. Pharmacologyonline 2020, 2, 314–320. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, J.D. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; Mcginnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucl. Acids Res. 2008, 36 (Suppl. 2), W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetic Analysis across computing platforms. Molec. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Dopazo, J. Estimating errors and confidence intervals for branch lengths in phylogenetic trees by a bootstrap approach. J. Mol. Evol. 1994, 38, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F.; Nei, M. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1984, 1, 269–285. [Google Scholar] [PubMed] [Green Version]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, 2019; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 15 January 2022).
Figure 1.
Pure fungal cultures on PDA obtained from samples of apples with dieback symptoms. (a,g) = Trametes versicolor; (b,h) = Diaporthe eres; (c,i) = Diaporthe feoniculina; (d,j) = Pestalotiopsis funerea; (e,k) = Phomopsis spp.; (f,l) = Neofusicoccum parvum.
Figure 1.
Pure fungal cultures on PDA obtained from samples of apples with dieback symptoms. (a,g) = Trametes versicolor; (b,h) = Diaporthe eres; (c,i) = Diaporthe feoniculina; (d,j) = Pestalotiopsis funerea; (e,k) = Phomopsis spp.; (f,l) = Neofusicoccum parvum.

Figure 2.
Molecular phylogenetic tree obtained through the neighbor-joining (NJ) method, based on the 58 ITS region sequences data (658 bp) from fungal isolates in the present study and published sequences. Five fungal species (Diaporthella corylina, Valsa japonica, Sordaria alcina, Grifola fondosa and Diplodia seriata) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 1.38532669 is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.
Figure 2.
Molecular phylogenetic tree obtained through the neighbor-joining (NJ) method, based on the 58 ITS region sequences data (658 bp) from fungal isolates in the present study and published sequences. Five fungal species (Diaporthella corylina, Valsa japonica, Sordaria alcina, Grifola fondosa and Diplodia seriata) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 1.38532669 is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.

Figure 3.
Molecular phylogenetic tree obtained through the neighbor-joining (NJ) method, based on the 39 tub-2 gene sequences data (500 bp) from fungal isolates in the present study and published sequences. The fungal species (D. corylina, B. dothidea and S. alcina) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 1.08594872 is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.
Figure 3.
Molecular phylogenetic tree obtained through the neighbor-joining (NJ) method, based on the 39 tub-2 gene sequences data (500 bp) from fungal isolates in the present study and published sequences. The fungal species (D. corylina, B. dothidea and S. alcina) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 1.08594872 is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.

Figure 4.
Molecular phylogenetic tree obtained through neighbor-joining (NJ) method based on the 24 ACT gene sequences data (298 bp) from fungal isolates in the present study and published sequences. The fungal species (D. helianthi and B. dothidea) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 0.90526841is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.
Figure 4.
Molecular phylogenetic tree obtained through neighbor-joining (NJ) method based on the 24 ACT gene sequences data (298 bp) from fungal isolates in the present study and published sequences. The fungal species (D. helianthi and B. dothidea) were used as outgroups in the analysis. The optimal tree with the sum of branch length = 0.90526841is shown. The confidence probability estimated using the bootstrap test (1000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tajima–Nei method and are in the units of the number of base substitutions/site. Scientific names of the fungi along with collection place, isolate abbreviation and GenBank AC number are shown in the trees.

Figure 5.
Results of artificial inoculation of the apple twigs with the six fungal isolates investigated. The length of the lesions developed was measured 1 month after inoculation. The experiment was carried out only once with three replications (twigs) and four wounds per replicate. Columns indicate the average length of the lesions with standard errors. Means followed by different letters are significantly different according to Tukey’s test (p = 0.01).
Figure 5.
Results of artificial inoculation of the apple twigs with the six fungal isolates investigated. The length of the lesions developed was measured 1 month after inoculation. The experiment was carried out only once with three replications (twigs) and four wounds per replicate. Columns indicate the average length of the lesions with standard errors. Means followed by different letters are significantly different according to Tukey’s test (p = 0.01).

Figure 6.
Apple trees located in the Val d’Agri area showing die-back symptoms (a–c) and death of tree caused by die-back (d). Figure a—courtesy of Dr. Camilla Nigro, ALSIA, Basilicata Region.
Figure 6.
Apple trees located in the Val d’Agri area showing die-back symptoms (a–c) and death of tree caused by die-back (d). Figure a—courtesy of Dr. Camilla Nigro, ALSIA, Basilicata Region.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fungal isolates obtained during this study with their respective GenBank accession numbers and percentage of identity when compared to reference nucleotide sequences for the same species from the NCBI nucleotide database.
Table 1.
Fungal isolates obtained during this study with their respective GenBank accession numbers and percentage of identity when compared to reference nucleotide sequences for the same species from the NCBI nucleotide database.
| Isolate Name | Species | GenBank Accession Number * | Identity (%) * | ||||
|---|---|---|---|---|---|---|---|
| ITS ** | TUB-2 ** | ACT ** | ITS ** | TUB-2 ** | ACT ** | ||
| Noefp1 | Neofusicoccum parvum | LR757960 | OU022063 | OU023206 | >99–100 | >99 | 100 |
| Neofp2 | N. parvum | LR757961 | OU022064 | OU023207 | -//- | -//- | -//- |
| Neofp3 | N. parvum | LR757962 | OU022065 | OU023208 | -//- | -//- | -//- |
| Neofp4 | N. parvum | LR757963 | OU022066 | OU023209 | -//- | -//- | -//- |
| Neofp5 | N. parvum | LR757964 | OU022067 | OU023210 | -//- | -//- | -//- |
| Neofp6 | N. parvum | LR757965 | OU022068 | OU023211 | -//- | -//- | -//- |
| Neofp7 | N. parvum | LR757966 | OU022069 | OU023212 | -//- | -//- | -//- |
| Tramtv1 | Trametes versicolor | LR759930 | - | - | >99–100 | - | - |
| Tramtv2 | T. versicolor | LR759931 | - | - | -//- | - | - |
| Tramtv3 | T. versicolor | LR759932 | - | - | -//- | - | - |
| Tramtv4 | T. versicolor | LR759933 | - | - | -//- | - | - |
| Tramtv5 | T. versicolor | LR759934 | - | - | -//- | - | - |
| Tramtv6 | T. versicolor | LR759935 | - | - | -//- | - | - |
| Diapore1 | Diaporthe eres | OU020696 | OU022056 | OU023199 | >99 | >99–100 | >99 |
| Diapore2 | D. eres | OU020697 | OU022057 | OU023200 | -//- | -//- | -//- |
| Diapore3 | D. eres | OU020698 | OU022058 | OU023201 | -//- | -//- | -//- |
| Diapore4 | D. eres | OU020699 | OU022059 | OU023202 | -//- | -//- | -//- |
| Diaporf1 | Diaporthe foeniculina | OU020700 | OU022060 | OU023203 | >99–100 | 100 | >99 |
| Diaporf2 | D. foeniculina | OU020701 | OU022061 | OU023204 | -//- | -//- | -//- |
| Diaporf3 | D. foeniculina | OU020702 | OU022062 | OU023205 | -//- | -//- | -//- |
| Pestf1 | Pestalotiopsis funerea | OU020703 | OU022070 | - | >99 | >99 | - |
| Pestf2 | P. funerea | OU020704 | OU022071 | - | -//- | -//- | - |
| Pestf3 | P. funerea | OU020705 | OU022072 | - | -//- | -//- | - |
| Phomp1 | Phomopsis sp. | OU026160 | - | - | >99 | - | - |
| Phomp2 | Phomopsis sp. | OU026161 | - | - | -//- | - | - |
| Phomp3 | Phomopsis sp. | OU026162 | - | - | -//- | - | - |
Note: * The percentage of identity was established after comparing the nucleotide sequences from this study with at least two of the reference species existent in the database for each fungal species. ** ITS = Nuclear ribosomal internal transcribed spacer regions; TUB-2 = β-tubulin 2 gene; ACT = actin gene. “-” = no data were obtained/or exist in the GenBank nucleotide database. “-//-” = identical values as those in the previous row are reported.
Table 2.
Details of primers pairs used in this study for the amplification and sequencing of fungal DNA.
Table 2.
Details of primers pairs used in this study for the amplification and sequencing of fungal DNA.
| Locus * | Primer | Sequences 5′→3′ | Reference |
|---|---|---|---|
| ITS | ITS5 | 5′-GGA AGT AAA AGT CGT AAC AAG G-3′ | White et al., 1990 |
| ITS4 | 5′-TCC TCC GCT TAT TGA TAT GC-3′ | ||
| TUB-2 | Bt2a | 5′-GGT AAC CAA ATC GGT GCT GCT TTC-3′ | Glass and Donaldson, 1995 |
| Bt2b | 5′-ACC CTC AGT GTA GTG ACC CTT GGC-3′ | ||
| ACT | ACT-512F | 5′-ATG TGC AAG GCC GGT TTC GC-3′ | Carbone and Kohn, 1999 |
| ACT-783R | 5′-TAC GAG TCC TTC TGG CCC AT-3′ |
* ITS: internal transcribed spacer regions and intervening 5.8S rRNA gene; TUB-2: partial beta tubulin gene; ACT: actin gene.
Table 3.
List of taxa, fungal isolates and GenBank accession numbers of the genes analyzed in this study and used for phylogenetic analysis.
Table 3.
List of taxa, fungal isolates and GenBank accession numbers of the genes analyzed in this study and used for phylogenetic analysis.
| Taxa | Isolate Name | Culture No. | Gene | GenBank Accession Number * | Reference | ||
|---|---|---|---|---|---|---|---|
| ITS | TUB-2 | ACT | ITS | TUB-2 | ACT | ||
| Diaporthe eres | CBS 186.37 | CPC 30116 | CBS:145040 | MH855881 | MG281261 | MK442634 | Wu et al., 2019 |
| D. eres | SS48 | NEFF 3-23-4 | Pho12 | KP903620 | MW208555 | JN230370 | GenBank |
| D. eres | STEU 8322 | DB14AGO27 | MIFCC 316 | KY312645 | MH063919 | MN136112 | GenBank |
| D. eres | STEU 8323 | Nc1 | CBS:587.79 | KY312646 | LC316667 | KJ420770 | GenBank |
| D. eres | Fi2333 | MJL13 | DNP128 | KR023623 | MT109632 | KJ420762 | GenBank |
| Diaporthe foeniculina | ISPaVe 2156 | ColPat-560 | ISPaVe 2156 | LN651172 | MK522116 | LN651174 | GenBank a Lopez-Moral et al., 2020 |
| D. foeniculina | A1907B | Av-1 | ISPaVe 2157 | MT230444 | MT374093 | LN651175 | GenBank c Udayanga et al., 2014 |
| D. foeniculina | P101b | CAA133 | MEP12891 | MT735646 | KY435665 | KC843283 | GenBank a Mathioudakis et al., 2020 c Udayanga et al., 2014 |
| ** Diaporthella corylina | CBS121124 | CBS121124 | CBS:592.81 | KC343004 | KC343972 | N/A | Gomes et al., 2013 |
| ** Diaporthe helianthi | N/A | N/A | AR4131 | N/A | N/A | KF199885 | c GenBank |
| Neofusicoccum parvum | B32 | ACBA15 | YELO-21a | KJ499738 | MG970291 | MH393619 | GenBank |
| N. parvum | B65 | AKKA308.2 | CMW 7773 | KJ499740 | MH221123 | - e | GenBank c Hunter et al.,2006 |
| d N. ribis | - e | - e | CMW 7773 | - e | - e | DQ267605 | GenBank |
| N. parvum | B135 | 11215_3 | N/A | KJ499742 | JX398944 | N/A | GenBank |
| N. parvum | B146 | MFLUCC_12-0380 | N/A | KJ499743 | MN643160 | N/A | GenBank |
| N. parvum | Fi2326 | GDTCMF23 | N/A | KR002830 | MT424811 | N/A | GenBank |
| N. parvum | JRad16 | CBS:130994 | N/A | KY680281 | a MT529709 | N/A | GenBank a Zhang et al., 2021 |
| N. parvum | CMW9081; ICMP 8003 | CMW994 | N/A | NR119487 | a AY236912 | N/A | GenBank a Slippers et al., 2004 |
| ** Diplodia seriata | ASJ297 | HL1 | N/A | MH221102 | JF4040814 | N/A | GenBank |
| b Botryosphaeria dothidea | N/A | N/A | HPLW1 | N/A | N/A | JF440940 | a Tang et al., 2012 |
| Pestalotiopsis funerea | ML4DY | ML4DY | N/A | EF055197 | EF055234 | N/A | GenBank |
| Pestalotiopsis sp. | SGSGf16 | TAP18N030 | N/A | EU715650 | LC427211 | N/A | GenBank |
| Pestalotiopsis sp. | ZC-W-1-1 | BRIP 66615 | N/A | KR822153 | MK977634 | N/A | GenBank |
| Pestalotiopsis sp. | NW-FWA2867 | ZX18A | N/A | MG098325 | MW218534 | N/A | GenBank |
| ** Sordaria alcina | CBS 109460 | CBS 109460 | N/A | AY681198 | a AY681232 | N/A | Liu et al., 2010 a Cai et al., 2006 |
| Phomopsis sp. | FN-1-N1-2-2 | N/A | N/A | KJ465317 | N/A | N/A | GenBank |
| Phomopsis sp. | blx-2-51 | N/A | N/A | MN944543 | N/A | N/A | GenBank |
| Phomopsis sp. | Peach | N/A | N/A | MK934328 | N/A | N/A | GenBank |
| Phomopsis sp. | 136.1-COLP-L | N/A | N/A | KT182885 | N/A | N/A | GenBank |
| **Valsa japonica | CBS375.29 | N/A | N/A | AF191185 | N/A | N/A | Adams et al., 2002 |
| Trametes versicolor | D. HaelewF-1599pt.2a | N/A | N/A | MN749366 | N/A | N/A | GenBank |
| T. versicolor | STEU 8295 | N/A | N/A | KY312629 | N/A | N/A | GenBank |
| T. versicolor | STEU 8296 | N/A | N/A | KY312630 | N/A | N/A | GenBank |
| T. versicolor | 2473 | N/A | N/A | AM269814 | N/A | N/A | GenBank |
| ** Grifola frondosa | WC835 | N/A | N/A | AY049140 | N/A | N/A | Shen et al., 2002 |
Notes: * ITS-Nuclear ribosomal internal transcribed spacer regions; TUB- (tub-2) gene; ACT-actin gene. ** This fungal species has been used as outgroup for the phylogenetic analysis. The sign “-” in the column table indicates either that the nucleotide sequences do not exist in the GenBank or were not obtained in this study for that particular fungal isolate. a This study was considered only for the tub-2 nucleotide gene sequence. b This fungal species was used only for the phylogenetic analysis involving the tub-2 gene nucleotide sequences. c This GenBank origin of the accession regards only ACT gene nucleotide sequences used for the phylogenetic analysis. d This fungal species was used only for the phylogenetic analysis involving the ACT gene. N/A: not available.e Data not considered.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mang, S.M.; Marcone, C.; Maxim, A.; Camele, I. Investigations on Fungi Isolated from Apple Trees with Die-Back Symptoms from Basilicata Region (Southern Italy). Plants 2022, 11, 1374. https://doi.org/10.3390/plants11101374
AMA Style
Mang SM, Marcone C, Maxim A, Camele I. Investigations on Fungi Isolated from Apple Trees with Die-Back Symptoms from Basilicata Region (Southern Italy). Plants. 2022; 11(10):1374. https://doi.org/10.3390/plants11101374
Chicago/Turabian StyleMang, Stefania Mirela, Carmine Marcone, Aurel Maxim, and Ippolito Camele. 2022. "Investigations on Fungi Isolated from Apple Trees with Die-Back Symptoms from Basilicata Region (Southern Italy)" Plants 11, no. 10: 1374. https://doi.org/10.3390/plants11101374
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.