
A Revised Phylogenetic Classification for Viola (Violaceae)
by
 , , , and
, , , and
Thomas Marcussen
1,*,
Harvey E. Ballard
2,
Jiří Danihelka
3,4,
Ana R. Flores
5,
Marcela V. Nicola
6 and
John M. Watson
5 1
Department of Biosciences, Centre for Ecological and Evolutionary Synthesis (CEES), University of Oslo, P.O. Box 1066 Blindern, NO-0316 Oslo, Norway
2
Department of Environmental and Plant Biology, Ohio University, Athens, OH 45701, USA
3
Department of Botany and Zoology, Masaryk University, Kotlářská 2, CZ-61137 Brno, Czech Republic
4
Czech Academy of Sciences, Institute of Botany, Zámek 1, CZ-252 43 Průhonice, Czech Republic
5
Independent Researcher, Casilla 161, Los Andes 2100412, Chile
6
Instituto de Botánica Darwinion (IBODA, CONICET-ANCEFN), Labardén 200, Casilla de Correo 22, San Isidro, Buenos Aires B1642HYD, Argentina
*
Author to whom correspondence should be addressed.
Plants 2022, 11(17), 2224; https://doi.org/10.3390/plants11172224
Submission received: 10 July 2022
/
Revised: 15 August 2022
/
Accepted: 16 August 2022
/
Published: 27 August 2022
(This article belongs to the Special Issue Advances in Violaceae Research)
Abstract
:The genus Viola (Violaceae) is among the 40–50 largest genera among angiosperms, yet its taxonomy has not been revised for nearly a century. In the most recent revision, by Wilhelm Becker in 1925, the then-known 400 species were distributed among 14 sections and numerous unranked groups. Here, we provide an updated, comprehensive classification of the genus, based on data from phylogeny, morphology, chromosome counts, and ploidy, and based on modern principles of monophyly. The revision is presented as an annotated global checklist of accepted species of Viola, an updated multigene phylogenetic network and an ITS phylogeny with denser taxon sampling, a brief summary of the taxonomic changes from Becker’s classification and their justification, a morphological binary key to the accepted subgenera, sections and subsections, and an account of each infrageneric subdivision with justifications for delimitation and rank including a description, a list of apomorphies, molecular phylogenies where possible or relevant, a distribution map, and a list of included species. We distribute the 664 species accepted by us into 2 subgenera, 31 sections, and 20 subsections. We erect one new subgenus of Viola (subg. Neoandinium, a replacement name for the illegitimate subg. Andinium), six new sections (sect. Abyssinium, sect. Himalayum, sect. Melvio, sect. Nematocaulon, sect. Spathulidium, sect. Xanthidium), and seven new subsections (subsect. Australasiaticae, subsect. Bulbosae, subsect. Clausenianae, subsect. Cleistogamae, subsect. Dispares, subsect. Formosanae, subsect. Pseudorupestres). Evolution within the genus is discussed in light of biogeography, the fossil record, morphology, and particular traits. Viola is among very few temperate and widespread genera that originated in South America. The biggest identified knowledge gaps for Viola concern the South American taxa, for which basic knowledge from phylogeny, chromosome counts, and fossil data is virtually absent. Viola has also never been subject to comprehensive anatomical study. Studies into seed anatomy and morphology are required to understand the fossil record of the genus.
Keywords:
Viola; Violaceae; taxonomic revision; nomenclature; fossils; morphology; phylogeny; monophyletic; polyploidy1. Introduction
Viola L. is one of the largest angiosperm genera but has not been subject to taxonomic revision for nearly a century [1]. The genus comprises violets and pansies and is one of two temperate genera in the otherwise neotropical Violaceae Batsch family [2,3,4], besides Cubelium Raf. ex Britton & A. Br. for C. concolor (T. F. Forst.) Raf. ex Britton & A. Br. With its c. 664 species, Viola is the largest genus in the family, the fourth largest within Malpighiales (after Euphorbia with 2400 species, Croton with at least 1300 species, and Phyllanthus with 1200 species [5]) and among the 40–50 largest among angiosperms, despite not being among the genera listed by Frodin [6]. Viola is one of very few Malpighiales genera with large radiations in the temperate zone, next to Hypericum L., Linum L., Salix L. and Populus L.
Violets and pansies are well-known plants and have a long history in European folklore and the first records describing the use of Viola in Europe are from Ancient Greece [7]. Fragrant violets were sold in the Athenian agora, praised by Greek poets, such as in the writings of Sappho, used in medicine, had an active role in myths, such as in the abduction of Persephone, were used in garlands, and were present in The Odyssey’s garden of Calypso [7]. Viola continued to be used throughout the Middle Ages and species like V. odorata (Figure 1), V. elatior, and V. tricolor were described as medicinal plants in early modern period herbals (e.g., Matthiolus 1562 [8]). In Renaissance paintings and Christian traditions, violets were commonly associated with the Virgin Mary and had a symbolic meaning connected with humility [7].
Dried flowering shoots of Viola arvensis and V. tricolor are included in the European Pharmacopoeia as Violae tricoloris herba cum flore [9]. They are used as comminuted herbal substances for infusions for cutaneous and internal use, mainly in the treatment of various skin disorders. Viola and Violaceae in general are rich in cyclotides, a family of cyclic plant peptides involved in host defence (e.g., [10,11,12]). Given the chemical stability of the cyclotide framework, there is interest in using these peptides as scaffolds in drug design [13], and many species of Viola have been screened (e.g., [10,14,15]). Viola odorata (Figure 1) in particular has been cultivated for the production of essential oil for the perfume industry [16,17] but nowadays the fragrant compound, ionone, is usually synthesised chemically or endogenously from β-carotene [18]. From the leaves of the same species, absolutes with scent with floral and green notes, reminiscent of cucumber, are extracted and used in the perfume industry [17]. Several species of Viola are grown as ornamentals, such as the pansy hybrids V. ×williamsii and V. ×wittrockiana [19], and certain cultivars of V. sororia, V. palmata and V. prionantha for their floral display. Others are grown for their colourful or variegated decorative foliage, such as V. variegata and V. riviniana f. purpurea (often as V. labradorica hort. non Schrank). Some are grown for their fragrant flowers, such as V. odorata, filled forms of V. alba subsp. dehnhardtii known as ‘Parma violets’ [7,20], and V. suavis [21,22]. Pansy flowers have been used as garnishes on salads and cakes. Since ancient times the petals of blue- or purple-flowered species have been used to make syrups and jellies, and the young leaves of various species have been boiled as a vegetable [23]. Viola sororia is the state flower of the USA states of Illinois, Rhode Island, New Jersey, and Wisconsin. In Canada, V. cucullata is the provincial flower of New Brunswick. In the United Kingdom, V. riviniana is the county flower of Lincolnshire.
All phylogenetic studies to date have recovered Viola as monophyletic [3,4,24]. Unlike most other genera in the family, Viola is usually herbaceous and with a temperate distribution and is defined by several apomorphies with few exceptions, including the non-articulated peduncles (i.e., lacking an abscission zone at the level of bracteoles), solitary flowers, calycine appendages, bottom petal that is distinctly spurred (rarely scarcely saccate or gibbose), and with the blade shorter than to not much longer than the lateral and upper petals [25]. The spurred bottom petal is a shared feature with its sister lineage, the monotypic shrubby genera Noisettia and Schweiggeria, but this character is not unique within the family [3,25]. Cleistogamy is widespread and common in the genus (as well as in the family), and many of the lineages in the northern hemisphere have evolved seasonal cleistogamy [26,27].
Viola is distributed in most ice-free regions of the world except Antarctica, mainly in the temperate zones of both hemispheres and at high elevations in the mountain systems of the tropics [2,28] (Figure 2). The genus has its centres of taxonomic and morphological diversity in the Andes, in the Mediterranean area of Europe, in eastern Asia, and in North America. Three species, i.e., V. biflora, V. suecica, and V. selkirkii, have nearly circumboreal distributions. Viola rostrata is disjunctly distributed in Japan and eastern North America. Viola palustris is Amphiatlantic. Viola arvensis, V. odorata, and V. tricolor are near cosmopolites as a result of introductions.
Viola, like Violaceae as a whole, is assumed to have originated in South America [2,4,28,29]. Dating analysis associates the origin and beginning diversification of Viola with the Eocene-Oligocene cooling event [30,31,32] which, in combination with the formation of the Andes during the Eocene [33,34,35,36], may have given this temperate lineage opportunities to diversify [4,28].
An inherent feature of Viola is the lack of barriers against hybridisation, which occurs commonly between closely related species, especially in disturbed or transitional habitats, and which can make species identification difficult [37,38,39,40]. Speciation by allopolyploidisation, which occurs as a consequence of genome duplication in a hybrid, has been estimated to occur with a higher proportion in Viola (67% to 88% [28]) than in angiosperms in general (15% to 30% [41,42]). It is therefore no coincidence that the first polyploid series of chromosome numbers was discovered in Viola, with n = 6, 12, 18, 24, 36, 48 (Miyaji 1913 [43,44]). Allopolyploidisation has been instrumental in at least three major radiations within the genus, i.e., the first following dispersal into the northern hemisphere 18–20 Ma ago and the associated diversification into at least nine allopolyploid endemic lineages [28], the second following dispersal into North America c. 10 Ma ago and formation of the endemic allodecaploid sect. Nosphinium [45], and the third since c. 10 Ma within sect. Melanium in the western Palearctic [28].
The first taxonomic treatments of Viola were contributed by Frédéric C. J. Gingins de la Sarraz (1790–1873) in 1823 [46] and in the chapter on Violarieae in de Candolle’s Prodromus in 1824 [47]. Gingins realised that the shape of the style was a variable and reliable character to subdivide the genus, and based on that he grouped the 105 species known at the time into five sections, sect. Nomimium (=sect. Viola), sect. Dischidium, sect. Chamaemelanium, sect. Melanium, and sect. Leptidium. All but the last section covered the northern hemisphere taxa.
By the end of the 19th century, the number of known Viola species had doubled to 200. The treatment of Viola by Karl Reiche (1860–1922) for the first edition of Engler & Prantl’s Die Natürlichen Pflanzenfamilien [48] was the first to take into account the morphological distinction of the rosulate violets of South America (subg. Neoandinium in our circumscription). Reiche placed them in sect. Rosulatae, while uniting all of Gingins’ sections in sect. Sparsifoliae (subg. Viola in our circumscription). In addition, he erected sect. Confertae for five morphologically deviating species of both subgenera.
The treatment of Viola by Wilhelm Becker (1874–1928) for the second edition of Engler’s Die Natürlichen Pflanzenfamilien in 1925 [1] represented a leap forward in the understanding and classification of the genus, for which c. 400 species were known at the time. Summarising more than two decades of his taxonomic work on Viola, Becker recognised a total of 14 sections based on general morphology and biogeography, including five of Gingins’s [46] but, for some reason, none of Reiche’s [48]. Hence, Becker erected sect. Delphiniopsis, sect. Nosphinium, sect. Sclerosium, and sect. Xylinosium for northern hemisphere taxa, sect. Andinium (an illegitimate name for Reiche’s sect. Rosulatae), sect. Chilenium, sect. Rubellium, and sect. Tridens for South American taxa, and sect. Erpetion for the Australian taxa. In addition, he noted the need for additional sections to accommodate a few more, divergent species not included in his system, namely V. abyssinica and relatives in Africa, V. filicaulis in New Zealand, and V. papuana in New Guinea. Notably, Becker subdivided the large and heterogeneous sect. Nomimium (=sect. Viola) into a total of 17 unranked greges, denoted A through R, many of which have since been combined at the subsection or section level.
The treatment of Viola by Wilhelm Becker (1874–1928) for the second edition of Engler’s Die Natürlichen Pflanzenfamilien in 1925 [1] represented a leap forward in the understanding and classification of the genus, for which c. 400 species were known at the time. Summarising more than two decades of his taxonomic work on Viola, Becker recognised a total of 14 sections based on general morphology and biogeography, including five of Gingins’s [46] but, for some reason, none of Reiche’s [48]. Hence, Becker erected sect. Delphiniopsis, sect. Nosphinium, sect. Sclerosium, and sect. Xylinosium for northern hemisphere taxa, sect. Andinium (an illegitimate name for Reiche’s sect. Rosulatae), sect. Chilenium, sect. Rubellium, and sect. Tridens for South American taxa, and sect. Erpetion for the Australian taxa. In addition, he noted the need for additional sections to accommodate a few more, divergent species not included in his system, namely V. abyssinica and relatives in Africa, V. filicaulis in New Zealand, and V. papuana in New Guinea. Notably, Becker subdivided the large and heterogeneous sect. Nomimium (=sect. Viola) into a total of 17 unranked greges, denoted A through R, many of which have since been combined at the subsection or section level.
Becker’s taxonomic treatment from 1925 [1] remains the last comprehensive taxonomic treatment of Viola. Although comprehensive, it was only a summary, with very short descriptions of infrageneric taxa only and incomplete lists of taxa. Becker probably considered this treatment provisional, as it is known that he was working on a monograph of the genus when he died after a short illness in 1928, aged only 54 [49,50]. His notes were lost and never published. His Viola herbarium, containing approximately 4300 specimens and acquired by the Herbarium berolinense (B) in 1929, was destroyed by a fire in early March 1943 after a bombing by Allied forces [51,52]. These unfortunate events, along with the mere size of Viola which renders the genus difficult to study in its entirety, are likely reasons why Viola has not been subject to full revision in nearly a century.
In the late 1920s and early 1930s, numerous studies on chromosome cytology were published on Viola in the northern hemisphere [29,43,44,53,54,55,56]. Based on these findings, along with observations on general morphology, biogeography, and crossing experiments [57,58], Jens C. Clausen (1891–1969) suggested two considerable changes to Becker’s system [29,56,59]. The first was introducing the concept of a widely defined sect. Chamaemelanium that united all yellow-flowered taxa having the base chromosome number x = 6, i.e., including sect. Dischidium and greges Orbiculares and Memorabiles of sect. Nomimium. The second change was splitting in two the large and heterogeneous sect. Nomimium, i.e., into sect. Plagiostigma, having a margined style and the base chromosome number x = 12, and sect. Rostellatae (=sect. Viola), having an unmargined, rostellate style and x = 10. Although this subdivision was backed up by substantial evidence and later also confirmed phylogenetically, Clausen’s revision was not implemented in any treatment of the genus for the next 90 years [2,28,45,60,61].
Only a few monographs have been published dealing comprehensively with particular groups, i.e., on sect. Chilenium [62,63], sect. Melanium [64,65,66], subsect. Borealiamericanae [67], and most recently on subg. Neoandinium [68]. The remaining major post-Beckerian taxonomic treatments of Viola by specialists have been regional, e.g., for North America [69,70], Peru [71], the former Soviet Union and Russia [21,61,72], Europe [73], Malesia [74], China and Taiwan [75,76,77,78], Iran and parts of adjacent countries [79], Norden [19], and Argentina [80]. In general, the Russian and Asian taxonomic treatments have combined Becker’s sections at the subgenus level and used higher taxonomic ranks for all the infrageneric groups of Viola. There is currently no taxonomic consensus.
Of the numerous phylogenetic studies that have been published for Viola [2,28,45,60,81,82,83,84,85,86,87,88,89,90,91,92,93,94] only two have been near-comprehensive in terms of sampling of infrageneric groups [2,28]. The ITS phylogeny of Ballard et al. [2] was the first phylogeny for Viola and covered eight of Becker’s 14 sections. The species-level phylogenies of Marcussen et al. [28,45] covered all of Becker’s sections, and based on three low-copy genes and a chloroplast marker, allowed also for the reconstruction of reticulate, allopolyploid history of the genus. Among other things, the phylogenetic findings lended support to Clausen’s [29,56,59] suggestions for a re-circumscription of the large and heterogeneous sect. Nomimium and to Reiche’s [48] early suggestion to recognise the South American rosulate violets at a higher taxonomic level. In addition, numerous new infrageneric segregates have been identified (or confirmed) in recent years that require taxonomic recognition, i.e., V. abyssinica and relatives and V. decumbens in Africa [28], the recently discovered V. hybanthoides in China, which has been assigned to the monotypic sect. Danxiaviola [90], V. kunawurensis for which a reference genome is on the way (NCBI accession PRJNA805692, as V. “kunawarensis”), and V. spathulata and relatives [28] in Eurasia, and a large clade of North American and Hawaiian allodecaploids provisionally referred to as sect. Nosphinium s.lat. [2,45,81].
In summary, the knowledge that has been accumulating for nearly a century, since the last revision of Viola by Becker in 1925 [1], has not been revised and systematised. This has beyond doubt hindered the testing of new hypotheses and obtaining new knowledge. Since the last revision in 1925, the number of known species in Viola has increased by 60% and numerous new infrageneric segregates have been identified using molecular methods and morphology. Among the amended classifications that do exist, no consensus exists for use of rank, delimitation, or nomenclature, mostly because each of these classifications covered only a small part of the genus and taxon delimitation and rank had not been defined in the context of the total variation within Viola. Furthermore, none of the hitherto proposed classifications have been phylogenetic by nature and aimed at reconciling taxon monophyly and the extensive reticulate evolution due to allopolyploidy [28] in the genus. Finally, it is now known that a substantial proportion of the known species of Viola are narrow endemics and endangered species and are as such at risk of extinction due to human-induced changes in land use and climate [95]. Viola (sect. Melanium) cryana, is considered extinct in Europe and globally [96] and V. (sect. Plagiostigma) stoloniflora is considered extinct in the wild in the Ryukyus Islands [97], and it is to be feared that up to 27 species within subg. Neoandinium, most of which have not been seen since the type collection, have become extinct [68].
The aim of this revision was to generate an updated infrageneric taxonomy for Viola based on modern principles of phylogenetics and monophyly and the accumulated information since Becker’s previous morphology-based classification from 1925 [1]. The revision is presented as (1) a global checklist of species of Viola accepted by us and annotated with infrageneric taxonomy, (2) an updated multigene phylogenetic network and an ITS phylogeny with denser taxon sampling, (3) a brief summary of the taxonomic changes from Becker’s classification and their justification, (4) an account of each infrageneric group with justifications for delimitation and rank including a description, a list of apomorphies, molecular phylogenies where possible or relevant, and a list of accepted species, and (5) a morphological binary key to the accepted subgenera, sections and subsections. It is our intention and hope that this synthesis, by summarising what is known and what remains to be known for Viola, will serve as both a foundation and an inspiration for further studies on this large, diverse and insufficiently understood genus.
2. Results and Discussion
2.1. Phylogeny and Classification
We recognise 664 known species of Viola, 43 of which have not yet been described. The global species checklist, annotated with infrageneric taxonomy, is presented in Appendix A. We subdivide the genus into two subgenera, 31 sections, and 20 subsections. Subgenus Neoandinium comprises 139 species in 11 sections, and subg. Viola, 525 species in 20 sections and 20 subsections (Table 1; Figure 3). Photographs of representatives of each accepted infrageneric segregate are shown in Figure 3; these images are also presented in a downloadable and printable poster in Supplementary File S1. Section Plagiostigma is by far the most species-rich section with 142 species, followed by sect. Melanium with 110 species. Nearly half of the sections, 15 of 31, include three species or less (Figure 4). We propose subg. Neoandinium as a replacement name for the illegitimate subg. Andinium (W. Becker) Marcussen, and erect 13 new infrageneric taxa within subg. Viola, i.e., six new sections (sect. Abyssinium, sect. Himalayum, sect. Melvio, sect. Nematocaulon, sect. Spathulidium, and sect. Xanthidium), and seven new subsections (subsects. Australasiaticae, Bulbosae, and Formosanae within sect. Plagiostigma, subsect. Clausenianae within sect. Nosphinium, and subsects. Cleistogamae, Dispares, and Pseudorupestres within sect. Melanium). Justifications for erecting new taxa are given under each taxon in the taxonomic section and in the form of a binary key (Chapter 5).
2.1.1. Genus Phylogeny
We updated the allopolyploid phylogenetic network obtained by Marcussen et al. (2015 [28]), based on homoeologs of three low-copy nuclear genes, with new information on chromosome counts and sequences (Figure 5). A dated phylogeny of the ITS marker, with denser sampling for selected taxa, is shown in Figure 6. New ITS sequences provided a new and older crown node age for subg. Neoandinium (c. 20.3 Ma) compared to Marcussen et al. [28], and also allowed placing the two novel sections Danxiaviola [90] and Himalayum as distinct lineages within the North Hemisphere CHAM + MELVIO allopolyploid tangle in Figure 5. We have re-evaluated the phylogenetic placement of Viola (sect. Melvio) decumbens (Figure 5), after discarding the erroneous trnL-trnF sequence that placed it next to V. arborescens in sect. Xylinosium [28]. Viola decumbens appears to be hexaploid, as each of the three low-copy genes analysed by [28] has three MELVIO homoeologs that coalesce around 17–22 Ma. These homoeologs coalesce slightly shallower (on average 1.6 Ma) with one another than with the rest of the MELVIO clade, suggesting that the subgenomes of V. decumbens constitute a monophyletic sister to the rest of the MELVIO lineage. No chromosome counts exist for V. decumbens. The updated and corrected chromosome counts on V. (sect. Erpetion) banksii (2n = 50, not 60) and V. (sect. Tridens) tridentata (2n = 40, not 80) are reconcilable with the molecular data without the need to formulate complex hypotheses of homoeolog loss and duplication (cf. [28]). Both homoeolog number and chromosome count for sect. Erpetion indicate that this lineage is allo-octoploid (Figure 5); the recent count of 2n = 50 in V. banksii [98] is very close indeed to the expected 2n = 48 based on x = 6 in the diploid ancestor of sects. Chamaemelanium and Rubellium. Similarly, for sect. Tridens both homoeolog number and chromosome count agree with allohexaploidy (Figure 5); the count of 2n = 40 [99] is very close to the expected 2n = 38 based on x = 6 in the two genomes shared with sect. Erpetion and x = 7 in the one shared with sect. Leptidium.
2.1.2. Justification for Taxonomic Levels and Classification
The phylogenetic history of Viola is reticulated to such an extent that monophyletic groups can be delimited at three hierarchical levels only (Figure 5). The highest hierarchical level corresponds to subgenus in our treatment and delimitates two monophyletic taxa, i.e., subg. Neoandinium and subg. Viola. The intermediate hierarchical level corresponds to the section level. The lowest hierarchical level delimits subsections. Below the level of subsection, taxa are interconnected by allopolyploidy and the taxonomic level of series is not applicable as a result of non-monophyly. In addition, we have chosen to apply the levels of subgenus, section and subsection because this use maximises taxonomic stability by minimising the number of changes from Becker’s [1] treatment and by allowing us to keep most of his sections.
The alternative to treating Neoandinium at the subgenus level would be to recognise it as a separate genus (e.g., as Andinium). This could have been justified both morphologically and phylogenetically. However, this change would be phylogenetically unnecessary, as monophyly is not affected, and there is also no need for additional taxonomic levels within Viola, considering that we here abandon the taxonomic level of series for reasons of monophyly. Recognising a separate genus for subg. Neoandinium would further disrupt taxonomic stability and require numerous new taxonomic combinations to be made.
In our taxonomic treatment, sect. Nosphinium is the only exception to the rule of strict monophyly, which cannot be enforced due to the conceptual conflict between reticulate evolution, as a result of allopolyploidy, and the hierarchical system of classification. This conflict occurs because sect. Nosphinium is an allodecaploid lineage that originated by hybridisation between taxa deeply nested within the sections Chamaemelanium, Plagiostigma, and Viola, and that during its diversification acquired several additional Plagiostigma genomes by further allopolyploidisation [45]. Enforcing strict monophyly in this case would, by a domino effect, have the undesirable consequence that all sections within subg. Viola were rendered non-monophyletic.
2.1.3. Changes to Becker’s Original System for Viola
The comprehensive classification of Viola presented here is the first since that proposed by Becker [1] nearly a century ago. Changes in classification from Becker’s system are summarised and displayed as a “wire” diagram in Figure 7. We give justifications for these changes under each taxon in Chapter 2.4. (Taxonomic Treatment of Viola).
Becker [1] recognised 14 sections and numerous infrasectional greges within Viola. Here, we suggest recognising two subgenera, subg. Neoandinium (Becker [1]: sect. Andinium) with 11 sections and subg. Viola with 20 sections and 18 subsections. Recently, Watson et al. [68] proposed a provisional classification of subg. Neoandinium (as subg. Andinium) with 11 sections based on general morphology. In the absence of phylogenetic data and a good understanding of character polarity in the two subgenera, we tentatively follow this classification. Within subg. Viola, we make the largest changes in circumscription to Becker’s sections Nomimium, Dischidium, Nosphinium, and Chamaemelanium, where Becker’s [1] species groups are now re-distributed among six sections. Section Chamaemelanium now comprises the former sect. Dischidium and greges Memorabiles and Orbiculares of sect. Nomimium. Section Viola corresponds to the former sect. Nomimium s.str. and unites greges Repentes, Umbraticolae and Rostratae in subsect. Rostratae, and greges Uncinatae, Lignosae and Serpentes (pro parte) in subsect. Viola. Section Plagiostigma unites greges Serpentes pro parte, Vaginatae, Bilobatae and Stolonosae in subsect. Australasiaticae and subsect. Stolonosae, retains grex Diffusae as subsect. Diffusae and retains most of grex Adnatae as subsect. Patellares. Section Nosphinium, which in the original sense comprised the Hawaiian Viola only, is here considerably expanded and comprises subsect. Borealiamericanae, subsect. Mexicanae, subsect. Pedatae, and subsect. Langsdorffianae (all previously greges of sect. Nomimium), next to subsect. Nosphinium (Becker [1]: sect. Nosphinium) and subsect. Clausenianae.
In subg. Viola six new sections have been erected to accommodate the following taxa: sect. Abyssinium for the African species V. abyssinica and allies (Becker [1]: mentioned but not formally classified); sect. Himalayum for V. kunawarensis in the Himalayas (Becker [1]: sect. Nomimium grex Adnatae); sect. Melvio for the South African Cape endemic V. decumbens (Becker [1]: sect. Xylinosium); sect. Nematocaulon for the New Zealand endemic V. filicaulis (Becker [1]: mentioned but not formally classified); sect. Spathulidium for V. spathulata and allies in southwestern Asia (Becker [1]: sect. Nomimium grex Adnatae); and sect. Xanthidium for the V. flavicans group in southern South America (Becker: not included in the monograph [1] but mentioned elsewhere [100,101,102]). Section Danxiaviola has already been published to accommodate the newly described V. hybanthoides endemic to Yunnan, China [90]. These six new sections comprise in total about 11 species only, indicating that Becker’s [1] century-old classification provided a remarkably good overview of the genus.
2.2. Patterns of Evolution within Viola
2.2.1. Historical Biogeography of Viola
We reconstructed the historical biogeography of Viola (Figure 8) using a simplified approach based on four biogeographic categories, a single-rate transition model, and 50 operational taxonomic units as defined in the diploid multilabelled phylogenetic timetree that is the counterpart of the phylogenetic allopolyploid network in Figure 5. Our result gives the strongest possible support (pp = 1.0) to the previously proposed, but never actually tested, hypothesis that Viola originated in South America [2,28,29]. Subgenus Neoandinium has remained within the ancestral range in South America. Within subg. Viola, it is inferred that the CHAM and MELVIO lineages dispersed independently into the Northern Hemisphere 20–25 Ma ago where they eventually met and formed allopolyploids. Intersectional biogeographic relationships within the Northern Hemisphere are not resolvable due to the basal polytomy. However, it seems likely that the diploid CHAM lineage dispersed northwards from South America into North America, where it gave rise to sect. Chamaemelanium which at present has its diversity centre along the Pacific coast of North America; this scenario was proposed already by Clausen nearly a century ago [29]. The dispersal history of the diploid MELVIO lineage remains unknown, as it is represented by a single species (V. (sect. Melvio) decumbens) that occurs allopatrically in the Cape of South Africa. It seems clear, however, that members of CHAM and MELVIO both dispersed into Eurasia where they by hybridisation gave rise to numerous allopolyploid lineages, most of which correspond to sections in our treatments. Western Eurasia appears to have been the cradle of early allopolyploid diversification, as the majority of these sections are endemic or have diversity centres here; only three sections have diversity centres in eastern Eurasia (sects. Danxiaviola, Himalayum, and Plagiostigma). Both the ancestral diploids (CHAM and MELVIO) have since become extinct in western Eurasia.
Concerning the several dispersals out of South America, it would not be surprising if Viola was shown to have migrated northwards within the continent, following the progressive rise of the Andes and the advent of new alpine habitats, as known for many other taxa. Although the current mountain elevations were reached relatively recently during the Plio-Pleistocene [104,105,106], the southern and central Andes date back to the Cretaceous, and the northern Andes to the Eocene [107,108]. Regarding the dispersal of Viola from South America to North America, first, there is evidence that the Isthmus of Panama in Central America was an uninterrupted chain above sea level from the late Eocene until at least the late Miocene [109]. Second, although the distances were presumably too long to allow for direct dispersal between North and South America, comparing these patterns with those involving dispersal to the Pacific Islands, Carlquist [110] suggested that dispersal by birds could account for many of the disjunct distributions between North and South America. Third and finally, even though Viola species do not have palatable fruits or seeds, a large number of genera showing disjunctions do not have obvious effective long-distance dispersal mechanisms either. Some birds may eat and thus internally carry fruits or seeds other than those that are big, fleshy, and strikingly coloured [110].
Regardless of mode, Violas are apparently quite capable of long-distance dispersal and have successfully colonised remote oceanic islands like Hawaii, New Zealand, and the Azores, the temperate “sky islands” of tropical Africa, and the South African Cape province, or have even dispersed back to southern South America (V. huidobrii). A few extant species occur on more than one continent (e.g., V. biflora, V. palustris, V. rostrata, V. selkirkii, V. suecica).
2.2.2. Hybridisation and Allopolyploidy
Interspecific hybridisation is common in Viola and is well studied for the sections in the Northern Hemisphere. Hybridisation occurs most commonly between pairs of closely related species, especially among those that share a genome due to allopolyploidy, such as V. epipsila (4x) and V. palustris (8x) and European members of subsect. Rostratae (4x/8x/12x) [19,38,39,111,112,113,114,115,116]. As a result, spontaneous hybrids occur nearly exclusively between taxa within the same subsection, more rarely between species belonging in different subsections, and only occasionally between species in different sections. The most phylogenetically distant taxa to form spontaneous hybrids are members of sect. Plagiostigma subsect. Patellares and sect. Viola subsect. Rostratae, which are estimated to have diverged some 19 Ma ago (Figure 5). Their hybrids are extremely rare and have been reported from single individuals only, of which V. japonica × V. rostrata is the only one that has been confirmed by DNA data [61,117,118]. Artificial hybrids are, however, easily made between members of these two sections and also with sect. Nosphinium subsect. Borealiamericanae, to a lesser degree with sect. Chamaemelanium [57,58]. The genomic compatibility of these lineages most likely reflects their comparatively slow evolutionary rates [28].
The symplesiomorphic, retained ability of taxa to interbreed for millions of years after they diverged has evidently played an important role in the phylogenetic history of the genus by allowing for extensive allopolyploid speciation (Figure 5; [28,45,60]. Although historically most allopolyploidisations have involved recently diverged parental taxa, their divergence may have been more than 10 Ma for mesopolyploids such as sect. Leptidium and sect. Tridens (Figure 5) and widespread neo-octoploids such as V. blanda, V. incognita, V. pluviae, and V. palustris [45,93]. All these four neo-octoploids have Boreal distributions and their origins coincide with the climate cooling and repeated glaciations in the last 5 Ma [119]. More than anything, this shows that the ability to hybridise and speciate by allopolyploidisation can be a rapid mode of diversification to fill vacant niches (e.g., [120]).
The association of long-distance dispersal with polyploidy is striking in Viola. In each of the seven cases of long-distance dispersals older than a few Ma (Figure 5 and Figure 8), the colonist taxon has a higher ploidy than its sister taxon or, if known, ancestor. This is seen on a massive scale in connection with the colonisation of the Northern Hemisphere by the CHAM and MELVIO lineages, which occurred c. 19 Ma ago and gave rise to more than 400 species [28], and with the decaploidisation that gave rise to sect. Nosphinium following independent dispersal to North America of its ancestors in sect. Plagiostigma and sect. Viola, which occurred c. 10 Ma ago and gave rise to 61 species [45]. The same pattern of increased ploidy in the colonist taxon is seen on a smaller scale for sect. Erpetion in Australia within the last 7 Ma (11 species), for subsect. Nosphinium in the Hawaiian islands within the last 5 Ma (9 species), for sect. Abyssinium in tropical African mountains within the last 5 Ma (3 species), for sect. Melvio (i.e., V. decumbens) in South Africa possibly 20 Ma ago, and for sect. Nematocaulon (i.e., V. filicaulis) in New Zealand, age unknown. In the four cases where there is sufficient phylogenetic resolution, polyploidisation seems to have occurred after colonisation (CHAM + MELVIO, sect. Nosphinium, sect. Erpetion, sect. Melvio). This indicates that polyploidy is linked with colonisation rather than dispersal, an association that is general across angiosperms and may reflect that speciation by polyploidy gets to dominate during phases of colonisation because it is a much faster process than homoploid speciation (e.g., [120]).
The phylogenetic network for Viola (Figure 5) contains 13 homoploid speciations and 23 allopolyploid speciations, which means that allopolyploidy may have accounted for 64% (=23/(13 + 23)) of the speciations above the section level. This proportion is lower than the estimate of 67–88% by Marcussen et al. [28] as a result of new and re-interpreted information for numerous sections, as well as an expanded set of taxa, but the estimate is still far higher than that for angiosperms as a whole, estimated to 15% [41] or 30% [42].
The reason why polyploidisation is more common in Viola than in other lineages is probably linked to the ability to hybridise in combination with cleistogamy. The retained ability for lineages to form hybrids, in some cases up to 15 Ma or more, provides the raw material for allopolyploid formation. Regular selfing through cleistogamy might help the nascent allopolyploid in the early phases of establishment.
2.2.3. Base Chromosome Numbers in Viola
The limited number of chromosome counts appears to indicate that x = 7 may be the base chromosome number for Viola as a whole. The two counts in subg. Neoandinium both show 2n = 14 [121]. For subg. Viola, x = 6 was long assumed because 2n = 12 is shared by its two diploid sections, Chamaemelanium [29,43,44] and sect. Rubellium [60]. However, the two deepest lineages of subg. Viola, which are now extinct as diploids, may rather have had x = 7, which is indicated by ploidy and chromosome counts for the two polyploid sections Leptidium (x = 6.75, based on 2n = 54 [53] and 8x; Figure 5) and Tridens (x = 6.67, based on 2n = 40 [99] and 6x; Figure 5). The reduction from x = 7 to x = 6 may therefore be a synapomorphy for the most recent common ancestor of sects. Chamaemelanium and Rubellium. This hypothesis needs to be tested with additional counts for the South American lineages of Viola, and also from the sister genera, Noisettia and Schweiggeria, for which data are lacking but would be relevant for understanding character polarity.
2.2.4. Morphology, Anatomy, Adaptations
Perhaps the most striking finding in our phylogeny of Viola is the lack of a clear correspondence between macromorphology and phylogeny, with the exception of the distinction between subg. Viola and subg. Neoandinium, and style morphology to some extent (Figure 9). There are two likely causes for this—the highly reticulate phylogeny, which has allowed for the redistribution of apomorphies and adaptations, and the large polytomy at the base of the Northern Hemisphere taxa, which precludes the existence of synapomorphies among these taxa (Figure 8).
Style shape is variable in Viola (Figure 9) and has historically been a key character to subdivide the genus [1,29,46,47,48,59,61,68,90,122,130]. While broad diversity of style morphologies have been used previously for limited studies of taxa within subsections or sections of the genus, we sought to greatly expand the sampling to encompass the main “phenotypes” of style morphology within the two subgenera and all sections and subsections, and to evaluate the efficacy of style traits for delimiting higher-level taxa in addition to morphology. We recognised broad types of styles, first as “undifferentiated” (styles cylindrical, often straight, lacking apical ridges or processes and terminating in the stigmatic orifice) and “differentiated” (clavate or capitate, with processes or apical ridges or lobes, the stigmatic orifice on a rostellum). Additional traits were noted, such as the presence/absence of papillae or trichomes; the shape, orientation and fusion of apical ridges or lobes; and the thickness, prolongation and orientation of the rostellum supporting the stigmatic orifice. In subg. Neoandinium, the bulk of species display conspicuous and remarkable types of crests and processes, each species often dramatically distinct in these stylar adornments. We speculate that the divergent stylar crests or processes among related species serve a role in pollinator specificity, in a region where the paucity of pollination vectors could drive selection for diverse pollinator behaviours to reduce hybridisation. In subg. Viola, the range of style morphologies within some larger sections such as Chamaemelanium and Plagiostigma is very broad, whereas the range within Leptidium, Melanium, and Viola is generally quite narrow. Variability within subsections is generally quite narrow and readily characterised. In all higher-level taxa (sections and subgenera), the range of style morphologies can be discretely described and used to support the recognition of higher-level taxa based on morphology and chromosome number. In particular, style morphology can provide distinctive apomorphies where certain morphological features may be homoplasious in comparing some higher-level taxa, especially in sect. Nosphinium and sect. Plagiostigma.
Some of the variations in style morphology are geographically structured and might reflect adaptation to special modes of pollination and/or pollinators. Undifferentiated, filiform styles occur exclusively in tropical-montane and south-temperate taxa, i.e., sect. Erpetion, sect. Leptidium, sect. Tridens, sect. Nematocaulon, and in single species within sect. Chilenium (V. commersonii), and sect. Viola (V. papuana). Trichomatous-bearded styles occur exclusively in north-temperate taxa, i.e., sects. Chamaemelanium, Melanium, and Viola.
Shoot morphology has been given much attention in previous classifications, at least among the herbaceous Northern Hemisphere taxa, notably the presence or absence of leaf rosettes, aerial stems, or stolons. Taxa have accordingly been described and classified as rosulate or arosulate, caulescent or acaulescent, and stolonose or estolonose (e.g., [1,131,132,133,134]). This classification is, however, artificial and does not reflect phylogenetic relationships. In addition, this classification is problematic because of the logical flaw of defining taxa based on the absence of a structure (e.g., acaulescence), and it also eludes the possibility that aerial stems in one “caulescent” taxon could be homologous with stolons in another “acaulescent stolonose” taxon, as otherwise suggested by the intermediate morphology of interspecific hybrids (e.g., V. canina × V. uliginosa, V. odorata × V. riviniana [58], V. epipsila × V. riviniana; T.M., unpublished data from crossing experiments). In any case, our data show that shoot morphology is quite labile and that loss, gain, or transitions among character states have occurred repeatedly in the four sections Nosphinium, Plagiostigma, Viola, and Chamaemelanium to the extent that it is not possible to infer which state(s) was ancestral; the exception is sect. Chamaemelanium where nearly all species have aerial stems and this character state seems to be ancestral. The loss of lateral stems presumably has a simple genetic basis, but these structures appear to be gained almost as easily. For instance, within sect. Plagiostigma, aerial stems have been invented from an ancestor that lacked them in subsect. Diffusae within the last 3 Ma (V. guangzhouensis) and in subsect. Stolonosae within the last 5 Ma (V. moupinensis). Similarly, stolons have been invented de novo in sect. Erpetion within the last 7 Ma. Within sect. Viola subsect. Rostratae all character states (i.e., aerial stem, stolon, or absence of both) may have evolved within the last 7 Ma.
Another conspicuous characteristic is woodiness. This was most obviously the ancestral character state at the stem node of the genus, given that the sister lineage of Viola (Noisettia and Schweiggeria) and nearly all other genera in Violaceae are woody. However, the most recent common ancestor of Viola was probably not a lignose. Shrubby and subshrubby taxa occur scattered throughout the genus, and the fact that shrubbiness is most definitely derived in the taxa of subsect. Nosphinium, which arrived in the Hawaiian Islands some 5 Ma ago (see Chapter 5) [45,81,85], indicates that this too is a plastic character. Furthermore, none of the shrubby taxa of Viola (except for the Hawaiian ones) have retained the differentiated shoot architecture found in Noisettia and Schweiggeria as well as woody seed plants in general, with growth axes differentiated in orthotropic vegetative axes and plagiotropic reproductive axes [135].
A suite of characters appears to have evolved in the ancestor of the Northern Hemisphere taxa, perhaps in part as adaptations to increased seasonality as compared to South America. These include a shoot architecture with differentiated growth axes, seasonal cleistogamy, and a bearded style. All three characters are expressed in the diploid sect. Chamaemelanium and might therefore be adaptations associated with the ancestral CHAM genome, but they are not expressed in all of the allopolyploids having CHAM and MELVIO genomes. In sect. Chamaemelanium shoot differentiation is extreme, with the perennating axis usually being a deep-buried rhizome and lateral stems annual, aerial and floriferous; this differentiation is less extreme, but present in large sections such as Viola, Plagiostigma and Nosphinium. Another character is cleistogamy, which is common in Viola. Viola has the type of cleistogamy referred to as dimorphic, i.e., the primordial bud is already predetermined to develop into either a chasmogamous or cleistogamous flower [27]. Cleistogamy is facultative in the Southern Hemisphere lineages in sects. Leptidium, Chilenium, and Nematocaulon, and at least in the last two may occur as reproductive assurance under unfavourable conditions [26,136]. Many of the Northern Hemisphere lineages have instead evolved seasonal cleistogamy by which production of flower type is determined by photoperiod and temperature: during long-day conditions, cleistogamous flowers are produced and during short-day conditions, chasmogamous flower buds are produced that remain dormant until the following spring [137,138,139,140,141,142,143,144]. Seasonal cleistogamy is known from sects. Chamaemelanium, Himalayum, Melanium, Nosphinium, Plagiostigma, and Viola.
There have been no comprehensive anatomical studies of Viola (cf. [145]), but investigations have been conducted on particular species or groups of species (e.g., [145,146,147,148,149,150,151,152,153,154]). Shoot architecture has been studied for a few European species [155].
Pollen in Violaceae is generally tricolporate [156]. In Viola, however, about one third of the species are heteromorphic with regard to pollen aperture number, which has been explained as a consequence of neopolyploidy [157]. Hence, up to five and six apertures occur in the high-polyploids (4x to 20x) of sect. Melanium whereas three and four apertures occur in the other investigated sections [157]. It may be noted that [157] severely underestimated the ploidy of most of the investigated taxa; e.g., the 12-ploid V. tricolor, 16-ploid V. arvensis, and 18-ploid V. langsdorffii were all interpreted to be diploid. Gavrilova & Nikitin [147] found that East European species in the sections Chamaemelanium, Plagiostigma, and Viola have 3–(4)-colp(oroide)ate pollen with long colpa and mostly complex exine ornamentation, while sect. Melanium has (3–)4–5(–6)-colporate pollen with shorter colpa and simple exine ornamentation. No palynological data exist on South American members of the genus.
Viola comprises numerous metallophytes, i.e., species or populations of species with the capacity to tolerate metal toxicities as well as survive and reproduce on metalliferous soils. While the situation in subg. Neoandinium is not known, within subg. Viola at least 20 taxa are known to hyperaccumulate heavy metals and other toxic elements (As, Cd, Ni, Pb, Sb, Tl, Zn) [158,159]. Most of these taxa belong in sect. Melanium, but all the four largest sections in the northern hemisphere are represented, i.e., sect. Chamaemelanium (V. cuneata, V. brevistipulata), sect. Melanium (V. albanica, V. allchariensis, V. arsenica, V. beckiana, V. dukadjinica, V. elegantula, V. kopaonikensis, V. lutea, V. raunsiensis, V. tricolor, V. vourinensis, V. ×wittrockiana), sect. Nosphinium (V. howellii), sect. Plagiostigma (V. baoshanensis, V. philippica, V. principis), and sect. Viola (V. sacchalinensis, V. kizildaghensis). Additionally, some members of sect. Erpetion (V. banksii, V. serpentinicola) may well prove to be hyperaccumulators [98,160]. The high prevalence of metallophytes in sect. Melanium reflects the general ability of members of this section to adapt to extreme abiotic conditions.
Although no species of subg. Neoandinium are known to be hyperaccumulators, it is reasonable to assume that several species are, especially those known from metalliferous soils in the immediate proximity of Andean copper mines in Chile, such as V. escarapela, V. exilis, and V. gelida (sect. Rosulatae), V. godoyae (sect. Relictium), V. uniquissima (sect. Triflabellium), and V. vallenarensis (sect. Subandinium) [68,161,162]. In fact, V. godoyae and V. uniquissima were both discovered during the initial surveys of the immediate areas where their respective mines are located. It is also noteworthy that many Neoandinium taxa have an affinity for extreme abiotic conditions in much the same way as sect. Melanium.
In Violaceae, hyperaccumulators occur within another four genera outside of Viola, i.e., Afrohybanthus, Agatea, Pigea, and Rinorea [163,164,165,166,167]. These five genera are phylogenetically scattered and not closely related [3], indicating that the ability of metal hyperaccumulation evolved independently in each lineage, presumably from a set of shared preadaptations. The possible occurrence of hyperaccumulators in both subgenera of Viola points to a set of preadaptations being shared across the genus.
2.2.5. Biotic Interactions
The chasmogamous flowers of Viola are visited and pollinated primarily by solitary bees (Anthophoridae, Halictidae, Andrenidae; Hymenoptera), but also bumblebees (Apidae, Hymenoptera), hoverflies (Syrphidae, Diptera), bee flies (Bombyliidae, Diptera), butterflies (Lepidoptera), and in some species beetles (Coleoptera) [168,169,170,171,172,173]. Many species appear to be generalists but the degree of specialisation varies considerably among species and even populations and appears to be evolutionary plastic. For instance, among the closely related species of subsect. Rostratae, V. adunca and V. striata are visited mostly by solitary bees, V. reichenbachiana mostly by hoverflies, and the longer-spurred V. rostrata mostly by bee flies [169]. Populations in natural and disturbed sites can attract widely differ pollinators, as shown for V. (sect. Rubellium) portalesia [173]. Among the most highly specialised are V. (sect. Leptidium) arguta [174] which has a red corolla and a saccate spur and is the only Viola pollinated by hummingbirds (Trochilidae, Aves), and the three species of sect. Delphiniopsis which are characterised by bright pink corollas with a very long, thin spur and are pollinated by a single species of day-flying hawk-moth (Macroglossum stellatarum, Sphingidae, Lepidoptera) [175,176].
Spurred, nectar-producing flowers occur also in the sister genera of Viola, Noisettia and Schweiggeria, and must therefore be considered the ancestral state in Viola. In certain sections of subg. Viola with species adapted to pollination by solitary bees there has been a transition from nectar to pollen as a pollinator reward. Contrary to “nectar flowers”, which produce nectar from the nectariferous appendage of the two bottom anthers and store it in the spur of the bottom petal, these “pollen flowers” do not produce nectar and have reduced anther appendages and petal spur [171,177,178]. This transition has occurred independently in sect. Leptidium [171] and sect. Erpetion [177,178], but morphology suggests that several other short-spurred groups may have evolved into “pollen” flowers; at least V. sumatrana (sect. Plagiostigma, subsect. Australasiaticae) appears to produce no nectar [171]. In sect. Leptidium the bottom pair of stamens have prolonged “u”-shaped anther connective appendages that the female bee (Anthrenoides, Andrenidae) holds onto while harvesting pollen by vibration (“buzz-pollination”) [171]; curiously, the only bird-pollinated Viola species, V. arguta, belongs to the same section and does produce copious amounts of nectar (4 µL per 24 h) [174].
Caterpillars of Nymphalidae butterflies, subfam. Heliconiinae, are specialised to feeding on the foliage of members of the parietal Malpighiales, which includes Violaceae, Passifloraceae, and Salicaceae [179]. Viola are frequently predated by the larvae of tribe Argynnini whose diversification in time and space appears to have been tracking that of Viola. The ancestral Argynnini used Passiflora and Violaceae as larval host plants but specialising on Violaceae appears to have occurred prior to c. 23 Ma on the branch subtending the stem node of Yramea [180,181,182] in South America, where Yramea occurs today. The crown node of Viola is estimated to be just a few Ma older, c. 31 Ma [28]. Dispersal into Eurasia of the common ancestor of Boloria, Issoria, Brenthis, Argynnis (Argynnina clade) may have happened around c. 15 Ma ago [181,182,183], which again corresponds well with the appearance of Viola seeds in the Eurasian fossil record 17–18 Ma [184,185]. Finally, further dispersal and diversification of Argynnis (Speyera subclade) into North America is estimated to have occurred c. 5 Ma ago [183,186], apparently tracking the diversification of the decaploid sect. Nosphinium lineage [45]. Host switches from Viola to other cold-temperate taxa have occurred in Boloria (to, e.g., Dryas, Vaccinium, Salix) and in Brenthis (to mainly Rosaceae) [180,186].
While tribe Argynnini have larvae that feed mostly on Violaceae, the other lineages within subfam. Heliconiinae have specialised on the closely related Passifloraceae (the clade consisting of tribes Heliconiini and Acraeini) and Salicaceae (tribe Vagrantini). A Malpighialean larval host appears to be ancestral in the common ancestor of Heliconiinae and its sister lineage, subfam. Limenitidinae [179].
2.2.6. Fossil Record of Viola
Viola is represented in the fossil record of Eurasia from the Miocene onwards, by both pollen [187,188,189,190,191,192,193] and seeds [185,194,195,196,197,198,199,200,201,202]. There are in addition unconfirmed records of Viola macrofossils from the Pliocene and Pleistocene of North America [203,204,205,206]. Viola has no known fossil record in South America although this continent is where the genus has the longest history.
Records of Viola fossils older than Miocene are deemed questionable [200,207]. Significantly, one of the most frequently cited fossils (e.g., [208]), “Viola” rimosa P. Nikit. from the Oligocene and Miocene of western Eurasia, was reidentified as Poliothyrsis (Salicaceae) [207].
Seeds of Viola can be recognised by the relatively large chalaza, the transverse cellular pattern of the inner surface of the testa, and the existence of a layer with rhomboid crystals within the testa [197,199,200]. Fossil seeds of Viola are common in western Eurasian sediments from the Miocene onwards, where a total of c. 19 extinct morphotypes have been described [185,194,195,196,197,198,199,200,201,202]. Most of these are known from single fossil sites only but two have a wide stratigraphic range, i.e., V. miocenica (20.44–5.333 Ma, western Siberia [202]) and V. neogenica (15.97–2.58 Ma, Germany and Italy [200,201,209]). The oldest fossils of Viola are seeds from the Lower Miocene of Europe and comprise several unnamed morphotypes, one from the Burdigalian (18–17 Ma [184]) of Austria [185] and three from the Upper Karpatian (17.5–16.5 Ma [210]) of Poland [197]. Additionally four morphotypes, two of which are closely similar to one of the Polish ones [197], have been described from western Siberia [196,202] from about the same time interval (20.44–11.63 Ma [211]).
Seed fossils closely similar to, and possibly attributable to, extant species of Viola are known back to the Pliocene (5.333–2.58 Ma) of Europe, i.e., V. palustris back to the Lower Pliocene (5.333–3.6 Ma) of Germany [198,200] and European Russia [202], V. tricolor back to the Upper Pliocene (3.6–2.58 Ma) of Germany [198], and V. cf. uliginosa back to the Pliocene of Poland [194,197]. Seeds attributed to the extant V. canina and V. rupestris (probably incorrectly so) have been reported from the Tortonian (10–9 Ma [212]) of Germany [199]. Seed morphotypes comparable to sect. Viola have been reported from the Miocene of western Siberia [202], i.e., V. miocenica and Viola [Arbuzova] sp. 6 (both compared to V. alba, V. collina, V. mirabilis, V. riviniana, and V. suavis). Seed morphotypes comparable to either of the two subsections of sect. Viola are younger, from the Pliocene (5.333–2.58 Ma) of the southern Urals [202]; i.e., Viola [Arbuzova] sp. 1 to 3 are compared to species of subsect. Viola (V. alba, V. ambigua, V. collina, and V. suavis); and Viola [Arbuzova] sp. 4 is compared to species of subsect. Rostatae (V. mirabilis, V. reichenbachiana, and V. tanaitica). Three among the oldest seed morphotypes (20.44–11.63 Ma) from western Siberia were reported to bear no similarity to extant taxa, i.e., Viola [Arbuzova] sp. 5, Viola [Arbuzova] sp. 8, and V. kireevskiana [202].
The assignments of these fossils to extant infrageneric taxa of Viola should be considered tentative as none has been justified by apomorphies or phylogenetic analysis. As noted by Łańcucka-Środoniowa [197], the taxonomic distinction of species in the genus Viola is difficult because the structure of seeds is very similar, at least among the European sections. Indeed, in a survey of seed morphology in East European angiosperms, Bojňanský & Fargašová [213] found no significant differences in seed morphology among the four sections of Viola studied by them, based on 28 species. However, their survey employed rather superficial morphological features observable using a light microscope, and it is therefore possible that more detailed studies using scanning electron microscope (SEM) micrographs on a more comprehensive sample of Viola sections could reveal apomorphies, e.g., such as seen within subsect. Borealiamericanae [214]. The only infrageneric group that stands out as distinct is the obligate myrmecochorous [215] subsect. Viola with its apomorphic large seeds, 2.0–3.0 × 1.3–2.0 mm (vs. 1.3–2.9 × 0.7–1.7 mm in other species), with a large elaiosome covering about half of the length of the raphe (vs. <1/3 in other taxa) [57,113,213]. The three fossil seed morphotypes with possible affinity to subsect. Viola, from the Pliocene of the southern Urals, are somewhat smaller (1.8–2.4 × 1.3–1.6 mm [202]) than seeds of extant species of this subsection [213]. However, at least within sect. Viola, seeds derived from chasmogamous flowers are often larger and heavier than seeds from cleistogamous flowers [216], up to almost twice as heavy in V. odorata [19].
The sudden appearance of Viola in the fossil record of western Eurasia and its almost immediate diversification into several recognisable morphotypes [185,196,197,202] agree with both the rapid radiation inferred from nuclear gene sequences [28,45] and the reconstruction of historical biogeography for both Viola (Figure 8) and Violaceae [4].
The perceived absence of Viola fossils in South America must be seen in the light of fossil recovery rates not being constant in time and space and across lineages. In fact, the exception in Violaceae is the “burst” in occurrence rates of Viola fossils in Eurasia from 17–18 Ma. Apart from that, the fossil record of Violaceae is practically non-existent. There are for instance no records from North America from the same period even though we must assume that Viola was present there. Possible explanations for the lack of identified, older Viola fossils in South America, despite the existence of geological formations of an appropriate age (e.g., Abanico from Eocene-Miocene, Río Turbio from Eocene-Oligocene, La Leona from Oligocene, etc.), include possibly low fossilisation probabilities due to Viola growing far from the fossilisation sites, and a lack of paleobotanical studies enforced by the absence of reference anatomical studies on extant taxa.
2.3. The “Known Unknowns”: Outstanding Research in Viola
The level of knowledge of the genus Viola has a strong geographic bias towards the northern hemisphere, primarily Europe, where taxonomic research has the longest history and where taxa have been most intensively studied. This has resulted in a “eurocentric” understanding of the diversity of the genus, its evolution, and its classification. The most significant gaps in our knowledge of Viola are for the South American taxa, notably subg. Neoandinium, for which classification, diversity and phylogeny are still poorly (or not) understood, all being based on morphological characters and geography. Because Viola originated in South America, understanding the evolutionary patterns here is key to understanding patterns within the genus as a whole.
This is the first, comprehensive taxonomy for Viola in the last 97 years, since that of Becker (1925 [1]). It is beyond doubt that the century-long absence of systematised information that an updated classification would have represented has hindered the formation and testing of new hypotheses—and therefore accumulation of new knowledge. Below we discuss the most imminent gaps in our knowledge of Viola.
2.3.1. Phylogeny of Viola
Phylogenetic data are completely lacking for the monotypic sect. Nematocaulon from New Zealand (V. filicaulis), sect. Xanthidium (V. flavicans) from South America, both in subg. Viola, and for most of subg. Neoandinium from South America. As subg. Neoandinium comprises a minimum of 140 known species and currently makes up some 21% of the diversity within the genus, this is beyond comparison the biggest knowledge gap within the genus. In addition, a large proportion of the species are narrow endemics that are critically endangered [68]. The monotypic sect. Danxiaviola is known from ITS and chloroplast sequences only which means that its ploidy and exact placement within the polyploid CHAM × MELVIO tangle remain unknown. While the occurrence of the polyploid CHAM × MELVIO tangle in the Northern Hemisphere has been well established, the same can not be said about the occurrence of similar tangles in the southern hemisphere involving the polyploid sections Chilenium, Tridens, Leptidium, Erpetion, and probably also Nematocaulon and Xanthidium. For these taxa inference of the species-level phylogeny in the study of Marcussen et al. [28] was rendered difficult by gene duplication and loss, even though three low-copy nuclear genes were used, and the lack of supporting data on chromosome numbers and ploidy. Though there is a large number of chromosome counts within the species-rich and probably also highly polyploid sect. Melanium, these numbers do not allow for reliable inferences on ploidy level in particular taxa. This lack of knowledge is combined with very limited information about the phylogeny of this section as the phylogenetic analyses, using a combination of ITS and ISSR markers [217] and more recently a combination of nuclear ITS and ETS and plastid trnS–trnG intergenic spacer sequences [94], have yielded poor resolution.
2.3.2. Chromosome Counts and Ploidy
Chromosome number is an important taxonomic character and also gives information on ploidy. Chromosome counts are completely lacking for the sections Chilenium, Melvio, Spathulidium, and Xanthidium, and for most of subg. Neoandinium. Numerous other sections are represented only by a single count that is in need of confirmation (i.e., sects. Abyssinium, Danxiaviola, Erpetion, Himalayum, Leptidium, Nematocaulon, and Rubellium). Genome size has been measured by flow-cytometry mainly on European taxa [218,219,220,221] but is ploidy-informative within sections only.
2.3.3. Fossil Record
Despite Viola having a rich seed fossil record from the Miocene (18–17 Ma) onwards of Europe and western Siberia, interpretations on phylogeny, evolution, and biogeography are limited by the lack of detailed knowledge of variation and apomorphies among extant species and sections of the genus, e.g., based on SEM micrographs. To this date, the only comparative study of seed morphology [213] covered only parts of the European territory and taxa and did not use SEM. Furthermore, the seed fossil record outside of western Eurasia is limited to unconfirmed records from the Pliocene and Pleistocene of North America, and there are no seed fossil records for Viola in South America although the genus has its longest history there. There are several geological formations in or near the Andes with fossiliferous horizons assignable to the Eocene-Oligocene boundary onwards. However, there are also no comprehensive studies on the morphology and anatomy of pollen, seeds, and other plant structures on the extant South American species of Viola that can serve as a solid basis for fossil surveys.
2.3.4. Alpha Taxonomy
In recent years, a better understanding has been acquired of difficult groups such as subg. Neoandinium in South America (e.g., [68,80,222,223,224]), sect. Nosphinium subsect. Borealiamericanae in North America [67,214,225], sect. Erpetion in Australia [98,160,177,226,227], as well as the genus as a whole in China [76,78]. The last remaining blank spot seems to be the southeastern Asian and Malayan species, which comprises relatively few, but morphologically specialised and probably not closely related species that do not fit seamlessly with the taxonomic system, as indicated by the few treatments available [74,228,229,230,231].
2.3.5. Transcriptomes and Genomes
Thus far, reference sequence genome has been published for the diploid Viola (sect. Chamaemelanium) pubescens [232] and the octoploid V. (sect. Himalayum) kunawurensis (as V. “kunawarensis”; NCBI accession PRJNA805692), but numerous Viola genomes are planned sequenced by the Earth Biogenome Project during the next decade [233]. Transcriptomes have been published for at least the four most widespread sections within subg. Viola, i.e., sects. Chamaemelanium, Melanium, Plagiostigma, and Viola (e.g., [234,235,236]), but to date no transcriptomes exist for taxa from outside of Eurasia and North America.
2.4. Taxonomic Treatment of Viola
Viola
Description.—Annual or perennial acaulescent or caulescent herbs, shrubs or very rarely treelets. Axes morphologically differentiated or not. Stipules free or adnate, small or foliaceous, margin entire, laciniate, dentate, or fimbriate. Lamina linear to reniform, more or less petiolate, margin entire, crenulate, serrate, pinnate, or pedate. Flowers axillary and solitary, rarely in cymes. Peduncle non-articulated, lacking an abscission zone at the level of the bracteoles. Corolla white to yellow, orange or violet or multicoloured with or without yellow throat, strongly zygomorphic. Calycine appendages present. Bottom petal slightly to much shorter than others and weakly differentiated, rarely larger than others. Spur scarcely exserted to very long, rarely absent. Filaments free, two lowest stamens calcarate, dorsal connective appendage large, oblong-ovate, entire. Style filiform, clavate, or capitate, variously crested or not, bearded or not, often rostellate at tip. Capsule thick-walled. Seeds few to many per carpel, obovoid to globose, often arillate. Cleistogamous flowers often produced. Base chromosome numbers x = 6, 7.
Diagnostic characters.—Flowers axillary and solitary AND peduncle non-articulated AND plant herbaceous AND temperate distribution AND bottom petal slightly to much shorter than others and weakly differentiated.
Ploidy and accepted chromosome counts.—2x, 4x, 6x, 8x, 10x, 12x, 14x, 16x, 18x, 20x, >20x. 2n = 4, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 34, 36, 40, 44, c. 44, 46, 48, 50, 52, 54, 58, 60, c. 64, 72, 76, 80, c. 80, 82, c. 96, 102, c. 120, 128.
Age.—Crown node age 30.9 (29.8–31.3) Ma [28].
Included species.—664.
Distribution.—Temperate regions and montane areas in the tropics worldwide; all continents except Antarctica (Figure 2).
Discussion.—The two main lineages of Viola are here treated as subgenera, Neoandinium and Viola. The two subgenera differ rather consistently in aspects of growth form, leaf shape, degree of emargination of the bottom petal, shape of the anther appendages, style shape, and also in base chromosome number for the diploids investigated so far. Reiche [48,122,130] was the first to notice the fundamental distinction between these two sublineages of the genus. He recognised three sections, the first corresponding to subg. Viola (as sect. Sparsifoliae), the second to subg. Neoandinium (as sect. Rosulatae), and a third small section with four deviant taxa from both subgenera (sect. Confertae) [48]. Becker [1], however, treated subg. Neoandinium as one of 14 sections of the genus (as sect. Andinium).
2.4.1. Key to the Subgenera, Sections, and Subsections of Viola
Conventions and definition of terms:
- An “M” dash (“—”) is used to identify uncommonly expressed traits/separate characters that have no counterpart in the antithesis.
- Arosulate acaulescent: with leaves scattered on stem, not in rosettes. Aerial stems and stolons (e.g., V. filicaulis).
- Arosulate caulescent: with leaves on aerial stems. Rosettes and stolons absent (e.g., V. abyssinica, V. arborescens, V. stagnina).
- Beard: tuft of hairs on the lateral petals (and sometimes upper or bottom petals) located at the throat of the chasmogamous flower, also a tuft of trichomes near the apex of the style in some species or groups. Organs with or without a beard are referred to as bearded or glabrous, respectively.
- Calycine appendage: Appendage at base of the sepal; synonymous with “sepal auricle” or “sepal appendage”.
- Caulescent/acaulescent: with/without aerial stems.
- Flower colour: base colour of the petals in living plants excluding the nectar guides, unless otherwise noted.
- Foliaceous: used to describe stipules that are green and often large and leaf-like (e.g., V. elatior, V. raddeana, V. tricolor).
- Papilla: lateral expansion of the cell wall to form a short conical structure up to 3 times as long as wide. For instance, a pad of papillae is found on the lateral petals of sect. Erpetion in place of a beard of trichomes exhibited in some other lineages.
- Rosulate/arosulate: with/without leaves in rosette.
- Rosulate acaulescent: with leaves in rosettes. Aerial stems and stolons absent (e.g., V. hirta, V. pedata, V. selkirkii).
- Rosulate caulescent: with leaves in rosettes, aerial stems present. Stolons absent (e.g., V. canadensis, V. riviniana).
- Rosulate stoloniferous: with leaves in rosettes, stolons present. Aerial stems absent (e.g., V. banksii, V. odorata, V. palustris).
- Stolon: lateral, specialised procumbent stem producing adventitious roots and new plantlets. We restrict the term to taxa in which the shoot axes are differentiated.
- Trichome: elongate hair-like structure usually more than 3 times as long as wide and typically linear or distinctly broader above the base.
- Violet: colour of the corolla and petal striation in many species. In the literature, this colour is often referred to, rather ambiguously, as “blue” or “purple”.
- 1a.
- Herbs, usually forming subacaulous imbricate or loose rosettes, very rarely erect-cauline, rarely woody based, or dwarf ericoid shrublets. Margin of juvenile laminas flat, not involute. Peduncle shorter or as long as mature lamina. Bottom petal usually cleft, more rarely emarginate or entire. Nectariferous appendage of the two bottom stamens filiform. Style at apex capitate, beardless, usually crested; crest 1–3 lobes or flanges at sides or top of style apex, or a continuous sharp dorsolateral rim, very rarely crest absent. Cleistogamous flowers not produced. (Subg. Neoandinium) ................................................................................................................................................................................................................................................................................................. 2.
- 1b.
- Herbs, subshrubs or shrubs, with leaves scattered on stem or in rosette, rarely cushions with imbricated distichous leaves (sect. Tridens). Margin of juvenile laminas usually involute. Peduncle often longer than mature lamina. Bottom petal entire or emarginate, very rarely cleft. Nectariferous appendage of the two bottom stamens various in shape, very rarely filiform. Style filiform, clavate or (sub)capitate, not crested (lateral lobes present: sect. Sclerosium) but top of style apex often flattened or concave with more or less raised edges, sometimes bearded. Cleistogamous flowers often produced. (Subg. Viola) .............................................................................................................................................................................................................................................................................................. 13.
- 2a.
- Underground part of stems conspicuously elongated, leafless and stolon-like, branching or not. ........................................................................................ sect. Rhizomandinium
- 2b.
- Stems without basal stolon-like segment. ........................................................................................................................................................................................................................ 3.
- 3a.
- Leaves glabrous, except occasionally for minute cilia on margins, rarely glabrescent or pubescent. Lamina usually more or less rigid, thick or coriaceous; margins usually entire, rarely crenulate. ....................................................................................................................................................................................................................................................... 4.
- 3b.
- Leaves with indumentum, or if glabrous, then with prominently raised veins above. Lamina flexible, thick or thin; margins usually crenate or incised, rarely entire. ...... 7.
- 4a.
- Plant a dwarf ericoid shrublet. ...................................... ................................................................................................................................................sect. Ericoidium (V. fluehmannii)
- 4b.
- Plants other. ......................................................................................................................................................................................................................................................................... 5.
- 5a.
- Plant caulescent. ............................................................................................................................................................................................................... sect. Confertae (V. nassauvioides)
- 5b.
- Plants subacaulous, rosulate. ............................................................................................................................................................................................................................................. 6.
- 6a.
- Bottom petal longer than or equal to the other petals. ........................................................................................................................................................................ sect. Sempervivum
- 6b.
- Bottom petal much shorter than the other petals. ............................................................................................................................ sect. Inconspicuiflos, in part (V. membranacea)
- 7a.
- (4). Style crest as one apical and two lateral lobes. .............................................................................................................................................................................. sect. Triflabellium
- 7b.
- Style crest lateral, or lateral and frontal, or apical only, or a sharp dorsolateral rim. .................................................................................................................................................. 8.
- 8a.
- Plant with short woody aerial stems. ........................................................................................................................................................................................... sect. Xylobasis (V. beati)
- 8b.
- Plants completely herbaceous............................................................................................................................................................................................................................................ 9.
- 9a.
- Corolla large, four times wider than lamina width or more. ................................................................................................................................................................. sect. Grandiflos
- 9b.
- Corolla small, usually as wide or up to twice as wide as lamina width, exceptionally up to four times wider than lamina width. ................................................................................................................................................................................................................................................................................................ 10.
- 10a.
- Cilia long, surrounding entire lamina margin, strongly deflexed. ....................................................................................................................................................... sect. Relictium
- 10b.
- Cilia short, more or less patent. ..................................................................................................................................................................................................................................... 11.
- 11a.
- Bottom petal much smaller than the other petals. .............................................................................................................................................. sect. Inconspicuiflos, in largest part
- 11b.
- Bottom petal not smaller than the other petals.............................................................................................................................................................................................................. 12.
- 12a.
- Annuals. Lamina linear, oblanceolate or obovate; margin entire or shallowly and remotely crenulate. ................................................................................................................................................................................................................................................................. sect. Subandinium
- 12b.
- Annuals or perennials. Lamina elliptical, narrowly to broadly obovate, orbicular, or rhomboid; margin deeply to shallowly crenate, sinuate, incised, pinnatifid, or rarely entire when plant perennial. ...................................................................................................................................................................................................................... sect. Rosulatae
- 13a (1).
- Style slender and slightly clavate, with a pair of apical or subapical lateral lobes. Corolla white to violet with yellow-green throat. Stipules minute. Annual herbs or subshrubs. (northeastern Africa, southern and eastern Arabia, southwestern Asia) ...................................................................................................................sect. Sclerosium
- 13b.
- Style tubular, clavate or (sub)capitate, lacking lateral processes, but sometimes at apex bearded, or margined with +/− raised edges, or bilobate. Corolla yellow throughout, or white to violet with syncolorous or yellow-green throat, or multicoloured. Stipules prominent. Perennial herbs, sometimes annuals (in sect. Melanium), occasionally shrubs or subshrubs. (more widely distributed) .................................................................................................................................................................................. 14.
- 14a.
- Bottom petal (excluding spur) more than twice as long and broad as lateral and upper petals. Subshrub. (southern China: Guangdong) ....................................................................................................................................................................................................................................... sect. Danxiaviola (V. hybanthoides)
- 14b.
- Bottom petal (excluding spur) subequal to somewhat smaller or larger than lateral and upper petals. Herbs, subshrubs or shrubs. (not restricted to China) .................. 15.
- 15a.
- Spur 12–30 mm long. Petals pink to magenta. Arosulate caulescent. Leaves sessile, seemingly ternate to palmate, with 3–5 lanceolate, entire segments (lamina and 2 or 4 stipule segments similar). (southern Europe) .................................................................................................................................................................................. sect. Delphiniopsis
- 15b.
- Spur <20 mm long (to 16 mm in sect. Melanium and in subsect. Rostratae). Petals of various colours, very rarely pink to magenta. Rosulate or arosulate, caulescent or acaulescent. Leaves commonly petiolate. (not restricted to southern Europe) ..................................................................................................................................................... 16.
- 16a.
- Lamina subulate, somewhat succulent, margin entire. Style sigmoid, dorsiventrally flattened at base, tapering in width and becoming filiform towards apex, with an apical stigmatic opening. Subshrub. (South Africa) ......................................................................................................................................................... sect. Melvio (V. decumbens)
- 16b.
- Lamina broader, margin usually crenate. Style filiform, clavate or capitate. Herbs, subshrubs, or shrubs. (not South Africa) .......................................................................... 17.
- 17a.
- Style filiform, protruding, straight or somewhat geniculate at base, with an apical stigmatic opening. Spur reduced to a swelling (gibba), or short, 0.5–1.5 mm, as long as tall (spur 4 mm long and as tall, half length of petal blade, and corolla bright red: V. (sect. Leptidium) arguta). Herbs, more rarely subshrubs or shrubs. ........................ 18.
- 17b.
- Style clavate or (sub)capitate, monosymmetric (style filiform and spur 4–9 mm long: V. (sect. Viola) papuana). Spur well developed, as long as tall or longer. Herbs or subshrubs............................................................................................................................................................................................................................................................................ 21.
- 18a.
- Leaves 2–10 mm long. Petiole indistinct. Lamina obovate, at apex tridentate, sometimes bilobate or entire. Phyllotaxis distichous. (s South America) ......................... ..................................................................................................................................................................................................................................................... sect. Tridens (V. tridentata)
- 18b.
- Leaves >10 mm long. Petiole distinct. Lamina of various shapes, crenate. Phyllotaxis polystichous. .................................................................................................................... 19.
- 19a.
- Stipules long, densely short-fimbriate, broad and sheathing the stem. Subshrubs or herbs. Arosulate caulescent, with reclining or weakly ascending to erect stems. Corolla with a white throat, rarely throat red (V. arguta). (Latin America) ......................................................................................................................................... sect. Leptidium
- 19b.
- Stipules rather small, entire or sparingly lacerate to laciniate with few long processes, not sheathing the stem. Herbs. Rosulate stoloniferous or arosulate acaulescent. Corolla with a yellow throat or with a green blotch on bottom petal. ..................................................................................................................................................................... 20.
- 20a.
- Stem creeping, remotely noded, branched. Stolons absent. Corolla with a yellow throat. Spur distinct, 0.5–1.5 mm long, yellow. Lateral petals sparsely bearded. Cleistogamous flowers produced. (New Zealand) ................................................................................................................................................... sect. Nematocaulon (V. filicaulis)
- 20b.
- Stem usually densely noded (usually rosettes). Stolons present, sympodial. Corolla without a yellow throat, but bottom petal with a green blotch inside. Spur absent, reduced to a swelling (gibba). Lateral petals with a broad dense pad of papillae. Cleistogamous flowers not produced, but some species have flowers with a small corolla. (Australia) ......................................................................................................................................................................................................................................... sect. Erpetion
- 21a
- (17). Corolla white on the inside, rarely pale violet, lacking violet striation. Shrubs, usually with lateral, leafless 1–few-flowered inflorescences, rarely herbs with solitary flowers (V. kauaensis). (Hawaiian Islands).—Lower stipules ovate or triangular, partially sheathing the stem. Style apex with weak subapical dorsolateral swelling (where distinct rim occurs in several other groups), rostellum formed by bent apex tall and blunt at tip. ....................................................... sect. Nosphinium subsect. Nosphinium
- 21b.
- Corolla variously coloured, usually with violet striation. Herbs or subshrubs. Flowers solitary, not in inflorescences. (not Hawaiian Islands) ............................................................................................................................................................................................................................................................................................. 22.
- 22a.
- Small subshrubs. Lamina lanceolate or spathulate. .................................................................................................................................................................................................... 23.
- 22b.
- Herbs, sometimes with a woody rhizome. Lamina shape and style shape variable. Cleistogamous flowers produced or not. ....................................................................... 24.
- 23a.
- Leaf base decurrent. Petiole indistinct. Stipules entire or with one or two basal segments, sometimes foliaceous. Corolla violet, white or yellow. Style apex scarcely to weakly bent ventrad. (Mediterranean) ................................................................................................................................................................................................. sect. Xylinosium
- 23b.
- Leaf base cuneate. Petiole distinct. Stipules small, bract-shaped, fimbriate. Corolla violet or magenta. Style apex strongly bent ventrad or with stigma on ventral side. (Chile) ........................................................................................................................................................................................................................................................... sect. Rubellium
- 24a.
- Corolla with a yellow throat. Petals yellow or variously coloured. Style clavate or (sub)capitate. (Throat white, corolla white with reddish-violet striation, stipules free, spur as long as tall, style more or less filiform: V. commersonii. Throat white or cream, petals violet, style capitate: V. nummulariifolia, V. cornuta, V. orthoceras.) .............. 25.
- 24b.
- Corolla with a white or cream, violet, or yellowish-green throat. Petals usually violet or white, occasionally pink, never yellow. Style clavate or cylindrical, rarely filiform, never capitate. .................................................................................................................................................................................................................................................... 32.
- 25a.
- Usually caulescent. Perennial or annual. Stipules entire or with a few irregular teeth, or deeply pinnatifid. Petals yellow or variously coloured. Style usually capitate and bearded. (Northern Hemisphere, naturalised elsewhere) ........................................................................................................................................................................................... 26.
- 25b.
- Acaulescent. Perennial. Stipules glandular-lacerate to glandular-laciniate. Petals yellow (white in V. (sect. Chilenium) commersonii). Style usually concave or flattened at apex, glabrous or bearded. (Style ellipsoid with broadly rounded apex when fresh in sect. Xanthidium, bearded: V. flavicans.) (South America) ......................................... 31.
- 26a.
- Perennial. Rosulate, caulescent, rarely stoloniferous (V. kusnezowiana) or acaulescent (V. barroetana). Perennating stem a monopodial rhizome, often deeply buried. Stipules not distinctly foliaceous, margins entire or with 1–2(–4) irregular shallow teeth on either margin. Spur usually very short to short (less than twice as long as tall), rarely longer (in 2 Asian species). Calycine appendages short (<2 mm). Bottom petal (including spur) typically <15 mm. Style various at apex, often (sub)capitate and bearded, occasionally bifid, but lacking shallow reflexed lateral lobes. Lamina margin subentire, crenate, lobed or divided.—................................... sect. Chamaemelanium
- 26b.
- Perennial to annual. Arosulate caulescent, sometimes indistinctly caulescent (V. alpina). Perennating stem a sympodially branching pleiocorm. Stipules usually large and foliaceous, pinnatifid or palmately divided, rarely small with entire or dentate margins. Spur very short to very long (0.9–16 mm, often much longer than tall). Calycine appendages very short to very long (0.3–4.7 mm). Bottom petal (including spur) 2–34 mm. Style capitate and bearded at apex, with a pair of inconspicuous or prominent shallow reflexed lateral lobes. Lamina margin entire or crenate, never lobed or deeply divided. (sect. Melanium) ...................................................................................................................................................................................................................................................................................... 27.
- 27a.
- Cleistogamous flowers produced in summer. Annual or biennial. (eastern North America) ........................................ sect. Melanium subsect. Cleistogamae (V. rafinesquei)
- 27b.
- Cleistogamous flowers not produced. Annual to perennial. (Palaearctic, naturalised elsewhere) ......................................................................................................................... 28.
- 28a.
- Corolla violet, with a cream-coloured throat. Stipules ovate-lanceolate, dentate. Bottom petal 9.5–10.5 mm. Low, high-Alpine perennial. (southwestern Alps and Corsica) ......................................................................................................................................................................................... sect. Melanium subsect. Pseudorupestres (V. nummulariifolia)
- 28b.
- Corolla colour various, often yellow or violet, with a bright yellow throat (if throat cream or white, then lateral petals directed horizontally or downwards: V. cornuta and V. orthoceras). Stipules variable, often foliaceous, rarely dentate. Bottom petal 2–34 mm. Annual or perennial. ......................................................................................... 29.
- 29a.
- Annual. Basal leaves entire or indistinctly crenulate. Bottom petal 2–11.5 mm. Spur 0.9–3 mm. ............................................................. sect. Melanium subsect. Ebracteatae
- 29b.
- Annual to perennial. Leaves crenate or entire, but in annual species basal leaves crenate. Bottom petal 5–34 mm. Spur 1–16 mm. ................................................................. 30.
- 30a.
- Calycine appendages 0.3–1.0 mm. Bottom petal 5–13 mm. Spur 1–3.5 mm. (Mediterranean area) .................................................................. sect. Melanium subsect. Dispares
- 30b.
- Calycine appendages 0.9–4.7 mm. Bottom petal 5.4–34 mm. Spur 1.8–16 mm. ............................................................................................ sect. Melanium subsect. Bracteolatae
- 31a.
- Rosulate, perennating stem a short monopodial rhizome. Style ellipsoid and broadly rounded at apex when fresh, when dried clavate with flattened apex, bearded (V. flavicans) or at most occasionally papillate (V. pallascaensis). Stipules adnate at base or for most of their length, narrow, shallowly glandular-lacerate. Bracteoles narrow, shallowly glandular-lacerate. ................................................................................................................................................................................................................. sect. Xanthidium
- 31b.
- Variably rosulate or arosulate, perennating stems multiple, elongate and deeply buried. Style clavate or straight fresh or dried, apex concave, flattened or slightly acute with sharp dorsolateral rim, sometimes with a short subapical ventrad or incurved rostellum bearing the stigma, usually beardless (white-hairy in V. rudolphii). Stipules free, broad, deeply glandular-laciniate, rarely entire. Bracteoles broad, deeply glandular-laciniate. ............................................................................................. sect. Chilenium
- 32a
- (24). Stipules adnate at least in the lower 1/3, rarely in the lower 1/4 or less (subsect. Clausenianae). Rosulate acaulescent, estoloniferous. ..................................................... 33.
- 32b.
- Stipules free or adnate at base only. Rosulate caulescent, rosulate stoloniferous, rosulate acaulescent, or arosulate caulescent. (Stipules partly adnate and plant stoloniferous: subsect. Bulbosae and V. (subsect. Rostratae) uliginosa. Stipules (1/2–)2/3 or more adnate and flowers white with bottom petal blade densely striated, spur shorter than tall: V. (subsect. Mexicanae) humilis.) ....................................................................................................................................................................................................... 34.
- 33a.
- Lamina of various shape but not spathulate, undivided with margin crenulate or serrate, or incised to dissected. ......................................................................................... 34.
- 33b.
- Lamina spathulate, undivided, margin entire or indistinctly and remotely crenulate (southern and western Asia). .......................................................................................... 36.
- 34a.
- Lamina deeply pedately dissected. Calycine appendages entire. Spur short, as long as tall. Style with long dorsolateral margin closely following style body as a narrowly rounded rim running laterally and ventrally at an acute angle from dorsum of apex to a more proximal point on the ventral surface, the stigma hidden in the narrow cavity created by the rim. Cleistogamous flowers not produced. .................................................................................................... sect. Nosphinium subsect. Pedatae (V. pedata)
- 34b.
- Lamina undivided, incised to pinnatifid, or ternately to triternately dissected. Calycine appendages dentate or entire. Spur as long as tall or longer. Style with dorsolateral margin obsolete, or short and more or less perpendicular to dorsum, or produced as a thick or swollen continuous rim at an acute angle from dorsum of apex to the centre of the ventral surface. Cleistogamous flowers produced. ............................................................................................................................................................................................................................................................................................. 35.
- 35a.
- Stipules adnate in lower 1/4, margins glandular-lacerate. Calycine appendages short, triangular, narrowly rounded at apex, entire. Spur short, as long as tall. Style apex protruded dorsally as a thickened broadly truncate or slightly emarginate rim, continuous laterally and ventrally at an acute angle from dorsum of apex to a proximal point on the ventral surface, ending in a strongly incurved rostellum. Lamina deltoid-triangular. (western North America: Utah) .............................................................................................................................................................................................. sect. Nosphinium subsect. Clausenianae (V. clauseniana)
- 35b.
- Stipules 1/3–3/4 adnate to petiole, margins entire or indistinctly crenulate. Calycine appendages short or elongated, usually oblong, truncate or emarginate at apex, usually dentate. Spur longer than tall, usually 1/5 to 1/2 of total length of bottom petal, 2–10 mm. Style apex with dorsolateral margin obsolete, or dorsolateral margin slightly thickened or produced as a pair of short lobes more or less perpendicular to dorsum but not continuous laterally to the straight ventrad rostellum. Lamina of various shape, undivided, deeply incised, lobed or dissected. (not restricted to North America) ................................. sect. Plagiostigma subsect. Patellares, in largest part
- 36a
- (33). Petiole indistinct, about as long as lamina. Style apex with thickened dorsal margin and a ventral rostrum. Cleistogamous flowers not produced. Spur 1.5–4 mm, longer than tall. (southwestern Asia) .................................................................................................................................................................................................. sect. Spathulidium
- 36b.
- Petiole distinct, at least twice as long as lamina. Style lacking distinct margins. Cleistogamous flowers produced. ........................................................................................................................................................................................................................................................................................... 37.
- 37a.
- Spur c. 1.5 mm, as long as tall. Plant with stems subterranean from deeply buried rhizome, appearing aboveground as proximal or tufted rosettes. (Rim of the Tibetan Plateau) ........................................................................................................................................................................................................................ sect. Himalayum (V. kunawurensis)
- 37b.
- Spur 3–7.5 mm, longer than tall. Rhizome usually at soil surface, with leaf rosette. ...... Sect. Plagiostigma subsect. Patellares, in part (V. alaica, V. dolichocentra, V. turkestanica)
- 38a
- (32). Spur longer than tall. ........................................................................................................................................................................................................................................39.
- 38b.
- Spur shorter than tall. ................................................................................................................................................................................................................................................ 42.
- 39a.
- Bottom petal (excluding spur) conspicuously longer than the other petals, emarginate, 6–11 mm. Stolons leafless, terminated by a leafy rosette. (Taiwan, Ryukyu islands) .......................................................................................................................................................................................................................... sect. Plagiostigma subsect. Formosanae
- 39b.
- Bottom petal (excluding spur) not longer than the other petals. Stolons, if present, with scattered leaves along the length. (not restricted to southeastern Asia) ........... 39.
- 40a.
- Spur saccate, less than twice as long as tall. Calycine appendages very short or obsolete, 0–0.5 mm. Arosulate caulescent. Stems creeping to reclining or suberect, proximally rooting. Style clavate, apex sharply bent 90° ventrad into a prolonged rostrum, beardless. Cleistogamous flowers not produced. (Africa) .........sect. Abyssinium
- 40b.
- Spur not saccate, pronounced to very long, (much) more than twice as long as tall, 2–20 mm long. Calycine appendages short or long, >0.5 mm long. Rosulate caulescent, rosulate stoloniferous, rosulate acaulescent, or arosulate caulescent. Style cylindrical or subclavate, apex straight to slightly curved or abruptly bent ventrad, bearded or beardless above. Cleistogamous flowers usually produced (sect. Viola). ............................................................................................................................................................. 41.
- 41a.
- Capsule trigonous-ellipsoid, usually glabrous, forcibly ejecting the seeds after dehiscence, borne on erect peduncles at maturity. Style often bearded above, nearly straight to weakly bent at apex with rostellum. Usually rosulate caulescent, more rarely rosulate stoloniferous, rosulate acaulescent, or arosulate caulescent. .................. ............................................................................................................................................................................................................................................... sect. Viola subsect. Rostratae
- 41b.
- Capsule globose, usually hairy, non-dehiscent, borne on decumbent to prostrate peduncles at maturity (cleistogamous flowers and capsules often underground). Style beardless, often strongly bent at apex with pronounced rostrum. Rosulate acaulescent or rosulate stoloniferous. ................................................... sect. Viola subsect. Viola
- 42a
- (38). Corolla pale pink or pale violet, rarely white. Bottom petal 2.5–12 mm long (including spur), conspicuously shorter and narrower than the others, usually acute, with distinct violet striation or reticulation. Style apex bilobate. Stipules linear to broadly lanceolate, densely or remotely fimbriate, free or 1/3 adnate. Stolons produced. .............................................................................................................................................................................................................................................................................................. 43.
- 42b.
- Corolla white or violet, occasionally pink. Bottom petal 7–25 mm (including spur), not usually conspicuously smaller than the others. Style apex bilobate or distinctly margined. Stipules lanceolate to ovate, entire or remotely denticulate to fimbriate-dentate, free or adnate. Stolons produced or not. ........................................................... 44
- 43a.
- Lateral petals not bearded. Peduncles glabrous; plant usually glabrous or nearly so. Rhizome long and remotely noded or short and densely noded. Stolons present or rarely absent, with (many) scattered leaves. Stipules free or adnate at base only, often brownish, long-fimbriate to laciniate. Corolla usually pale violet to whitish, without a greenish throat. Perennials. ............................................................................................................. sect. Plagiostigma subsect. Australasiaticae
- 43b.
- Lateral petals usually bearded. Peduncles with patent hairs, rarely glabrous (in V. nanlingensis); plant usually hairy. Rhizome short, densely noded. Stolons with 1–2 (smaller) leaves and a leaf rosette at apex. Stipules adnate in the lower 1/3 (stipules on stems free in V. guangzhouensis), remotely or rarely densely fimbriate. Corolla usually pale pink to pale violet, with a greenish throat. Perennials or rarely annuals (V. diffusa). ......................................................... sect. Plagiostigma subsect. Diffusae
- 44a.
- Bottom petal 7–12 mm including the spur. Corolla usually white with violet striation. Style strongly bilobate or distinctly margined all around. ...................................... 45.
- 44b.
- Bottom petal 12–25 mm including the spur. Corolla violet, rarely white (V. grahamii, some V. hookeriana, some V. moupinensis, V. oxyodontis, V. brevipes, some V. thomsonii) or rose-violet (V. rossii). Style with weak to pronounced dorsolateral rim or not, not strongly bilobate. .............................................................................................................. 47.
- 45a.
- Stem vertical, growing from underground bulbil. Stolons underground, branched, leafless, with cleistogamous flowers. Outer stipules adnate, inner stipules free.—Style bilobate. ................................................................................................................................................................................................................. sect. Plagiostigma subsect. Bulbosae
- 45b.
- Bulbils absent, rhizome oblique to vertical. Stolons different than above, or absent. Stipules usually free. ............................................................................................................................................................................................................................................................................................ 46.
- 46a.
- Lateral stems creeping, ascending or erect. Stipules green, margins entire, remotely denticulate, or 1–3-toothed on either side, teeth eglandular. Style apex bilobate. ................................................................................................................................................................................................................................. sect. Plagiostigma subsect. Bilobatae
- 46b.
- Lateral stems absent, or present as stolons. Stipules membranous, glandular-lacerate. Style apex margined or rarely bilobate. .................................................................................................................................................................................................. sect. Plagiostigma subsect. Stolonosae, in largest part
- 47a
- (44). Lateral petals glabrous, rarely with a few hairs. Calycine appendages dentate or entire. Style with or without a distinct dorsolateral rim, if present this short and weakly spreading or oriented apically, usually not extending much laterally. .............. sect. Plagiostigma subsect. Stolonosae, in part (V. suecica, V. bissettii, V. brevipes, V. diamantiaca, V. epipsila, V. moupinensis, V. palustris, V. pluviae, V. rossii, V. thomsonii, V. vaginata)
- 47b.
- Lateral petals densely bearded (glabrous or sparsely bearded in certain species of subsect. Mexicanae). Calycine appendages entire (dentate in a few species of subsect. Borealiamericanae). Style apex sharp-edged without a distinct dorsolateral rim or with a pronounced and thickened spreading rim commonly extending laterally to the rostellum. ........................................................................................................................................................................................................................................................................ 48.
- 48a.
- Aerial stems present. Lower stipules ovate, shallowly glandular-fimbriate, sheathing the stem. Style apex broadly rounded, with or without a weak dorsal or dorsolateral swelling in place of a distinct rim. (Amphiberingian) .................................................................................................. sect. Nosphinium subsect. Langsdorffianae
- 48b.
- Aerial stems absent. Stipules linear-lanceolate to lanceolate (ovate and glandular-laciniate in V. guatemalensis and V. nubicola of subsect. Mexicanae). Style apex abruptly flattened or concave with a sharp edge, or flanked by a prominent truncate to emarginate or bilobate spreading to dorsad thickened rim. .................................................................................................................................................................................................................................................................................... 49.
- 49a.
- Stolons present or absent, if absent then lateral petals glabrous or sparsely bearded. Lateral petals glabrous or sparsely bearded (densely bearded in stoloniferous white-flowered V. grahamii and V. oxyodontis). Corolla white or violet, rarely dark violet (V. beamanii). Stipules in some species basally or mostly adnate. Calycine appendages short and entire. Bottom petal glabrous. Style apex merely sharp-edged or scarcely thickened, apically oriented or slightly inrolled, not prolonged, not strongly thickened or spreading and not extending much laterally (somewhat prolonged and slightly thickened dorsally in V. hookeriana). (Mexico to northern South America) ........................................................................................................................................................................................................................... sect. Nosphinium subsect. Mexicanae
- 49b.
- Stolons absent. Lateral petals densely bearded. Corolla violet to dark violet. Stipules free. Calycine appendages short or elongated, entire or dentate. Bottom petal glabrous or bearded. Style apex with pronounced thickened ascending to spreading rounded or strongly emarginate dorsolateral rim extending ventrad partly or fully to rostellum. (North America, V. nuevoleonensis in northern Mexico) ............................................................................................ sect. Nosphinium subsect. Borealiamericanae
2.4.2. Descriptions of Subgenera, Sections, and Subsections of Viola
Viola subg. Neoandinium Marcussen, Nicola, Danihelka, H. E. Ballard, A. R. Flores, J. S. Watson, subg. nov.—Type: Viola rosulata Poepp. & Endl.
Description.—Perennial or annual herbs, usually forming subacaulous imbricate or loose rosettes, very rarely either caulescent, woody based, or dwarf ericoid subshrublet (in sect. Ericoidium). Axes not morphologically differentiated. Stems vertical, branched or not, occasionally arising from a buried branching “rhizome” (stolon-like persistent axes). Stipules inconspicuous or sometimes absent. Lamina usually spathulate, tapering into the petiole (pseudopetiole); margin entire, hyaline, crenulate, or lobed to pinnate; margin of juvenile laminas flat, not involute. Peduncle shorter or as long as mature laminas. Bottom petal usually cleft, rarely emarginate or entire. Spur present or rarely absent. Nectariferous appendage of the two bottom stamens filiform. Style at apex capitate and crested; crest 1–3 lobes or flanges at sides or top of style apex, or a continuous sharp dorsolateral rim, very rarely crest absent. Cleistogamous flowers not produced. Diploid. Base chromosome number x = 7.
Diagnostic characters.—Margin of juvenile laminas not involute OR peduncles not longer than mature leaves OR style capitate and crested OR cleistogamous flowers absent.
Ploidy and accepted chromosome counts.—2x; 2n = 14.
Age.—Crown node age c. 20.3 Ma (Figure 6).
Included species.—139.
Distribution.—From the equator (Ecuador) to southern Patagonia (Argentina) (Figure 10).
Etymology.—The well-established but illegitimate sectional name Andinium refers to the majority of species of the subgenus (90%) inhabiting the Andes mountains (Figure 10). Instead of combining the little used name Viola sect. Rosulatae to the subgenus level, we are deliberately describing a new subgenus, Neoandinium, with a name that clearly indicates a connection to Becker’s sect. Andinium.
Discussion.—The subgenus state of subg. Neoandinium is justified by its phylogenetic sister position to the rest of Viola and by its morphological distinctness, notably in the frequently imbricate rosettes and conspicuously and variably crested style. In spite of the high species diversity (21% of the total diversity of Viola) and wide distribution in the Andes, subg. Neoandinium is incompletely known. Dozens of species await description [68] and the subgenus lacks both a phylogeny and until recently a taxonomic treatment. The data presented here are a synopsis of the recent monograph by Watson et al. [68] who recognised 11 morphological sections within subg. Neoandinium. Hitherto all species studied have proven diploid (four species in two sections) but unpublished data on gene homoeolog numbers indicate allopolyploidy at least within sect. Sempervivum (T.M., unpublished).
Both Reiche [48] and Becker [1] subdivided subg. Neoandinium in annual and perennial species, but this classification does not appear to be natural [68]. However, this difference in life cycles is reflected in a difference in the growth form. Annual species have a taproot and only one rosette, while perennial species present a taproot usually branching below the ground, and various degrees of transition between rosettes, pleiocorm, and alpine cushion plants. Stolon-like persistent axes can also rarely be found among perennial species (sect. Rhizomandinium). A constant character within the subgenus is the margins of the leaf lamina. On the one hand, there is a group of species that present entire margins (sects. Confertae, Ericoidium, Rhizomandinium, and Sempervivum) and, on the other hand, another group of species with crenulate, crenate, lobed, even incised margins (sects. Grandiflos, Inconspicuiflos, Relictium, Rosulatae, Subandinium, Triflabellium, and Xylobasis). Generally, hairiness and the presence/absence of glands are correlated with this character; the entire leaves being generally glabrous without glands, and the leaves with non-entire margins often having hairs, glands, and raised veins. Because several characters are correlated, it can be hypothesised that these two morphological groups reflect phylogeny at least to some degree, but it is currently not known whether they are phylogenetic sisters or whether one is nested within the other.
The undescribed Viola quasichilenium J. M. Watson & A. R. Flores, ined., is superficially similar to sect. Chilenium of subg. Viola in having an extended petiole and in corolla colour and shape, but belongs in subg. Neoandinium on the basis of having abaxial lamina glands and a style with a significant crest, apparently apical. The specimen is known from photograph only, without geographical information.
Viola sect. Confertae Reiche in Nat. Pflanzenfam., ed. 1 [Engler & Prantl], 3(6): 335. 1895.—Lectotype (Watson et al. [68], page: 189): Viola nassauvioides Phil.
Diagnostic characters.—Perennial erect, caulescent, glabrous herb. Fertile stem enveloped in short, acaulous laminas, apex as expanded, imbricate rosette. Sterile rosettes basal, subacaulous, imbricate.
Included species.—1. Viola nassauvioides Phil.
Distribution.—Unknown (probably central Chile) [68].
Viola sect. Ericoidium J. M. Watson, A. R. Flores & Marcussen in Watson et al., Viola Subg. Andinium: 189. 2021.—Type: Viola fluehmannii Phil.
Diagnostic characters.—Perennial dwarf ericoid shrublets.
Included species.—1. Viola fluehmannii Phil.
Distribution.—Southern Chile, central-western Argentina.
Viola sect. Grandiflos J. M. Watson, A. R. Flores & Marcussen in Watson et al., Viola Subg. Andinium: 190. 2021.—Type: Viola truncata Meyen.
Diagnostic characters.—Perennial subacaulous, rosette-forming herbs. Rosette loose, irregular, not imbricated, radiating, not depressed. Lamina narrow, oblanceolate-spathulate, flexible, acute, entire, dentate or pinnatifid, never crenate. Corolla large, prominent, c. 15 × 15 mm, twice as wide as lamina or more.
Included species.—6. Viola acanthophylla Leyb. ex Reiche, V. angustifolia Phil., V. belovorum J. M. Watson & A. R. Flores, ined., V. bustillosia Gay, V. cheeseana J. M. Watson, V. truncata Meyen
Distribution.—Central Chile.
Viola sect. Inconspicuiflos J. M. Watson & A. R. Flores in Watson et al., Viola Subg. Andinium: 192. 2021.—Type: Viola lilliputana Iltis & H. E. Ballard
Diagnostic characters.—Dwarf, cushion forming plants, glabrous or with indumentum. Corolla notably small, the upper and lateral petals distinctly larger than the bottom one.
Included species.—8. Viola blefescudiana, ined., V. diminutiva, ined., V. enmae P. Gonzáles, V. lilliputana Iltis & H. E. Ballard, V. membranacea W. Becker, V. quasimelanium H. Beltrán & J. M. Watson, ined., V. quercifolia, ined., V. weibelii J. F. Macbr.
Distribution.—Peru.
Viola sect. Relictium J. M. Watson, A. R. Flores & Marcussen in Watson et al., Viola Subg. Andinium: 193. 2021.—Type: Viola huesoensis Martic.
Diagnostic characters.—Annual rosulate herbs. Cilia long, surrounding entire lamina margin, strongly deflexed.
Distribution.—Northern Chile.
Included species.—8. Violadandoisiorum J. M. Watson & A. R. Flores, V. deflexa, ined., V. godoyae Phil., V. huesoensis Martic., V. johnstonii W. Becker, V. marcelorosasii J. M. Watson & A. R. Flores, V. ovalleana Phil., V. simulans, ined.
Viola sect. Rhizomandinium J. M. Watson, A. R. Flores & Marcussen in Watson et al., Viola Subg. Andinium: 193. 2021 (“Rhizomandimium”).—Type: Viola escondidaensis W. Becker
Diagnostic characters.—Perennial herbs. Stem arising from the apex of long, creeping, stolon-like segment.
Distribution.—Northern Argentine Patagonia.
Included species.—2. Violaanitae J. M. Watson, V. escondidaensis W. Becker
Viola sect. Rosulatae Reiche in Nat. Pflanzenfam., ed. 1 [Engler & Prantl], 3(6): 335. 1895.—Type (Shenzhen Code Art. 22.2): Viola rosulata Poepp. & Endl.
≡ Viola sect. Andinium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 374. 1925, nom. illeg. superfl. (Shenzhen Code Art. 52.1).—Type (Shenzhen Code Art. 7.5): Viola rosulata Poepp. & Endl.
Diagnostic characters.—Perennial or annual subacaulous, more or less hairy rosette-forming herbs. Lamina flexible, elliptical, narrowly to broadly obovate, orbicular, or rhomboid, deeply to shallowly crenate, sinuous-crenate, dentate, incised, pinnatifid, or rarely entire when plant perennial.
Ploidy and accepted chromosome counts.—2x (V. congesta); 2n = 14 (V. montagnei, V. roigii).
Age.—Crown node unknown; stem node c. 13.9 Ma (Figure 6).
Distribution.—Central-northern Peru to northern Patagonia.
Included species.—55. Viola(Rosulatae) sp. 02, ined., V. (Rosulatae) sp. 04, ined., V. (Rosulatae) sp. 05, ined., V. (Rosulatae) sp. 06, ined., V. argentina W. Becker, V. aurantiaca Leyb., V. calchaquiensis W. Becker, V. chamaedrys Leyb., V. cistanthe, ined., V. congesta Gillies ex Hook. & Arn., V. decipiens Reiche, V. escarapela J. M. Watson & A. R. Flores, V. evae Hieron. ex W. Becker, V. exilis Phil., V. exsul J. M. Watson & A. R. Flores, V. farkasiana J. M. Watson & A. R. Flores, V. ferreyrae P. Gonzáles, V. friderici W. Becker, V. frigida Phil., V. gelida J. M. Watson, M. P. Cárdenas & A. R. Flores, V. glechomoides Leyb., V. granulosa Wedd., V. hillii W. Becker, V. hippocratica J. M. Watson & A. R. Flores, ined., V. imbricata J. M. Watson & A. R. Flores (et al.), ined., V. kermesina W. Becker, V. lanifera W. Becker, V. lilloana W. Becker, V. llullaillacoensis W. Becker, V. longibracteata P. Gonzáles & J. M. Watson, ined., V. montagnei Gay, V. multiflora, ined., V. nazarenoensis, ined., V. neuquenensis J. M. Watson & A. R. Flores, ined., V. niederleinii W. Becker, V. ornata D. Montesinos & J. M. Watson (et al.), ined., V. philippiana Greene, V. philippii Leyb., V. replicata W. Becker, V. rhombiloba H. E. Ballard, ined. [Monheim s. n.], V. rodriguezii W. Becker, V. roigii Rossow, V. rosulata Poepp. & Endl., V. rubromarginata J. M. Watson & A. R. Flores, V. rugosa Phil. ex W. Becker, V. singularis J. M. Watson & A. R. Flores, V. spegazzinii W. Becker, V. stellaris, ined., V. tectiflora W. Becker, V. tholiformis, ined., V. tovarii P. Gonzáles & Molina-Alor, V. trochlearis J. M. Watson & A. R. Flores, V. umbrina, ined., V. volcanica Gillies ex Hook. & Arn., V. xanthopotamica J. M. Watson & A. R. Flores
Viola sect. Sempervivum J. M. Watson & A. R. Flores in Watson et al., Viola Subgenus Andinium: 188. 2021.—Type: Viola atropurpurea Leyb.
Diagnostic characters.—Perennial or annual subacaulous, glabrous, imbricated rosette-forming herbs. Lamina entire or shallowly subcrenulate, apex acute to obtuse.
Ploidy and accepted chromosome counts.—Unknown; gene homoeolog numbers indicate allopolyploidy in some species (T.M., unpubl.).
Age.—Crown node c. 13.3 Ma; stem node c. 20.3 Ma (Figure 6).
Distribution.—Ecuador to southern Patagonia.
Included species.—34. Violaabbreviata J. M. Watson & A. R. Flores, V. aizoon Reiche, V. atropurpurea Leyb., V. auricolor Skottsb., V. bangii Rusby, V. beckeriana J. M. Watson & A. R. Flores, V. columnaris Skottsb., V. comberi W. Becker, V. coronifera W. Becker, V. cotyledon Ging., V. cupuliformis H. E. Ballard, ined. [T. Hofreiter & T. Franke 1/104], V. dasyphylla W. Becker, V. hieronymi W. Becker, V. leyboldiana Phil., V. lologensis (W. Becker) J. M. Watson, V. marcelae, ined., V. micranthella Wedd., V. nigriflora H. E. Ballard, ined. [T. Hofreiter & T. Franke 1/103], V. nobilis W. Becker, V. obituaria J. M. Watson & A. R. Flores, V. pachysoma M. Sheader & J. M. Watson, V. petraea W. Becker, V. polycephala H. E. Ballard & P. Jørg., V. portulacea Leyb., V. pusillima Wedd., V. pygmaea Juss. ex Poir., V. regina J. M. Watson & A. R. Flores, V. rossowiana J. M. Watson & A. R. Flores, V. sacculus Skottsb., V. santiagonensis W. Becker, V. sempervivum Gay, V. skottsbergiana W. Becker, V. turritella J. M. Watson & A. R. Flores, V. vortex, ined.
Viola sect. Subandinium J. M. Watson & A. R. Flores in Watson et al., Viola Subg. Andinium: 193. 2021.—Type: Viola subandina J. M. Watson
Diagnostic characters.—Annual rosulate herbs. Lamina flexible, linear, oblanceolate or obovate, entire or shallowly long-crenulate. Diploid.
Ploidy and accepted chromosome counts.—2x (Viola pusilla); no chromosome counts.
Age.—Crown node c. 4.8 Ma; stem node c. 13.9 Ma (Figure 6).
Distribution.—Southern Chile to southern Peru.
Included species.—15. Violaaraucaniae W. Becker, V. aurata Phil., V. auricula Leyb., V. domeikoana Gay, V. minutiflora Phil., V. nubigena Leyb., V. polypoda Turcz., V. pulvinata Reiche, V. pusilla Poepp., V. rhombifolia Leyb., V. subandina J. M. Watson, V. taltalensis W. Becker, V. vallenarensis W. Becker, V. weberbaueri W. Becker, V. yrameae J. M. Watson & A. R. Flores, ined.
Viola sect. Triflabellium J. M. Watson, A. R. Flores & Marcussen in Watson et al., Viola Subg. Andinium: 192. 2021.—Type: Viola triflabellata W. Becker
Diagnostic characters.—Perennial rosette-forming herbs. Style crest as one apical and two lateral extended lobes.
Distribution.—Northern Chile to northwestern Argentina.
Included species.—7. Viola(Triflabellium) sp. 1, ined., V. flos-idae Hieron., V. joergensenii W. Becker, V. mesadensis W. Becker, V. triflabellata W. Becker, V. tucumanensis W. Becker, V. uniquissima J. M. Watson & A. R. Flores
Viola sect. Xylobasis J. M. Watson & A. R. Flores in Watson et al., Viola Subg. Andinium: 191. 2021.—Type: Viola beati J. M. Watson & A. R. Flores
Diagnostic characters.—Perennial hairy, rosette-forming herbs. Stem shortly woody-branched.
Distribution.—Northwestern Argentina.
Included species.—1. Viola beati J. M. Watson & A. R. Flores
=Viola sect. Sparsifoliae Reiche in Nat. Pflanzenfam., ed. 1 [Engler & Prantl], 3(6): 334. 1895, nom. inval. (Shenzhen Code Art. 22.2; Viola odorata L.)
Description.—Annual or perennial herbs, subshrubs or very occasionally treelets. Axes morphologically differentiated or not. Leaves scattered on stems or in rosettes, very occasionally imbricated with distichous phyllotaxy (sect. Tridens). Stipules free or partially adnate, sometimes large and foliaceous. Lamina usually petiolate; young laminas with involute margins (rarely folded in narrow leaves). Peduncles often longer than mature leaves. Bottom petal usually entire or shallowly emarginate, very rarely cleft. Spur absent to very long (34 mm). Nectariferous appendage of the two bottom stamens of various shape, rarely filiform. Style filiform, clavate, or capitate, not crested (but lateral lobes present in sect. Sclerosium) but top of style apex often flattened or with more or less raised edges, bearded or beardless. Cleistogamous flowers often produced.
Diagnostic characters.—Young laminas with involute margins OR peduncles longer than mature leaves OR style not crested OR cleistogamous flowers present.
Ploidy and accepted chromosome counts.—2x, 4x, 6x, 8x, 10x, 12x, 14x, 16x, 18x, 20x, >20x. 2n = 4, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 34, 36, 40, 44, c. 44, 46, 48, 50, 52, 54, 58, 60, c. 64, 72, 76, 80, c. 80, 82, c. 96, 102, c. 120, 128.
Age.—Crown node age 29.0 (28.3–29.4) Ma; stem node age 30.9 (29.8–31.3) Ma [28].
Included species.—525.
Distribution.—All continents except Antarctica. Diversity centres in e Asia, w Eurasia and N America.
Discussion.—Within subg. Viola we recognise 20 sections which can be grouped in three well-separated biogeographic clusters and allopolyploid tangles. The first cluster occurs in South and Central America and Australasia and comprises 43 species in 7 sections (sects. Chilenium, Erpetion, Leptidium, Nematocaulon, Rubellium, Tridens, and Xanthidium). The second cluster occurs primarily in the northern hemisphere and comprises 470 species in 12 sections (sects. Abyssinium, Chamaemelanium, Danxiaviola, Delphiniopsis, Himalayum, Melanium, Nosphinium, Plagiostigma, Sclerosium, Spathulidium, Viola, and Xylinosium). The third cluster occurs in South Africa with a single allopolyploid section and species (sect. Melvio; V. decumbens). The last two clusters are phylogenetically nested within the first one. Sections Chamaemelanium and Rubellium (2n = 12) are the only diploid lineages within subg. Viola (no data for sect. Xanthidium).
Viola sect. Abyssinium Marcussen, sect. nov.—Type: Viola abyssinica Steud. ex Oliv.
Description.—Perennial herbs. Axes not morphologically differentiated. All stems ascending or trailing, rooting at proximal nodes. Stipules deeply dentate-laciniate to entire. Lamina crenulate, petiolate. Flowers c. 1 cm, peduncles produced only from some leaf axils. calycine appendages very short or absent. Corolla violet or white, with a white throat, bottom petal with violet striations. Spur saccate. Style clavate, laterally compressed, at base geniculate, at apex galeiform and distally margined, beardless. Cleistogamous flowers not produced. Allododecaploid (CHAM + MELVIO). Secondary base chromosome number x’ = c. 36. ITS sequence of MELVIO type.
Diagnostic characters.—All stems ascending or trailing AND corolla violet or white with white throat AND style clavate.
Ploidy and accepted chromosome counts.—12x; 2n = c. 72 (Viola abyssinica).
Age.—Crown node age c. 2 Ma; stem node age 3.6 (1.8–5.0) Ma [28].
Included species.—3. Violaabyssinica Steud. ex Oliv., V. eminii (Engl.) R. E. Fr., V. nannae R. E. Fr.
Distribution.—High mountains of central and eastern Africa and Madagascar (Figure 11): Viola abyssinica throughout the range; V. eminii in eastern Congo, Rwanda, Burundi, Uganda to central and southern Kenya, northern Tanzania south to the Uluguru Mountains; V. nannae in central and southern Kenya [238].
Etymology.—The name Abyssinium refers to the main distribution area in and around Ethiopia (=Abyssinia).
Discussion.—Sect. Abyssinium is one of just two endemic African lineages of Viola (the other is the South African sect. Melvio). The count of 2n = c. 72 in V. abyssinica [239] is the only count for the section and needs confirmation. Section Abyssinium has an African distribution but is phylogenetically nested within the north hemisphere tangle of allopolyploid lineages. It appears to have originated in the Pliocene, from an allopolyploid of sect. Spathulidium (8x) and one of the 4x ancestors of that lineage (Figure 2 and [28]), which is distributed in southwestern Asia. The relatively recent origin of sect. Abyssinium from Eurasian ancestors fits a pattern commonly observed in Afrotemperate/Afromontane floral elements [240]. Becker [1] made a note that this group of species would merit a separate section, but he did not provide one. Possible hybridisation among the three species of sect. Abyssinium is briefly discussed by Grey-Wilson [238].
Viola sect. Chamaemelanium Ging. in Mém. Soc. Phys. Genève 2(1): 28. 1823 ≡ Viola subg. Chamaemelanium (Ging.) Juz. in Schischk. & Bobrov, Flora URSS 15: 446. 1949—Type: Viola canadensis L.
≡Lophion Spach, Hist. Nat. Vég. [Spach] 5: 516. 1836 ≡ Lophion subg. Eulophion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 215. 1914, nom. inval. (Shenzhen Code Art. 22.2)—Type: Viola canadensis L.
=Viola sect. Dischidium Ging. in Mém. Soc. Phys. Genève 2(1): 28. 1823 ≡ Dischidium (Ging.) Opiz in Bercht. & Opiz, Oekon.-Techn. Fl. Böhm. [Berchtold & al.] 2(2): 7. 1839 ≡ Viola subg. Dischidium (Ging.) Peterm., Deutschl. Fl.: 65. 1846; (Ging.) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 172. 1909 (isonym); (Ging.) Juz. in Schisk. & Bobrov, Flora URSS 15: 441. 1949 (isonym) ≡ Viola [unranked] (”Gruppe”) Dischidium W. Becker in Beih. Bot. Centralbl., Abt. 2, 36: 38. 1918—Type: Viola biflora L.
≡Chrysion Spach, Hist. Nat. Vég. [Spach] 5: 509. 1836.—Type: Viola biflora L.
≡Viola [unranked] §.5. Dischidieae Boiss., Fl. Orient. 1: 452. 1867 ≡ Viola subsect. Dischidieae (Boiss.) Rouy & Foucaud, Fl. France [Rouy & Foucaud] 3: 36. 1896—Type: Viola biflora L.
=Crocion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 215. 1914—Type: V. pubescens Aiton
=Viola (sect. Nomimium) [unranked] (“Gruppa”) Memorabiles W. Becker in B. Fedtsch., Fl. Aziat. Ross. 8: 19. 1915 ≡ Viola sect. Memorabiles (W. Becker) Juz. in Schischk. & Bobrov, Flora URSS 15: 407. 1949—Type: Viola kusnezowiana W. Becker
=Viola “class” Orbiculares Pollard in Bot. Gaz. 26: 330. 1898, nom. inval. (Shenzhen Code Art. 33.9) ≡ Viola [unranked] Orbiculares W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 369. 1925 ≡ Viola subsect. Orbiculares (“Pollard“) Brizicky in J. Arnold Arb. 42: 326. 1961, nom inval. (Shenzhen Code Art. 41.5)—Type: Viola orbiculata Geyer ex Holz.
=Viola [unranked] D. Erectae W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 370. 1925 ≡ Viola sect. Erectae (W. Becker) Ching J.Wang, Fl. Reipubl. Popularis Sin. 51: 123. 1991.—Lectotype (designated here): Viola acutifolia (Kar. & Kir.) W. Becker
=Viola [unranked] (Untergruppe) Longicalcaratae W. Becker in Beih. Bot. Centralbl. 36(2): 38. 1918 ≡ Viola [sect. Dischidium; unranked] A. Longicalcaratae W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 370. 1925 ≡ Viola subsect. Longicalcaratae (W. Becker) W. Becker in Acta Horti Gothob. 2: 288. 1926 ≡ Viola subsect. Longicalcaratae (W. Becker) Ching J. Wang, Fl. Reipubl. Popularis Sin. 51: 119. 1991.—Lectotype (designated here): Viola wallichiana Ging.
Description.—Perennial herbs. Axes usually morphologically differentiated into a perennial rhizome and annual aerial stems. Rhizome usually deep-buried with a few-leaved apical rosette. Lateral stems aerial, rarely stolons, sometimes reduced or absent. Stipules partially to completely herbaceous or rarely membranous, margins entire or irregularly dentate with a few teeth. Lamina cordate to lanceolate, margin crenate, lobed, or pedately divided, usually long-petiolate. Corolla yellow, white, or violet, always with a yellow throat. Spur very short, rarely longer in a few Asian species. Style clavate or capitate, variable, usually bearded at apex. Cleistogamous flowers usually produced; cleistogamy seasonal. Diploid. Base chromosome number x = 6. ITS sequence of CHAM type.
Diagnostic characters.—Corolla with a yellow throat AND base chromosome number x = 6.
Ploidy and accepted chromosome counts.—2x, 4x, 6x, 8x, 12x; 2n = 12, 24, 36, 48, 72.
Age.—Crown node age 19.0 (18.0–19.3) Ma [28].
Included species.—69. Viola acutifolia (Kar. & Kir.) W. Becker, V. alliariifolia Nakai, V. allochroa Botsch., V. angkae Craib, V. aurea Kell., V. bakeri Greene, V. barroetana W. Schaffn. ex Hemsl., V. beckwithii Torr. & A. Gray, V. biflora L., V. brevistipulata (Franch. & Sav.) W. Becker, V. californica M. S. Baker, V. cameleo H. Boissieu, V. canadensis L., V. caucasica (Rupr.) Kolen. ex Juz., V. charlestonensis M. S. Baker & J. C. Clausen, V. coahuilensis H. E. Ballard, ined. [P. Fryxell 2692], V. confertifolia C. C. Chang, V. crassa (Makino) Makino, V. cuneata S. Watson, V. delavayi Franch., V. dimorphophylla Y. S. Chen & Q. E. Yang, V. douglasii Steud., V. eriocarpa Schwein., V. fischeri W. Becker, V. flagelliformis Hemsl., V. flettii Piper, V. franksmithii N. H. Holmgren, V. galeanaensis M. S. Baker, V. glabella Nutt., V. glaberrima (Ging. ex Chapm.) House, V. guadalupensis A. M. Powell & Wauer, V. hallii A. Gray, V. hastata Michx., V. hediniana W. Becker, V. kitamiana Nakai, V. kusnezowiana W. Becker, V. lithion N. H. Holmgren & P. K. Holmgren, V. lobata Benth., V. majchurensis Pissjauk., V. muehldorfii Kiss, V. muliensis Y. S. Chen & Q. E. Yang, V. nuttallii Pursh, V. ocellata Torr. & A. Gray, V. orbiculata Geyer ex Holz., V. orientalis (Maxim.) W. Becker, V. painteri Rose & House, V. pedunculata Torr. & A. Gray, V. pinetorum Greene, V. praemorsa Douglas, V. pubescens Aiton, V. purpurea Kellogg, V. quercetorum M. S. Baker & J. C. Clausen, V. rockiana W. Becker, V. rotundifolia Michx., V. rugulosa Greene, V. scopulorum (A. Gray) Greene, V. sempervirens Greene, V. sheltonii Torr., V. szetschwanensis W. Becker & H. Boissieu, V. tenuipes Pollard, V. tenuissima C. C. Chang, V. tomentosa M. S. Baker & J. C. Clausen, V. trinervata (Howell) Howell ex A. Gray, V. tripartita Elliott, V. uniflora L., V. urophylla Franch., V. utahensis M. S. Baker & J. C. Clausen, V. vallicola A. Nelson, V. wallichiana Ging.
Discussion.—Sect. Chamaemelanium is the only diploid representative of the CHAM genome; intrasectional allopolyploids are frequent but there was no hybridisation with the MELVIO lineage. The lineage is characterised karyologically by the base chromosome number x = 6 and morphologically by a plesiomorphic yellow corolla (variously coloured but always with a yellow throat in the Canadenses and Chrysanthae greges), shoots differentiated in a perennial (often deep-buried) rhizome with an apical (often few-leafed) leaf rosette and annual lateral floriferous stems, and the presence of seasonal cleistogamy. The lateral stems are usually more or less erect and aerial, in some reclining or prostrate and leafy or leafless (V. kusnezowiana in northeastern Asia, V. orbiculata, V. rotundifolia and V. sempervirens in North America), or entirely missing (V. barroetana in Mexico). Stipules in some species are semi-membranous or membranous, and are commonly entire or with one to few irregular teeth on one or both margins. Leaf lamina is usually crenate or crenulate but deeply divided in some taxa (greges Chrysanthae and Nudicaules in North America, the V. biflora group in northeastern Asia). Style shape is variable [29] but most species groups have a capitate, bearded style. Members of the V. biflora group (the former sect. Dischidium) have a bilobate style, while a few other species have style shapes resembling those found in other sections, such as sect. Viola (V. kitamiana and V. kusnezowiana in northeastern Asia) or sect. Plagiostigma (V. rotundifolia in eastern North America). Elaiosomes are highly reduced to obsolete in at least some species of the Canadenses grex. Cleistogamous flowers are missing in some taxa adapted to arid habitats (notably grex Chrysanthae and V. guadalupensis).
We recognise a broadly defined sect. Chamaemelanium that includes sect. Dischidium Ging. (i.e., the V. biflora group), grex Orbiculares Pollard (i.e., V. orbiculata, V. sempervirens and V. rotundifolia) and grex Memorabiles W. Becker (i.e., V. kusnezowiana) previously placed in sect. Nomimium by Becker [1], and V. kitamiana. The inclusion of Dischidium and Orbiculares in sect. Chamaemelanium, first suggested nearly a century ago by Clausen [29,59], is supported by morphology, chromosome counts, and by phylogeny (Figure 13) [28,60]. Viola kusnezowiana is included in sect. Chamaemelanium on basis of flower and stipule characters [61]; the somewhat emarginate lamina apex is particularly reminiscent of the V. biflora group. Viola kitamiana is included in sect. Chamaemelanium based on its corolla with a yellow throat (otherwise white) and the diploid chromosome number 2n = 12 [61].
We do not recognise infrasectional groups within sect. Chamaemelanium because its extant sublineages, at least the North American ones (Figure 13), are interconnected by allopolyploidy and therefore non-monophyletic [59,60,241,242]. Furthermore, the 7–8 diploid deep lineages do not correspond to any recognised morphological greges [1] and their interrelationships are deep and largely unresolved. For instance, both the capitate-bearded style shape and the rhizomatous habit with lateral, aerial floriferous stems, the two characters that define Becker’s grex Erectae, appear to be ancestral and plesiomorphic within sect. Chamaemelanium (Figure 13).
The initial radiation of sect. Chamaemelanium appears to have coincided with that of the CHAM + MELVIO allopolyploids in the northern hemisphere c. 19 Ma ago [28]. It has not been established whether the CHAM genomes involved in these allopolyploidisations were derived from within the extant sect. Chamaemelanium or from a lineage sister to it. It is, however, clear that the version of the CHAM genome present in the southern hemisphere sect. Chilenium and sect. Erpetion is sister to all other CHAM genomes.
The report of 2n = 20 in V. kusnezowiana [243] is at odds with the other counts in the section, all of which are based on x = 6, and in need of confirmation.
Viola sect. Chilenium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 376. 1925.—Lectotype (designated here): Viola maculata Cav.—Note: Viola maculata is indicated as “Haupttypus” in the protologue (Becker 1925: 376).
≡Viola [unranked] § I. Bicaules Reiche in Fl. Chile [Reiche] 1: 139. 1896—Lectotype (designated here): Viola maculata Cav.
Description.—Perennial herbs. Axes not morphologically differentiated. Stem creeping, more or less densely noded, deeply seated. Stipules free, broad and glandular-laciniate, rarely entire. Lamina oblong, elliptical or rhombic-lanceolate to reniform, margin crenate, long-petiolate. Bracteoles broad, deeply glandular-laciniate. Corolla yellow, rarely white (V. commersonii). Bottom petal at least twice as broad as top petals, rarely only slightly broader than the top petals (V. rudolphii), with brown striation, rarely reddish-violet striation (V. commersonii). Spur shorter than tall, rarely much longer than tall (V. rudolphii and V. stuebelii). Style clavate or straight, concave, flattened or slightly acute apically with a continuous sharp dorsolateral rim, the rim truncate and hiding the stigma (V. stuebelii) or slightly prolonged on the upper side with an upcurved visible rostellum bearing the stigma (other species), usually beardless (white-hairy in V. rudolphii). Cleistogamous flowers produced; cleistogamy facultative. Allopolyploid.
Diagnostic characters.—All stems rhizomes AND corolla yellow with brown striation or white with reddish-violet striation AND facultative cleistogamy.
Ploidy and accepted chromosome counts.—≥4x; no chromosome counts.
Age.—Crown node age unknown, stem node age 7.4 (6.5–7.7) Ma [28].
Included species.—7. Violacommersonii DC. ex Ging., V. germainii Sparre, V. maculata Cav., V. magellanica G. Forst., V. reichei Skottsb. ex Macloskie, V. rudolphii Sparre, V. stuebelii Hieron.
Distribution.—Disjunct in southern (Argentina and Chile) and northern South America (Colombia, Ecuador, and Peru) (Figure 14).
Discussion.—We here modify Becker’s [1] original delimitation of sect. Chilenium by including V. stuebelii (=V. glandularis H. E. Ballard & P. Jørg.) based on shared diagnostic characters, and excluding V. huidobrii (=V. brachypetala Gay). Reiche [122,130] was the first to recognise this group, which he circumscribed under an invalid taxonomic rank (i.e., the unranked Bicaules within the invalid “Divisio” Sparsifoliae). Later, Sparre [62,63] revised the section and recognised eight southern South American species (some of them were later synonymised), which he distributed among three subsections, Maculatae (V. germainii, V. maculata, V. reichei), Magellanicae (V. commersonii, V. magellanica), and Lanatae (V. rudolphii), based on characteristics of the spur, style, and nectariferous appendages. We transfer the distinctive, violet-flowered V. huidobrii (Sparre as subsect. Coeruleae) to sect. Viola subsect. Rostratae. The new delimitation of sect. Chilenium renders the section geographically disjunct, with V. stuebelii in northern South America and the rest of the species in southern South America. Section Chilenium comprises only seven species, some closely related (e.g., V. maculata and V. reichei) and others known only from the type specimen (V. germainii and V. rudolphii), and in the absence of molecular data we choose not to keep Sparre’s subsections.
Viola sect. Danxiaviola W. B. Liao & Q. Fan in Phytotaxa 197: 19. 2015—Type: Viola hybanthoides W. B. Liao & Q. Fan
Description.—Subshrub. Axes not morphologically differentiated. All stems erect or ascending. Stipules free, conspicuous, oblong-lanceolate, remotely long-fimbriate. Lamina elliptic or ovate-lanceolate, margin serrate, short-petiolate. Corolla whitish to pale violet. Bottom petal clawed, much larger than the other, reduced petals, whitish to pale violet with a yellowish green blotch at base. Spur short and saccate. Style capitate, at apex slightly bilobate, beardless, not beaked and with a stigmatic opening in front and with a lamellar processus below the opening. Cleistogamous flowers not produced. Chromosome number x = 10. ITS sequence of CHAM type.
Diagnostic characters.—Bottom petal clawed, much larger than the other petals.
Ploidy and accepted chromosome counts.—Probably 4x; 2n = 20.
Age.—Crown node age not applicable (monotypic section), stem node age probably 17.8–19.3 Ma.
Included species.—1. Violahybanthoides W. B. Liao & Q. Fan
Distribution.—Southeastern China (northern Guangdong). Known only from two sites on Mt. Danxia (Figure 15).
Discussion.—The single species in the section, V. hybanthoides, is phylogenetically isolated within the north hemisphere allopolyploid tangle, based on both ITS and chloroplast sequences [90]. The combined features of the larger bottom petal and much smaller lateral and upper petals is unique in Viola but found recurrently in most bilaterally symmetrical genera of the Violaceae, such as genera currently being segregated from the former polyphyletic Hybanthus, and sisters to Viola, Noisettia and Schweiggeria [3]. We infer that V. hybanthoides is probably a CHAM + MELVIO meso-allotetraploid, judging from its chromosome number (2n = 20) which in sect. Viola and sect. Delphiniopsis reflects 4x, the small size of its chromosomes and tricolporate pollen which both reflect a certain time since the polyploidisation, and its phylogenetic placement nested within a tetraploid clade [90] (Figure 6).
This morphologically highly unusual species was discovered as late as 2012 and published in 2015 [90] and was therefore included neither in the morphological treatment of Becker [1] nor in the phylogeny of Marcussen et al. [28].
Viola sect. Delphiniopsis W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 373. 1925—Type (Shenzhen Code Art. 10.8): Viola delphinantha Boiss.
≡Viola [unranked] §.1. Delphinoideae Boiss., Fl. Orient. 1: 451. 1867.—Type: Viola delphinantha Boiss.
≡Viola [unranked] c. Lobulariae Nyman, Consp. Fl. Eur. 1: 79. 1878.—Type: Viola delphinantha Boiss.
Description.—Perennial herbs with a woody base. Axes not morphologically differentiated. All stems aerial, annual, growing in fascicles from a woody and sometimes thick rhizome (pleiocorm). Leaves sessile, consisting of 3–5 lanceolate, entire segments, lamina and stipule segments similar. Corolla pink to magenta. Spur 12–30 mm, down-curved. Style clavate, glabrous, unmargined, with a simple, wide stigmatic opening. Cleistogamous flowers not produced. Allotetraploid (CHAM + MELVIO). Secondary base chromosome number x’ = 10. ITS sequence of MELVIO type.
Diagnostic characters.—Corolla pink to magenta AND spur 12–30 mm, down-curved.
Ploidy and accepted chromosome counts.—4x; 2n = 20.
Age.—Crown node age unknown, stem node age 14.7 (6.3–18.6) Ma [28].
Included species.—3. Violacazorlensis Gand., V. delphinantha Boiss., V. kosaninii (Degen) Hayek.
Distribution.—Disjunct in southern Europe: southern Spain (V. cazorlensis) and the Balkans (V. delphinantha, V. kosaninii) (Figure 16).
Discussion.—Section Delphiniopsis is highly distinct, phylogenetically, karyologically (x’ = 10), and morphologically. The species are specialised to be pollinated by a single species of day-flying hawk-moth (Macroglossum stellatarum, Sphingidae, Lepidoptera) [175,176]. The disjunct distribution of sect. Delphiniopsis, between V. delphinantha and V. kosaninii in the Balkans and V. cazorlensis in southern Spain, has been suggested to result from vicariance and to date from the Early Pliocene, 3.6–5.3 Ma [244]. The crown age of the section have so far not been phylogenetically dated, but the idea that the species are young is further supported by their morphological similarity and reports of their being able to hybridise in culture (plants of V. cazorlensis × V. delphinantha were displayed at the Midland AGS SHOW 2012). Phylogenetic analysis confirms that sect. Delphiniopsis is an isolated and highly specialised lineage on a rather long branch, indicating rapid evolution [28]. The characters once considered ”primitive” [244,245], such as woodiness, entire leaves and stipules, small and uniform chromosomes and rather common chromosome number, relict distribution, and lack of cleistogamy, should rather be interpreted as secondary specialisations, just like the highly specialised pollination. The low and apparently young diversity of the section may be explained by its high level of specialisation in both pollination syndrome and choice of habitat: the species inhabit limestone crevices, a rare habitat that minimises competition but at the same time limits the population size and dispersal, thereby increasing the risk of extinction.
Viola sect. Erpetion (DC. ex Sweet) W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 376. 1925 ≡ Erpetion DC. ex Sweet, Brit. Fl. Gard. 2: nr. 170. 1826 ≡ Viola subg. Erpetion (DC. ex Sweet) Y. S. Chen in Raven & Hong, Fl. China 13: 111. 2007—Type: Viola hederacea Labill.
Description.—Perennial herbs. Axes seemingly morphologically differentiated in a perennial stem with lateral sympodial stolons. Perennating stem densely or occasionally remotely noded. Sympodial stolons with a pair of bracts between each cluster of leaves. Stipules small, lanceolate. Lamina ovate-rhomboid to broadly reniform, margin crenate, long-petiolate. Corolla white to dark violet, often with a darker throat; corolla sometimes highly reduced. Spur reduced to a gibba and a green blotch on the inside of the bottom petal. Lateral petals with a broad dense pad of papillae. Style filiform, beardless. Cleistogamous flowers not produced. Allo-octoploid. Secondary base chromosome number x’ = 25.
Diagnostic characters.—Sympodial stolons present. Spur reduced to a gibba. Lateral petals with a broad dense pad of papillae.
Ploidy and accepted chromosome counts.—8x, 16x, 24x; 2n = 50 (V. banksii).
Age.—Crown node age 3.7 (3.2–3.9) Ma, stem node age 7.4 (6.5–7.7) Ma [28].
Included species.—11. Violabanksii K. R. Thiele & Prober, V. cleistogamoides (L. G. Adams) Seppelt, V. curtisiae (L. G. Adams) K. R. Thiele, V. eminens K. R. Thiele & Prober, V. fuscoviolacea (L. G. Adams) T. A. James, V. hederacea Labill., V. improcera L. G. Adams, V. perreniformis (L. G. Adams) R. J. Little & Leiper, V. serpentinicola de Salas, V. sieberiana Spreng., V. silicestris K. R. Thiele & Prober
Distribution.—Southern and eastern Australia; Tasmania (Figure 17).
Discussion.—Phylogenetically, sect. Erpetion is an allo-octoploid lineage with two CHAM genomes and another two genomes in common with sect. Chilenium, indicating that sect. Erpetion experienced a second genome duplication after the two sections diverged. There is no indication that this ancestral tetraploid Erpetion still exists. Section Erpetion is characterised karyologically by the secondary base chromosome number x’ = 25 [98]. The estimate of 10x for sect. Erpetion by Marcussen et al. [28] was based on unconfirmed (and probably erroneous) counts of 2n = 60 and 2n = 120 on “representatives of the Viola hederacea complex in the Kosciusko area” by Moore in [246].
Members of sect. Erpetion can be recognised immediately by two unique synapomorphies, i.e., the presence of sympodial stolons, which differ from true stolons by their clustered leaves and bibracteolate stem segments, and the pad of papillae on the lateral petals in place of the beard of trichomes some members of other lineages exhibit. Anatomically, the sympodial stolon consists of a potentially infinite chain of bibracteolate stem segments each ending in a leaf rosette, which in turn produces a new segment from the axil of its lowermost leaf. Adventitious roots are produced at the base of each rosette only. In Fragaria (Rosaceae), both sympodial and monopodial stolons can be found among closely related species (e.g., F. viridis vs. F. vesca, respectively), suggesting that the underlying genetics can be quite simple.
We follow the original delimitation of Becker [1] for the section. At the time only one variable species was recognised, Viola hederacea, but c. 11 species are now recognised [160,226]. Genome size data (2C DNA) indicate that sect. Erpetion forms a polyploid series based on 8x, i.e., with V. banksii at the 8x level (two accessions with 1.26 and 1.27 pg), V. fuscoviolacea at the 16x level (2.57 pg), and V. hederacea at the 24x level (3.45 pg; T.M., unpublished data, and [98]). Indeed, the occurrence of autogamous taxa with very small corollas, i.e., V. cleistogamoides and V. fuscoviolacea, agree with the observation of high ploidy in this section. A cultivar attributed to V. banksii is frequently grown as an ornamental, but appears to be a hybrid, based on having low pollen fertility [98].
The sister lineage of sect. Erpetion is the South American sect. Chilenium [28], from which it may have diverged c. 7.4 Ma ago [28]. This relationship is surprisingly from a morphological perspective, as the two taxa are rather dissimilar and lack obvious synapomorphies.
Viola sect. Himalayum Marcussen, sect. nov.—Type: Viola kunawurensis Royle
Description.—Dwarf perennial herb. Axes not morphologically differentiated. Stems subterranean from deeply buried pleiocorm, appearing aboveground as proximal or tufted rosettes. Stipules adnate to ¾ of their length. Lamina subentire with 0–2 shallow crenulae, spathulate, gradually tapering in a long petiole. Corolla c. 10 mm, light violet with dark striations. Lateral petals beardless. Spur as long as tall, saccate, 1–1.5 mm, obtuse. Style clavate, unmargined. Cleistogamous flowers produced; cleistogamy seasonal. Allo-octoploid (CHAM + MELVIO). Secondary base chromosome number x’ = 10 (needs confirmation). ITS sequence of MELVIO type.
Diagnostic characters.—Stipules adnate AND lamina spathulate and subentire AND spur as long as tall, 1–1.5 mm AND cleistogamous flowers produced.
Ploidy and accepted chromosome counts.—8x; 2n = 20?
Age.—Crown node age not applicable (monotypic section), stem node age probably 17.8–19.3 Ma.
Included species.—1. Viola kunawurensis Royle
Distribution.—High mountains surrounding the Tibetan Plateau: Tian Shan, Pamir, the Himalayas, Hengduan Shan, and Qilian Shan (Figure 18).
Etymology.—The name Himalayum refers to the distribution in the Himalayas and adjacent mountain ranges.
Discussion.—Section Himalayum comprises a single species, V. kunawurensis (=V. “kunawarensis”, V. thianschanica Maxim.), occurring at high elevations (3000–5000 m) in the Central Asian high mountains surrounding the Tibetan plateau (Figure 18). Viola kunawurensis differs from similar species of sect. Plagiostigma subsect. Patellares in having a very short spur and frequently elongated internodes arising from the deep-buried pleiocorm, as well as in chromosome number, and from sect. Spathulidium in style shape and in producing cleistogamous flowers.
Mining GPI sequences from the sequence reads archive of the reference sequence genome of V. kunawurensis (as V. “kunawarensis”; NCBI accession PRJNA805692) strongly indicates the presence of four homoeologs, confirming that sect. Himalayum is an independent CHAM + MELVIO allotetraploid lineage and further suggesting that the extant species is octoploid as a result of a secondary autopolyploidisation (Figure 6). The single chromosome count of 2n = 20 [247] is doubtful as this number reflects 4x in sect. Viola and sect. Delphiniopsis and therefore seems at odds with the octoploid condition of V. kunawurensis.
Becker originally placed Viola kunawurensis in grex Gmelinianae [248], later in grex Adnatae [1]; see note under sect. Plagiostigma subsect. Patellares. Sun and coworkers placed V. kunawurensis in sect. Viola subsect. Rostratae based on the (allegedly) shared chromosome number 2n = 20 [247] and numerical taxonomy of 58 traits [77].
Viola sect. Leptidium Ging. in Mém. Soc. Phys. Genève 2(1): 28. 1823 ≡ Viola subg. Leptidium (Ging.) Peterm., Deutschl. Fl.: 66. 1846—Type: Viola stipularis Sw.
Description.—Subshrubs or perennial herbs. Axes not morphologically differentiated. Stems reclining to erect, sometimes branched (in Viola scandens and V. stipularis at least 1 m long). Stipules lanceolate to ovate, laciniate, partially sheathing the stem. Lamina linear-lanceolate to reniform, margin crenate, short- to long-petiolate. Corolla whitish to violet with a white throat (corolla entirely red in V. arguta). Spur short and saccate (spur thick and bulbous in V. arguta). Bottom pair of stamens with apical “u”-shaped connective appendage. Style filiform, straight undifferentiated, with a simple stigmatic opening. Cleistogamous flowers produced, sometimes subterranean in V. arguta and possibly other species; cleistogamy facultative. Allotetraploid. Inferred secondary base chromosome number [x’ = 13.5].
Diagnostic characters.—Aerial stems AND laciniate sheathing stipules AND short saccate or thick bulbous spur AND “u”-shaped connective appendage on bottom pair of stamens AND filiform style.
Ploidy and accepted chromosome counts.—4x, 8x; 2n = 54 (V. dombeyana).
Age.—Crown node age 8.7 (3–16) Ma [28].
Included species.—18. Violaarguta Humb. & Bonpl. ex Schult., V. atroseminalis H. E. Ballard, ined., V. boliviana Britton, V. bridgesii Britton, V. cerasifolia A. St.-Hil., V. dombeyana DC. ex Ging., V. fuscifolia W. Becker, V. gracillima A. St.-Hil., V. lehmannii W. Becker ex H. E. Ballard & P. Jørg., V. mandonii W. Becker, V. saccata Melch., V. scandens Humb. & Bonpl. ex Schult., V. steinbachii W. Becker, V. stipularis Sw., V. subdimidiata A. St.-Hil., V. thymifolia Britton, V. uleana W. Becker, V. veronicifolia Planch. & Linden
Distribution.—Southeastern Mexico to Bolivia; northwestern Venezuela; southeastern Brazil (Figure 19).
Discussion.—Section Leptidium is an allotetraploid (4x) lineage, derived from ancient hybridisation and chromosome doubling of the common ancestor of subgenus Viola and the most recent common ancestor of sect. Leptidium and sect. Tridens; this allopolyploidisation may have happened c. 15 Ma ago [28]. A comprehensive phylogeny of sect. Leptidium has not been published. While V. arguta appears to be 4x, further allopolyploidisation has occurred in V. stipularis (8x). The count of n = 27 in V. dombeyana (as V. humboldtii Tr. & Pl. [53]), presumably referring to the 8x level as well, is the only count for the section and needs confirmation.
This widely distributed Latin American lineage encompasses 17 species and possibly the mysterious V. producta W. Becker. Viola scandens and V. stipularis account for the Mesoamerican and Antillean portions of the range of the section, with four species in southeastern Brazil and 13 (14?) occupying middle and higher elevations of the northern and central Andean Mountains in South America. All species have petals glabrous within, and all share a peculiar synapomorphy of prolonged “u”-shaped anther connective appendages on the bottom pair of stamens, first documented in two Brazilian species by Freitas and Sazima [171]. The Mesoamerican and southeastern Brazilian lineages may have diverged 8.7 (3–16) Ma ago [28].
A transition from nectar to pollen flowers and “buzz” pollination has been suggested for the majority of the species within sect. Leptidium; the unique “u” shape of the connective stamen appendages appears to be an adaptation to this [171]. The flowers are pollinated by Anthrenoides bees (Andrenidae) that hold onto the connective appendages while harvesting pollen by vibration (“buzz-pollination”) [171], and may thus be analogous to similar structures in unrelated genera (e.g., Arbutus, Ericaceae). Curiously, a secondary transition to hummingbird pollination, unique in the genus, seems to have occurred in V. arguta; this species produces copious amounts of nectar, 4 µL per 24 h [174] but it has also preserved the “u”-shaped connective appendages indicative of ancestal “buzz”-pollination.
Viola sect. Melanium Ging. in Mém. Soc. Phys. Genève 2(1): 28. 1823 ≡ Viola subg. Melanium (Ging.) Peterm., Deutschl. Fl.: 65. 1846; (Ging.) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 221. 1909 (isonym)—Type: Viola tricolor L.
≡Mnemion Spach, Hist. Nat. Vég. [Spach] 5: 510. 1836—Lectotype (Nieuwland & Kaczmarek 1914 [249], page 210): Viola tricolor L.
≡Viola sect. Pogonostylos Godron, Fl. Lorraine, ed. 2, 1: 90. 1857, nom. illeg. superfl. (Szhenzhen Code Art. 52.1; Viola tricolor L.)
≡Viola sect. Novercula Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 225. 1909, nom illeg. superfl. (Szhenzhen Code Art. 52.1; Viola tricolor L.)
=Jacea Opiz in Bercht. & Opiz, Oekon.-Techn. Fl. Böhm. [Berchtold & al.] 2(2): 8. 1839, nom. illeg., non Mill., Gard. Dict. Abr., ed. 4: [not paginated]. 1754 (=Centaurea)
Description.—Annual to perennial herbs. Taproot preserved, in perennials often deeply buried and thickened. Axes not morphologically differentiated. All stems more or less aerial; in perennials proximal portion rhizome-like. Stipules usually foliaceous, pinnately or palmately lobed with leaflike segments. Lamina entire, crenulate or crenate, petiolate. Corolla small or large (bottom petal 2–34 mm), often varicoloured and/or variegated, nearly always with a yellow throat. Spur short or long and slender (0.9–16 mm). Calycine appendages short or long (0.5–5.3 mm). Style capitate, bearded. Cleistogamous flowers usually not produced; if produced, then cleistogamy seasonal (V. rafinesquei). Allotetraploid (CHAM + MELVIO). ITS sequence of MELVIO type. Aneuploid.
Diagnostic characters.—All stems more or less aerial AND stipules usually foliaceous AND corolla small to very large, nearly always with a yellow throat.
Ploidy and accepted chromosome counts.—4x, 8x, 12x, 16x, 20x, >20x; 2n = 4, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 34, 36, 40, 48, 52, c.64, c.96, 120, c. 128.
Age.—Crown node age 12.5 (11.8–12.8) Ma [28].
Included species.—112.
Distribution.—Western Eurasia; one species in eastern North America (Figure 20). Mainly in mountainous areas, with a centre of diversity in the mountains of the Balkans, Apennine Peninsula and Sicily, seven species in northwestern Africa (three of them endemic) and one species in eastern North America (Viola rafinesquei). A few species are widespread in the lowlands, nearly all annuals or biennials (e.g., V. arvensis, V. tricolor, and V. rafinesquei).
Discussion.—Section Melanium is phylogenetically an allotetraploid CHAM + MELVIO lineage having retained the MELVIO homoeolog for ITS (Figure 6). Morphologically the section is highly characteristic, primarily by the annual or perennial habit with undifferentiated stems, the often large and leaf-like, usually deeply divided stipules, the orbicular, ovate, lanceolate or linear, crenate (or entire) laminas, often also by pronounced heterophylly, the small to very large, often multicoloured, corolla with a yellow throat (cream throat in, e.g., V. argenteria, V. cornuta, and V. orthoceras), the unique capitate-bearded style, and the absence of cleistogamous flowers (present in V. rafinesquei). Section Melanium is morphologically distinct and has by numerous authors been ranked as subgenus or even genus (Mnemion Spach and Jacea Opiz non Mill.). However, molecular data place it firmly among the other north hemisphere allotetraploid lineages, albeit with very long branches, suggesting that its morphological differentiation is a result of rapid evolution rather than deep divergence. The subtending branch of Melanium is also long and its diversification did not start until 12–13 Ma ago, corresponding with the onset of a global climatic cooling trend from c. 14 Ma ago [31]. Subsection Bracteolatae, which comprises most of the species, started diversifying 9–10 Ma ago [28]. In line with a relatively recent origin, the detailed evolutionary relationships within sect. Melanium remain elusive when based on markers such as ITS, chloroplast loci, and ISSRs [94,217]. The low-copy genes used by Marcussen et al. [28] revealed high ploidy levels for the three species sampled. These findings are corroborated also by the occurrence of numerous loci for ribosomal DNA in the section [250,251,252]. At present (2022), transcriptome data exist for Viola tricolor only [236].
Section Melanium is characterised by an extraordinarily high karyological diversity and plasticity which imply that ploidy can not be inferred from chromosome numbers alone. Here, we estimate the ploidy of 12 taxa in sect. Melanium (Table 2) from the number of homoeologs of the low-copy nuclear gene GPI [28] and genome size estimated from flow-cytometry [218,219,220,221,253] and for the Earth Biogenome Project (EBP [233]). These data in combination indicate that subsect. Ebracteatae is 4x (V. dirimliensis), subsect. Cleistogamae is 8x (V. rafinesquei), and that subsect. Bracteolatae comprises several ploidy levels, at least 8x (V. kitaibeliana, V. beckiana, V. elegantula, V. cornuta), 12x (V. tricolor), 16x (V. arvensis, V. calcarata), and 20x (V. lutea, V. bubanii). Given that the 1Cx-values within subsect. Bracteolatae are stable around 0.27–0.30 pg, changes in chromosome number in these taxa are due to chromosome fusions rather than to loss or deletions. Still, karyotype homology seems preserved at least in some allopolyploids containing the homologous ancestral 4x genomes, because considerably good chromosome pairing during the meiosis and fertility occurs in some heteroploid hybrids (e.g., [37,66,254,255]). Such chromosome fusions appear to occur throughout sect. Melanium, and are at the most extreme in subsect. Ebracteatae: in the presumably tetraploid V. modesta (2n = 4) the ancestral four monoploid genomes have probably fused to just four chromosomes. The highest widespread chromosome number is 2n = 52 (20x), found in six species of subsect. Bracteolatae, while chromosome numbers above this value (e.g., 2n = 64, 96, 120, and 128, the last two in V. bubanii) may have been counted in hybrids or aberrant individuals and require confirmation. It seems futile to try estimating the base chromosome number x for sect. Melanium, knowing that the nascent Melanium allotetraploid likely started out with n = 10 to 12 chromosomes just like the other CHAM + MELVIO tetraploids, and knowing that reductions in chromosome number have occurred independently in different sublineages of this section. Not surprisingly, each attempt until now has produced a different x, i.e., x = 5 [217], x = 6 [56], x = 7 [217], x = 10 [56], and x = 11 [255].
Although a detailed revision of sect. Melanium must await comprehensive phylogenomic study of the lineage, the already available data from phylogeny [28,94,217], morphology, and genome size and pollen aperture number which reflects ploidy [157], yield sufficient resolution to delimit five sublineages within sect. Melanium (Table 3). These lineages form morphologically and biogeographically recognisable units but do not conform with previous classifications (e.g., [1,21,72,133]). Below we formally introduce them as subsections, i.e., (1) subsect. Bracteolatae, (2) subsect. Cleistogamae, (3) subsect. Dispares, (4) subsect. Ebracteatae, and (5) subsect. Pseudorupestres. The vast majority of species belong in subsect. Bracteolatae and only a dozen in the other four subsections which may well be considered relictual and phylogenetically isolated.
Key to the subsections of sect. Melanium:
- 1a.
- Cleistogamous flowers produced. Annual or biennial. (eastern North America) ..................................................................................................................................................................................................................... subsect. Cleistogamae (V. rafinesquei)
- 1b.
- Cleistogamous flowers not produced. Annual to perennial. (Palaearctic, elsewhere alien) .................................................................................................................................................................................................................................................................................. 2.
- 2a.
- Corolla violet, with a cream-coloured throat. Stipules ovate-lanceolate, dentate. Bottom petal 9.5–10.5 mm. Low, high-Alpine perennial. (western Alps and Corsica) ....................................................................................................................................................................................................................... subsect. Pseudorupestres (V. nummulariifolia)
- 2b.
- Corolla colour various, with a bright yellow throat (if throat cream or white and lateral petals directed horizontally or downwards: V. cornuta and V. orthoceras). Stipules variable, often foliaceous, rarely dentate. Bottom petal 2–34 mm. Annual or perennial. ..................................................................................................................................................................................................................................................................... 3.
- 3a.
- Annual. Basal leaves entire or indistinctly crenulate. Bottom petal 2–11.5 mm. Spur 0.9–3 mm. ......................................................................................................................................................................................................................................................... subsect. Ebracteatae
- 3b.
- Annual to perennial. Lamina crenate or entire, but if annual then lamina of basal leaves crenate. Bottom petal 5–34 mm. Spur 1–16 mm. .................................................................................................................................................................................................................................................................................... 4.
- 4a.
- Calycine appendages 0.3–1.0 mm long. Bottom petal 5–13 mm. Spur 1–3.5 mm. .......................................................................................................................................................................................................................................................... subsect. Dispares
- 4b.
- Calycine appendages 0.9–4.7 mm long. Bottom petal 5.4–34 mm. Spur 1.8–16 mm. ........................................................................................................................................................................................................................................................ subsect. Bracteolatae
Viola subsect. Bracteolatae Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 228. 1909—Lectotype (designated here): Viola tricolor L.
≡Viola subsect. Melanium (Ging.) Vl. V. Nikitin in Bot. Zhurn. (Moscow & Leningrad) 83(3): 135. 1998—Type: Viola tricolor L.
=Viola sect. Pseudonovercula Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3: 222. 1909—Type: Viola cornuta L.
Description.—Annual to perennial. Lamina of basal leaves entire or crenate, but if plants annual then lamina crenate. Calycine appendages 0.9–4.7 mm. Corolla with bright yellow, rarely pale yellow throat. Bottom petal (spur included) 5.5–34 mm. Spur 1.8–16 mm. Cleistogamous flowers not produced. Pollen apertures 4 or 5 heteromorphic. Ploidy 8x, 12x, 16x, 20x, >20x.
Diagnostic characters.—See Table 3 and key.
Ploidy and accepted chromosome counts.—8x, 12x, 16x, 20x, >20x; 2n = 16, 18, 20, 22, 24, 26, 28, 34, 36, 40, 48, 52, c. 64, c. 96, 120, c. 128.
Age.—Crown node c. 4 Ma (Figure 6), probably an underestimate; stem node age 9.8 (9.1–10.0) Ma [28].
Included species.—98 (in addition the two ornamental hybrids Viola ×williamsii Wittr. and V. ×wittrockiana Gams). Viola acrocerauniensis Erben, V. aethnensis (Ging.) Strobl, V. aetolica Boiss. & Heldr., V. albanica Halácsy, V. allchariensis Beck, V. alpina Jacq., V. altaica Ker Gawl., V. arsenica Beck, V. arvensis Murray, V. athois W. Becker, V. babunensis Erben, V. beckiana Fiala ex Beck, V. bertolonii Pio, V. bornmuelleri Erben, V. brachyphylla W. Becker, V. bubanii Timb.-Lagr., V. calcarata L., V. cenisia L., V. cephalonica Bornm., V. cheiranthifolia Bonpl., V. comollia Massara, V. cornuta L., V. corsica Nyman, V. crassifolia Fenzl, V. crassiuscula Bory, V. cryana Gillot, V. culminis F. Fen. & Moraldo, V. dacica Borbás, V. declinata Waldst. & Kit., V. dichroa Boiss., V. diversifolia (Ging.) W. Becker, V. doerfleri Degen, V. dubyana Burnat ex Gremli, V. dukadjinica W. Becker & Košanin, V. elegantula Schott, V. epirota (Halácsy) Raus, V. etrusca Erben, V. euboea Halácsy, V. eugeniae Parl., V. eximia Formánek, V. ferrarinii Moraldo & Ricceri, V. fragrans Sieber, V. frondosa (Velen.) Velen., V. ganiatsasii Erben, V. gostivariensis Bornm., V. gracilis Sm., V. graeca (W. Becker) Halácsy, V. grisebachiana Vis., V. guaxarensis M. Marrero, Docoito Díaz & Martín Esquivel, V. heldreichiana Boiss., V. henriquesii (Willk. ex Cout.) W. Becker, V. herzogii (W. Becker) Bornm., V. hispida Lam., V. hymettia Boiss. & Heldr., V. ivonis Erben, V. kitaibeliana Schult., V. kopaonikensis Pančić ex Tomović & Niketić, V. langeana Valentine, V. lutea Huds., V. magellensis Porta & Rigo ex Strobl, V. merxmuelleri Erben, V. minuta M. Bieb., V. montcaunica Pau, V. munbyana Boiss. & Reut., V. nana (DC. ex Ging.) Le Jol., V. nebrodensis C. Presl, V. odontocalycina Boiss., V. orbelica Pančić, V. oreades M. Bieb., V. orphanidis Boiss., V. orthoceras Ledeb., V. palmensis (Webb & Berthel.) Sauer, V. paradoxa Lowe, V. parnonia Kit Tan, Sfikas & Vold, V. perinensis W. Becker, V. phitosiana Erben, V. pseudaetolica Tomović, Melovski & Niketić, V. pseudogracilis (A. Terracc.) Strobl ex Degen & Dörfl., V. pseudograeca Erben, V. raunsiensis W. Becker & Košanin, V. rausii Erben, V. rhodopeia W. Becker, V. roccabrunensis Espeut, V. samothracica (Degen) Raus, V. schariensis Erben, V. serresiana Erben, V. sfikasiana Erben, V. slavikii Formánek, V. stojanowii W. Becker, V. striis-notata (J. Wagner) Merxm. & W. Lippert, V. subatlantica (Maire) Ibn Tattou, V. tineorum Erben & Raimondo, V. tricolor L., V. ucriana Erben & Raimondo, V. valderia All., V. velutina Formánek, V. voliotisii Erben, V. vourinensis Erben
Distribution.—Western Eurasia.
Discussion.—Sect. Melanium subsect. Bracteolatae comprises the vast majority of the species in the section and is difficult to describe (Table 3). The lineage is phylogenetically characterised by being at least 8-ploid (Table 2), karyologically by a variety of chromosome numbers, and morphologically by the sometimes very large corollas with bottom petal up to 32 mm long, and usually heteromorphic mostly 4-colporate, rarely 5-colporate pollen [157]. The diversification in subsect. Bracteolatae is evidently recent and may at least partly have been driven by geographic isolation in combination with homoploid and heteroploid hybrid speciation [254], as indicated from chromosome counts, crossing experiments [254], genome size variation (Table 2), and subcloning of nuclear genes and ribotypes [28,250,252]. Not surprisingly, the two phylogenies of sect. Melanium [94,217], both using ITS, showed little variation among species. The evolutionary relationships within subsect. Bracteolatae are poorly understood. However, our preliminary interpretation based on all available lines of evidence is that the subsection comprises at least 3–4 independent homoeologous genome lineages that occur in different variants, numbers and combinations among the different species. In some cases the shared subgenomes are similar enough to allow for gene flow among different species despite differences in ploidy, such as between V. tricolor (2n = 26; 12x) and V. arvensis (2n = 34; 16x), whereas in other species pairs the subgenomes are too dissimilar to allow for gene flow or even hybrid formation, such as between V. tricolor and V. cornuta (2n = 22; 8x) [254]. The available morphological, genetical [254] and molecular evidence from ITS [94] and 5S-IGS [252] suggest that, for instance, V. heldreichiana, V. kitaibeliana, V. hymettia (all 2n = 16; 8x), V. tricolor (12x) and V. arvensis (16x) form a polyploid series. Furthermore, the species with 2n = 20 (8x) and 2n = 40 (16x) of the Alps, Dinarids, Apeninnes and Sicily, traditionally referred to as the V. calcarata group [94], are probably closely related. The Pyrenean V. cornuta and the Caucasian V. orthoceras (both with several shared, rather unique character states; 2n = 22) are probably geographic isolates. Viola tricolor and V. arvensis are cosmopolitan weeds. Viola ×williamsii and V. ×wittrockiana are grown as ornamentals.
Viola subsect. Cleistogamae Marcussen & Danihelka, subsect. nov.—Type: Viola rafinesquei Greene (=V. bicolor Pursh non Hoffm.)
Description.—Annual to biennial. Lamina of basal leaves crenate. Calycine appendages 0.5–2 mm. Corolla with bright yellow throat. Bottom petal (spur included) 8–10 mm. Spur 1–1.5 mm. Cleistogamous flowers produced. Pollen apertures 4. Ploidy 8x. Chromosome number n = 17.
Diagnostic characters.—Cleistogamous flowers produced.
Ploidy and accepted chromosome counts.—8x; 2n = 34.
Age.—Crown node age not applicable (monotypic subsection), stem node age 9.8 (9.1–10.0) Ma [28].
Included species.—1. Viola rafinesquei Greene
Distribution.—Eastern North America.
Etymology.—The name Cleistogamae refers to the occurrence of seasonal cleistogamy in the type species.
Distribution.—Section Melanium subsect. Cleistogamae comprises Viola rafinesquei (=V. bicolor Pursh non Hoffm.) only, which differs from all other pansies in two key respects: it has seasonal cleistogamy, i.e., produces chasmogamous flowers in spring (after vernalisation) and cleistogamous ones later in summer, and its native range is in eastern North America while all the other Melanium species have their native ranges in the western Palearctic. Cleistogamy in V. rafinesquei involves reduction of the four lower anthers, unlike in other Viola where the three upper anthers are reduced [256], suggesting cleistogamy in these lineages is not entirely homologous. Viola rafinesquei has the chromosome number 2n = 34 and 4-colporate pollen, and is an octoploid [28]. Different views on the nomenclature of V. rafinesquei have been proposed [257,258] but we hold that of Shinners [257] to be correct. For general taxonomy, see Clausen et al. [256].
The subsections Cleistogamae and Bracteolatae appear to be monophyletic at the octoploid level and may have split 9–10 Ma ago [28]. The two are, however, genetically distant and cannot be crossed successfully [256].
Viola subsect. Dispares Marcussen & Danihelka, subsect. nov.—Type: Viola dyris Maire
Description.—Ephemeral annual or dwarf perennial. Lamina of basal leaves entire or crenate. Calycine appendages 0.3–1 mm. Corolla with bright yellow throat. Bottom petal (spur included) 5–13 mm. Spur 1–3.5 mm. Cleistogamous flowers not produced. Pollen apertures 3 or 4. Ploidy probably 4x, 8x.
Diagnostic characters.—See Table 3 and key.
Ploidy and accepted chromosome counts.—Probably 4x, 8x; 2n = 12 (Viola poetica), 20, 22 (V. dyris), 24 (V. demetria).
Included species.—3. Violademetria Prolongo ex Boiss., V. dyris Maire, V. poetica Boiss. & Spruner
Distribution.—Disjunctly distributed in the Mediterranean area of southern Europe and northern Africa: Viola dyris in Morocco (High Atlas), V. demetria in southernmost Spain (Andalusia), and V. poetica in central Greece (Parnassos).
Etymology.—The name Dispares refers to the strikingly different general habits and life histories, and few apomorphic characters for this subsection.
Discussion.—Section Melanium subsect. Dispares is the third and last lineage nested within the basal polytomy of sect. Melanium (Figure 6). We infer that the subsection comprises three species, V. demetria, V. dyris, and V. poetica. The last species has not been investigated phylogenetically, but monophyly is strongly supported for the other two species using both ITS and chloroplast sequence data [94]. The very short calycine appendages (<1 mm) are an apomorphy for the subsection. Furthermore, all three species have stipules with the main segment resembling the lamina (crenulate in V. demetria with 0–3 narrow basal segments [i.e., palmate], entire and undivided in the other two) and small corollas (c. 5 mm in V. dyris, up to 13 mm in the other two). In both V. demetria and V. poetica the spur is intensively violet, and thicker and almost saccate in V. demetria. In other respects the three species are morphologically disparate, which probably reflects their adaptations to different extreme environments, i.e., to high-Alpine habitats in the perennials V. dyris (scree) and V. poetica (rock crevices and screes) as opposed to summer-dry habitats with a short growing season in the ephemeral annual V. demetria. The three species are also highly disjunct. Viola poetica (2n = 12) has 3-colporate pollen and is probably 4x, while V. dyris (2n = 20, 22) and V. demetria (2n = 24) both have 4-colporate pollen [157] and are probably 8x. The chromosome numbers 2n = 12 (V. poetica) and 2n = 24 (V. demetria) form a polyploid series; the former is unique and the latter extremely rare among pansies [65,66]. The divergence of V. demetria and V. dyris may have been relatively recent, only c. 2.7 Ma (Figure 2).
Viola subsect. Ebracteatae Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3: 225. 1909 ≡ Viola ser. Ebracteatae (Kupffer) Vl. V. Nikitin in Bot. Zhurn. (Moscow & Leningrad) 83(3): 135. 1998—Lectotype (Nikitin 1998 [72], page 135): Viola modesta Fenzl
Description.—Ephemeral annuals. Lamina of basal leaves entire or subcrenate. Calycine appendages 0.5–5.3 mm. Corolla with bright yellow throat. Bottom petal (spur included) 2–11.5 mm. Spur 0.9–3 mm. Cleistogamous flowers not produced. Pollen apertures 3 or heteromorphic 4. Ploidy 4x, 8x, >8x.
Diagnostic characters.—Annuals AND lamina of basal leaves entire or subcrenate.
Ploidy and accepted chromosome counts.—4x, 8x, >8x; 2n = 4, 8, 10, 20, 36.
Included species.—9. Violadenizliensis O. D. Düsen, Göktürk, U. Sarpkaya & B. Gürcan, V. dirimliensis Blaxland, V. ermenekensis Yıld. & Dinç, V. mercurii Orph. ex Halácsy, V. modesta Fenzl, V. occulta Lehm., V. parvula Tineo, V. pentadactyla Fenzl, V. rauliniana Erben
Distribution.—Western Eurasia. Diversity centre in the eastern Mediterranean area.
Discussion.—Section Melanium subsect. Ebracteatae is the second lineage nested within the basal polytomy of sect. Melanium (Figure 6). This lineage is characterised phylogenetically by being partly tetraploid [28], karyologically by having very low chromosome numbers (2n = 4, 8, 10; polyploid 2n = 20, 36), and morphologically by being small-flowered ephemeral annuals (bottom petal 2–11.5 mm) with entire or subcrenate basal leaves. In most species the appendages of the two lower sepals are conspicuously longer than those of the other sepals (not in V. denizliensis and V. dirimliensis). The tetraploids have small, monomorphic 3-colporate pollen [157].
Viola subsect. Pseudorupestres (W. Becker) Marcussen & Danihelka, comb. et stat. nov.—Basionym: Viola [sect. Melanium; unranked] “γ” Pseudorupestres W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 372. 1925 (“Pseudo-rupestres”).—Type: Viola nummulariifolia Vill.
Description.—Dwarf perennial. Stipules dentate, not foliaceous. Lamina of basal leaves entire. Calycine appendages 1.2–1.5 mm. Corolla violet with cream throat. Bottom petal (spur included) 9.5–11.5 mm. Spur 2.3–2.5 mm. Cleistogamous flowers not produced. Pollen apertures 3. Ploidy probably 4x. Chromosome number n = 7.
Diagnostic characters.—Dwarf perennial AND stipules dentate, not foliaceous AND corolla violet with cream throat.
Ploidy and accepted chromosome counts.—Probably 4x; 2n = 14.
Age.—Crown node age not applicable (monotypic subsection), stem node age c. 7.2 Ma (Figure 6).
Included species.—1. Viola nummulariifolia Vill.
Distribution.—Southern Europe: Maritime Alps and Corsica.
Discussion.—Section Melanium subsect. Pseudorupestres comprises a single species, Viola nummulariifolia Vill. (=V. argenteria Moraldo & Forneris). Chloroplast and ITS data place it in (or as sister to) the basal polytomy within sect. Melanium [94] (Figure 2), and in a PCO of genomic ISSR data the species ends up ‘close’ to the outgroup [217]. Furthermore, other attributes seem to suggest an isolated phylogenetic position. The low chromosome number of 2n = 14 and the 3-colporate pollen [157] indicate low ploidy level, presumably 4x, the ancestral condition in sect. Melanium [28]. Morphologically, V. nummulariifolia has a suite of character states that might be interpreted as plesiomorphic, e.g., perenniality, the flower having a cream throat (not bright yellow as in most other pansies) and simple, entire to dentate stipules, reminiscent of V. rupestris (sect. Viola), and not large and foliaceous as in many other pansies. Viola nummulariifolia has a relictual distribution at high elevations (1800–2900 m) on crystalline rocks in the Maritime Alps and in Corsica [259].
Viola sect. Melvio Marcussen, sect. nov.—Type: Viola decumbens L. f.
Description.—Perennial subshrubs. Axes not morphologically differentiated. All stems aerial. Stipules somewhat adnate, green, linear, with 1–2 basal teeth. Lamina entire, linear, subapiculate and somewhat succulent. Bracteoles persistent, 1–3 mm. Corolla violet with a white throat. Spur slender, yellow or orange. Style dorsiventrally flattened and tapering towards the tip, in lateral view filiform and sigmoid. Cleistogamous flowers not produced. Allopolyploid (MELVIO).
Diagnostic characters.—Style dorsiventrally flattened and tapering towards the tip, in lateral view filiform and sigmoid. Allopolyploid (MELVIO).
Ploidy and accepted chromosome counts.—Allopolyploid, possibly 6x; chromosome number unknown.
Age.—Crown node age not applicable (monotypic section), stem node age 20.5–22.6 Ma [28].
Included species.—1. Viola decumbens L. f.
Distribution.—South Africa: Cape region (Figure 21).
Etymology.—Section Melvio is named after the lineage to which it belongs, the diploid MELVIO lineage, for which Viola decumbens is the only extant species. The name was originally applied by T.M. [88] to delimit a clade in the ITS phylogeny of Ballard et al. [2] which comprised sect. Melanium (“MEL”) and sect. Viola (“VIO”) only, as a result of ITS homoeolog loss and limited sampling.
Discussion.—Section Melvio comprises a single species, Viola decumbens (Figure 4), a shrublet with an isolated distribution in the fynbos of the southern Cape of South Africa [260]. It is the sole member of the otherwise extinct Eurasian MELVIO clade and sister to the taxa involved in the dozen of allopolyploidisations that occurred in Eurasia 15–19 Ma ago. Viola decumbens may have been isolated in South Africa since Early Miocene, 20–25 Ma ago [28]. Viola decumbens is allopolyploid, possibly paleohexaploid, based on gene copy number for two nuclear genes [28]. The species was previously included in sect. Xylinosium [1,28], to which it is superficially similar in shrubby habit. It differs, however, from sect. Xylinosium in several key traits. These include the style, which in V. decumbens is characteristically dorsiventrally flattened and tapering towards the tip, in lateral view filiform and sigmoid, vs. clavate in sect. Xylinosium; the leaves which in V. decumbens are entire, linear, subapiculate and somewhat succulent vs. lanceolate and usually crenate in sect. Xylinosium; the bracteoles which are 3–5 mm and persistent in V. decumbens vs. 1–2 mm or caducous in sect. Xylinosium; and the indument of stems and leaves, minutely papillate in V. decumbens and distinctly longer in sect. Xylinosium (sometimes glabrous or ciliate). The inclusion of V. decumbens in sect. Xylinosium by Marcussen et al. [28] was a mistake relating to a chloroplast sequence of V. arborescens (sect. Xylinosium) that had been erroneously assigned to V. decumbens (trnL-trnF; KJ138159). Indeed, another chloroplast sequence (rbcL; AM235165) places this species in agreement with the nuclear homoeologs, of which none are shared between these two taxa.
Viola sect. Nematocaulon Marcussen, Nicola, J. M. Watson, A. R. Flores, H. E. Ballard, sect. nov.—Type: Viola filicaulis Hook. f.
Description.—Perennial herbs. Axes not morphologically differentiated: all stems creeping, branched and remotely noded. Stipules ovate, free, remotely long-fimbriated. Lamina reniform to ovate, few-crenate, long-petiolate. Corolla small, white with violet striations, with a golden yellow throat. Spur short, yellow. Style filiform, terminated in a quadrangular stigmatic opening. Cleistogamous flowers produced; cleistogamy facultative. Chromosome number n = 36.
Diagnostic characters.—Corolla with a yellow throat AND style filiform.
Ploidy and accepted chromosome counts.—Ploidy unknown; 2n = 72.
Age.—Unknown.
Distribution.—New Zealand (Figure 22).
Included species.—1. Viola filicaulis Hook. f.
Etymology.—The name Nematocaulon is a Greek translation of the species epithet of the type species, Viola filicaulis, which refers to the creeping stems of that species.
Discussion.—Viola filicaulis is distinct from all other groups and species of violets, as noted already by Hooker [261] in the protologue. Becker [1] noted in the introduction to his treatment of Viola that V. filicaulis was sufficiently distinct to be placed in a section of its own, although he did not erect one. DNA samples of V. filicaulis have not been available for phylogenetic analysis. However, its morphological affinities are clearly with the other southern hemisphere sections of subg. Viola. In having a filiform style it is most similar to the species of sect. Tridens, sect. Erpetion, and sect. Leptidium. In the violet-striate pigmentation and shape of the corolla it approaches sect. Tridens (which, however, lacks the yellow throat) and in expressing facultative cleistogamy it is similar to sect. Chilenium and sect. Leptidium. The high chromosome number of V. filicaulis (2n = 72 [262]) also agrees with polyploidy in all of these sections. At the same time, style shape, stem not differentiated in a rhizome and lateral stems, and facultative cleistogamy effectively exclude an affinity of V. filicaulis to the morphologically superficially similar sections in the northern hemisphere (i.e., Chamaemelanium, Nosphinium, Plagiostigma, and Viola).
Viola filicaulis produces cleistogamous flowers in abundance, both seasonally (during summer) and facultatively under unfavourable conditions. These are, however, more morphologically variable and appear less specialised (petals reduced but not absent, number of fertile stamens variable) than in the sympatric V. cunninghamii which belongs in sect. Plagiostigma subsect. Bilobatae and which has a north-temperate origin [26,263].
Viola sect. Nosphinium W. Becker in Engler, Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 374. 1925 ≡ Viola subg. Nosphinium (W. Becker) Espeut in Botanica Pacifica 9(1): 34. 2020.—Lectotype (Espeut 2020 [61], page 34): Viola chamissoniana Ging.
Description.—Perennial herbs (subshrubs or treelets in most species of subsect. Nosphinium). Axes in some species morphologically differentiated into a perennating stem and annual aerial stems (subsect. Langsdorffianae, modified in subsect. Nosphinium) or stolons (some species of subsect. Mexicanae). Perennating stem usually a rhizome, deep or shallow, vertical or horizontal, terminating in an apical rosette. Stipules membranous or partially herbaceous, free (basally or strongly adnate in a few species of subsect. Mexicanae), glandular-fimbriate to glandular-laciniate. Lamina various, long-petiolate. Corolla violet (white in a few species of subsect. Mexicanae and subsect. Nosphinium), throat white. Calycine appendages short and rounded or elongate, quadrate and often dentate, often elongated somewhat in cleistogamous fruit. Petals large, lateral petals glabrous or sparsely to densely bearded within (spurred petal bearded in some species of subsect. Borealiamericanae). Spur thick, as long as tall (sometimes nearly twice as long as tall in V. langsdorffii). Style cylindrical with slight subapical swelling (subsect. Langsdorffianae), or clavate with apex flanked by a dorsolateral sharp edge or protruding thickened apically oriented or spreading rim, stigma on a pronounced rostellum. Cleistogamous flowers produced, mostly seasonal (cleistogamy absent in subsect. Pedatae and in most species of subsect. Nosphinium). Allodecaploid with one 2x genome from sect. Chamaemelanium, one or more 4x genomes from sect. Plagiostigma, and one 4x genome from sect. Viola. ITS sequence of MELVIO (sect. Viola) type. Inferred secondary base chromosome number [x’ = 28].
Diagnostic characters.—Rosulate habit (rarely stoloniferous or with aerial stems) AND rhizome thick AND stipules free (rarely adnate) AND corolla violet (rarely white) AND petals large AND spur thick and short AND style clavate with dorsolateral edge or thickened rim and pronounced rostellum (rarely with merely a weak dorsolateral swelling) AND allodecaploid with one 2x genome from sect. Chamaemelanium, one or more 4x genomes from sect. Plagiostigma, and one 4x genome from sect. Viola.
Ploidy and accepted chromosome counts.—10x, 14x, 18x, ~22x; 2n = c. 44, 54, c. 76, 80, c. 80, c. 85, c. 86, c. 96, 102, c. 120.
Age.—Crown node 8.4 (7.5–8.8) Ma [28].
Included species.—62.
Distribution.—North America, Hawaiian Islands, Mexico and Central America, a few species in northern South America, one species in northeastern Asia (Figure 23).
Discussion.—Sect. Nosphinium is a young and nearly exclusively North American radiation. The lineage is allodecaploid and derived from successive hybridisation between North American members of sect. Chamaemelanium grex Nudicaules (2x), sect. Plagiostigma subsect. Stolonosae (4x), and sect. Viola (4x; Figure 24); it has retained the ITS homoeolog of the sect. Viola parent (Figure 6) and the chloroplast of the sect. Plagiostigma parent [2,45,81]. Section Nosphinium comprises five of Becker’s [1] infrageneric taxa, i.e., sect. Nosphinium in the strict sense (the Hawaiian violets sensu Becker) and sect. Nomimium greges Borealiamericanae, Pedatae, Mexicanae, and Langsdorffianae (excluding V. moupinensis). These five taxa, in addition to V. clauseniana, represent different lineages that we recognise at the subsection level. Section Nosphinium consists of two principal lineages, a western, Pacific lineage and an eastern lineage. The western lineage gave rise to subsects. Nosphinium and Langsdorffianae by independent allopolyploidisations with various sect. Plagiostigma taxa, bringing the ploidy of these lineages to 14x and 18x, respectively (Figure 24). The eastern lineage gave rise to subsects. Borealiamericanae, Clausenianae, and Pedatae without change in ploidy (10x) and subsect. Mexicanae (14x) by yet another allopolyploidisation with another sect. Plagiostigma taxon (Figure 24).
Morphologically, the members of sect. Nosphinium are a “compromise” among the three parental sections, except for their larger stature which probably reflects higher ploidy. The predominantly violet corolla is shared with sect. Viola and the short spur with the other two parents. The style shape is intermediate between sects. Plagiostigma and Viola. The ability of forming lobed or dissected leaves is shared with sect. Chamaelenanium. The lanceolate sepals are more similar to sects. Plagiostigma and Viola than to sect. Chamaemelanium which generally has narrow-lanceolate sepals.
Apart from the unique decaploidisation that gave rise to sect. Nosphinium, the section is difficult to characterise with unique morphological synapomorphies, given that some lineages were produced by secondary allopolyploidisations involving ancestors of diverse subsections in sect. Plagiostigma. It is much easier to distinguish the subsections recognised here. Generally, the section is distinguished by a rather thick rhizome, typically large stature, near absence of stolons (present only in some Mexicanae), and a short thick spur. Caulescent subsections Langsdorffianae and Nosphinium (woody except V. kauaensis) have broad semi-sheathing stipules and a broadly rounded or convex style apex bent into a slender or thick rostellum but lacking a distinct thickened dorsolateral rim; acaulescent subsections Clausenianae and Pedatae have partially to almost completely adnate stipules, the former with a style strongly protruded and conspicuously thickened dorsally and a short strongly incurved ventral rostellum, the latter with a style lacking dorsal elongation and merely with a thin dorsolateral margin surrounding a concavity which hides the ventral stigmatic orifice. Subsection Borealiamericanae lacks stolons, has lateral petals always densely bearded and bottom petal bearded in some species, and a style with a well developed spreading conspicuously thickened dorsolateral rim, while subsect. Mexicanae often produces stolons, has lateral petals beardless or sparsely bearded, bottom petal beardless, and a style with a weak dorsolateral rim oriented forward.
Key to the subsections of sect. Nosphinium
- 1a.
- Plant caulescent with aerial stems. Lower stipules broad, triangular or ovate, sheathing the stem. Style apex bent to form a short slender or broad blunt rostellum or tip, the rostellum flanked by a weak dorsolateral swelling. ............................................................................................................................................................................................................................................................................. 2
- 1b.
- Plant acaulescent, stolons present or absent. Stipules narrow, linear-lanceolate, not sheathing the stem. Style apex with a pronounced thickened dorsolateral margin flanking the prominent rostellum (dorsolateral margin thin and rounded with stigmatic orifice hidden in the cavity in subsect. Pedatae). ............................................................................................................................................................................................................................................................................... 3
- 2a.
- Flowers white or nearly so on the inside, lacking violet striation. Shrubs or subshrubs, rarely rhizomatous herb with reclining to ascending aerial stems (V. kauaensis). Lower stipules triangular, acute to acuminate at apex, margins glandular-lacerate. Flowers often 1–4 together on lateral stems with reduced or absent leaves. Cleistogamous flowers not produced (present in V. kauaensis). (Hawaii) ........................................................................................................................................................................................................................................... subsect. Nosphinium
- 2b.
- Flowers violet, with dark violet striation. Herbs. Lower stipules broadly ovate, obtuse at apex, margins shortly glandular-fimbriate. Flowers solitary with well developed subtending leaves. Cleistogamous flowers produced. (northern Pacific region) ..................................................................................................................................................................................................................................................... subsect. Langsdorffianae
- 3a.
- At least the outer stipules adnate in basal 1/3 or nearly entirely to petiole. Petals beardless. ...................................................................................................................................................................................................................................................................................... 4
- 3b.
- Stipules free (outer mostly adnate to petiole in V. humilis). Lateral petals commonly bearded (bottom petal also bearded in some Borealiamericanae). ...................................................................................................................................................................................................................................................................................... 5
- 4a.
- Rhizome stout, vertical, and barrel-like. Stipules adnate for most of their length. Lamina typically deeply pedately divided. Style apex narrowly rounded from above, with dorsolateral margin as a narrowly rounded rim continuing to the ventral surface, the stigma hidden in the narrow triangular cavity created by the rim. Cleistogamous flowers not produced. ....................................................................................................................................................................... subsect. Pedatae (V. pedata)
- 4b.
- Rhizome relatively slender, oblique or somewhat horizontal, not barrel-like. Stipules adnate for up to 1/3 of their length. Lamina not divided, margins merely crenate. Style apex obtriangular from above, with dorsolateral margin protruding as a thickened broadly truncate or slightly emarginate rim continuing to the rostellum on the ventral surface, the rostellum apically oriented or incurved. Cleistogamous flowers produced. ............................................................................................................................................................................................................................................... subsect. Clausenianae (V. clauseniana)
- 5a.
- Stolons absent. Stipules free. Laminas in some species lobed or dissected. Calycine appendages elongate and dentate in some species. Corollas violet to dark violet. Lateral petals densely bearded, bottom petal bearded in some species. Style apex with pronounced thickened spreading broadly rounded, sometimes weakly trilobate dorsolateral rim, sides or lateral lobes continuing to the rostellum. (North America, V. nuevoleonensis in northern Mexico) ........................................................................................................................................................................................................................................ subsect. Borealiamericanae
- 5b.
- Stolons usually present (absent in V. beamanii, V. cuicochensis, V. hemsleyana, V. hookeriana, and V. humilis). Stipules free, or basally or mostly adnate. Laminas undivided. Calycine appendages short and entire. Corollas white or violet. Lateral petals beardless or sparsely bearded (sometimes densely in V. nubicola with violet corollas, and in V. grahamii and V. oxydontis with white corollas), bottom petal beardless. Style apex with weakly thickened apically oriented dorsolateral rim (somewhat prolonged and somewhat thickened dorsally in V. hookeriana) continuing partly or completely to rostellum. (Mexico to northern South America) ..................................................................................................................................................................................................................................... subsect. Mexicanae
Viola subsect. Borealiamericanae (W. Becker) Gil-ad in Bossiera 53: 42. 1997 (“Boreali-Americanae”; Shenzhen Code Art. 41.8) ≡ Viola [sect. Nomimium; unranked] Borealiamericanae W. Becker in Repert. Spec. Nov. Regni Veg. 19: 396. 1923 (“Boreali-Americanae”) ≡ Viola [sect. Plagiostigma] subsect. Borealiamericanae (W. Becker) Brizicky in J. Arnold Arb. 42: 327. 1961, nom. inval. (Shenzhen Code Art. 41.5; “Boreali-Americanae”) ≡ Viola [sect. Plagiostigma] subsect. Borealiamericanae (W. Becker) Val. Tikhom. in Bot. Zhurn. 100: 497. 2015, nom. inval. (Shenzhen Code Art. 41.8; “Boreali-Americanae”) ≡ Viola sect. Borealiamericanae (W. Becker) Espeut in Botanica Pacifica 9(1): 35. 2020 (“Boreali-Americanae”)—Type (only species cited): Viola nuevoleonensis W. Becker
=Viola subg. Hesperion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 211. 1914—Type: Viola palmata L.
Description.—Perennial herbs. Axes not morphologically differentiated; stem a perennial rhizome terminating in an apical rosette. Stipules narrow, free, glandular-lacerate. Laminas in some species lobed or dissected. Calycine appendages various. Petals violet (rarely whitish), lateral and often the spurred petal densely bearded. Style clavate with a pronounced thickened spreading broadly rounded sometimes weakly trilobate dorsolateral rim with sides or lateral lobes continuing to the ventrally oriented rostellum. Cleistogamous flowers produced, seasonal (in temperate species) or simultaneous (in subtropical species). Base chromosome number x = 27.
Diagnostic characters.—Habit strictly rosulate AND stipules free AND petals violet AND lateral (sometimes spurred) densely bearded AND style with pronounced thickened spreading broadly rounded sometimes weakly trilobate dorsolateral rim and ventrally oriented rostellum AND cleistogamy present AND base chromosome number x = 27.
Ploidy and accepted chromosome counts.—10x; 2n = 54.
Included species.—38. Violaaffinis Leconte, V. baxteri House, V. brittoniana Pollard, V. calcicola R. A. McCauley & H. E. Ballard, V. chalcosperma Brainerd, V. communis Pollard, V. cucullata Aiton, V. edulis Spach, V. egglestonii Brainerd, V. emarginata (Nutt.) Leconte, V. fimbriatula Sm., V. floridana Brainerd, V. hirsutula Brainerd, V. impostor R. Burwell & H. E. Ballard, ined. [H. E. Ballard 18-002], V. langloisii Greene, V. latiuscula Greene, V. lovelliana Brainerd, V. missouriensis Greene, V. monacanora J. L. Hastings & H. E. Ballard, ined. [H. E. Ballard 21-015], V. nephrophylla Greene, V. novae-angliae House, V. nuevoleonensis W. Becker, V. palmata L., V. pectinata E. P. Bicknell, V. pedatifida G. Don, V. pedatiloba (Brainerd) Burwell & H. E. Ballard, ined., V. pratincola Greene, V. retusa Greene, V. rosacea Brainerd, V. sagittata Aiton, V. septemloba Leconte, V. septentrionalis Greene, V. sororia Willd., V. stoneana House, V. subsinuata (Greene) Greene, V. tenuisecta Zumwalde & H. E. Ballard, ined. [Ballard 21-017], V. viarum Pollard, V. villosa Walter
Distribution.—North America.
Discussion.—This endemic North American lineage retains the initial allodecaploid genome constitution of the ancestor to sect. Nosphinium. A suite of traits delimits the subsection, including a thickish rhizome, strictly rosulate habit, free stipules, undivided or lobed to dissected leaf laminas, large violet to dark violet, rarely whitish corolla, densely bearded lateral petals and often bearded bottom petal, and a style with a spreading conspicuously thickened dorsolateral rim and distinct rostellum. Species express a wide range of diagnostic features in cleistogamous capsule and seed morphology. The centre of diversity is in the Appalachian Mountain range and adjacent uplands. Ezra Brainerd and others conducted many studies of interspecific hybridisation in the subsection, including long-term garden observations and cultivation of F3 and F4 generations (summarised in Brainerd [264]). Hybridisation is extensive among locally co-occurring species, with hybrids, typically vigorous, failing in chasmogamous reproduction, commonly producing either underdeveloped capsules or capsules with a reduced proportion of viable seeds relative to parental species, and progeny of hybrids express recombinant phenotypic traits of the parental taxa in the plants derived from seeds of the cleistogamous capsules. All species but one occur north of Mexico, whereas V. nuevoleonensis is confined to northeastern Mexico.
Despite gradually increasing synonymy by specialists since Brainerd [69], recent studies by HEB and collaborators are revealing many overlooked new species (including some local and regional endemics) and resurrecting previously synonymised species, making it is one of the more diverse subsectional lineages in the genus, and the second largest in the Western Hemisphere (minimum 38 species, possibly as many as 60). Viola communis Pollard thrives in lawns and fencerows, and a few species have been inadvertently introduced into Europe [73,265,266,267,268].
Viola subsect. Clausenianae H. E. Ballard, subsect. nov.—Type: Viola clauseniana M. S. Baker
Description.—Perennial herbs. Axes not morphologically differentiated; stem a perennial rhizome terminating in an apical rosette. Stipules narrow, adnate in lowest 1/3. Laminas undivided. Calycine appendages short and truncate to rounded. Petals violet, beardless. Style clavate, the apex triangular from above, the pronounced thickened dorsolateral rim protruding apically as a broadly truncate or weakly emarginate margin continuing down to the rostellum, the rostellum oriented apically or incurved. Cleistogamous flowers produced, seasonal.
Diagnostic characters.—Habit strictly rosulate AND stipules basally adnate AND petals violet AND all petals beardless AND style with apically protruding broadly truncate dorsolateral rim and forward-pointing to incurved rostellum AND cleistogamy present.
Ploidy and accepted chromosome counts.—10x; 2n = c. 44 (needs confirmation).
Age.—Crown node not applicable (monotypic subsection), stem node age 5.0–11.5 Ma [45].
Included species.—1. Viola clauseniana M. S. Baker
Distribution.—USA (Utah).
Discussion.—A monotypic subsection for the anomalous Utah endemic Viola clauseniana. Phylogenetic analyses place it firmly among other Nosphinium groups but indicate only ambiguous placement otherwise. Genetically it appears to retain the initial allodecaploid constitution of the ancestor of the section [45], which puts into question the count of n = c. 22 reported by Clausen [59] from the type locality; we would rather expect n = 27 as in the subsections Borealiamericanae and Pedatae. While most similar morphologically to the Borealiamericanae, the absence of petal beards, basally adnate stipules, and style with dorsally protruding and very thickened dorsolateral rim and a forward-pointing to incurved rostellum delimit it uniquely in the section. T.M. has observed unusual morphology in the cleistogamous flowers. The species is known from a single area, Zion National Park, and occurs in isolated “hanging gardens”, seasonally moist to wet cliff ledges.
Viola subsect. Langsdorffianae (W. Becker) W. Becker in Acta Horti Gothob. 2: 286. 1926 ≡ Viola [sect. Nomimium; unranked] Langsdorffianae W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 368. 1925 (excl. V. moupinensis) ≡ Viola sect. Langsdorffianae (W. Becker) Espeut in Botanica Pacifica 9(1): 35. 2020—Type (Shenzhen Code Art. 10.8): Viola langsdorffii Fisch. ex Ging.
=Viola sect. Arction Juz. in Schischk. & Bobrov, Fl. URSS 15: 437, 1949, nom. inval. (Shenzhen Code Art. 39.1, descr. rossica); Viola sect. Arction Juz. ex Zuev in Peschkova, Fl. Sibiri 10: 96. 1996, nom. inval. (Shenzhen Code Art. 40.1, without type)
Description.—Perennial, herbs. Axes morphologically differentiated into a perennial rhizome with or without a terminating apical rosette and lateral, annual floriferous stems. Stipules ovate, free, sheathing the stem, shortly glandular-fimbriate. Laminas undivided. Calycine appendages short and truncate to rounded. Petals violet, lateral bearded. Style cylindrical or slightly clavate with a weak dorsolateral swelling and ventrally oriented rostellum. Cleistogamous flowers produced, seasonal. Allo-14-ploid or allo-18-ploid (10x with additional 4x genomes from sect. Plagiostigma). Secondary base chromosome number x’ = 40.
Diagnostic characters.—Herbaceous AND aerial stems AND stipules ovate, obtuse, shortly glandular-fimbriate and sheathing the stem AND cleistogamy present.
Ploidy and accepted chromosome counts.—14x, 18x, ~22x; 2n = c. 80 (V. howellii), c. 96, 102 (V. langsdorffii), c. 120 (V. “langsdorffii” sensu Taylor & Mulligan [269]).
Age.—Crown node not known, stem node age 1.3–8.8 Ma [45].
Included species.—3. Viola(Langsdorffianae) sp., ined. [J. A. Calder & Roy L. Taylor 36425; or: J. A. Calder, Roy L. Taylor & L. C. Sherk 34963], V. howellii A. Gray, V. langsdorffii Fisch. ex Ging.
Distribution.—Western North America and northeastern Asia.
Discussion.—Subsect. Langsdorffianae is a young high-polyploid lineage that has diversified in response to climate cooling in the Pleistocene [45]. The patterns of variation within subsect. Langsdorffianae are poorly understood, but available information from phylogenetics and chromosome counts indicate that the subsection comprises three ploidy levels, each of which we tentatively refer to as species. Viola howellii (14x) occurs in the North American Pacific Northwest, V. langsdorffii (18x) occurs on both sides of Beringia from California to Japan, and a yet undescribed taxon (20x) occurs at least in the Queen Charlotte Islands of British Columbia [269].
The only phylogenetically investigated species, V. langsdorffii (18x), arose from successive allopolyploidisations involving the allodecaploid ancestor common to all Nosphinium and V. suecica (4x) of the Stolonosae and an unknown member of the Bilobatae (4x) [45]. The expected chromosome number for V. langsdorffii is [2n = 104], and the closest actual count is of 2n = 102 chromosomes in plants from Hokkaido [270]. As pointed out by Marcussen et al. [45], the numerous counts of 2n = (ca.) 96 chromosomes [44,271,272,273,274] “presumably reflect partly the great difficulty in counting many small chromosomes, and partly the wish to align counts with multiples of x = 12, the base number attributed to Langsdorffianae by early authors” [44,59]. Reports of lower chromosome counts for V. langsdorffii (2n = c. 60, c. 64, c. 72) must be rejected on the basis of being incompatible with the phylogenetic history of this species and section [45].
The lower chromosome number of Viola howellii (2n = 14x = 80) suggests that it lacks either the Stolonosae or the Bilobatae genome present in V. langsdorffii. Clausen [59] reported “tetraploid” (n = 20) and “octoploid” (n = 40) counts from Oregon, but whether these refer to the same taxon has not been confirmed; we think the counts of n = 20 may rather refer to the sympatric V. (subsect. Rostratae) aduncoides.
The counts of n = 60 and 2n = c. 120 chromosomes in plants of “V. langsdorffii” from the Queen Charlotte Islands, British Columbia [269], most likely represent the 22x level and a yet undescribed species. Presumably this taxon has acquired yet another 4x genome from sect. Plagiostigma.
Several taxa have been distinguished from Viola langsdorffii in foliage and style traits, including V. superba [275] and the acaulescent V. simulata [276] in western North America and V. kamtschadalorum in eastern Asia [61,277], but no studies have confirmed the distinctness of these taxa.
Viola subsect. Mexicanae (W. Becker) Marcussen & H. E. Ballard, stat. nov. ≡ Basionym: Viola [sect. Nomimium; unranked] Mexicanae W. Becker in Repert. Spec. Nov. Regni Veg. 19: 396. 1923 ≡ Viola sect. Mexicanae (W. Becker) Espeut in Botanica Pacifica 9(1): 35. 2020.—Lectotype (designated here): Viola humilis Kunth
Description.—Perennial herbs. Axes usually morphologically differentiated into a perennial rhizome terminating in an apical rosette and lateral stolons which are often absent. Stipules narrow, free or basally to mostly adnate, glandular-lacerate. Laminas undivided. Calycine appendages mostly short and rounded. Petals violet or whitish, lateral glabrous or sparsely bearded (sometimes densely bearded in V. grahamii, V. nubicola, and V. oxyodontis). Style clavate with a sharp-edged or sometimes weakly thickened apically oriented or slightly incurved dorsolateral rim (somewhat prolonged on the upper side in V. hookeriana) continuing partly or fully to the ventrally oriented rostellum. Cleistogamous flowers produced, simultaneous. Allo-14-ploid (10x with an additional 4x genome from sect. Plagiostigma). Secondary base chromosome number x’ = 40.
Diagnostic characters.—Habit rosulate or stoloniferous AND stipules free or adnate AND petals violet or whitish AND lateral petals glabrous or sparsely (rarely densely) bearded AND style with weakly thickened apically oriented (rarely prolonged) dorsolateral rim and ventrally oriented rostellum AND cleistogamy present.
Ploidy and accepted chromosome counts.—14x; 2n = 80 (Viola nannei).
Included species.—10. Violabeamanii Calderón, V. cuicochensis Hieron., V. grahamii Benth., V. guatemalensis W. Becker, V. hemsleyana Calderón, V. hookeriana Kunth, V. humilis Kunth, V. nannei Pol., V. nubicola H. E. Ballard, ined. [J. H. Beaman 2976], V. oxyodontis H. E. Ballard
Distribution.—Mexico to Ecuador.
Discussion.—This subsection currently comprises 10 species expressing diverse morphologies but which appear to belong to a single lineage (an unpublished ITS phylogeny by HEB including most species was monophyletic with strong support among other lineages of the genus). It arose from a secondary allopolyploidisation event from the ancestor of the Nosphinium lineage and an early sister lineage to the North American sublineage of Stolonosae [45]. Slightly more than half produce above-ground stolons, two non-stoloniferous species often produce adventitious shoots on roots (V. beamanii and V. hookeriana), a few species have white flowers (V. grahamii, V. oxyodontis, and central Mexican populations of V. hookeriana), and most species have lateral petals beardless or with sparse beards. One species has strongly adnate outer stipules (V. humilis) while two others have basally adnate stipules (V. grahamii, V. oxyodontis). The style apex has the thin short dorsolateral rim erect (rather than spreading as in the Borealiamericanae). Most species are restricted to Mexico, a few extend into Central America, and two are found in northern South America.
Viola subsect. Nosphinium (W. Becker) Marcussen & H. E. Ballard, stat. nov. ≡ Basionym: Viola sect. Nosphinium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 374. 1925—Lectotype (Espeut 2020 [61], page 34): Viola chamissoniana Ging.
≡Viola [unranked] (”Gruppe”) Sandvicenses W. Becker in Beih. Bot. Centralbl., Abt. 2, 34: 209. 1917.—Lectotype (designated here): Viola chamissoniana Ging.
Description.—Branching or non-branching shrubs or treelets, rarely perennial herbs (Viola kauaensis). Axes morphologically differentiated into erect stems, rarely rhizomes (V. kauaensis), and lateral floriferous stems or branches (very rarely absent). Leaves of floriferous stems in most species reduced to a pair of stipules, giving the floriferous stem the appearance of a leafless, bracteose, 1–4-flowered inflorescence; rarely floriferous stems with normal-sized leaf laminas (V. chamissoniana and V. kauaensis) or reduced leaf laminas (V. tracheliifolia). Stipules triangular, free, sheathing the stem, glandular-lacerate. Laminas crenulate, undivided. Calycine appendages short and truncate to rounded. Petals on the inside violet or whitish, concolourous and lacking violet striation, lateral sometimes bearded; petals often violet on the back side. Style cylindrical or slightly clavate with a weak dorsolateral swelling and thick blunt or short conic rostellum. Cleistogamous flowers produced in V. kauaensis only. Allo-14-ploid (10x with one additional 4x genome from sect. Plagiostigma). Inferred secondary base chromosome number [x’ = 40].
Diagnostic characters.—Woody (rarely herbaceous) AND aerial stems AND stipules triangular, acute or acuminate, glandular-lacerate and sheathing the stem AND cleistogamy absent (rarely present).
Ploidy and accepted chromosome counts.—14x; 2n = c. 76, c. 85, c. 86.
Included species.—9. Viola chamissoniana Ging., V. helena C. N. Forbes & Lydgate, V. kauaensis A. Gray, V. lanaiensis W. Becker, V. maviensis H. Mann, V. oahuensis C. N. Forbes, V. robusta Hillebr., V. tracheliifolia Ging., V. wailenalenae (Rock) Skottsb.
Distribution.—Hawaiian Islands.
Discussion.—This endemic Hawaiian Island subsection arose from a secondary allopolyploidisation including genomes of the allodecaploid ancestor of the Nosphinium lineage and a Pacific sublineage of allotetraploid subsect. Stolonosae (different from that leading to the Mexicanae) [45]. Subsection Nosphinium is represented by nine species, most of which are woody and produce lateral 1-4-flowered leafless inflorescences. These species have entirely rayless wood, which agrees with the phylogenetic inference that woodiness is secondary [45,81,278]. Viola tracheliifolia, the largest species, is a branched shrub or treelet with lateral inflorescences with reduced (but not absent) leaf laminas. Only V. kauaensis has retained the presumably ancestral, herbaceous habit and lateral floriferous stems with solitary flowers in the axil of normal leaves (i.e., peduncles not clustered together on leafless lateral axes) and is the only species producing cleistogamous flowers. The predominantly woody habit and racemose inflorescence, broad semi-sheathing stipules, style with apex bent into a tall short rostellum, and near-absence of cleistogamy define the subsection. An initial phylogenetic study using ITS [81] indicated V. langsdorffii erroneously as a direct sister taxon to subsect. Nosphinium, but the relationships were later shown to be more complex due to separate allopolyploid origins in the Langsdorffianae and Nosphinium lineages [45].
Ballard et al. [81] indicated that the initial diversification occurred on the oldest island of Kauai, with speciation occurring along ecological gradients, and later dispersal and further speciation to younger islands eastward. Havran et al. [85] reanalysed biogeography of subsect. Nosphinium with more sophisticated models and arrived at a scenario involving initial dispersal to Maui Nui. A reanalysis of the molecular data set by T.M. arrived at the original finding of colonisation beginning on Kauai (Figure 25), as supported by both ancestral state reconstruction and inferred node ages, and subsequent dispersal and diversification proceeding eastward per the Progression Rule, i.e., hypotheses of phylogeographic congruence among codistributed taxa that track the ages of the islands [279]. This scenario receives further support from the facts that Kauai is home to the only species that has retained the ancestral herbaceous morphology (V. kauaensis) and that the average branch length is higher for taxa on Kauai than for taxa on any other Hawaiian island.
Viola subsect. Pedatae (Pollard ex W. Becker) Brizicky ex Marcussen & H. E. Ballard, stat. nov. ≡ Basionym: Viola [unranked] Pedatae Pollard ex W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 369. 1925 ≡ Viola “class” Pedatae Pollard in Bot. Gaz. 26: 237. 1898, nomen inval. (Shenzen Code Art. 37.6) ≡ Viola subsect. Pedatae “(Pollard) Brizicky” in J. Arnold Arb. 42: 327. 1961, nom. inval. (Shenzhen Code Art. 41.5) ≡ Viola sect. Pedatae (Pollard ex W. Becker) Espeut in Botanica Pacifica 9(1): 35. 2020—Type (Shenzhen Code Art. 10.8): Viola pedata L.
≡Oionychion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 210. 1914.—Type: Viola pedata L.
Description.—Perennial herbs. Axes not morphologically differentiated; stem a rhizome terminating in an apical rosette. Rhizome thick, vertical and barrel-like. Stipules narrow, long-adnate to petiole. Laminas deeply pedately divided (rare variations with triternate or merely apically incised laminas). Calycine appendages prominent, truncate or dentate. Petals violet, beardless. Style clavate, apex narrowly rounded from above, with dorsolateral margin as a narrowly rounded rim continuing to the ventral surface, the stigma hidden in the narrow triangular cavity created by the rim. Cleistogamous flowers not produced. Secondary base chromosome number x’ = 27.
Diagnostic characters.—Rosulate acaulescent AND stipules long-adnate AND laminas deeply pedately divided (rarely otherwise) AND petals violet AND all petals beardless AND style with narrowly rounded dorsolateral rim and hidden stigma AND cleistogamous flowers not produced. Allodecaploid. n = 27.
Ploidy and accepted chromosome counts.—10x; 2n = 54.
Age.—Crown node not applicable (monotypic subsection), stem node age 5.0–6.0 Ma [45].
Included species.—1. Viola pedata L.
Distribution.—Eastern North America.
Discussion.—A monotypic subsection for Viola pedata, a widely distributed eastern North American species of dry oak woodlands, oak savannas and dry prairies. The subsection (and species) are unusual in having a short vertical barrel-like rhizome pulled below the soil surface by contractile roots, long-adnate stipules, a clavate or narrowly ellipsoid style lacking a noticeable to prominent thickened dorsolateral rim (this simply a thin non-spreading margin), and absence of cleistogamy. The type variety produces deeply pedately divided laminas with linear segments; populations with narrowly flabellate laminas in the Sandhills region of the southeastern U.S. and populations with triternately divided laminas in the east-central Piedmont, are treated as varieties. This species is unusual also in maintaining a presumably balanced polymorphism in corolla colour pattern, with individuals with dark violet-black upper petals increasingly common further south in the range, and individuals with all petals light violet increasingly common to the north. Finally, V. pedata is the only member of the genus reported to be self-incompatible [280]. Phylogenetic studies involving all North American lineages have shown that, like V. clauseniana, V. pedata does belong to the Nosphinium lineage but has ambiguous relationships among the other species. It has retained the initial allodecaploid constitution of the ancestor of the Nosphinium lineage [45] but has obviously diverged considerably from the other subsections.
Viola sect. Plagiostigma Godr., Fl. Lorraine, ed. 2, 1: 90. 1857 ≡ Viola [unranked] (“Gruppe”) Plagiostigma (Godr.) Kupffer in Oesterr. Bot. Z. 53: 329. 1903 ≡ Viola [sect. Nomimium] subsect. Plagiostigma (Godr.) J. C. Clausen in Ann. Bot. (Oxford) 43: 751. 1929; (Godr.) P. Y. Fu, Fl. Pl. Herb. Chin. Bor.-Or. 6: 89. 1977 (isonym)—Type: Viola palustris L.
Description.—Perennial herbs, very rarely annuals. Axes morphologically differentiated in a rhizome and lateral stems; sometimes only a rhizome present. Rhizome densely or sometimes remotely noded, with an apical leaf rosette. Lateral stems annual aerial stems or stolons. Stipules free or adnate to petiole. Lamina extremely variable, entire or deeply divided, petiolate. Corolla violet, pink or white, with violet striations, and a white or yellow-green throat. Spur short and saccate to very long and slender. Style clavate, at apex flattened above, laterally and distally margined, or bilobate, beardless. Cleistogamous flowers produced; cleistogamy seasonal. Allotetraploid (CHAM + MELVIO). ITS sequence of CHAM type. Secondary base chromosome number x’ = 12.
Diagnostic characters.—Corolla violet, pink or white with a white or yellow-green throat AND style clavate, at tip flattened above, laterally and distally margined, or bilobate AND base chromosome number x’ = 12.
Ploidy and accepted chromosome counts.—4x, 8x, 12x; 2n = 20, 22, 24, 26, 44, 48, 72, 74.
Age.—Crown node age 16.6 (15.4–17.0) Ma [28].
Included species.—142.
Distribution.—Throughout the northern temperate region, with a few species south of the equator in Australasia and South America; centre of diversity in eastern Asia (Figure 26).
Discussion.—Sect. Plagiostigma is phylogenetically an allotetraploid CHAM + MELVIO lineage; unlike all other sections except sect. Danxiaviola it has retained the CHAM homoeolog for ITS (Figure 6). It is karyologically characterised by the secondary base chromosome number x = 12, and morphologically by the clavate, margined or bilobate, beardless style, and the occurrence of seasonal cleistogamy. Here, we recognise a narrowly circumscribed sect. Plagiostigma that comprises the six Beckerian greges [1] having a ‘plagiostigmate’ style shape and a secondary base chromosome number x’ = 12, i.e., sect. Nomimium greges Adnatae p.p., Bilobatae, Diffusae, Serpentes p.p., Stolonosae, and Vaginatae. In this respect our classification approaches Clausen’s [29,59] but we further exclude the North American allodecaploid lineage, herein transferred to sect. Nosphinium [28,45,61].
With its 139 known species and a crown node of 16.6 (15.4–17.0) Ma, sect. Plagiostigma is both the oldest and the most species-rich of all Viola sections. It could be justified to treat subsect. Diffusae and subsect. Patellares as separate sections. We keep them within sect. Plagiostigma because of at least two synapomorphies, the style shape and the base chromosome number x = 12. We recognise seven subsections within sect. Plagiostigma (Figure 27), each monophyletic and morphologically characterised, i.e., subsect. Australasiaticae, subsect. Bilobatae, subsect. Bulbosae, subsect. Diffusae, subsect. Patellares, and subsect. Stolonosae. Diffusae and Patellares are sisters (or sister) to the lineage comprising Bilobatae, Bulbosae, and Stolonosae. The phylogenetic placement of subsect. Australasiaticae within the section is unknown, as this taxon is represented by ITS sequences only and this marker (Figure 6) poorly reflects the genome phylogeny (Figure 27).
While 2n = 24 is retained in most of the subsections, 2n = 22 is apomorphic in subsect. Formosanae and, possibly, 2n = 46 in subsect. Austalasiaticae.
There is little agreement between Becker’s [1] greges and the subsections proposed herein. This is discussed briefly under each subsection.
Key to the subsections of Viola sect. Plagiostigma
- 1a.
- Rhizome annual, vertical, growing from deep-buried bulbils, aerial part more remotely noded. Lateral stolons present, underground, leafless. Outer stipules adnate, inner stipules free. ................................................................................................................................................................................................................................. subsect. Bulbosae
- 1b.
- Rhizome perennial or very rarely plant annual (in V. diffusa), horizontal or vertical, bulbils absent. Lateral stolons absent or when present not underground and leafless. Stipules free or adnate. ..................................................................................................................................................................................................................................................................... 2.
- 2a.
- Rhizomatous herbs lacking lateral stolons and aerial stems but sometimes with adventitious buds on roots. Stipules adnate to petiole in the lower 1/3–1/4. Leaf margin crenulate to deeply divided. Spur 1–10 mm, usually slender and longer than the calycine appendages. Corolla white to deep (bluish or reddish) violet. ...................................................................................................................................................................................................................................................... subsect. Patellares
- 2b.
- Rhizomatous herbs, usually with lateral stolons and/or aerial stems. Stipules free or up to 1/3 adnate to petiole. Leaf margin crenulate or crenate, never deeply divided. Spur usually short and saccate, 1–4 mm, rarely 5–7 mm (in V. formosana). Corolla usually white or pale violet ......................................................................................................................................................................................................................................................... 3.
- 3a.
- Bottom petal longer than the other petals, deeply emarginate or cleft. Spur longer than tall, 1.5–7 mm. .................................................................................................................................................................................................................................................... subsect. Formosanae
- 3b.
- Bottom petal shorter or subequal to the other petals, acute, obtuse or rarely emarginate. Spur as long as tall, 1–4 mm. ................................................................................................................................................................................................................................................................................ 4.
- 4a.
- Lateral stems aerial, decumbent or erect, rarely short or absent (in V. cunninghamii). Stipules foliaceous, free or adnate at base only, dentate or entire. Lamina semilunate to triangular or hastate. Style margined and bilobate at apex. ................................................................................................................................................................................................................................................. subsect. Bilobatae
- 4b.
- Lateral stems stolons, rarely aerial stems or absent. Stipules pale or purple-brown, rarely greenish, free or partially adnate, fimbriate or entire. Lamina reniform to narrowly lanceolate. Style apex bilobate or flattened, distinctly margined. ...................................................................................................................................................................................................................................................................................... 5.
- 5a.
- Sepals usually broadly lanceolate to broadly ovate, rarely lanceolate (but then with long denticulate sepal appendages: V. thomsonii). Sepal appendages up to 2 mm, sometimes denticulate. Bottom petal 7–25 mm, usually not conspicuously shorter than the other petals, sometimes longer, truncate or emarginate, rarely acute, violet-striate. Style apex flattened above and margined, rarely bilobate. Stipules lanceolate to ovate, entire or remotely denticulate to fimbriate-dentate, free or adnate at base only. Corolla commonly white with a yellow-green throat, rarely violet or pink. ............................................................................................................................................................................................................................................... subsect. Stolonosae
- 5b.
- Sepals linear-lanceolate to lanceolate, rarely ovate-lanceolate (in V. (Diffusae) guangzhouensis), with short appendages, 0.4–1 mm, rounded or slightly denticulate (absent in V. kwangtungensis). Bottom petal 5–12 mm, shorter and narrower than the others, usually acute, with conspicuous violet striation or reticulation. Style apex bilobate. Stipules linear to broadly lanceolate, densely or remotely fimbriate, free or adnate in the lower 1/3. Corolla pale pink or pale violet, rarely white. ....................................................................................................................................................................................................................................................... 6
- 6a.
- Lateral petals not bearded. Peduncles glabrous; plant usually glabrous or nearly so. Rhizome long and remotely noded or short and densely noded. Stolons present or rarely absent, with (many) scattered leaves. Stipules free or adnate at base only, often brownish, long-fimbriate to laciniate. Corolla usually pale violet to whitish, without a greenish throat. Lamina margin crenate, occasionally with conspicuous mucronules. Perennials. ....................................................................................... ......................................................................................................................................................... subsect. Australasiaticae
- 6b.
- Lateral petals usually bearded. Peduncles with patent hairs, rarely glabrous (in V. nanlingensis); plant usually hairy. Rhizome short, densely noded. Stolons with 1–2 (smaller) leaves and a leaf rosette at apex. Stipules adnate in the lower 1/3 (stipules on aerial stems free in V. guangzhouensis), remotely or rarely densely fimbriate. Corolla usually pale pink to pale violet, with a greenish throat. Lamina margin crenulate, never with mucronules. Perennials or rarely annuals (V. diffusa). ................................................................................................................................................................................................................................................... subsect. Diffusae
Viola subsect. Australasiaticae (M. Okamoto) Marcussen, comb. et stat. nov.—Basionym: Viola ser. Australasiaticae M. Okamoto in Taxon 42(4): 784. 1993.—Type: Viola sumatrana Miq.
Description.—Rhizome perennial; bulbils absent. Lateral stems usually present: aboveground stolons, most leaves scattered. Stipules free or adnate at base only, brown, linear-lanceolate to broadly lanceolate, long-fimbriate. Lamina triangular-ovate to reniform, base cuneate to deeply cordate, apex obtuse to acuminate, margin crenate or mucronulate. Corolla white or pale violet. Sepals linear-lanceolate to lanceolate; appendages short or absent (0–1.4 mm), rounded or slightly denticulate. Lateral petals not bearded; bottom petal shorter than the other petals (5–12 mm), acute to obtuse; spur short (1–2.5 mm) and saccate. Style at apex margined and bilobate.
Diagnostic characters.—Plants usually stoloniferous AND stolons with most leaves scattered AND sepals linear-lanceolate to lanceolate AND lateral petals glabrous AND bottom petal shorter than the others AND style margined and bilobate at apex.
Ploidy and accepted chromosome counts.—4x? 8x; 2n = 46.
Age.—Crown node age c. 12.0 Ma; stem node c. 16.3 Ma (Figure 6).
Included species.—10. Violaannamensis Baker f., V. austrosinensis Y. S. Chen & Q. E. Yang, V. balansae Gagnep., V. duclouxii W. Becker, V. hossei W. Becker, V. kwangtungensis Melch., V. mucronulifera Hand.-Mazz., V. shiweii Xiao Chen Li & Z. W. Wang, V. sikkimensis W. Becker, V. sumatrana Miq.
Distribution.—Southeastern Asia and Malesia.
Discussion.—Becker [1] erected (sect. Nomimium) grex Serpentes as a catch-all taxon for stoloniferous species from subtropical Asia. This group was highly heterogeneous and the constituent species have later been redistributed among sect. Viola subsects. Rostratae and Viola, and sect. Plagiostigma subsects. Australasiaticae, Diffusae, Patellares, and Stolonosae [86,126,229,231]. Wang [76] expanded Becker’s greges, as sect. Serpentes, to include numerous Stolonosae species. Okamoto et al. [229] showed that the type species of grex Serpentes (V. serpens Blume, a synonym of V. pilosa) belongs in subsect. Viola and they therefore designated ser. Australasiaticae (type: V. sumatrana) as a replacement name for the remaining species not belonging in subsect. Viola. However, also Okamoto’s [229] Australasiaticae proved heterogeneous and including taxa from different sections and subsections. The type, V. sumatrana, was however not analysed phylogenetically before the recent study by C. Li et al. [231] which clearly identified the Australasiaticae in the strict sense as a separate lineage within sect. Plagiostigma (Figure 6). We here define subsect. Australasiaticae narrowly as comprising all known Plagiostigma species having stolons with scattered leaves, linear-lanceolate or lanceolate sepals, unbearded lateral petals, and a bilobate style.
The only chromosome counts for subsect. Australasiaticae are of 2n = 46 in V. sumatrana and V. annamensis (as V. rheophila Okamoto) and were reported without metadata by H. Okada in Okamoto et al. [229] and are therefore in need of confirmation. If proved correct, they presumably reflect the 8x level and present a unique number in the genus and a possible apomorphy for subsect. Australasiaticae. It is not known whether this chromosome number and ploidy are shared by all the members of the subsection.
Spinulose or mucronulate leaf margins (as an adaptation to guttation?) occur only in this subsection within Viola but have apparently originated twice. In Viola balansae and V. kwangtungensis the mucronules are extensions of the apex of each leaf tooth and are in the plane of the leaf. In V. mucronulifera the mucronules are adaxial extensions of the invagination between leaf teeth and are perpendicular to the plane of the leaf [231].
Viola subsect. Bilobatae (W. Becker) W. Becker in Acta Horti Gothob. 2: 288. 1926 ≡ Viola [sect. Nomimium; unranked] (“Gruppe”) Bilobatae W. Becker in Beih. Bot. Centralbl., Abt. 2, 34: 226. 1917 ≡ Viola ser. Bilobatae (W. Becker) Steenis in Bull. Jard. Bot. Buitenzorg, ser. 3, 13 (1933–1936): 260. 1934 ≡ Viola sect. Bilobatae (W. Becker) Juz. in Schischk. & Bobrov, Fl. URSS 15: 439. 1949—Lectotype (Espeut 2020 [61], page 33): Viola arcuata Blume
Description.—Rhizome perennial; bulbils absent. Lateral stems present or rarely absent: aerial stems, decumbent or erect, leaves scattered. Stipules free or adnate at base only, green and foliaceous, up to 40 mm, linear-lanceolate to ovate, obtuse to acuminate, entire, remotely denticulate or lobed. Lamina ovate-triangular to narrowly triangular or nearly hastate, base truncate to broadly cordate, often with a lunate sinus, apex more or less acute, margin crenulate. Corolla white. Sepals linear to ovate-lanceolate; appendages short (c. 0.5 mm), rounded or slightly denticulate. Lateral petals bearded or not; bottom petal shorter than the other petals (6–8 mm), apex rounded; spur short (1–2 mm) and saccate. Style at apex margined and bilobate.
Diagnostic characters.—Stipules foliaceous AND style margined and distinctly bilobate at apex.
Ploidy and accepted chromosome counts.—4x, 8x; 2n = 24, 44, 48.
Included species.—8. Violaamurica W. Becker, V. caleyana G. Don, V. cunninghamii Hook. f., V. hamiltoniana D. Don, V. lyallii Hook. f., V. merrilliana W. Becker, V. raddeana Regel, V. triangulifolia W. Becker
Distribution.—Eastern Asia, Malesia, Australia, New Zealand.
Discussion.—The overall morphology of sect. Plagiostigma subsect. Bilobatae is superficially similar to that of the unrelated sect. Viola subsect. Rostratae, and conspicuously so in species such as V. raddeana (Bilobatae) and V. stagnina (Rostratae), which both are adapted to floodplain habitats. Reported chromosome counts of 2n = 20 in subsect. Bilobatae (cf. [61]) are likely errors.
Viola subsect. Bulbosae Marcussen, subsect. nov.—Type: Viola bulbosa Maxim.
Description.—Rhizome annual, growing from underground bulbil. Lateral stems present: underground stolons, usually leafless but with scattered nodes. Stipules outer stipules adnate, inner stipules free, pale, linear-lanceolate, remotely fimbriate. Lamina oblong-ovate, suborbicular or reniform, base cuneate or narrowly cordate, apex rounded or acute, margin crenulate. Corolla white. Sepals lanceolate to broadly lanceolate; appendages short (c. 0.8 mm), rounded. Lateral petals bearded or not; bottom petal shorter than the other petals (7–8 mm), apex rounded; spur short (1.2–1.7 mm) and saccate. Style at apex margined and bilobate.
Diagnostic characters.—Rhizome vertical, growing from deep-buried bulbils.
Ploidy and accepted chromosome counts.—4x; 2n = 24.
Age.—Crown node age unknown, stem node age 13.5 (12.2–14.0) Ma [28].
Included species.—2. Viola bulbosa Maxim., V. tuberifera Franch.
Distribution.—Eastern Himalaya and central China.
Discussion.—Section Plagiostigma subsect. Bulbosae comprises two species, Viola bulbosa and V. tuberifera [77,281]. The species are characterised by having small underground bulbs, a unique feature in Viola. The bulb is composed of 4–8 fleshy petiole bases along a condensed axial portion which apically elongates into the annual aerial stem and laterally produces underground, leafless stolons with cleistogamous flowers. The species were included in subsect. Patellares by both Becker [1] and Wang [76], as grex Adnatae and sect. Adnatae, respectively.
Viola subsect. Diffusae (W. Becker) Chang in Bull. Fan Mem. Inst. Biol., ser. n., 1(3): 249, 1949 [non vidimus] ≡ Viola [unranked] (“Gruppe”) Diffusae W. Becker in Beih. Bot. Centralbl., Abt. 2, 40: 113. 1924 ≡ Viola (sect. Nomimium) ser. Diffusae (W. Becker) Steenis in Bull. Jard. Bot. Buitenzorg, ser. 3, 13 (1933–1936): 260. 1934 ≡ Viola sect. Diffusae (W. Becker) Ching J.Wang, Fl. Reipubl. Popularis Sin. 51: 100. 1991.—Type (Shenzhen Code Art. 10.8): Viola diffusa Ging.
Description.—Rhizome perennial or rarely plant annual; bulbils absent. Lateral stems present: aboveground stolons, most leaves in apical rosette; rarely also aerial stems with scattered leaves. Stipules usually adnate in the lower ⅓, pale, greenish, or brown, subulate to lanceolate, acuminate, remotely long-fimbriate. Lamina ovate, ovate-oblong or elliptic, base cuneate to shallow-cordate, often decurrent, apex usually obtuse, margin crenate. Corolla usually pale pink or pale violet, with a greenish throat. Sepals linear to ovate-lanceolate; appendages short (0.3–0.8 mm), rounded or slightly denticulate. Lateral petals bearded or not; bottom petal shorter than the other petals (5–12 mm), apex acute; spur short (1–2.5 mm) and saccate. Style at apex margined and bilobate.
Diagnostic characters.—Stolons long with 1–2 leaves and a leaf rosette at apex AND stipules 1/3 adnate to petiole AND corolla mostly pale pink to pale violet, with a greenish throat AND style margined and bilobate at apex.
Ploidy and accepted chromosome counts.—4x, 8x, 12x; 2n = 24, 26, 48, 74.
Included species.—17. Viola(Diffusae) sp. 1, ined., V. (Diffusae) sp. 2, ined., V. (Diffusae) sp. 3, ined., V. amamiana Hatus., V. apoensis Elmer, V. changii J. S. Zhou & F. W. Xing, V. diffusa Ging., V. guangzhouensis A. Q. Dong, J. S. Zhou & F. W. Xing, V. huizhouensis Y. S. Huang & Q. Fan, V. jinggangshanensis Z. L. Ning & J. P. Liao, V. lucens W. Becker, V. nagasawae Makino & Hayata, V. nanlingensis J. S. Zhou & F. W. Xing, V. pricei W. Becker, V. tenuis Benth., V. wilsonii W. Becker, V. yunnanensis W. Becker & H. Boissieu
Distribution.—Southeastern Asia.
Discussion.—Section Plagiostigma subsect. Diffusae comprises a handful of southeast Asian species, characterisable by stolons with few internodes and a terminal leaf rosette, stipules adnate to the petiole in the lower third, and more or less lanceolate laminas with a narrow and shallow sinus. Some species have aerial stems. Most species are distinctly stiffly hairy and have pale violet or pink petals, often yellowish green at the base, with a short and narrow, pointed bottom petal and a very short spur. This subsection, although easily recognisable in most cases, is poorly understood owing to taxonomic confusion with the other stolonose subsections Australasiaticae and Stolonosae.
More than half of the species placed in subsect. Diffusae are narrow endemics native to southern China that have been discovered and described within the last 15 years [127,128,282,283,284]. These species grow on rock surfaces, often in inaccessible places, and numerous species are still undescribed in addition to the 17 cited here (Y.-S. Huang, pers. comm. [285]).
Viola subsect. Formosanae (J.-C. Wang & T.-C. Huang) Marcussen, comb. et stat. nov.—Basionym: Viola grex Formosanae J.-C. Wang & T.-C. Huang in Taiwania 35(1): 14. 1990.—Type (only species listed): Viola formosana Hayata
Description.—Rhizome perennial; bulbils absent. Lateral stems present: aboveground stolons, most leaves in apical rosette. Stipules free or adnate at base, purplish-brown, lanceolate or narrowly ovate, long fimbriate-laciniate. Lamina broadly triangular-ovate or oblong-orbicular, base deeply cordate to rounded, apex acute to rounded or obtuse, margin crenate. Corolla white or pale violet. Sepals narrowly lanceolate to oblong; appendages short (0.5–1 mm), rounded. Lateral petals not bearded; bottom petal longer than the other petals (8–15 mm), apex deeply emarginate or shallowly cleft; spur long and slender (1.5–7 mm). Style at apex margined and flattened, not bilobate. Secondary base chromosome number x’ = 11.
Diagnostic characters.—Bottom petal longer than the other petals AND stolons with most leaves in apical rosette AND chromosome number 2n = 22.
Ploidy and accepted chromosome counts.—4x; 2n = 22.
Age.—unknown.
Included species.—2. Viola formosana Hayata, V. stoloniflora Yokota & Higa
Distribution.—Southeastern Asia: the islands of Taiwan (V. formosana) and Okinawa (V. stoloniflora).
Discussion.—Becker was familiar with Viola formosana ([286], page 167), the only of the two species known at the time, but he did not mention it or place it systematically in his revision of the genus [1]. The second species, V. stoloniflora, has been placed in subsect. Australasiaticae [229] or in its predecessor, subsect. Serpentes, “on account of its procumbent stolons, almost free fimbriate stipules, and deplanate obtriangular-dilatate styles” [97]. In their revision of the violets of Taiwan, Wang & Huang (1990 [75]) recognised the distinctness of V. formosana and placed it in a provisional group of its own, Formosanae, one of eight unranked greges; their delimitation of greges is reconcilable with our classification.
The phylogenetic placement of subsect. Formosanae is unresolved, but published chloroplast DNA sequences of Viola formosana place it among the other stoloniferous subsections [287].
The two species Viola formosana and V. stoloniflora have never been grouped together, despite their close geographical proximity and several synapomorphies that set them apart from all other subsections of sect. Plagiostigma, including the long and emarginate bottom petal, the shape of the stolons (reminescent of subsect. Diffusae), and the rare chromosome number 2n = 22 [75,97].
Viola stoloniflora is extinct in the wild; its only known locality in Okinawa Island was destroyed by the construction of the Benoki Dam, which was completed in 1987 [97].
Viola subsect. Patellares (Boiss.) Rouy & Foucaud, Fl. France [Rouy & Foucaud] 3: 35. 1896 ≡ Viola [sect. Nomimium; unranked] §.3. Patellares Boiss., Fl. Orient. 1: 451. 1867, p.p. (excl. Viola uliginosa).—Lectotype (designated here): Viola kamtschatica Ging. (=V. selkirkii Pursh ex Goldie)
=Viola [sect. Nomimium; unranked] b. Patellariae Nyman, Consp. Fl. Eur. 1: 79. 1878, p.p.—Lectotype (designated here): Viola umbrosa Fr. (=Viola selkirkii Pursh ex Goldie)
=Viola subg. Violidium K. Koch in Linnaea 15: 251. 1841. ≡ Viola sect. Violidium (K. Koch) Juz. in Schischk. & Bobrov, Flora URSS 15: 408. 1949 ≡ Viola subsect. Violidium (K. Koch) P. Y. Fu in Fl. Pl. Herb. Chin. Bor.-Or. 6: 93. 1977.—Type: Viola somchetica K. Koch
=Viola [unranked] (“Gruppe”) Estolonosae Kupffer in Oesterr. Bot. Z. 53: 329. 1903 ≡ Viola subsect. Estolonosae (Kupffer) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 217. 1909 ≡ Viola sect. Estolonosae (Kupffer) Vl. V. Nikitin in Bot. Zhurn. (Moscow & Leningrad) 83(3): 132. 1998.—Lectotype (Nikitin 1998 [72], page 133): Viola purpurea Stev. (=V. somchetica K. Koch)
=Viola [sect. Nomimium; unranked] Adnatae W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 368. 1925 ≡ Viola subsect. Adnatae (W. Becker) W. Becker in Acta Horti Gothob. 2: 285. 1926 ≡ Viola ser. Adnatae (W. Becker) Steenis in Bull. Jard. Bot. Buitenzorg, ser. 3, 13 (1933–1936): 258. 1934 ≡ Viola sect. Adnatae (W. Becker) Ching J. Wang, Fl. Reipubl. Popularis Sin. 51: 41. 1991; (W. Becker) Vl. V. Nikitin in Bot. Zhurn. (Moscow & Leningrad) 83(3): 132. 1998 (isonym); (W. Becker) Vl. V. Nikitin in Novosti Sist. Vyssh. Rast. 31: 222. 1998 (isonym).—Lectotype (Nikitin 1998 [72], page 132): Viola selkirkii Pursh ex Goldie
=Viola [unranked] “Gruppe” Pinnatae W. Becker, Beih. Bot. Centralbl., Abt. 2. 40(2): 119. 1924 ≡ Viola sect. Pinnatae (W. Becker) Ching J. Wang, Fl. Reipubl. Popularis Sin. 51: 76. 1991 ≡ Viola subsect. Pinnatae (W. Becker) Vl. V. Nikitin, Novosti Sist. Vyssh. Rast. 34: 125. 2002.—Type (Shenzhen Code Art. 10.8): Viola pinnata L.
=Viola sect. Brachycerae Espeut in Botanica Pacifica 9(1): 32. 2020.—Type: Viola brachyceras Turcz.
Description.—Rhizome perennial; bulbils absent. Lateral stems absent. Stipules adnate in the lower 1/3 to 3/4, pale, greenish, or purple-brown, linear to ovate-lanceolate, acute or acuminate, entire or remotely denticulate-fimbriate. Lamina lanceolate to orbicular or triangular, sometimes 3–5-sect, base cuneate to deeply cordate, sometimes decurrent, apex obtuse to acuminate, margin subentire, crenulate, dentate, or deeply incised. Corolla white to deep violet. Sepals lanceolate to ovate; appendages short to very long (0.4–6 mm), rounded to 2–3-dentate. Lateral petals usually bearded; bottom petal usually longer than the other petals ((5–)10–23(–25) mm), apex rounded to emarginate; spur long (3–10 mm) and slender, rarely short (1–2 mm) and saccate. Style at apex margined and flattened, not bilobate.
Diagnostic characters.—All stems rhizomatous AND stipules 1/3 adnate to petiole AND spur slender, up to 10 mm AND cleistogamous flowers produced.
Ploidy and accepted chromosome counts.—4x, 8x, 12x; 2n = 22, 24, 48, 72.
Included species.—62. Violaalaica Vved., V. albida Palib., V. alexandrowiana (W. Becker) Juz., V. alexejana Kamelin & Junussov, V. bambusetorum Hand.-Mazz., V. baoshanensis W. S. Shu, W. Liu & C. Y. Lan, V. belophylla Boissieu, V. betonicifolia Sm., V. bhutanica H. Hara, V. boissieuana Makino, V. breviflora Jungsim Lee & M. Kim, V. cuspidifolia W. Becker, V. dactyloides Schult., V. forrestiana W. Becker, V. gmeliniana Schult., V. hancockii W. Becker, V. hirtipes S. Moore, V. inconspicua Blume, V. ingolensis Elisafenko, V. iwagawae Makino, V. japonica Langsd. ex Ging., V. jooi Janka, V. keiskei Miq., V. lactiflora Nakai, V. macroceras Bunge, V. magnifica C. J. Wang & X. D. Wang, V. mandshurica W. Becker, V. maximowicziana Makino, V. miaolingensis Y. S. Chen, V. microcentra W. Becker, V. mongolica Franch., V. multifida Willd. ex Schult., V. nujiangensis Y. S. Chen & X. H. Jin, V. pacifica Juz., V. patrinii Ging., V. pekinensis (Regel) W. Becker, V. perpusilla Boissieu, V. phalacrocarpa Maxim., V. philippica Cav., V. pinnata L., V. prionantha Bunge, V. rupicola Elmer, V. selkirkii Pursh ex Goldie, V. senzanensis Hayata, V. seoulensis Nakai, V. sieboldii Maxim., V. somchetica K. Koch, V. sphaerocarpa W. Becker, V. tashiroi Makino, V. tenuicornis W. Becker, V. tienschiensis W. Becker, V. tokaiensis Sugim., nom. nud., V. tokubuchiana Makino, V. trichopetala C. C. Chang, V. turkestanica Regel & Schmalh., V. ulleungdoensis M. Kim & J. Lee, V. umphangensis S. Nansai, Srisanga & Suwanph., V. variegata Fisch. ex Link, V. violacea Makino, V. yezoensis Maxim., V. yunnanfuensis W. Becker, V. yuzufeliensis A. P. Khokhr.
Distribution.—North-temperate, with a diversity centre in northeastern Asia; only four species in Europe and one in North America, the scattered circumboreal V. selkirkii.
Discussion.—Section Plagiostigma subsect. Patellares is species-rich and easily characterised by the absence of stolons, and stipules adnate to the petiole in the lower third. The corolla can be of a deep lilac tone, sometimes fragrant but with a fragrance somewhat different from that of sect. Viola (e.g., V. odorata), and the spur of the bottom petal is usually relatively longer than in the other subsections of Plagiostigma. The lamina shape is extremely variable, from spathulate to cordate in outline, and with margins subentire to crenate or variously deeply divided. Some species form adventitious shoots from roots and have the ability to regenerate from cut roots (e.g., V. prionantha). Many species of the subsection have seeds that germinate directly without stratification.
Phylogenetic relationships within subsect. Patellares are contradictory. There is poor correspondence in patterns obtained from ITS sequences, cpDNA sequences, and morphology [77], but also among studies [82,86,87,89]. This may on one side indicate the presence of real genealogical conflicts resulting from incomplete lineage sorting, allopolyploidisation, and chloroplast introgression, but also taxonomic confusion and misidentifications.
Nested within subsect. Patellares is a pair of dwarf species from the Ryukyus Archipelago (Japan) with dwarf habit and 2n = 22, Viola tashiroi and V. iwagawae. These species form adventitious shoots from roots that superficially look like stolons [288].
Becker [248] erected grex Gmelinianae for a heterogeneous group of Central Asian rosette plants with cuneate or spathulate leaves and adnate stipules, which he later incorporated in grex Adnatae [1]. The Gmelinianae is, however, polyphyletic and here we redistribute its members among three sections: sect. Plagiostigma subsect. Patellares with V. gmeliniana, V. perpusilla, and V. turkestanica; sect. Spathulidium with V. spathulata; and sect. Himalayum with V. kunawurensis. The group consisting of V. perpusilla, V. turkestanica, and the similar V. alata, are atypical within subsect. Patellares in having subentire leaves and unmargined style; they however have the characteristic long spurs of that subsection while sect. Himalayum has a short spur.
Viola subsect. Stolonosae (Kupffer) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 217. 1909 ≡ Viola [unranked; “Gruppe”] Stolonosae Kupffer in Oesterr. Bot. Z. 53: 329. 1903.—Lectotype (designated here): Viola palustris L.
=Viola subg. Verbasculum Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 213. 1914.—Type: Viola primulifolia L.
=Viola [unranked] (”Gruppe”) Vaginatae W. Becker in Beih. Bot. Centralbl., Abt. 2, 36: 29. 1918 ≡ Viola ser. Vaginatae Taken in J. Sci. N.-E. Norm. Univ., Biol. 1: 86. 1955 ≡ Viola subsect. Vaginatae (W. Becker) P.Y.Fu, Fl. Pl. Herb. Chin. Bor.-Or. 6: 91. 1977 ≡ Viola sect. Vaginatae (W. Becker) Ching J.Wang, Fl. Reipubl. Popularis Sin. 51: 85. 1991.—Type (Shenzhen Code Art. 10.8): Viola vaginata Maxim.
Description.—Rhizome perennial; bulbils absent. Lateral stems present or absent: aboveground stolons, most leaves scattered; or rarely aerial stems with leaves in apical rosette (in V. moupinensis). Stipules free or occasionally up to 1/2 adnate (in V. brachyceras), pale, greenish, or brown, (linear-lanceolate to) lanceolate to ovate, acuminate, entire or remotely denticulate-fimbriate. Lamina lanceolate to reniform, base cuneate to deeply cordate, apex rounded to acuminate, margin subentire to crenate. Corolla white or pale violet. Sepals lanceolate to ovate; appendages short or long (0.5–2 mm), rounded or dentate. Lateral petals bearded or not; bottom petal shorter than, or subequal to, the other petals (6–20 mm), apex acute to emarginate; spur short (1–5 mm) and saccate. Style at apex margined and flattened, rarely bilobate.
Diagnostic characters.—Stolons (if present) with most leaves scattered AND sepals lanceolate to ovate AND stipules usually lanceolate to ovate AND style apex margined and flattened, rarely bilobate.
Ploidy and accepted chromosome counts.—4x, 8x; 2n = 20, 24, 44, 48.
Included species.—41. Violaadenothrix Hayata, V. binayensis Okamoto & K. Ueda, V. bissetii Maxim., V. blanda Willd., V. brachyceras Turcz., V. brevipes (M. S. Baker) Marcussen, ined., V. cochranei H. E. Ballard, V. davidii Franch., V. diamantiaca Nakai, V. epipsila Ledeb., V. fargesii H. Boissieu, V. glaucescens Oudem., V. grandisepala W. Becker, V. hultenii W. Becker, V. incognita Brainerd, V. jalapaensis W. Becker, V. javanica W. Becker, V. kjellbergii Melch., V. lanceolata L., V. macloskeyi F. E. Lloyd, V. maoershanensis Y. S. Chen & Q. E. Yang, V. mearnsii Merr., V. minuscula Greene, V. moupinensis Franch., V. nitida Y. S. Chen & Q. E. Yang, V. nuda W. Becker, V. occidentalis (A. Gray) Howell, V. palustris L., V. petelotii W. Becker ex Gagnep., V. pluviae Marcussen, H. E. Ballard & Blaxland, V. primulifolia L., V. principis Boissieu, V. renifolia A. Gray, V. rossii Hemsl., V. shikokiana Makino, V. striatella H. Boissieu, V. suecica Fr., V. thomsonii Oudem., V. vaginata Maxim., V. vittata Greene, V. yazawana Makino
Distribution.—North-temperate; one species (Viola lanceolata) in northern South America. Viola suecica (=V. achyrophora Greene, V. epipsiloides Á. Löve & D. Löve, V. epipsila subsp. repens W. Becker) is circumboreal.
Discussion.—The delimitation of this subsection is “locked” by the existence of allopolyploids between distantly related internal lineages, one of which happens to be the type of the subsection (Viola palustris). The polyploids include the North American V. blanda and V. incognita (8x) which are allopolyploids of V. renifolia or perhaps more likely V. brachyceras (4x) and a taxon within the V. primulifolia group (4x); the Amphiatlantic V. palustris (8x) which is the alloploid of V. minuscula (=V. pallens auct., non (Banks) Brainerd; 4x) and V. epipsila (4x); the Pacific American V. pluviae (8x) which is the alloploid of V. macloskeyi/occidentalis (4x) and V. suecica (4x); and presumably also the North American V. brevipes [45,93]. These five allo-octoploids are no older than 2.5–5 Ma, and their marked boreal distributions suggest they originated in response to the climate cooling and repeated glaciations in the Pleistocene [93].
Disregarding allopolyploidy, at least four informal species groups are nevertheless recognisable at the 4x level based on published phylogenetic studies (Figure 8; [45,82,86,87,287]). These include (1) a clade comprising the Chinese species V. davidii and V. grandisepala; (2) a clade of mostly hairy species occurring in eastern Asia and northern North America comprising V. principis, V. renifolia, V. yazawana, and presumably also V. adenothrix and V. brachyceras; (3) a clade of mostly large species with acuminate laminas and larger pale violet to pink corollas and broad somewhat sheathing denticulate stipules comprising the circumboreal V. epipsila-suecica complex, V. moupinensis, and most of Becker’s [1] grex Vaginatae, i.e., V. bissetii, V. diamantiaca, V. vaginata, etc.; and, finally, (4) the North American stoloniferous species comprising V. primulifolia, V. lanceolata, V. macloskeyi, V. minuscula, etc., by Marcussen et al. [45] referred to as “grex Primulifoliae”.
The group of species having a creeping, remotely noded rhizome and which was previously informally designated as the Palustres grex comprises a subset of the species in clade 3, i.e., V. epipsila and V. suecica, and their allopolyploids, i.e., V. palustris, V. pluviae, and V. brevipes, formed with species in clade 4.
Phylogenetic studies of the north-temperate species of subsect. Stolonosae [45,93] indicate that a narrow species concept coinciding with morphological-geographic units best applies to these taxa. This concept challenges in particular the traditional classification of the North American taxa into a few, broadly defined species based on lamina shape [289,290,291]. Hence, we consider V. minuscula distinct from V. macloskeyi, V. occidentalis distinct from V. primulifolia, V. vittata distinct from V. lanceolata, and V. suecica distinct from V. epipsila. Among the octoploids V. brevipes and V. pluviae are distinct from V. palustris, and V. incognita is distinct from V. blanda. Taxonomy, variation, and phylogeography in this circumboreal complex are poorly understood and require further study throughout its range. Certain characters traditionally considered diagnostic, such as leaf shape and pubescence in V. palustris, have proven variable even within single specimens [292,293].
The chromosome number 2n = 20, apparently at odds with the predominance of 2n = 24 in this subsection, has been reported several times in Viola brachyceras and also in the closely related V. yazawana, for which also 2n = 40 has been reported (cf. [61] and references therein); this number could also explain 2n = 44 (not 48) in the octoploids V. blanda and V. incognita, and possibly also in V. maoershanensis [294]. Counts of 2n = 20 outside of this species group within subsect. Stolonosae are probably errors.
Viola sect. Rubellium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 374. 1925.—Type (Shenzhen Code Art. 10.8): Viola rubella Cav.
=Viola [unranked] § II. Tri(-Pluri-)Caules Reiche in Fl. Chile [Reiche] 1: 140. 1896, nom. inval. (Shenzhen Code Art. 21.2)
Description.—Perennial subshrubs. Axes morphologically (weakly) differentiated in a perennial monopodial aerial stem and lateral monopodial aerial elongated stems bearing flowers; lateral stems with distichous phyllotaxy in Viola portalesia. Stipules small, bract-shaped, fimbriate. Lamina oblong to lanceolate, base cuneate, margin crenate, short-petiolate. Peduncle long. Corolla violet to whitish inward with a greenish throat, or magenta to pink throughout (V. rubella). Spur short. Style clavate, at apex neither margined nor bearded, bent into a simple, ventrad rostellum, or apex rounded with the rostellum on the ventral surface. Cleistogamous flowers not produced. Diploid. Base chromosome number x = 6.
Diagnostic characters.—Subshrubs AND corolla magenta or violet AND style apex strongly bent ventrad or with stigma on ventral side AND diploid with 2n = 12.
Ploidy and accepted chromosome counts.—2x; 2n = 12 (V. rubella).
Age.—Crown node age 1.6 (0.4–2.2) Ma; stem node 26.5 (25.7–26.8) Ma [28].
Included species.—3. Violacapillaris Pers., V. portalesia Gay, V. rubella Cav.
Distribution.—Central Chile (Figure 28).
Discussion.—Section Rubellium is phylogenetically isolated and the only subshrubby diploid lineage within subg. Viola [60]. The original delimitation was established by Becker (1925). Previously, Reiche [122,130] circumscribed the group under an invalid taxonomic rank (i.e., the unranked Tri(-Pluri-)Caules within the invalid Division Sparsifoliae). Sparre [63] included in sect. Rubellium also the herbaceous V. huidobrii, by us reclassified in sect. Viola subsect. Rostratae.
Viola sect. Sclerosium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 374. 1925.—Lectotype (designated here): Viola cinerea Boiss.
=Viola [sect. Nomimium; unranked] §.2. Cinereae Boiss., Fl. Orient. 1: 451. 1867, p. p. (excl. V. spathulata) ≡ Viola [unranked] (”Gruppe”) Cinereae Boiss. em. W. Becker in Beih. Bot. Centralbl., Abt. 2, 36: 36. 1918
Description.—Annual herbs or perennial subshrubs, glabrous or densely short-pubescent. Axes morphologically differentiated in aerial stems and short axillary branches bearing cleistogamous flowers. Stipules small, lanceolate. Lamina ovate to lanceolate, remotely denticulate, petiolate. Corolla pink with a green throat. Spur short and thick. Style slender and cylindrical or slightly clavate, crested; crest a pair of apical or subapical lateral ear-like processes. Simultaneous production of chasmogamous in upper leaf axils and cleistogamous flowers on short branches in lower leaf axils. Allotetraploid (CHAM + MELVIO). Secondary base chromosome number x’ = 11.
Diagnostic characters.—Style with a pair of apical or subapical lateral ear-like processes. Base chromosome number x = 11.
Ploidy and accepted chromosome counts.—4x, 8x; 2n = 22 (V. stocksii).
Age.—Crown node 3.5–10 Ma [150].
Included species.—7. Violabehboudiana Rech. f. & Esfand., V. cinerea Boiss., V. erythraea (Fiori) Chiov., V. etbaica Schweinf., V. kouliana Bhellum & Magotra, V. somalensis Engl., V. stocksii Boiss.
Distribution.—Northeastern Africa to southwestern Asia (Figure 29). Disjunctly distributed in the monsoon region on both sides of the Red Sea, Sokotra and the Arabic coast of the Indian Ocean, southern Iran, most of Pakistan, and northwestern India.
Discussion.—Variation patterns within sect. Sclerosium are poorly understood. It contains closely related races that are difficult to delimit but differ in distribution, life history traits (annual or perennial), pubescence, and style shape. Nine allopatric taxa have been described [1,150,295,296] but most authorities have interpreted the variation as more or less continuous and have retained only one or two variable species [79,297]. However, a detailed study of the Iranian taxa [91,150] revealed three morphologically discrete species and allopolyploid relationships among them (V. stocksii 4x; V. cinerea 8x; V. behboudiana 8x), which may suggest more taxa warrant recognition within the section. Section Sclerosium may have started to diversify in Late Miocene 3.5–10 Ma ago [150]. The young age corroborates the low morphological differentiation among taxa. The crown group age coincides with the initiation (or intensification) of the Indian monsoon system, caused by the uplift of the Himalayas and the East African mountain plateaus [298,299]. The precipitation brought by the monsoon plays an important role for the flora in this otherwise arid region.
Section Sclerosium is vegetatively somewhat similar to sect. Xylinosium (especially Viola scorpiuroides) but the sections are distantly related, allopatric, they differ in several important characters, and any similarity must be interpreted as parallel adaptation to arid environments.
Viola sect. Spathulidium Marcussen, sect. nov.—Type: Viola spathulata Willd.
Description.—Perennial herbs. Axes not morphologically differentiated. All stems rhizomatous, forming cushions. Stipules ¾ adnate to petiole. Lamina spathulate to lanceolate, subentire, tapering into short and indistinct petiole. Corolla pale violet, pink or whitish. Spur 1.5–4 mm, longer than tall. Style clavate, geniculate at base, at apex 2-lobed, with a distinct dorsolateral margin and ventral rostrum. Cleistogamous flowers not produced. Allo-octoploid (CHAM + MELVIO). ITS sequence of MELVIO type.
Diagnostic characters.—Lamina spathulate to lanceolate, subentire, tapering into short and indistinct petiole AND style clavate, at apex 2-lobed, with a distinct dorsolateral margin AND cleistogamous flowers not produced.
Ploidy and accepted chromosome counts.—[Section by origin 8x], 16x (V. spathulata). Chromosome number unknown.
Age.—Crown node c. 1 Ma; stem node 5.0 (4.2–5.3) Ma [28].
Included species.—3. Violamaymanica Grey-Wilson, V. pachyrrhiza Boiss. & Hohen., V. spathulata Willd. ex Schult.
Distribution.—Disjunctly distributed in the high mountains of southwestern Asia (Figure 30): Viola pachyrrhiza in northeastern Iraq and southern Iran; V. spathulata in northern Iran (Elburs mountains); and V. maymanica in northwestern Afghanistan.
Etymology.—The name Spathulidium refers to the distinctive spathulate leaves.
Discussion.—Section Spathulidium is an allooctoploid CHAM + MELVIO lineage and has retained the MELVIO homoeolog for ITS (Figure 2). The lineage is morphologically recognisable on being cushion plants, inhabiting rock fissures, with spathulate short-petiolate leaves, a somewhat bilobed style, and the absence of cleistogamous flowers. The Spathulidium lineage is inferred to be the alloploid of two unknown tetraploid lineages; further allopolyploidy based on 8x may have happened in V. spathulata (16x) [28]. The three species of sect. Spathulidium have traditionally been grouped within sect. Plagiostigma subsect. Patellares based on being violet-flowered rosette plants with narrow leaves and adnate stipules [1,248]. However, sect. Spathulidium differs from subsect. Patellares in being cushion plants, having leaves with entire or subcrenate margins, in lacking cleistogamy, and in ploidy. Section Spathulidium differs from sect. Himalayum in being cushion plants, in having a margined style apex and a much longer spur, and in lacking cleistogamous flowers. Both sections are 8x but have different allopolyploid origins.
Section Spathulidium is most closely related to the African sect. Abyssinium (see note under the latter).
Viola sect. Tridens W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 376. 1925.—Type (Shenzhen Code Art. 10.8): Viola tridentata Sm.
Description.—Perennial procumbent herb, forming perennial herbaceous mats with branched stems. Axes morphologically differentiated in elongated rhizome and lateral, short floriferous stems with distichous phyllotaxy. Stipules completely adnate to the pseudopetiole or only with the free end forming a short tooth. Leaves tridentate on floriferous shoots, bilobate or entire on sterile shoots, small, imbricated, fleshy. Corolla small, white with violet striation. Spur short. Anthers with scattered hairs. Style cylindrical, at base curved, slightly tapering towards apex, filiform. Cleistogamous flowers not produced. Allohexaploid. Secondary base chromosome number x’ = 20.
Diagnostic characters.—Leaves tridentate, distichous, imbricate.
Ploidy and accepted chromosome counts.—6x; 2n = 40.
Age.—Crown node age not applicable (monotypic lineage); stem node 9.2 (1.0–14.7) Ma [28].
Included species.—1. Viola tridentata Sm.
Distribution.—Southernmost South America: Argentina, Chile, Falkland/Malvinas Islands (Figure 31).
Discussion.—Section Tridens is immediately recognisable by the tridentate, distichous, and imbricate leaves. Phylogenetically, sect. Tridens is allohexaploid and two of its diploid genomes are shared with other polyploid southern hemisphere lineages, i.e., Leptidium on the one side, and Chilenium/Erpetion on the other (Figure 5). The original inference by Marcussen et al. [28] that Tridens is 12x was based on incorrect counts for sect. Erpetion and sect. Tridens which overestimated the ploidy.
The delimitation of sect. Tridens is the same as Becker’s [1] except for the inclusion of V. muscoides Phil. as a synonym of V. tridentata based on shared diagnostic characters. Viola muscoides was erroneously synonymised with Myrteola nummularia (Poir.) O. Berg (Myrtaceae) by Kausel [300].
≡Viola sect. Nomimium Ging., p.p. in Mém. Soc. Phys. Genève 2(1): 28. 1823, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.) ≡ Viola subg. Nomimium (Ging.) Peterm., Deutschl. Fl.: 64. 1846, nom. inval. (Szhenzhen Code Art. 22.2)
≡Viola [sect. Nomimium; unranked] §.4. Rostellatae Boiss., Fl. Orient. 1: 451. 1867, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.) ≡ Viola subsect. Rostellatae (Boiss.) Rouy & Foucaud, Fl. France [Rouy & Foucaud] 3: 3. 1896, nom. inval. (Szhenzhen Code Art. 22.2). ≡ Viola sect. Rostellatae (Boiss.) J. C. Clausen in Madroño 17: 196. 1964, nom. inval. (Shenzhen Code Art. 22.2)
≡Viola [sect. Nomimium; unranked] a. Rostellata Nyman, Consp. Fl. Eur. 1: 76. 1878, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.)
Description.—Perennial herbs. Axes morphologically differentiated in a perennial rhizome with lateral stems; sometimes only one type of stem produced. Rhizome creeping or vertical, branched or not, with apical rosette of leaves. Lateral stems annual aerial stems, stolons, or absent. Stipules usually free, entire, dentate, laciniate or fimbriate, sometimes large and foliaceous. Lamina reniform to rhomboid, crenulate, petiolate. Flowers scented or scentless. Corolla violet to white, with a white throat. Spur (much) longer than tall, up to 16 mm. Style clavate or rarely filiform, at apex not margined, bearded or not. Capsule trigonous and explosive or globose and non-explosive. Cleistogamous flowers usually produced; cleistogamy seasonal, rarely facultative. Allotetraploid (CHAM + MELVIO). Secondary base chromosome number x’ = 10. ITS sequence of MELVIO type.
Diagnostic characters.—Perennial herbs AND corolla with a white throat AND style clavate, unmargined AND base chromosome number x = 10.
Ploidy and accepted chromosome counts.—4x, 8x; 12x; 2n = 20, 40, 58, 60.
Age.—Crown node 11.8 (10.1–12.4) Ma [28].
Included species.—75.
Distribution.—Throughout the temperate zone of the northern hemisphere; one species in southern South America (Figure 32). Diversity centre in western Eurasia.
Discussion.—Section Viola is phylogenetically an allotetraploid CHAM + MELVIO lineage and has retained the MELVIO homoeolog for ITS (Figure 6). Karyologically it is characterised by the secondary base chromosome number x’ = 10, and morphologically by the clavate unmargined style. Section Viola is one of three species-rich segregates of Becker’s widely delimited sect. Nomimium, which comprised nearly all the temperate herbaceous, violet- or white-flowered taxa with seasonal cleistogamy. Section Viola differs from both sect. Plagiostigma and sect. Nosphinium in having the base chromosome number x = 10 and a unmargined style, sometimes bearded above.
Section Viola is phylogenetically subdivided into two morphologically well-defined groups (Figure 6 and Figure 33), here treated as subsect. Rostratae and subsect. Viola.
Key to the subsections of sect. Viola
- 1a.
- Capsules globose, often hairy, non-dehiscent, on decumbent peduncles. Seeds large, with conspicuous elaiosome more than half the length of the seed (myrmecochory). Lateral stems stolons or absent. Style glabrous. ........................................................................................................................................................................................................................................................................ subsect. Viola
- 1b.
- Capsule elongate, trigonous, glabrous, forcibly ejecting seeds after dehiscence, on erect peduncles at maturity. The elaiosome much less than half the length of the seed (diplochory). Lateral stems usually aerial (occasionally stolons or absent). Style bearded above or beardless. ............................................................................................................................................................................................................................................................... subsect. Rostratae
Viola subsect. Rostratae (Kupffer) W. Becker in Acta Horti Gothob. 2: 285. 1926 ≡ Viola [unranked] (“Gruppe”) Rostratae Kupffer in Oesterr. Bot. Z. 53: 328. 1903 ≡ Viola sect. Rostratae (Kupffer) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 193. 1909 ≡ Viola [sect. Nomimium) [unranked] Rostratae (Kupffer) Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 365. 1925.—Lectotype (designated here): Viola riviniana Rchb.
≡Viola sect. Trigonocarpea Godron, Fl. Lorraine, ed. 2, 1: 88. 1857 ≡ Viola subsect. Trigonocarpea (Godr.) P. Y. Fu, Fl. Pl. Herb. Chin. Bor.-Or. 6: 82. 1977; Vl. V. Nikitin in Novosti Sist. Vyssh. Rast. 33: 178. 2001 (isonym).—Lectotype (Nikitin 1996 [301], page 189): Viola riviniana Rchb.
=Viola [unranked] Rosulantes Borbás in Hallier & Wohlfarth, Syn. Deutsch. Schweiz. Fl., ed. 3, 1: 196. 1892 ≡ Viola subsect. Rosulantes (Borbás) J. C. Clausen in Madroño 17: 196. 1964, nom. inval. (Shenzhen Code Art. 41.5)
=Lophion subg. Eucentrion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 216. 1914.—Type: Viola rostrata Pursh
=Lophion subg. Rhabdotion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 216. 1914.—Type: Viola striata Aiton
=Viola [sect. Nomimium; unranked] Umbraticolae W. Becker in Repert. Spec. Nov. Regni Veg. 19: 396. 1923.—Type (Shenzhen Code Art. 10.8): Viola umbraticola Kunth
=Viola [unranked] Repentes Kupffer in Oesterr. Bot. Z. 53: 329. 1903 ≡ Viola subsect. Repentes (Kupffer) Juz. in Schisk. & Bobrov, Fl. URSS 15: 401.—Type: Viola uliginosa Besser
≡Viola sect. Icmasion Juz. ex Tzvelev in Cvelev, Opred. Sosud. Rast. Severo-Zapadn. Rossii: 679. 2000.—Type: Viola uliginosa Besser
=Viola subsect. Grypocerae Espeut in Botanica Pacifica 9(1): 16. 2020.—Type: Viola grypoceras A. Gray
=Viola [unranked] Mirabiles Nyman Syll. Fl. Eur.: 226. 1855, nom. inval. (Shenzhen Code Art. 38.1) ≡ Viola [unranked] b2 Mirabiles Nyman ex Borbás Syn. Deutsch. Schweiz. Fl., ed. 3, 1: 195. 1890 ≡ Viola subsect. Mirabiles (Nyman ex Borbás) Juz. in Schischk. & Bobrov, Flora URSS 15: 375. 1949 ≡ Viola sect. Mirabiles (Nyman ex Borbás) Vl. V. Nikitin in Bot. Zhurn. (Moscow & Leningrad) 83(3): 130. 1998.—Type (Shenzhen Code Art. 10.8): Viola mirabilis L.
=Viola [sect. Chilenium] subsect. Coeruleae Sparre in Lilloa 17: 414. 1949.—Type: Viola huidobrii Gay
Description.—Rhizome with an apical leaf rosette and lateral aerial stems or stolons, or all stems rhizomatous, or all stems aerial. Stipules often large and foliaceous. Style bearded or not. Capsule trigonous, forcibly ejecting seeds after dehiscence. Seeds with a small elaiosome.
Diagnostic characters.—Capsules trigonous, erect at maturity, explosive; seeds with small elaiosome covering less than 1/2 of the raphe.
Ploidy and accepted chromosome counts.—4x, 8x, 12x; 2n = 20, 40, 58, 60.
Included species.—51. Violaacuminata Ledeb., V. adunca Sm., V. aduncoides Á. Löve & D. Löve, V. anagae Gilli, V. appalachiensis L. K. Henry, V. canina L., V. caspia (Rupr.) Freyn, V. dirphya Tiniakou, V. elatior Fr., V. faurieana W. Becker, V. ganpinensis W. Becker, ined. [E. Bodinier 2176], V. grayi Franch. & Sav., V. grypoceras A. Gray, V. henryi H. Boissieu, V. huidobrii Gay, V. jordanii Hanry, V. kosanensis Hayata, V. kusanoana Makino, V. labradorica Schrank, V. lactea Sm., V. laricicola Marcussen, V. mariae W. Becker, V. mauritii Tepl., V. mirabilis L., V. obtusa (Makino) Makino, V. oligyrtia Tiniakou, V. ovato-oblonga (Miq.) Makino, V. papuana W. Becker & Pulle, V. pendulicarpa W. Becker, V. percrenulata H. E. Ballard, ined. [H. S. Gentry 7247], V. pseudomirabilis H. J. Coste, V. pumila Chaix, V. reichenbachiana Jord. ex Boreau, V. riviniana Rchb., V. rostrata Pursh, V. rupestris F. W. Schmidt, V. sacchalinensis H. Boissieu, V. serrula W. Becker, V. shinchikuensis Yamam., V. sieheana W. Becker, V. stagnina Kit. ex Schult., V. stewardiana W. Becker, V. striata Aiton, V. tanaitica Grosset, V. thibaudieri Franch. & Sav., V. uliginosa Besser, V. umbraticola Kunth, V. utchinensis Koidz., V. walteri House, V. websteri Hemsl., V. willkommii R. Roem. ex Willk..
Distribution.—North-temperate, except for Viola huidobrii in southern South America and V. papuana in New Guinea. Viola riviniana is naturalised in western North America, Australia, and New Zealand.
Discussion.—Within sect. Viola, this lineage is characterised by the explosive capsules, borne on erect peduncles at maturity (in fact a plesiomorphic trait within Viola). Subsection Rostratae is widely distributed in the temperate zone of Eurasia and North America; one species occurs in southern South America, and one in New Guinea. Becker [1] included in grex Rostratae only species with aerial floriferous stems but subsequent studies have shown that the subsection should be more inclusive.
Subsect. Rostratae has often been further subdivided based on shoot system but none of the segregates delimit monophyletic units, due to extensive allopolyploidy and presumably also parallel evolution. The vast majority of the species in subsect. Rostratae have a basal leaf rosette and lateral floriferous stems; these have traditionally been referred to as grex Rosulantes Borbás. In a few species the chasmogamous flowers are produced from the leaf rosette and the aerial stems develop after chasmogamous anthesis; these have been referred to as grex Mirabiles Nym. (i.e., V. mirabilis, V. pseudomirabilis, and V. willkommii). Other species have lateral stems that are stolon-like (e.g., V. anagae, V. appalachiensis, V. papuana, V. walteri; grex Repentes Kupffer: V. uliginosa) or absent altogether (e.g., V. ganpinensis, V. pendulicarpa, and V. shinchikuensis; grex Umbraticolae W. Becker: V. percrenulata and V. umbraticola). Finally, a fourth group of species that lack the basal rosette and instead have a sympodial growth system from annual floriferous stems has been referred to as grex Arosulatae Borbás (i.e., V. canina, V. elatior, V. lactea, V. pumila, V. stagnina). The formal greges Arosulatae, Mirabiles, and Rosulantes are superfluous as they are de facto synonyms of the higher taxon subsect. Rostratae as a result of being interconnected by allopolyploids (Figure 33, [84,92,302]). Among these, only the group formerly referred to as “Arosulatae” may merit recognition on ecological grounds. These western Eurasian species are ecological specialists to floodplains [303,304] and each possesses at least one stagnina genome. We suggest that this group be referred to informally as the V. stagnina group. Becker included here V. acuminata and V. jordanii by mistake: neither has a sympodial growth system lacking a basal rosette nor possesses a stagnina genome (Figure 33, [84,302]).
Morphologically, the southern South American Viola huidobrii (including its synonym V. brachypetala) belongs in subsect. Rostratae, based on having a rhizome with a terminal leaf rosette and lateral floriferous stems, violet corolla, long spur, and the characteristic rostellate style (Figure 9el). Viola huidobrii was previously included in sect. Chilenium [1,62] or sect. Rubellium [63]. It is the only species of sect. Viola native to the southern hemisphere. The Taiwanese endemic V. shinchikuensis (2n = 20) is (erroneously?) reported to be similar to subsect. Viola in having globose capsules borne on prostrate peduncles when mature [75,305] but it is phylogenetically placed in subsect. Rostratae [86] (Figure 2) with which it also shares numerous typical traits, e.g., bearded style, acute sepals with dentate appendages, bracteoles in the uppermost part of the peduncle, and thick non-hyaline stipules. The New Guinean endemic V. papuana has an unusual filiform style (which puzzled Becker; Figure 9eo) and isolated distribution but is a good match for subsect. Rostratae in other morphological characters, including the 4–9 mm long, upcurved spur and a pale violet corolla, and lateral stems or stolons. The reported chromosome count of 2n = 48 [74] is dubious.
Subsect. Rostratae, and sect. Viola as a whole, appears to have originated in western Eurasia. Only one clade, grex Rosulantes s.str., has dispersed into eastern Asia, North America, and South America.
A read-leaved mutant of V. riviniana, f. purpurea auct., is sometimes grown as an ornamental, often under the erroneous name V. labradorica hort. non Schrank.
=Viola sect. Odoratae Boiss. in Diagn. Pl. Orient. 8: 51. 1849, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.)
=Viola sect. Hypocarpea Godron, Fl. Lorraine, ed. 2, 1: 86. 1857 ≡ Viola subsect. Hypocarpea (Godron) P. Y. Fu, Fl. Pl. Herb. Chin. Bor.-Or. 6: 82. 1977, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.)
=Viola [unranked] (”Gruppe”) Uncinatae Kupffer in Oesterr. Bot. Z. 53: 328. 1903, nom inval. (Szhenzhen Code Art. 22.2; Viola odorata L.) ≡ Viola sect. Uncinatae (Kupffer) Kupffer in Kusnezow et al., Fl. Caucas. Crit. 3(9): 174. 1909, nom. inval. (Szhenzhen Code Art. 22.2)
=Viola [unranked] a) Curvato-pedunculatae W. Becker in Beih. Bot. Centralbl., Abt. 2, 26: 1. 1910, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.)
=Viola subg. Euion Nieuwl. & Kaczm. in Amer. Midl. Naturalist 3: 211. 1914, nom. inval. (Szhenzhen Code Art. 22.2; Viola odorata L.)
=Viola [unranked] α Lignosae W. Becker in Beih. Bot. Centralbl., Abt. 2, 26: 1. 1910 ≡ Viola [unranked] (“Gruppe”) D. Lignosae W. Becker in Nat. Pflanzenfam. ed. 2 [Engler & Prantl], 21: 367. 1925.—Lectotype (designated here): Viola chelmea Boiss.
=Viola [unranked] (“Gruppe”) Serpentes W. Becker in Beih. Bot. Centralbl., Abt. 2, 40: 102. 1924—Viola subsect. Serpentes (W. Becker) W. Becker in Acta Horti Gothoburg. 2: 287. 1926—Viola ser. Serpentes (W. Becker) Steenis in Bull. Jard. Bot. Buitenzorg, ser. 3, 13 (1933–1936): 259. 1934—Viola sect. Serpentes Ching J. Wang, Fl. Reipubl. Popularis Sin. 51: 88. 1991.—Type (Shenzhen Code Art. 10.8): Viola serpens Wall. ex Ging. (=V. pilosa Blume)
Description.—Rhizome with apical rosette of leaves. Lateral stolons present or absent. Stipules free, not foliaceous. Style beardless. Capsule globose, non-explosive. Seeds with large elaiosome.
Diagnostic characters.—Capsules globose, usually hairy, decumbent at maturity, non-dehiscent. Seeds with a large elaiosome covering 1/2–3/4 of the raphe.
Ploidy and accepted chromosome counts.—4x, 8x; 2n = 20, 40.
Included species.—24. Violaalba Besser, V. ambigua Waldst. & Kit., V. barhalensis G. Knoche & Marcussen, V. bocquetiana Yıld., V. canescens Wall., V. chelmea Boiss., V. collina Besser, V. hirta L., V. hondoensis W. Becker & H. Boissieu, V. indica W. Becker, V. isaurica Contandr. & Quézel, V. jangiensis W. Becker, V. jaubertiana Marès & Vigin., V. kizildaghensis Dinç & Yıld., V. libanotica Boiss., V. odorata L., V. pilosa Blume, V. pyrenaica Ramond ex DC., V. sandrasea Melch., V. sintenisii W. Becker, V. suavis M. Bieb., V. thomasiana Songeon & E. P. Perrier, V. vilaensis Hayek, V. yildirimlii Dinç & Bagci
Distribution.—Eurasia; diversity centre in southern Europe. Viola odorata is naturalised throughout the temperate zone.
Discussion.—The principal apomorphy of subsect. Viola is the globose and non-explosive capsules borne on decumbent peduncles, containing large seeds with a conspicuous elaiosome, an adaptation to obligate myrmecochory. Subsection Viola as circumscribed here comprises three of Becker’s [1] greges. These include grex Uncinatae W. Becker (V. odorata, etc.) with both stolonose and estolonose temperate taxa, grex Lignosae W. Becker (V. chelmea, etc.) with estolonose taxa from the northeastern Mediterranean region, and parts of grex Serpentes W. Becker (V. pilosa, etc.) with stolonose taxa from southern Asia. The presence or absence of stolons has been used to classify species within the subsection but does not delimitate monophyletic groups [116]. At least in European species, the transitions from the stolonose condition (ser. Flagellatae Kittel) to the estolonose condition (ser. Eflagellatae Kittel) seems to have occurred several times and by different genetic mechanisms, and the two morphological groups are also linked by allopolyploidy, i.e., V. suavis (8x) [116]. Grex Serpentes has been demonstrated to be an artificial aggregate of species [229], most of them belonging in sect. Viola subsect. Viola or in various sect. Plagiostigma subsections.
A few species are grown as ornamentals, primarily for their fragrant flowers, i.e., V. odorata and filled forms of V. alba subsp. dehnhardtii (Ten.) W. Becker referred to as ‘Parma’ violets or ‘Violette de Toulouse’ [7,20]. The former (Figure 1) has been cultivated for the production of essential oil for the perfume industry [16,17].
Viola sect. Xanthidium Marcussen, Nicola, J. M. Watson, A. R. Flores & H. E. Ballard, sect. nov.—Type: Viola flavicans Wedd.
Description.—Perennial herbs. Axes not morphologically differentiated. All stem rhizomatous, with leaves in loose apical rosettes. Stipules partially or largely adnate to the petiole, narrow, shallowly glandular-lacerate. Lamina lanceolate, remotely crenate, petiolate. Bracteoles narrow, shallowly glandular-lacerate. Corolla yellow with brown striation. Spur short. Style clavate, geniculate at the base, when fresh ellipsoid with broadly rounded apex (in dried condition with flattened apex), the stigmatic orifice on a small rostellum on ventral surface, bearded (Viola flavicans; Figure 9fb) or beardless (V. pallascaensis; Figure 9fc). Cleistogamous flowers apparently produced; type of cleistogamy unknown.
Diagnostic characters.—Rosulate herbs AND bracteoles glandular-lacerate AND corolla yellow AND style ellipsoid with broadly rounded apex when fresh, flattened when dry.
Distribution.—Disjunct in central-western South America (northwestern Argentina and Bolivia, central-eastern Peru) (Figure 34).
Included species.—2. Viola flavicans Wedd., V. pallascaensis W. Becker
Etymology.—The name Xanthidium is based on the Greek translation of the species epithet of the type species, Viola flavicans, which refers to its yellow corolla.
Discussion.—Section Xanthidium has not yet been subject to phylogenetic analysis nor has it been characterised at the chromosomal level. Becker placed neither of these species (nor their current synonyms) in any section. He identified the taxa as related, but did not include them in his genus treatment [1]. Later, Sparre ([63], page: 348) viewed this group (as the “V. flavescens-group”) as “intermediary between the sections Chilenium and Andinium”. Nicola [80] placed V. flavicans in sect. Nomimium Ging., an artificial aggregate of numerous northern hemisphere lineages and sections.
Viola sect. Xylinosium W. Becker in Nat. Pflanzenfam., ed. 2 [Engler & Prantl], 21: 373. 1925.—Lectotype (designated here): Viola arborescens L.
=Viola [sect. Nomimium; unranked] Fruticulosa Nyman, Consp. Fl. Eur. 1: 76. 1878, nom. inval. (Shenzhen Code Art. 32.1)
Description.—Perennial subshrubs. Axes not morphologically differentiated. All stems aerial, decumbent or ascendent. Stipules green, linear, with 0–2 basal, lateral, smaller segments. Lamina lanceolate, crenate or subentire, sessile or indistinctly petiolated. Bracteoles minute or caducous (0–2 mm). Corolla violet to whitish with a white throat or corolla bright yellow throughout. Spur stout or saccate, longer than calycine appendages. Style clavate, unmargined, beardless. Cleistogamous flowers not produced. Allopolyploid (CHAM + MELVIO). Secondary base chromosome number x’ = 26. ITS sequence of MELVIO type.
Diagnostic characters.—Subshrubs AND lamina lanceolate, remotely crenate, indistinctly petiolated AND base chromosome number x = 26.
Ploidy and accepted chromosome counts.—≥4x; 2n = 52 (V. arborescens, V. saxifraga).
Age.—Crown node age 5.6 (3.9–6.2) Ma; [28].
Included species.—3. Viola arborescens L., V. saxifraga Maire, V. scorpiuroides Coss.
Distribution.—Three disjunct species in the Mediterranean region (Figure 35): Viola saxifraga in the high Atlas, V. arborescens in the western Mediterranean, V. scorpiuroides in the southeastern Mediterranean.
Discussion.—Section Xylinosium is phylogenetically an allopolyploid CHAM + MELVIO lineage and has retained the MELVIO homoeolog for ITS (Figure 2). Karyologically it is characterised by the secondary base chromosome number x’ = 26, and morphologically by the subshrubby habit in combination with the minute bracteoles (caducous in V. arborescens and V. saxifraga; 1–2 mm in V. scorpiuroides). The exact ploidy and genomic constitution of the section are obscured by gene loss and duplication [28]. Presumably, 2n = 52 reflects octoploidy. Pollen in both V. arborescens and V. saxifraga is monomorphic 3-colporate, and in V. scorpiuroides heteromorphic 3–4-colporate which indicates secondary polyploidy in this species [157]. Both Becker [1] and Marcussen et al. [28] included in sect. Xylinosium also the South African V. decumbens. Here, we place this last taxon in the monotypic sect. Melvio; see there for justification. The species of sect. Xylinosium have sometimes been confused with those of the vegetatively similar, allopatric sect. Sclerosium (e.g., [295,306]), which may explain the erroneous report of cleistogamous flowers in sect. Xylinosium [73].
3. Materials and Methods
To generate a comprehensive taxonomy for Viola, we first compiled a list of accepted species. Morphological and chromosome count data for these species were reconciled with phylogenetic data and used to infer monophyletic groups and define apomorphies based on which, using a set of predefined criteria, the classification was based. A downloadable and printable poster with the Viola images presented in Figure 3 is available in Supplementary File S1. The data (scripts and analysis files) presented in this study are available in Supplementary File S2.
3.1. Species Checklist
To generate a global species checklist for Viola we first downloaded the list of accepted species names for Viola from the Plants of the World Online database [307] and further revised this list of species according to our expert knowledge and based on published taxonomic treatments. This included adding numerous published names, most of which we accept as species, along with some that we consider synonymous but which are accepted by other authorities. We also included the accepted fossil species. Where relevant, protologues and type specimens were inspected. We classified taxa as either “accepted”, for the species recognised by us including entities not yet published, “hybrid” for interspecific hybrids, “included” for infraspecific taxa of an accepted species such as a subspecies or variety, “synonym” for species synonyms, “unresolved” in the rare case that rank or validity of a taxon could not be determined, and “rejected” for rejected names. Subspecies and varieties are included only in the cases that they are mentioned in the synonymy. For synonyms, instead of including all names ever published for Viola, most of which have been out of use for a long time, we included those names cited in the updated literature covering the distribution range of Viola, i.e., Africa [308], Australia [226], central Asia [79,309,310], eastern Asia [61,74,78,311], Europe [22,73,312,313,314], North America [291,315], and South America [68,80,316]. The original list downloaded from POWO comprised 761 entries; the edited list, including many new species which we individually confirm as distinct and requiring publication, comprises 1792 entries, 664 of which we accept as species.
3.2. Morphology Data
A wide range of morphological traits of flowering and fruiting plants were examined on herbarium specimens, including online images verified as to identity, for several to many representative and morphologically diverse species in larger infrageneric groups and for most or all species in smaller groups, where available. Protologues and recently published descriptions for many species (where identifications were confirmed) were also consulted. Of particular value in delimiting or distinguishing infrageneric groups or suggesting relationships among groups were growth form; duration, habit of rhizomes, stems, or stolons; stipule size and shape, adnation, and margins; leaf lamina features; calycine appendage size, shape, and margins; corolla throat and petal colour pattern; shape of bottom (anterior) petal and its size relative to lateral and upper petals; spur size and shape; presence or absence of beards (indument within) on lateral or bottom petals; style features; capsule dehiscence behaviour; and ability/inability to produce cleistogamous flower and whether cleistogamy is seasonal or not. Previous classifications [1,29,46,47] have highlighted style morphology as particularly important in diagnosing and comparing groups, and other studies have shown details of style morphology to be effective species-diagnostic traits [317]. We made special efforts to survey styles from numerous species across all infrageneric groups (Figure 9), from specimens and from the literature, developed a rubric for interpreting and describing particular features, developed descriptions of styles for individual species, then created summary descriptions for all groups.
3.3. Chromosome Number Data
Base chromosome numbers within Viola differ among sublineages (e.g., x = 6 in sect. Chamaemelanium, x = 10 in sect. Viola, x = 12 in sect. Plagiostigma). In order to systematise this information, we first downloaded data on chromosome counts for all species from the Chromosome Counts Database (CCDB) [318] and from primary literature sources. We then evaluated the reliability of individual counts and discarded counts that did not fit other counts on the same species or lineage in terms of ploidy and base number.
3.4. Criteria and Principles for an Updated Infrageneric Classification of Viola
We proposed a phylogenetic classification, based on previously published data (primarily [28,45]). Criteria for defining the formal infrageneric taxa were that they are monophyletic and/or possess apomorphies (morphological or other). Taxonomic levels and taxon names were chosen to maximise taxonomic stability and continuity. Allopolyploidy is widespread in Viola and its phylogeny has the topology of a network rather than a tree. Such reticulate phylogenies are not always reconcilable with a hierarchical classification. To accommodate for the conflicting situations we have chosen to accept the three infrageneric segregate taxa (e.g., sections) A, B, and X even if X is the allopolyploid of A and B. This affected sect. Chamaemelanium (which is diploid and possibly contributed genomes to a dozen of allotetraploid sections/lineages) and sect. Nosphinium (which is 10x and combines genomes from three other sections/lineages). In the case that an infrageneric segregate taxon (e.g., section) is known to contain internal polyploids, we have chosen to delimit it so that A, B, and X, as defined above, are monophyletic. For example, subsect. Stolonosae is typified with V. palustris, and because V. palustris (8x) is the alloploid of V. epipsila (4x) and V. minuscula (4x) [45], subsect. Stolonosae by definition has to comprise at least these three species.
3.5. Generating Distributional Maps for Viola Sections
Occurrence data for each Viola section was downloaded from the Global Biodiversity Information Facility (GBIF) database [319] using a custom R [320] script using the packages rgbif [321], tidyverse [322], and raster [323], and cleaned using speciesgeocodeR [324]. All the occurrence datasets were accessed via https://GBIF.org (accessed on 11 December 2021) and have the following DOIs: subg. Neoandinium https://doi.org/10.15468/dl.6a3dvh, sect. Abyssinium https://doi.org/10.15468/dl.utndtm, sect. Chamaemelanium https://doi.org/10.15468/dl.wr7kd5 and https://doi.org/10.15468/dl.fg8kk8, sect. Chilenium https://doi.org/10.15468/dl.5ugyp9, sect. Danxiaviola https://doi.org/10.15468/dl.9v545h, sect. Delphiniopsis https://doi.org/10.15468/dl.ct87uy, sect. Erpetion https://doi.org/10.15468/dl.r7tjd3, sect. Himalayum https://doi.org/10.15468/dl.rhqf8q, sect. Leptidium https://doi.org/10.15468/dl.wscdns, sect. Melanium https://doi.org/10.15468/dl.p6ysnh, sect. Melvio and sect. Nematocaulon https://doi.org/10.15468/dl.v5nrqx, sect. Nosphinium https://doi.org/10.15468/dl.stx66g and https://doi.org/10.15468/dl.fhw4xu, sect. Plagiostigma https://doi.org/10.15468/dl.jsftmz, sect. Rubellium https://doi.org/10.15468/dl.a9cpek, sect. Sclerosium https://doi.org/10.15468/dl.mvahfp, sect. Spathulidium https://doi.org/10.15468/dl.x5btdu, sect. Tridens https://doi.org/10.15468/dl.ufbaqp, sect. Viola https://doi.org/10.15468/dl.efxwyy, sect. Xanthidium https://doi.org/10.15468/dl.vn5t5f, and sect. Xylinosium https://doi.org/10.15468/dl.d48ncv. To each dataset we added further records that had not been uploaded to public databases, from, e.g., literature, herbarium specimens, and field surveys. Maps were constructed using a custom R [320] script using the packages maptools [325], rgdal [326], and reader [327].
3.6. Monoploid Phylogeny of Viola (Figure 5)
We reinterpreted the phylogenetic network of Marcussen et al. [28] based on new information, i.e., new chromosome count for Viola banksii (2n = 50 not 60 [98]), correction of chromosome count for V. tridentata (2n = 40 not 80 [99]), correction of the interpretation of homoeologs in V. decumbens, and sequences of new taxa (e.g., [90,231]). The species checklist is available in Appendix A.
3.7. ITS Phylogeny for Viola (Figure 6)
In order to obtain a phylogeny with denser taxon sampling, we downloaded sequences of the ribosomal internal transcribed spacers 1 and 2 (ITS) for 87 representative species from GenBank, including one outgroup, and obtained another three sequences by PCR following the protocol of Ballard et al. [2] (Table 4). Sequences were combined in cases where ITS1 and ITS2 had been sequenced separately for the same species. The resulting 90 sequences were aligned in AliView [328] and terminal gaps were coded as “?”. Indels were coded by Simple Indel Coding [329] in SeqState v1.4.1 [330]. The analysis was set up in BEAUTi v1.10.4 and analysed in BEAST v1.10.4 [331] with substitution model GTR + G for the nucleotide partition and a 1-rate + G model (equivalent to JC + G) for the indel partition, a common uncorrelated lognormal clock, a Yule tree prior. The MCMC chain was run for 20 million generations with subsampling every 10,000 generations and monitored in Tracer v1.7.1 [332] to ensure all parameters reached convergence and the recommended effective sample size of at least 200. After removal of a 10% burn-in, the maximum credibility tree was calculated in TreeAnnotator v1.10.4 [331] and visualised in FigTree [333]. Normal age priors, specified as N(µ,σ), were obtained from the appendix of Marcussen et al. [28] and applied to five crown nodes, i.e., Viola N(30.9,0.38), the CHAM lineage N(18.98,0.35), the MELVIO lineage N(18.71,0.34), sect. Plagiostigma (16.62,0.45), and sect. Melanium N(12.51,0.25). Section Plagiostigma and Viola subg. Viola were each constrained as monophyletic.
3.8. Historical Biogeography of Viola (Figure 8)
We reconstructed the discrete historical biogeography of Viola (Figure 8) using a simplified approach based on stochastic character mapping [103] of four biogeographic categories, a single-rate transition model, and 50 operational taxonomic units as defined in the diploid multilabelled phylogenetic timetree [334] that is the counterpart of the phylogenetic allopolyploid network in Figure 5. Each section of the genus was given either of four biogeographic categories (Australia, northern hemisphere, South Africa, and South America) in correspondence with the area shared by 90% of its species. Stochastic character mapping was performed with 1000 simulations using the R [320] package phytools [335].
3.9. Multigene Phylogeny for Sect. Chamaemelanium (Figure 13)
The Chamaemelanium phylogeny was generated based on concatenated sequences of the nuclear regions GPI, NRPD2a, and ITS, and the chloroplast region trnL-trnF (Table 5). The analysis was set up in BEAUTi v1.10.4 and analysed in BEAST v1.10.4 [331] with substitution model GTR + G for each of the nucleotide partitions, a common uncorrelated lognormal clock, and a Yule tree prior. The MCMC chain was run for 10 million generations with subsampling every 10,000 generations and monitored in Tracer v1.7.1 [332] to ensure all parameters reached convergence and the recommended effective sample size of at least 200. After removal of a 10% burn-in, the maximum credibility tree was calculated in TreeAnnotator v1.10.4 [331] and visualised in FigTree [333]. The ingroup (sect. Chamaemelanium) was constrained as monophyletic.
3.10. Historical Biogeography and Age of the Hawaiian Violets, Subsect. Nosphinium (Figure 25)
The historical biogeography and age of subsect. Nosphinium (Figure 25) was estimated by simultaneous analysis of ITS sequence data, island biogeography, and node dating. Available sequences of the Hawaiian taxa and outgroups (Table 6) were downloaded from GenBank and aligned in AliView [328]. The dating analysis was set up in BEAUTi v1.10.4 and analysed in BEAST v1.10.4 [331] with substitution model GTR + G for the nucleotide partition with useAmbiguities set to “true”, an uncorrelated lognormal clock, and a Yule tree prior. Biogeography (i.e., island), obtained from the original publications [81,85,336,337], was added as a discrete trait and analysed under a symmetrical model and a strict clock; the biogeography of outgroup taxa was scored as missing (“?”). The MCMC chain was run for 100 million generations with subsampling every 10,000 generations and monitored in Tracer v1.7.1 [332] to ensure all parameters reached convergence and the recommended effective sample size of at least 200. After removal of a 10% burn-in, the maximum credibility tree was calculated in TreeAnnotator v1.10.4 [331] and visualised in FigTree [333]. A normal age prior, N(8.44,0.34) Ma, obtained from the appendix of Marcussen et al. [28], was applied to the crown node of sect. Nosphinium. Subgenus Viola and sect. Nosphinium were each constrained as monophyletic.
3.11. Multigene Phylogeny for Sect. Plagiostigma and Sect. Viola (Figure 27 and Figure 33)
The multigene phylogeny for sect. Plagiostigma and sect. Viola (Figure 27 and Figure 33) was generated based on concatenated sequences of the eight nuclear regions GPI-C (CHAM homoeolog), GPI-M (MELVIO homoeolog), NRPD2a-C (CHAM homoeolog), NRPD2a-M (MELVIO homoeolog), ITS-C (CHAM homoeolog), ITS-M (MELVIO homoeolog), SDH-C (CHAM homoeolog), and SDH-M (MELVIO homoeolog; Table 7). Sequence reads for the genomic sequences (V. acuminata, V. dissecta, V. grypoceras, V. raddeana) were obtained by BLAST searching the NCBI Sequence Reads Archive (SRA) database and were assembled in BioEdit [338] by CAP alignment and in AliView [328]. The phylogenetic analysis was set up in BEAUTi v1.10.4 and analysed in BEAST v1.10.4 [331] with substitution model HKY + G for each of the four nucleotide partitions (i.e., GPI, ITS, NRPD2a, SDH), a common uncorrelated lognormal clock, and a Yule tree prior. Two MCMC chains were run for a total of 150 million generations with subsampling every 10,000 generations and monitored in Tracer v1.7.1 [332] to ensure all parameters reached convergence and the recommended effective sample size of at least 200. After removal of a 1% burn-in of each chain, the chains were merged with LogCombiner v.1.10.4 [331] and the maximum credibility tree was calculated in TreeAnnotator v1.10.4 [331] and visualised in FigTree [333]. The ingroup (sect. Plagiostigma + sect. Viola) was constrained as monophyletic.
Supplementary Materials
File S1: Photographs of representatives of each accepted infrageneric segregate; File S2: The data (scripts and analysis files) presented in this study. The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11172224/s1.
Author Contributions
Conceptualisation, all coauthors; methodology, T.M.; software, T.M., H.E.B., J.D. and M.V.N.; validation, all coauthors; formal analysis, T.M., H.E.B., J.D. and M.V.N.; investigation, all coauthors.; resources, T.M., H.E.B., J.D. and M.V.N.; data curation, all coauthors; writing—original draft preparation, T.M.; writing—review and editing, T.M., H.E.B., J.D. and M.V.N.; visualization, T.M.; supervision, n/a; project administration, T.M.; funding acquisition, J.D. and M.V.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by Agencia Nacional de Promoción Científica y Tecnológica, Argentina (ANPCyT), grant number PICT 2017-1939 to M.V.N. and by a long-term research development project No. RVO 67985939 of the Czech Academy of Sciences to J.D.
Data Availability Statement
The data (scripts and analysis files) presented in this study are available in the Supplementary Material.
Acknowledgments
Elżbieta Kuta, Gerd Knoche, Kevin R. Thiele, Masashi Igari, Narjes Yousefi, Qiang Fan, Yan-Shuang Huang, and Yousheng Chen are thanked for taxonomic advice and discussions. Gerhard Karrer is thanked for consulation on morphology and life forms, Kanchi Gandhi for advice on nomenclatural matters, Andrea S. Meseguer for sharing R scripts, and Alexander Sennikov for locating type material. Chris Blaxland is thanked for giving permission to use Kim Blaxland’s excellent photos and drawings of Viola styles. Elżbieta Kuta is thanked for providing images of styles. We thank the following for contributing images of Viola to Figure 3: Abdelmonaim Homrani Bakali, Alan Keohane, Albert Keshet and Ori Fragman-Sapir (Flora of Israel Online), Andrew Townsend, Dewald du Plessis, Dieter Zschummel, Instituto de Botánica Darwinion, Jerome Viard, Jonathan Goff, Karl Magnacca, Kuan-Chieh (Chuck) Hung, larsonek, Magriet Brink, Mário Duchoň, Mary Ann Yaich, Masashi Igari, Neptalí Ramírez Marcial, Pablo Silva, Qiang Fan, Robert v. Blittersdorff (www.eastafricanplants.org), Sam Wilson, Santiago Martín-Bravo, Sylvain Piry, Toshihiro Nagata, Vladimir Epiktetov, and Yanshuang Huang.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Global species checklist for Viola. Provisional, unpublished names are not included. Accepted species are indicated in boldface. Out of a total of 1796 cited taxa, 664 are extant and accepted at the species level, and 4 are fossil.
- Crocion nuttallii (Pursh) Nieuwl. & Lunell—synonym of Viola nuttallii (sect. Chamaemelanium)
- Crocion vallicola (A. Nelson) Nieuwl. & Lunell—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Erpetion hederaceum (Labill.) G. Don.—synonym of V. hederacea (sect. Erpetion)
- Erpetion reniforme Sweet—synonym of V. banksii (sect. Erpetion)
- Ionidium stipulare (Sw.) Schult.—synonym of V. stipularis (sect. Leptidium)
- Lophion aduncum (Sm.) Nieuwl. & Lunell—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Lophion rostratum (Pursh) Nieuwl. & Kaczmarek—synonym of V. rostrata (sect. Viola, subsect. Rostratae)
- Lophion rugulosum (Greene) Lunell—synonym of V. rugulosa (sect. Chamaemelanium)
- Lophion striatum (Aiton) Nieuwl. & Kaczmarek—synonym of V. striata (sect. Viola, subsect. Rostratae)
- Mnemion arvense (Murray) Nieuwl. & Kaczmarek—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola abbreviata J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola aberrans (Greene ex W. Stone) House—hybrid (V. communis × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola abulensis Fern. Casado & Nava—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola abundans House—hybrid (V. fimbriatula × V. sagittata), synonym of V. subsagittata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola abyssinica Steud. ex Oliv.—(sect. Abyssinium)
- Viola abyssinica var. eminii Engl.—synonym of V. eminii (sect. Abyssinium)
- Viola abyssinica var. ulugurensis Engl.—synonym of V. eminii (sect. Abyssinium)
- Viola acanthophylla Leyb. ex Reiche—(sect. Grandiflos)
- Viola acanthophylla var. tontalensis W. Becker—synonym of V. flos-idae (sect. Triflabellium)
- Viola accrescens Klokov—synonym of V. pumila (sect. Viola, subsect. Rostratae)
- Viola achyrophora Greene—synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola acrocerauniensis Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola acuminata Ledeb.—(sect. Viola, subsect. Rostratae)
- Viola acutifolia (Kar. & Kir.) W. Becker—(sect. Chamaemelanium)
- Viola acutilabella Hayata—synonym of V. nagasawae (sect. Plagiostigma, subsect. Diffusae)
- Viola adenothrix Hayata—(sect. Plagiostigma, subsect. Stolonosae)
- Viola adriatica Freyn—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola adulterina Godr.—hybrid (V. alba × V. hirta; sect. Viola, subsect. Viola)
- Viola adunca Sm.—(sect. Viola, subsect. Rostratae)
- Viola adunca Sm. subsp. adunca—(sect. Viola, subsect. Rostratae)
- Viola adunca subsp. ashtoniae M. S. Baker—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola adunca subsp. bellidifolia (Greene) H. E. Ballard, ined.—(sect. Viola, subsect. Rostratae)
- Viola adunca var. bellidifolia (Greene) H. D. Harrington—synonym of V. adunca subsp. bellidifolia (sect. Viola, subsect. Rostratae)
- Viola adunca var. minor (Hook.) Fernald—synonym of V. labradorica (sect. Rostratae)
- Viola aduncoides Á. Löve & D. Löve—(sect. Viola, subsect. Rostratae)
- Viola aethnensis (DC.) Strobl—(sect. Melanium, subsect. Bracteolatae)
- Viola aethnensis (DC.) Strobl subsp. aethnensis—(sect. Melanium, subsect. Bracteolatae)
- Viola aethnensis subsp. messanensis (W. Becker) Merxm. & Lippert—(sect. Melanium, subsect. Bracteolatae)
- Viola aethnensis subsp. splendida (W. Becker) Merxm. & Lippert—(sect. Melanium, subsect. Bracteolatae)
- Viola aetolica Boiss. & Heldr.—(sect. Melanium, subsect. Bracteolatae)
- Viola affinis Leconte—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola affinis var. subarctica J. Rousseau—synonym of V. affinis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola aizoon Reiche—(sect. Sempervivum)
- Viola alabamensis Pollard—synonym of V. villosa (sect. Nosphinium, subsect. Borealiamericanae)
- Viola alaica Vved.—(sect. Plagiostigma, subsect. Patellares)
- Viola alata Burgersd.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola alba Besser—(sect. Viola, subsect. Viola)
- Viola alba Besser subsp. alba—(sect. Viola, subsect. Viola)
- Viola alba subsp. cretica (Boiss. & Heldr.) Marcussen—(sect. Viola, subsect. Viola)
- Viola alba subsp. dehnhardtii (Ten.) W. Becker—(sect. Viola, subsect. Viola)
- Viola alba subsp. scotophylla (Jord.) Nyman—synonym of V. alba subsp. alba (sect. Viola, subsect. Viola)
- Viola alba subsp. sintenisii (W. Becker) W. Becker—synonym of V. sintenisii (sect. Viola, subsect. Viola)
- Viola alba subsp. thessala (Boiss. & Spruner) Hayek—synonym of V. alba subsp. alba (sect. Viola, subsect. Viola)
- Viola albanica Halácsy—(sect. Melanium, subsect. Bracteolatae)
- Viola albida Palib.—(sect. Plagiostigma, subsect. Patellares)
- Viola albimaritima Vl. V. Nikitin (pro hybr.)—synonym of V. suecica subsp. suecica (sect. Plagiostigma, subsect. Stolonosae)
- Viola albovii Vl. V. Nikitin—hybrid (V. oreades × V. orthoceras; sect. Melanium, subsect. Bracteolatae)
- Viola alburnica Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola alexandrowiana (W. Becker) Juz.—(sect. Plagiostigma, subsect. Patellares)
- Viola alexejana Kamelin & Junussov—(sect. Plagiostigma, subsect. Patellares)
- Viola aliceae M. S. Baker—synonym of V. umbraticola (sect. Viola, subsect. Rostratae)
- Viola alisoviana f. intermedia (Kitag.) Taken.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola alisoviana Kiss—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola allchariensis Beck—(sect. Melanium, subsect. Bracteolatae)
- Viola allchariensis Beck subsp. allchariensis—synonym of V. allchariensis (sect. Melanium, subsect. Bracteolatae)
- Viola allchariensis subsp. gostivarensis W. Becker & Bornm.—synonym of V. gostivariensis (sect. Melanium, subsect. Bracteolatae)
- Viola allegheniensis L. K. Henry, non Roem. & Schult.—unresolved
- Viola alliariifolia Nakai—(sect. Chamaemelanium)
- Viola allochroa Botsch.—(sect. Chamaemelanium)
- Viola alpina Jacq.—(sect. Melanium, subsect. Bracteolatae)
- Viola alpina Ruiz & Pav. ex Ging.—synonym of V. pygmaea (sect. Sempervivum)
- Viola altaica Ker Gawl.—(sect. Melanium, subsect. Bracteolatae)
- Viola altaica subsp. oreades (M. Bieb.) W. Becker—synonym of V. oreades (sect. Melanium, subsect. Bracteolatae)
- Viola amamiana Hatus.—(sect. Plagiostigma, subsect. Diffusae)
- Viola ambigua Waldst. & Kit.—(sect. Viola, subsect. Viola)
- Viola amiatina Ricceri & Moraldo—synonym of V. etrusca (sect. Melanium, subsect. Bracteolatae)
- Viola amorphophylla Pollard—synonym of V. fimbriatula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola ampliata Greene—synonym of V. pedata subsp. pedata (sect. Nosphinium, subsect. Pedatae)
- Viola amurica W. Becker—(sect. Plagiostigma, subsect. Bilobatae)
- Viola anagae Gilli—(sect. Viola, subsect. Rostratae)
- Viola angellae Pollard—hybrid (V. palmata var. triloba × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola angkae Craib—(sect. Chamaemelanium)
- Viola angustifolia Phil.—(sect. Grandiflos)
- Viola angustistipulata C. C. Chang—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola anitae J. M. Watson—(sect. Rhizomandinium)
- Viola annamensis Baker f.—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola apoensis Elmer [“apoense”]—(sect. Plagiostigma, subsect. Diffusae)
- Viola appalachiensis L. K. Henry—(sect. Viola, subsect. Rostratae)
- Viola araucaniae W. Becker—(sect. Subandinium)
- Viola arborescens L.—(sect. Xylinosium)
- Viola arbuscula Phil.—synonym of V. philippii (sect. Rosulatae)
- Viola arcuata Blume—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola arcuata Miq., non Blume—synonym of V. javanica (sect. Plagiostigma, subsect. Stolonosae)
- Viola arcuata Ooststr. & H. J. Lam, non Blume—synonym of V. kjellbergii (sect. Plagiostigma, subsect. Stolonosae)
- Viola arenaria DC.—synonym of V. rupestris subsp. rupestris (sect. Viola, subsect. Rostratae)
- Viola argenteria Moraldo & Forneris—synonym of V. nummulariifolia (sect. Melanium, subsect. Pseudorupestres)
- Viola argentina W. Becker—(sect. Rosulatae)
- Viola arguta Humb. & Bonpl. ex Schult.—(sect. Leptidium)
- Viola arguta subsp. meridionalis W. Becker—synonym of V. arguta (sect. Leptidium)
- Viola arguta subsp. typica W. Becker—synonym of V. arguta (sect. Leptidium)
- Viola arguta var. glaberrima W. Becker—synonym of V. arguta (sect. Leptidium)
- Viola arguta var. meridionalis (W. Becker) L. B. Sm. & A. Fernández—synonym of V. arguta (sect. Leptidium)
- Viola arisanensis W. Becker—synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola arizonica Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola arsenica Beck—(sect. Melanium, subsect. Bracteolatae)
- Viola arvensioides Strobl—synonym of V. hymettia (sect. Melanium, subsect. Bracteolatae)
- Viola arvensis Murray—(sect. Melanium, subsect. Bracteolatae)
- Viola arvensis Murray subsp. arvensis—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola arvensis subsp. megalantha Nauenb.—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola asterias Hook. & Arn.—synonym of V. pusilla (sect. Subandinium)
- Viola asterias var. atacamensis Phil.—synonym of V. pusilla (sect. Subandinium)
- Viola asterias var. caulescens Phil.—synonym of V. pusilla (sect. Subandinium)
- Viola asterias var. depauperata Phil.—synonym of V. pusilla (sect. Subandinium)
- Viola asterias var. genuina Reiche—synonym of V. pusilla (sect. Subandinium)
- Viola asterias var. glabra Phil. ex Reiche—synonym of V. pusilla (sect. Subandinium)
- Viola athois W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola atlantica Britton—synonym of V. brittoniana (sect. Nosphinium, subsect. Borealiamericanae)
- Viola atriplicifolia Greene—synonym of V. purpurea subsp. atriplicifolia (sect. Chamaemelanium)
- Viola atropurpurea Leyb.—(sect. Sempervivum)
- Viola atroviolacea—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola aurantiaca Leyb.—(sect. Rosulatae)
- Viola aurata Phil.—(sect. Subandinium)
- Viola aurea Kellogg—(sect. Chamaemelanium)
- Viola aurea Kellogg subsp. aurea—(sect. Chamaemelanium)
- Viola aurea subsp. arizonensis M. S. Baker—(sect. Chamaemelanium)
- Viola aurea subsp. mohavensis M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola aurea var. venosa (S. Watson) S. Watson—synonym of V. purpurea subsp. venosa (sect. Chamaemelanium)
- Viola auricolor Skottsb.—(sect. Sempervivum)
- Viola auricula Leyb.—(sect. Subandinium)
- Viola auritella W. Becker—synonym of V. sacculus (sect. Sempervivum)
- Viola austiniae Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola austrosinensis Y. S. Chen & Q. E. Yang—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola austroussuriensis (W. Becker) Kom.—synonym of V. acuminata subsp. grandistipulata (sect. Viola, subsect. Rostratae)
- Viola avatschensis—synonym of V. crassa subsp. avatschensis (sect. Chamaemelanium)
- Viola awagatakensis T. Yamaz., I. Ito & Ageishi—synonym of V. rostrata (sect. Viola, subsect. Rostratae)
- Viola babunensis Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola bachtschisaraensis Vl. V. Nikitin—hybrid (V. alba × V. caspia; sect. Viola)
- Viola baicalensis W. Becker—synonym of V. variegata subsp. primorskajensis (sect. Plagiostigma, subsect. Patellares)
- Viola bakeri Greene—(sect. Chamaemelanium)
- Viola balansae Gagnep.—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola balcanica Delip.—synonym of V. dacica (sect. Melanium, subsect. Bracteolatae)
- Viola balsaminoides Gardn.—synonym of V. cerasifolia (sect. Leptidium)
- Viola baltica W. Becker—hybrid (V. canina × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola bambusetorum Hand.-Mazz.—(sect. Plagiostigma, subsect. Patellares)
- Viola bangiana W. Becker—synonym of V. boliviana (sect. Leptidium)
- Viola bangii Rusby—(sect. Sempervivum)
- Viola banksii K. R. Thiele & Prober—(sect. Erpetion)
- Viola baoshanensis W. S. Shu, W. Liu & C. Y. Lan—(sect. Plagiostigma, subsect. Patellares)
- Viola barhalensis G. Knoche & Marcussen—(sect. Viola, subsect. Viola)
- Viola barkalovii Bezd.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola barroetana W. Schaffn. ex Hemsl.—(sect. Chamaemelanium)
- Viola bavarica Schrank—hybrid (V. reichenbachiana × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola baxteri House—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola beamanii Calderón—(sect. Nosphinium, subsect. Mexicanae)
- Viola beati J. M. Watson & A. R. Flores—(sect. Xylobasis)
- Viola beckeriana J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola beckiana Beck subsp. beckiana—synonym of V. beckiana (sect. Melanium, subsect. Bracteolatae)
- Viola beckiana Fiala ex Beck—(sect. Melanium, subsect. Bracteolatae)
- Viola beckiana subsp. pascua (W. Becker) Trinajstić—synonym of V. eximia subsp. eximia (sect. Melanium, subsect. Bracteolatae)
- Viola beckwithii Torr. & A. Gray—(sect. Chamaemelanium)
- Viola beckwithii Torr. & A. Gray var. beckwithii—(sect. Chamaemelanium)
- Viola beckwithii var. cachensis C. P. Sm.—(sect. Chamaemelanium)
- Viola beckwithii var. trinervata Howell—synonym of V. trinervata (sect. Chamaemelanium)
- Viola begoniifolia Benth.—synonym of V. stipularis (sect. Leptidium)
- Viola behboudiana Rech. f. & Esfand.—(sect. Sclerosium)
- Viola bellidifolia Greene—synonym of V. adunca subsp. bellidifolia (sect. Viola, subsect. Rostratae)
- Viola belophylla H. Boissieu—(sect. Plagiostigma, subsect. Patellares)
- Viola bernardii (Greene) Greene—hybrid (V. pedatifida × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola bertolonii Pio—(sect. Melanium, subsect. Bracteolatae)
- Viola bertolonii Pio subsp. bertolonii—synonym of V. bertolonii (sect. Melanium, subsect. Bracteolatae)
- Viola bertolonii subsp. messanensis (W. Becker) A. Schmidt—synonym of V. aethnensis subsp. messanensis (sect. Melanium, subsect. Bracteolatae)
- Viola bethkeana Borbás—hybrid (V. reichenbachiana × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola betonicifolia f. pubescens H. Hara—synonym of V. trichopetala (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia Sm.—(sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia Sm. subsp. betonicifolia—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia subsp. australis W. Becker—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia subsp. dielsiana W. Becker—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia subsp. jaunsariensis (W. Becker) H. Hara—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia subsp. nepalensis (Ging.) W. Becker—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia subsp. novaguineensis D. M. Moore—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola betonicifolia var. oblongosagittata (Nakai) F. Maek. & T. Hashimoto—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola bezdelevae Vorosch.—synonym of V. kitamiana (sect. Chamaemelanium)
- Viola bhutanica H. Hara—(sect. Plagiostigma, subsect. Patellares)
- Viola bicolor Pursh—synonym of V. rafinesquei (sect. Melanium, subsect. Cleistogamae)
- Viola bicolor Reiche—synonym of V. montagnei (sect. Rosulatae)
- Viola biflora L.—(sect. Chamaemelanium)
- Viola biflora L. var. biflora—synonym of V. biflora (sect. Chamaemelanium)
- Viola biflora subsp. carlottae Calder & Roy L. Taylor—synonym of V. crassa (sect. Chamaemelanium)
- Viola biflora var. acuminata Maxim.—synonym of V. biflora (sect. Chamaemelanium)
- Viola biflora var. acutifolia H. Boissieu—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola biflora var. acutifolia Kar. & Kir.—synonym of V. acutifolia (sect. Chamaemelanium)
- Viola biflora var. carlottae (Calder & Roy L. Taylor) B. Boivin—synonym of V. crassa (sect. Chamaemelanium)
- Viola biflora var. ciliicalyx H. Boissieu—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola biflora var. hirsuta W. Becker—synonym of V. biflora (sect. Chamaemelanium)
- Viola biflora var. nudicaulis W. Becker—synonym of V. biflora (sect. Chamaemelanium)
- Viola biflora var. platyphylla Franch.—synonym of V. confertifolia (sect. Chamaemelanium)
- Viola biflora var. rockiana (W. Becker) Y. S. Chen—synonym of V. rockiana (sect. Chamaemelanium)
- Viola biflora var. valdepilosa Hand.-Mazz.—synonym of V. biflora (sect. Chamaemelanium)
- Viola binayensis Okamoto & K. Ueda—(sect. Plagiostigma, subsect. Stolonosae)
- Viola binchuanensis S. H. Huang—synonym of V. grandisepala (sect. Plagiostigma, subsect. Stolonosae)
- Viola bissellii House—hybrid (V. communis × V. cucullata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola bissetii Maxim.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola blanda subsp. macloskeyi (F. E. Lloyd) A. E. Murray—synonym of V. macloskeyi (sect. Plagiostigma, subsect. Stolonosae)
- Viola blanda var. macloskeyi (F. E. Lloyd) Jeps.—synonym of V. macloskeyi (sect. Plagiostigma, subsect. Stolonosae)
- Viola blanda var. renifolia (A. Gray) A. Gray—synonym of V. renifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola blanda Willd.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola blandiformis Nakai—synonym of V. brachyceras (sect. Plagiostigma, subsect. Stolonosae)
- Viola blaxlandiae J. M. Watson & A. R. Flores—hybrid (V. cotyledon × V. pachysoma; sect. Sempervivum)
- Viola bocquetiana Yıld.—(sect. Viola, subsect. Viola)
- Viola boissieuana Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola boissieui H. Lév. & Maire—synonym of V. delavayi (sect. Chamaemelanium)
- Viola bolivari Pau—synonym of V. pachyrrhiza (sect. Spathulidium)
- Viola boliviana Britton—(sect. Leptidium)
- Viola boliviana W. Becker, non Britton—synonym of V. steinbachii (sect. Leptidium)
- Viola bonnevillensis Cottam—hybrid (V. beckwithii × V. utahensis; sect. Chamaemelanium)
- Viola borchersii Phil.—synonym of V. frigida (sect. Rosulatae)
- Viola borealis Weinm.—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola bornmuelleri Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola borussica (Borbás) W. Becker—hybrid (V. canina × V. reichenbachiana; sect. Viola, subsect. Rostratae)
- Viola brachyantha Stapf—synonym of V. kitaibeliana (sect. Melanium, subsect. Bracteolatae)
- Viola brachycentra Hayata—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola brachyceras Turcz.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola brachypetala Gay—synonym of V. huidobrii (sect. Viola, subsect. Rostratae)
- Viola brachyphylla W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola brachysepala Maxim.—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola brainerdii Greene—synonym of V. renifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola brauniae Grover ex Cooperr.—hybrid (V. rostrata × V. striata; sect. Viola, subsect. Rostratae)
- Viola braunii Borbás—hybrid (V. canina × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola breviflora Jungsim Lee & M. Kim—(sect. Plagiostigma, subsect. Patellares)
- Viola brevipes (M. S. Baker) Marcussen, ined.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola brevistipulata (Franch. & Sav.) W. Becker—(sect. Chamaemelanium)
- Viola bridgesii Britton—(sect. Leptidium)
- Viola brittoniana Pollard—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola brittoniana var. pectinata (E. P. Bicknell) Alexander—synonym of V. pectinata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola brunneostipulosa Hand.-Mazz.—synonym of V. grandisepala (sect. Plagiostigma, subsect. Stolonosae)
- Viola bubanii Timb.-Lagr.—(sect. Melanium, subsect. Bracteolatae)
- Viola buchtieniana W. Becker—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola bulbosa Maxim.—(sect. Plagiostigma, subsect. Bulbosae)
- Viola bulbosa subsp. tuberifera (Franch.) W. Becker—synonym of V. tuberifera (sect. Plagiostigma, subsect. Bulbosae)
- Viola bulbosa var. franchetii H. Boissieu—synonym of V. tuberifera (sect. Plagiostigma, subsect. Bulbosae)
- Viola burgersdijkii Oud.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola burnatii Gremli—hybrid (V. riviniana × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola bustillosia Gay—(sect. Grandiflos)
- Viola cadevallii Pau—synonym of V. alba subsp. dehnhardtii (sect. Viola, subsect. Viola)
- Viola caesariensis House—hybrid (V. palmata var. triloba × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola caespitosa D. Don—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola calabra (A. Terracc.) Ricceri & Moraldo—synonym of V. aethnensis subsp. messanensis (sect. Melanium, subsect. Bracteolatae)
- Viola calaminaria (DC.) Lej.—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola calcarata L.—(sect. Melanium, subsect. Bracteolatae)
- Viola calcarata L. subsp. calcarata—(sect. Melanium, subsect. Bracteolatae)
- Viola calcarata subsp. cavillieri (W. Becker) Negodi—(sect. Melanium, subsect. Bracteolatae)
- Viola calcarata subsp. villarsiana (Roem. & Schult.) Merxm.—(sect. Melanium, subsect. Bracteolatae)
- Viola calcarata subsp. zoysii (Wulfen) Merxm.—(sect. Melanium, subsect. Bracteolatae)
- Viola calchaquiensis W. Becker—(sect. Rosulatae)
- Viola calcicola R. A. McCauley & H. E. Ballard—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola calderensis W. Becker—synonym of V. polypoda (sect. Subandinium)
- Viola caleyana G. Don—(sect. Plagiostigma, subsect. Bilobatae)
- Viola californica M. S. Baker—(sect. Chamaemelanium)
- Viola callosa Benth.—synonym of V. stipularis (sect. Leptidium)
- Viola cameleo H. Boissieu—(sect. Chamaemelanium)
- Viola canadensis L.—(sect. Chamaemelanium)
- Viola canadensis subsp. scopulorum (A. Gray) House—synonym of V. scopulorum (sect. Chamaemelanium)
- Viola canadensis var. rugulosa (Greene) C. L. Hitchc.—synonym of V. rugulosa (sect. Chamaemelanium)
- Viola canadensis var. rydbergii (Greene) House—synonym of V. canadensis (sect. Chamaemelanium)
- Viola canadensis var. scopulorum A. Gray—synonym of V. scopulorum (sect. Chamaemelanium)
- Viola canadensis var. sitchensis Bong. ex Ledeb.—synonym of V. glabella (sect. Chamaemelanium)
- Viola candidula Nieuwl.—synonym of V. missouriensis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola canescens f. glabrescens W. Becker—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola canescens H. Boissieu & Capit., non Wall.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola canescens subsp. lanuginosa W. Becker—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola canescens Wall.—(sect. Viola, subsect. Viola)
- Viola canina L.—(sect. Viola, subsect. Rostratae)
- Viola canina L. subsp. canina—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola canina subsp. montana auct.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola canina subsp. ruppii (All.) Schübl. & G. Martens—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola canina subsp. schultzii (Billot) Kirschl.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola canina var. adunca (Sm.) A. Gray—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola canina var. kamtschatica Ging.—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola canina var. muhlenbergii (Torr.) A. Gray—synonym of V. labradorica (sect. Rostratae)
- Viola canobarbata Leyb.—synonym of V. montagnei (sect. Rosulatae)
- Viola canobarbata var. albiflora W. Becker—synonym of V. montagnei (sect. Rosulatae)
- Viola capillaris Pers.—(sect. Rubellium)
- Viola carlesii Nakai—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola carnosula W. Becker—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola cascadensis M. S. Baker—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola caspia (Rupr.) Freyn—(sect. Viola, subsect. Rostratae)
- Viola cassinensis Strobl—synonym of V. pseudogracilis subsp. cassinensis (sect. Melanium, subsect. Bracteolatae)
- Viola castillonii (W. Becker) Xifreda & Sanso—synonym of V. flavicans (sect. Xanthidium)
- Viola catalonica W. Becker—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola caucasica (Rupr.) Kolen. ex Juz.—(sect. Chamaemelanium)
- Viola caviahuensis M. Sheader & A.-L. Sheader—synonym of V. pachysoma (sect. Sempervivum)
- Viola cavillieri W. Becker—synonym of V. calcarata subsp. cavillieri (sect. Melanium, subsect. Bracteolatae)
- Viola cazorlensis Gand.—(sect. Delphiniopsis)
- Viola celebica W. Becker—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola cenisia L.—(sect. Melanium, subsect. Bracteolatae)
- Viola cephalonica Bornm.—(sect. Melanium, subsect. Bracteolatae)
- Viola cerasifolia A. St.-Hil.—(sect. Leptidium)
- Viola cerasifolia subsp. conferta (A. St.-Hil.) W. Becker—synonym of V. cerasifolia (sect. Leptidium)
- Viola cerasifolia subsp. selloiana W. Becker—synonym of V. cerasifolia (sect. Leptidium)
- Viola cerasifolia subsp. typica W. Becker—synonym of V. cerasifolia (sect. Leptidium)
- Viola cerasifolia var. intermedia A. St.-Hil.—synonym of V. cerasifolia (sect. Leptidium)
- Viola cervatiana Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola cestrica House—hybrid (V. emarginata × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola chaerophylloides (Regel) W. Becker—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola chalcosperma Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola chamaedrys Leyb.—(sect. Rosulatae)
- Viola chamissoniana Ging.—(sect. Nosphinium, subsect. Nosphinium)
- Viola chamissoniana subsp. robusta (Hillebr.) W. L. Wagner, D. R. Herbst & Sohmer—synonym of V. robusta (sect. Nosphinium, subsect. Nosphinium)
- Viola chamissoniana subsp. tracheliifolia (Ging.) W. L. Wagner, D. R. Herbst & Sohmer—synonym of V. tracheliifolia (sect. Nosphinium, subsect. Nosphinium)
- Viola champlainensis House—hybrid (V. affinis × V. septentrionalis; sect. Nosphinium, subsect. Borealiamericanae)
- Viola changii J. S. Zhou & F. W. Xing—(sect. Plagiostigma, subsect. Diffusae)
- Viola charlestonensis M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola chassanica Kork.—synonym of V. yazawana (sect. Plagiostigma, subsect. Stolonosae)
- Viola cheeseana J. M. Watson—(sect. Grandiflos)
- Viola cheiranthifolia Bonpl.—(sect. Melanium, subsect. Bracteolatae)
- Viola chejuensis Y. N. Lee & Y. C. Oh—hybrid (V. albida (V. chaerophylloides) × V. phalacrocarpa; sect. Plagiostigma, subsect. Patellares)
- Viola chelmea Boiss.—(sect. Viola, subsect. Viola)
- Viola chelmea Boiss. & Heldr. subsp. chelmea—synonym of V. chelmea (sect. Viola, subsect. Viola)
- Viola chelmea subsp. vratnikensis Gáyer & Degen—synonym of V. vilaensis (sect. Viola, subsect. Viola)
- Viola chiapasiensis W. Becker—synonym of V. nannei (sect. Nosphinium, subsect. Mexicanae)
- Viola chillanensis Phil.—synonym of V. congesta (sect. Rosulatae)
- Viola chinensis f. alboviolacea Skvortsov—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola chinensis f. anomala Skvortsov—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola chinensis f. communis Skvortsov—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola chinensis f. dissecta Skvortsov—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola chinensis f. glabra Skvortsov—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola chinensis G. Don—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola chingiana W. Becker—synonym of V. biflora (sect. Chamaemelanium)
- Viola chrysantha Phil.—synonym of V. philippiana (sect. Rosulatae)
- Viola chrysantha Schrad. ex Rchb.—hybrid (V. calcarata × V. lutea; sect. Melanium, subsect. Bracteolatae)
- Viola cilentana Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola ciliaris Willd. ex W. Becker—synonym of V. stipularis (sect. Leptidium)
- Viola ciliata Schltdl., non Willd. ex Schult.—synonym of V. grahamii (sect. Nosphinium, subsect. Mexicanae)
- Viola cilicica Contandr. & Quézel—synonym of V. jordanii (sect. Viola, subsect. Rostratae)
- Viola cinerea Boiss.—(sect. Sclerosium)
- Viola cinerea var. erythraea Fiori—synonym of V. erythraea (sect. Sclerosium)
- Viola cinerea var. stocksii (Boiss.) W. Becker—synonym of V. stocksii (sect. Sclerosium)
- Viola clauseniana M. S. Baker—(sect. Nosphinium, subsect. Clausenianae)
- Viola cleistogamoides (L. G. Adams) Seppelt—(sect. Erpetion)
- Viola cochranei H. E. Ballard—(sect. Plagiostigma, subsect. Stolonosae)
- Viola cognata Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola collina Besser—(sect. Viola, subsect. Viola)
- Viola colombiana W. Becker—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola colubrina Wedd. ex W. Becker—synonym of V. fuscifolia (sect. Leptidium)
- Viola columbiana House—hybrid (V. affinis × V. communis; sect. Nosphinium, subsect. Borealiamericanae)
- Viola columnaris Skottsb.—(sect. Sempervivum)
- Viola comberi W. Becker—(sect. Sempervivum)
- Viola commersonii DC. ex Ging.—(sect. Chilenium)
- Viola communis Pollard—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola comollia Massara—(sect. Melanium, subsect. Bracteolatae)
- Viola concordifolia C. J. Wang—synonym of V. yunnanfuensis (sect. Plagiostigma, subsect. Patellares)
- Viola concordifolia var. hirtipedicellata Ching J. Wang—synonym of V. japonica (sect. Plagiostigma, subsect. Patellares)
- Viola conferta (W. Becker) Nakai—synonym of V. orientalis (sect. Chamaemelanium)
- Viola conferta A. St.-Hil.—synonym of V. cerasifolia (sect. Leptidium)
- Viola confertifolia C. C. Chang—(sect. Chamaemelanium)
- Viola confusa Champ. ex Benth.—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola confusa H. Boissieu & Capit., non Champ.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola congener Leconte—synonym of V. palmata var. triloba (sect. Nosphinium, subsect. Borealiamericanae)
- Viola congesta Gillies ex Hook. & Arn.—(sect. Rosulatae)
- Viola conilii Franch. & Sav.—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola conjugens Greene—hybrid (V. communis × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola consobrina House—hybrid (V. affinis × V. hirsutula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola consocia House—hybrid (V. affinis × V. cucullata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola consona House—hybrid (V. affinis × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola conspersa Rchb.—synonym of V. labradorica (sect. Rostratae)
- Viola conspersa var. masonii Farwell—synonym of V. striata (sect. Viola, subsect. Rostratae)
- Viola contempta Jord.—hybrid (V. arvensis × V. tricolor; sect. Melanium, subsect. Bracteolatae)
- Viola conturbata House—hybrid (V. cucullata × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola convicta House—hybrid (V. fimbriatula × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola cooperrideri H. E. Ballard—hybrid (V. striata × V. walteri; sect. Viola, subsect. Rostratae)
- Viola copahuensis M. Sheader & A.-L. Sheader—synonym of V. pachysoma (sect. Sempervivum)
- Viola corchorifolia Dombey ex Ging.—synonym of V. arguta (sect. Leptidium)
- Viola cordifolia (Nutt.) Schwein.—hybrid (V. hirsutula × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola cordifolia W. Becker, non Schwein.—synonym of V. yunnanfuensis (sect. Plagiostigma, subsect. Patellares)
- Viola coreana H. Boissieu—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola cornuta L.—(sect. Melanium, subsect. Bracteolatae)
- Viola coronifera var. minoriflora W. Becker—synonym of V. coronifera (sect. Sempervivum)
- Viola coronifera W. Becker—(sect. Sempervivum)
- Viola corralensis Phil.—synonym of V. rubella (sect. Rubellium)
- Viola corsica Nyman—(sect. Melanium, subsect. Bracteolatae)
- Viola corsica Nyman subsp. corsica—(sect. Melanium, subsect. Bracteolatae)
- Viola corsica subsp. ilvensis (W. Becker) Merxm.—(sect. Melanium, subsect. Bracteolatae)
- Viola corsica subsp. limbarae Merxm. & Lippert—(sect. Melanium, subsect. Bracteolatae)
- Viola cotyledon Ging.—(sect. Sempervivum)
- Viola cotyledon subsp. lologensis W. Becker—synonym of V. lologensis (sect. Sempervivum)
- Viola crassa (Makino) Makino—(sect. Chamaemelanium)
- Viola crassa (Makino) Makino subsp. crassa—(sect. Chamaemelanium)
- Viola crassa subsp. alpicola Takah.—(sect. Chamaemelanium)
- Viola crassa subsp. avatschensis (W. Becker & Hultén) Espeut—(sect. Chamaemelanium)
- Viola crassa subsp. borealis Takah.—(sect. Chamaemelanium)
- Viola crassa subsp. yatsugatakeana Takah.—(sect. Chamaemelanium)
- Viola crassa var. shikkensis Miyabe & Tatew.—synonym of V. crassa subsp. avatschensis (sect. Chamaemelanium)
- Viola crassa var. vegeta Nakai—synonym of V. biflora (sect. Chamaemelanium)
- Viola crassicalcarata Ching J. Wang—synonym of V. japonica (sect. Plagiostigma, subsect. Patellares)
- Viola crassicornis W. Becker & Hultén—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola crassifolia Fenzl—(sect. Melanium, subsect. Bracteolatae)
- Viola crassiuscula Bory—(sect. Melanium, subsect. Bracteolatae)
- Viola crassula Greene—hybrid (V. nephrophylla × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola crenulata Greene—hybrid (V. affinis × V. nephrophylla), synonym of V. venustula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola cretica Boiss. & Heldr.—synonym of V. alba subsp. cretica (sect. Viola, subsect. Viola)
- Viola crinita Wedd. ex W. Becker—synonym of V. steinbachii (sect. Leptidium)
- Viola cryana Gillot—(sect. Melanium, subsect. Bracteolatae)
- Viola cuatrecasasii L. B. Sm. & A. Fernández—synonym of V. scandens var. integristipula (sect. Leptidium)
- Viola cucullata Aiton—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola cucullata var. leptosepala (Greene) W. Stone—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola cucullata var. macrotis (Greene) W. Stone—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola cucullata var. microtitis Brainerd—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola cuicochensis Hieron.—(sect. Nosphinium, subsect. Mexicanae)
- Viola culminis F. Fen. & Moraldo—(sect. Melanium, subsect. Bracteolatae)
- Viola cummingii W. Becker—synonym of V. bridgesii (sect. Leptidium)
- Viola cuneata S. Watson—(sect. Chamaemelanium)
- Viola cunninghamii Hook. f.—(sect. Plagiostigma, subsect. Bilobatae)
- Viola curicoensis W. Becker—synonym of V. aurantiaca (sect. Rosulatae)
- Viola curtisiae (L. G. Adams) K. R. Thiele—(sect. Erpetion)
- Viola curvistylis H. Boissieu & Capit.—synonym of V. yunnanensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola cuspidifolia W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola cyathiformis W. Becker—synonym of V. aizoon (sect. Sempervivum)
- Viola czemalensis Zuev—synonym of V. macroceras (sect. Plagiostigma, subsect. Patellares)
- Viola dacica Borbás—(sect. Melanium, subsect. Bracteolatae)
- Viola dactyloides Schult.—(sect. Plagiostigma, subsect. Patellares)
- Viola dactyloides var. multipartita W. Becker—synonym of V. dactyloides (sect. Plagiostigma, subsect. Patellares)
- Viola dageletiana Nakai—synonym of V. kusanoana (sect. Viola, subsect. Rostratae)
- Viola dalatensis Gagnep.—synonym of V. hossei (sect. Plagiostigma, subsect. Stolonosae)
- Viola dandoisiorum J. M. Watson & A. R. Flores—(sect. Relictium)
- Viola danielae Pînzaru—replacement name for V. mariae Pinzaru, status uncertain; probably a synonym of V. dubiana or a hybrid (sect. Melanium, subsect. Bracteolatae)
- Viola dasyphylla W. Becker—(sect. Sempervivum)
- Viola davidii Franch.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola davidii var. paucicrenata W. Becker—synonym of V. davidii (sect. Plagiostigma, subsect. Stolonosae)
- Viola davisii House—hybrid (V. affinis × V. brittoniana; sect. Nosphinium, subsect. Borealiamericanae)
- Viola decipiens Reiche—(sect. Rosulatae)
- Viola declinata Waldst. & Kit.—(sect. Melanium, subsect. Bracteolatae)
- Viola decumbens L. f.—(sect. Melvio)
- Viola dehnhardtii Ten.—synonym of V. alba subsp. dehnhardtii (sect. Viola, subsect. Viola)
- Viola delavayi f. depauperata Diels—synonym of V. delavayi (sect. Chamaemelanium)
- Viola delavayi Franch.—(sect. Chamaemelanium)
- Viola delavayi var. villosa W. Becker—synonym of V. delavayi (sect. Chamaemelanium)
- Viola delphinantha Boiss.—(sect. Delphiniopsis)
- Viola delphiniifolia Nutt. ex Torr. & A. Gray—synonym of V. pedatifida (sect. Nosphinium, subsect. Borealiamericanae)
- Viola deltoidea Yatabe—synonym of V. raddeana (sect. Plagiostigma, subsect. Bilobatae)
- Viola demetria Prolongo ex Boiss.—(sect. Melanium, subsect. Dispares)
- Viola denizliensis O. D. Düsen, Göktürk, U. Sarpkaya & B. Gürcan—(sect. Melanium, subsect. Ebracteatae)
- Viola dentariifolia H. Boissieu—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola deseglisei Jord. ex Boreau—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola diamantiaca Nakai—(sect. Plagiostigma, subsect. Stolonosae)
- Viola diamantiaca var. glabrior Kitag.—synonym of V. diamantiaca (sect. Plagiostigma, subsect. Stolonosae)
- Viola dichotoma Ging.—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola dichroa Boiss.—(sect. Melanium, subsect. Bracteolatae)
- Viola dicksonii Greene—synonym of V. sororia (sect. Nosphinium, subsect. Borealiamericanae)
- Viola diffusa Ging.—(sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa subsp. apoensis (Elmer) D. M. Moore—synonym of V. apoensis (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa subsp. tenuis (Benth.) W. Becker—synonym of V. tenuis (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa var. brevibarbata C. J. Wang—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa var. brevisepala W. Becker—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa var. glabella H. Boissieu—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa var. glaberrima W. Becker—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusa var. tomentosa W. Becker—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola diffusoides C. J. Wang—synonym of V. diffusa (sect. Plagiostigma, subsect. Diffusae)
- Viola digitata Pursh—synonym of V. pedata subsp. pedata (sect. Nosphinium, subsect. Pedatae)
- Viola dimissa House—hybrid (V. emarginata × V. hirsutula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola dimorphophylla Y. S. Chen & Q. E. Yang—(sect. Chamaemelanium)
- Viola dirimliensis Blaxland—(sect. Melanium, subsect. Ebracteatae)
- Viola dirphya Tiniakou—(sect. Viola, subsect. Rostratae)
- Viola discors House—hybrid (V. affinis × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola discurrens Greene—synonym of V. canadensis (sect. Chamaemelanium)
- Viola disjuncta W. Becker—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola dissecta f. pubescens (Regel) Kitag.—synonym of V. dissecta (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta f. sieboldiana (Maxim.) Makino—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta Ledeb.—synonym of V. multifida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta subvar. albida (Palib.) Makino—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta var. albida (Palib.) Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta var. chaerophylloides (Regel) Makino—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta var. pubescens (Regel) Kitag.—synonym of V. dissecta (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta var. sieboldiana (Maxim.) Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissecta var. takahashii Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola dissena House—hybrid (V. affinis × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola dissita House—hybrid (V. hirsutula × V. palmata var. triloba; sect. Nosphinium, subsect. Borealiamericanae)
- Viola distans Wall.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola diversifolia (Ging.) W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola doerfleri Degen—(sect. Melanium, subsect. Bracteolatae)
- Viola doii Taken.—hybrid (V. mandshurica × V. violacea; sect. Plagiostigma, subsect. Patellares)
- Viola dolichoceras C. J. Wang—synonym of V. pekinensis (sect. Plagiostigma, subsect. Patellares)
- Viola dombeyana DC. ex Ging.—(sect. Leptidium)
- Viola domeikoana Gay—(sect. Subandinium)
- Viola donetzkiensis Klokov—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola douglasii Steud.—(sect. Chamaemelanium)
- Viola dowelliana House—hybrid (V. affinis × V. hirsutula), synonym of V. consobrina (sect. Nosphinium, subsect. Borealiamericanae)
- Viola dubia Wiesb.—hybrid (V. reichenbachiana × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola dubyana Burnat ex Gremli—(sect. Melanium, subsect. Bracteolatae)
- Viola duclouxii W. Becker—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola dukadjinica W. Becker & Košanin—(sect. Melanium, subsect. Bracteolatae)
- Viola dumetorum Phil.—synonym of V. capillaris (sect. Rubellium)
- Viola dusenii W. Becker & Sam.—synonym of V. uleana (sect. Leptidium)
- Viola dyris Maire—(sect. Melanium, subsect. Dispares)
- Viola eamesii House—hybrid (V. brittoniana × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola ebracteolata Fenzl—synonym of V. modesta (sect. Melanium, subsect. Ebracteatae)
- Viola eclipes H. E. Ballard—hybrid (V. labradorica × V. striata; sect. Viola, subsect. Rostratae)
- Viola ecuadorensis W. Becker—synonym of V. humilis (sect. Nosphinium, subsect. Mexicanae)
- Viola edanoii W. Becker—synonym of V. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola edulis Spach—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola effusa W. Becker—synonym of V. rupicola (sect. Plagiostigma, subsect. Patellares)
- Viola egglestoniana House—hybrid (V. fimbriatula × V. latiuscula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola egglestonii Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola egregia House—hybrid (V. brittoniana × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola eizanensis (Makino) Makino—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola eizasieboldii Sugim. ex T. Shimizu—hybrid (V. albida (V. eizanensis) × V. sieboldii; sect. Plagiostigma, subsect. Patellares)
- Viola elatior Fr.—(sect. Viola, subsect. Rostratae)
- Viola elegantula Schott—(sect. Melanium, subsect. Bracteolatae)
- Viola elisabethae Klokov—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola emarginata (Nutt.) Leconte—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola emarginata var. subsinuata Greene—synonym of V. subsinuata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola emeiensis C. J. Wang—synonym of V. bambusetorum (sect. Plagiostigma, subsect. Patellares)
- Viola eminens K. R. Thiele & Prober—(sect. Erpetion)
- Viola eminii (Engl.) R. E. Fr.—(sect. Abyssinium)
- Viola eminii var. duriprati (R. E. Fr.) Stork—synonym of V. eminii (sect. Abyssinium)
- Viola emirnensis Bojer—synonym of V. abyssinica (sect. Abyssinium)
- Viola enmae P. Gonzáles—(sect. Inconspicuiflos)
- Viola epipsila Ledeb.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola epipsila var. hyperborea Rupr.—synonym of V. suecica subsp. suecica (sect. Plagiostigma, subsect. Stolonosae)
- Viola epipsila subsp. repens W. Becker—synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola epipsila var. repens (W. Becker) R. J. Little—synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola epipsiloides Á. Löve & D. Löve—synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola epirota (Halácsy) Raus—(sect. Melanium, subsect. Bracteolatae)
- Viola erdneri Gerstl.—hybrid (V. odorata × V. suavis; sect. Viola, subsect. Viola)
- Viola erectifolia A. Nelson—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola eriksoniana W. Becker—hybrid (V. riviniana × V. uliginosa; sect. Viola, subsect. Rostratae)
- Viola eriocarpa Schwein.—(sect. Chamaemelanium)
- Viola ermenekensis Yıld. & Dinç—(sect. Melanium, subsect. Ebracteatae)
- Viola erratica House—hybrid (V. emarginata × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola erythraea (Fiori) Chiov.—(sect. Sclerosium)
- Viola escarapela J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola escondidaensis W. Becker—(sect. Rhizomandinium)
- Viola esculenta Elliott ex Greene—synonym of V. edulis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola etbaica Schweinf.—(sect. Sclerosium)
- Viola etrusca Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola euboea Halácsy—(sect. Melanium, subsect. Bracteolatae)
- Viola eugeniae Parl.—(sect. Melanium, subsect. Bracteolatae)
- Viola eugeniae Parl. subsp. eugeniae—(sect. Melanium, subsect. Bracteolatae)
- Viola eugeniae subsp. levieri (Parl.) A. Schmidt—(sect. Melanium, subsect. Bracteolatae)
- Viola evae Hieron. ex W. Becker—(sect. Rosulatae)
- Viola evae var. flossdorfii (Hicken) Nicola—synonym of V. evae (sect. Rosulatae)
- Viola excerpta House—hybrid (V. baxteri × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola excisa Hance—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola exigua var. castillonii W. Becker—synonym of V. flavicans (sect. Xanthidium)
- Viola exigua W. Becker—synonym of V. flavicans (sect. Xanthidium)
- Viola exilis Phil.—(sect. Rosulatae)
- Viola eximia Formánek—(sect. Melanium, subsect. Bracteolatae)
- Viola eximia Formánek subsp. eximia—(sect. Melanium, subsect. Bracteolatae)
- Viola eximia subsp. tringiana Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola exsul J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola extremiorientalis Vorosch. & N. S. Pavlova—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola falcata Greene—synonym of V. palmata var. dilatata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola falconeri Hook. f. & Thomson—synonym of V. jordanii (sect. Viola, subsect. Rostratae)
- Viola fallacissima Greene—hybrid (V. pedatifida × V. sororia), synonym of V. bernardii (sect. Nosphinium, subsect. Borealiamericanae)
- Viola fargesii H. Boissieu—(sect. Plagiostigma, subsect. Stolonosae)
- Viola farkasiana J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola faurieana W. Becker—(sect. Viola, subsect. Rostratae)
- Viola fedtschenkoana W. Becker—synonym of V. caspia (sect. Viola, subsect. Rostratae)
- Viola fennica F. Nyl.—synonym of V. epipsila (sect. Plagiostigma, subsect. Stolonosae)
- Viola ferdinandea Ricceri & Moraldo—synonym of V. aethnensis subsp. messanensis (sect. Melanium, subsect. Bracteolatae)
- Viola fernaldii House—hybrid (V. fimbriatula × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola ferrarinii Moraldo & Ricceri—(sect. Melanium, subsect. Bracteolatae)
- Viola ferreyrae P. Gonzáles—(sect. Rosulatae)
- Viola festata House—hybrid (V. cucullata × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola fibrillosa W. Becker—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola filicaulis Hook. f.—(sect. Nematocaulon)
- Viola filicetorum Greene—hybrid (V. affinis × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola filifera Kom.—synonym of V. bulbosa (sect. Plagiostigma, subsect. Bulbosae)
- Viola fimbriata Steud.—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola fimbriatula Sm.—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola fischeri Sweet, non W. Becker—synonym of V. gmeliniana (sect. Plagiostigma, subsect. Patellares)
- Viola fischeri W. Becker—(sect. Chamaemelanium)
- Viola fissifolia Kitag.—hybrid (V. ingolensis × [unknown]; sect. Plagiostigma, subsect. Patellares)
- Viola flagelliformis Hemsl.—(sect. Chamaemelanium)
- Viola flagelliformis var. glabrescens W. Becker—synonym of V. flagelliformis (sect. Chamaemelanium)
- Viola flavicans Wedd.—(sect. Xanthidium)
- Viola flavida Bureau & Franch., non Schur—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola flaviflora Nakai—synonym of V. brevistipulata (sect. Chamaemelanium)
- Viola flavovirens Pollard—synonym of V. praemorsa subsp. flavovirens (sect. Chamaemelanium)
- Viola flettii Piper—(sect. Chamaemelanium)
- Viola floridana Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola flos-idae Hieron.—(sect. Triflabellium)
- Viola flos-idae var. pseudovolcanica Hieron.—synonym of V. flos-idae (sect. Triflabellium)
- Viola flos-mariae Hieron.—synonym of V. montagnei (sect. Rosulatae)
- Viola flos-mariae var. nivea Hieron.—synonym of V. montagnei (sect. Rosulatae)
- Viola flos-mariae var. virescens Hieron.—synonym of V. montagnei (sect. Rosulatae)
- Viola fluehmannii Phil.—(sect. Ericoidium)
- Viola foliosa Čelak.—hybrid (V. hirta × V. suavis; sect. Viola, subsect. Viola)
- Viola formosana Hayata—(sect. Plagiostigma, subsect. Formosanae)
- Viola formosana var. stenopetala (Hayata) J. C. Wang, T. C. Huang & T. Hashim.—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola formosana var. tozanensis (Hayata) C. F. Hsieh—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola forrestiana W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola forskaalii Greuter—synonym of V. stocksii (sect. Sclerosium)
- Viola fragrans Sieber—(sect. Melanium, subsect. Bracteolatae)
- Viola franchetii H. Boissieu—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola franksmithii N. H. Holmgren—(sect. Chamaemelanium)
- Viola friderici W. Becker—(sect. Rosulatae)
- Viola frigida Phil.—(sect. Rosulatae)
- Viola frigida var. borchersii (Phil.) Reiche—synonym of V. frigida (sect. Rosulatae)
- Viola frondosa (Velen.) Velen.—(sect. Melanium, subsect. Bracteolatae)
- Viola frusinatae Ricceri & Moraldo—synonym of V. cassinensis subsp. cassinensis (sect. Melanium, subsect. Bracteolatae)
- Viola fruticosa W. Becker—synonym of V. stipularis (sect. Leptidium)
- Viola fujisanensis S. Watan.—hybrid (V. albida (V. eizanensis) × V. shikokiana; sect. Plagiostigma, subsect. Patellares)
- Viola fukienensis W. Becker—synonym of V. kosanensis (sect. Viola, subsect. Rostratae)
- Viola funghuangensis P. Y. Fu & Y. C. Teng—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola fuscifolia W. Becker—(sect. Leptidium)
- Viola fuscoviolacea (L. G. Adams) T. A. James—(sect. Erpetion)
- Viola fusiformis Sm.—synonym of V. gmeliniana (sect. Plagiostigma, subsect. Patellares)
- Viola galacifolia Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola galeanaensis M. S. Baker—(sect. Chamaemelanium)
- Viola ganchouenensis—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola ganeschinii Vl. V. Nikitin—hybrid (V. mauritii × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola ganiatsasii Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola gaviolii Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola gelida J. M. Watson, M. P. Cárdenas & A. R. Flores—(sect. Rosulatae)
- Viola geminiflora Greene—synonym of V. canadensis (sect. Chamaemelanium)
- Viola genevensis W. Becker & Cheneverd—hybrid (V. canina (V. nemoralis) × V. stagnina (“V. persicifolia”); sect. Viola, subsect. Rostratae)
- Viola germainii Sparre—(sect. Chilenium)
- Viola glabella Nutt.—(sect. Chamaemelanium)
- Viola glabella var. remotifolia Suksd.—synonym of V. glabella (sect. Chamaemelanium)
- Viola glaberrima (Ging. ex Chapm.) House—(sect. Chamaemelanium)
- Viola glaberrima (Murb.) Ye. V. Serg., non (Ging.) House—synonym of V. rupestris (sect. Viola, subsect. Rostratae)
- Viola glacialis Poepp. & Endl.—synonym of V. truncata (sect. Grandiflos)
- Viola glandularis H. E. Ballard & P. Jørg.—synonym of V. stuebelii (sect. Chilenium)
- Viola glauca M. Bieb.—synonym of V. rupestris (sect. Viola, subsect. Rostratae)
- Viola glaucescens Oudem.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola glechomoides Leyb.—(sect. Rosulatae)
- Viola gmeliniana Schult.—(sect. Plagiostigma, subsect. Patellares)
- Viola gmeliniana var. albiflora W. Becker—synonym of V. gmeliniana (sect. Plagiostigma, subsect. Patellares)
- Viola gmeliniana var. glabra Ledeb.—synonym of V. gmeliniana (sect. Plagiostigma, subsect. Patellares)
- Viola gmeliniana var. hispida Ledeb.—synonym of V. gmeliniana (sect. Plagiostigma, subsect. Patellares)
- Viola godoyae Phil.—(sect. Relictium)
- Viola gomphopetala Greene—synonym of V. praemorsa var. linguifolia (sect. Chamaemelanium)
- Viola gostivariensis Bornm.—(sect. Melanium, subsect. Bracteolatae)
- Viola gotlandica W. Becker—hybrid (V. pumila × V. stagnina; sect. Viola, subsect. Rostratae)
- Viola gracilis Sm.—(sect. Melanium, subsect. Bracteolatae)
- Viola gracillima A. St.-Hil.—(sect. Leptidium)
- Viola gracillima var. incisa W. Becker—synonym of V. gracillima (sect. Leptidium)
- Viola graeca (W. Becker) Halácsy—(sect. Melanium, subsect. Bracteolatae)
- Viola grahamii Benth.—(sect. Nosphinium, subsect. Mexicanae)
- Viola grandidentata Kalela—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola grandis Greene—hybrid (V. communis × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola grandisepala W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola granulosa Wedd.—(sect. Rosulatae)
- Viola grayi Franch. & Sav.—(sect. Viola, subsect. Rostratae)
- Viola grayi var. candida H. Boissieu—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola greatrexii Nakai & F. Maek.—hybrid (V. phalacrocarpa × V. variegata; sect. Plagiostigma, subsect. Patellares)
- Viola greenei House—hybrid (V. communis × V. emarginata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola greenmanii House—hybrid (V. cucullata × V. palmata var. triloba; sect. Nosphinium, subsect. Borealiamericanae)
- Viola griffithiana Boiss.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola grisebachiana Vis.—(sect. Melanium, subsect. Bracteolatae)
- Viola grubovii Vl. V. Nikitin—hybrid (V. variegata × V. dactyloides; sect. Plagiostigma, subsect. Patellares)
- Viola grypoceras A. Gray—(sect. Viola, subsect. Rostratae)
- Viola grypoceras var. barbata W. Becker—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola grypoceras var. pubescens Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola guadalupensis A. M. Powell & Wauer—(sect. Chamaemelanium)
- Viola guangzhouensis A. Q. Dong, J. S. Zhou & F. W. Xing—(sect. Plagiostigma, subsect. Diffusae)
- Viola guatemalensis W. Becker—(sect. Nosphinium, subsect. Mexicanae)
- Viola guaxarensis M. Marrero, Docoito Díaz & Martín Esquivel—(sect. Melanium, subsect. Bracteolatae)
- Viola guestphalica Nauenb.—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola halacsyana Degen & Dörfl.—hybrid (V. allchariensis × V. arsenica; sect. Melanium, subsect. Bracteolatae)
- Viola hallii A. Gray—(sect. Chamaemelanium)
- Viola hamiltoniana D. Don—(sect. Plagiostigma, subsect. Bilobatae)
- Viola hancockii var. fangshanensis J. W. Wang—synonym of V. hancockii (sect. Plagiostigma, subsect. Patellares)
- Viola hancockii W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola harae Miyabe & Tatew.—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola hastata Michx.—(sect. Chamaemelanium)
- Viola hastata var. tripartita (Elliott) A. Gray—synonym of V. tripartita (sect. Chamaemelanium)
- Viola haynaldii Wiesb.—hybrid (V. ambigua × V. suavis; sect. Viola, subsect. Viola)
- Viola hebeiensis J. W. Wang & T. G. Ma—synonym of V. mongolica (sect. Plagiostigma, subsect. Patellares)
- Viola hederacea Labill.—(sect. Erpetion)
- Viola hederacea subsp. cleistogamoides L. G. Adams—synonym of V. cleistogamoides (sect. Erpetion)
- Viola hederacea subsp. curtisiae L. G. Adams—synonym of V. curtisiae (sect. Erpetion)
- Viola hederacea subsp. fuscoviolacea L. G. Adams—synonym of V. fuscoviolacea (sect. Erpetion)
- Viola hederacea subsp. hederacea—synonym of V. hederacea (sect. Erpetion)
- Viola hederacea subsp. perreniformis L. G. Adams—synonym of V. perreniformis (sect. Erpetion)
- Viola hederacea subsp. seppeltiana L. G. Adams—probable synonym of V. sieberiana (sect. Erpetion)
- Viola hederacea subsp. sieberiana (Spreng.) L. G. Adams—synonym of V. sieberiana (sect. Erpetion)
- Viola hederacea var. elatines DC.—synonym of V. banksii (sect. Erpetion)
- Viola hederacea var. genuina Domin.—synonym of V. hederacea (sect. Erpetion)
- Viola hediniana W. Becker—(sect. Chamaemelanium)
- Viola heldreichiana Boiss.—(sect. Melanium, subsect. Bracteolatae)
- Viola helena C. N. Forbes & Lydgate—(sect. Nosphinium, subsect. Nosphinium)
- Viola helioscopia Hillebr.—synonym of V. chamissoniana (sect. Nosphinium, subsect. Nosphninium)
- Viola hemsleyana Calderón—(sect. Nosphinium, subsect. Mexicanae)
- Viola henriquesii (Willk. ex Cout.) W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola henryi H. Boissieu—(sect. Viola, subsect. Rostratae)
- Viola herbivaga Ridl.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola herzogii (W. Becker) Bornm.—(sect. Melanium, subsect. Bracteolatae)
- Viola heterocarpa Borbás—hybrid (V. mirabilis × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola heterophylla subsp. euboa (Halácsy) W. Becker—synonym of V. euboea (sect. Melanium, subsect. Bracteolatae)
- Viola heterophylla subsp. graeca (W. Becker) W. Becker—synonym of V. graeca (sect. Melanium, subsect. Bracteolatae)
- Viola heterosepala Boiss. & Heldr.—synonym of V. aetolica (sect. Melanium, subsect. Bracteolatae)
- Viola hichitoana Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola hidakana Nakai—synonym of V. brevistipulata (sect. Chamaemelanium)
- Viola hideoi Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola hieronymi W. Becker—(sect. Sempervivum)
- Viola hillii W. Becker—(sect. Rosulatae)
- Viola himalayensis W. Becker—synonym of V. rupestris (sect. Viola, subsect. Rostratae)
- Viola hirsutula Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola hirta L.—(sect. Viola, subsect. Viola)
- Viola hirta var. collina (Besser) Regel—synonym of V. collina (sect. Viola, subsect. Viola)
- Viola hirta var. japonica Maxim.—synonym of V. hondoensis (sect. Viola, subsect. Viola)
- Viola hirtiformis Wiesb.—hybrid (V. ambigua × V. hirta; sect. Viola, subsect. Viola)
- Viola hirtipedoides W. Becker—synonym of V. hirtipes (sect. Plagiostigma, subsect. Patellares)
- Viola hirtipes S. Moore—(sect. Plagiostigma, subsect. Patellares)
- Viola hispida Lam.—(sect. Melanium, subsect. Bracteolatae)
- Viola hissarica Juz.—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola hollickii House—hybrid (V. affinis × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola holmiana House—hybrid (V. brittoniana × V. emarginata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola hondoensis W. Becker & H. Boissieu—(sect. Viola, subsect. Viola)
- Viola hookeri Franch., non Thomson ex Hook. f.—synonym of V. bulbosa (sect. Plagiostigma, subsect. Bulbosae)
- Viola hookeri Thomson ex Hook. f.—synonym of V. sikkimensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola hookeriana Kunth—(sect. Nosphinium, subsect. Mexicanae)
- Viola hosakae H. St. John—synonym of V. kauaensis var. hosakae (sect. Nosphinium, subsect. Nosphinium)
- Viola hosoii Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola hossei W. Becker—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola howellii A. Gray—(sect. Nosphinium, subsect. Langsdorffianae)
- Viola hsinganensis Taken.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola huanucoensis W. Becker—synonym of V. boliviana (sect. Leptidium)
- Viola huesoensis Martic.—(sect. Relictium)
- Viola huidobrii Gay—(sect. Viola, subsect. Rostratae)
- Viola huizhouensis Y. S. Huang & Q. Fan—(sect. Plagiostigma, subsect. Diffusae)
- Viola hultenii W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola humboldtii Triana & Planch.—synonym of V. dombeyana (sect. Leptidium)
- Viola humboldtii var. cuneata W. Becker—synonym of V. dombeyana (sect. Leptidium)
- Viola humboldtii var. renifolia Britton—synonym of V. mandonii (sect. Leptidium)
- Viola humilis Kunth—(sect. Nosphinium, subsect. Mexicanae)
- Viola hunanensis Hand.-Mazz.—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola hungarica Degen & Sabr.—hybrid (V. ambigua × V. odorata; sect. Viola, subsect. Viola)
- Viola hupeiana W. Becker—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola hybanthoides W. B. Liao & Q. Fan—(sect. Danxiaviola)
- Viola hymettia Boiss. & Heldr.—(sect. Melanium, subsect. Bracteolatae)
- Viola hyperborea (Rupr.) Vl. V. Nikitin (pro hybr.)—synonym of V. suecica subsp. suecica (sect. Plagiostigma, subsect. Stolonosae)
- Viola hypoleuca Hayata—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola hyrcanica Vl. V. Nikitin—hybrid (V. reichenbachiana × V. caspia (“V. sieheana”); sect. Viola, subsect. Rostratae)
- Viola ibukiana Makino—hybrid (V. albida (V. chaerophylloides) × V. violacea; sect. Plagiostigma, subsect. Patellares)
- Viola ignobilis Rupr.—synonym of V. odorata (sect. Viola, subsect. Viola)
- Viola igoschinae Vl. V. Nikitin—hybrid (V. canina × V. mauritii; sect. Viola, subsect. Rostratae)
- Viola ikedaeana W. Becker ex Taken.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola ilvensis (W. Becker) Arrigoni—synonym of V. corsica subsp. ilvensis (sect. Melanium, subsect. Bracteolatae)
- Viola imberbis Ledeb.—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola impatiens H. Lév.—synonym of V. delavayi (sect. Chamaemelanium)
- Viola improcera L. G. Adams—(sect. Erpetion)
- Viola improvisa Vl. V. Nikitin—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola incisa Turcz.—hybrid (V. ingolensis × [unknown]; sect. Plagiostigma, subsect. Patellares)
- Viola incissecta Vl. V. Nikitin—hybrid (V. ingolensis × [unknown] (V. incisa) × V. multifida; sect. Plagiostigma, subsect. Patellares)
- Viola incognita Brainerd—(sect. Plagiostigma, subsect. Stolonosae)
- Viola inconspicua Blume—(sect. Plagiostigma, subsect. Patellares)
- Viola inconspicua subsp. dielsiana W. Becker—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola inconspicua subsp. nagasakiensis (W. Becker) J. C. Wang & T. C. Huang—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola indica W. Becker—(sect. Viola, subsect. Viola)
- Viola indivisa Greene—hybrid (V. pedatifida × V. pratincola; sect. Nosphinium, subsect. Borealiamericanae)
- Viola ingolensis Elisafenko—(sect. Plagiostigma, subsect. Patellares)
- Viola insessa House—hybrid (V. cucullata × V. nephrophylla; sect. Nosphinium, subsect. Borealiamericanae)
- Viola insignis Pollard, non K. Richt.—synonym of V. septemloba (sect. Nosphinium, subsect. Borealiamericanae)
- Viola insolita House—hybrid (V. brittoniana × V. communis; sect. Nosphinium, subsect. Borealiamericanae)
- Viola insularis Nakai—synonym of V. kusanoana (sect. Viola, subsect. Rostratae)
- Viola integerrima Phil.—synonym of V. portalesia (sect. Rubellium)
- Viola interjecta Borbás—hybrid (V. ambigua × V. hirta; sect. Viola, subsect. Viola)
- Viola intermedia Rchb.—hybrid (V. reichenbachiana × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola interposita Kitag.—synonym of V. tenuicornis subsp. tenuicornis (sect. Plagiostigma, subsect. Patellares)
- Viola intersita Beck—hybrid (V. canina × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola ircutiana Turcz.—synonym of V. tenuicornis subsp. ircutiana (sect. Plagiostigma, subsect. Patellares)
- Viola irinae Zolot.—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola isaurica Contandr. & Quézel—(sect. Viola, subsect. Viola)
- Viola iselensis W. Becker—hybrid (V. reichenbachiana × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola ishidoyana Nakai—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola isopetala Juz.—synonym of V. caspia (sect. Viola, subsect. Rostratae)
- Viola ivonis Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola iwagawae Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola jagellonica Zapal.—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola jalapaensis W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola jangiensis W. Becker—(sect. Viola, subsect. Viola)
- Viola japonica Korth., non Langsd. ex Ging.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola japonica Langsd. ex Ging.—(sect. Plagiostigma, subsect. Patellares)
- Viola japonica var. stenopetala Franch. ex H. Boissieu—synonym of V. japonica (sect. Plagiostigma, subsect. Patellares)
- Viola jaubertiana Marès & Vigin.—(sect. Viola, subsect. Viola)
- Viola javanica W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola jeniseensis Zuev—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola jettmarii Hand.-Mazz.—hybrid (V. ingolensis × [unknown]; sect. Plagiostigma, subsect. Patellares)
- Viola jindoensis M. Kim—hybrid (V. albida (V. chaerophylloides) × V. violacea; sect. Plagiostigma, subsect. Patellares)
- Viola jinggangshanensis Z. L. Ning & J. P. Liao—(sect. Plagiostigma, subsect. Diffusae)
- Viola jizushanensis S. H. Huang—synonym of V. rockiana (sect. Chamaemelanium)
- Viola joannis-wagneri Kárpáti—hybrid (V. hirta × V. riviniana; sect. Viola)
- Viola joergensenii W. Becker—(sect. Triflabellium)
- Viola johnstonii W. Becker—(sect. Relictium)
- Viola jooi Janka—(sect. Plagiostigma, subsect. Patellares)
- Viola jordanii Hanry—(sect. Viola, subsect. Rostratae)
- Viola josephii J. M. Watson & A. R. Flores—hybrid (V. triflabellata × [unknown]; sect. Triflabellium)
- Viola jugalis Ridl.—synonym of V. sumatrana (sect. Plagiostigma, subsect. Australasiaticae)
- Viola juzepczukii Vl. V. Nikitin—hybrid (V. dissecta × V. mandshurica; sect. Plagiostigma, subsect. Patellares)
- Viola kalbreyeri W. Becker—synonym of V. humilis (sect. Nosphinium, subsect. Mexicanae)
- Viola kalksburgensis Wiesb.—hybrid (V. alba × V. suavis; sect. Viola, subsect. Viola)
- Viola kamtschadalorum W. Becker & Hultén—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola kamtschatica var. pekinensis Regel—synonym of V. pekinensis (sect. Plagiostigma, subsect. Patellares)
- Viola kamtschatika Ging.—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola kanoi Sasaki—synonym of V. biflora (sect. Chamaemelanium)
- Viola kansuensis var. oblonga W. Becker—synonym of V. pendulicarpa (sect. Viola, subsect. Rostratae)
- Viola kansuensis W. Becker—synonym of V. pendulicarpa (sect. Viola, subsect. Rostratae)
- Viola kapsanensis Nakai—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola karakalensis Klokov—synonym of V. kitaibeliana (sect. Melanium, subsect. Bracteolatae)
- Viola karakulensis Vl. V. Nikitin & O. Baranova—hybrid (V. canina × V. collina; sect. Viola)
- Viola karlreicheana Sanso, M. Seo & Xifreda—synonym of V. montagnei (sect. Rosulatae)
- Viola kashmiriana W. Becker—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola kauaensis A. Gray—(sect. Nosphinium, subsect. Nosphinium)
- Viola kauaensis A. Gray var. kauaensis—(sect. Plagiostigma, subsect. Patellares)
- Viola kauaensis var. hosakae (H. St. John) Havran & Ching Harbin—(sect. Plagiostigma, subsect. Patellares)
- Viola kawakamii Hayata—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola kawakamii var. stenopetala Hayata—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola keiskei f. hirsutior W. Becker—synonym of V. sphaerocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola keiskei Miq.—(sect. Plagiostigma, subsect. Patellares)
- Viola keiskei subsp. transmaritima W. Becker—synonym of V. pacifica (sect. Plagiostigma, subsect. Patellares)
- Viola kermesina W. Becker—(sect. Rosulatae)
- Viola kerneri Wiesb.—hybrid (V. hirta × V. suavis; sect. Viola, subsect. Viola)
- Viola kiangsiensis W. Becker—synonym of V. kosanensis (sect. Viola, subsect. Rostratae)
- Viola kireevskiana Arbuzova—fossil
- Viola kishidae Nakai—synonym of V. brevistipulata (sect. Chamaemelanium)
- Viola kisoana Nakai—hybrid (V. mandshurica × V. variegata var. nipponica; sect. Plagiostigma, subsect. Patellares)
- Viola kitaibeliana Schult.—(sect. Melanium, subsect. Bracteolatae)
- Viola kitaibeliana var. rafinesquei (Greene) Fernald—synonym of V. rafinesquei (sect. Melanium, subsect. Cleistogamae)
- Viola kitamiana Nakai—(sect. Chamaemelanium)
- Viola kizildaghensis Dinç & Yıld.—(sect. Viola, subsect. Viola)
- Viola kjellbergii Melch.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola klineana (Kupff.) W. Becker—hybrid (V. canina (V. nemoralis) × V. uliginosa; sect. Viola, subsect. Rostratae)
- Viola klingeana Ronniger—hybrid (V. canina × V. uliginosa; sect. Viola, subsect. Rostratae)
- Viola klopotovi W. Becker—hybrid (V. epipsila × V. selkirkii; sect. Plagiostigma)
- Viola klossii Ridl.—synonym of V. papuana (sect. Viola, subsect. Rostratae)
- Viola komarovii W. Becker—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola kopaonikensis Pančić ex Tomović & Niketić—(sect. Melanium, subsect. Bracteolatae)
- Viola koraiensis Nakai—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola korinchensis Ridl.—synonym of V. sumatrana (sect. Plagiostigma, subsect. Australasiaticae)
- Viola kosanensis Hayata—(sect. Viola, subsect. Rostratae)
- Viola kosaninii (Degen) Hayek—(sect. Delphiniopsis)
- Viola kouliana Bhellum & Magotra—(sect. Sclerosium)
- Viola kozo-poljanskii Grosset—hybrid (V. odorata × V. suavis; sect. Viola, subsect. Viola)
- Viola krascheninnikoviorum Vl. V. Nikitin—hybrid (V. pumila × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola krugiana W. Becker—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola kunawarensis auct.—synonym of V. kunawurensis (sect. Himalayum)
- Viola kunawarensis f. longifolia C. J. Wang—synonym of V. kunawurensis (sect. Himalayum)
- Viola kunawarensis var. angustifolia W. Becker—synonym of V. kunawurensis (sect. Himalayum)
- Viola kunawurensis Royle—(sect. Himalayum)
- Viola kupfferi Klokov—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola kupfferiana W. Becker—hybrid (V. canina × V. uliginosa; sect. Viola, subsect. Rostratae)
- Viola kurilensis Nakai—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola kusanoana Makino—(sect. Viola, subsect. Rostratae)
- Viola kusnezowiana W. Becker—(sect. Chamaemelanium)
- Viola kwangtungensis Melch.—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola labradorica Schrank—(sect. Viola, subsect. Rostratae)
- Viola laciniata (H. Boissieu) Koidz.—synonym of V. brevistipulata (sect. Chamaemelanium)
- Viola laciniosa A. Gray—synonym of V. acuminata subsp. grandistipulata (sect. Viola, subsect. Rostratae)
- Viola lacmonica Hausskn.—hybrid (V. aetolica × V. orphanidis; sect. Melanium, subsect. Bracteolatae)
- Viola lactea Sm.—(sect. Viola, subsect. Rostratae)
- Viola lactiflora Nakai—(sect. Plagiostigma, subsect. Patellares)
- Viola lagaipensis D. M. Moore—synonym of V. kjellbergii (sect. Plagiostigma, subsect. Stolonosae)
- Viola lainzii P. Monts.—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola lanaiensis W. Becker—(sect. Nosphinium, subsect. Nosphinium)
- Viola lanceolata L.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola lanceolata subsp. occidentalis (A. Gray) N. H. Russell—synonym of V. occidentalis (sect. Plagiostigma, subsect. Stolonosae)
- Viola lanceolata subsp. vittata (Greene) N. H. Russell—synonym of V. vittata (sect. Plagiostigma, subsect. Stolonosae)
- Viola lanceolata var. vittata (Greene) Weath. & Griscom—synonym of V. vittata (sect. Plagiostigma, subsect. Stolonosae)
- Viola langeana Valentine—(sect. Melanium, subsect. Bracteolatae)
- Viola langloisii Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola langsdorffii Fisch. ex Ging.—(sect. Nosphinium, subsect. Langsdorffianae)
- Viola lanifera W. Becker—(sect. Rosulatae)
- Viola laricicola Marcussen—(sect. Viola, subsect. Rostratae)
- Viola lasczinskyi (Zuev) Baikov—synonym of V. uniflora (sect. Chamaemelanium)
- Viola lasiostipes Nakai—synonym of V. muehldorfii (sect. Chamaemelanium)
- Viola latisepala Wettst.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola latistipula Hemsl.—synonym of V. flagelliformis (sect. Chamaemelanium)
- Viola latiuscula Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola lavandulacea E. P. Bicknell—hybrid (V. cucullata × V. emarginata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola lavrenkoana Klokov—synonym of V. hymettia (sect. Melanium, subsect. Bracteolatae)
- Viola lechlerii Griseb.—synonym of V. huidobrii (sect. Viola, subsect. Rostratae)
- Viola leconteana G. Don—synonym of V. blanda (sect. Plagiostigma, subsect. Stolonosae)
- Viola lehmannii subsp. cordifolia W. Becker—synonym of V. atroseminalis H. E. Ballard, ined. (sect. Leptidium)
- Viola lehmannii subsp. ovalifolia W. Becker—synonym of V. lehmanii (sect. Leptidium)
- Viola lehmannii W. Becker ex H. E. Ballard & P. Jørg.—(sect. Leptidium)
- Viola lepida Jord.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola leptosepala Greene—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola leucopetala Greene—synonym of V. labradorica (sect. Rostratae)
- Viola leveillei H. Boissieu—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola leyboldiana Phil.—(sect. Sempervivum)
- Viola lianhuashanensis C. J. Wang & K. Sun—synonym of V. bambusetorum (sect. Plagiostigma, subsect. Patellares)
- Viola liaosiensis P. Y. Fu & Y. C. Teng—synonym of V. pekinensis (sect. Plagiostigma, subsect. Patellares)
- Viola libanotica Boiss.—(sect. Viola, subsect. Viola)
- Viola lii Kitag.—synonym of V. dissecta (sect. Plagiostigma, subsect. Patellares)
- Viola lilascens Heldr. ex Boiss.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola lilliputana Iltis & H. E. Ballard—(sect. Inconspicuiflos)
- Viola lilloana W. Becker—(sect. Rosulatae)
- Viola limbarae (Merxm. & W. Lippert) Arrigoni—synonym of V. corsica subsp. limbarae (sect. Melanium, subsect. Bracteolatae)
- Viola limprichtiana W. Becker—synonym of V. lactiflora (sect. Plagiostigma, subsect. Patellares)
- Viola lindeniana Turcz.—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola linguifolia Nutt.—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola lithion N. H. Holmgren & P. K. Holmgren—(sect. Chamaemelanium)
- Viola litoralis Phil.—synonym of V. huesoensis (sect. Relictium)
- Viola litoralis Spreng.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola livonica Vl. V. Nikitin—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola llullaillacoensis W. Becker—(sect. Rosulatae)
- Viola lobata Benth.—(sect. Chamaemelanium)
- Viola lobata Benth. subsp. lobata—(sect. Chamaemelanium)
- Viola lobata subsp. integrifolia (S. Watson) R. J. Little—(sect. Chamaemelanium)
- Viola lobata subsp. psychodes (Greene) Munz—synonym of V. lobata subsp. lobata (sect. Chamaemelanium)
- Viola lobata var. integrifolia S. Watson—synonym of V. lobata subsp. integrifolia (sect. Chamaemelanium)
- Viola lologensis (W. Becker) J. M. Watson—(sect. Sempervivum)
- Viola longipedunculata Franch. & Sav.—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola longipetiolata Ricceri & Moraldo—synonym of V. eugeniae (sect. Melanium, subsect. Bracteolatae)
- Viola longistipulata Hayata—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola lovelliana Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola lucens W. Becker—(sect. Plagiostigma, subsect. Diffusae)
- Viola luciae Skottsb.—hybrid (V. chamissoniana × V. maviensis; sect. Nosphinium, subsect. Nosphinium)
- Viola lucida Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola lucidifolia Newbro—synonym of V. missouriensis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola lunata Ridl.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola lunellii Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola lungtungensis S. S. Ying—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola lutchuensis Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola lutea Huds.—(sect. Melanium, subsect. Bracteolatae)
- Viola lutea Huds. subsp. lutea—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola lutea subsp. calaminaria (DC.) Rothm.—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola lutea subsp. sudetica (Willd.) W. Becker—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola lutea var. westfalica A. A. H. Schulz—synonym of V. lutea (sect. Melanium, subsect. Bracteolatae)
- Viola lyallii Hook. f.—(sect. Plagiostigma, subsect. Bilobatae)
- Viola macedonica Boiss. & Heldr.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola macloskeyi F. E. Lloyd—(sect. Plagiostigma, subsect. Stolonosae)
- Viola macloskeyi subsp. pallens (Banks ex Ging.) M. S. Baker—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola macloskeyi var. pallens (Banks ex Ging.) C. L. Hitchc.—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola macloviana Gand.—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola macroceras Bunge—(sect. Plagiostigma, subsect. Patellares)
- Viola macrotis Greene—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola maculata Cav.—(sect. Chilenium)
- Viola maculata Cav. var. maculata—(sect. Chilenium)
- Viola maculata f. chilensis Weibel—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata f. constitucionensis Sparre—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata f. fimbriata (Steud.) Weibel—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola maculata f. grandidentata (Kalela) Weibel—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata f. munozii Sparre—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata f. septentrionalis (Kalela) Weibel—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola maculata f. typica Weibel—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata subsp. microphyllos (Poir.) Weibel—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola maculata var. buchtienii (Gand.) Weibel—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata var. megaphylla Ging.—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola maculata var. microphyllos Poir.—(sect. Chilenium)
- Viola maculata var. pubescens Reiche—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola magellanica G. Forst.—(sect. Chilenium)
- Viola magellensis Porta & Rigo ex Strobl—(sect. Melanium, subsect. Bracteolatae)
- Viola magnifica C. J. Wang & X. D. Wang—(sect. Plagiostigma, subsect. Patellares)
- Viola magniflora Molina—synonym of V. magellanica (sect. Chilenium)
- Viola mainlingensis S. Y. Chen—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola mairei H. Lév.—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola majchurensis Pissjauk.—(sect. Chamaemelanium)
- Viola makranica Omer & Qaiser—synonym of V. behboudiana (sect. Sclerosium)
- Viola malteana House—hybrid (V. labradorica × V. rostrata; sect. Viola, subsect. Rostratae)
- Viola malvina Ridl.—synonym of V. yunnanensis (sect. Plagiostigma, subsect. Diffusae)
- Viola manaslensis F. Maek.—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola mandonii W. Becker—(sect. Leptidium)
- Viola mandshurica f. albiflora P. Y. Fu & Y. C. Teng—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica f. ciliata (Nakai) F. Maek.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica f. glabra (Nakai) Hiyama ex Maek.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica f. macrantha (Maxim.) Nakai & Kitag.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica subsp. nagasakiensis W. Becker—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica var. ciliata Nakai—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica var. glabra Nakai—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola mandshurica W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola maoershanensis Y. S. Chen & Q. E. Yang—(sect. Plagiostigma, subsect. Stolonosae)
- Viola marcelorosasii J. M. Watson & A. R. Flores—(sect. Relictium)
- Viola mariae Pînzaru—synonym of V. danielae (sect. Melanium, subsect. Bracteolatae)
- Viola mariae W. Becker—(sect. Viola, subsect. Rostratae)
- Viola marihelenae Vl. V. Nikitin—hybrid (V. arvensis × V. tricolor; sect. Melanium, subsect. Bracteolatae)
- Viola maritima (Schweigg.) Tzvelev—synonym of V. tricolor (subsp. curtisii; sect. Melanium, subsect. Bracteolatae)
- Viola markgrafii W. Becker—hybrid (V. dukadjinica × V. albanica; sect. Melanium, subsect. Bracteolatae)
- Viola maroccana (Maire) Maire—synonym of V. alba subsp. dehnhardtii (sect. Viola, subsect. Viola)
- Viola martinicensis L. Hahn—synonym of V. stipularis (sect. Leptidium)
- Viola marylandica House—hybrid (V. brittoniana × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola matczkasensis Vl. V. Nikitin—hybrid (V. collina × V. mirabilis; sect. Viola)
- Viola matsudae Hayata—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola matutina Klokov—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola mauritii Tepl.—(sect. Viola, subsect. Rostratae)
- Viola maviensis H. Mann—(sect. Nosphinium, subsect. Nosphinium)
- Viola maximowicziana Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola maymanica Grey-Wilson—(sect. Spathulidium)
- Viola mccabeiana M. S. Baker—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola mearnsii Merr.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola mearnsii Standl.—synonym of V. eminii (sect. Abyssinium)
- Viola melissifolia Greene—hybrid (V. cucullata × V. septentrionalis; sect. Nosphinium, subsect. Borealiamericanae)
- Viola membranacea W. Becker—(sect. Inconspicuiflos)
- Viola menitzkii Vl. V. Nikitin—hybrid (V. arvensis × V. occulta; sect. Melanium)
- Viola mercurii Orph. ex Halácsy—(sect. Melanium, subsect. Ebracteatae)
- Viola merkensteinensis Wiesb.—hybrid (V. collina × V. odorata; sect. Viola, subsect. Viola)
- Viola merrilliana W. Becker—(sect. Plagiostigma, subsect. Bilobatae)
- Viola merxmuelleri Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola mesadensis W. Becker—(sect. Triflabellium)
- Viola methodiana Const. & Gand.—synonym of V. scorpiuroides (sect. Xylinosium)
- Viola miaolingensis Y. S. Chen—(sect. Plagiostigma, subsect. Patellares)
- Viola micrantha Turcz., non J. & C. Presl—synonym of V. acuminata subsp. acuminata (sect. Viola, subsect. Rostratae)
- Viola micranthella Wedd.—(sect. Sempervivum)
- Viola microcentra W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola microceras Rupr.—synonym of V. biflora (sect. Chamaemelanium)
- Viola microdonta Chang—synonym of V. collina (sect. Viola, subsect. Viola)
- Viola microphylla Phil.—synonym of V. philippii (sect. Rosulatae)
- Viola microphyllos f. septentrionalis Kalela—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola microphyllos Poir.—synonym of V. maculata var. microphyllos (sect. Chilenium)
- Viola miersii Bertero ex Steud.—synonym of V. pusilla (sect. Subandinium)
- Viola milanae Vl. V. Nikitin—synonym of V. dissecta (sect. Plagiostigma, subsect. Patellares)
- Viola minima Arbuzova, non M. S. Baker —fossil, in need of a replacement name
- Viola minima M. S. Baker—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola minor (Makino) Makino—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola minuscula Greene—(sect. Plagiostigma, subsect. Stolonosae)
- Viola minuta M. Bieb.—(sect. Melanium, subsect. Bracteolatae)
- Viola minuta M. Bieb. subsp. minuta—synonym of V. minuta (sect. Melanium, subsect. Bracteolatae)
- Viola minuta subsp. dagestanica (Rupr.) Vl. V. Nikitin—synonym of V. minuta (sect. Melanium, subsect. Bracteolatae)
- Viola minuta subsp. meyeriana (Rupr.) Vl. V. Nikitin—synonym of V. minuta (sect. Melanium, subsect. Bracteolatae)
- Viola minutiflora Phil.—(sect. Subandinium)
- Viola miocenica Arbuzova—fossil
- Viola mirabilis f. latisepala W. Becker—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis L.—(sect. Viola, subsect. Rostratae)
- Viola mirabilis subsp. subglabra (Ledeb.) Zuev—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis var. brachysepala (Maxim.) Regel—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis var. brevicalcarata Nakai—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis var. glaberrima W. Becker—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis var. platysepala Kitag.—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola mirabilis var. subglabra Ledeb.—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola miranda W. Becker—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola missouriensis Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola mistassinica Greene—synonym of V. renifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola mistura House—hybrid (V. sagittata × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola mixta A. Kern.—hybrid (V. canina × V. reichenbachiana; sect. Viola, subsect. Rostratae)
- Viola miyabei Makino—synonym of V. hirtipes (sect. Plagiostigma, subsect. Patellares)
- Viola miyajiana Koidz.—hybrid (V. albida (V. eizanensis) × V. yezoensis; sect. Plagiostigma, subsect. Patellares)
- Viola miyakei Nakai—synonym of V. kusanoana (sect. Viola, subsect. Rostratae)
- Viola modesta Fenzl—(sect. Melanium, subsect. Ebracteatae)
- Viola modestula Klokov—synonym of V. modesta (sect. Melanium, subsect. Ebracteatae)
- Viola modica House—hybrid (V. communis × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola molfinoana W. Becker—synonym of V. frigida (sect. Rosulatae)
- Viola molisana Ricceri & Moraldo—synonym of V. eugeniae (sect. Melanium, subsect. Bracteolatae)
- Viola mollicula House—hybrid (V. minuscula × V. primulifolia; sect. Plagiostigma, subsect. Stolonosae)
- Viola monbeigii—synonym of V. belophylla (sect. Plagiostigma, subsect. Patellares)
- Viola mongolica f. longisepala P. F. Fu & Y. C. Teng—synonym of V. pekinensis (sect. Plagiostigma, subsect. Patellares)
- Viola mongolica Franch.—(sect. Plagiostigma, subsect. Patellares)
- Viola montagnei Gay—(sect. Rosulatae)
- Viola montagnei var. glandulosa Phil.—synonym of V. montagnei (sect. Rosulatae)
- Viola montana auct., non L.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola montana L.—rejected name; synonym of V. elatior, conserved name (sect. Viola, subsect. Rostratae)
- Viola montanensis Rydb.—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola montcaunica Pau—(sect. Melanium, subsect. Bracteolatae)
- Viola montivaga House—hybrid (V. septentrionalis × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola moupinensis Franch.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola moupinensis var. lijiangensis C. J. Wang—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola mrkvickiana Velen.—synonym of V. hertzogii (sect. Melanium, subsect. Bracteolatae)
- Viola mucronulifera Hand.-Mazz.—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola muehldorfii Kiss—(sect. Chamaemelanium)
- Viola muhlenbergiana Ging.—synonym of V. labradorica (sect. Rostratae)
- Viola muhlenbergiana var. minor Hook.—synonym of V. labradorica (sect. Rostratae)
- Viola muhlenbergii Torr.—synonym of V. labradorica (sect. Rostratae)
- Viola mulfordiae Pollard—hybrid (V. brittoniana × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola muliensis Y. S. Chen & Q. E. Yang—(sect. Chamaemelanium)
- Viola multifida Willd. ex Schult.—(sect. Plagiostigma, subsect. Patellares)
- Viola multistolonifera C. J. Wang—synonym of V. bulbosa (sect. Plagiostigma, subsect. Bulbosae)
- Viola munbyana Boiss. & Reut.—(sect. Melanium, subsect. Bracteolatae)
- Viola munozensis W. Becker—synonym of V. rodriguezii (sect. Rosulatae)
- Viola muriculata Greene—synonym of V. canadensis (sect. Chamaemelanium)
- Viola murronensis Ricceri & Moraldo—synonym of V. eugeniae (sect. Melanium, subsect. Bracteolatae)
- Viola muscoides Phil.—synonym of V. tridentata (sect. Tridens)
- Viola mutsuensis W. Becker—synonym of V. kusanoana (sect. Viola, subsect. Rostratae)
- Viola nagamiana T. Hashimoto—synonym of V. kosanensis (sect. Viola, subsect. Rostratae)
- Viola nagasawae Makino & Hayata—(sect. Plagiostigma, subsect. Diffusae)
- Viola nagasawae var. acutilabella (Hayata) Nakai—synonym of V. nagasawae (sect. Plagiostigma, subsect. Diffusae)
- Viola najadum Wein—hybrid (V. riviniana × V. stagnina; sect. Viola, subsect. Rostratae)
- Viola nana (DC. ex Ging.) Le Jol.—(sect. Melanium, subsect. Bracteolatae)
- Viola nanlingensis J. S. Zhou & F. W. Xing—(sect. Plagiostigma, subsect. Diffusae)
- Viola nannae R. E. Fr.—(sect. Abyssinium)
- Viola nannei Pol.—(sect. Nosphinium, subsect. Mexicanae)
- Viola nannei var. glaberrima W. Becker—synonym of V. nannei (sect. Nosphinium, subsect. Mexicanae)
- Viola nantouensis S. S. Ying—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola napae House—hybrid (V. communis × V. sororia), synonym of V. grandis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola napellifolia Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola napellifolia var. sieboldiana (Maxim.) Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola nassauvioides Phil.—(sect. Confertae)
- Viola nebrodensis C. Presl—(sect. Melanium, subsect. Bracteolatae)
- Viola neglecta F. W. Schmidt—hybrid (V. canina (V. nemoralis) × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola neglecta M. Bieb.—synonym of V. caspia (sect. Viola, subsect. Rostratae)
- Viola neglectiformis Vl. V. Nikitin—hybrid (V. canina × V. tanaitica; sect. Viola, subsect. Rostratate)
- Viola nelsonii W. Becker—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola nemoralis Kütz.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola neogenica Mai & Walther—fossil
- Viola neomexicana Greene—synonym of V. canadensis (sect. Chamaemelanium)
- Viola nephrophylla Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola nephrophylla var. arizonica (Greene) Kearney & Peebles—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola nephrophylla var. cognata (Greene) C. L. Hitchc.—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola niederleinii W. Becker—(sect. Rosulatae)
- Viola niijimensis Nakai—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola nikitinii Vasjukov—hybrid (V. mirabilis × V. sacchalinensis; sect. Viola, subsect. Rostratae)
- Viola nikkoensis Nakai—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola nipponica Makino—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola nitida Y. S. Chen & Q. E. Yang—(sect. Plagiostigma, subsect. Stolonosae)
- Viola nivalis Benth.—synonym of V. bangii (sect. Sempervivum)
- Viola nobilis W. Becker—(sect. Sempervivum)
- Viola norvegica Wittr.—hybrid (V. arvensis × V. tricolor; sect. Melanium, subsect. Bracteolatae)
- Viola notabilis E. P. Bicknell—hybrid (V. brittoniana × V. cucullata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola novae-angliae House—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola nubigena Leyb.—(sect. Subandinium)
- Viola nuda W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola nudicaulis (W. Becker) S. Y. Chen—synonym of V. biflora (sect. Chamaemelanium)
- Viola nuevoleonensis W. Becker—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola nujiangensis Y. S. Chen & X. H. Jin—(sect. Plagiostigma, subsect. Patellares)
- Viola nummulariifolia All.—isonym of V. nummulariifolia Vill. (sect. Melanium, subsect. Pseudorupestres)
- Viola nummulariifolia Vill.—(sect. Melanium, subsect. Pseudorupestres)
- Viola nuttallii Pursh—(sect. Chamaemelanium)
- Viola nuttallii subsp. vallicola (A. Nelson) Roy L. Taylor & MacBryde—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Viola nuttallii var. bakeri (Greene) C. L. Hitchc.—synonym of V. bakeri (sect. Chamaemelanium)
- Viola nuttallii var. linguifolia (Nutt.) Piper—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola nuttallii var. vallicola (A. Nelson) H. St. John—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Viola nuttallii var. venosa S. Watson—synonym of V. purpurea subsp. venosa (sect. Chamaemelanium)
- Viola oahuensis C. N. Forbes—(sect. Nosphinium, subsect. Nosphinium)
- Viola obituaria J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola obliqua Aiton—to be rejected as a nomen ambiguum, identity cannot be determined (sect. Nosphinium, subsect. Borealiamericanae)
- Viola obliquifolia Turcz.—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola oblonga Blatt.—probable syn. of V. jordanii (sect. Rostratae)
- Viola oblongosagittata f. ishizakii Yamam.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola oblongosagittata Nakai—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola oblongosagittata var. violascens Nakai—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola obtusa (Makino) Makino—(sect. Viola, subsect. Rostratae)
- Viola obtusoacuminata T. Hashim. ex T. Shimizu—hybrid (V. acuminata × V. obtusa; sect. Viola, subsect. Rostratae)
- Viola obtusogrypoceras Makino—hybrid (V. grypoceras × V. obtusa; sect. Viola, subsect. Rostratae)
- Viola occidentalis (A. Gray) Howell—(sect. Plagiostigma, subsect. Stolonosae)
- Viola occulta Lehm.—(sect. Melanium, subsect. Ebracteatae)
- Viola ocellata Torr. & A. Gray—(sect. Chamaemelanium)
- Viola odontocalycina Boiss.—(sect. Melanium, subsect. Bracteolatae)
- Viola odorata L.—(sect. Viola, subsect. Viola)
- Viola okinawensis K. Nakaj., nom. nud.—synonym of V. utchinensis (sect. Viola, subsect. Rostratae)
- Viola okuharae F. Maek. ex T. Shimizu—hybrid (V. hirtipes × V. violacea var. makinoi; sect. Plagiostigma, subsect. Patellares)
- Viola oldhamiana Nakai—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola oligoceps Chang—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola oligyrtia Tiniakou—(sect. Viola, subsect. Rostratae)
- Viola olimpia Beggiat.—hybrid (V. odorata × V. reichenbachiana; sect. Viola)
- Viola orbelica Pančić—(sect. Melanium, subsect. Bracteolatae)
- Viola orbiculata Geyer ex Holz.—(sect. Chamaemelanium)
- Viola orbignyana J. Rémy—synonym of V. pygmaea (sect. Sempervivum)
- Viola oreades M. Bieb.—(sect. Melanium, subsect. Bracteolatae)
- Viola oreocallis Greene—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola orientalis (Maxim.) W. Becker—(sect. Chamaemelanium)
- Viola orientalis (Maxim.) W. Becker subsp. orientalis—(sect. Chamaemelanium)
- Viola orientalis subsp. xanthopetala (Nakai) Espeut—(sect. Chamaemelanium)
- Viola orientalis var. conferta W. Becker—synonym of V. orientalis subsp. orientalis (sect. Chamaemelanium)
- Viola orophila Wiesb.—hybrid (V. mirabilis × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola orphanidis Boiss.—(sect. Melanium, subsect. Bracteolatae)
- Viola orphanidis Boiss. subsp. orphanidis—(sect. Melanium, subsect. Bracteolatae)
- Viola orphanidis subsp. crinita Delip.—(sect. Melanium, subsect. Bracteolatae)
- Viola orphanidis subsp. nicolai (Pant.) Valentine—synonym of V. orphanidis subsp. orphanidis (sect. Melanium, subsect. Bracteolatae)
- Viola orthoceras Ledeb.—(sect. Melanium, subsect. Bracteolatae)
- Viola ovalifolia W. Becker—synonym of V. yunnanensis (sect. Plagiostigma, subsect. Diffusae)
- Viola ovalleana Phil.—(sect. Relictium)
- Viola ovata Nutt.—synonym of V. fimbriatula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola ovata var. hicksii (Pollard) Pollard—synonym of V. fimbriatula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola ovato-oblonga (Miq.) Makino—(sect. Viola, subsect. Rostratae)
- Viola oxyceras (S. Watson) Greene—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola oxyodontis H. E. Ballard—(sect. Nosphinium, subsect. Mexicanae)
- Viola pachyrrhiza Boiss. & Hohen.—(sect. Spathulidium)
- Viola pachysoma M. Sheader & J. M. Watson—(sect. Sempervivum)
- Viola pacifica Juz.—(sect. Plagiostigma, subsect. Patellares)
- Viola painteri Rose & House—(sect. Chamaemelanium)
- Viola palatina Y. N. Lee—hybrid (V. albida (V. chaerophylloides) × V. japonica; sect. Plagiostigma, subsect. Patellares)
- Viola palentina Losa—synonym of V. bubanii (sect. Melanium, subsect. Bracteolatae)
- Viola pallascaensis W. Becker—(sect. Xanthidium)
- Viola pallens (Banks ex Ging.) Brainerd—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola pallens subsp. macloskeyi (F. E. Lloyd) M. S. Baker—synonym of V. macloskeyi (sect. Plagiostigma, subsect. Stolonosae)
- Viola pallens var. subreptans J. Rousseau—synonym of V. minuscula (sect. Plagiostigma, subsect. Stolonosae)
- Viola palmaris Buch.-Ham. ex Ging.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola palmata L.—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmata L. var. palmata—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmata var. heterophylla Elliott—synonym of V. edulis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmata var. pedatifida (G. Don) Cronquist—synonym of V. pedatifida (sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmata var. sororia (Willd.) Pollard—synonym of V. sororia (sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmata var. triloba (Schwein.) Ging.—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola palmensis (Webb & Berthel.) Sauer—(sect. Melanium, subsect. Bracteolatae)
- Viola palustris L.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola palustris L. subsp. palustris—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola palustris subsp. brevipes M. S. Baker—synonym of V. brevipes (sect. Plagiostigma, subsect. Stolonosae)
- Viola palustris subsp. juressii (Link ex K. Wein) Coutinho—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola palustris subsp. pubifolia Kuta—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola palustris var. moupinensis (Franch.) Franch.—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola palustroides (W. Becker) Tzvelev—synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola papilionacea Pursh—misapplied, type is V. affinis [needs rejection proposal] (sect. Nosphinium, subsect. Borealiamericanae)
- Viola papillaris Willd.—synonym of V. spathulata (sect. Spathulidium)
- Viola papuana W. Becker & Pulle—(sect. Viola, subsect. Rostratae)
- Viola paradoxa Lowe—(sect. Melanium, subsect. Bracteolatae)
- Viola paravaginata H. Hara—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola parca House—hybrid (V. fimbriatula × V. septentrionalis; sect. Nosphinium, subsect. Borealiamericanae)
- Viola parnonia Kit Tan, Sfikas & Vold—(sect. Melanium, subsect. Bracteolatae)
- Viola parviflora Mutis ex L. f.—synonym of Pombalia parviflora (Mutis ex L. fil.) Paula-Souza (Violaceae)
- Viola parvifolia Benth.—synonym of V. polycephala (sect. Sempervivum)
- Viola parvula Tineo—(sect. Melanium, subsect. Ebracteatae)
- Viola pascua W. Becker—synonym of V. eximia subsp. eximia (sect. Melanium, subsect. Bracteolatae)
- Viola patagonica W. Becker—synonym of V. sacculus (sect. Sempervivum)
- Viola patrinii f. glabra (Nakai) F. Maek.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii f. hispida W. Becker p. p.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii Ging.—(sect. Plagiostigma, subsect. Patellares)
- Viola patrinii H. Boissieu & Capit., non DC.—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii Merr., non DC.—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. brevicalcarata Skvortsov—synonym of V. patrinii (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. caespitosa (D. Don) Ridl.—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. chinensis Ging.—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. laotiana H. Boissieu—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. macrantha Maxim.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. minor Makino—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola patrinii var. nepaulensis Ging.—synonym of V. betonicifolia (sect. Plagiostigma, subsect. Patellares)
- Viola peckiana House—hybrid (V. sororia × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola pectinata E. P. Bicknell—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola pedata L.—(sect. Nosphinium, subsect. Pedatae)
- Viola pedata L. subsp. pedata—(sect. Nosphinium, subsect. Pedatae)
- Viola pedata subsp. cuneatiloba (Brainerd ex Fernald) H. E. Ballard, ined.—(sect. Nosphinium, subsect. Pedatae)
- Viola pedata subsp. flabellata (D. Don) H. E. Ballard, ined.—(sect. Nosphinium, subsect. Pedatae)
- Viola pedata var. atropurpurea Ging.—synonym of V. pedata subsp. pedata (sect. Nosphinium, subsect. Pedatae)
- Viola pedatifida G. Don—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola pedatifida subsp. brittoniana (Pollard) L. E. McKinney—synonym of V. brittoniana (sect. Nosphinium, subsect. Borealiamericanae)
- Viola pedatifida var. brittoniana (Pollard) R. J. Little & L. E. McKinney—synonym of V. brittoniana (sect. Nosphinium, subsect. Borealiamericanae)
- Viola pedunculata subsp. tenuifolia M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola pedunculata Torr. & A. Gray—(sect. Chamaemelanium)
- Viola pedunculata Torr. & A. Gray subsp. pedunculata—(sect. Chamaemelanium)
- Viola pekinensis (Regel) W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola pendulicarpa W. Becker—(sect. Viola, subsect. Rostratae)
- Viola pensylvanica Michx.—synonym of V. pubescens (sect. Chamaemelanium)
- Viola pentadactyla Fenzl—(sect. Melanium, subsect. Ebracteatae)
- Viola pentelica Vierh.—synonym of V. alba subsp. dehnhardtii (sect. Viola, subsect. Viola)
- Viola peramoena Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola perinensis W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola permixta Jord.—hybrid (V. hirta × V. odorata; sect. Viola, subsect. Viola)
- Viola perpensa Greene—hybrid (V. pedatifida × V. sororia), synonym of V. bernardii (sect. Nosphinium, subsect. Borealiamericanae)
- Viola perpera House—hybrid (V. hirsutula × V. sororia), synonym of V. cordifolia (sect. Nosphinium, subsect. Borealiamericanae)
- Viola perplexa Gremli—hybrid (V. mirabilis × V. reichenbachiana; sect. Viola, subsect. Rostratae)
- Viola perpusilla H. Boissieu—(sect. Plagiostigma, subsect. Patellares)
- Viola perreniformis (L. G. Adams) R. J. Little & Leiper—(sect. Erpetion)
- Viola persicariifolia Poir.—synonym of V. stipularis (sect. Leptidium)
- Viola persicifolia Schreb.—synonym of V. elatior/stagnina (sensu Nikitin 2006; sect. Viola, subsect. Rostratae)
- Viola petelotii W. Becker ex Gagnep.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola petraea f. albida W. Becker—synonym of V. petraea (sect. Sempervivum)
- Viola petraea W. Becker—(sect. Sempervivum)
- Viola phalacrocarpa Maxim.—(sect. Plagiostigma, subsect. Patellares)
- Viola phalacrocarpoides Makino—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola philippiana Greene—(sect. Rosulatae)
- Viola philippica Cav.—(sect. Plagiostigma, subsect. Patellares)
- Viola philippica f. candida (Kitag.) Kitag.—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola philippica f. intermedia (Kitag.) Kitag.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola philippica subsp. malesica W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola philippica subsp. munda W. Becker—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola philippica subsp. philippica—(sect. Plagiostigma, subsect. Patellares)
- Viola philippica var. yunnanfuensis W. Becker—synonym of V. trichopetala (sect. Plagiostigma, subsect. Patellares)
- Viola philippii Leyb.—(sect. Rosulatae)
- Viola philippii var. arbuscula (Phil.) Reiche—synonym of V. philippii (sect. Rosulatae)
- Viola phitosiana Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola physalodes Greene—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Viola pichinchensis Turcz.—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola pilosa Blume—(sect. Viola, subsect. Viola)
- Viola pinetorum Greene—(sect. Chamaemelanium)
- Viola pinetorum Greene subsp. pinetorum—(sect. Chamaemelanium)
- Viola pinetorum subsp. grisea (Jeps.) R. J. Little—(sect. Chamaemelanium)
- Viola pinetorum var. grisea (Jeps.) R. J. Little—synonym of V. pinetorum subsp. grisea (sect. Chamaemelanium)
- Viola pinnata L.—(sect. Plagiostigma, subsect. Patellares)
- Viola pinnata subsp. multifida W. Becker—synonym of V. multifida (sect. Plagiostigma, subsect. Patellares)
- Viola pinnata var. angustisecta W. Becker—synonym of V. multifida (sect. Plagiostigma, subsect. Patellares)
- Viola pinnata var. chaerophylloides Regel—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola pinnata var. dissecta (Ledeb.) Regel—synonym of V. multifida (sect. Plagiostigma, subsect. Patellares)
- Viola pinnata var. sieboldiana Maxim.—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola pitouchaoensis S. S. Ying—synonym of V. confusa (sect. Plagiostigma, subsect. Patellares)
- Viola placida W. Becker—unresolved: protologue incomplete; type not in B (destroyed?). (sect. Plagiostigma)
- Viola pluricaulis Borbás—hybrid (V. alba × V. odorata; sect. Viola, subsect. Viola)
- Viola pluviae Marcussen, H. E. Ballard & Blaxland—(sect. Plagiostigma, subsect. Stolonosae)
- Viola pobedimovae Ye. V. Serg.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola poelliana Murr—hybrid (V. collina × V. hirta × V. odorata; sect. Viola, subsect. Viola)
- Viola poetica Boiss. & Spruner—(sect. Melanium, subsect. Dispares)
- Viola pogonantha W. W. Smith—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola poltavensis Vl. V. Nikitin—hybrid (V. elatior × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola polycephala H. E. Ballard & P. Jørg.—(sect. Sempervivum)
- Viola polymorpha C. C. Chang—synonym of V. pendulicarpa (sect. Viola, subsect. Rostratae)
- Viola polyodonta W. Becker—synonym of V. dacica (sect. Melanium, subsect. Bracteolatae)
- Viola polypoda Turcz.—(sect. Subandinium)
- Viola pontica W. Becker—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola popovae Vl. V. Nikitin—hybrid (V. canina (V. nemoralis subsp. nemoralis) × V. caspia (“V. sieheana”); sect. Viola, subsect. Rostratae)
- Viola populifolia Greene—hybrid (V. palmata var. triloba × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola porphyrea R. Uechtr.—hybrid (synonym of V. collina; sect. Viola, subsect. Viola)
- Viola portalesia Gay—(sect. Rubellium)
- Viola porteriana Pollard—hybrid (V. cucullata × V. fimbriatula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola portulacea Leyb.—(sect. Sempervivum)
- Viola praemorsa Douglas—(sect. Chamaemelanium)
- Viola praemorsa Douglas subsp. praemorsa—(sect. Chamaemelanium)
- Viola praemorsa subsp. arida M. S. Baker—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola praemorsa subsp. flavovirens (Pollard) Fabijan—(sect. Chamaemelanium)
- Viola praemorsa subsp. linguifolia (Nutt. ex Torr. & A. Gray) M. S. Baker—(sect. Chamaemelanium)
- Viola praemorsa subsp. major (Hook.) M. S. Baker—synonym of V. vallicola var. major (sect. Chamaemelanium)
- Viola praemorsa subsp. oregona M. S. Baker—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola praemorsa var. altior Blankinship—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola praemorsa var. flavovirens (Pollard) R. J. Little—synonym of V. praemorsa subsp. flavovirens (sect. Chamaemelanium)
- Viola praemorsa var. linguifolia (Nutt.) M. Peck—synonym of V. praemorsa subsp. linguifolia (sect. Chamaemelanium)
- Viola praemorsa var. major (Hook.) M. Peck—synonym of V. vallicola var. major (sect. Chamaemelanium)
- Viola pratincola Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola prattii W. Becker—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola prenja Beck—synonym of V. pyrenaica (sect. Viola, subsect. Viola)
- Viola priceana Pollard—synonym of V. communis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola pricei W. Becker—(sect. Plagiostigma, subsect. Diffusae)
- Viola primorskajensis (W. Becker) Vorosch.—synonym of V. variegata subsp. primorskajensis (sect. Plagiostigma, subsect. Patellares)
- Viola primulifolia L.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola primulifolia Loureiro, non L.—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola primulifolia subsp. occidentalis (A. Gray) L. E. McKinney & R. J. Little—synonym of V. occidentalis (sect. Plagiostigma, subsect. Stolonosae)
- Viola primulifolia subsp. villosa (Eaton) N. H. Russell—synonym of V. primulifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola primulifolia var. acuta (Bigelow) Torr. & A. Gray—synonym of V. primulifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola primulifolia var. glabra Nakai—synonym of V. patrinii (sect. Plagiostigma, subsect. Patellares)
- Viola primulifolia var. villosa Eaton—synonym of V. primulifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola principis H. Boissieu—(sect. Plagiostigma, subsect. Stolonosae)
- Viola principis var. acutifolia C. J. Wang—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola prionantha Bunge—(sect. Plagiostigma, subsect. Patellares)
- Viola prionantha subsp. jaunsariensis W. Becker—synonym of V. tienschiensis (sect. Plagiostigma, subsect. Patellares)
- Viola prionantha var. incisa Kitag.—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola prionantha var. sylvatica Kitag.—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola prionantha var. trichantha C. J. Wang—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola prionosepala Greene—synonym of V. cucullata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola producta W. Becker—unresolved. Type Fiebrig 2529 in B, probably destroyed.
- Viola prunellifolia Fisch. ex Ging.—synonym of V. patrinii (sect. Plagiostigma, subsect. Patellares)
- Viola prunellifolia Kunth—synonym of V. humilis (sect. Nosphinium, subsect. Mexicanae)
- Viola pruniflora Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola psammophila Phil.—synonym of V. polypoda (sect. Subandinium)
- Viola pseudaetolica Tomović, Melovski & Niketić—(sect. Melanium, subsect. Bracteolatae)
- Viola pseudasterias Reiche—synonym of V. polypoda (sect. Subandinium)
- Viola pseudasterias var. psammophila (Phil.) Reiche—synonym of V. polypoda (sect. Subandinium)
- Viola pseudoarcuata C. C. Chang—synonym of V. pendulicarpa (sect. Viola, subsect. Rostratae)
- Viola pseudobambusetorum C. C. Chang—synonym of V. bambusetorum (sect. Plagiostigma, subsect. Patellares)
- Viola pseudogracilis (A. Terracc.) Strobl ex Degen & Dörfl.—(sect. Melanium, subsect. Bracteolatae)
- Viola pseudogracilis Strobl subsp. pseudogracilis—(sect. Melanium, subsect. Bracteolatae)
- Viola pseudogracilis subsp. cassinensis (Strobl) Merxm.—(sect. Melanium, subsect. Bracteolatae)
- Viola pseudograeca Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola pseudojaponica Nakai—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola pseudomakinoi M. Mizush. ex T. Shimizu—hybrid (V. sieboldii f. variegata × V. violacea var. makinoi; sect. Plagiostigma, subsect. Patellares)
- Viola pseudomirabilis H. J. Coste—(sect. Viola, subsect. Rostratae)
- Viola pseudomonbeigii Chang—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola pseudoprionantha W. Becker—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola pseudovulcanica W. Becker—synonym of V. volcanica (sect. Rosulatae)
- Viola pteropoda Hemsl.—synonym of V. umbraticola (sect. Viola, subsect. Rostratae)
- Viola pubescens Aiton—(sect. Chamaemelanium)
- Viola pubescens var. eriocarpa Nutt.—synonym of V. pubescens (sect. Chamaemelanium)
- Viola pubescens var. peckii House—synonym of V. pubescens (sect. Chamaemelanium)
- Viola pubescens var. scabriuscula Schwein. ex Torr. & A. Gray—synonym of V. eriocarpa (sect. Chamaemelanium)
- Viola pubifolia (Kuta) G. H. Loos—synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola pulchella Leyb. ex Reiche—synonym of V. escarapela (sect. Rosulatae)
- Viola pulla W. Becker—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola pulvinata Reiche—(sect. Subandinium)
- Viola pumila Chaix—(sect. Viola, subsect. Rostratae)
- Viola purpurea Kellogg—(sect. Chamaemelanium)
- Viola purpurea Kellogg subsp. purpurea—(sect. Chamaemelanium)
- Viola purpurea Kellogg var. purpurea—synonym of V. purpurea subsp. purpurea (sect. Chamaemelanium)
- Viola purpurea subsp. atriplicifolia (Greene) M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea subsp. aurea (Kellogg) J. C. Clausen—synonym of V. aurea subsp. aurea (sect. Chamaemelanium)
- Viola purpurea subsp. dimorpha M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea subsp. geophyta M. S. Baker—(sect. Chamaemelanium)
- Viola purpurea subsp. integrifolia M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea subsp. mesophyta M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea subsp. mohavensis (M. S. Baker & J. C. Clausen) J. C. Clausen—synonym of V. aurea subsp. mohavensis (sect. Chamaemelanium)
- Viola purpurea subsp. quercetorum (M. S. Baker & J. C. Clausen) R. J. Little—synonym of V. quercetorum (sect. Chamaemelanium)
- Viola purpurea subsp. venosa (S. Watson) M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea subsp. xerophyta M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola purpurea var. aurea (Kellogg) M. S. Baker ex Jeps.—synonym of V. aurea subsp. aurea (sect. Chamaemelanium)
- Viola purpurea var. charlestonensis (M. S. Baker & J. C. Clausen) S. L. Welsh & Reveal—synonym of V. charlestonensis (sect. Chamaemelanium)
- Viola purpurea var. dimorpha (M. S. Baker & J. C. Clausen) J. T. Howell—synonym of V. purpurea subsp. dimorpha (sect. Chamaemelanium)
- Viola purpurea var. grisea Jeps.—synonym of V. pinetorum subsp. grisea (sect. Chamaemelanium)
- Viola purpurea var. integrifolia (M. S. Baker & J. C. Clausen) J. T. Howell—synonym of V. purpurea subsp. integrifolia (sect. Chamaemelanium)
- Viola purpurea var. mesophyta (M. S. Baker & J. C. Clausen) J. T. Howell—synonym of V. purpurea subsp. mesophyta (sect. Chamaemelanium)
- Viola purpurea var. mohavensis (M. S. Baker & J. C. Clausen) J. T. Howell—synonym of V. aurea subsp. mohavensis (sect. Chamaemelanium)
- Viola purpurea var. venosa (S. Watson) Brainerd—synonym of V. purpurea subsp. venosa (sect. Chamaemelanium)
- Viola pusilla Hook. & Arn.—synonym of V. subandina (sect. Subandinium)
- Viola pusilla Poepp.—(sect. Subandinium)
- Viola pusillima Wedd.—(sect. Sempervivum)
- Viola pygmaea Juss. ex Poir.—(sect. Sempervivum)
- Viola pynzarii Vl. V. Nikitin—hybrid (V. reichenbachiana × V. tanaitica; sect. Viola, subsect. Rostratae)
- Viola pyrenaica Ramond ex DC.—(sect. Viola, subsect. Viola)
- Viola pyrolifolia Poir.—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola quercetorum M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola raddeana Regel—(sect. Plagiostigma, subsect. Bilobatae)
- Viola rafinesquei Greene—(sect. Melanium, subsect. Cleistogamae)
- Viola ramiflora K. O. Yoo—synonym of V. hirtipes (sect. Plagiostigma, subsect. Patellares)
- Viola ramosiana W. Becker—synonym of V. sikkimensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola ramosissima Turcz.—synonym of V. dombeyana (sect. Leptidium)
- Viola rauliniana Erben—(sect. Melanium, subsect. Ebracteatae)
- Viola raunsiensis W. Becker & Košanin—(sect. Melanium, subsect. Bracteolatae)
- Viola rausii Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola ravida House—hybrid (V. hirsutula × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola redacta House—hybrid (V. fimbriatula × V. hirsutula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola redunca House—synonym of V. pedata subsp. pedata (sect. Nosphinium, subsect. Pedatae)
- Viola regina J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola reichei f. gunckelii Sparre—synonym of V. reichei (sect. Chilenium)
- Viola reichei Skottsb. ex Macloskie—(sect. Chilenium)
- Viola reichenbachiana Jord. ex Boreau—(sect. Viola, subsect. Rostratae)
- Viola reinii W. Becker—synonym of V. phalacrocarpa (sect. Plagiostigma, subsect. Patellares)
- Viola renifolia A. Gray—(sect. Plagiostigma, subsect. Stolonosae)
- Viola renifolia var. brainerdii (Greene) Fernald—synonym of V. renifolia (sect. Plagiostigma, subsect. Stolonosae)
- Viola reniformis (Sweet) Endl.—synonym of V. banksii (sect. Erpetion)
- Viola repens Schwein.—synonym of V. striata (sect. Viola, subsect. Rostratae)
- Viola repens Turcz. ex Trautv. & C. A. Mey., non Schwein. —synonym of V. suecica subsp. repens (sect. Plagiostigma, subsect. Stolonosae)
- Viola replicata W. Becker—(sect. Rosulatae)
- Viola reschetnikovae Vl. V. Nikitin—hybrid (V. rupestris × V. selkirkii)
- Viola reticulata Schltdl.—synonym of V. humilis (sect. Nosphinium, subsect. Mexicanae)
- Viola retusa Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola rheophila Okamoto—synonym of V. annamensis (sect. Plagiostigma, subsect. Australasiaticae)
- Viola rhizomata Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola rhodopeia W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola rhodosepala Kitag.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola rhombifolia Leyb.—(sect. Subandinium)
- Viola rimosa P. Nikit. — fossil, synonym of Poliothyrsis maii Arbuzova & Zhil. (Salicaceae)
- Viola ritschliana W. Becker—hybrid (V. canina × V. stagnina; sect. Viola, subsect. Rostratae)
- Viola riviniana Rchb.—(sect. Viola, subsect. Rostratae)
- Viola riviniana subsp. minor (Greg.) Valentine—synonym of V. riviniana (sect. Viola, subsect. Rostratae)
- Viola robinsoniana House—hybrid (V. fimbriatula × V. palmata var. triloba; sect. Nosphinium, subsect. Borealiamericanae)
- Viola robinsonii Ridl.—synonym of V. sumatrana (sect. Plagiostigma, subsect. Australasiaticae)
- Viola robusta Hillebr.—(sect. Nosphinium, subsect. Nosphinium)
- Viola roccabrunensis Espeut—(sect. Melanium, subsect. Bracteolatae)
- Viola rockiana W. Becker—(sect. Chamaemelanium)
- Viola rodriguezii W. Becker—(sect. Rosulatae)
- Viola roigii Rossow—(sect. Rosulatae)
- Viola rosacea Brainerd—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola rossii Hemsl.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola rossowiana J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola rosthornii E. Pritzel—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola rostrata Pursh—(sect. Viola, subsect. Rostratae)
- Viola rosulata Poepp. & Endl.—(sect. Rosulatae)
- Viola rotundifolia Michx.—(sect. Chamaemelanium)
- Viola rotundifolia Michx. var. pallens Banks ex Ging.—probable synonym of V. palustris (sect. Plagiostigma, subsect. Stolonosae)
- Viola rubella Cav.—(sect. Rubellium)
- Viola rubella var. angustifolia Ging.—synonym of V. rubella (sect. Rubellium)
- Viola rubella var. latifolia Ging.—synonym of V. rubella (sect. Rubellium)
- Viola rubromarginata J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola rudolfii Vl. V. Nikitin—synonym of V. collina (sect. Viola, subsect. Viola)
- Viola rudolphii Sparre—(sect. Chilenium)
- Viola rugosa Phil. ex W. Becker—(sect. Rosulatae)
- Viola rugulosa Greene—(sect. Chamaemelanium)
- Viola rupestriformis Vl. V. Nikitin—hybrid (V. elatior × V. rupestris; sect. Viola, subsect. Rostratae)
- Viola rupestris F. W. Schmidt—(sect. Viola, subsect. Rostratae)
- Viola rupestris F. W. Schmidt subsp. rupestris—(sect. Viola, subsect. Rostratae)
- Viola rupestris nothosubsp. glabrescens (Neum.) Vl. V. Nikitin—synonym of V. rupestris subsp. rupestris (sect. Viola, subsect. Rostratae)
- Viola rupestris subsp. glaberrima (Murb.) Vl. V. Nikitin—synonym of V. rupestris subsp. rupestris (sect. Viola, subsect. Rostratae)
- Viola rupestris subsp. licentii W. Becker—(sect. Viola, subsect. Rostratae)
- Viola rupestris subsp. relicta Jalas—(sect. Viola, subsect. Rostratae)
- Viola rupicola Elmer—(sect. Plagiostigma, subsect. Patellares)
- Viola ruppii All.—synonym of V. canina (sect. Viola, subsect. Rostratae)
- Viola ruprechtiana Borbás—hybrid (V. epipsila × V. palustris; sect. Plagiostigma, subsect. Stolonosae)
- Viola ruralis Jord. ex Boreau—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola russellii B. Boivin—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Viola rydbergii Greene—synonym of V. canadensis (sect. Chamaemelanium)
- Viola ryoniae House—hybrid (V. cucullata × V. subsinuata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola sabulosa (DC.) Boreau—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola saccata Melch.—(sect. Leptidium)
- Viola sacchalinensis H. Boissieu—(sect. Viola, subsect. Rostratae)
- Viola sacculus Skottsb.—(sect. Sempervivum)
- Viola sagittata Aiton—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola sagittata var. emarginata Nutt.—synonym of V. emarginata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sagittata var. hicksii Pollard—synonym of V. fimbriatula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sagittata var. ovata (Nutt.) Torr. & A. Gray—synonym of V. fimbriatula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sagittata var. subsagittata (Greene) Pollard—synonym of V. sagittata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola samothracica (Degen) Raus—(sect. Melanium, subsect. Bracteolatae)
- Viola sandrasea Melch.—(sect. Viola, subsect. Viola)
- Viola sandrasea subsp. cilicica Contandr. & Quézel—synonym of V. sandrasea (sect. Viola, subsect. Viola)
- Viola santiagonensis W. Becker—(sect. Sempervivum)
- Viola sarmentosa Burgersd., non M. Bieb.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola sarmentosa Douglas, non M. Bieb.—synonym of V. sempervirens (sect. Chamaemelanium)
- Viola sarmentosa var. orbiculata A. Gray—synonym of V. orbiculata (sect. Chamaemelanium)
- Viola savatieri Makino—hybrid (V. albida (V. eizanensis) × V. tokubuchiana; sect. Plagiostigma, subsect. Patellares)
- Viola savatieri var. detonsa Kitag.—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola saxatilis F. W. Schmidt—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola saxifraga Maire—(sect. Xylinosium)
- Viola scabra F. Braun—hybrid (V. hirta × V. odorata; sect. Viola, subsect. Viola)
- Viola scabrida Nakai—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola scandens Humb. & Bonpl. ex Schult.—(sect. Leptidium)
- Viola scandens var. hirtella L. B. Sm. & A. Fernández—synonym of V. scandens var. scandens (sect. Leptidium)
- Viola scandens var. integristipula Benoist—(sect. Leptidium)
- Viola scandens Willd. ex Humb. & Bonpl. var. scandens—(sect. Leptidium)
- Viola schachimardanica Khalk.—synonym of V. biflora (sect. Chamaemelanium)
- Viola schaffneriana W. Becker—synonym of V. grahamii (sect. Nosphinium, subsect. Mexicanae)
- Viola schariensis Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola schauloi Vl. V. Nikitin—hybrid (V. rupestris × V. sacchalinensis; sect. Viola, subsect. Rostratae)
- Viola schensiensis W. Becker—probable syn. of V. striatella (sect. Plagiostigma, subsect. ?Stolonosae)
- Viola schneideri W. Becker—synonym of V. davidii (sect. Plagiostigma, subsect. Stolonosae)
- Viola schulzeana W. Becker—synonym of V. biflora (sect. Chamaemelanium)
- Viola scopulorum (A. Gray) Greene—(sect. Chamaemelanium)
- Viola scorpiuroides Coss.—(sect. Xylinosium)
- Viola scrotiformis DC. ex Ging.—synonym of V. decumbens (sect. Melvio)
- Viola seleriana W. Becker—synonym of V. grahamii (sect. Nosphinium, subsect. Mexicanae)
- Viola selkirkii H. Boissieu & Capit., non Pursh—synonym of V. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii Pursh ex Goldie—(sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. albiflora Nakai—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. angustistipulata W. Becker p. p.—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. brevicalcarata (W. Becker) W. Becker—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. glabrescens W. Becker & Hultén—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. subbarbata W. Becker—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola selkirkii var. variegata Nakai—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola semilunaris (Maxim.) W. Becker—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola sempervirens Greene—(sect. Chamaemelanium)
- Viola sempervirens Greene subsp. sempervirens—(sect. Chamaemelanium)
- Viola sempervirens subsp. orbiculoides M. S. Baker—(sect. Chamaemelanium)
- Viola sempervirens var. orbiculata (A. Gray) J. K. Henry—synonym of V. orbiculata (sect. Chamaemelanium)
- Viola sempervivum Gay—(sect. Sempervivum)
- Viola senzanensis Hayata—(sect. Plagiostigma, subsect. Patellares)
- Viola seoulensis Nakai—(sect. Plagiostigma, subsect. Patellares)
- Viola septemloba Leconte—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola septemloba subsp. egglestonii (Brainerd) L. E. McKinney—synonym of V. egglestonii (sect. Nosphinium, subsect. Borealiamericanae)
- Viola septentrionalis Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola sequeirae Capelo, R. Jardim, J. C. Costa, Lousã & Rivas Mart.—synonym of V. riviniana (sect. Viola, subsect. Rostratae)
- Viola sequoiensis Kellogg—synonym of V. lobata subsp. lobata (sect. Chamaemelanium)
- Viola sergievskiae Tzvelev—synonym of V. rupestris (sect. Viola, subsect. Rostratae)
- Viola sermenika Formánek—hybrid (V. aetolica × V. tricolor (“V. macedonica”); sect. Melanium, subsect. Bracteolatae)
- Viola serpens subsp. gurhwalensis W. Becker—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola serpens var. confusa (Benth.) J. D. Hooker & Thomson—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola serpens var. pseudoscotophylla H. Boissieu—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola serpens Wallich ex Ging.—synonym of V. pilosa (sect. Viola, subsect. Viola)
- Viola serpentinicola de Salas—(sect. Erpetion)
- Viola serresiana Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola serrula W. Becker—(sect. Viola, subsect. Rostratae)
- Viola sfikasiana Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola shaoyoukengensis S. S. Ying—synonym of V. nagasawae (sect. Plagiostigma, subsect. Diffusae)
- Viola sheltonii Torr.—(sect. Chamaemelanium)
- Viola sheltonii var. biternata (Greene) A. Nelson—synonym of V. sheltonii (sect. Chamaemelanium)
- Viola shikokiana Makino—(sect. Plagiostigma, subsect. Stolonosae)
- Viola shinchikuensis Yamam.—(sect. Viola, subsect. Rostratae)
- Viola shiweii Xiao Chen Li & Z. W. Wang—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola sieberiana Spreng.—(sect. Erpetion)
- Viola sieboldiana (Maxim.) Makino—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola sieboldiana var. chaerophylloides (Regel) Nakai—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola sieboldii Maxim.—(sect. Plagiostigma, subsect. Patellares)
- Viola sieheana W. Becker—(sect. Viola, subsect. Rostratae)
- Viola sikkimensis var. acuminatifolia W. Becker—synonym of V. sumatrana (sect. Plagiostigma, subsect. Australasiaticae)
- Viola sikkimensis var. debilis W. Becker—synonym of V. kwangtungensis (sect. Plagiostigma, subsect. Australasiaticae)
- Viola sikkimensis W. Becker—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola silicestris K. R. Thiele & Prober—(sect. Erpetion)
- Viola silvestriformis W. Becker—synonym of V. kusanoana (sect. Viola, subsect. Rostratae)
- Viola simulata M. S. Baker—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola singularis J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola sintenisii W. Becker—(sect. Viola, subsect. Viola)
- Viola sirinica Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola skofitziana Wiesb.—hybrid (V. elatior × V. pumila; sect. Viola, subsect. Rostratae)
- Viola skottsbergiana W. Becker—(sect. Sempervivum)
- Viola slavikii Formánek—(sect. Melanium, subsect. Bracteolatae)
- Viola smithiana W. Becker—synonym of V. davidii (sect. Plagiostigma, subsect. Stolonosae)
- Viola somalensis Engl.—(sect. Sclerosium)
- Viola somchetica K. Koch—(sect. Plagiostigma, subsect. Patellares)
- Viola sororia subsp. affinis (Leconte) R. J. Little—synonym of V. affinis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sororia var. affinis (Leconte) L. E. McKinney—synonym of V. affinis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sororia var. missouriensis (Greene) L. E. McKinney—synonym of V. missouriensis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sororia var. novae-angliae (House) L. E. McKinney—synonym of V. novae-angliae (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sororia Willd.—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola spathulata Willd. ex Schult.—(sect. Spathulidium)
- Viola speciosa Schrad. ex Rchb.—synonym of V. elegantula (sect. Melanium, subsect. Bracteolatae)
- Viola spegazzinii W. Becker—(sect. Rosulatae)
- Viola sphaerocarpa W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola splendida W. Becker—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola squamulosa W. Becker—synonym of V. sacculus (sect. Sempervivum)
- Viola stagnina Kit. ex Schult.—(sect. Viola, subsect. Rostratae)
- Viola steinbachii W. Becker—(sect. Leptidium)
- Viola stellata Miers—synonym of V. pusilla (sect. Subandinium)
- Viola stenocentra Hayata ex Nakai—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola stewardiana W. Becker—(sect. Viola, subsect. Rostratae)
- Viola stipularis Sw.—(sect. Leptidium)
- Viola stocksii Boiss.—(sect. Sclerosium)
- Viola stojanowii W. Becker—(sect. Melanium, subsect. Bracteolatae)
- Viola stolonifera J. J. Rodr.—synonym of V. odorata (sect. Viola, subsect. Viola)
- Viola stoloniflora Yokota & Higa—(sect. Plagiostigma, subsect. Formosanae)
- Viola stoneana House—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola striata Aiton—(sect. Viola, subsect. Rostratae)
- Viola striatella H. Boissieu—(sect. Plagiostigma, subsect. Stolonosae)
- Viola striis-notata (J. Wagner) Merxm. & W. Lippert—(sect. Melanium, subsect. Bracteolatae)
- Viola stuebelii Hieron.—(sect. Chilenium)
- Viola suaviflora Borbás & Heinr. Braun—hybrid (V. collina × V. suavis; sect. Viola, subsect. Viola)
- Viola suavis M. Bieb.—(sect. Viola, subsect. Viola)
- Viola suavis subsp. adriatica (Freyn) Haesler—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola suavis subsp. austrodalmatica Mereďa & Hodálová—synonym of V. suavis (sect. Viola, subsect. Viola)
- Viola subaequiloba (Franch. & Sav.) Nakai—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola subaffinis House—hybrid (V. affinis × V. nephrophylla), synonym of V. venustula (sect. Nosphinium, subsect. Borealiamericanae)
- Viola subandina J. M. Watson—(sect. Subandinium)
- Viola subasica (Fiori) Ricceri & Moraldo—synonym of V. eugeniae (sect. Melanium, subsect. Bracteolatae)
- Viola subatlantica (Maire) Ibn Tattou—(sect. Melanium, subsect. Bracteolatae)
- Viola subdelavayi S. H. Huang—synonym of V. urophylla (sect. Chamaemelanium)
- Viola subdimidiata A. St.-Hil.—(sect. Leptidium)
- Viola subglabra (Ledeb.) Baikov ex A. V. Grebenjuk & Chepinoga—synonym of V. mirabilis (sect. Viola, subsect. Rostratae)
- Viola subjuncta Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola sublanceolata House—hybrid (V. lanceolata × V. minuscula; sect. Nosphinium, subsect. Borealiamericanae)
- Viola subsagittata Greene—hybrid (V. fimbriatula × V. sagittata; sect. Nosphinium, subsect. Borealiamericanae)
- Viola subsagittifolia Suksd.—synonym of V. vallicola var. vallicola (sect. Chamaemelanium)
- Viola subsinuata (Greene) Greene—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola subviscosa Greene—synonym of V. septentrionalis (sect. Nosphinium, subsect. Borealiamericanae)
- Viola suecica Fr.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola suecica Fr. subsp. suecica—(sect. Plagiostigma, subsect. Stolonosae)
- Viola suecica subsp. repens W. Becker—(sect. Plagiostigma, subsect. Stolonosae)
- Viola sukaczewii Vl. V. Nikitin—hybrid (V. canina × V. mirabilis; sect. Viola, subsect. Rostratae)
- Viola sumatrana Miq.—(sect. Plagiostigma, subsect. Australasiaticae)
- Viola superba M. S. Baker—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola sylvestris var. candida (H. Boissieu) H. Lév.—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola sylvestris var. grypoceras (A. Gray) Maxim.—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola sylvestris var. puberula E. Sheld.—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola szetschwanensis var. kangdienensis Chang—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola szetschwanensis var. nudicaulis W. Becker—synonym of V. szetschwanensis (sect. Chamaemelanium)
- Viola szetschwanensis W. Becker & H. Boissieu—(sect. Chamaemelanium)
- Viola taischanensis Ching J. Wang—synonym of V. prionantha (sect. Plagiostigma, subsect. Patellares)
- Viola taiwanensis W. Becker—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola taiwaniana Nakai—synonym of V. philippica subsp. malesica (sect. Plagiostigma, subsect. Patellares)
- Viola takahashii (Nakai) Taken.—synonym of V. albida (sect. Plagiostigma, subsect. Patellares)
- Viola takasagoensis Koidz.—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola takedana Makino—synonym of V. tokubuchiana (sect. Plagiostigma, subsect. Patellares)
- Viola takesimana Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola talmensis Vl. V. Nikitin—hybrid (V. rupestris × V. stagnina (“V. persicifolia”); sect. Viola, subsect. Rostratae)
- Viola taltalensis var. glaberrima W. Becker—synonym of V. taltalensis (sect. Subandinium)
- Viola taltalensis W. Becker—(sect. Subandinium)
- Viola tanaitica Grosset—(sect. Viola, subsect. Rostratae)
- Viola tarbagataica Klokov—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tashiroi Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola tatrae Borbás—hybrid (V. tricolor (subsp. saxatilis) × V. lutea (subsp. sudetica); sect. Melanium, subsect. Bracteolatae)
- Viola tayemonii Hayata—synonym of V. biflora (sect. Chamaemelanium)
- Viola taynensis Elisafenko—synonym of V. odorata (sect. Viola, subsect. Viola)
- Viola tectiflora W. Becker—(sect. Rosulatae)
- Viola tenuicornis subsp. ircutiana (Turcz.) Espeut—(sect. Plagiostigma, subsect. Patellares)
- Viola tenuicornis subsp. primorskajensis W. Becker—synonym of V. variegata subsp. primorskajensis (sect. Plagiostigma, subsect. Patellares)
- Viola tenuicornis subsp. tenuicornis—(sect. Plagiostigma, subsect. Patellares)
- Viola tenuicornis subsp. trichosepala W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola tenuicornis var. brachytricha W. Becker—synonym of V. tenuicornis subsp. tenuicornis (sect. Plagiostigma, subsect. Patellares)
- Viola tenuicornis W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola tenuipes Pollard—(sect. Chamaemelanium)
- Viola tenuis Benth.—(sect. Plagiostigma, subsect. Diffusae)
- Viola tenuis var. geralensis W. Becker—synonym of V. uleana (sect. Leptidium)
- Viola tenuis W. Becker—synonym of V. uleana (sect. Leptidium)
- Viola tenuissima C. C. Chang—(sect. Chamaemelanium)
- Viola teplouchovii Juz.—synonym of V. mauritii (sect. Viola, subsect. Rostratae)
- Viola teshioensis Miyabe & Tatew.—synonym of V. collina (sect. Viola, subsect. Viola)
- Viola thasia W. Becker—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola thianschanica Maxim.—synonym of V. kunawurensis (sect. Himalayum)
- Viola thibaudieri Franch. & Sav.—(sect. Viola, subsect. Rostratae)
- Viola thomasiana Songeon & E. P. Perrier—(sect. Viola, subsect. Viola)
- Viola thomsonii Oudem.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola thrichopoda Hayata—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola thymifolia Britton—(sect. Leptidium)
- Viola tichomirovii Vl. V. Nikitin—hybrid (V. canina × V. elatior; sect. Viola, subsect. Rostratae)
- Viola tienschiensis W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola tigirekica Vl. V. Nikitin—hybrid (V. altaica × V. tricolor; sect. Melanium, subsect. Bracteolatae)
- Viola tineorum Erben & Raimondo—(sect. Melanium, subsect. Bracteolatae)
- Viola tokaiensis Sugim., nom. nud.—(sect. Plagiostigma, subsect. Patellares)
- Viola tokubuchiana Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola tomentosa M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola tomentosifolia Makino ex U. Ohga—synonym of V. collina (sect. Viola, subsect. Viola)
- Viola toppingii Elmer—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola toroensis Martic.—synonym of V. philippiana (sect. Rosulatae)
- Viola torslundensis W. Becker—hybrid (V. elatior × V. stagnina; sect. Viola, subsect. Viola)
- Viola tosaensis Nakai—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola tovarii P. Gonzáles & Molina-Alor—(sect. Rosulatae)
- Viola tozanensis Hayata—synonym of V. formosana (sect. Plagiostigma, subsect. Formosanae)
- Viola tracheliifolia Ging.—(sect. Nosphinium, subsect. Nosphinium)
- Viola triangulifolia W. Becker—(sect. Plagiostigma, subsect. Bilobatae)
- Viola trichopetala C. C. Chang—(sect. Plagiostigma, subsect. Patellares)
- Viola trichosepala (W. Becker) Juz.—synonym of V. tenuicornis subsp. trichosepala (sect. Plagiostigma, subsect. Patellares)
- Viola tricolor L.—(sect. Melanium, subsect. Bracteolatae)
- Viola tricolor L. subsp. tricolor—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. alpestris (Ging.) Ces.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. ammotropha Wittr.—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. curtisii (V. Forster) Syme—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. macedonica (Boiss. & Heldr.) A. Schmidt—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. matutina (Klokov) Valentine—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. polychroma (A. Kern.) Nyman—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor subsp. subalpina Gaudin—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor var. arvensis (Murray) DC.—synonym of V. arvensis (sect. Melanium, subsect. Bracteolatae)
- Viola tricolor var. hortensis DC.—probable synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola tridentata Sm.—(sect. Tridens)
- Viola triflabellata W. Becker—(sect. Triflabellium)
- Viola triloba Schwein.—synonym of V. palmata var. triloba (sect. Nosphinium, subsect. Borealiamericanae)
- Viola triloba var. dilatata (Elliott) Brainerd—synonym of V. palmata var. palmata (sect. Nosphinium, subsect. Borealiamericanae)
- Viola trinervata (Howell) Howell ex A. Gray—(sect. Chamaemelanium)
- Viola trinervis Korth.—synonym of V. inconspicua (sect. Plagiostigma, subsect. Patellares)
- Viola tripartita Elliott—(sect. Chamaemelanium)
- Viola trochlearis J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola truncata Meyen—(sect. Grandiflos)
- Viola truncata var. glaberrima W. Becker—synonym of V. truncata (sect. Grandiflos)
- Viola truncata var. glandulifera W. Becker—synonym of V. cheeseana (sect. Grandiflos)
- Viola truncata W. Becker, non Meyen—synonym of V. boliviana (sect. Leptidium)
- Viola tsugitakaensis Masamune—probable synonym of V. fargesii (sect. Plagiostigma, subsect. Stolonosae)
- Viola tuberifera Franch.—(sect. Plagiostigma, subsect. Bulbosae)
- Viola tuberifera var. brevipedicellata S. Y. Chen—synonym of V. tuberifera (sect. Plagiostigma, subsect. Bulbosae)
- Viola tuberifera var. pseudopalustris H. Lév.—synonym of V. tuberifera (sect. Plagiostigma, subsect. Bulbosae)
- Viola tucumanensis W. Becker—(sect. Triflabellium)
- Viola turczaninowii Juz.—synonym of V. acuminata subsp. acuminata (sect. Viola, subsect. Rostratae)
- Viola turkestanica Regel & Schmalh.—(sect. Plagiostigma, subsect. Patellares)
- Viola turritella J. M. Watson & A. R. Flores—(sect. Sempervivum)
- Viola tuvinica Vl. V. Nikitin—hybrid (V. sacchalinensis (V. komarovii) × V. mauritii; sect. Viola, subsect. Rostratae)
- Viola tzvelevii Vl. V. Nikitin—hybrid (V. pumila × V. riviniana; sect. Viola, subsect. Rostratae)
- Viola ucriana Erben & Raimondo—(sect. Melanium, subsect. Bracteolatae)
- Viola uechtritziana Borbás—hybrid (V. mirabilis × V. reichenbachiana; sect. Viola)
- Viola uleana W. Becker—(sect. Leptidium)
- Viola uliginosa Besser—(sect. Viola, subsect. Rostratae)
- Viola ulleungdoensis M. Kim & J. Lee—(sect. Plagiostigma, subsect. Patellares)
- Viola umbraticola Kunth—(sect. Viola, subsect. Rostratae)
- Viola umbraticola var. glaberrima W. Becker—synonym of V. umbraticola (sect. Viola, subsect. Rostratae)
- Viola umbrosa (Wahlb.) Fr.—synonym of V. selkirkii (sect. Plagiostigma, subsect. Patellares)
- Viola umemurae Makino—synonym of V. variegata subsp. primorskajensis (sect. Plagiostigma, subsect. Patellares)
- Viola umphangensis S. Nansai, Srisanga & Suwanph.—(sect. Plagiostigma, subsect. Patellares)
- Viola uncinulata Greene—synonym of V. adunca subsp. adunca (sect. Viola, subsect. Rostratae)
- Viola unica J. M. Watson & A. R. Flores, nom. inval.—synonym of V. uniquissima (sect. Triflabellium)
- Viola uniflora L.—(sect. Chamaemelanium)
- Viola uniflora subsp. lasczinskyi Zuev—synonym of V. uniflora (sect. Chamaemelanium)
- Viola uniflora var. orientalis Maxim.—synonym of V. orientalis subsp. orientalis (sect. Chamaemelanium)
- Viola uniquissima J. M. Watson & A. R. Flores—(sect. Triflabellium)
- Viola unwinii W. Becker—synonym of V. hossei (sect. Plagiostigma, subsect. Stolonosae)
- Viola urophylla Franch.—(sect. Chamaemelanium)
- Viola urophylla var. densivillosa C. J. Wang—synonym of V. urophylla (sect. Chamaemelanium)
- Viola ursina Kom.—synonym of V. langsdorffii (sect. Nosphinium, subsect. Langsdorffianae)
- Viola utahensis M. S. Baker & J. C. Clausen—(sect. Chamaemelanium)
- Viola utchinensis Koidz.—(sect. Viola, subsect. Rostratae)
- Viola vadimii Vl. V. Nikitin—synonym of V. elatior (sect. Viola, subsect. Rostratae)
- Viola vaginata Maxim.—(sect. Plagiostigma, subsect. Stolonosae)
- Viola vaginata subsp. alata W. Becker—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola vaginata var. sutchuensis Franch. ex H. Boissieu—synonym of V. moupinensis (sect. Plagiostigma, subsect. Stolonosae)
- Viola vagula Greene—synonym of V. nephrophylla (sect. Nosphinium, subsect. Borealiamericanae)
- Viola valderia All.—(sect. Melanium, subsect. Bracteolatae)
- Viola valdiviana Kalela—synonym of V. maculata var. maculata (sect. Chilenium)
- Viola vallenarensis W. Becker—(sect. Subandinium)
- Viola vallicola A. Nelson—(sect. Chamaemelanium)
- Viola vallicola var. major (Hook.) Fabijan—(sect. Chamaemelanium)
- Viola vallicola var. vallicola—(sect. Chamaemelanium)
- Viola vanroyenii H. St. John—synonym of V. kauaensis (sect. Nosphinium, subsect. Nosphinium)
- Viola variegata Fisch. ex Link—(sect. Plagiostigma, subsect. Patellares)
- Viola variegata Fisch. ex Link subsp. variegata—(sect. Plagiostigma, subsect. Patellares)
- Viola variegata subsp. chinensis (Bunge ex Regel) W. Becker—synonym of V. tenuicornis subsp. tenuicornis (sect. Plagiostigma, subsect. Patellares)
- Viola variegata subsp. primorskajensis (W. Becker) Espeut—(sect. Plagiostigma, subsect. Patellares)
- Viola variegata var. chinensis Bunge ex Regel—synonym of V. tenuicornis subsp. tenuicornis (sect. Plagiostigma, subsect. Patellares)
- Viola variegata var. nipponica Makino—synonym of V. variegata subsp. primorskajensis (sect. Plagiostigma, subsect. Patellares)
- Viola variegata var. viridis Kitag.—synonym of V. variegata (sect. Plagiostigma, subsect. Patellares)
- Viola vectoris Ricceri & Moraldo—synonym of V. eugeniae (sect. Melanium, subsect. Bracteolatae)
- Viola velutina Formánek—(sect. Melanium, subsect. Bracteolatae)
- Viola venusta Nakai—synonym of V. sacchalinensis (sect. Viola, subsect. Rostratae)
- Viola venustula Greene—hybrid (V. affinis × V. nephrophylla; sect. Nosphinium, subsect. Borealiamericanae)
- Viola verecunda A. Gray—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola vermontana House—hybrid (V. latiuscula × V. sororia; sect. Nosphinium, subsect. Borealiamericanae)
- Viola veronicifolia Planch. & Linden—(sect. Leptidium)
- Viola vespertina Klokov—synonym of V. tricolor (sect. Melanium, subsect. Bracteolatae)
- Viola viarum Pollard—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola viatkensis Vl. V. Nikitin—hybrid (V. canina × V. selkirkii)
- Viola vilaensis Hayek—(sect. Viola, subsect. Viola)
- Viola villaquensis Benz—hybrid (V. canina (“V. montana”) × V. rupestris (var. arenaria); sect. Viola, subsect. Rostratae)
- Viola villosa Walter—(sect. Nosphinium, subsect. Borealiamericanae)
- Viola vilnaensis W. Becker—hybrid (V. rupestris × V. stagnina; sect. Viola, subsect. Rostratae)
- Viola vindobonensis Wiesb.—hybrid (V. odorata × V. suavis; sect. Viola, subsect. Viola)
- Viola violacea Makino—(sect. Plagiostigma, subsect. Patellares)
- Viola vittata Greene—(sect. Plagiostigma, subsect. Stolonosae)
- Viola volcanica Gillies ex Hook. & Arn.—(sect. Rosulatae)
- Viola volcanica var. exilis (Phil.) Reiche—synonym of V. exilis (sect. Rosulatae)
- Viola volcanica var. truncata (Meyen) Reiche—synonym of V. truncata (sect. Grandiflos)
- Viola voliotisii Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola vorobievii Bezd.—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola vourinensis Erben—(sect. Melanium, subsect. Bracteolatae)
- Viola vulturis Ricceri & Moraldo—synonym of V. aethnensis subsp. splendida (sect. Melanium, subsect. Bracteolatae)
- Viola wailenalenae (Rock) Skottsb.—(sect. Nosphinium, subsect. Nosphinium)
- Viola wallichiana Ging.—(sect. Chamaemelanium)
- Viola wallichiana subsp. brevicornis W. Becker—synonym of V. biflora (sect. Chamaemelanium)
- Viola walteri House—(sect. Viola, subsect. Rostratae)
- Viola wansanensis Y. N. Lee—hybrid (V. albida (V. chaerophylloides) × V. mandshurica; sect. Plagiostigma, subsect. Patellares)
- Viola weberbaueri W. Becker—(sect. Subandinium)
- Viola websteri Hemsl.—(sect. Viola, subsect. Rostratae)
- Viola weddellii W. Becker—synonym of V. thymifolia (sect. Leptidium)
- Viola weibelii J. F. Macbr.—(sect. Inconspicuiflos)
- Viola weixiensis C. J. Wang—synonym of V. pendulicarpa (sect. Viola, subsect. Rostratae)
- Viola werdermannii f. glandulifera W. Becker—synonym of V. polypoda (sect. Subandinium)
- Viola werdermannii var. glaberrima W. Becker—synonym of V. polypoda (sect. Subandinium)
- Viola werdermannii var. typica W. Becker—synonym of V. polypoda (sect. Subandinium)
- Viola werdermannii W. Becker—synonym of V. polypoda (sect. Subandinium)
- Viola wettsteinii Richt.—synonym of V. reichenbachiana (sect. Viola, subsect. Rostratae)
- Viola wichurae Nakai—synonym of V. grypoceras (sect. Viola, subsect. Rostratae)
- Viola wikipedia J. M. Watson & A. R. Flores—nom. illeg. ; synonym of V. angustifolia (sect. Grandiflos)
- Viola wilczekiana Beauverd—hybrid (V. hirta × V. rupestris; sect. Viola)
- Viola wilhelmii Vl. V. Nikitin—hybrid (V. oreades × V. tricolor (subsp. saxatilis); sect. Melanium, subsect. Bracteolatae)
- Viola wilibaldii Vl. V. Nikitin—hybrid (V. pumila × V. reichenbachiana; sect. Viola, subsect. Rostratae)
- Viola williamsii Wittr.—hybrid (garden cultivar; V. wittrockiana × V. cornuta; sect. Melanium, subsect. Bracteolatae)
- Viola willkommii R. Roem. ex Willk.—(sect. Viola, subsect. Rostratae)
- Viola wilmattiae Pollard & Cockerell—hybrid (V. nephrophylla × V. pedatifida; sect. Nosphinium, subsect. Borealiamericanae)
- Viola wilsonii W. Becker—(sect. Plagiostigma, subsect. Diffusae)
- Viola wittrockiana Gams ex Nauenb. & Buttler—hybrid (garden cultivar involving at least V. altaica, V. lutea, and V. tricolor; sect. Melanium, subsect. Bracteolatae)
- Viola woosanensis Y. N. Lee & J. Kim—hybrid (V. albida (V. chaerophylloides) × V. ulleungdoensis; sect. Plagiostigma, subsect. Patellares)
- Viola woroschilovii Bezd.—synonym of V. prionantha (sect. Plagiostigma)
- Viola wujekii H. E. Ballard—hybrid (V. appalachensis × V. striata; sect. Viola, subsect. Rostratae)
- Viola wulingensis S. S. Ying—synonym of V. senzanensis (sect. Plagiostigma, subsect. Patellares)
- Viola xanthopetala Nakai—synonym of V. orientalis subsp. xanthopetala (sect. Chamaemelanium)
- Viola xanthopotamica J. M. Watson & A. R. Flores—(sect. Rosulatae)
- Viola yakusimana Nakai—synonym of V. hamiltoniana (sect. Plagiostigma, subsect. Bilobatae)
- Viola yamatsutae Ishidoya ex Kitag.—synonym of V. mongolica (sect. Plagiostigma, subsect. Patellares)
- Viola yazawana Makino—(sect. Plagiostigma, subsect. Stolonosae)
- Viola yedoensis f. candida Kitag.—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola yedoensis f. intermedia Kitag.—synonym of V. mandshurica (sect. Plagiostigma, subsect. Patellares)
- Viola yedoensis Makino—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola yedoensis var. pseudojaponica (Nakai) T. Hashimoto ex E. Hama & K. Nakai—synonym of V. philippica subsp. philippica (sect. Plagiostigma, subsect. Patellares)
- Viola yezoensis Maxim.—(sect. Plagiostigma, subsect. Patellares)
- Viola yezoensis var. hebeiensis (J. W. Wang & T. G. Ma) J. W. Wang & J. Yang [“hopeiensis”]—synonym of V. mongolica (sect. Plagiostigma, subsect. Patellares)
- Viola yildirimlii Dinç & Bagci—(sect. Viola, subsect. Viola)
- Viola yubariana Nakai—synonym of V. brevistipulata (sect. Chamaemelanium)
- Viola yunnanensis W. Becker & H. Boissieu—(sect. Plagiostigma, subsect. Diffusae)
- Viola yunnanfuensis W. Becker—(sect. Plagiostigma, subsect. Patellares)
- Viola yurii Vl. V. Nikitin—hybrid (V. collina × V. riviniana; sect. Viola)
- Viola yuzufeliensis A. P. Khokhr.—(sect. Plagiostigma, subsect. Patellares)
- Viola zongia Tul.—synonym of V. abyssinica (sect. Abyssinium)
- Viola zophodes K. R. Thiele & Prober—hybrid (V. eminens × V. fuscoviolacea; sect. Erpetion)
- Viola zwienenii J. M. Watson & A. R. Flores—hybrid (V. atropurpurea × V. beckeriana; sect. Sempervivum)
References
- Becker, W. Viola L. In Die Natürlichen Pflanzenfamilien 21. Parietales und Opuntiales; Engler, A., Ed.; Wilhelm Engelmann: Leipzig, Germany, 1925; Volume 21, pp. 363–376. [Google Scholar]
- Ballard, H.E.; Sytsma, K.J.; Kowal, R.R. Shrinking the violets: Phylogenetic relationships of infrageneric groups in Viola (Violaceae) based on internal transcribed spacer DNA sequences. Syst. Bot. 1999, 23, 439–458. [Google Scholar] [CrossRef]
- Wahlert, G.A.; Marcussen, T.; de Paula-Souza, J.; Feng, M.; Ballard, H.E. A phylogeny of the Violaceae (Malpighiales) inferred from plastid DNA sequences: Implications for generic diversity and intrafamilial taxonomy. Syst. Bot. 2014, 39, 239–252. [Google Scholar] [CrossRef]
- Marcussen, T.; Ballard, H.E.; Wahlert, G.A.; de Paula-Souza, J. Dated phylogeny and historical biogeography of Violaceae. Unpublished manuscript.
- Stevens, P.F. Angiosperm Phylogeny Website. Version 14, July 2017 [and more or less continuously updated since]. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 15 August 2022).
- Frodin, D.A. History and concepts of big plant genera. Taxon 2004, 53, 753–776. [Google Scholar] [CrossRef]
- De Carvalho, L.M.; Fernandes, F.M.; de Fátima Nunes, M.; Brigola, J.; Casbas, N.; Groves, C. History and cultivation of parma violets (Viola, Violaceae) in the United Kingdom and France in the nineteenth century. Harv. Pap. Bot. 2013, 18, 137–145. [Google Scholar] [CrossRef]
- Matthiolus, P.A. Herbarz: Ginak Bylinář Welmi Vžitečný a Figůrami Pieknymi y Zřetedlnymi Podlé Praweho a Yako Ziwého Zrostu Bylin Ozdobeny y Také Mnohymi a Zkussenymi Lékarzstwijmi Rozhognieny Gessto Takowy Nikdá w Ziádnem Yazyku prwé Wydán Nebyl; u Girzijka Melantrycha z Awentynu: W Starém Miestie Pražském [The Old City of Prague, Kingdom of Bohemia], 1562. [Google Scholar]
- Mills, S.; Hutchins, R. Violae Herba Cum Flore—Wild Pansy (Flowering Aerial Parts); ESCOP Monographs Online Series; The Scientific Foundation for Herbal Medicinal Products: Exeter, UK, 2015; pp. 1–14. [Google Scholar]
- Park, S.; Yoo, K.-O.; Marcussen, T.; Backlund, A.; Jacobsson, E.; Rosengren, K.J.; Doo, I.; Göransson, U. Cyclotide evolution: Insights from the analyses of their precursor sequences, structures and distribution in violets (Viola). Front. Plant Sci. 2017, 8, 2058. [Google Scholar] [CrossRef]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar] [CrossRef]
- Gilding, E.K.; Jackson, M.A.; Poth, A.G.; Henriques, S.T.; Prentis, P.J.; Mahatmanto, T.; Craik, D.J. Gene coevolution and regulation lock cyclic plant defence peptides to their targets. New Phytol. 2016, 210, 717–730. [Google Scholar] [CrossRef]
- Camarero, J.A.; Campbell, M.J. The potential of the cyclotide scaffold for drug development. Biomedicines 2019, 7, 31. [Google Scholar] [CrossRef]
- Rajendran, S.; Slazak, B.; Mohotti, S.; Strömstedt, A.A.; Göransson, U.; Hettiarachchi, C.M.; Gunasekera, S. Tropical vibes from Sri Lanka—Cyclotides from Viola betonicifolia by transcriptome and mass spectrometry analysis. Phytochemistry 2021, 187, 112749. [Google Scholar] [CrossRef]
- Slazak, B.; Kaltenböck, K.; Steffen, K.; Rogala, M.; Rodríguez-Rodríguez, P.; Nilsson, A.; Shariatgorji, R.; Andrén, P.E.; Göransson, U. Cyclotide host-defense tailored for species and environments in violets from the Canary Islands. Sci. Rep. 2021, 11, 12452. [Google Scholar] [CrossRef]
- Von Soden, H. Über ätherische Öle, welche durch Extraktion frischer Blüten mit flüchtigen Lösungsmitteln gewonnen werden (ätherische Blütenextraktöle). J. Für Prakt. Chem. 1904, 69, 256–272. [Google Scholar] [CrossRef]
- Saint-Lary, L.; Roy, C.; Paris, J.-P.; Tournayre, P.; Berdagué, J.-L.; Thomas, O.P.; Xavier Fernandez, X. Volatile compounds of Viola odorata absolutes: Identification of odorant active markers to distinguish plants originating from France and Egypt. Chem. Biodivers. 2014, 11, 843–860. [Google Scholar] [CrossRef]
- Aloum, L.; Alefishat, E.; Adem, A.; Petroianu, G. ionone is more than a violet’s fragrance: A review. Molecules 2020, 25, 5822. [Google Scholar] [CrossRef]
- Marcussen, T.; Karlsson, T. Violaceae. In Flora Nordica; Karlsson, T., Ed.; Swedish Royal Academy of Sciences: Stockholm, Sweden, 2010; Volume 6, pp. 12–52. [Google Scholar]
- Malécot, V.; Marcussen, T.; Munzinger, J.; Yockteng, R.; Henry, M. On the origin of the sweet-smelling Parma Violet cultivars (Violaceae): Wide intraspecific hybridization, sterility, and sexual reproduction. Amer. J. Bot. 2007, 94, 29–41. [Google Scholar] [CrossRef]
- Yuzepchuk, S.V. Viola L. In Flora of the U.S.S.R.; Shishkin, B.K., Ed.; Israel Program for Scientific Translations: Jerusalem, Israel, 1949; Volume 15, pp. 263–360. [Google Scholar]
- Muñoz Garmendia, F.; Montserrat, P.; Laínz, M.; Aldasoro, J.J. Viola L. In Flora iberica; Castroviejo, S., Aedo, C., Cirujano, S., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Navarro, C., Paiva, J., Soriano, C., Eds.; Real Jardín Botánico—C.S.I.C.: Madrid, Spain, 1993; Volume 3, pp. 276–298. [Google Scholar]
- Gibbons, E. Stalking the Healthful Herbs; David McKay, Co.: Philadelphia, PA, USA, 1972. [Google Scholar]
- Tokuoka, T. Molecular phylogenetic analysis of Violaceae (Malpighiales) based on plastid and nuclear DNA sequences. J. Plant Res. 2008, 121, 253–260. [Google Scholar] [CrossRef]
- Ballard, H.E.; de Paula-Souza, J.; Wahlert, G.A. Violaceae. In Flowering Plants. Eudicots: Malpighiales; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 303–322. [Google Scholar]
- Thomson, G.M. Notes on cleistogamic flowers of the genus Viola. Trans. Proc. N. Zeal. Inst. 1878, 11, 415–417. [Google Scholar]
- Culley, T.M.; Klooster, M.R. The cleistogamous breeding system: A review of its frequency, evolution, and ecology in angiosperms. Bot. Rev. 2007, 73, 1–30. [Google Scholar] [CrossRef]
- Marcussen, T.; Heier, L.; Brysting, A.K.; Oxelman, B.; Jakobsen, K.S. From gene trees to a dated allopolyploid network: Insights from the angiosperm genus Viola (Violaceae). Syst. Biol. 2015, 64, 84–101. [Google Scholar] [CrossRef]
- Clausen, J. Chromosome number and relationship of some North American species of Viola. Ann. Bot. 1929, 63, 741–764. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Westerhold, T.; Marwan, N.; Drury, A.J.; Liebrand , D.; Agnini, C.; Anagnostou, E.; Barnet, J.S.K.; Bohaty, S.M.; de Vleeschouwer, D.; Florindo, F.; et al. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 2020, 369, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Sauermilch, I.; Whittaker, J.M.; Klocker, A.; Munday, D.R.; Hochmuth, K.; Bijl, P.K.; LaCasce, J.H. Gateway-driven weakening of ocean gyres leads to Southern Ocean cooling. Nat. Commun. 2021, 12, 6465. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Nylander, J.A.A.; Persson, C.; Sanmartín, I. Tracing the impact of the Andean uplift on Neotropical plant evolution. Proc. Natl. Acad. Sci. USA 2009, 106, 9749–9754. [Google Scholar] [CrossRef] [PubMed]
- Aragón, E.; D’Eramo, F.; Castro, A.; Pinotti, L.; Brunelli, D.; Rabbia, O.; Rivalenti, G.; Varela, R.; Spakman, W.; Demartis, M.; et al. Tectono-magmatic response to major convergence changes in the North Patagonian suprasubduction system; the Paleogene subduction–transcurrent plate margin transition. Tectonophysics 2011, 509, 218–237. [Google Scholar] [CrossRef]
- Armijo, R.; Lacassin, R.; Coudurier-Curveur, A.; Carrizo, D. Coupled tectonic evolution of Andean orogeny and global climate. Earth-Sci. Rev. 2015, 143, 1–35. [Google Scholar] [CrossRef]
- Pérez-Escobar, O.A.; Zizka, A.; Bermúdez, M.A.; Meseguer, A.S.; Condamine, F.L.; Hoorn, C.; Hooghiemstra, H.; Pu, Y.; Bogarín, D.; Boschman, L.M.; et al. The Andes through time: Evolution and distribution of Andean floras. Trends Plant Sci. 2022, 27, 364–378. [Google Scholar] [CrossRef]
- Krahulcová, A.; Krahulec, F.; Kirschner, J. Introgressive hybridization between a native and an introduced species: Viola lutea subsp. sudetica versus V. tricolor. Folia Geobot. Phytotax. 1996, 31, 319–344. [Google Scholar] [CrossRef]
- Brandrud, K.H. Viola epipsila, V. palustris og V. epipsila × palustris i Sørøst-Norge. En undersøkelse av cytologi, morfologi og økologi. In Faculty of Mathematics and Natural Sciences; University of Oslo: Oslo, Norway, 1989; pp. 1–86. [Google Scholar]
- Røren, V.; Stabbetorp, O.E.; Borgen, L. Hybridization between Viola canina and V. persicifolia in Norway. Nord. J. Bot. 1994, 14, 165–172. [Google Scholar] [CrossRef]
- Cortés-Palomec, A.C.; Ballard, H.E. Evidence for hybridization between two sympatric violet species, Viola grahamii and V. hookeriana (Violaceae), in central Mexico. IDA Contrib. Bot. 2006, 22, 1119–1133. [Google Scholar]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef]
- Mayrose, I.; Zhan, S.H.; Rothfels, C.J.; Magnuson-Ford, K.; Barker, M.S.; Rieseberg, L.H.; Otto, S.P. Recently formed polyploid plants diversify at lower rates. Science 2011, 333, 1257. [Google Scholar] [CrossRef]
- Miyaji, Y. Untersuchungen über die Chromosomenzahlen bei einigen Viola-Arten. Bot. Mag. Tokyo 1913, 27, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Miyaji, Y. Studien über die Zahlenverhältnisse der Chromosomen bei der Gattung Viola. Cytologia 1929, 1, 28–58. [Google Scholar] [CrossRef]
- Marcussen, T.; Jakobsen, K.S.; Danihelka, J.; Ballard, H.E.; Blaxland , K.; Brysting, A.K.; Oxelman, B. Inferring species networks from gene trees in high-polyploid North American and Hawaiian violets (Viola, Violaceae). Syst. Biol. 2012, 61, 107–126. [Google Scholar] [CrossRef]
- Gingins de la Sarraz, F.C.J. Mémoire sur la famille des Violacées. Mém. Soc. Phys. Hist. Nat. Genève 1823, 2, 1–27. [Google Scholar]
- Gingins de la Sarraz, F.C.J.; de Candolle, A.P. Viola . In Prodromus Systematis Naturalis Regni Vegetabilis; De Candolle, A.P., Ed.; Treuttel et Würtz: Parisiis, France, 1824; Volume 1, pp. 291–306. [Google Scholar]
- Reiche, K.F.; Taubert, P. Violaceae. In Die Natürlichen Pflanzenfamilien, 3 (6); Engler, A., Prantl, K., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1895; pp. 322–336. [Google Scholar]
- Görz, R. Wilhelm Becker. Verh. Bot. Vereins Prov. Brandenbg. 1929, 71, 142–150. [Google Scholar]
- Wein, K. Wilhelm Becker †. Mitth. Thüring. Bot. Vereins N.F. 1930, 39, 14–16. [Google Scholar]
- Merrill, E.D. Destruction of the Berlin herbarium. Science 1943, 98, 490–491. [Google Scholar] [CrossRef]
- Hiepko, P. The collections of the Botanical Museum Berlin-Dahlem (B) and their history. Englera 1987, 7, 219–252. [Google Scholar] [CrossRef]
- Heilborn, O. Bidrag til Violaceernas cytologi. Sven. Bot. Tidskr. 1926, 20, 414–419. [Google Scholar]
- Gershoy, A. Studies in North American violets I. General considerations. Bull. Vermont Agric. Exp. Sta. 1928, 279, 3–18. [Google Scholar]
- Clausen, J. Genetical and cytological investigations on Viola tricolor L. and V. arvensis Murr. Berlingska Boktr. 1926, 8, 1–156. [Google Scholar] [CrossRef]
- Clausen, J. Chromosome number and the relationship of species in the genus Viola. Ann. Bot. 1927, 61, 677–714. [Google Scholar] [CrossRef]
- Gershoy, A. Studies in North American violets III. Chromosome numbers and species characters. Bull. Vermont Agric. Exp. Sta. 1934, 367, 3–91. [Google Scholar]
- Dodd, J.D.; Gershoy, A. Experiments in the grafting of species in the genus Viola. Bull. Torrey Bot. Club 1943, 70, 91–103. [Google Scholar] [CrossRef]
- Clausen, J. Cytotaxonomy and distributional ecology of western North American violets. Madroño 1964, 17, 173–197. [Google Scholar]
- Marcussen, T.; Blaxland , K.; Windham, M.D.; Haskins, K.E.; Armstrong, F. Establishing the phylogenetic origin, history and age of the narrow endemic Viola guadalupensis (Violaceae). Amer. J. Bot. 2011, 98, 1978–1988. [Google Scholar] [CrossRef]
- Espeut, M. Revision of the genus Viola L. (Violaceae) in the Russian Far East with notes on adjacent territories. Bot. Pac. 2020, 9, 3–52. [Google Scholar] [CrossRef]
- Sparre, B.B. Systematical and nomenclaturistical studies on the genus Viola. I. An attempt at a revision of the section “Chilenium”. Lilloa 1949, 17, 377–416. [Google Scholar]
- Sparre, B.B. Systematical and nomenclatorial studies on the genus Viola. II. Further annotations on the section Chilenium. Lilloa 1950, 23, 347–364. [Google Scholar]
- Merxmüller, H. Veilchenstudien I–IV. Phyton 1974, 16, 137–158. [Google Scholar]
- Merxmüller, H.; Lippert, W. Veilchenstudien V–VII. Mitt. Bot. Staatssamml. München 1977, 13, 503–534. [Google Scholar]
- Erben, M. Cytotaxonomische Untersuchungen an südosteuropäischen Viola-Arten der Section Melanium. Mitt. Bot. Staatssamml. München 1985, 21, 339–740. [Google Scholar]
- Gil-ad, N.L. Systematics of Viola subsection Boreali-Americanae. Boissiera 1997, 53, 1–130. [Google Scholar]
- Watson, J.M.; Flores, A.R.; Nicola, M.V.; Marcussen, T. Viola subgenus Andinium, Preliminary Monograph; Scottish Rock Garden Club with International Rock Gardener: Glasgow, UK, 2021; pp. 1–215. [Google Scholar]
- Brainerd, E. Violets of North America. Bull. Vermont. Agric. Exp. Station 1921, 224, 1–172. [Google Scholar]
- Baird, V.B. Wild Violets of North America; University of California Press: Berkeley, CA, USA, 1942; p. 225. [Google Scholar]
- Baehni, C.; Weibel, R. Violaceae. In Flora of Peru; Macbride, J.F., Ed.; Field Museum of Natural History, Botanical Series: Chicago, IL, USA, 1941; Volume 13 (part 4, number 1), pp. 56–82. [Google Scholar]
- Nikitin, V.V. The system of the genus Viola (Violaceae) of Eastern European and Caucasian flora. Bot. Zhurn. 1998, 83, 123–137. (In Russian) [Google Scholar]
- Valentine, D.H.; Merxmüller, H.; Schmidt, A. Viola L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; University Press: Cambridge, UK, 1968; Volume 2, pp. 270–282. [Google Scholar]
- Jacobs, M.; Moore, D.M. Violaceae. In Flora Malesiana; Naturalis journals and series: Leiden, The Netherlands, 1971; Volume 7, pp. 178–212. [Google Scholar]
- Wang, J.C.; Huang, T.C. Notes on the flora of Taiwan 9. The Viola L. Violaceae. Taiwania 1990, 35, 12–56. [Google Scholar] [CrossRef]
- Wang, C. Violaceae. Flora Reipublicae Pop. Sin. 1991, 51, 1–148. (In Chinese) [Google Scholar]
- Sun, K.; Wang, Q.; Wang, Y. Numerical taxonomic study of Viola in China. J. Northwest Normal Univ. (Nat. Sci. Ed.) 1997, 33, 69–73, (In Chinese, with English Abstract). [Google Scholar]
- Chen, Y.S.; Quin-er, Y.; Ohba, H.; Nikitin, V.V. Violaceae. In Flora of China; Science Press, Beijing and Missouri Botanical Garden Press: St. Louis, USA, 2007; Volume 13, pp. 74–111. [Google Scholar]
- Schmidt, A. Violaceae. In Flora Iranica; Rechinger, K.H., Ed.; Akademische Druck- und Verlagssantalt: Graz, Austria, 1992; Volume 169, pp. 1–29. [Google Scholar]
- Nicola, M.V. Viola L. In Flora Vascular de la República Argentina: Dicotyledoneae-Celastrales, Cucurbitales, Fagales, Malpighiales, Oxalidales; Zuloaga, F.O., Belgrano, M.J., Eds.; Instituto de Botánica Darwinion: San Isidro, Buenos Aires, Argentina, 2017; Volume 17, pp. 371–408. [Google Scholar]
- Ballard, H.E.; Sytsma, K.J. Evolution and biogeography of the woody Hawaiian violets (Viola, Violaceae): Arctic origins, herbaceous ancestry and bird dispersal. Evolution 2000, 54, 1521–1532. [Google Scholar] [CrossRef]
- Yoo, K.-O.; Jang, S.-K.; Lee, W.-T. Phylogeny of Korean Viola based on ITS sequences. Korean J. Plant Taxon. 2005, 35, 7–23. [Google Scholar] [CrossRef]
- Yoo, K.-O.; Jang, S.-K. Phylogenetic relationships of Viola based on matK and atpB-rbcL sequences of chloroplast DNA. Korean J. Plant Taxon. 2007, 37, 1–15. [Google Scholar] [CrossRef]
- Van den Hof, K.; Van den Berg, R.G.; Gravendeel, B. Chalcone synthase gene lineage diversification confirms allopolyploid evolutionary relationships of European rostrate violets. Mol. Biol. Evol. 2008, 25, 2099–2108. [Google Scholar] [CrossRef]
- Havran, J.C.; Sytsma, K.J.; Ballard, H.E. Evolutionary relationships, interisland biogeography, and molecular evolution in the Hawaiian violets (Viola: Violaceae). Amer. J. Bot. 2009, 96, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Zhou, J.S.; Zhang, Y.X.; Liang, G.X.; Chen, H.F.; Xing, F.W. Molecular systematics of genus Viola L. in China. J. Trop. Subtrop. Bot. 2010, 18, 633–642. [Google Scholar] [CrossRef]
- Liang, G.X.; Xing, F.W. Infrageneric phylogeny of the genus Viola (Violaceae) based on trnL-trnF, psbA-trnH, rpL16, ITS sequences, cytological and morphological data. Acta Bot. Yunn. 2010, 32, 477–488. [Google Scholar] [CrossRef]
- Marcussen, T.; Oxelman, B.; Skog, A.; Jakobsen, K.S. Evolution of plant RNA polymerase IV/V genes: Evidence of subneofunctionalization of duplicated NRPD2/NRPE2-like paralogs in Viola (Violaceae). BMC Evol. Biol. 2010, 10, 1–15. [Google Scholar] [CrossRef]
- Yoo, K.-O.; Jang, S.-K. Infrageneric relationships of Korean Viola based on eight chloroplast markers. J. Syst. Evol. 2010, 48, 474–481. [Google Scholar] [CrossRef]
- Fan, Q.; Chen, S.; Wang, L.; Chen, Z.; Liao, W. A new species and new section of Viola (Violaceae) from Guangdong, China. Phytotaxa 2015, 197, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi Shahrestani, M.; Marcussen, T.; Saeidi Mehrvarz, S.; Yousefi, N. Morphological and phylogenetic relationships within Viola sect. Sclerosium (Violaceae) in Iran. Flora 2015, 215, 67–74. [Google Scholar] [CrossRef]
- Małobęcki, A.; Marcussen, T.; Bohdanowicz, J.; Migdałek, G.; Słomka, A.; Kuta, E. Cleistogamy and phylogenetic position of Viola uliginosa (Violaceae) re-examined. Bot. J. Linn. Soc. 2016, 182, 180–194. [Google Scholar] [CrossRef]
- Blaxland , K.; Ballard, H.E.; Marcussen, T. Viola pluviae sp. nov. (Violaceae), a member of subsect. Stolonosae in the Pacific Northwest region of North America. Nord. J. Bot. 2018, 36, e01931. [Google Scholar] [CrossRef]
- Krause, S.; Kadereit, J.W. Identity of the Calcarata species complex in Viola sect. Melanium (Violaceae). Willdenowia 2020, 50, 195–206. [Google Scholar] [CrossRef]
- Cowie, R.H.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: Fact, fiction or speculation? Biol. Rev. 2022, 97, 640–663. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Yokota, M.; Higa, S.; Yoshioka, H.; Shimabuku, K.-I. Viola stoloniflora (Violaceae), a new species from the Ryukyus. Bot. Mag. Shokubutsu-Gaku-Zasshi 1988, 101, 1–8. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Żabicka, J.; Migdałek, G.; Żabicki, P.; Cubała, M.; Bohdanowicz, J.; Słomka, A.; Jedrzejczyk-Korycinska, M.; Sliwinska, E.; Sychta, K.; et al. Comprehensive characteristics and genetic diversity of the endemic Australian Viola banksii (section Erpetion, Violaceae). Aust. J. Bot. 2019, 67, 81–98. [Google Scholar] [CrossRef]
- Moore, D.M. Chromosome numbers of Falkland islands angiosperms. Br. Antarct. Surv. Bull. 1967, 14, 69–82. [Google Scholar]
- Becker, W. Violae andinae. Bot. Jahrb. Syst. Pflanzengesch. Pflanzengeogr. 1906, 37, 587–592. [Google Scholar]
- Becker, W. Violae novae Americanae meridionalis. Repert. Spec. Nov. Regni Veg. 1922, 18, 180–186. [Google Scholar] [CrossRef]
- Becker, W. Beiträge zur Veilchenflora Argentiniens und Chiles. Repert. Spec. Nov. Regni Veg. 1926, 23, 222–227. [Google Scholar]
- Huelsenbeck, J.P.; Nielsen, R.; Bollback, J.P. Stochastic mapping of morphological characters. Syst. Biol. 2003, 52, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.B. Pleistocene changes in the flora of the high tropical Andes. Paleobiology 1975, 1, 273–294. [Google Scholar] [CrossRef]
- Van der Hammen, T.; Cleef, A.M. High Altitude Tropical Biogeography; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Gregory-Wodzicki, K.M. Uplift history of the Central and Northern Andes: A review. Bull. Geol. Soc. Am. 2000, 112, 1091–1105. [Google Scholar] [CrossRef]
- Taylor, D.W. Paleobiogeographic relationships of Andean angiosperms of Cretaceous to Pliocene age. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1991, 88, 69–84. [Google Scholar] [CrossRef]
- Boschman, L.M. Andean mountain building since the Late Cretaceous: A paleoelevation reconstruction. Earth-Sci. Rev. 2021, 220, 103640. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; McFadden, R.; Morón, S.E.; Silva, C.A.; Restrepo-Moreno, S.; Ramírez, D.A.; Hoyos, N.; Wilson, J.; Farris, D.; et al. Evidence for middle Eocene and younger land emergence in central Panama: Implications for Isthmus closure. GSA Bull. 2012, 124, 780–799. [Google Scholar] [CrossRef]
- Carlquist, S. The biota of long-distance dispersal. V. Plant dispersal to pacific islands. Bull. Torrey Bot. Club 1967, 94, 129–162. [Google Scholar] [CrossRef]
- Kuta, E. Biosystematic studies on the genus Viola L; section Plagiostigma Godr. I. Karyological analysis of V. epipsila Ledeb.; V. palustris L. and their hybrids from Poland. Acta Biol. Crac. Ser. Bot. 1989, 31, 29–44. [Google Scholar]
- Kuta, E. Biosystematic studies on the genus Viola L; section Plagiostigma Godr. II. Embryological analysis of V. epipsila Ledeb.; V. palustris L. and their hybrids from Poland. Acta Biol. Crac. Ser. Bot. 1989, 31, 45–62. [Google Scholar]
- Schmidt, A. Zytotaxonomische Untersuchungen an europäischen Viola-Arten der Sektion Nomimium. Oesterr. Bot. Z. 1961, 108, 20–88. [Google Scholar] [CrossRef]
- Moore, D.M. Population studies on Viola lactea Sm. and its wild hybrids. Evolution 1959, 13, 318–332. [Google Scholar] [CrossRef]
- Marcussen, T.; Borgen, L. Species delimitation in the Ponto-Caucasian Viola sieheana complex, based on evidence from allozymes, morphology, ploidy levels, and crossing experiments. Plant Syst. Evol. 2011, 201, 183–196. [Google Scholar] [CrossRef]
- Marcussen, T.; Borgen, L. Allozymic variation and relationships within Viola subsection Viola (Violaceae). Plant Syst. Evol. 2000, 223, 29–57. [Google Scholar] [CrossRef]
- Yoshida, M.; Takeda, S.; Yokoyama, J. A new and noteworthy hybrid of Viola (Violaceae) from Iwate, northern Japan: Viola rostrata var. japonica × V. violacea var. makinoi. Acta Phytotax. Geobot. 2016, 67, 115–122. [Google Scholar] [CrossRef]
- Nikitin, V.V. New species and hybrids in the genus Viola L. (Violaceae)/Нoвые виды и гибриды рoда Viola L. (Violaceae). Novosti Sist. Vyssh. Rast. 2003, 35, 135–146. (In Russian) [Google Scholar]
- Gradstein, F.M.; Ogg, J.G.; Smith, A.G. A Geologic Time Scale 2004; Cambridge University Press: Cambridge, NY, UK, 2005; p. 589. [Google Scholar]
- Rice, A.; Šmarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Sanso, A.M.; Seo, M.N. Chromosomes of some Argentine angiosperms and their taxonomic significance. Caryologia 2005, 58, 171–177. [Google Scholar] [CrossRef]
- Reiche, K.F. Violae chilenses Ein Beitrag zur Systematik der Gattung Viola. Bot. Jahrb. 1893, 16, 405–452. [Google Scholar]
- Watson, J.M.; Flores, A.R.; Arroyo-Leuenberger, S.C. A new Viola (Violaceae) from the Argentinian Andes. Willdenowia 2019, 49, 35–41. [Google Scholar] [CrossRef]
- Maximowicz, C.J. Enumeratio Plantarum Hucusque in Mongolia Nec Non Adjacente Parte Turkestaniae Sinensis Lectarum. Fasc. 1. Thalamiflorae et Disciflorae; Typis Academiae Imperialis Scientiarum Petropolitanae: St. Petersburg, Russia, 1889. [Google Scholar]
- Blaxland , K. A new species of Viola (Violaceae) from south-west Turkey. Bot. J. Linn. Soc. 2004, 145, 505–509. [Google Scholar] [CrossRef]
- Chen, Y.S.; Yang, Q.E. A new species of Viola (Violaceae) from southern China. Bot. J. Linn. Soc. 2008, 158, 755–761. [Google Scholar] [CrossRef]
- Huang, Y.-S.; Kang, N.; Zhong, X.-J.; Liao, W.-B.; Fan, Q. A new species of Viola (Violaceae) from Guangdong Province, China. PhytoKeys 2021, 176, 67. [Google Scholar] [CrossRef]
- Ning, Z.-L.; Zeng, Z.-X.; Chen, L.; Xu, B.-Q.; Liao, J.-P. Viola jinggangshanensis (Violaceae), a new species from Jiangxi, China. Ann. Bot. Fenn. 2012, 49, 383–386. [Google Scholar] [CrossRef]
- Knoche, K.G.; Marcussen, T. Viola barhalensis (Violaceae), a new species from northeastern Turkey. Phytotaxa 2016, 275, 14–22. [Google Scholar] [CrossRef]
- Reiche, K.F. Viola. In Flora de Chile; Imprenta Cervantes: Santiago, Chile, 1896; Volume 1, pp. 137–162. [Google Scholar]
- Linnaeus, C. Species Plantarum; Laurentius Salvius: Holmiae [Stockholm], Sweden, 1753; Volume 2, pp. 561–1200. [Google Scholar]
- Borbás, V.V. Viola . In Synopsis der Deutschen und Schweizer Flora, Enthaltend die Genauer Bekannten Phanerogamischen Gewächse, So Wie die Cryptogamischen Gefäss-Pflanzen, Welche in Deutschland, der Schweiz, in Preussen und Istrien Wild Wachsen, 3rd ed.; Hallier, E.H., Brand , A., Eds.; O. R. Reisland: Leipzig, Germany, 1892; Volume 1, pp. 161–226. [Google Scholar]
- Kupffer, K.R. Violaceae. In Flora Caucasica Critica; Kusnezow, N.I., Busch, N.A., Fomin, A.V., Eds.; Tipografija K. Mattisena: Jurjev [Tartu], Russia, 1909; Volume 3, pp. 158–248. [Google Scholar]
- Gerstlauer, L. Vorschläge zur Systematik der einheimischen Veilchen. Ber. Bayer. Bot. Ges. Erforsch. Heim. Flora 1943, 26, 12–55. [Google Scholar]
- Barthélémy, D.; Caraglio, Y. Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann. Bot. 2007, 99, 375–407. [Google Scholar] [CrossRef]
- Seguí, J.; Lázaro, A.; Traveset, A.; Salgado-Luarte, C.; Gianoli, E. Phenotypic and reproductive responses of an Andean violet to environmental variation across an elevational gradient. Alp. Bot. 2018, 128, 59–69. [Google Scholar] [CrossRef]
- Holdsworth, M. Cleistogamy of Viola cunninghamii. Trans. Roy. Soc. N. Z. Bot. 1966, 3, 169. [Google Scholar]
- Evans, L.T. Chasmogamous flowering in Viola palustris L. Nature 1956, 178, 1301. [Google Scholar] [CrossRef]
- Mayers, A.M.; Lord, E.M. Comparative flower development in the cleistogamous species Viola odorata. I. A growth rate study. Am. J. Bot. 1983, 70, 1548–1555. [Google Scholar] [CrossRef]
- Mayers, A.M.; Lord, E.M. Comparative flower development in the cleistogamous species Viola odorata. II. An organographic study. Am. J. Bot. 1983, 70, 1556–1563. [Google Scholar] [CrossRef]
- Mayers, A.M.; Lord, E.M. Comparative flower development in the cleistogamous species Viola odorata. III. A histological study. Bot. Gaz. 1984, 145, 83–91. [Google Scholar] [CrossRef]
- Borgström, G. Formation of cleistogamic and chasmogamic flowers in wild violets as a photoperiodic response. Nature 1939, 144, 514–515. [Google Scholar] [CrossRef]
- Russell, N.H. Studies in the photoperiodic responses of violets (Viola). S. W. Naturalist 1960, 5, 177–186. [Google Scholar] [CrossRef]
- Sternberger, A.L.; Ruhil, A.V.S.; Rosenthal, D.M.; Ballard, H.E.; Wyatt, S.E. Environmental impact on the temporal production of chasmogamous and cleistogamous flowers in the mixed breeding system of Viola pubescens. PLoS ONE 2020, 15, e0229726. [Google Scholar] [CrossRef]
- Rubin, G.; Paolillo, D.J., Jr. Vascular and general anatomy of the rootstocks of three stemless Viola species. Ann. Bot. 1978, 42, 981–988. [Google Scholar] [CrossRef]
- Bağcı, Y.; Dinç, M.; Öztürk, M. Morphological, anatomical and ecological study on Turkish endemic Viola yildirimlii M. Dinç & Y. Bağcı. Int. J. Nat. Eng. Sci. 2008, 2, 1–5. [Google Scholar]
- Gavrilova, O.; Nikitin, V.V. Palynological characteristics of East European Viola species (Violaceae) according to scanning electron microscope data. Bot. Zhurn. 2012, 97, 100–115. [Google Scholar]
- Yousefi, N.; Saeidi Mehrvarz, S.; Marcussen, T. Anatomical studies on selected species of Viola (Violaceae). Nord. J. Bot. 2012, 30, 461–469. [Google Scholar] [CrossRef]
- Saeidi Mehrvarz, S.; Vafi, M.; Marcussen, T. Taxonomic and anatomical notes on Viola sect. Viola (Violaceae) in Iran. Wulfenia 2013, 20, 73–79. [Google Scholar]
- Mohammadi Shahrestani, M.; Saeidi Mehrvarz, S.; Marcussen, T.; Yousefi, N. Taxonomy and comparative anatomical studies of Viola sect Sclerosium (Violaceae) in Iran. Acta Bot. Gall. 2014, 161, 343–353. [Google Scholar] [CrossRef]
- Colombo, P.; Spadaro, V.; Raimondo, F.M. Morpho-anatomical analysis of Viola tineorum and V. ucriana (Violaceae) endemic to the mountains around Palermo (NW-Sicily). Bocconea 2007, 21, 233–247. [Google Scholar]
- Mereacre, A.; Toniuc, A.; Toma, C. Histo-anatomical observations regarding Viola L. Species in the Gârboavele reserve (county of Galaţi). Analele Şti. Univ. "Al. I. Cuza" Iaşi Sect. II A Biol. Veget. 2014, 60, 13–24. [Google Scholar]
- Pilberg, C.; Ricco, M.V.; Alvarez, M.A. Foliar anatomy of Viola maculata growing in Parque Nacional Los Alerces, Chubut, Patagonia, Argentina. Rev. Bras. De Farmacogn. 2016, 26, 459–463. [Google Scholar] [CrossRef]
- Chen, M.; Gao, M.-F.; Liu, L.-L.; Wang, L.-H.; Wang, K.-M.; Hou, Y.-T. Comparative study on micro-morphology of the leaf surfaces from the genus Viola L. (Violaceae) in Shandong. J. Wuhan Bot. Res. 2010, 28, 268–278. [Google Scholar]
- Meusel, H.; Kästner, A. Zur Wuchsform einiger Veilchenarten. Phyton 1974, 16, 127–135. [Google Scholar]
- Mark, P.J.L.; Wortley, A.H.; Furness, C.A. Not a shrinking violet: Pollen morphology of Violaceae (Malpighiales). Grana 2012, 51, 181–193. [Google Scholar] [CrossRef]
- Nadot, S.; Ballard, H.E.; Creach, J.B.; Dajoz, I. The evolution of pollen heteromorphism in Viola: A phylogenetic approach. Plant Syst. Evol. 2000, 223, 155–171. [Google Scholar] [CrossRef]
- Tomović, G.; Đurović, S.; Buzurović, U.; Niketić, M.; Milanović, Đ.; Mihailović, N.; Jakovljević, K. Accumulation of potentially toxic elements in Viola L. (Sect. Melanium Ging.) from the ultramafic and non-ultramafic soils of the Balkan Peninsula. Water Air Soil Pollut. 2021, 232, 46. [Google Scholar] [CrossRef]
- Aksoy, A. Accumulation of cadmium, copper and zinc in selected natural Viola taxa in Turkish Mediterranean serpentine soils. Anatol. J. Bot. 2021, 5, 138–143. [Google Scholar] [CrossRef]
- De Salas, M.F. Viola serpentinicola (Violaceae), a new Tasmanian species endemic to serpentinised ultramafic soils. Muelleria 2018, 36, 112–117. [Google Scholar] [CrossRef]
- Watson, J.M.; Cárdenas, M.; Flores, A.R.; Macaya, J.; Jimenez, H.G.; Barría, J. Viola gelida, a new, rare and vulnerable rosulate species from the high Andes of Atacama Region, Chile. Gayana Bot. 2013, 70, 390–394. [Google Scholar] [CrossRef]
- Watson, J.M.; Flores, A.R. Meet our most unique viola and its Andean habitat. A renamed new species endemic to the northern semi-desert Altiplano of Chile. Int. Rock Gard. 2020, 128, 3–52. [Google Scholar]
- Severne, B.C.; Brooks, R.R. A nickel-accumulating plant from Western Australia. Planta 1972, 103, 91–94. [Google Scholar] [CrossRef]
- Cole, M.M. Geobotanical and biogeochemical investigations in the sclerophyllous woodland and shrub associations of the Eastern Goldfield Area of Western Australia, with particular reference to the role of Hybanthus floribundus (Lindl.) F. Muell. as a nickel indicator and accumulator plant. J. Appl. Ecol. 1973, 10, 269–320. [Google Scholar] [CrossRef]
- Weerasinghe, H.A.S.; Iqbal, M.C.M. Plant diversity and soil characteristics of the Ussangoda serpentine site. J. Natl. Sci. Found. Sri Lanka 2011, 39, 355–363. [Google Scholar] [CrossRef]
- Fernando, E.S.; Quimado, M.O.; Doronila, A.I. Rinorea niccolifera (Violaceae), a new, nickel-hyperaccumulating species from Luzon Island, Philippines. PhytoKeys 2014, 37, 1–13. [Google Scholar] [CrossRef]
- Paul, A.; Gei, V.; Isnard, S.; Fogliani, B.; Echevarria, G.; Erskine, P.; Jaffré, T.; Munzinger, J.; van der Ent, A. Nickel hyperaccumulation in New Caledonian Hybanthus (Violaceae) and occurrence of nickel-rich phloem in Hybanthus austrocaledonicus. Ann. Bot. 2020, 126, 905–914. [Google Scholar] [CrossRef]
- Müller, H. The Fertilisation of Flowers. Translated and Edited by D’Arcy W. Thompson, B.A.; Scholar of Trinity College, Cambridge, with a Preface by Charles Darwin. with Illustrations; Macmillan and Co.: London, UK, 1883; pp. 1–669. [Google Scholar]
- Beattie, A.J. Floral evolution in Viola. Ann. Missouri Bot. Gard. 1974, 61, 781–793. [Google Scholar] [CrossRef]
- Culley, T.M. Reproductive biology and delayed selfing in Viola pubescens (Violaceae), an understory herb with chasmogamous and cleistogamous flowers. Int. J. Plant Sci. 2002, 163, 113–122. [Google Scholar] [CrossRef]
- Freitas, L.; Sazima, M. Floral biology and pollination in two Viola species—From nectar to pollen flowers? Ann. Bot. 2003, 91, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Son, H.D.; Im, H.T.; Hon, E.J.; Park, S.J. Species biology study of pollination and myrmecochory on two Viola species. Korean J. Nat. Conserv. 2008, 6, 143–147. (In Korean) [Google Scholar] [CrossRef]
- Murúa, M.; Espinoza, C.; Bustamante, R.; Marín, V.H.; Medel, R. Does human-induced habitat transformation modify pollinator-mediated selection? A case study in Viola portalesia (Violaceae). Oecologia 2010, 163, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, B.A.; Graham, C.H.; Aguilar, J.M.; Schleuning, M. Effects of hummingbird morphology on specialization in pollination networks vary with resource availability. Oikos 2017, 126, 52–60. [Google Scholar] [CrossRef]
- Herrera, C.M. Selection on floral morphology and environmental determinants of fecundity in a hawk moth-pollinated violet. Ecol. Monogr. 1993, 63, 251–275. [Google Scholar] [CrossRef]
- McPavlis. Viola delphinantha and its sisters. Int. Rock Gard. 2010, 2010, 11–12. [Google Scholar]
- Thiele, K.R.; Prober, S.M. New species and a new hybrid in the Viola hederacea species complex, with notes on Viola hederacea Labill. Muelleria 2003, 18, 7–25. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Bohdanowicz, J.; Cubała, M.; Słomka, A.; Żabicka, J.; Żabicki, P.; Migdałek, G.; Marcussen, T.; Thiele, K.; Kuta, E. A new pollination system in non-cleistogamous species of Viola results from nyctinastic (night-closing) petal movements—A mixed outcrossing-selfing strategy. Flora 2019, 253, 1–9. [Google Scholar] [CrossRef]
- Nylin, S.; Slove, J.; Janz, N. Host plant utilization, host range oscillations and diversification in Nymphalid butterflies: A phylogenetic investigation. Evolution 2014, 68, 105–124. [Google Scholar] [CrossRef]
- Simonsen, T.J. Fritillary phylogeny, classification, and larval host plants: Reconstructed mainly on the basis of male and female genitalic morphology (Lepidoptera: Nymphalidae: Argynnini). Biol. J. Linn. Soc. 2006, 89, 627–673. [Google Scholar] [CrossRef]
- Wahlberg, N.; Leneveu, J.; Kodandaramaiah, U.; Peña, C.; Nylin, S.; Freitas, A.V.L.; Brower, A.V.Z. Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proc. R. Soc. B Biol. Sci. 2009, 276, 4295–4302. [Google Scholar] [CrossRef] [PubMed]
- Chazot, N.; Wahlberg, N.; Freitas, A.V.L.; Mitter, C.; Labandeira, C.; Sohn, J.-C.; Sahoo, R.K.; Seraphim, N.; de Jong, R.; Heikkilä, M. Priors and posteriors in Bayesian timing of divergence analyses: The age of butterflies revisited. Syst. Biol. 2019, 68, 797–813. [Google Scholar] [CrossRef]
- De Moya, R.S.; Savage, W.K.; Tenney, C.; Bao, X.; Wahlberg, N.; Hill, R.I. Interrelationships and diversification of Argynnis Fabricius and Speyeria Scudder butterflies. Syst. Entomol. 2017, 42, 635–649. [Google Scholar] [CrossRef]
- Kovar-Eder, J.; Kvaček, Z.; Meller, B. Comparing Early to Middle Miocene floras and probable vegetation types of Oberdorf N Voitsberg (Austria), Bohemia (Czech Republic), and Wackersdorf (Germany). Rev. Palaeobot. Palynol. 2001, 114, 83–125. [Google Scholar] [CrossRef]
- Kovar-Eder, J.; Meller, B. Plant assemblages from the hanging wall sequence of the opencast mine Oberdorf N Voitsberg, Styria (Austria, Early Miocene, Ottnangian). Palaeontogr. Abt. B Paläophytol. 2001, 259, 65–112. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Wahlberg, N.; Warren, A.D.; Sperling, F.A.H. The evolutionary history of Boloria (Lepidoptera: Nymphalidae): Phylogeny, zoogeography and larval–foodplant relationships. Syst. Biodivers. 2010, 8, 513–529. [Google Scholar] [CrossRef]
- Oszast, J.; Stuchlik, L. The Neogene vegetation of the Podhale (West Carpathians, Poland) [Roślinność Podhala w neogenie]. Acta Palaeobot. 1977, 18, 45–86. [Google Scholar]
- Li, W.-Y.; Wu, H.-F. A palynological investigation on the Late Tertiary and Early Quaternary and its significance in the paleogeographical study in Central Yunnan. Acta Geogr. Sin. (Ti Li Lsueh Pao) 1978, 33, 142–155, (In Chinese with English Summary). [Google Scholar]
- Bolotnikova, M.D. Spore and Pollen Assemblages from Tertiary Deposits of the Western Shore of the Japanese Sea [Sporovo-Pyltsevye Kompleksy Tretichnykh Otlozheniy Zapadnogo Poberezhya Yaponskogo Morya]; Nauka: Moscow, Russia, 1979; pp. 1–195. [Google Scholar]
- Zheng, Y.-H.; Zhou, S.-F.; Liu, X.-Q.; Wang, L.-Y.; Xu, S.-J.; Wang, X.-Z. Neogene sporo-pollen grains from northern Jiangsu and South Yellow Sea Basin. Bull. Nanjing Inst. Geol. Palaeontol. (Acad. Sin.) 1981, 3, 29–90, (In Chinese with English Summary). [Google Scholar]
- Tomskaya, A.I. Palynology of Cenozoic Deposits of Yakutia [Palinologiya Kaynozoya Yakutii]; Nauka: Novosibirsk, Russia, 1981; pp. 1–221. [Google Scholar]
- Knobloch, E. Paleobotanic evidence on the age of the Plio-Pleistocene deposits between Mohelnice and Kroměříž [Paleobotanické doklady ke stáří pliocenních–pleistocenních uloženin mezi Mohelnicí a Kroměříží]. Věstn. Ústřed. Ústavu Geol. [Bull. Geol. Surv. Prague] 1987, 62, 25–33. [Google Scholar]
- Utescher, T.; Ivanov, D.; Harzhauser, M.; Bozukov, V.; Ashraf, A.R.; Rolf, C.; Urbat, M.; Mosbrugger, V. Cyclic climate and vegetation change in the late Miocene of western Bulgaria. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 272, 99–114. [Google Scholar] [CrossRef]
- Szafer, W. Flora plioceńska z Krościenka n/Dunajcem, II (The Pliocene flora of Krościenko in Poland). Rozpr. Wydz. Mat. Przyr. PAU Dz. B 1947, 72, 1–213. [Google Scholar]
- Szafer, W. Miocenska flora ze Starych Gliwic na Śląsku (Miocene Flora of Stare Gliwice in Upper Silesia). Pr. Inst. Geol. 1961, 33, 1–205. [Google Scholar]
- Dorofeev, P.I. Tretichnye flory zapadnoi Sibiri [The Tertiary floras of western Siberia]; Izdatel’stvo Akademii Nauk SSSR: Moscow/Leningrad, Russia, 1963; p. 343. [Google Scholar]
- Łańcucka-Środoniowa, M. Macroscopic plant remains from the freshwater Miocene of the Nowy Sącz Basin (West Carpathians, Poland)/Szczątki makroskopowe roślin z miocenu słodkowodnego Kotliny Sądeckiej (Karpaty Zachodnie, Polska). Acta Palaeobot. 1979, 20, 3–117. [Google Scholar]
- Van der Burgh, J. Allochthonous seed and fruit floras from the Pliocene of the lower Rhine Basin. Rev. Palaeobot. Palynol. 1983, 40, 33–90. [Google Scholar] [CrossRef]
- Van der Burgh, J. Miocene floras in the lower Rhenish Basin and their ecological interpretation. Rev. Palaeobot. Palynol. 1987, 52, 299–366. [Google Scholar] [CrossRef]
- Mai, D.H.; Walther, H. Die pliozänen Floren von Thüringen, Deutsche Demokratische Republik. Quartärpalaeontologie 1988, 7, 55–297. [Google Scholar]
- Mai, D.H. Die mittelmiozänen und obermiozänen Floren aus der Meuroer und Raunoer Folge in der Lausitz. III. Fundstellen und Palaeobiologie. Palaeontograph. Abt. B 2001, 258, 1–85. [Google Scholar]
- Arbuzova, O.N. Viola L. In Iskopaemye Tsvetkovye Rastenija Rossii I Sopredel’nyh Gosudarstv [Fossil Flowering Plants of Russia and Adjacent States]. Nyctaginaceae—Salicaceae; Budantsev, L., Ed.; Komarov Botanical Institute of the Russian Academy of Sciences: Moscow, Russia, 2005; Volume 4, pp. 48–52. [Google Scholar]
- Hollick, A. Plant remains from a Pleistocene lake deposit in the Upper Connecticut River Valley. Brittonia 1931, 1, 35–55. [Google Scholar] [CrossRef]
- Bennike, O. News from the Plio-Pleistocene Kap København Formation, North Greenland. Polar Res. 1987, 5, 339–340. [Google Scholar] [CrossRef]
- Farlow, J.O.; Sunderman, J.A.; Havens, J.J.; Swinehart, A.L.; Holman, J.A.; Richards, R.L.; Miller, N.G.; Martin, R.A.; Hunt, R.M.; Storrs, G.W.; et al. The Pipe Creek Sinkhole biota, a diverse late Tertiary continental fossil assemblage from Grant County, Indiana. Am. Midl. Nat. 2001, 145, 367–378. [Google Scholar] [CrossRef]
- Fletcher, T.L.; Telka, A.; Rybczynski, N.; Matthews, J.V., Jr. Neogene and early Pleistocene flora from Alaska, USA and Arctic/Subarctic Canada: New data, intercontinental comparisons and correlations. Palaeontol. Electron. 2021, 24, a08. [Google Scholar] [CrossRef]
- Mai, D.H. Zur Bedeutung von Relikten in der Florengeschichte. In 100-Jahre Arboretum Berlin Jubiläumsschrift; Vent, W., Ed.; Humboldt-Universität Berlin: Berlin, Germany, 1980; pp. 281–307. [Google Scholar]
- Nikitin, V.V. Trends of morphological evolution and relationships of taxa within genus Viola (Violaceae) of Eastern European and Caucasian Flora. Bot. Zhurnal 2002, 87, 31–55. (In Russian) [Google Scholar]
- Ciangherotti, A.; Esu, D.; Martinetto, E.; Giuntelli, P. The remarkable Middle Pliocene non-marine mollusc record from Ceresole d’Alba, Piedmont, north-west Italy: Biochronology, palaeobiogeography and palaeoecology supported by fossil plants. Geobios 2007, 40, 573–587. [Google Scholar] [CrossRef]
- Nemcok, M.; Pospisil, L.; Lexa, J.; Donelick, R.A. Tertiary subduction and slab break-off model of the Carpathian–Pannonian region. Tectonophysics 1998, 295, 307–340. [Google Scholar] [CrossRef]
- Babushkin, A.E.; Volkova, V.S.; Gnibidenko, Z.N. Unified Regional Stratigraphic Charts of Neogene and Paleogene Deposits of the West Siberian Plain and Explanatory Note; SNIIGGiMS: Novosibirsk, Russia, 2001; pp. 1–82. (In Russian) [Google Scholar]
- Schäfer, A.; Utescher, T.; Mörs, T. Stratigraphy of the Cenozoic Lower Rhine Basin, northwestern Germany. Newsl. Stratigr. 2004, 40, 73–110. [Google Scholar] [CrossRef]
- Bojňanský, V.; Fargašová, A. Atlas of Seeds and Fruits of Central and East-European Flora. In The Carpathian Mountains Region; Springer: Dordrecht, The Netherlands, 2007; pp. 1–1046. [Google Scholar]
- Gil-ad, N.L. The micromorphologies of seed coats and petal trichomes of the taxa of Viola subsect. Boreali-Americanae (Violaceae) and their utility in discerning orthospecies from hybrids. Brittonia 1998, 50, 91–121. [Google Scholar] [CrossRef]
- Beattie, A.J.; Lyons, N. Seed dispersal in Viola (Violaceae): Adaptations and strategies. Am. J. Bot. 1975, 62, 714–722. [Google Scholar] [CrossRef]
- Eckstein, R.L.; Otte, A. Effects of cleistogamy and pollen source on seed production and offspring performance in three endangered violets. Basic Appl. Ecol. 2005, 6, 339–350. [Google Scholar] [CrossRef]
- Yockteng, R.; Ballard, H.E.; Mansion, G.; Dajoz, I.; Nadot, S. Phylogenetic relationships among pansies (Viola section Melanium) investigated using ITS and ISSR markers. Plant Syst. Evol. 2003, 241, 153–170. [Google Scholar] [CrossRef]
- Šmarda, P.; Knápek, O.; Březinová, A.; Horová, L.; Grulich, V.; Danihelka, J.; Veselý, P.; Šmerda, J.; Rotreklová, O.; Bureš, P. Genome sizes and genomic guanine+cytosine (GC) contents of the Czech vascular flora with new estimates for 1700 species. Preslia 2019, 91, 117–142. [Google Scholar] [CrossRef]
- Pustahija, F.; Brown, S.C.; Bogunić, F.; Bašić, N.; Muratović, E.; Ollier, S.; Hidalgo, O.; Bourge, M.; Stevanović, V.; Siljak-Yakovlev, S. Small genomes dominate in plants growing on serpentine soils in West Balkans, an exhaustive study of 8 habitats covering 308 taxa. Plant Soil 2013, 373, 427–453. [Google Scholar] [CrossRef]
- Siljak-Yakovlev, S.; Pustahija, F.; Šolić, E.M.; Bogunić, F.; Muratović, E.; Bašić, N.; Catrice, O.; Brown, S.C. Towards a genome size and chromosome number database of Balkan Flora: C-Values in 343 taxa with novel values for 242. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Cires, E.; Cuesta, C.; Fernández Casado, M.A.; Nava, H.S.; Vázquez, V.M.; Fernández Prieto, J.A. Isolation of plant nuclei suitable for flow cytometry from species with extremely mucilaginous compounds: An example in the genus Viola L. (Violaceae). Anales Jard. Bot. Madrid 2011, 68, 139–154. [Google Scholar] [CrossRef]
- Gonzáles, P.; Cano, A. Two new species of Viola (Violaceae) named in honor of preceding Peruvian botanists. Phytotaxa 2016, 283, 83–90. [Google Scholar] [CrossRef]
- Gonzáles, P.; Molina-Alor, J. Viola tovarii (Violaceae), a new species for Peruvian high Andes. Phytotaxa 2020, 451, 245–250. [Google Scholar] [CrossRef]
- Nicola, M.V.; Salomón, L.; Zuloaga, F.O. Nomenclatural and taxonomic study in species of Viola (Violaceae) from Argentina. Phytotaxa 2018, 338, 26. [Google Scholar] [CrossRef]
- McCauley, R.A.; Ballard, H.E. Viola calcicola (Violaceae), a new endemic violet from the Guadalupe Mountains of New Mexico and Texas. J. Bot. Res. Inst. Texas 2013, 7, 9–20. [Google Scholar]
- Thiele, K.; De Salas, M.F.; Walsh, N.G.; Messina, A.; Little, R.J.; Prober, S.M. Viola curtisiae, a new rank for a poorly understood species, with notes on V. hederacea subsp. seppeltiana. Muelleria 2018, 36, 107–111. [Google Scholar] [CrossRef]
- Thiele, K.R.; Prober, S.M. Viola silicestris, a new species in Viola section Erpetion from Australia. Telopea 2006, 11, 99–104. [Google Scholar] [CrossRef]
- Okamoto, M. A new species of Viola collected in East Kalimantan, with notes on the seed dispersal of the allied species concerned. Bull. Osaka Mus. Nat. Hist. 1984, 37, 9–15. [Google Scholar]
- Okamoto, M.; Okada, H.; Ueda, K. Morphology and chromosome number of Viola pilosa, and its systematic position. Taxon 1993, 42, 781–787. [Google Scholar] [CrossRef]
- Okamoto, M.; Ueda, K. A new species of violet from Seram (Moluccas). Acta Phytotax. Geobot. 1986, 37, 1–8. [Google Scholar] [CrossRef]
- Li, X.-C.; Wang, Z.-W.; Wang, Q.; Ge, B.-J.; Chen, B.; Yu, P.; Zhong, X. Viola shiweii, a new species of Viola (Violaceae) from karst forest in Guizhou, China. PhytoKeys 2022, 196, 63–89. [Google Scholar] [CrossRef]
- Sternberger, A.L.; Bowman, M.J.; Kruse, C.P.S.; Childs, K.L.; Ballard, H.E.; Wyatt, S.E. Transcriptomics identifies modules of differentially expressed genes and novel cyclotides in Viola pubescens. Front. Plant Sci. 2019, 10, 165. [Google Scholar] [CrossRef]
- Lewin, H.A.; Richards, S.; Lieberman Aiden, E.; Allende, M.L.; Archibald, J.M.; Bálint, M.; Barker, K.B.; Baumgartner, B.; Belov, K.; Bertorelle, G.; et al. The Earth BioGenome Project 2020: Starting the clock. Proc. Natl. Acad. Sci. USA 2022, 119, e2115635118. [Google Scholar] [CrossRef]
- Slazak, B.; Jacobsson, E.; Kuta, E.; Göransson, U. Exogenous plant hormones and cyclotide expression in Viola uliginosa (Violaceae). Phytochemistry 2015, 117, 527–536. [Google Scholar] [CrossRef]
- Gao, J.; Luo, M.; Zhu, Y.; He, Y.; Wang, Q.; Zhang, C. Transcriptome sequencing and differential gene expression analysis in Viola yedoensis Makino (Fam. Violaceae) responsive to cadmium (Cd) pollution. Biochem. Biophys. Res. Commun. 2015, 459, 60–65. [Google Scholar] [CrossRef]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Brainerd, E. Viola [Tourn.] L. In An Illustrated Flora of the Northern United States, Canada and the British Possessions: From Newfoundland to the Parallel of the Southern Boundary of Virginia, and from the Atlantic Ocean Westward to the 102d Meridian; Britton, N.L., Brown, A., Eds.; C. Scribner’s Sons: New York, NY, USA, 1913; Volume 2, pp. 545–563. [Google Scholar]
- Grey-Wilson, C. Notes on African Violaceae. Kew Bull. 1981, 36, 103–126. [Google Scholar] [CrossRef]
- Morton, J.K. Chromosome numbers and polyploidy in the flora of Cameroon Mountain. Opera Bot. 1993, 121, 159–172. [Google Scholar]
- Gizaw, A.; Brochmann, C.; Nemomissa, S.; Wondimu, T.; Masao, C.A.; Tusiime, F.M.; Abdi, A.A.; Oxelman, B.; Popp, M.; Dimitrov, D.; et al. Colonization and diversification in the African ‘sky islands’: Insights from fossil-calibrated molecular dating of Lychnis (Caryophyllaceae). New Phytol. 2016, 211, 719–734. [Google Scholar] [CrossRef]
- Stebbins, G.L.; Harvey, B.L.; Cox, E.L.; Rutger, J.N.; Jelencovic, G.; Yagil, E. Identification of the ancestry of an amphiploid Viola with the aid of paper chromatography. Am. J. Bot. 1963, 50, 830–839. [Google Scholar] [CrossRef]
- Fabijan, D.M.; Packer, J.G.; Denford, K.E. The taxonomy of the Viola nuttallii complex. Can. J. Bot. 1987, 65, 2562–2580. [Google Scholar] [CrossRef]
- Probatova, N.S.; Barkalov, V.Y.; Stepanov, N.V. Chromosome numbers in some vascular plant species from Siberia and the Russian Far East. Bot. Pac. 2017, 61, 51–55. [Google Scholar] [CrossRef]
- Melchior, H.; Cuatrecasas, J. La Viola cazorlensis, su distribución, sistemática y biología. Cavanillesia 1935, 7, 133–148. [Google Scholar]
- Schmidt, A. Zur systematischen Stellung von Viola chelmea Boiss. et Heldr. ssp. chelmea und V. delphinantha. Boiss. Ber. Deutsch. Bot. Ges. 1964, 77, 256–261. [Google Scholar]
- Smith-White, S. Cytological evolution in the Australian flora. Cold Spring Harb. Symp. Quant. Biol. 1959, 24, 273–289. [Google Scholar] [CrossRef]
- Sun, K.; Wang, C.-j. Notes on the Chinese Violaceae (II)—Cytological studies on some violets of China. Bull. Bot. Res. 1991, 11, 69–72. [Google Scholar]
- Becker, W. Violae Asiaticae et Australenses. II. Beih. Bot. Centralbl. Abt. 2 1917, 34, 373–433. [Google Scholar]
- Nieuwland , J.A.; Kaczmarek, R.M. Studies in Viola. I. Proposed segregates of Viola. Am. Midl. Nat. 1914, 3, 207–217. [Google Scholar] [CrossRef]
- Słomka, A.; Godzik, B.; Szarek-Łukaszewska, G.; Shuka, L.; Hoef-Emden, K.; Bothe, H. Albanian violets of the section Melanium, their morphological variability, genetic similarity and their adaptations to serpentine or chalk soils. J. Plant Physiol. 2015, 174, 110–123. [Google Scholar] [CrossRef]
- Słomka, A.; Wolny, E.; Kuta, E. Viola tricolor (Violaceae) is a karyologically unstable species. Plant Biosyst. 2014, 148, 602–608. [Google Scholar] [CrossRef]
- Scoppola, A.; Cardoni, S.; Marcussen, T.; Simeone, M.C. Complex scenarios of reticulation, polyploidization, and species diversity within annual pansies of subsect. Bracteolatae (Viola sect. Melanium, Violaceae) in Italy: Insights from 5S-IGS high-throughput sequencing and plastid DNA variation. Plants 2022, 11, 1294. [Google Scholar]
- Leitch, I.J.; Johnston, E.; Pellicer, J.; Hidalgo, O.; Bennett, M.D. Angiosperm DNA C-Values Database, Release 9.0. 2019. Available online: https://cvalues.science.kew.org/ (accessed on 1 January 2022).
- Clausen, J. Cyto-genetic and taxonomic investigations on Melanium violets. Hereditas 1931, 15, 219–308. [Google Scholar] [CrossRef]
- Erben, M. The significance of hybridization on the forming of species in the genus Viola. Bocconea 1996, 5, 113–118. [Google Scholar]
- Clausen, J.; Channel, R.B.; Nur, U. Viola rafinesquii, the only Melanium violet native to North America. Rhodora 1964, 66, 32–46. [Google Scholar]
- Shinners, L.H. Viola rafinesquii: Nomenclature and native status. Rhodora 1961, 63, 327–335. [Google Scholar]
- Kartesz, J.T.; Gandhi, K.N. Nomenclatural notes for the North American flora. III. Phytologia 1990, 69, 129–137. [Google Scholar]
- Casazza, G.; Grassi, F.; Zecca, G.; Minuto, L. Phylogeographic insights into a peripheral refugium: The importance of cumulative effect of glaciation on the genetic structure of two endemic plants. PLoS ONE 2016, 11, e0166983. [Google Scholar] [CrossRef]
- Roux, J.P. Flora of South Africa; South African Biodiversity Institute (SANBI): Gauteng, South Africa, 2003; Available online: https://plants.jstor.org/collection/FLOSA (accessed on 1 January 2022).
- Hooker, J.D. The Botany of the Antarctic Voyage of H.M. Discovery Ships Erebus and Terror in the Years 1839–1843; Reeve Brothers: London, UK, 1844; Volume 2, p. 1. [Google Scholar]
- Hair, J.B.; Beuzenberg, E.J. Contributions to a chromosome atlas of the New Zealand flora—2. Miscellaneous families. N. Z. J. Sci. (Wellington) 1959, 2, 148–156. [Google Scholar]
- Laing, R.M.; Blackwell, E.W. Plants of New Zealand ; Whitcombe and Tombs, Ltd.: Christchurch, New Zealand , 1906; Volume 1906. [Google Scholar]
- Brainerd, E. Some natural violet hybrids of North America. Bull. Vermont Agric. Exp. Sta. 1924, 239, 1–255. [Google Scholar]
- Dakskobler, I.; Trnkoczy, I. Viola sororia Willd. = V. cucullata auct.; non Aiton = Viola obliqua Hill. Hladnikia 2010, 25, 53–55. [Google Scholar]
- Maslo, S.; Sarajlić, N.; Kotrošan, D. Viola sororia Willd. (Violaceae): New alien species in the flora of Bosnia and Herzegovina. Glas. Hrvat. Bot. Druš. 2018, 6, 20–24. [Google Scholar]
- Wirth, T.; Kovács, D.; Csiky, J. Contributions to the escaped, naturalised and potentially invasive species of the Hungarian adventive flora. Kitaibelia 2020, 25, 111–156. [Google Scholar] [CrossRef]
- Vitek, E.; Danihelka, J.; Adler, W. Viola ×palmata (Violaceae) new for Austria and Europe. Ann. Naturhist. Mus. Wien B 2012, 113, 264–265. [Google Scholar]
- Taylor, R.L.; Mulligan, G.A. Flora of the Queen Charlotte Islands. Part 2. Cytological Aspects of the Vascular Plants; Roger Duhamel, F.R.S.C.: Ottawa, ON, Canada, 1968. [Google Scholar]
- Nishikawa, T. Chromosome counts of flowering plants of Hokkaido (11). J. Hokkaido Univ. Educ. Sect. IIB 1988, 38, 33–40. [Google Scholar]
- Sokolovskaya, A.P. [Geographical distribution of polyploidy in plants: Investigation of the flora of Sakhalin]. Vestn. Leningr. Univ. Ser. Biol. 1960, 4, 42–58. (In Russian) [Google Scholar]
- Sokolovskaya, A.P. [Geographical distribution of polyploidy in plants: Investigation of the flora of the Kamchatka Peninsula]. Vestn. Leningr. Univ. Ser. Biol. 1963, 15, 38–52. (In Russian) [Google Scholar]
- Volkova, S.A.; Boyko, E.V.; Antusheva, I.V. [Chromosome numbers of some species of Kamchatka and Sakhalin flora]. Bot. Zhurnal 2003, 88, 154–155. (In Russian) [Google Scholar]
- Probatova, N.S.; Barkalov, V.Y.; Rudyka, E.G. Caryology of the Flora of Sakhalin and the Kurile Islands: Chromosome Numbers, Taxononomic and Phytogeographical Comments; Dal’nauka: Vladivostok, Russia, 2007. [Google Scholar]
- Baker, M.S. Studies in western violets–III. Madroño 1940, 5, 218–231. [Google Scholar]
- Baker, M.S. Studies in western violets–II. New species and varieties. Madroño 1936, 3, 232–239. [Google Scholar]
- Becker, W.; Hultén, E. The genus Viola in Kamtchatka. Ark. Bot. 1929, 22, 1–8. [Google Scholar]
- Carlquist, S. Comparative Wood Anatomy: Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood, 2nd ed.; Springer Series in Wood Science; Timell, T.E., Ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Shaw, K.L.; Gillespie, R.G. Comparative phylogeography of oceanic archipelagos: Hotspots for inferences of evolutionary process. Proc. Natl. Acad. Sci. USA 2016, 113, 7986–7993. [Google Scholar] [CrossRef]
- Becker, W.A.; Ewart, L.C. Pollination, seed set and pollen tube growth investigations in Viola pedata L. Acta Hortic. 1990, 272, 33–36. [Google Scholar] [CrossRef]
- Su, X.; Hou, Y.-Y.; Gao, T.; Wang, Y.-L.; Sun, K. Taxonomic position of Viola tuberifera based on morphological variation of the populations and digital specimen. Bull. Bot. Res. 2018, 38, 189–194. [Google Scholar] [CrossRef]
- Zhou, J.S.; Xing, F.W. Viola changii sp. nov. (Violaceae) from Guangdong, southern China. Nord. J. Bot. 2007, 25, 303–305. [Google Scholar] [CrossRef]
- Zhou, J.S.; Qin, G.; Xing, F.W. Viola nanlingensis (Violaceae), a new species from Guangdong, southern China. Acta Bot. Fenn. 2008, 45, 233–236. [Google Scholar] [CrossRef]
- Dong, A.Q.; Zhou, J.S.; Gong, Q.; Xing, F.W. A new species of Viola (Violaceae) from Guangdong, China. Novon 2009, 19, 457–470. [Google Scholar] [CrossRef]
- Huang, Y.-S.; (Sun Yat-sen University, Guangzhou, China). Personal communication, 2022.
- Becker, W. Violae Asiaticae et Australenses VI. Beih. Bot. Centralbl. Abt. 2 1924, 40, 119–171. [Google Scholar]
- Hu, H.-H.; Liu, B.; Liang, Y.-S.; Ye, J.-F.; Saqib, S.; Meng, Z.; Lu, L.-M.; Chen, Z.-D. An updated Chinese vascular plant tree of life: Phylogenetic diversity hotspots revisited. J. Syst. Evol. 2020, 58, 663–672. [Google Scholar] [CrossRef]
- Nakamura, K.; Denda, T.; Kokubugata, G.; Huang, C.-J.; Peng, C.-I.; Yokota, M. Phylogeny and biogeography of the Viola iwagawae-tashiroi species complex (Violaceae, section Plagiostigma) endemic to the Ryukyu Archipelago, Japan. Plant Syst. Evol. 2015, 301, 337–351. [Google Scholar] [CrossRef]
- Russell, N.H. The taxonomy of the North American acaulescent white violets. Am. Midl. Nat. 1955, 54, 481–494. [Google Scholar] [CrossRef]
- McKinney, L.E.; Russell, N.H. Violaceae of the Southeastern United States. Castanea 2002, 67, 369–379. [Google Scholar]
- Little, R.J.; McKinney, L.E. Violaceae Batsch. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA; Oxford, UK, 2016; Volume 6, Available online: http://www.efloras.org (accessed on 1 January 2022).
- Aedo, C.; Gallo, M.L. Sobre la pretendida subsp. “juressi” (Link. ex K. Wein) Coutinho de “Viola palustris” L. Anales Jard. Bot. Madrid 1991, 49, 145–146. [Google Scholar]
- Fernández Casado, M.d.l.A.; Nava Fernández, H.S. Variabilidad morfológica de Viola palustris L. s.l. en Asturias y territorios limítrofes (norte de España). Bol. Ci. Naturaleza I. D. E. A. 2015, 53, 189–198. [Google Scholar]
- Chen, Y.S.; Yang, Q.E. Two new stoloniferous species of Viola (Violaceae) from China. Bot. J. Linn. Soc. 2009, 159, 349–356. [Google Scholar] [CrossRef]
- Becker, W. Violae Asiaticae et Australenses. III. Beih. Bot. Centralbl. Abt. 2 1918, 36, 15–59. [Google Scholar]
- Bhellum, B.L.; Magotra, R. Viola kouliana (Violaceae: Section Sclerosium), a new species from India. Nord. J. Bot. 2022, e03648, in press. [Google Scholar] [CrossRef]
- Gilbert, M.G. Notes on Violaceae from Ethiopia. Nord. J. Bot. 1992, 12, 689–693. [Google Scholar] [CrossRef]
- Kroon, D.; Steens, T.; Troelstra, S.R. Onset of monsoonal related upwelling in the western Arabian sea as revealed by planktonic foraminifers. Proc. Ocean Drill. Program Sci. Results 1991, 117, 257–263. [Google Scholar] [CrossRef]
- Prell, W.L.; Kutzbach, J.E. Sensitivity of the Indian monsoon to forcing parameters and implications for its evolution. Nature 1992, 360, 647–652. [Google Scholar] [CrossRef]
- Kausel, E. Lista de las Mirtaceas y Leptospermaceas Argentinas. Lilloa 1966, 32, 323–368. [Google Scholar]
- Nikitin, V.V. Violaceae. In Flora Europae orientalis/Flora vostochnoj Evropy: Magnoliopsida; Tzvelev, N.N., Ed.; Nauka: St. Petersburg, Russia, 1996; Volume 9, pp. 180–205. [Google Scholar]
- Moore, D.M.; Harvey, M.J. Cytogenetic relationships of Viola lactea Sm. and other West European arosulate species. New Phytol. 1961, 60, 85–95. [Google Scholar]
- Eckstein, R.; O’Neill, R.; Danihelka, J.; Otte, A.; Kohler, W. Genetic structure among and within peripheral and central populations of three endangered floodplain violets. Mol. Ecol. 2006, 15, 2367–2379. [Google Scholar] [CrossRef]
- Eckstein, R.L.; Hölzel, N.; Danihelka, J. Biological Flora of Central Europe: Viola elatior, V. pumila, and V. stagnina. Perspect. Plant Ecol. Evol. Syst. 2006, 8, 45–66. [Google Scholar] [CrossRef]
- Peng, C.-I. Digital Flora of Taiwan. Available online: http://www.efloras.org (accessed on 1 January 2022).
- Greuter, W.; Rechinger, K.H. Flora der Insel Kythera: Gleichzeitig Beginn einer Nomenklatorischen Überprüfung der Griechischen Gefässpflanzenarten; Boissiera: Mémoires de Botanique Systématique, 1967; pp. 1–206. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://www.plantsoftheworldonline.org/ (accessed on 8 July 2021).
- African Plant Database, Version 3.4.0; Conservatoire et Jardin botaniques de la Ville de Genève and South African National Biodiversity Institute: Pretoria, South Africa, 2022; Available online: https://africanplantdatabase.ch (accessed on 1 January 2022).
- Zuev, V.V. Semejstvo Violaceae—Fialkovye [Family Violaceae]. In Flora Sibiri [Flora of Siberia]; Peschkova, G.A., Ed.; Nauka: Novosibirsk, Russia, 1996; Volume 10, pp. 83–101. (In Russian) [Google Scholar]
- Baikov, K.S. Conspectus Florae Sibiriae. Plantae Vasculares; Nauka: Novosibirsk, Russia, 2005; pp. 1–361. (In Russian) [Google Scholar]
- Bezdeleva, T.A. Fialkovye—Violaceae Batsch. In Plantae Vasculares Extremi Orienti Sovietici/Sosudistye Rastenija Sovetskogo Dal’nego Vostoka; Charkevicz, S.S., Ed.; Nauka: Leningrad, Russia, 1987; Volume 2, pp. 93–131. (In Russian) [Google Scholar]
- Nikitin, V.V. Violaceae Batsch, nom. cons. In Caucasian Flora Conspectus; Takhtajan, A.L., Ed.; KMK Scientific Press: Saint-Peterburg & Moscow, Russia, 2012; Volume 3, pp. 338–349. (In Russian) [Google Scholar]
- Tison, J.-M.; Foucalt, B. Flora Gallica: Flore de France; Biotope Éditions: Mèze, France, 2014. [Google Scholar]
- Raab-Straube, E.V.; Henning, T. Violaceae. In Euro+Med Plantbase—the Information Resource for Euro-Mediterranean Plant Diversity. Available online: https://www.europlusmed.org/ (accessed on 15 August 2021).
- Ballard, H.E. Violets (Violaceae) of the Great Plains and Eastern North America. Available online: https://people.ohio.edu/ballardh/vgpena (accessed on 1 January 2022).
- Sanso, A.M.; Xifreda, C.C.; Seo, M.N. Viola L. In Catálogo de las plantas vasculares del Cono Sur (Argentina, Sur de Brasil, Chile, Paraguay y Uruguay). Dicotyledoneae: Fabaceae (Senna-Zygia)–Zygophyllaceae 3. Monographs in Systematic Botany from the Missouri Botanical Garden; Zuloaga, F.O., Morrone, O., Belgrano, M.J., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; Volume 107, pp. 3158–3169. [Google Scholar]
- Ballard, H.E.; Wujek, D.E. Evidence for the recognition of Viola appalachiensis. Syst. Bot. 1994, 19, 523–538. [Google Scholar] [CrossRef]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)—A community resource of plant chromosome numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef]
- GBIF: The Global Biodiversity Information Facility. What Is GBIF? Available online: https://www.gbif.org/what-is-gbif (accessed on 13 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 12 February 2022).
- Chamberlain, S.A.; Boettiger, C. R Python, and Ruby clients for GBIF species occurrence data. PeerJ Preprints 2017, 5, e3304v1. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Etten, J.V.; Raster: Geographic Analysis and Modeling with Raster Data. R Package Version 2.0-12. 2012. Available online: http://CRAN.R-project.org/package=raster (accessed on 12 December 2021).
- Töpel, M.; Zizka, A.; Calió, M.F.; Scharn, R.; Silvestro, D.; Antonelli, A. SpeciesGeoCoder: Fast categorization of species occurrences for analyses of biodiversity, biogeography, ecology, and evolution. Syst. Biol. 2016, 66, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bivand , R.; Lewin-Koh, N.; Pebesma, E.; Archer, E.; Baddeley, A.; Bearman, N.; Bibiko, H.-J.; Brey, S.; Callahan, J.; Carrillo, G. Maptools: Tools for Handling Spatial Objects. R Package, Version 0.9. 2019. Available online: https://cran.r-project.org/web/packages/maptools/index.html (accessed on 8 November 2021).
- Keitt, T.H.; Bivand , R.; Pebesma, E.; Rowlingson, B.; Rgdal: Bindings for the Geospatial Data Abstraction Library. R Package, Version 0.6-27. 2010. Available online: https://cran.r-project.org/web/packages/rgdal/index.html (accessed on 13 December 2021).
- Cooper, N. Package ‘Reader’: Suite of Functions to Flexibly Read Data from Files. 2017. Available online: https://CRAN.R-project.org/package=reader (accessed on 16 June 2017).
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence-based phylogenetic analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Müller, K. SeqState—Primer design and sequence statistics for phylogenetic DNA data sets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer v1.5. 2009. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 10 October 2018).
- Rambaut, A. FigTree v1.4. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 6 December 2018).
- Huber, K.T.; Oxelman, B.; Lott, M.; Moulton, V. Reconstructing the evolutionary history of polyploids from multilabeled trees. Mol. Biol. Evol. 2006, 23, 1784–1791. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Havran, J.C.; Oppenheimer, H.; Keaton, J.; Piotrowski, K. Interisland range expansion of Viola lanaiensis (Violaceae: Malpighiales), an endangered Hawaiian violet. Pac. Sci. 2012, 66, 447–456. [Google Scholar] [CrossRef]
- Havran, J.C.; Harbin, S.C.; Portner, T. Viola kauaensis var. hosakae (Violaceae), a new variety of endemic Hawaiian violet. PhytoKeys 2014, 39, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
Figure 1.
Illustration of Viola odorata in the herbal of Matthiolus, printed in 1562 [8]. At least partly, the foliage appears to represent the common hybrid V. hirta × odorata. In the accompanying text (fol. CCCLIIII) the rooting stolons of V. odorata are described and compared to those of Fragaria and Pilosella officinarum.
Figure 1.
Illustration of Viola odorata in the herbal of Matthiolus, printed in 1562 [8]. At least partly, the foliage appears to represent the common hybrid V. hirta × odorata. In the accompanying text (fol. CCCLIIII) the rooting stolons of V. odorata are described and compared to those of Fragaria and Pilosella officinarum.

Figure 2.
Global distribution of Viola L. (Violaceae), showing the predominantly temperate distribution of the genus.
Figure 2.
Global distribution of Viola L. (Violaceae), showing the predominantly temperate distribution of the genus.

Figure 3.
Photos of representative members of the sections and subsections of Viola.—(a–l) Subg. Neoandinium.—(m–av) Subg. Viola.—(a) Sect. Confertae: V. nassauvioides.—(b) Sect. Ericoidium: V. fluehmannii (photo © John M. Watson).—(c) Sect. Grandiflos: V. cheeseana (photo © John M. Watson).—(d) Sect. Inconspicuiflos: V. weibelii (photo © John M. Watson).—(e) Sect. Inconspicuiflos: V. lilliputana (photo © Harvey E. Ballard).—(f) Sect. Relictium: V. dandoisiorum (photo © John M. Watson).—(g) Sect. Rhizomandinium: V. escondidaensis (photo © John M. Watson).—(h) Sect. Rosulatae: V. volcanica (photo © Ana R. Flores).—(i) Sect. Sempervivum: V. atropurpurea (photo © John M. Watson).—(j) Sect. Subandinium: V. subandina (photo © Ana R. Flores).—(k) Sect. Triflabellium: V. triflabellata (photo © Ana R. Flores).—(l) Sect. Xylobasis: V. beati (photo © Ana R. Flores).—(m) Sect. Abyssinium: V. abyssinica (photo © Robert v. Blittersdorff).—(n) Sect. Chamaemelanium: V. pubescens (photo © Kim Blaxland).—(o) Sect. Chilenium: V. maculata (photo © Instituto de Botánica Darwinion).—(p) Sect. Danxiaviola: V. hybanthoides (photo © Qiang Fan).—(q) Sect. Delphiniopsis: V. cazorlensis (photo © Santiago Martín-Bravo).—(r) Sect. Erpetion: V. banksii (photo © Dewald du Plessis).—(s) Sect. Himalayum: V. kunawurensis (photo © Vladimir Epiktetov).—(t) Sect. Leptidium: V. boliviana (photo © Sam Wilson).—(u) Sect. Melanium, subsect. Bracteolatae: V. tricolor and V. arvensis (photo © Thomas Marcussen).—(v) Sect. Melanium, subsect. Cleistogamae: V. rafinesquei (photo © Mary Ann Yaich).—(w) Sect. Melanium, subsect. Dispares: V. dyris (photo © Alan Keohane).—(x) Sect. Melanium, subsect. Ebracteatae: V. parvula (photo © Albert Keshet).—(y) Sect. Melanium, subsect. Pseudorupestres: V. nummulariifolia (photo © Sylvain Piry).—(z) Sect. Melvio: V. decumbens (photo © Magriet Brink).—(aa) Sect. Nematocaulon: V. filicaulis (photo © Andrew Townsend).—(ab)Sect. Nosphinium, subsect. Borealiamericanae: V. fimbriatula (photo © Kim Blaxland).—(ac) Sect. Nosphinium, subsect. Clausenianae: V. clauseniana (photo © Thomas Marcussen).—(ad) Sect. Nosphinium, subsect. Langsdorffianae: V. langsdorffii (photo © Jonathan Goff).—(ae) Sect. Nosphinium, subsect. Mexicanae: V. nannei (photo © Neptalí Ramírez Marcial).—(af) Sect. Nosphinium, subsect. Nosphinium: V. maviensis (photo © Karl Magnacca).—(ag) Sect. Nosphinium, subsect. Pedatae: V. pedata (photo © Kim Blaxland).—(ah) Sect. Plagiostigma, subsect. Australasiaticae: V. sumatrana (photo © Mário Duchoň).—(ai) Sect. Plagiostigma, subsect. Bilobatae: V. hamiltoniana (photo © Toshihiro Nagata).—(aj) Sect. Plagiostigma, subsect. Bulbosae: V. tuberifera (photo © Masashi Igari).—(ak) Sect. Plagiostigma, subsect. Diffusae: V. huizhouensis (photo © Yanshuang Huang).—(al) Sect. Plagiostigma, subsect. Formosanae: V. formosana (photo © Kuan-Chieh (Chuck) Hung).—(am) Sect. Plagiostigma, subsect. Patellares: V. tokubuchiana (photo © Masashi Igari).—(an) Sect. Plagiostigma, subsect. Stolonosae: V. pluviae (photo © Kim Blaxland).—(ao) Sect. Rubellium: V. rubella (photo © Pablo Silva).—(ap) Sect. Sclerosium: V. cinerea (photo © Jerome Viard).—(aq) Sect. Spathulidium: V. pachyrrhiza (photo © Dieter Zschummel).—(ar) Sect. Tridens: V. tridentata (photo © larsonek).—(as) Sect. Viola, subsect. Rostratae: V. canina (photo © Thomas Marcussen).—(at) Sect. Viola, subsect. Viola: V. collina (photo © Thomas Marcussen).—(au) Sect. Xanthidium: V. flavicans (photo © Instituto de Botánica Darwinion).—(av) Sect. Xylinosium: V. arborescens (photo © Abdelmonaim Homrani Bakali).—All images used in this figure were cropped and gamma-corrected.—Links to the online sources for the images used in this figure under a creative commons (CC) licence: V. abyssinica, http://www.westafricanplants.senckenberg.de/root/index.php?page_id=47&id=2203#image=22051, (accessed on 5 May 2022) © Robert v. Blittersdorff; V. arborescens, https://www.teline.fr/en/photos/violaceae/viola-arborescens#photo-7 (accessed on 11 May 2022), © Abdelmonaim Homrani Bakali, CC BY-NC 4.0; V. nummulariifolia, https://www.inaturalist.org/observations/89240299 (accessed on 5 May 2022), © Sylvain Piry, CC BY-NC 4.0; V. banksii, https://inaturalist.nz/observations/25048808 (accessed on 12 May 2022), © Dewald du Plessis, CC BY-NC 4.0; V. boliviana, https://ecuador.inaturalist.org/photos/57308380 (accessed on 5 May 2022), © Sam Wilson, CC BY-NC; V. cazorlensis, https://www.inaturalist.org/observations/109331467 (accessed on 5 May 2022), © Santiago Martín-Bravo, CC BY-NC 4.0; V. cinerea, https://www.inaturalist.org/observations/65108391 (accessed on 5 May 2022), © Jerome Viard, CC BY-NC 4.0; V. decumbens, https://www.inaturalist.org/observations/10807224 (accessed on 5 May 2022), © Magriet Brink, CC BY-NC 4.0; V. filicaulis, https://www.inaturalist.org/observations/67950856 (accessed on 5 May 2022), © Andrew Townsend, CC BY-NC; V. formosana, https://www.inaturalist.org/observations/25305342 (accessed on 5 May 2022), © Kuan-Chieh (Chuck) Hung, CC BY-NC 4.0; V. hamiltoniana, https://www.inaturalist.org/observations/74528783 (accessed on 5 May 2022), © Toshihiro Nagata, CC BY-NC 4.0; V. langsdorffii, https://www.inaturalist.org/observations/73470330 (accessed on 5 May 2022), © Jonathan Goff, CC BY-NC-SA 4.0; V. maviensis, https://www.inaturalist.org/observations/39272209 (accessed on 7 May 2022), © Karl Magnacca, CC BY-NC 4.0; V. nannei, https://www.inaturalist.org/observations/6527547 (accessed on 5 May 2022), © Neptalí Ramírez Marcial, CC BY-NC; V. rafinesquei, https://www.inaturalist.org/observations/111043498 (accessed on 5 May 2022), © Mary Ann Yaich, CC BY-NC 4.0; V. rubella, https://www.inaturalist.org/observations/93725094 (accessed on 5 May 2022), © Pablo Silva, CC BY-NC 4.0; and V. tridentata, https://www.inaturalist.org/observations/103467450 (accessed on 5 May 2022), © larsonek, CC BY-NC 4.0.—Links to the relevant CC licences: CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/; accessed on 5 May 2022), CC-BY-NC and CC BY-NC 4.0 (https://creativecommons.org/licenses/by-nc/4.0/; accessed on 5 May 2022), CC BY-NC-SA 4.0 (https://creativecommons.org/licenses/by-nc-sa/4.0/; accessed on 5 May 2022).
Figure 3.
Photos of representative members of the sections and subsections of Viola.—(a–l) Subg. Neoandinium.—(m–av) Subg. Viola.—(a) Sect. Confertae: V. nassauvioides.—(b) Sect. Ericoidium: V. fluehmannii (photo © John M. Watson).—(c) Sect. Grandiflos: V. cheeseana (photo © John M. Watson).—(d) Sect. Inconspicuiflos: V. weibelii (photo © John M. Watson).—(e) Sect. Inconspicuiflos: V. lilliputana (photo © Harvey E. Ballard).—(f) Sect. Relictium: V. dandoisiorum (photo © John M. Watson).—(g) Sect. Rhizomandinium: V. escondidaensis (photo © John M. Watson).—(h) Sect. Rosulatae: V. volcanica (photo © Ana R. Flores).—(i) Sect. Sempervivum: V. atropurpurea (photo © John M. Watson).—(j) Sect. Subandinium: V. subandina (photo © Ana R. Flores).—(k) Sect. Triflabellium: V. triflabellata (photo © Ana R. Flores).—(l) Sect. Xylobasis: V. beati (photo © Ana R. Flores).—(m) Sect. Abyssinium: V. abyssinica (photo © Robert v. Blittersdorff).—(n) Sect. Chamaemelanium: V. pubescens (photo © Kim Blaxland).—(o) Sect. Chilenium: V. maculata (photo © Instituto de Botánica Darwinion).—(p) Sect. Danxiaviola: V. hybanthoides (photo © Qiang Fan).—(q) Sect. Delphiniopsis: V. cazorlensis (photo © Santiago Martín-Bravo).—(r) Sect. Erpetion: V. banksii (photo © Dewald du Plessis).—(s) Sect. Himalayum: V. kunawurensis (photo © Vladimir Epiktetov).—(t) Sect. Leptidium: V. boliviana (photo © Sam Wilson).—(u) Sect. Melanium, subsect. Bracteolatae: V. tricolor and V. arvensis (photo © Thomas Marcussen).—(v) Sect. Melanium, subsect. Cleistogamae: V. rafinesquei (photo © Mary Ann Yaich).—(w) Sect. Melanium, subsect. Dispares: V. dyris (photo © Alan Keohane).—(x) Sect. Melanium, subsect. Ebracteatae: V. parvula (photo © Albert Keshet).—(y) Sect. Melanium, subsect. Pseudorupestres: V. nummulariifolia (photo © Sylvain Piry).—(z) Sect. Melvio: V. decumbens (photo © Magriet Brink).—(aa) Sect. Nematocaulon: V. filicaulis (photo © Andrew Townsend).—(ab)Sect. Nosphinium, subsect. Borealiamericanae: V. fimbriatula (photo © Kim Blaxland).—(ac) Sect. Nosphinium, subsect. Clausenianae: V. clauseniana (photo © Thomas Marcussen).—(ad) Sect. Nosphinium, subsect. Langsdorffianae: V. langsdorffii (photo © Jonathan Goff).—(ae) Sect. Nosphinium, subsect. Mexicanae: V. nannei (photo © Neptalí Ramírez Marcial).—(af) Sect. Nosphinium, subsect. Nosphinium: V. maviensis (photo © Karl Magnacca).—(ag) Sect. Nosphinium, subsect. Pedatae: V. pedata (photo © Kim Blaxland).—(ah) Sect. Plagiostigma, subsect. Australasiaticae: V. sumatrana (photo © Mário Duchoň).—(ai) Sect. Plagiostigma, subsect. Bilobatae: V. hamiltoniana (photo © Toshihiro Nagata).—(aj) Sect. Plagiostigma, subsect. Bulbosae: V. tuberifera (photo © Masashi Igari).—(ak) Sect. Plagiostigma, subsect. Diffusae: V. huizhouensis (photo © Yanshuang Huang).—(al) Sect. Plagiostigma, subsect. Formosanae: V. formosana (photo © Kuan-Chieh (Chuck) Hung).—(am) Sect. Plagiostigma, subsect. Patellares: V. tokubuchiana (photo © Masashi Igari).—(an) Sect. Plagiostigma, subsect. Stolonosae: V. pluviae (photo © Kim Blaxland).—(ao) Sect. Rubellium: V. rubella (photo © Pablo Silva).—(ap) Sect. Sclerosium: V. cinerea (photo © Jerome Viard).—(aq) Sect. Spathulidium: V. pachyrrhiza (photo © Dieter Zschummel).—(ar) Sect. Tridens: V. tridentata (photo © larsonek).—(as) Sect. Viola, subsect. Rostratae: V. canina (photo © Thomas Marcussen).—(at) Sect. Viola, subsect. Viola: V. collina (photo © Thomas Marcussen).—(au) Sect. Xanthidium: V. flavicans (photo © Instituto de Botánica Darwinion).—(av) Sect. Xylinosium: V. arborescens (photo © Abdelmonaim Homrani Bakali).—All images used in this figure were cropped and gamma-corrected.—Links to the online sources for the images used in this figure under a creative commons (CC) licence: V. abyssinica, http://www.westafricanplants.senckenberg.de/root/index.php?page_id=47&id=2203#image=22051, (accessed on 5 May 2022) © Robert v. Blittersdorff; V. arborescens, https://www.teline.fr/en/photos/violaceae/viola-arborescens#photo-7 (accessed on 11 May 2022), © Abdelmonaim Homrani Bakali, CC BY-NC 4.0; V. nummulariifolia, https://www.inaturalist.org/observations/89240299 (accessed on 5 May 2022), © Sylvain Piry, CC BY-NC 4.0; V. banksii, https://inaturalist.nz/observations/25048808 (accessed on 12 May 2022), © Dewald du Plessis, CC BY-NC 4.0; V. boliviana, https://ecuador.inaturalist.org/photos/57308380 (accessed on 5 May 2022), © Sam Wilson, CC BY-NC; V. cazorlensis, https://www.inaturalist.org/observations/109331467 (accessed on 5 May 2022), © Santiago Martín-Bravo, CC BY-NC 4.0; V. cinerea, https://www.inaturalist.org/observations/65108391 (accessed on 5 May 2022), © Jerome Viard, CC BY-NC 4.0; V. decumbens, https://www.inaturalist.org/observations/10807224 (accessed on 5 May 2022), © Magriet Brink, CC BY-NC 4.0; V. filicaulis, https://www.inaturalist.org/observations/67950856 (accessed on 5 May 2022), © Andrew Townsend, CC BY-NC; V. formosana, https://www.inaturalist.org/observations/25305342 (accessed on 5 May 2022), © Kuan-Chieh (Chuck) Hung, CC BY-NC 4.0; V. hamiltoniana, https://www.inaturalist.org/observations/74528783 (accessed on 5 May 2022), © Toshihiro Nagata, CC BY-NC 4.0; V. langsdorffii, https://www.inaturalist.org/observations/73470330 (accessed on 5 May 2022), © Jonathan Goff, CC BY-NC-SA 4.0; V. maviensis, https://www.inaturalist.org/observations/39272209 (accessed on 7 May 2022), © Karl Magnacca, CC BY-NC 4.0; V. nannei, https://www.inaturalist.org/observations/6527547 (accessed on 5 May 2022), © Neptalí Ramírez Marcial, CC BY-NC; V. rafinesquei, https://www.inaturalist.org/observations/111043498 (accessed on 5 May 2022), © Mary Ann Yaich, CC BY-NC 4.0; V. rubella, https://www.inaturalist.org/observations/93725094 (accessed on 5 May 2022), © Pablo Silva, CC BY-NC 4.0; and V. tridentata, https://www.inaturalist.org/observations/103467450 (accessed on 5 May 2022), © larsonek, CC BY-NC 4.0.—Links to the relevant CC licences: CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/; accessed on 5 May 2022), CC-BY-NC and CC BY-NC 4.0 (https://creativecommons.org/licenses/by-nc/4.0/; accessed on 5 May 2022), CC BY-NC-SA 4.0 (https://creativecommons.org/licenses/by-nc-sa/4.0/; accessed on 5 May 2022).




Figure 4.
Stacked bar plot showing species richness among the 31 sections of Viola. Species counts are indicated above each bar. Sections containing subsections are indicated as stacked bars with the distribution of species among subsections indicated in different colours. For details on each subsection, see Table 1.
Figure 4.
Stacked bar plot showing species richness among the 31 sections of Viola. Species counts are indicated above each bar. Sections containing subsections are indicated as stacked bars with the distribution of species among subsections indicated in different colours. For details on each subsection, see Table 1.

Figure 5.
Dated phylogeny for monophyletic sections of Viola, updated from Marcussen et al. [28]. Estimated base ploidy is indicated after the name of each Viola section. Curved lines indicate parental lineages of an allopolyploid lineage and filled diamonds indicate the estimated time of allopolyploidisation [28]. Daggers denote parental lineages that are extinct or unsampled and extant only as a subgenome of an allopolyploid lineage. Horizontal black triangles indicate the estimated crown node of a section (and is prone to increase as more taxa are added). No phylogenetic data exist for sect. Nematocaulon and sect. Xanthidium of subg. Viola and for another seven sections of subg. Neoandinium.
Figure 5.
Dated phylogeny for monophyletic sections of Viola, updated from Marcussen et al. [28]. Estimated base ploidy is indicated after the name of each Viola section. Curved lines indicate parental lineages of an allopolyploid lineage and filled diamonds indicate the estimated time of allopolyploidisation [28]. Daggers denote parental lineages that are extinct or unsampled and extant only as a subgenome of an allopolyploid lineage. Horizontal black triangles indicate the estimated crown node of a section (and is prone to increase as more taxa are added). No phylogenetic data exist for sect. Nematocaulon and sect. Xanthidium of subg. Viola and for another seven sections of subg. Neoandinium.

Figure 6.
Dated BEAST phylogeny of the Internal Transcribed Spacers (ITS) 1 and 2 in selected taxa of Viola and secondary age calibration of five internal nodes. Tip names are shown as section, subsection (if available), and species. Node bars indicate the posterior 95% credibility interval for node height; nodes with bars indicated in brown were used for age calibration. Mean ages are indicated on nodes. The outgroup (Melicytus) was pruned.
Figure 6.
Dated BEAST phylogeny of the Internal Transcribed Spacers (ITS) 1 and 2 in selected taxa of Viola and secondary age calibration of five internal nodes. Tip names are shown as section, subsection (if available), and species. Node bars indicate the posterior 95% credibility interval for node height; nodes with bars indicated in brown were used for age calibration. Mean ages are indicated on nodes. The outgroup (Melicytus) was pruned.

Figure 7.
Wire diagram illustrating the major taxonomic differences between the phylogenetic classification of Viola proposed here (“Marcussen et al. 2022”) compared to the morphological classification proposed by Becker (1925 [1]). Merging lines denote lumping of two or more of Becker’s infrageneric groups into one taxon, while splitting lines denote segregation into two or more taxa. Taxa placed on the same line and interlinked with a horizontal line are synonymous, but may differ in delimitation, rank, or name (for reasons of priority). Taxa indicated with a dagger have been reduced to synonymy under another taxon. Taxa indicated with an asterisk are new infrageneric segregates described here. We do not show infrasectional taxa for sect. Chamaemelanium, which are not accepted here, or for sect. Melanium and the sect. Andinium/subg. Neoandinium pair, for which our treatments differ substantially from that by Becker.
Figure 7.
Wire diagram illustrating the major taxonomic differences between the phylogenetic classification of Viola proposed here (“Marcussen et al. 2022”) compared to the morphological classification proposed by Becker (1925 [1]). Merging lines denote lumping of two or more of Becker’s infrageneric groups into one taxon, while splitting lines denote segregation into two or more taxa. Taxa placed on the same line and interlinked with a horizontal line are synonymous, but may differ in delimitation, rank, or name (for reasons of priority). Taxa indicated with a dagger have been reduced to synonymy under another taxon. Taxa indicated with an asterisk are new infrageneric segregates described here. We do not show infrasectional taxa for sect. Chamaemelanium, which are not accepted here, or for sect. Melanium and the sect. Andinium/subg. Neoandinium pair, for which our treatments differ substantially from that by Becker.

Figure 8.
Discrete historical biogeography of Viola sections, showing the South American origin of the genus and independent dispersal into the northern hemisphere by the CHAM and MELVIO lineages. Ancestral states were inferred by stochastic character mapping [103] using a 1-rate model and 1000 replicates, given the monoploid multilabelled timetree with 50 leaves that results from unfolding the network in Figure 5 to a tree. Sections Nematocaulon (New Zealand) and Xanthidium (South America), for which data are lacking, are not included.
Figure 8.
Discrete historical biogeography of Viola sections, showing the South American origin of the genus and independent dispersal into the northern hemisphere by the CHAM and MELVIO lineages. Ancestral states were inferred by stochastic character mapping [103] using a 1-rate model and 1000 replicates, given the monoploid multilabelled timetree with 50 leaves that results from unfolding the network in Figure 5 to a tree. Sections Nematocaulon (New Zealand) and Xanthidium (South America), for which data are lacking, are not included.

Figure 9.
Style shapes in Viola. (a–ae) Subg. Neoandinium. (af–fe) Subg. Viola.—Sect. Confertae: (a) V. nassauvioides [1].—Sect. Ericoidium: (b) V. fluehmannii [80].—Sect. Grandiflos: (c) V. acanthophylla [122].—Sect. Inconspicuiflos: (d) V. lilliputana, (e) V. membranacea [1].—Sect. Relictium: (f) V. ovalleana [48].—Sect. Rhizomandinium: (g) V. escondidaensis [80].—Sect. Rosulatae: (h) V. aurantiaca [1], (i) V. kermesina, (j) V. niederleinii [1], (k) V. replicata [1], (l) V. rugosa [1], (m) V. volcanica [1].—Sect. Sempervivum: (n) V. atropurpurea [80], (o) V. auricolor [80], (p) V. bangii, (q) V. coronifera [80], (r) V. cotyledon [1], (s) V. dasyphylla [80], (t) V. hieronymi [80], (u) V. micranthella, (v) V. pygmaea, (w) V. sacculus [80], (x) V. sempervivum.—Sect. Subandinium: (y) V. araucaniae [1], (z) V. polypoda, (aa) V. pusilla, (ab) V. subandina [80], (ac) V. weberbaueri.—Sect. Triflabellium: (ad) V. triflabellata [80].—Sect. Xylobasis: (ae) V. beati, tip of style [123].—Sect. Abyssinium: (af) V. abyssinica.—Sect. Chamaemelanium: (ag) V. beckwithii, (ah) V. biflora, (ai) V. canadensis, (aj) V. charlestonensis, (ak) V. cuneata, (al) V. douglasii, (am) V. flettii, (an) V. frank-smithii, (ao) V. guadalupensis, (ap) V. hallii, (aq) V. lithion, (ar) V. lobata, (as) V. nuttallii, (at) V. ocellata, (au) V. orientalis (redrawn from [78]), (av) V. scopulorum, (aw) V. sheltonii, (ax) V. trinervata, (ay) V. wallichiana (redrawn from [78]).—Sect. Chilenium: (az) V. commersonii [80], (ba) V. maculata, (bb) V. reichei, (bc) V. stuebelii.—Sect. Danxiaviola: (bd) V. hybanthoides (redrawn from [90]).—Sect. Delphiniopsis: (be) V. cazorlensis [1], (bf) V. delphinantha.—Sect. Erpetion: (bg) V. banksii.—Sect. Himalayum: (bh) V. kunawurensis (redrawn from [124]).—Sect. Leptidium: (bi) V. stipularis [1].—Sect. Melanium, subsect. Bracteolatae: (bj) V. cornuta [29], (bk) V. tricolor (redrawn from [125]).—Sect. Melanium, subsect. Cleistogamae: (bl) V. rafinesquei.—Sect. Melanium, subsect. Ebracteolatae: (bm) V. dirimliensis.—Sect. Melvio: (bn) V. decumbens.—Sect. Nematocaulon: (bo) V. filicaulis.—Sect. Nosphinium, subsect. Borealiamericanae: (bp) V. brittoniana, (bq) V. cucullata, (br) V. palmata, (bs) V. pedatifida, (bt) V. pratincola, (bu) V. sagittata, (bv) V. septemloba, (bw) V. sororia, (bx) V. viarum, (by) V. villosa.—Sect. Nosphinium, subsect. Clausenianae: (bz) V. clauseniana.—Sect. Nosphinium, subsect. Langsdorffianae: (ca) V. howellii, (cb) V. langsdorffii.—Sect. Nosphinium, subsect. Mexicanae: (cc) V. grahamii, (cd) V. guatemalensis, (ce) V. hookeriana, (cf) V. humilis, (cg) V. nannei, (ch) V. nubicola, (ci) V. oxyodontis.—Sect. Nosphinium, subsect. Nosphinium: (cj) V. kauaensis, (ck) V. maviensis [1].—Sect. Nosphinium, subsect. Pedatae: (cl) V. pedata.—Sect. Plagiostigma, subsect. Australasiaticae: (cm) V. austrosinensis (redrawn from [126]), (cn) V. annamensis, (co) V. kwangtungensis (redrawn from [78]), (cp) V. sumatrana (redrawn from [74]).—Sect. Plagiostigma, subsect. Bilobatae: (cq) V. arcuata (redrawn from [75]), (cs) V. merrilliana (redrawn from [74]), (ct) V. triangulifolia (redrawn from [78]), (cr) V. hamiltoniana [29].—Sect. Plagiostigma, subsect. Bulbosae: (cu) V. bulbosa (redrawn from [76]), (cv) V. tuberifera (redrawn from [76]).—Sect. Plagiostigma, subsect. Diffusae: (cw) V. diffusa [29], (cx) V. huizhouensis (redrawn from [127]), (cy) V. jinggangshanensis (redrawn from [128]), (cz) V. nagasawae (redrawn from [75]).—Sect. Plagiostigma, subsect. Formosanae: (da) V. formosana (redrawn from [75]), (db) V. stolonifolora (redrawn from [97]).—Sect. Plagiostigma, subsect. Patellares: (dc) V. dactyloides [1], (dd) V. japonica [29], (de) V. macroceras [1], (df) V. patrinii [1], (dg) V. pinnata [29], (dh) V. selkirkii.—Sect. Plagiostigma, subsect. Stolonosae: (di) V. suecica, (dj) V. blanda, (dk) V. brevipes, (dl) V. davidii (redrawn from [78]), (dm) V. fargesii (redrawn from [78]), (dn) V. grandisepala (redrawn from [78]), (do) V. jalapaensis, (dp) V. lanceolata, (dq) V. macloskeyi, (dr) V. minuscula, (ds) V. occidentalis, (dt) V. palustris, (du) V. pluviae, (dv) V. primulifolia, (dw) V. renifolia, (dx) V. rossii (redrawn from [76]).—Sect. Rubellium: (dy) V. capillaris [1], (dz) V. portalesia [1], (ea) V. rubella.—Sect. Sclerosium: (eb) V. stocksii, (ec) V. etbaica [1], (ed) V. somalensis [1].—Sect. Spathulidium: (ee) V. spathulata.—Sect. Tridens: (ef) V. tridentata [1].—Sect. Viola, subsect. Rostratae: (eg) V. acuminata (redrawn from [76]), (eh) V. adunca, (ei) V. appalachiensis, (ej) V. canina, (ek) V. elatior [29], (el) V. huidobrii [80], (em) V. jordanii, (en) V. labradorica, (eo) V. papuana (redrawn from [74]), (ep) V. rostrata, (eq) V. rupestris [29], (er) V. stagnina [29], (es) V. striata, (et) V. uliginosa [29], (eu) V. umbraticola, (ev) V. walteri.—Sect. Viola, subsect. Viola: (ew) V. barhalensis (redrawn from [129]), (ex) V. chelmea [1], (ey) V. hirta [29], (ez) V. odorata, (fa) V. pilosa [1].—Sect. Xanthidium: (fb) V. flavicans, (fc) V. pallascaensis.—Sect. Xylinosium: (fd) V. arborescens [1], (fe) V. scorpiuroides.—All drawings by Kim Blaxland, H.E.B, and T.M. except where indicated.
Figure 9.
Style shapes in Viola. (a–ae) Subg. Neoandinium. (af–fe) Subg. Viola.—Sect. Confertae: (a) V. nassauvioides [1].—Sect. Ericoidium: (b) V. fluehmannii [80].—Sect. Grandiflos: (c) V. acanthophylla [122].—Sect. Inconspicuiflos: (d) V. lilliputana, (e) V. membranacea [1].—Sect. Relictium: (f) V. ovalleana [48].—Sect. Rhizomandinium: (g) V. escondidaensis [80].—Sect. Rosulatae: (h) V. aurantiaca [1], (i) V. kermesina, (j) V. niederleinii [1], (k) V. replicata [1], (l) V. rugosa [1], (m) V. volcanica [1].—Sect. Sempervivum: (n) V. atropurpurea [80], (o) V. auricolor [80], (p) V. bangii, (q) V. coronifera [80], (r) V. cotyledon [1], (s) V. dasyphylla [80], (t) V. hieronymi [80], (u) V. micranthella, (v) V. pygmaea, (w) V. sacculus [80], (x) V. sempervivum.—Sect. Subandinium: (y) V. araucaniae [1], (z) V. polypoda, (aa) V. pusilla, (ab) V. subandina [80], (ac) V. weberbaueri.—Sect. Triflabellium: (ad) V. triflabellata [80].—Sect. Xylobasis: (ae) V. beati, tip of style [123].—Sect. Abyssinium: (af) V. abyssinica.—Sect. Chamaemelanium: (ag) V. beckwithii, (ah) V. biflora, (ai) V. canadensis, (aj) V. charlestonensis, (ak) V. cuneata, (al) V. douglasii, (am) V. flettii, (an) V. frank-smithii, (ao) V. guadalupensis, (ap) V. hallii, (aq) V. lithion, (ar) V. lobata, (as) V. nuttallii, (at) V. ocellata, (au) V. orientalis (redrawn from [78]), (av) V. scopulorum, (aw) V. sheltonii, (ax) V. trinervata, (ay) V. wallichiana (redrawn from [78]).—Sect. Chilenium: (az) V. commersonii [80], (ba) V. maculata, (bb) V. reichei, (bc) V. stuebelii.—Sect. Danxiaviola: (bd) V. hybanthoides (redrawn from [90]).—Sect. Delphiniopsis: (be) V. cazorlensis [1], (bf) V. delphinantha.—Sect. Erpetion: (bg) V. banksii.—Sect. Himalayum: (bh) V. kunawurensis (redrawn from [124]).—Sect. Leptidium: (bi) V. stipularis [1].—Sect. Melanium, subsect. Bracteolatae: (bj) V. cornuta [29], (bk) V. tricolor (redrawn from [125]).—Sect. Melanium, subsect. Cleistogamae: (bl) V. rafinesquei.—Sect. Melanium, subsect. Ebracteolatae: (bm) V. dirimliensis.—Sect. Melvio: (bn) V. decumbens.—Sect. Nematocaulon: (bo) V. filicaulis.—Sect. Nosphinium, subsect. Borealiamericanae: (bp) V. brittoniana, (bq) V. cucullata, (br) V. palmata, (bs) V. pedatifida, (bt) V. pratincola, (bu) V. sagittata, (bv) V. septemloba, (bw) V. sororia, (bx) V. viarum, (by) V. villosa.—Sect. Nosphinium, subsect. Clausenianae: (bz) V. clauseniana.—Sect. Nosphinium, subsect. Langsdorffianae: (ca) V. howellii, (cb) V. langsdorffii.—Sect. Nosphinium, subsect. Mexicanae: (cc) V. grahamii, (cd) V. guatemalensis, (ce) V. hookeriana, (cf) V. humilis, (cg) V. nannei, (ch) V. nubicola, (ci) V. oxyodontis.—Sect. Nosphinium, subsect. Nosphinium: (cj) V. kauaensis, (ck) V. maviensis [1].—Sect. Nosphinium, subsect. Pedatae: (cl) V. pedata.—Sect. Plagiostigma, subsect. Australasiaticae: (cm) V. austrosinensis (redrawn from [126]), (cn) V. annamensis, (co) V. kwangtungensis (redrawn from [78]), (cp) V. sumatrana (redrawn from [74]).—Sect. Plagiostigma, subsect. Bilobatae: (cq) V. arcuata (redrawn from [75]), (cs) V. merrilliana (redrawn from [74]), (ct) V. triangulifolia (redrawn from [78]), (cr) V. hamiltoniana [29].—Sect. Plagiostigma, subsect. Bulbosae: (cu) V. bulbosa (redrawn from [76]), (cv) V. tuberifera (redrawn from [76]).—Sect. Plagiostigma, subsect. Diffusae: (cw) V. diffusa [29], (cx) V. huizhouensis (redrawn from [127]), (cy) V. jinggangshanensis (redrawn from [128]), (cz) V. nagasawae (redrawn from [75]).—Sect. Plagiostigma, subsect. Formosanae: (da) V. formosana (redrawn from [75]), (db) V. stolonifolora (redrawn from [97]).—Sect. Plagiostigma, subsect. Patellares: (dc) V. dactyloides [1], (dd) V. japonica [29], (de) V. macroceras [1], (df) V. patrinii [1], (dg) V. pinnata [29], (dh) V. selkirkii.—Sect. Plagiostigma, subsect. Stolonosae: (di) V. suecica, (dj) V. blanda, (dk) V. brevipes, (dl) V. davidii (redrawn from [78]), (dm) V. fargesii (redrawn from [78]), (dn) V. grandisepala (redrawn from [78]), (do) V. jalapaensis, (dp) V. lanceolata, (dq) V. macloskeyi, (dr) V. minuscula, (ds) V. occidentalis, (dt) V. palustris, (du) V. pluviae, (dv) V. primulifolia, (dw) V. renifolia, (dx) V. rossii (redrawn from [76]).—Sect. Rubellium: (dy) V. capillaris [1], (dz) V. portalesia [1], (ea) V. rubella.—Sect. Sclerosium: (eb) V. stocksii, (ec) V. etbaica [1], (ed) V. somalensis [1].—Sect. Spathulidium: (ee) V. spathulata.—Sect. Tridens: (ef) V. tridentata [1].—Sect. Viola, subsect. Rostratae: (eg) V. acuminata (redrawn from [76]), (eh) V. adunca, (ei) V. appalachiensis, (ej) V. canina, (ek) V. elatior [29], (el) V. huidobrii [80], (em) V. jordanii, (en) V. labradorica, (eo) V. papuana (redrawn from [74]), (ep) V. rostrata, (eq) V. rupestris [29], (er) V. stagnina [29], (es) V. striata, (et) V. uliginosa [29], (eu) V. umbraticola, (ev) V. walteri.—Sect. Viola, subsect. Viola: (ew) V. barhalensis (redrawn from [129]), (ex) V. chelmea [1], (ey) V. hirta [29], (ez) V. odorata, (fa) V. pilosa [1].—Sect. Xanthidium: (fb) V. flavicans, (fc) V. pallascaensis.—Sect. Xylinosium: (fd) V. arborescens [1], (fe) V. scorpiuroides.—All drawings by Kim Blaxland, H.E.B, and T.M. except where indicated.



Figure 10.
Global distribution of Viola subg. Neoandinium.

Figure 11.
Global distribution of Viola sect. Abyssinium.

Figure 12.
Global distribution of Viola sect. Chamaemelanium.

Figure 13.
Ultrametric phylogeny of diploid taxa of Viola sect. Chamaemelanium, showing the basal irresolution among otherwise well-supported infrasectional lineages and the non-monophyly of Becker’s greges at both the diploid and allopolyploid level. Outgroups have been trimmed. The analysis was performed on a concatenated matrix of four loci (GPI, ITS, NRPD2a, trnL-trnF). The squares indicated on branches show the approximate phylogenetic placement of homoeologs of five North American allopolyploids [60], V. bakeri (8x), V. douglasii (8x), V. glabella (4x), and V. guadalupensis (4x). Branch support is given as posterior probabilities. * Viola biflora has a circumboreal distribution.
Figure 13.
Ultrametric phylogeny of diploid taxa of Viola sect. Chamaemelanium, showing the basal irresolution among otherwise well-supported infrasectional lineages and the non-monophyly of Becker’s greges at both the diploid and allopolyploid level. Outgroups have been trimmed. The analysis was performed on a concatenated matrix of four loci (GPI, ITS, NRPD2a, trnL-trnF). The squares indicated on branches show the approximate phylogenetic placement of homoeologs of five North American allopolyploids [60], V. bakeri (8x), V. douglasii (8x), V. glabella (4x), and V. guadalupensis (4x). Branch support is given as posterior probabilities. * Viola biflora has a circumboreal distribution.

Figure 14.
Global distribution of Viola sect. Chilenium.

Figure 15.
Global distribution of Viola sect. Danxiaviola.

Figure 16.
Global distribution of Viola sect. Delphiniopsis.

Figure 17.
Global distribution of Viola sect. Erpetion.

Figure 18.
Global distribution of Viola sect. Himalayum.

Figure 19.
Global distribution of Viola sect. Leptidium.

Figure 20.
Global distribution of Viola sect. Melanium.

Figure 21.
Global distribution of Viola sect. Melvio.

Figure 22.
Global distribution of Viola sect. Nematocaulon.

Figure 23.
Global distribution of Viola sect. Nosphinium.

Figure 24.
Reticulate allopolyploid phylogeny of Viola sect. Nosphinium, simplified from Marcussen et al. [45]. Allopolyploidisations are indicated by curved lines. Ploidy (x) is indicated.
Figure 24.
Reticulate allopolyploid phylogeny of Viola sect. Nosphinium, simplified from Marcussen et al. [45]. Allopolyploidisations are indicated by curved lines. Ploidy (x) is indicated.

Figure 25.
Historical biogeography of the Hawaiian violets, Viola subsect. Nosphinium, based on ITS sequences and simultaneous estimation of phylogeny and discrete biogeography using BEAST. Kauai is indicated as the most likely island of colonisation, based both on age, which excludes all the other islands, and on receiving the highest posterior probability (pp) by ancestral state reconstruction. The 95% credibility interval for node age is shown as a node bar on the crown node. Ancestral states are colour-coded according to island and indicated on each node along with the pp. Each island is shown as a silhouette and its age is indicated by a vertical line. Outgroups have been trimmed.
Figure 25.
Historical biogeography of the Hawaiian violets, Viola subsect. Nosphinium, based on ITS sequences and simultaneous estimation of phylogeny and discrete biogeography using BEAST. Kauai is indicated as the most likely island of colonisation, based both on age, which excludes all the other islands, and on receiving the highest posterior probability (pp) by ancestral state reconstruction. The 95% credibility interval for node age is shown as a node bar on the crown node. Ancestral states are colour-coded according to island and indicated on each node along with the pp. Each island is shown as a silhouette and its age is indicated by a vertical line. Outgroups have been trimmed.

Figure 26.
Global distribution of Viola sect. Plagiostigma.

Figure 27.
Phylogeny of Viola sect. Plagiostigma showing the delimitation of subsections (4x) with known allopolyploids (8x) superimposed, based on concatenated sequences of eight nuclear gene loci (GPI-C, GPI-M, ITS-C, ITS-M, NRPD2a-C, NRPD2a-M, SDH-C, and SDH-M). Outgroups have been pruned. The relative ages for polyploids are not to scale. Branch support is given as posterior probabilities. Abbreviations: bla, inc = V. blanda and V. incognita; pal = V. palustris; plu = V. pluviae.
Figure 27.
Phylogeny of Viola sect. Plagiostigma showing the delimitation of subsections (4x) with known allopolyploids (8x) superimposed, based on concatenated sequences of eight nuclear gene loci (GPI-C, GPI-M, ITS-C, ITS-M, NRPD2a-C, NRPD2a-M, SDH-C, and SDH-M). Outgroups have been pruned. The relative ages for polyploids are not to scale. Branch support is given as posterior probabilities. Abbreviations: bla, inc = V. blanda and V. incognita; pal = V. palustris; plu = V. pluviae.

Figure 28.
Global distribution of Viola sect. Rubellium.

Figure 29.
Global distribution of Viola sect. Sclerosium.

Figure 30.
Global distribution of Viola sect. Spathulidium.

Figure 31.
Global distribution of Viola sect. Tridens.

Figure 32.
Global distribution of Viola sect. Viola.

Figure 33.
Phylogeny of Viola sect. Viola showing the delimitation of subsections (4x) with known allopolyploids (8x) superimposed, based on concatenated sequences of eight nuclear gene loci (GPI-C, GPI-M, ITS-C, ITS-M, NRPD2a-C, NRPD2a-M, SDH-C, and SDH-M). Outgroups have been pruned. The ages and placements for polyploids are not to scale. Branch support is given as posterior probabilities. Abbreviations: amb = V. ambigua; can, lac, pum = V. canina, V. lactea, and V. pumila; pse, wil = V. pseudomirabilis and V. willkommii.
Figure 33.
Phylogeny of Viola sect. Viola showing the delimitation of subsections (4x) with known allopolyploids (8x) superimposed, based on concatenated sequences of eight nuclear gene loci (GPI-C, GPI-M, ITS-C, ITS-M, NRPD2a-C, NRPD2a-M, SDH-C, and SDH-M). Outgroups have been pruned. The ages and placements for polyploids are not to scale. Branch support is given as posterior probabilities. Abbreviations: amb = V. ambigua; can, lac, pum = V. canina, V. lactea, and V. pumila; pse, wil = V. pseudomirabilis and V. willkommii.

Figure 34.
Global distribution of Viola sect. Xanthidium.

Figure 35.
Global distribution of Viola sect. Xylinosium.


{kind=link}
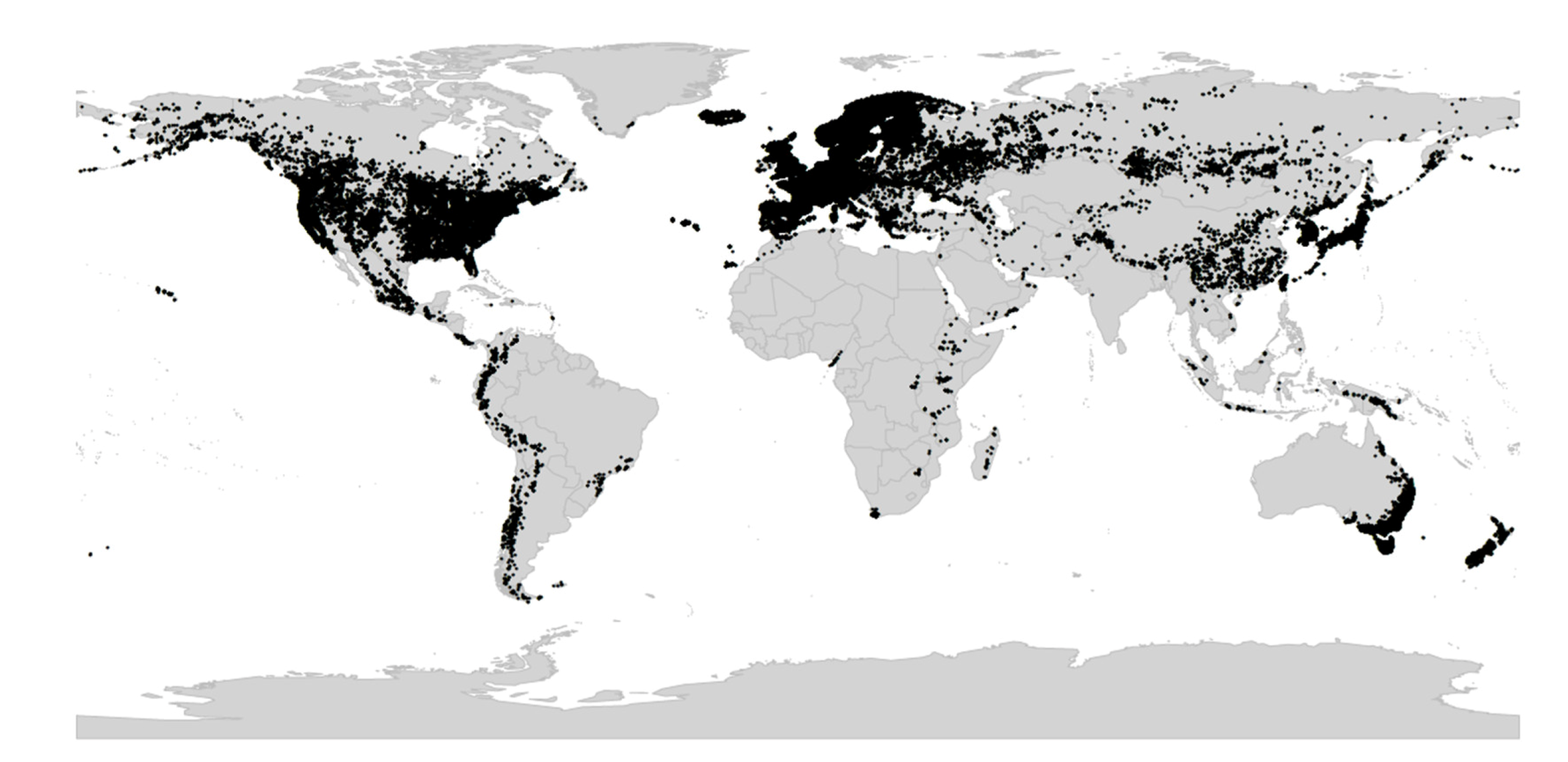{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
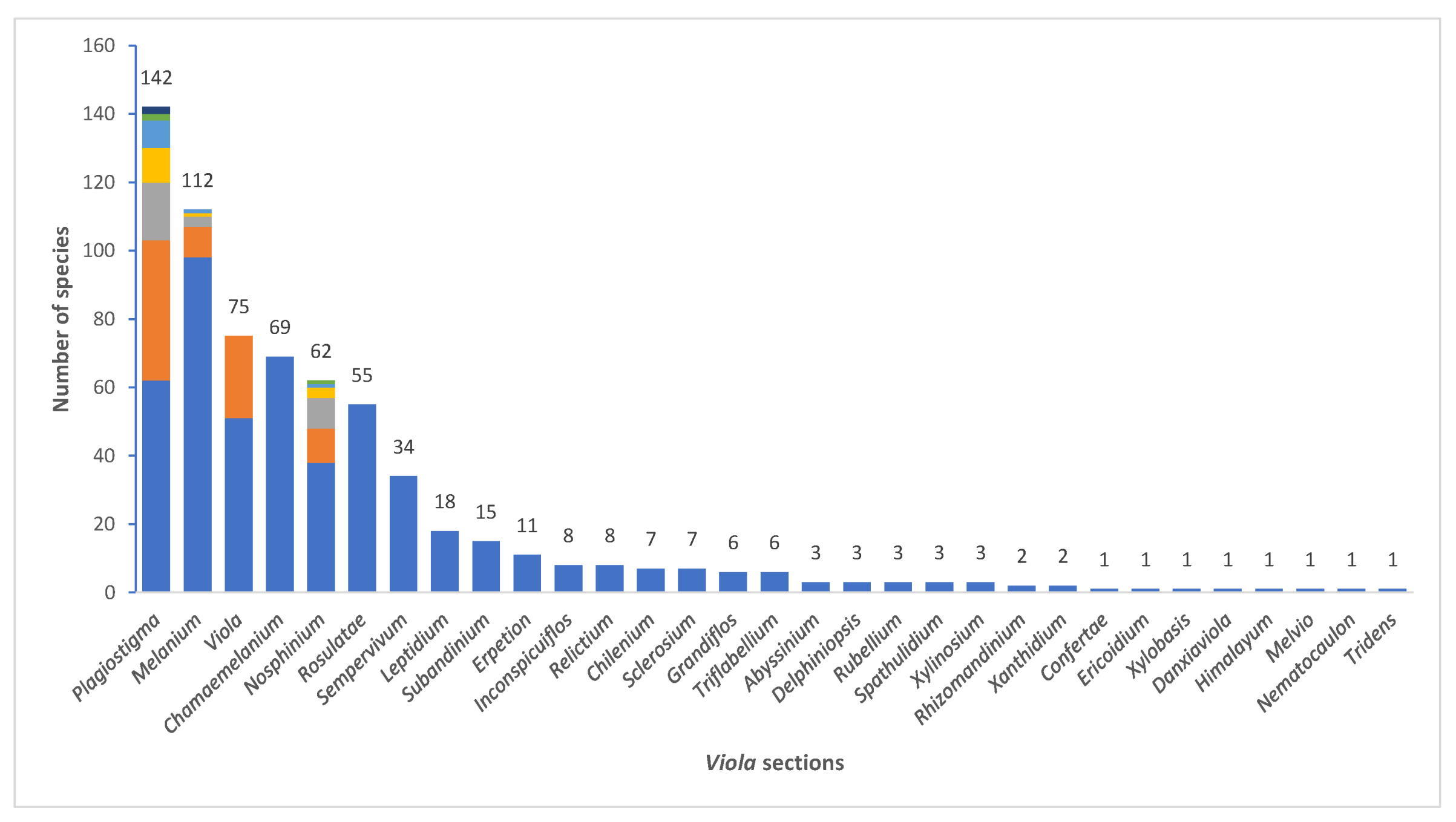{kind=link}
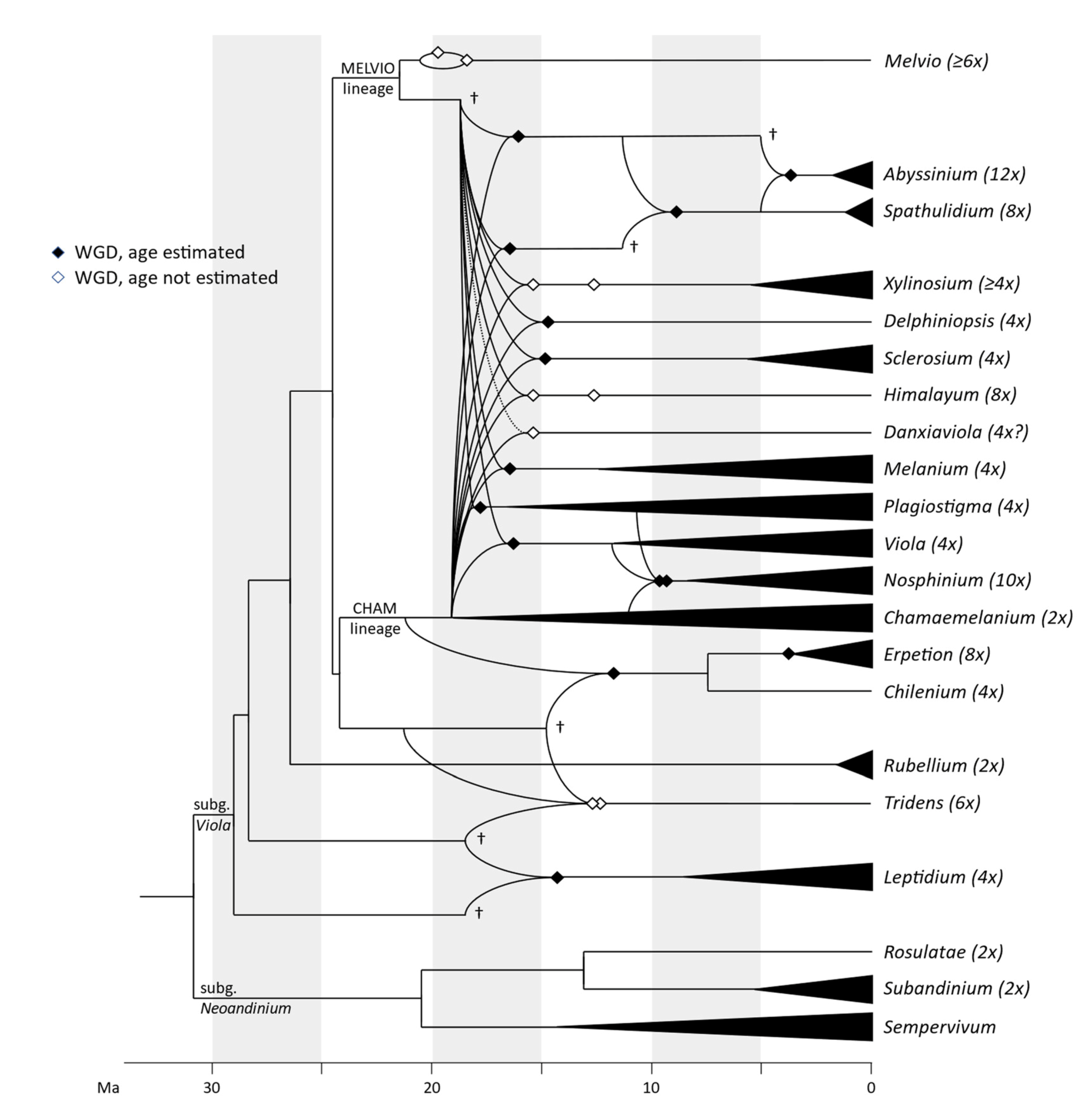{kind=link}
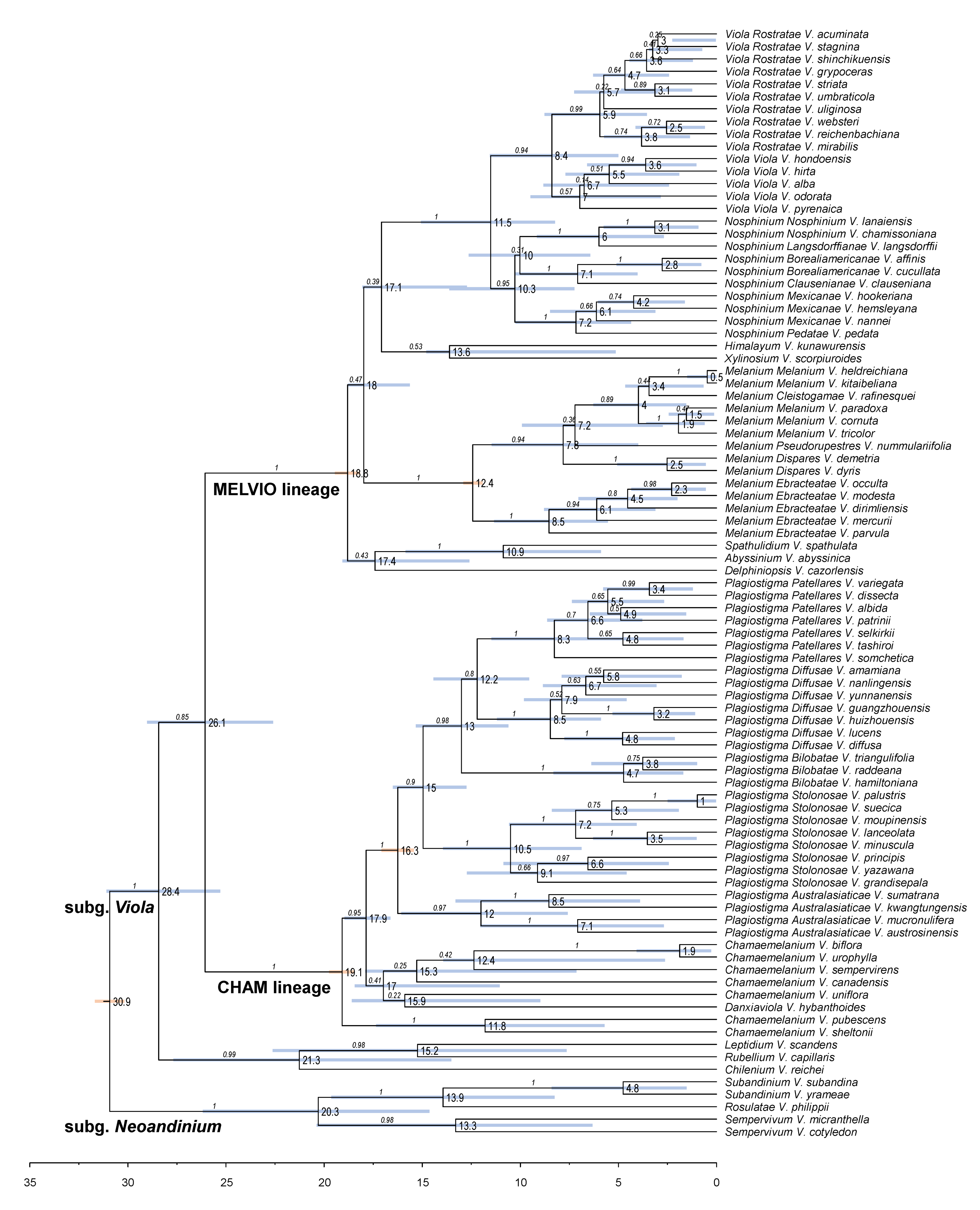{kind=link}
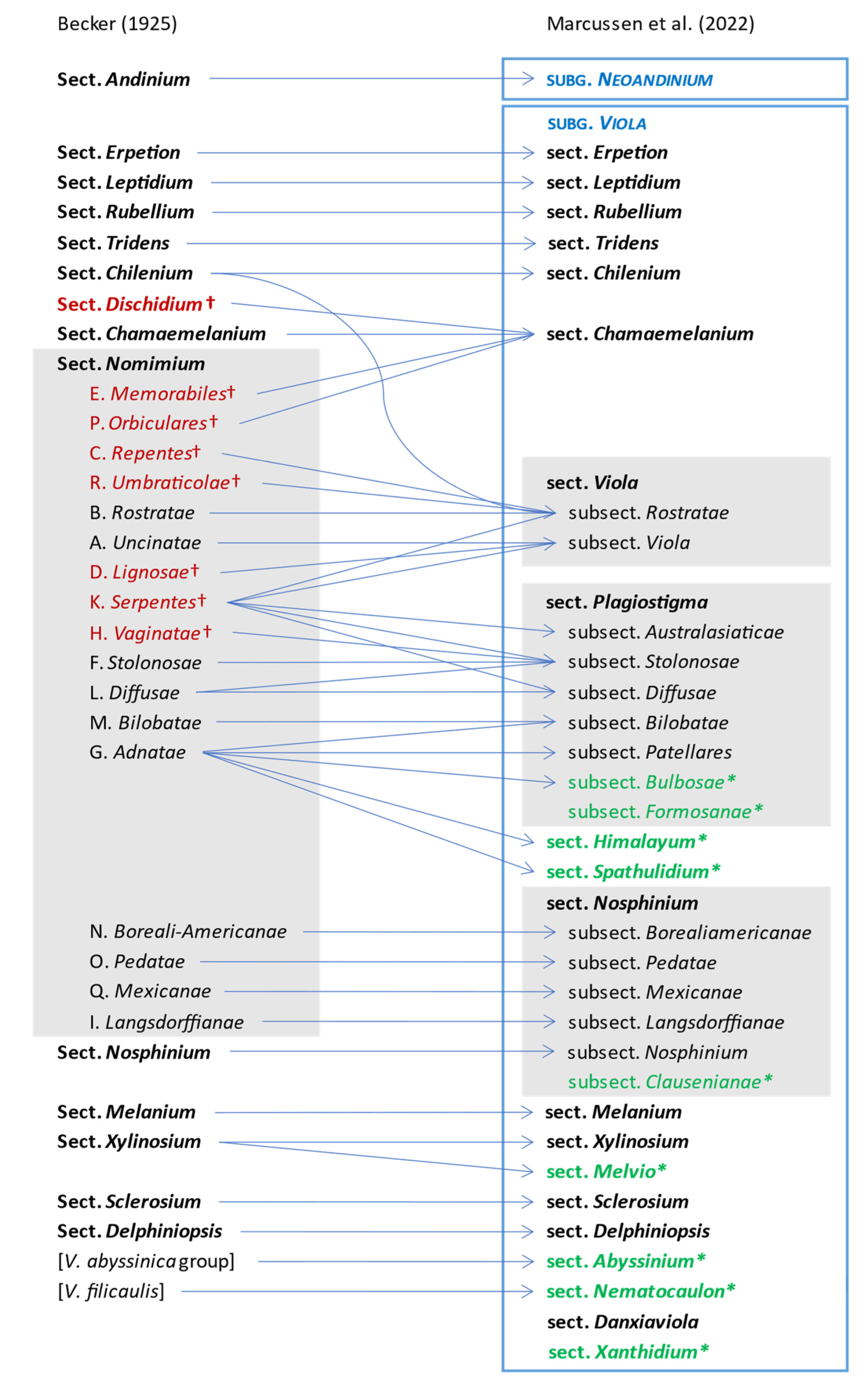{kind=link}
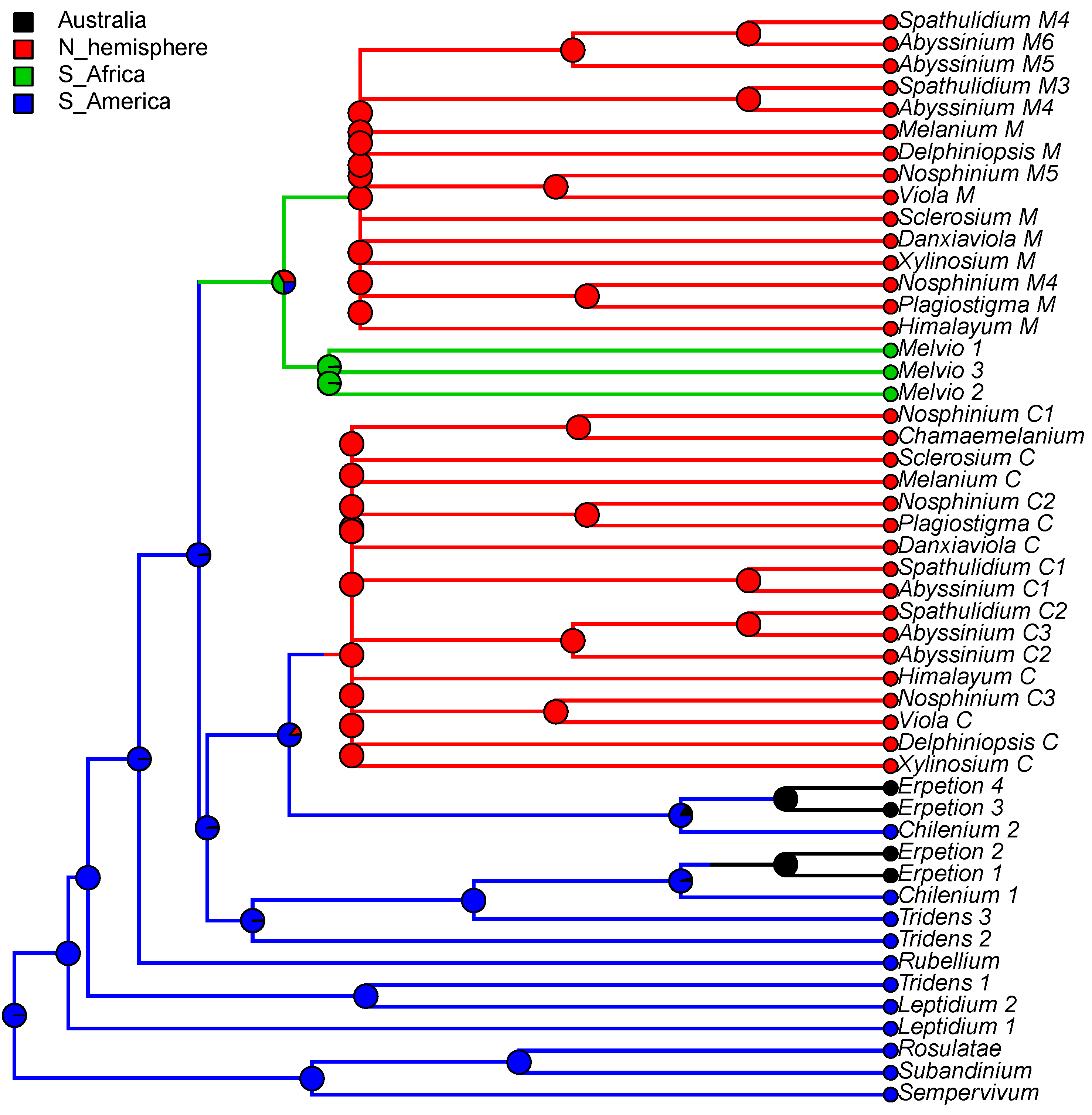{kind=link}
{kind=link}
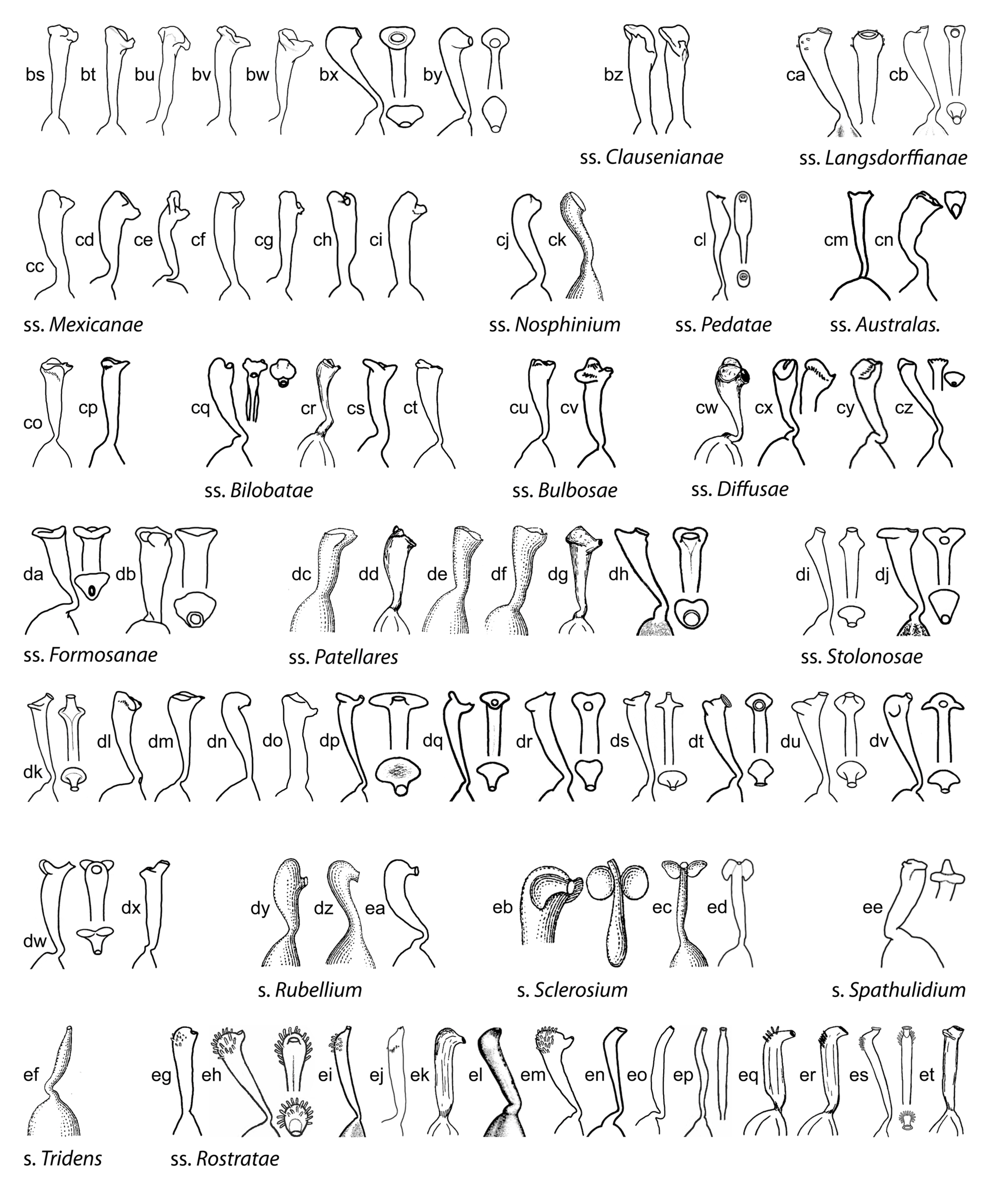{kind=link}
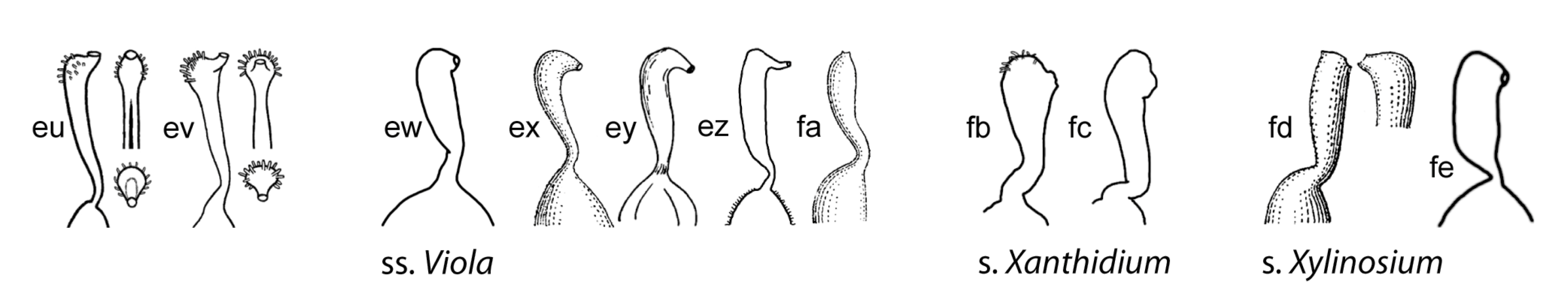{kind=link}
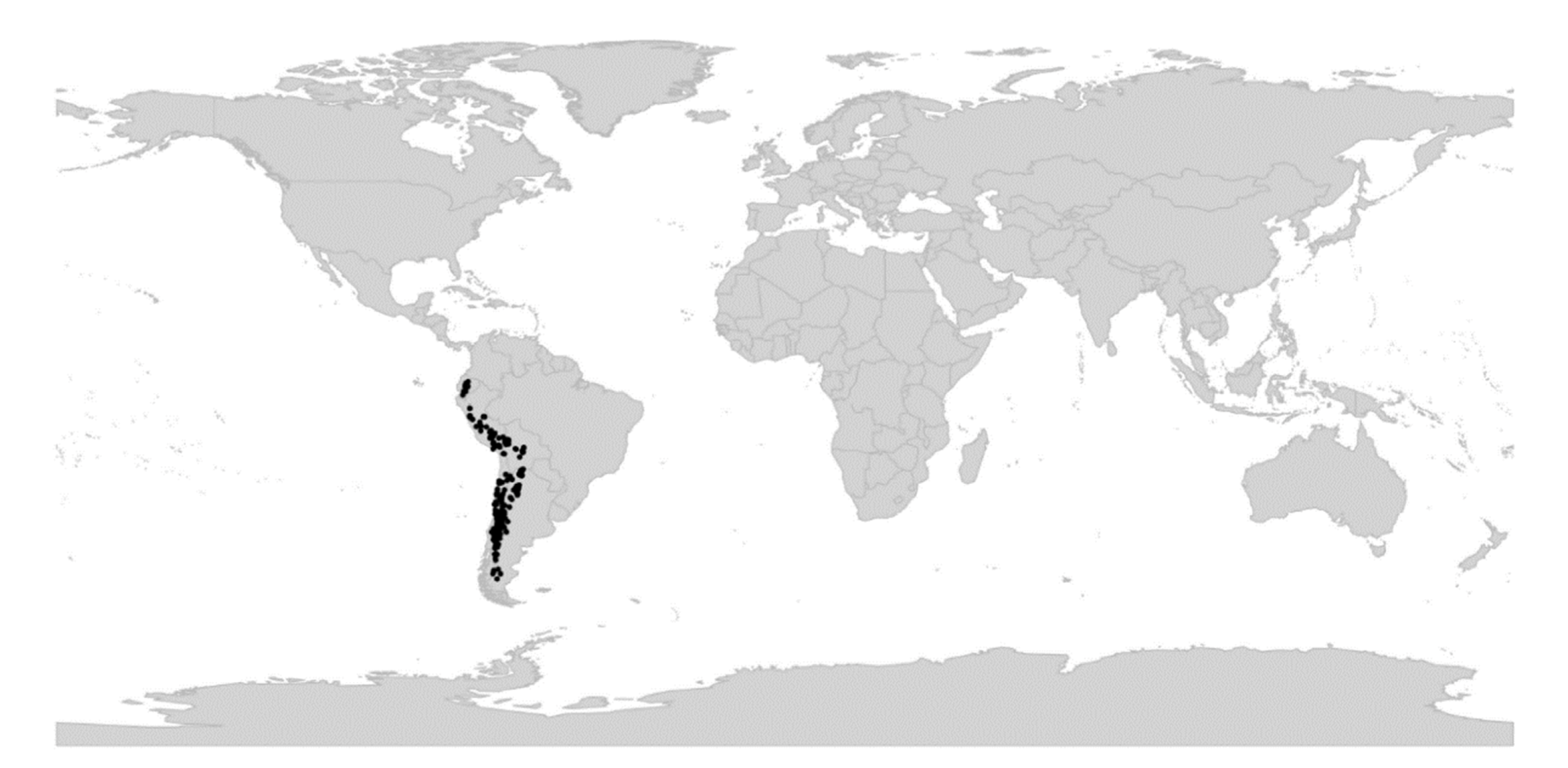{kind=link}
{kind=link}
{kind=link}
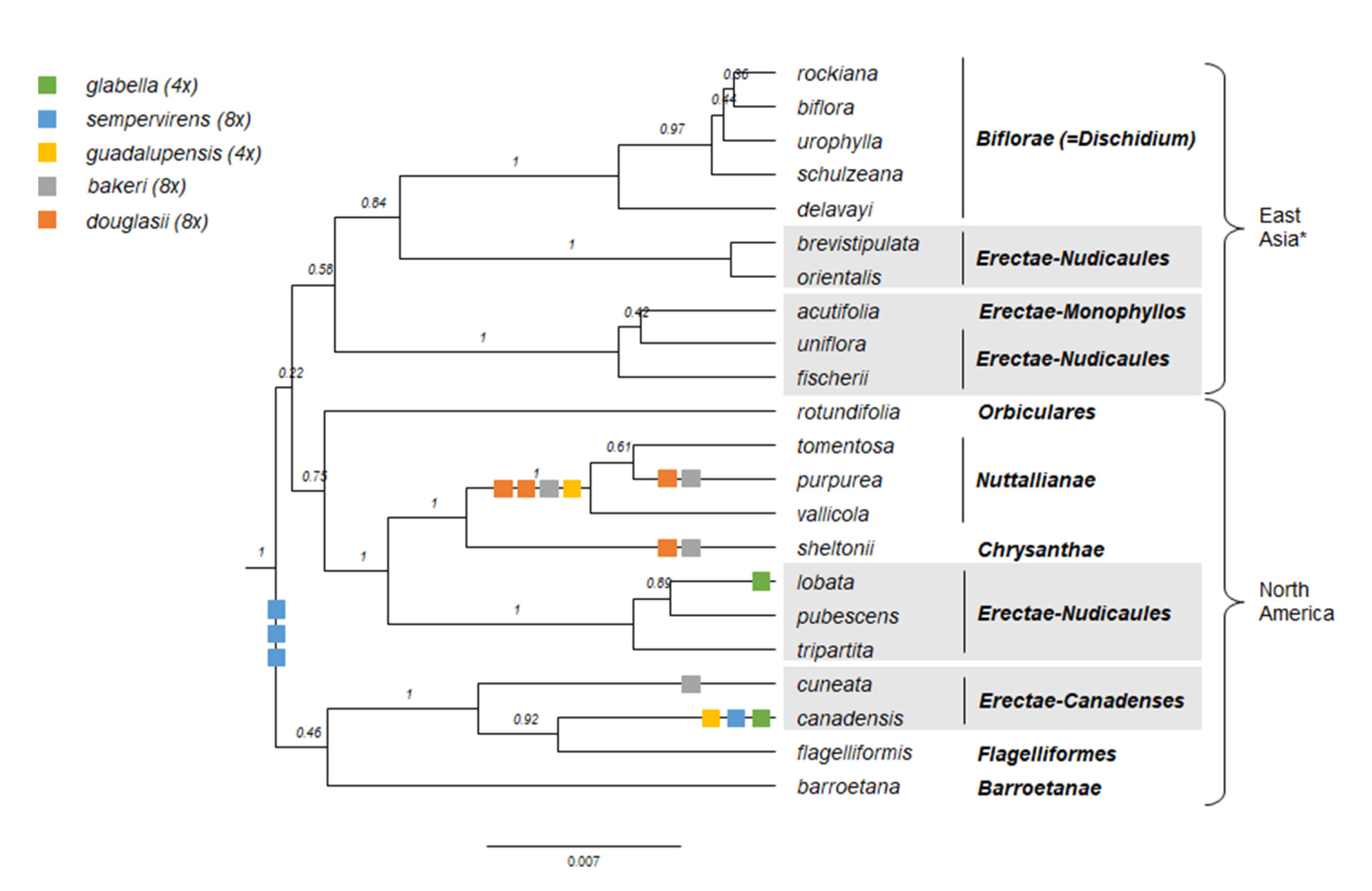{kind=link}
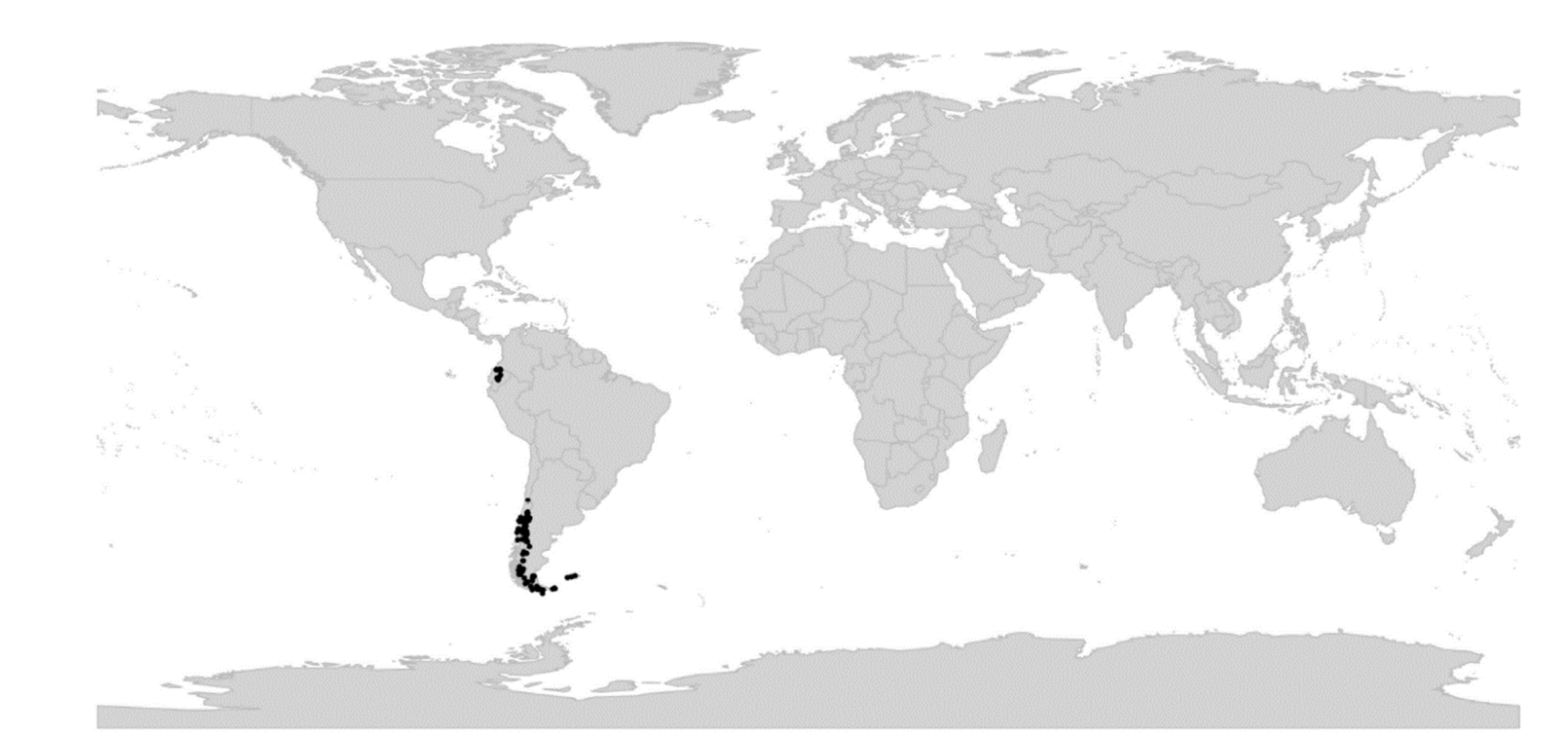{kind=link}
{kind=link}
{kind=link}
{kind=link}
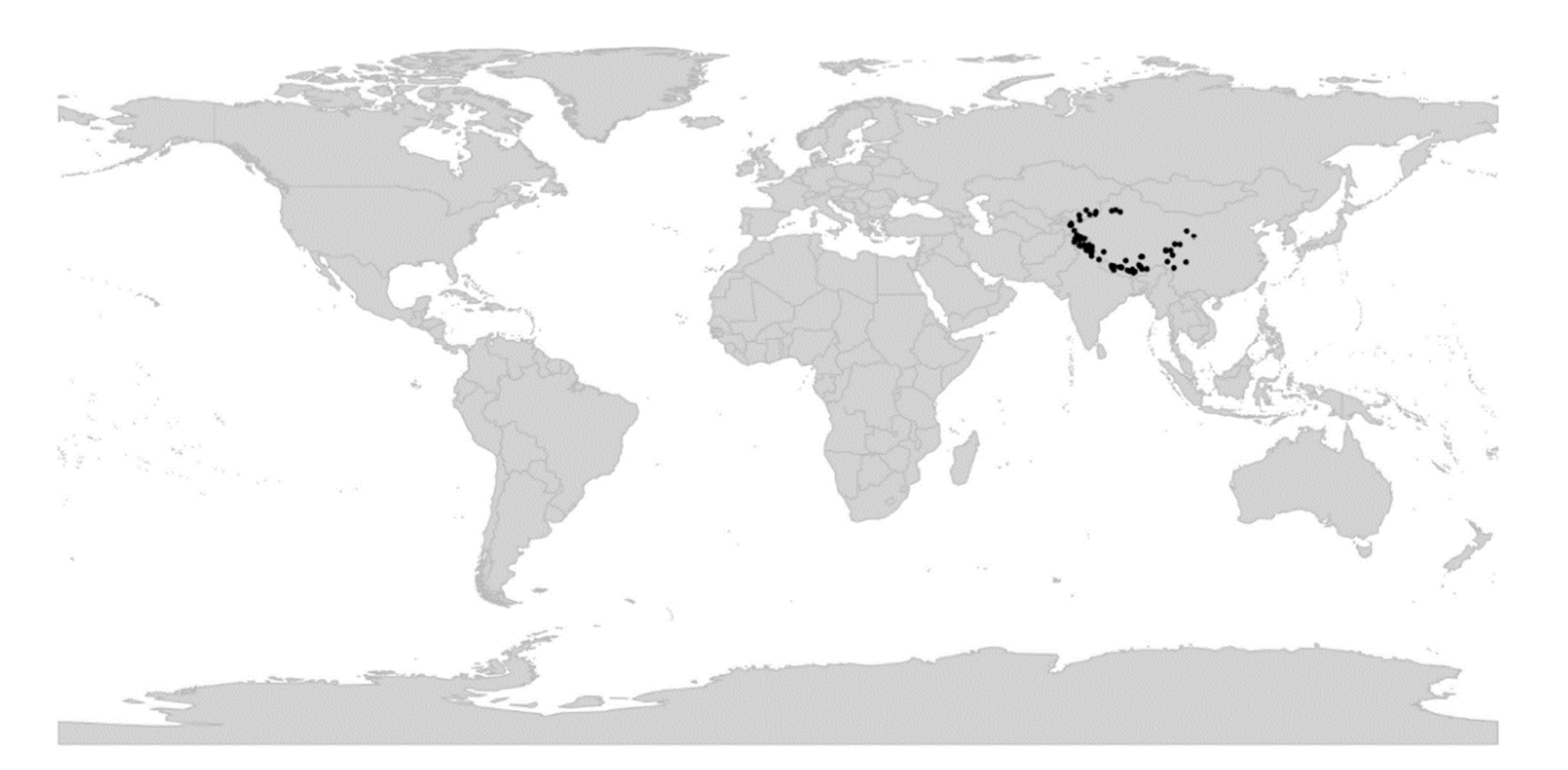{kind=link}
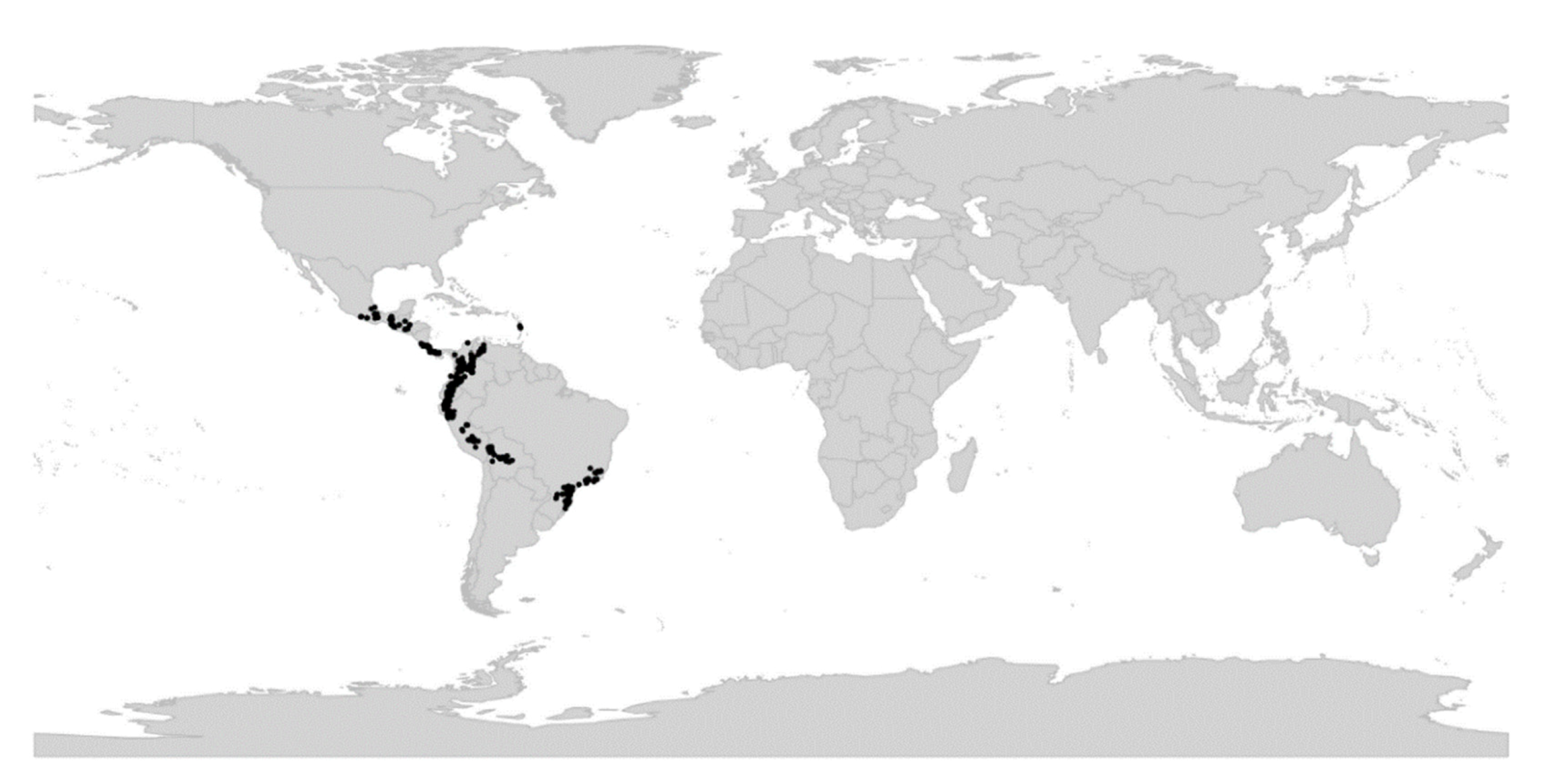{kind=link}
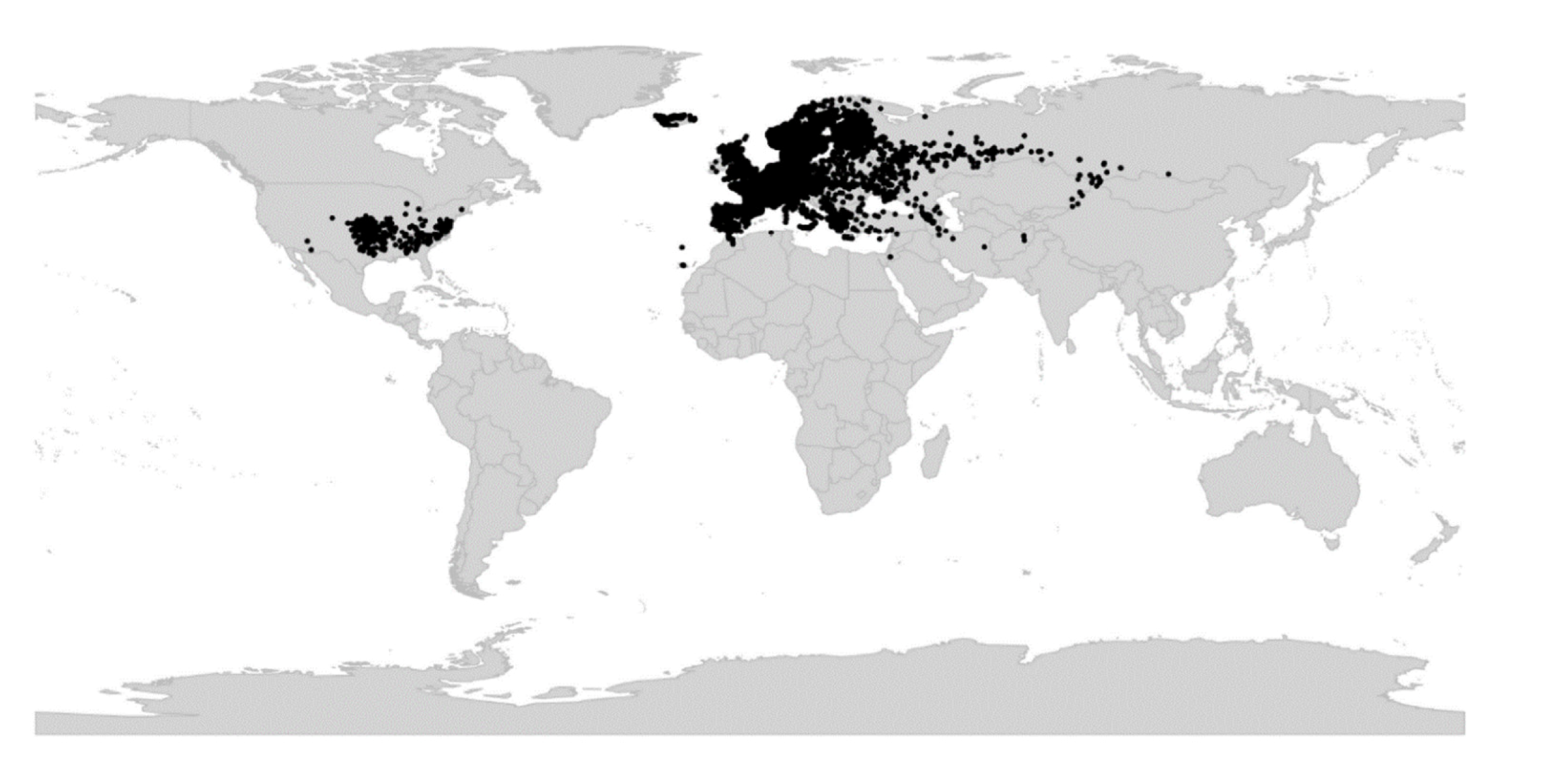{kind=link}
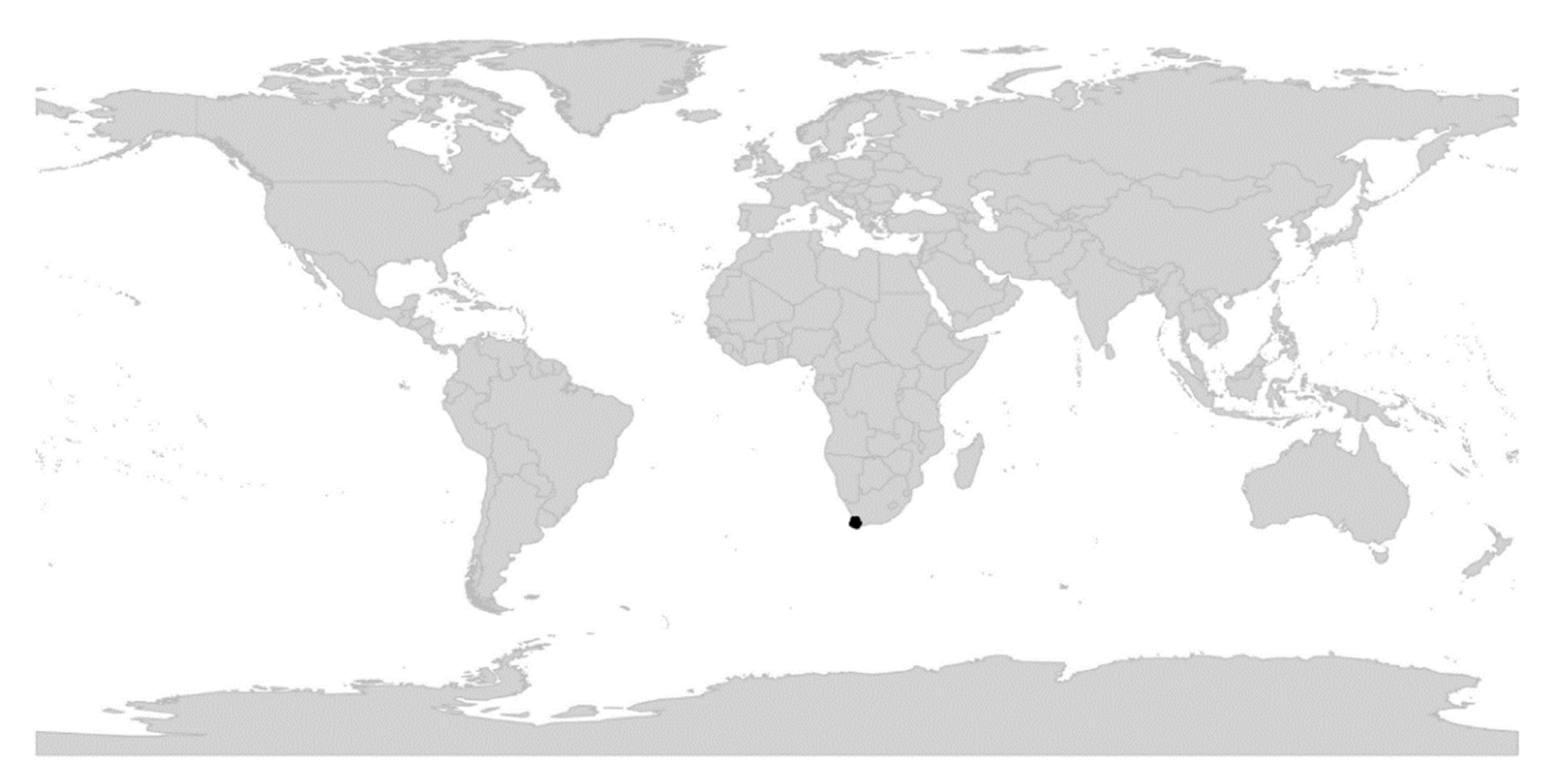{kind=link}
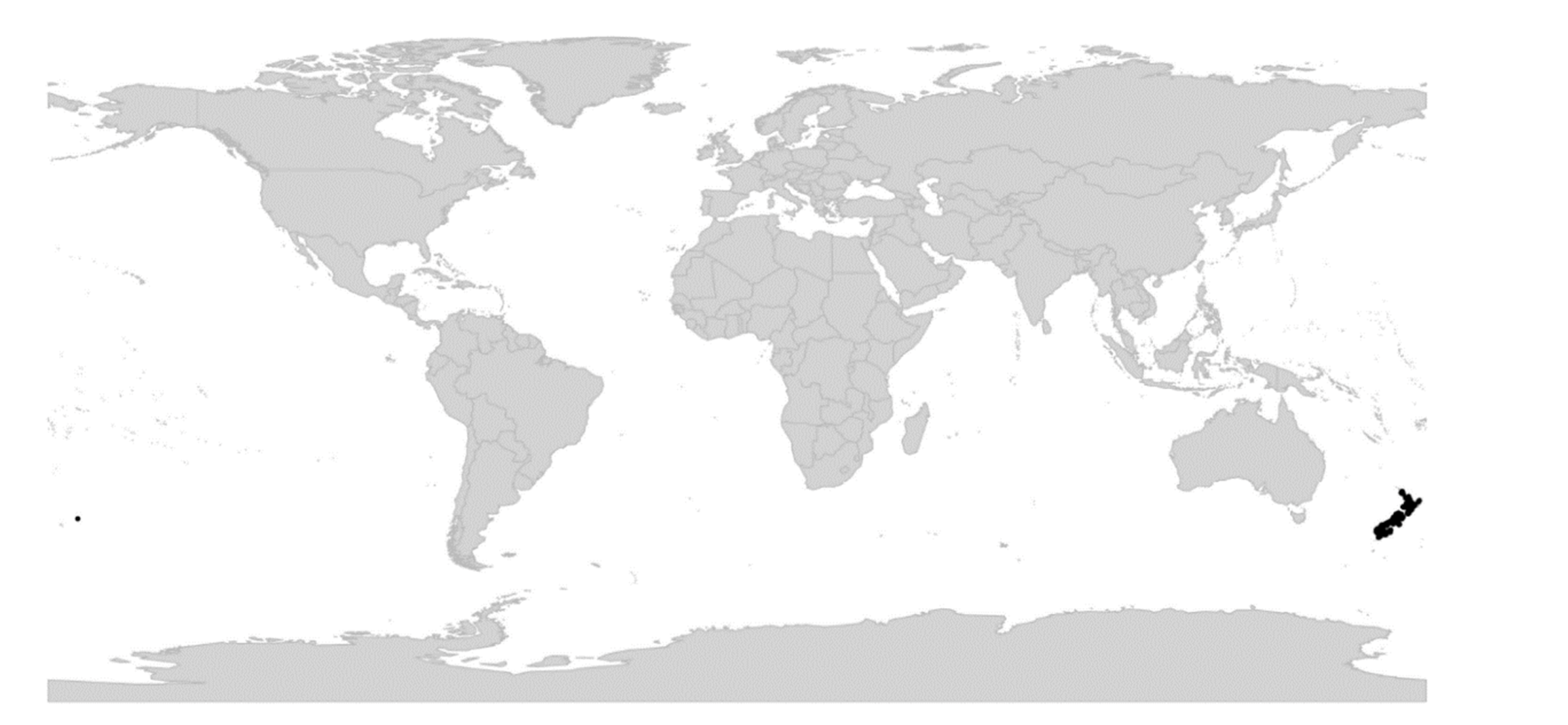{kind=link}
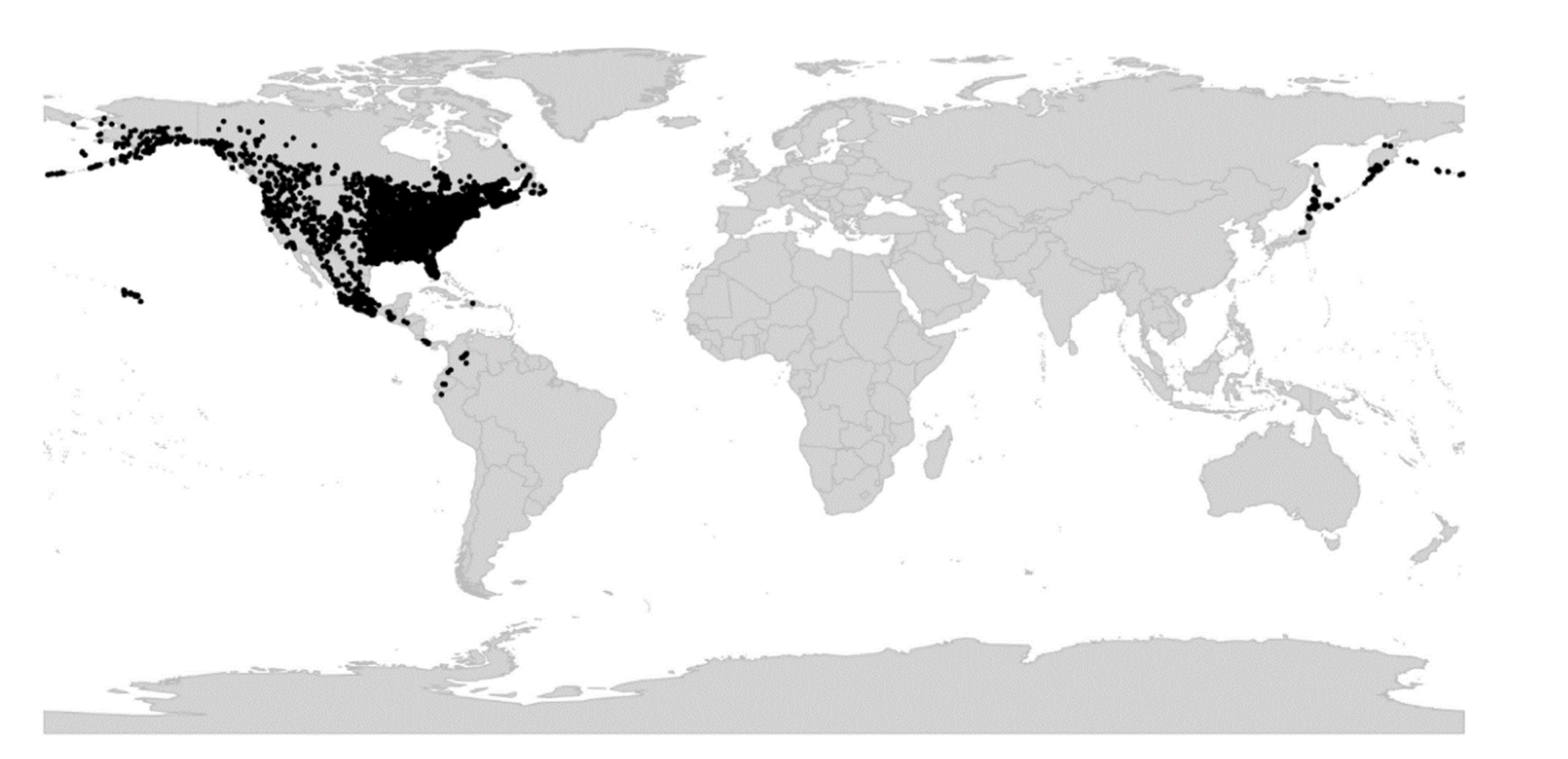{kind=link}
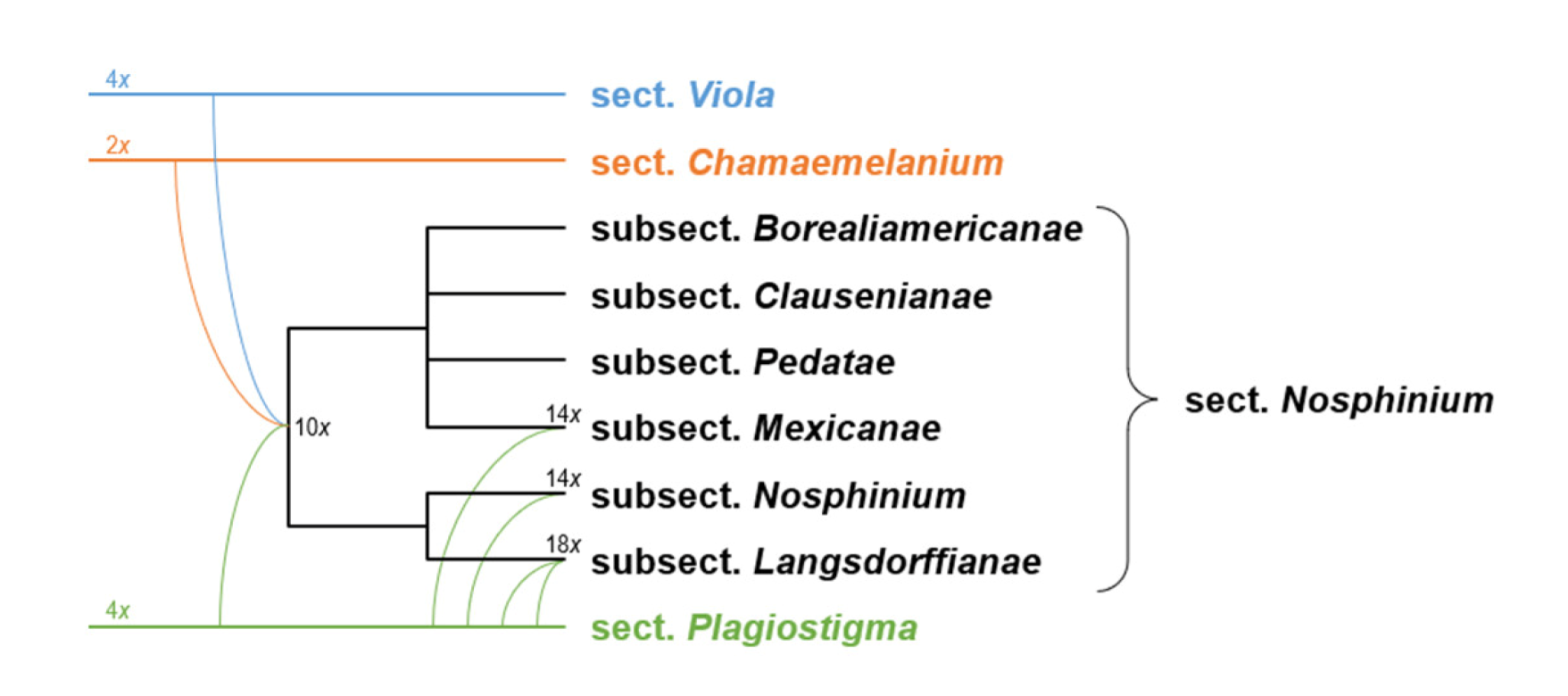{kind=link}
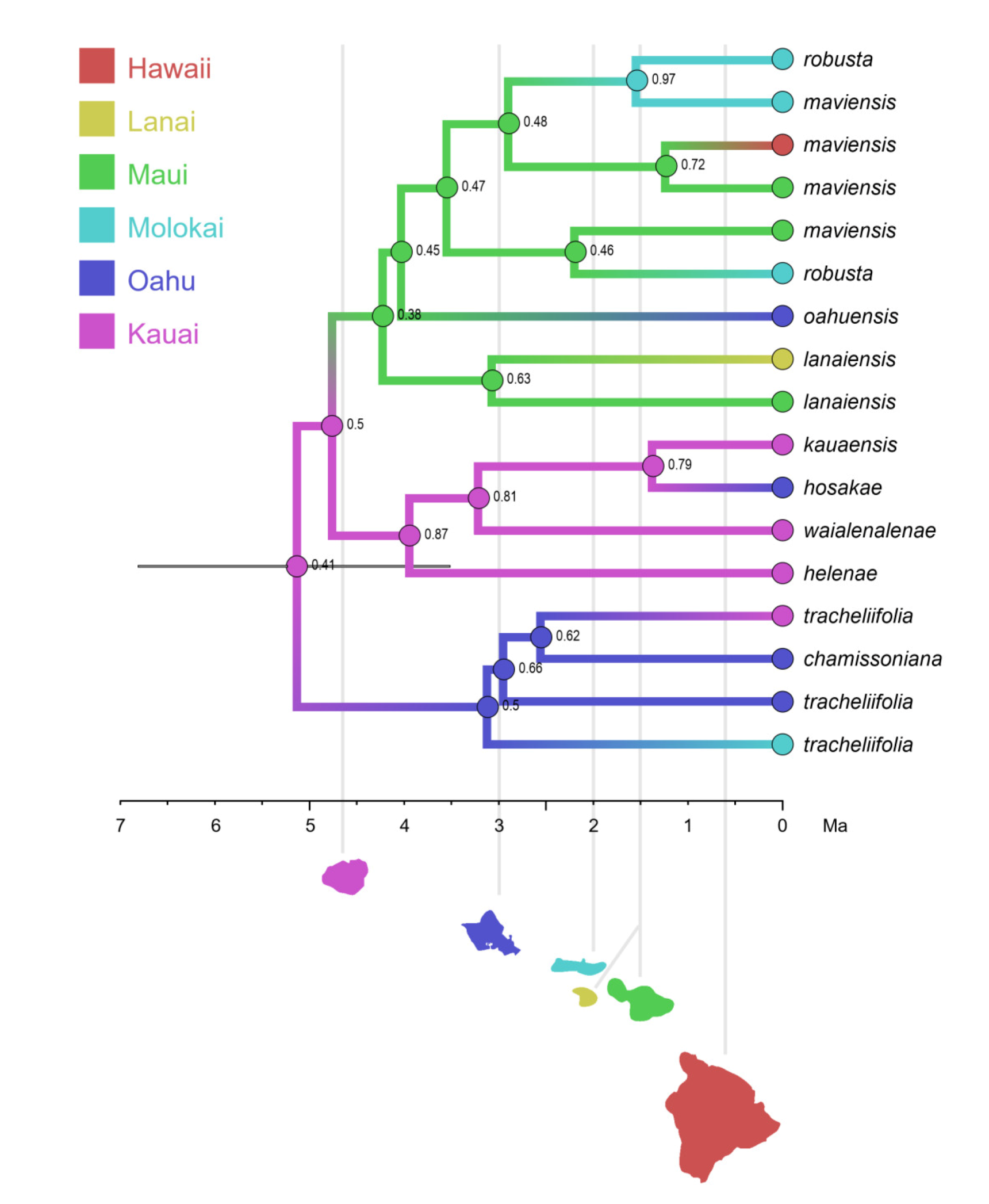{kind=link}
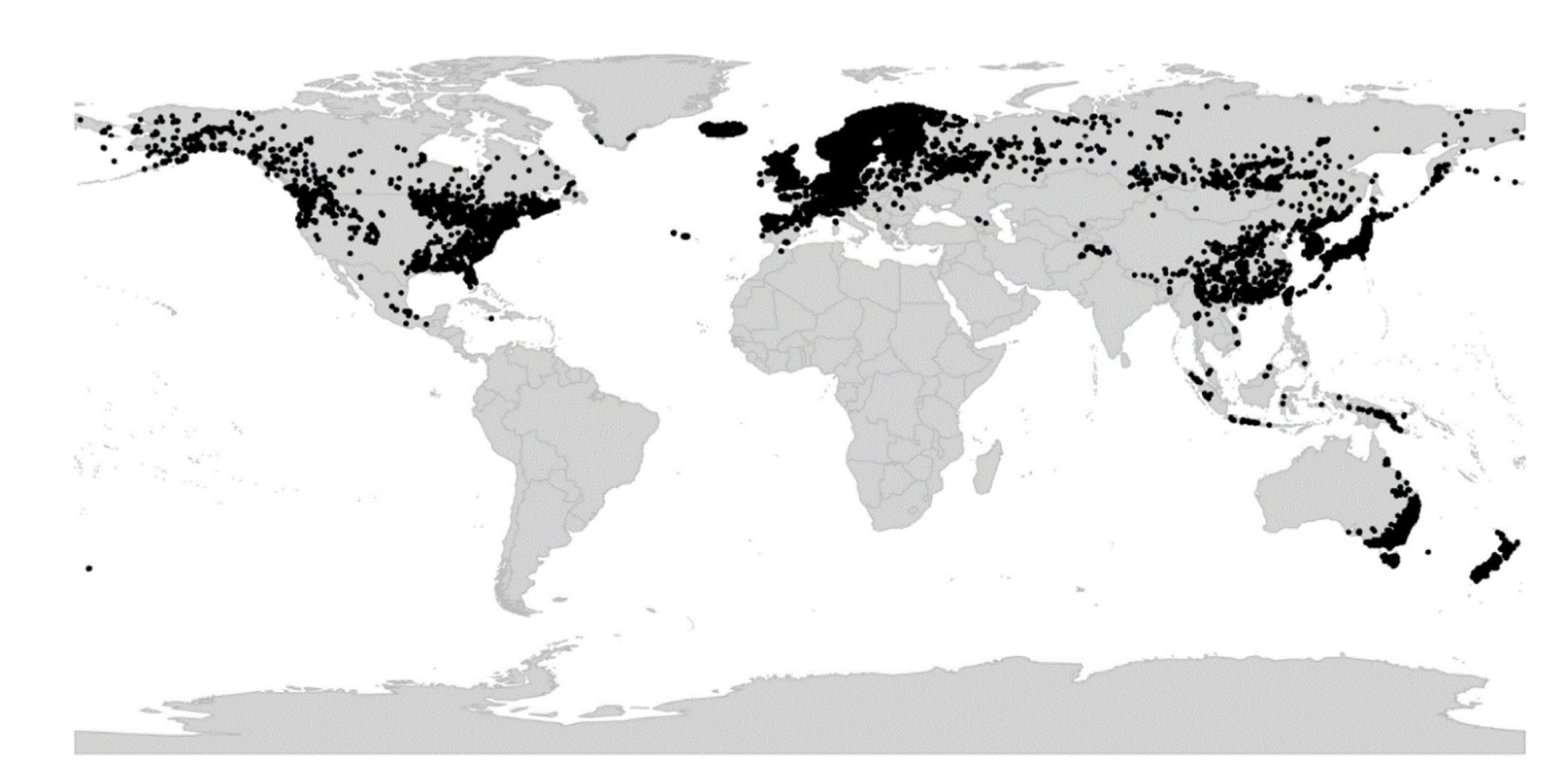{kind=link}
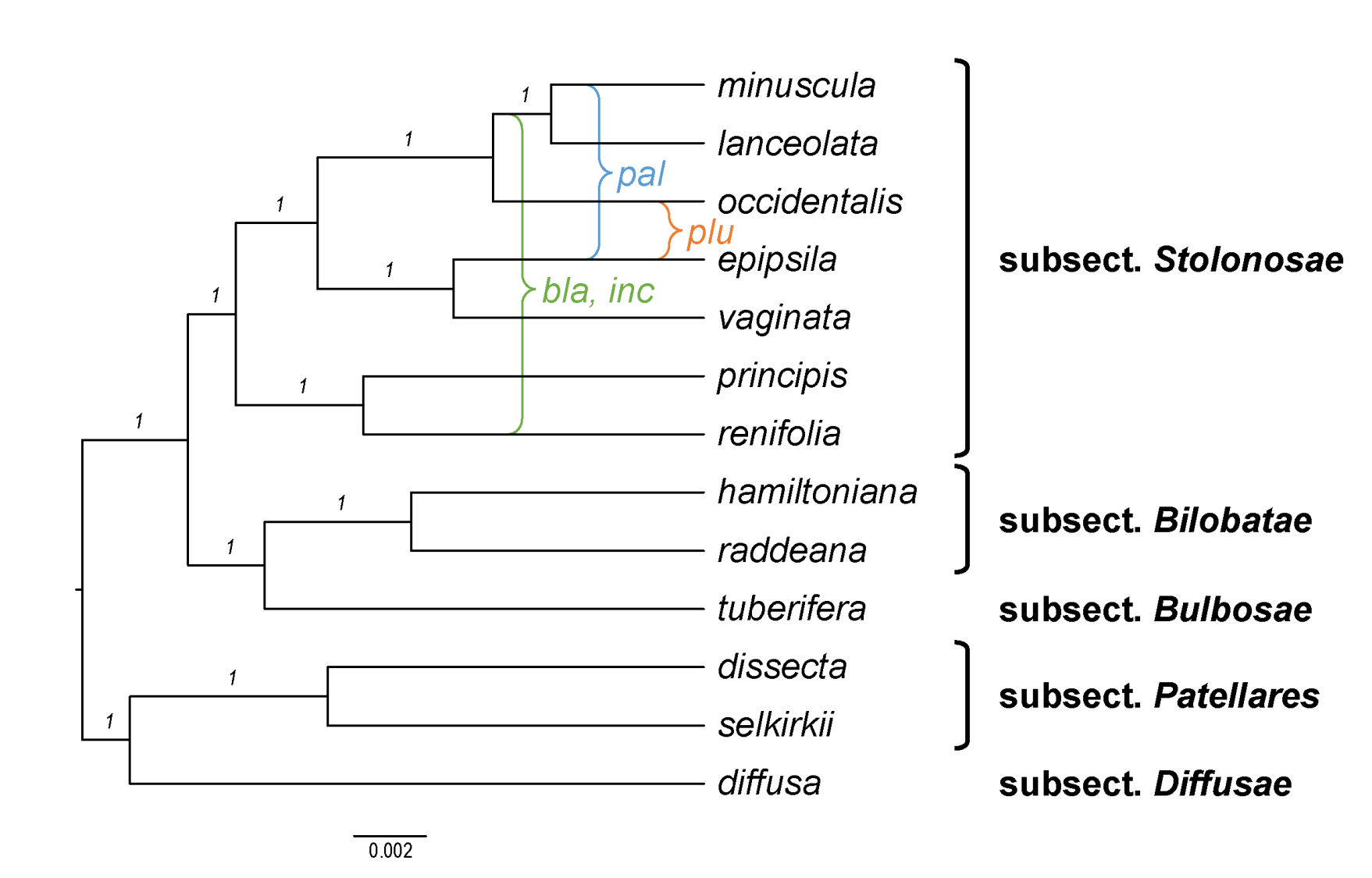{kind=link}
{kind=link}
{kind=link}
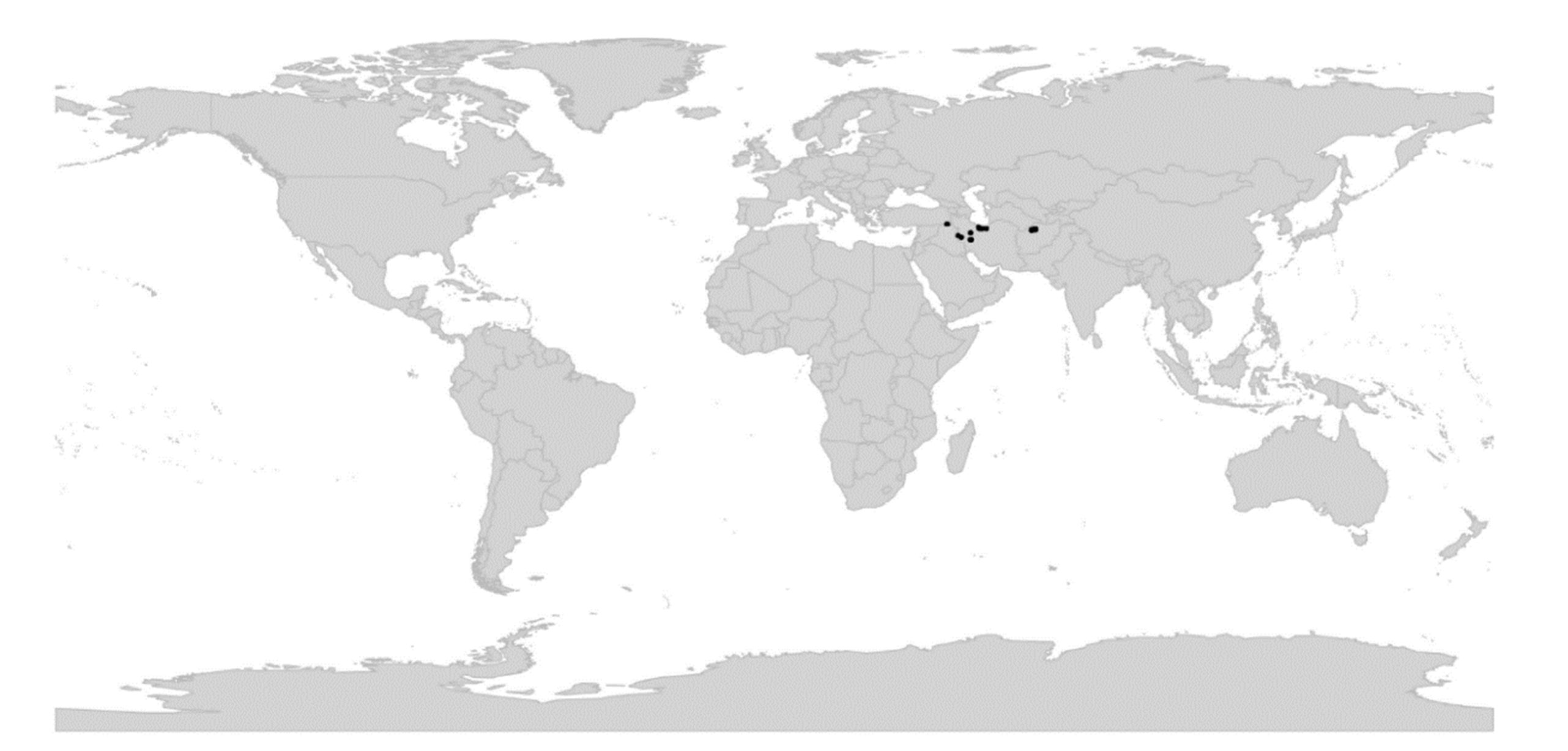{kind=link}
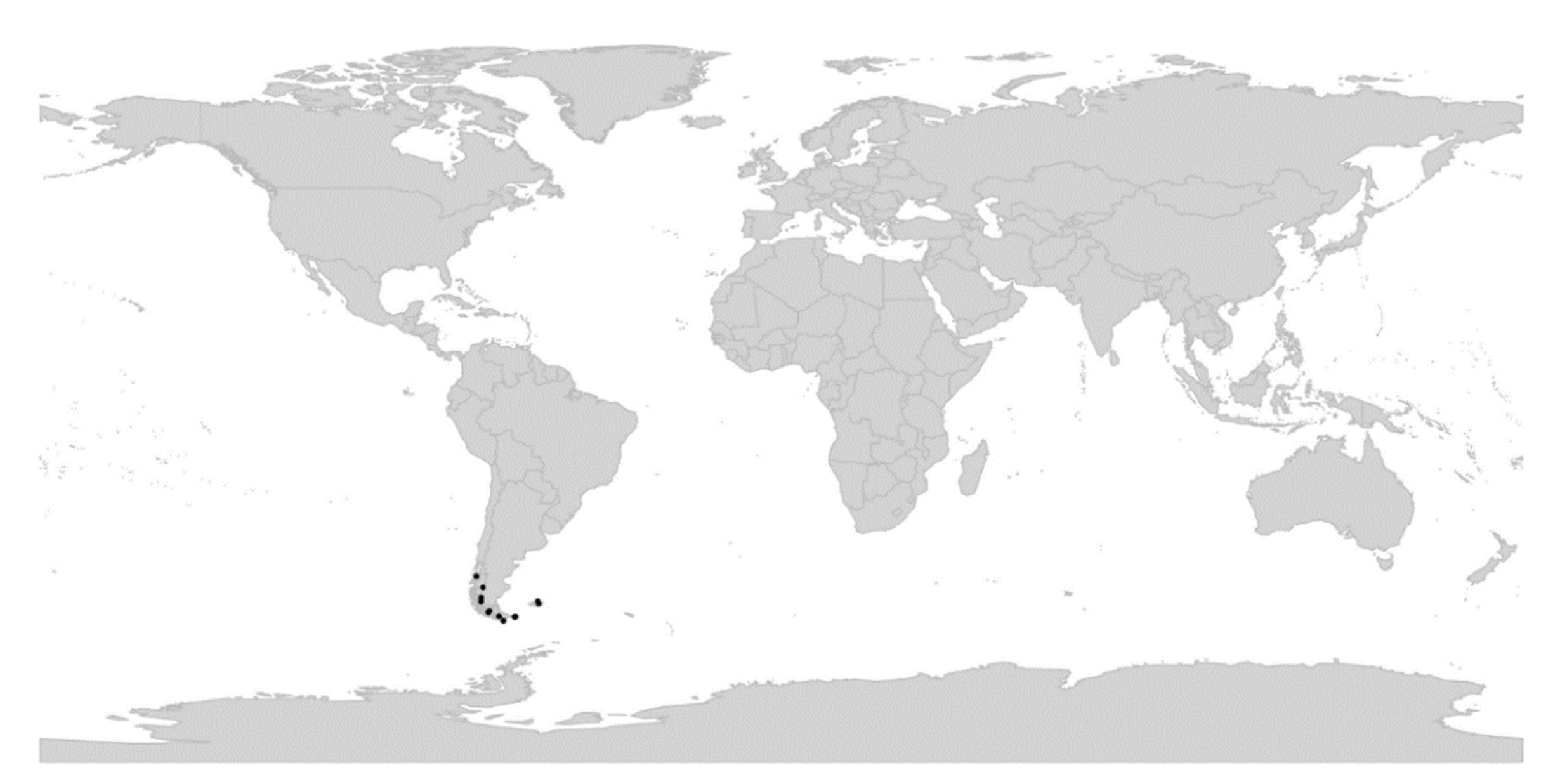{kind=link}
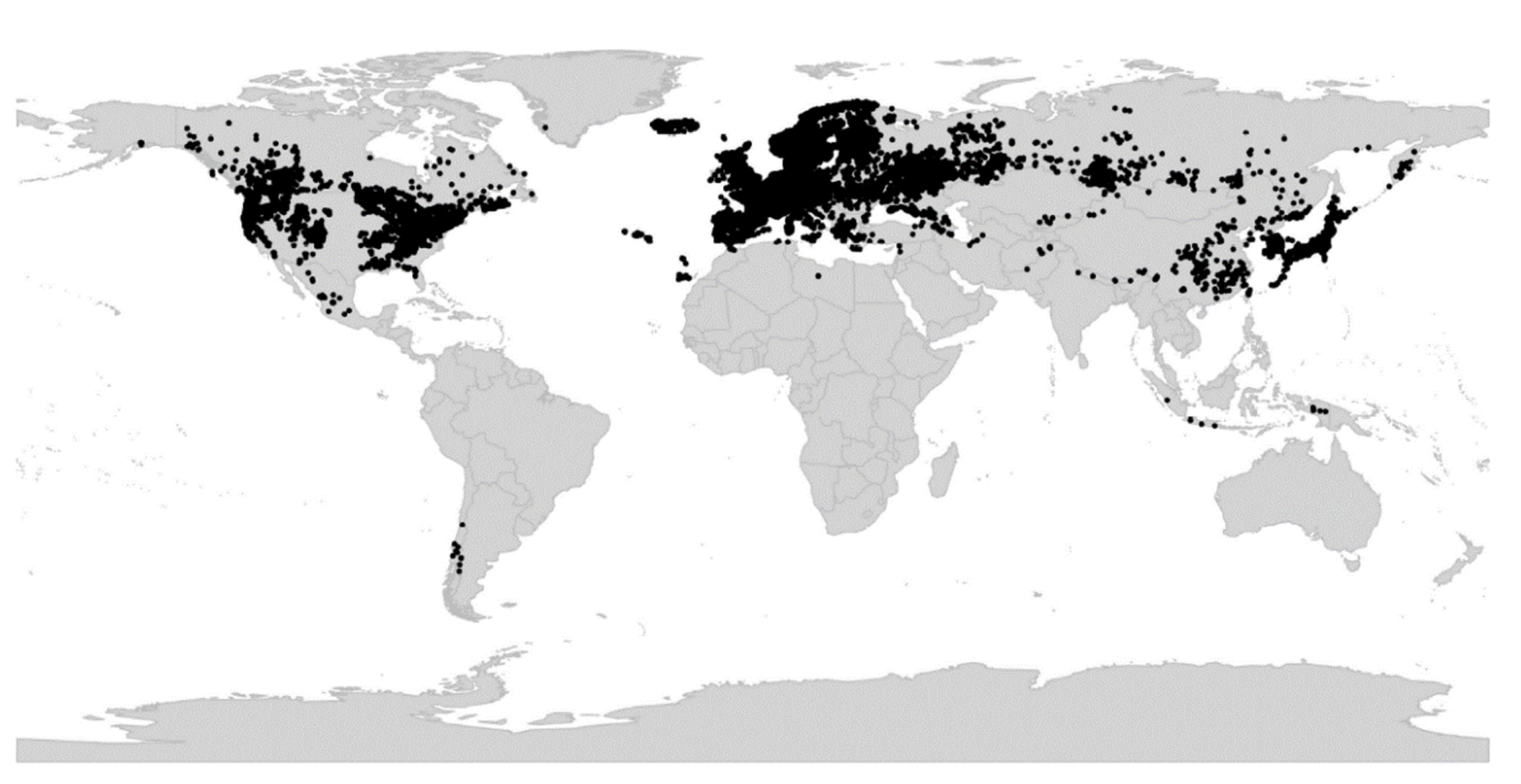{kind=link}
{kind=link}
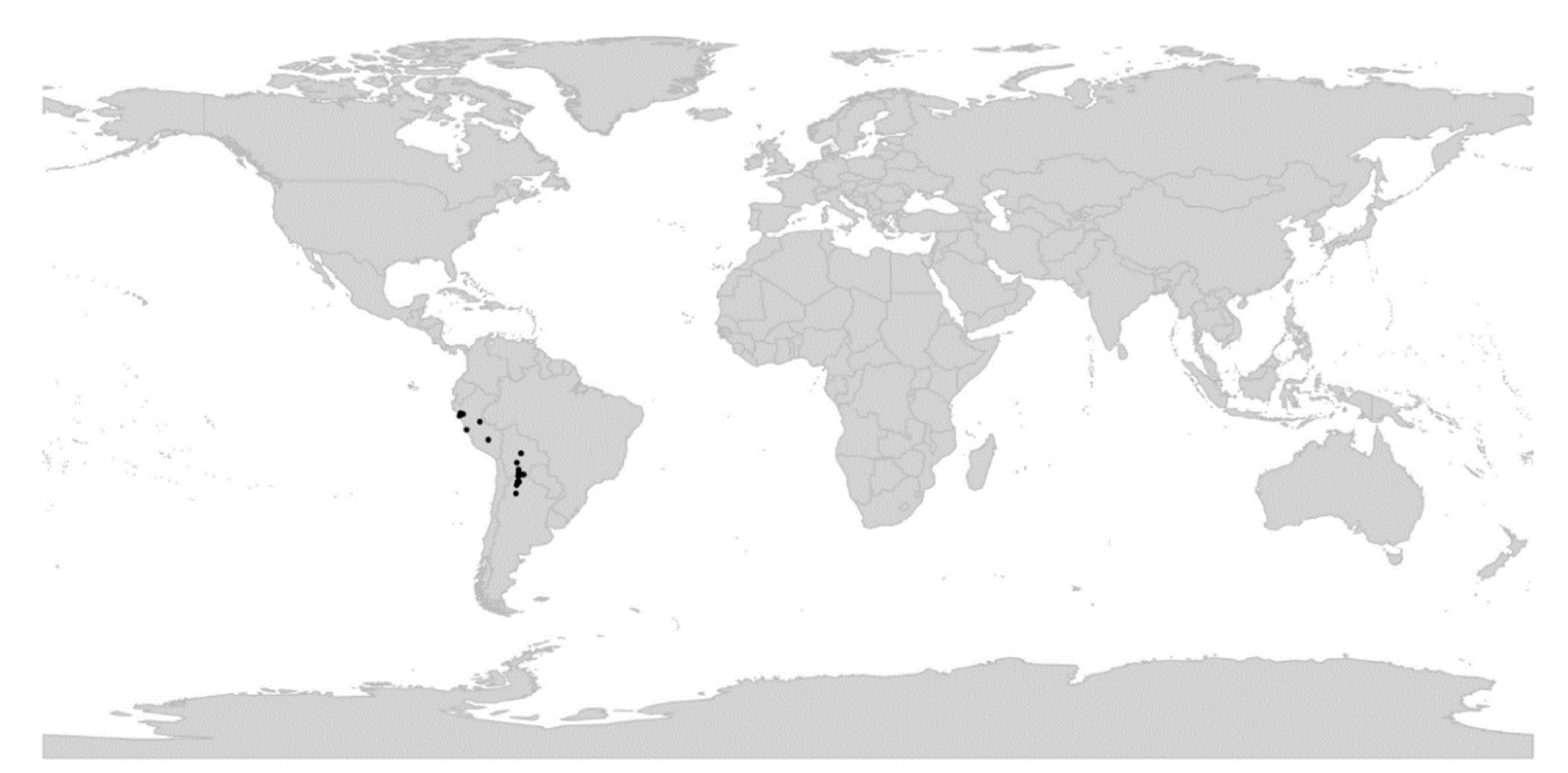{kind=link}
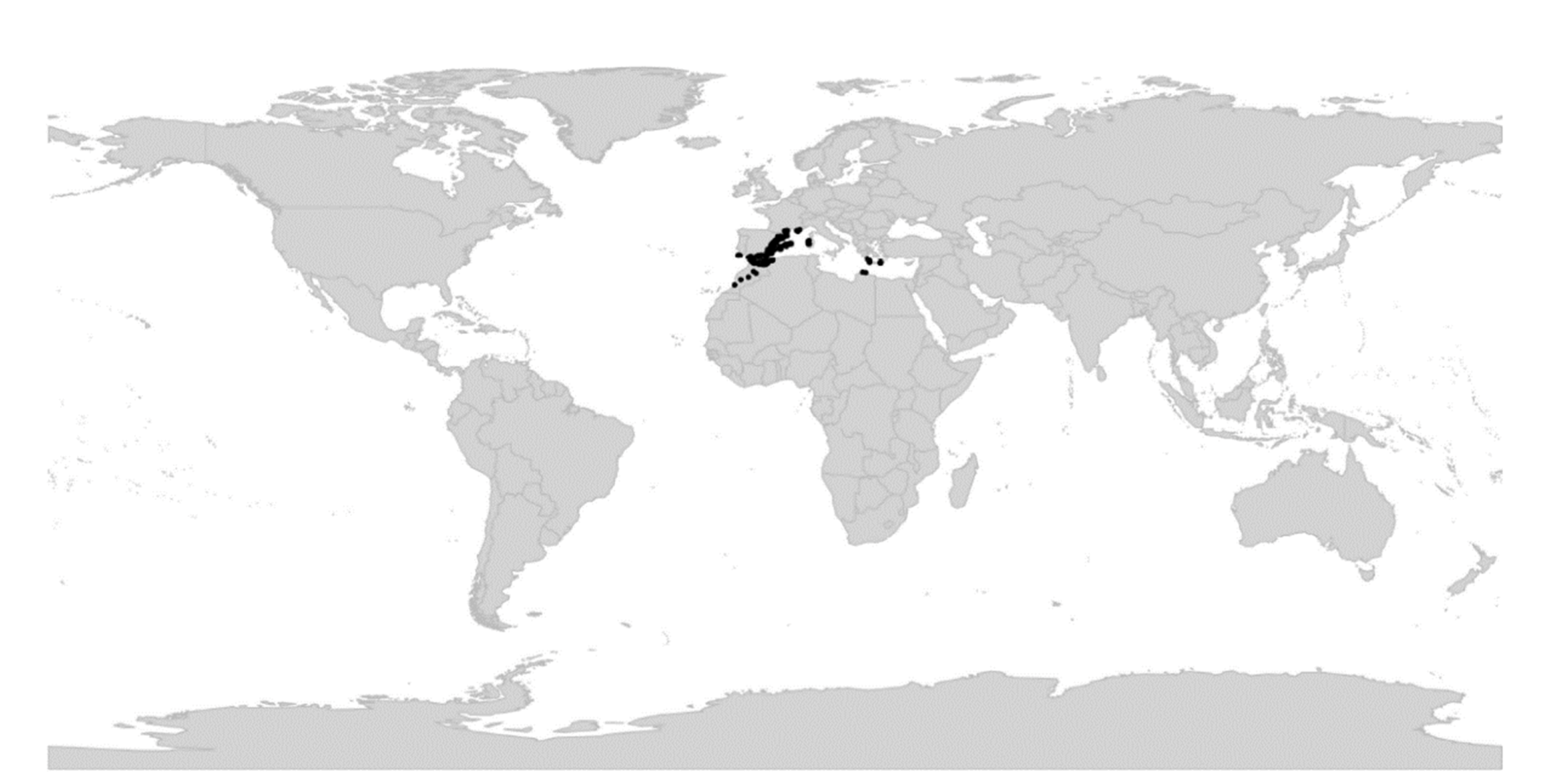{kind=link}
Table 1.
The proposed infrageneric classification of the 664 recognised species of Viola into two subgenera, 31 sections and 20 subsections. “-“ indicates missing data. “?” indicates unknown state. Chromosome numbers within square brackets indicate expected numbers based on ploidy and ancestry.
Table 1.
The proposed infrageneric classification of the 664 recognised species of Viola into two subgenera, 31 sections and 20 subsections. “-“ indicates missing data. “?” indicates unknown state. Chromosome numbers within square brackets indicate expected numbers based on ploidy and ancestry.
| Genus Segregate | Figure | Ploidy (x) | Base 2n | Species | Distribution | Type Species |
|---|---|---|---|---|---|---|
| Subg. Neoandinium | - | 2x | 14 | 139 | South America | Viola pygmaea |
| sect. Confertae | 3a | - | - | 1 | Chile? | V. nassauvioides |
| sect. Ericoidium | 3b | - | - | 1 | Argentina, Chile | V. fluehmannii |
| sect. Grandiflos | 3c | - | - | 6 | Argentina, Chile | V. truncata |
| sect. Inconspicuiflos | 3d–e | - | - | 8 | Peru | V. lilliputana |
| sect. Relictium | 3e | - | - | 8 | Chile | V. huesoensis |
| sect. Rhizomandinium | 3g | - | - | 2 | Argentina | V. escondidaensis |
| sect. Rosulatae | 3h | 2x | 14 | 55 | South America | V. rosulata |
| sect. Sempervivum | 3i | - | - | 34 | South America | V. atropurpurea |
| sect. Subandinium | 3j | 2x | - | 15 | South America | V. subandina |
| sect. Triflabellium | 3k | - | - | 7 | Argentina, Chile | V. triflabellata |
| sect. Xylobasis | 3l | - | - | 1 | Argentina | V. beati |
| Subg. Viola | - | 2x | 14? | 525 | cosmopolitan | V. odorata |
| sect. Abyssinium | 3m | 12x | c. 72 | 3 | Africa | V. abyssinica |
| sect. Chamaemelanium | 3n | 2x | 12 | 69 | northern hemisphere | V. canadensis |
| sect. Chilenium | 3o | ≥4x | - | 7 | South America | V. maculata |
| sect. Danxiaviola | 3p | 4x? | 20 | 1 | China: Guangdong | V. hybanthoides |
| sect. Delphiniopsis | 3q | 4x | 20 | 3 | southern Europe | V. delphinantha |
| sect. Erpetion | 3r | 8x | 50 | 11 | Australia | V. hederacea |
| sect. Himalayum | 3s | 8x | 20? | 1 | central Asia | V. kunawurensis |
| sect. Leptidium | 3t | 4x | 54 | 18 | Latin America | V. stipularis |
| sect. Melanium | - | 4x | ? | 112 | northern hemisphere | V. tricolor |
| —subsect. Bracteolatae | 3u | 8x | ? | 98 | western Palearctic | V. tricolor |
| —subsect. Cleistogamae | 3v | 8x | 34 | 1 | eastern North America | V. rafinesquei |
| —subsect. Dispares | 3w | 4x? | ? | 3 | Mediterranean region | V. dyris |
| —subsect. Ebracteatae | 3x | 4x | ? | 9 | Mediterranean region | V. modesta |
| —subsect. Pseudorupestres | 3y | 4x? | 14 | 1 | Alps & Corsica | V. nummulariifolia |
| sect. Melvio | 3z | 6x? | - | 1 | South Africa | V. decumbens |
| sect. Nematocaulon | 3aa | - | 72 | 1 | New Zealand | V. filicaulis |
| sect. Nosphinium | - | 10x | [56] | 62 | mainly North America | V. chamissoniana |
| —subsect. Borealiamericanae | 3ab | 10x | 54 | 38 | North America | V. cucullata |
| —subsect. Clausenianae | 3ac | 10x | c. 44? | 1 | USA: Utah | V. clauseniana |
| —subsect. Langsdorffianae | 3ad | 14x | [80] | 3 | Amphiberingian | V. langsdorffii |
| —subsect. Mexicanae | 3ae | 14x | [80] | 10 | Mexico to Ecuador and Venezuela | V. humilis |
| —subsect. Nosphinium | 3af | 14x | [80] | 9 | Hawaiian Islands | V. chamissoniana |
| —subsect. Pedatae | 3ag | 10x | 54 | 1 | eastern North America | V. pedata |
| sect. Plagiostigma | - | 4x | 24 | 142 | cosmopolite except Africa | V. palustris |
| —subsect. Australasiaticae | 3ah | 8x? | 46? | 10 | southeastern Asia and Malesia | V. sumatrana |
| —subsect. Bilobatae | 3ai | 4x | 24 | 8 | eastern Asia and Australasia | V. arcuata |
| —subsect. Bulbosae | 3aj | 4x | 24 | 2 | eastern Himalayas and central China | V. bulbosa |
| —subsect. Diffusae | 3ak | 4x | 24 | 17 | southeastern Asia | V. diffusa |
| —subsect. Formosanae | 3al | 4x | 22 | 2 | Taiwan and Okinawa | V. formosana |
| —subsect. Patellares | 3am | 4x | 24 | 62 | northern hemisphere | V. selkirkii |
| —subsect. Stolonosae | 3an | 4x | 24 | 41 | northern hemisphere | V. palustris |
| sect. Rubellium | 3ao | 2x | 12 | 3 | Chile | V. rubella |
| sect. Sclerosium | 3ap | 4x | 22 | 7 | Indian Ocean monsoon region | V. cinerea |
| sect. Spathulidium | 3aq | 8x | - | 3 | Iraq, Iran, Afganistan | V. spathulata |
| sect. Tridens | 3ar | 6x | 40 | 1 | southern South America | V. tridentata |
| sect. Viola | - | 4x | 20 | 75 | near cosmopolite | V. odorata |
| —subsect. Rostratae | 3as | 4x | 20 | 51 | near cosmopolite | V. riviniana |
| —subsect. Viola | 3at | 4x | 20 | 24 | Palearctic | V. odorata |
| sect. Xanthidium | 3av | - | - | 2 | central South America | V. flavicans |
| sect. Xylinosium | 3aw | 8x? | 52 | 3 | Mediterranean region | V. arborescens |
Table 2.
Inferred ploidy for 11 species of Viola sect. Melanium based on GPI homoeolog number [28] and estimated genome size as gigabases (Gb) and 1C. Genome size data were downloaded from the Plant DNA C-values Database [253] and Genomes on a Tree (GoaT; accessed on 10 March 2022), which presents genome-relevant metadata for Eukaryotic taxa to be sequenced by the Earth Biogenome Project [233]. “-” indicates missing data.
Table 2.
Inferred ploidy for 11 species of Viola sect. Melanium based on GPI homoeolog number [28] and estimated genome size as gigabases (Gb) and 1C. Genome size data were downloaded from the Plant DNA C-values Database [253] and Genomes on a Tree (GoaT; accessed on 10 March 2022), which presents genome-relevant metadata for Eukaryotic taxa to be sequenced by the Earth Biogenome Project [233]. “-” indicates missing data.
| Species | Inferred Ploidy | 2n = | GPI Homoeologs | Genome Size (Gb) | Genome Size (1C, pg) | Subsect. |
|---|---|---|---|---|---|---|
| V. dirimliensis | 4x | 8 | 2 | - | 1.07 (herein) | Ebracteatae |
| V. rafinesquei | 8x | 34 | 4 | - | - | Cleistogamae |
| V. beckiana | 8x | 20 | - | 1.32 | 1.35 [219] | Bracteolatae |
| V. cornuta | 8x | 22 | - | 1.25 | 1.18 [221] | Bracteolatae |
| V. elegantula | 8x | 20 | - | 1.32 | 1.35 [220] | Bracteolatae |
| V. kitaibeliana | 8x | 16 | - | - | 1.10 [218] | Bracteolatae |
| V. tricolor | 12x | 26 | 6 [236] | 2.07 | 1.76 to 1.78 [218] | Bracteolatae |
| V. arvensis | 16x | 34 | - | - | 2.23 [218] | Bracteolatae |
| V. calcarata | 16x | 40 | 8 | 2.82 | 2.89 [221] | Bracteolatae |
| V. lutea | 20x | 48 | - | - | 2.75 [218], 3.13 [221] | Bracteolatae |
| V. bubaniii | 20x | 52 | - | 3.32 | 3.39 [221] | Bracteolatae |
Table 3.
Taxonomic characters delimiting subsections within Viola sect. Melanium.
| Character\Subsection | Subsect. Pseudorupestres | Subsect. Ebracteatae | Subsect. Dispares | Subsect. Cleistogamae | Subsect. Bracteolatae |
|---|---|---|---|---|---|
| Life history/durancy | perennial | annual | annual or perennial | annual to biennial | annual to perennial |
| Cleistogamous flowers | not produced | not produced | not produced | produced | not produced |
| Lamina of basal leaves | entire | entire or subcrenate | entire or crenate | crenate | entire or crenate |
| Colour of corolla throat | cream | bright yellow | bright yellow | bright yellow | bright yellow, rarely cream |
| Ploidy | probably 4x | 4x, 8x, >8x | probably 4x, 8x | 8x | 8x, 12x, 16x, 20x |
| Bottom petal length, spur included (mm) | 9.5–11.5 | 2–11.5 | 5–13 | 8–10 | 5.5–34 |
| Spur length (mm) | 2.3–2.5 | 0.9–3 | 1–3.5 | 1–1.5 | 1.8–16 |
| Calycine appendage length (mm) | 1.2–1.5 | 0.5–5.3 | 0.3–1 | 0.5–2 | 0.9–4.7 |
| Pollen apertures | 3 | 3 or heteromorphic 4 | 3 or 4 | 4 | 4 or 5 heteromorphic |
Table 4.
GenBank sequence IDs for the ITS sequences used in the phylogeny in Figure 6. Taxon names have been edited to correspond to the nomenclature used herein. Metadata for each sequence are available at GenBank/NCBI.
Table 4.
GenBank sequence IDs for the ITS sequences used in the phylogeny in Figure 6. Taxon names have been edited to correspond to the nomenclature used herein. Metadata for each sequence are available at GenBank/NCBI.
| Infrageneric Classification | Species | GenBank Sequence IDs |
|---|---|---|
| sect. Rosulatae | Viola philippii | MH792062 |
| sect. Sempervivum | V. cotyledon | ON133602 |
| sect. Sempervivum | V. micranthella | AF097222, AF097268 |
| sect. Subandinium | V. subandina | MH781265 |
| sect. Subandinium | V. yrameae, ined. | ON133601 (as Viola sp. MVN-2022a) |
| sect. Abyssinium | V. abyssinica | MN723993 |
| sect. Chamaemelanium | V. biflora | DQ055348 |
| sect. Chamaemelanium | V. canadensis | AF097231, MG234951 |
| sect. Chamaemelanium | V. pubescens | DQ006044 |
| sect. Chamaemelanium | V. sempervirens | MG235908 |
| sect. Chamaemelanium | V. sheltonii | AF097226, AF097272 |
| sect. Chamaemelanium | V. uniflora | AY582167, AY541600 |
| sect. Chamaemelanium | V. urophylla | MH117805 |
| sect. Chilenium | V. reichei | AF097223, AF097269 |
| sect. Danxiaviola | V. hybanthoides | KF011244 (as Viola sp. LWB-2013a) |
| sect. Delphiniopsis | V. cazorlensis | AY148230, AY148250 |
| sect. Himalayum | V. kunawurensis | NCBI accession PRJNA805692 (as V. “kunawarensis”) |
| sect. Leptidium | V. scandens | AF097221, AF097267 |
| sect. Melanium subsect. Bracteolatae | V. cornuta | AY582166, MT367013 |
| sect. Melanium subsect. Bracteolatae | V. heldreichiana | MT367025 |
| sect. Melanium subsect. Bracteolatae | V. kitaibeliana | AY148235, KX166474, MT367029 |
| sect. Melanium subsect. Bracteolatae | V. paradoxa | MT367093 |
| sect. Melanium subsect. Bracteolatae | V. tricolor | DQ055396 |
| sect. Melanium subsect. Cleistogamae | V. rafinesquei | MG235080 (as V. bicolor) |
| sect. Melanium subsect. Dispares | V. demetria | MT367018 |
| sect. Melanium subsect. Dispares | V. dyris | MT367069 |
| sect. Melanium subsect. Ebracteatae | V. dirimliensis | ON129460 |
| sect. Melanium subsect. Ebracteatae | V. mercurii | MT367115 |
| sect. Melanium subsect. Ebracteatae | V. modesta | MT367084 |
| sect. Melanium subsect. Ebracteatae | V. occulta | HM851453 |
| sect. Melanium subsect. Ebracteatae | V. parvula | AY148240, AY148260 |
| sect. Melanium subsect. Pseudorupestres | V. nummulariifolia | MT367090 |
| sect. Nosphinium subsect. Borealiamericanae | V. affinis | AF097251, AF097297 |
| sect. Nosphinium subsect. Borealiamericanae | V. cucullata | AF097252, MG237103 |
| sect. Nosphinium subsect. Clausenianae | V. clauseniana | AF097300, AF097254 |
| sect. Nosphinium subsect. Langsdorffianae | V. langsdorffii | AF097259, MG235517 |
| sect. Nosphinium subsect. Mexicanae | V. hemsleyana | AF097258, AF097304 |
| sect. Nosphinium subsect. Mexicanae | V. hookeriana | AF097257, AF097303 |
| sect. Nosphinium subsect. Mexicanae | V. nannei | AF097255, AF097301 |
| sect. Nosphinium subsect. Nosphinium | V. chamissoniana | AF115955, AF115959 |
| sect. Nosphinium subsect. Nosphinium | V. lanaiensis | JN682058 |
| sect. Nosphinium subsect. Pedatae | V. pedata | AF097253, MG237117 |
| sect. Plagiostigma subsect. Australasiaticae | V. austrosinensis | OM406228 |
| sect. Plagiostigma subsect. Australasiaticae | V. kwangtungensis | OM406230 |
| sect. Plagiostigma subsect. Australasiaticae | V. mucronulifera | FJ002910 |
| sect. Plagiostigma subsect. Australasiaticae | V. sumatrana | OM406231 |
| sect. Plagiostigma subsect. Bilobatae | V. hamiltoniana | AY928283 (as V. verecunda) |
| sect. Plagiostigma subsect. Bilobatae | V. raddeana | AY928279 |
| sect. Plagiostigma subsect. Bilobatae | V. triangulifolia | FJ002912 |
| sect. Plagiostigma subsect. Diffusae | V. amamiana | JF830899 |
| sect. Plagiostigma subsect. Diffusae | V. diffusa | MH711723 |
| sect. Plagiostigma subsect. Diffusae | V. guangzhouensis | MW683479 |
| sect. Plagiostigma subsect. Diffusae | V. huizhouensis | MW683486 |
| sect. Plagiostigma subsect. Diffusae | V. lucens | FJ002913 |
| sect. Plagiostigma subsect. Diffusae | V. nanlingensis | FJ002916 |
| sect. Plagiostigma subsect. Diffusae | V. yunnanensis | FJ002915 |
| sect. Plagiostigma subsect. Patellares | V. albida | DQ787762 (as V. chaerophylloides) |
| sect. Plagiostigma subsect. Patellares | V. dissecta | JQ950564 |
| sect. Plagiostigma subsect. Patellares | V. patrinii | AY928298 |
| sect. Plagiostigma subsect. Patellares | V. selkirkii | AY928307 |
| sect. Plagiostigma subsect. Patellares | V. somchetica | HM851457 |
| sect. Plagiostigma subsect. Patellares | V. tashiroi | JF830885 |
| sect. Plagiostigma subsect. Patellares | V. variegata | KC330743 |
| sect. Plagiostigma subsect. Stolonosae | V. suecica | MG237736 (as V. epipsila) |
| sect. Plagiostigma subsect. Stolonosae | V. grandisepala | FJ002903 |
| sect. Plagiostigma subsect. Stolonosae | V. lanceolata | MG235616 |
| sect. Plagiostigma subsect. Stolonosae | V. minuscula | AF097236, AF097282 (as V. macloskeyi subsp. pallens) |
| sect. Plagiostigma subsect. Stolonosae | V. moupinensis | FJ002900 |
| sect. Plagiostigma subsect. Stolonosae | V. palustris | KX166144 |
| sect. Plagiostigma subsect. Stolonosae | V. principis | FJ002904 |
| sect. Plagiostigma subsect. Stolonosae | V. yazawana | AY928289 |
| sect. Rubellium | V. capillaris | AF097220, AF097266 |
| sect. Spathulidium | V. spathulata | HM851456 |
| sect. Viola subsect. Rostratae | V. acuminata | AY928273 |
| sect. Viola subsect. Rostratae | V. grypoceras | AY928280 |
| sect. Viola subsect. Rostratae | V. mirabilis | MK828560 |
| sect. Viola subsect. Rostratae | V. reichenbachiana | DQ055382 |
| sect. Viola subsect. Rostratae | V. shinchikuensis | FJ002885 |
| sect. Viola subsect. Rostratae | V. stagnina | KX166475 |
| sect. Viola subsect. Rostratae | V. striata | AF097247, MG234688 |
| sect. Viola subsect. Rostratae | V. uliginosa | KU949386 |
| sect. Viola subsect. Rostratae | V. umbraticola | AF097244, AF097290 |
| sect. Viola subsect. Rostratae | V. websteri | AY928274 |
| sect. Viola subsect. Viola | V. alba | EU413916 |
| sect. Viola subsect. Viola | V. hirta | EU413946 |
| sect. Viola subsect. Viola | V. hondoensis | AY928272 |
| sect. Viola subsect. Viola | V. odorata | EU413922 |
| sect. Viola subsect. Viola | V. pyrenaica | JF683824 |
| sect. Xylinosium | V. scorpiuroides | MT367099 |
| Outgroup | Melicytus obovatus | EF635462 |
Table 5.
Taxa and GenBank sequence IDs for the phylogenetic analysis of sect. Chamaemelanium. A dash indicates missing data. Metadata for each sequence are available at GenBank/NCBI.
Table 5.
Taxa and GenBank sequence IDs for the phylogenetic analysis of sect. Chamaemelanium. A dash indicates missing data. Metadata for each sequence are available at GenBank/NCBI.
| Species | GPI | ITS | NRPD2a | trnL-trnF |
|---|---|---|---|---|
| Viola barroetana | - | AF097224, AF097270 | - | - |
| V. biflora | JF767023 | AY928309 | GU289574 | JF767165 |
| V. brevistipulata | JF767032 | AY928275 | GU289575 | JF767167 |
| V. canadensis | JF767034 | AF097231, AF097277 | GU289576 | JF767163 |
| V. delavayi | - | FJ002908 | - | - |
| V. fischeri | - | AY582168, AY541601 | - | - |
| V. flagelliformis | - | AF097233, AF097279 | - | - |
| V. lobata | JF767080 | - | - | JF767161 |
| V. orientalis | - | AY541602, AY582169 | - | DQ085929 |
| V. pubescens | JF767117 | DQ006044 | GU289580 | JF767162 |
| V. purpurea | JF767118 | MG235177 | KJ138061 | JF767160 |
| V. rotundifolia | JF767122 | AF097241, AF097287 | KJ138062 | JF767168 |
| V. schulzeana | - | FJ002907 | - | - |
| V. sheltonii | JF767130 | AF097226, AF097272 | KJ138070 | JF767159 |
| V. tomentosa | JN620193 | - | - | JN620205 |
| V. tripartita | OP256029 | OP256030 | - | - |
| V. uniflora | JF767146 | AY582167, AY541600 | KJ138083 | JF767166 |
| Outgroup: V. congesta | JF767046 | MH781265 (V. subandina) | GU289564 | JF767154 |
| Outgroup: V. capillaris | JF767035 | AF097220, AF097266 | KJ138036 | JF767156 |
Table 6.
Taxa, island biogeography, and GenBank sequence IDs of ITS1 and ITS2 for the combined dating and biogeographic analysis of subsect. Nosphinium. Biogeography for the outgroup was not coded and was entered as missing (“?”) in the analysis. Metadata for each sequence are available at GenBank/NCBI.
Table 6.
Taxa, island biogeography, and GenBank sequence IDs of ITS1 and ITS2 for the combined dating and biogeographic analysis of subsect. Nosphinium. Biogeography for the outgroup was not coded and was entered as missing (“?”) in the analysis. Metadata for each sequence are available at GenBank/NCBI.
| Species | Biogeography | GenBank Sequence IDs |
|---|---|---|
| Viola chamissoniana | Oahu | AF115955, AF115959 |
| V. helenae | Kauai | AF097260, AF097306 |
| V. kauaensis var. hosakae | Oahu | AF115957, AF115961 |
| V. kauaensis var. kauaensis | Kauai | AF097262, AF097308 |
| V. lanaiensis | Lanai | FJ895310, FJ895319 |
| V. lanaiensis | Maui | JN682058 |
| V. maviensis | Maui | AF097263, AF097309 |
| V. maviensis | Molokai | FJ895311, FJ895320 |
| V. maviensis | Maui | FJ895312, FJ895321 |
| V. maviensis | Hawaii | FJ895313, FJ895322 |
| V. oahuensis | Oahu | FJ895314, FJ895323 |
| V. robusta | Molokai | AF115956, AF115960 |
| V. robusta | Molokai | FJ895315, FJ895324 |
| V. tracheliifolia | Kauai | AF097261, AF097307 |
| V. tracheliifolia | Oahu | FJ895316, FJ895325 |
| V. tracheliifolia | Molokai | FJ895317, FJ895326 |
| V. waialenalenae | Kauai | AF115958, AF115962 |
| Outgroup: V. selkirkii | ? | AY928307 |
| Outgroup: V. spathulata | ? | HM851456 |
| Outgroup: V. langsdorffii | ? | AF097259, AF097305 |
| Outgroup: V. langsdorffii | ? | FJ895309, FJ895318 |
| Outgroup: V. mirabilis | ? | DQ358858, DQ358835 |
| Outgroup: V. nannei | ? | AF097255, AF097301 |
| Outgroup: V. odorata | ? | EU413918 |
| Outgroup: V. pedata | ? | AF097253, AF097299 |
| Outgroup: V. reichenbachiana | ? | DQ055382 |
Table 7.
Taxa and GenBank sequence IDs for the combined phylogenetic analysis of sect. Plagiostigma and sect. Viola (Figure 27 and Figure 33). A dash indicates missing data. “ibid.” is used for taxa with genomic data whose loci have the same GenBank ID (given under GPI-C). Taxon names have been edited to correspond to the nomenclature used herein. Metadata for each sequence are available at GenBank/NCBI.
Table 7.
Taxa and GenBank sequence IDs for the combined phylogenetic analysis of sect. Plagiostigma and sect. Viola (Figure 27 and Figure 33). A dash indicates missing data. “ibid.” is used for taxa with genomic data whose loci have the same GenBank ID (given under GPI-C). Taxon names have been edited to correspond to the nomenclature used herein. Metadata for each sequence are available at GenBank/NCBI.
| Species | GPI-C | GPI-M | ITS-C | ITS-M | NRPD2a-C | NRPD2a-M | SDH-C | SDH-M |
|---|---|---|---|---|---|---|---|---|
| Sect. Plagiostigma | ||||||||
| V. dissecta | SRX11632715 | ibid. | ibid. | - | ibid. | ibid. | ibid. | - |
| V. diffusa | JF767047 | JF767048 | GQ434456 | - | KJ138043 | KJ138044 | KJ138112 | KJ138113 |
| V. epipsila | JF767049 | JF767050 | MG237736 | - | GU289587 | GU289588 | KJ138115 | KJ138116 |
| V. hamiltoniana | JF767150 | JF767151 | AY928283 | - | GU289591 | GU289592 | - | KJ138153 |
| V. lanceolata | JF767069 | JF767070 | MG235616 | - | KJ138051 | KJ138052 | KJ138119 | - |
| V. minuscula | JF767089 | JF767090 | AF097236, AF097282 | - | - | - | - | - |
| V. occidentalis | JF767088 | JF767087 | - | - | OP256031 1 | OP256032 1 | OP256033 1 | - |
| V. principis | JF767115 | JF767116 | FJ002904 | - | KJ138059 | KJ138060 | KJ138128 | - |
| V. raddeana | SRX9916745 | ibid. | ibid. | - | ibid. | ibid. | ibid. | ibid. |
| V. renifolia | JF767120 | JF767121 | JN999695 | - | - | - | - | - |
| V. selkirkii | JF767128 | JF767129 | MG234698 | - | GU289590 | GU289589 | KJ138143 | KJ138144 |
| V. tuberifera | JF767142 | JF767143 | - | - | OP256034 1 | OP256035 1 | OP256036 1 | OP256037 1 |
| V. vaginata | JF767148 | JF767149 | - | - | OP256038 1 | OP256039 1 | OP256040 1 | OP256041 1 |
| Sect. Viola | ||||||||
| V. acuminata | SRX11632718 | ibid. | - | ibid. | ibid. | ibid. | ibid. | ibid. |
| V. chelmea | JF767036 | JF767037 | - | - | KU949390 | KU949396 | KU949402 | KU949407 |
| V. collina | JF767044 | JF767045 | - | EU413938 | KU949389 | KU949395 | KU949401 | KU949406 |
| V. grypoceras | SRX14846970 | ibid. | - | ibid. | ibid. | ibid. | ibid. | ibid. |
| V. hirta | JF767065 | JF767066 | - | DQ358856, DQ358833 | GU289581 | GU289582 | KJ138117 | KJ138118 |
| V. laricicola | JF767078 | JF767079 | - | - | KU949387 | KU949393 | KU949399 | KU949404 |
| V. mirabilis | JF767085 | JF767086 | - | MK828558 | GU289583 | GU289584 | KJ138120 | KJ138121 |
| V. sintenisii | - | - | - | DQ358859, DQ358836 | KU949391 | KU949397 | - | - |
| V. stagnina | JF767133 | JF767134 | - | KX166475 | - | KU949392 | KU949398 | KU949403 |
| V. striata | JF767135 | JF767136 | - | AF097247, AF097293 | KU949388 | KU949394 | KU949400 | KU949405 |
| V. uliginosa | JF767144 | JF767145 | - | KU949386 | GU289585 | GU289586 | KJ138151 | KJ138152 |
| Outgroups | ||||||||
| V. congesta | JF767046 | JF767046 | MH781265 | MH781265 | GU289564 | GU289564 | KJ138104 | KJ138104 |
| V. capillaris | JF767035 | JF767035 | AF097220, AF097266 | AF097220, AF097266 | KJ138036 | KJ138036 | KJ138135 | KJ138135 |
1 Sequences published herein. Metadata are available in [45].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marcussen, T.; Ballard, H.E.; Danihelka, J.; Flores, A.R.; Nicola, M.V.; Watson, J.M. A Revised Phylogenetic Classification for Viola (Violaceae). Plants 2022, 11, 2224. https://doi.org/10.3390/plants11172224
AMA Style
Marcussen T, Ballard HE, Danihelka J, Flores AR, Nicola MV, Watson JM. A Revised Phylogenetic Classification for Viola (Violaceae). Plants. 2022; 11(17):2224. https://doi.org/10.3390/plants11172224
Chicago/Turabian StyleMarcussen, Thomas, Harvey E. Ballard, Jiří Danihelka, Ana R. Flores, Marcela V. Nicola, and John M. Watson. 2022. "A Revised Phylogenetic Classification for Viola (Violaceae)" Plants 11, no. 17: 2224. https://doi.org/10.3390/plants11172224
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.