

The Disease Progression and Molecular Defense Response in Chenopodium Quinoa Infected with Peronospora Variabilis, the Causal Agent of Quinoa Downy Mildew
 ,
,
Abstract
:1. Introduction
2. Results
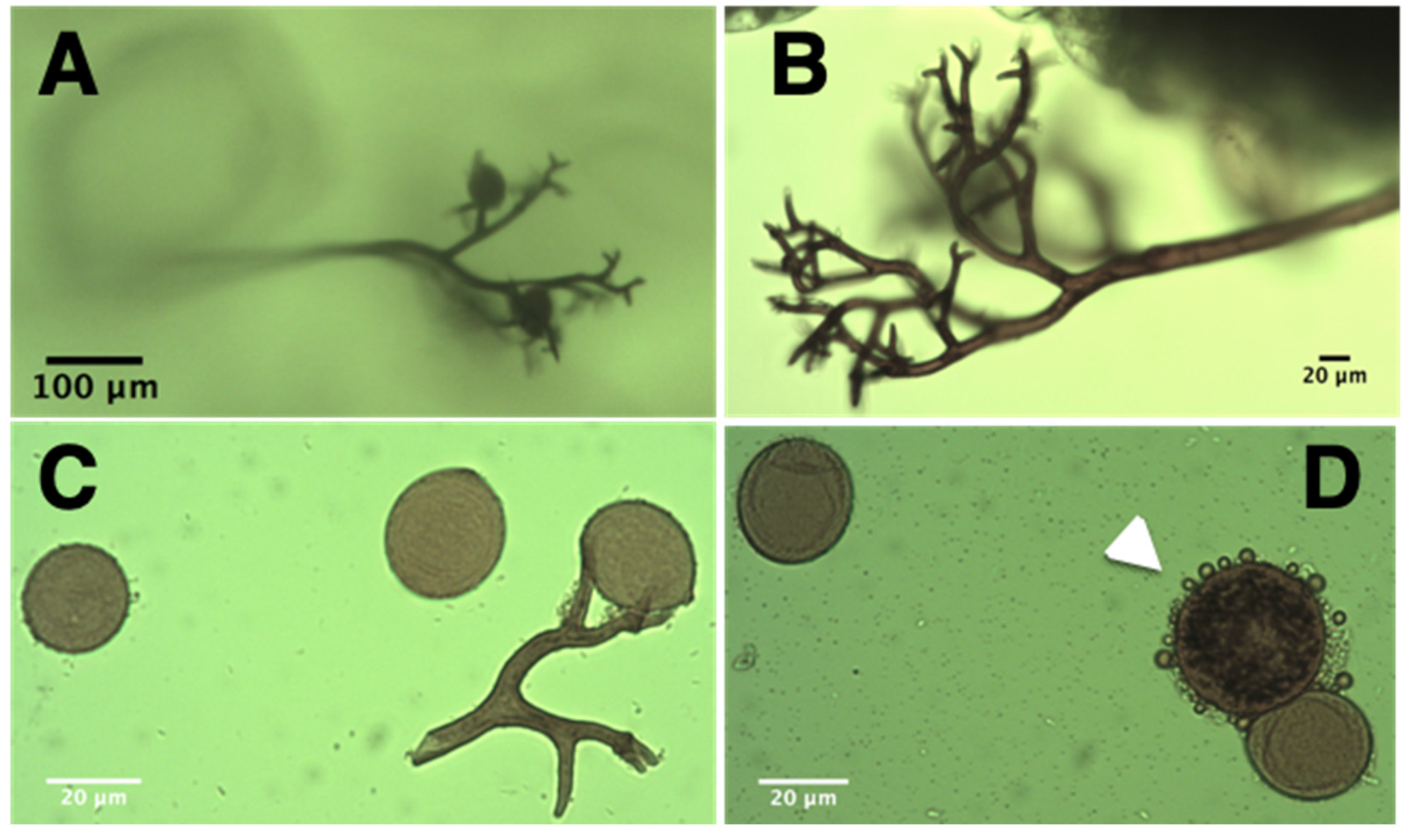
2.1. Isolation and Characterization of P. variabilis Isolate Kari
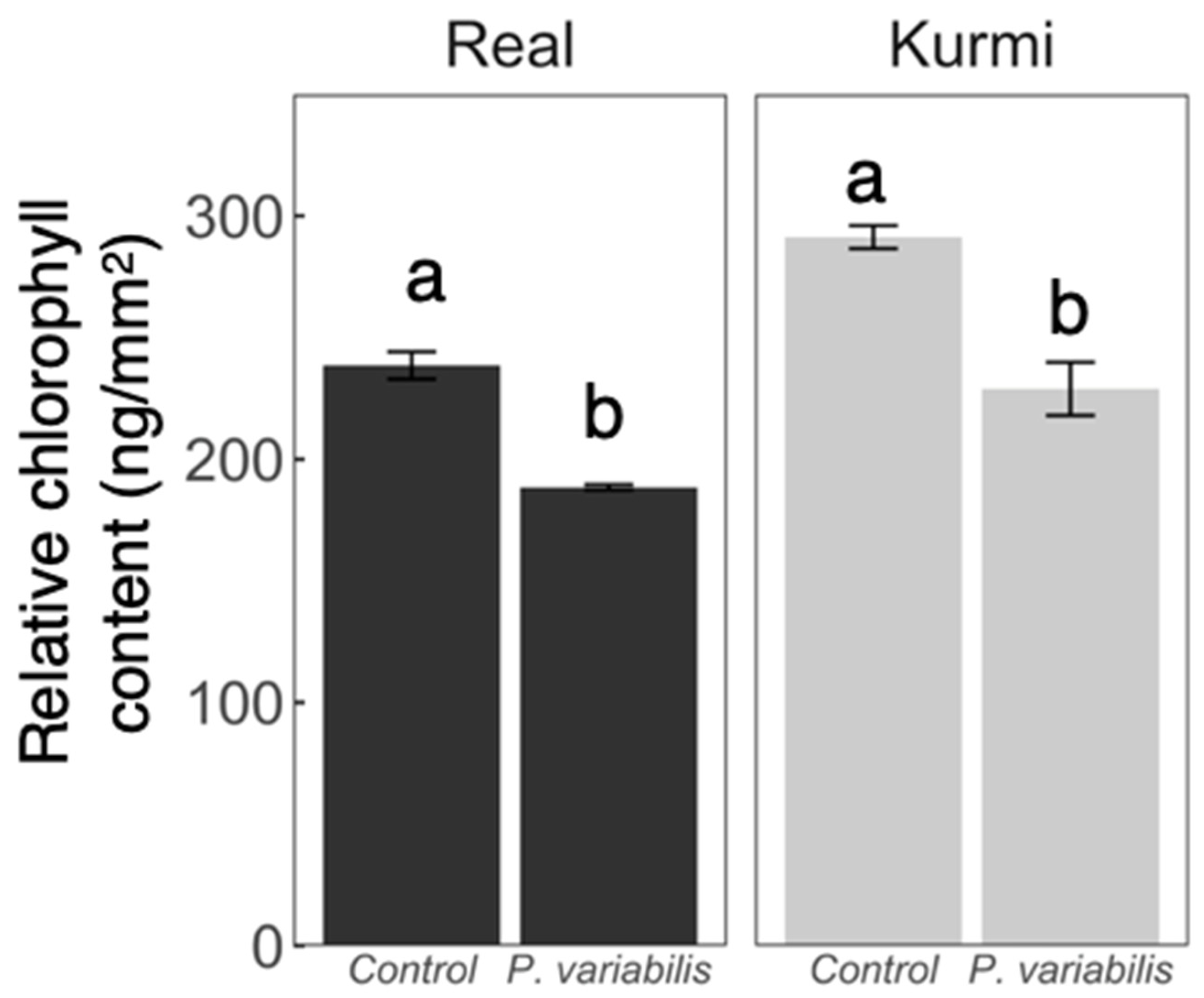
2.2. Downy Mildew Disease Progression in Two C. quinoa Cultivars
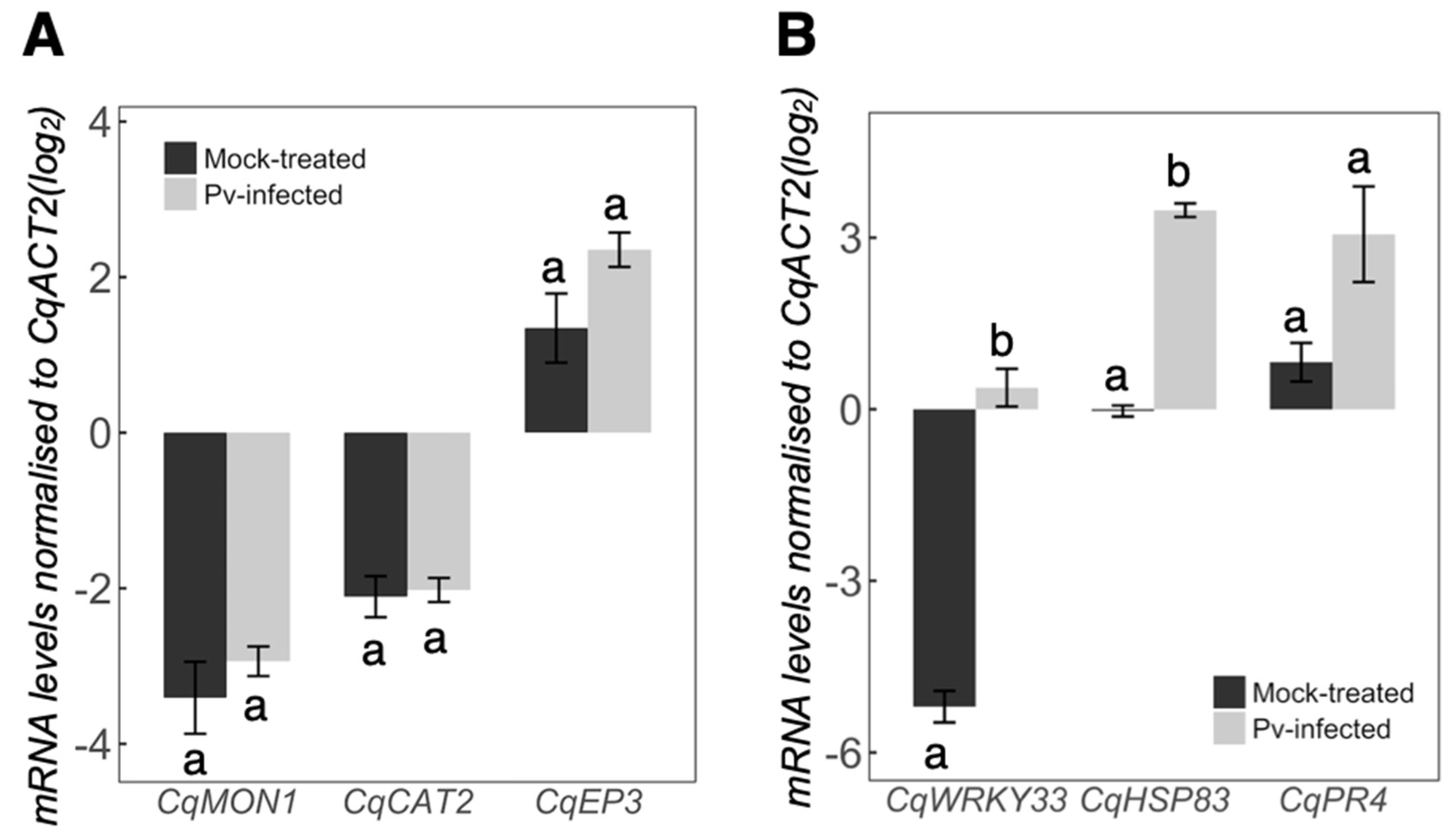
2.3. Quinoa Gene Expression Response against P. variabilis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Peronospora Variabilis Isolation
4.3. Peronospora Variabilis Maintenance
4.4. Microscopy of P. variabilis Structures
4.5. Molecular Identification of P. variabilis
4.6. Downy Mildew Disease Progression
4.7. RNA Isolation and cDNA Synthesis
4.8. Molecular Detection of P. variabilis PvCOX2
4.9. Gene Expression Analysis
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Bazile, D.; Jacobsen, S.E.; Verniau, A. The Global Expansion of Quinoa: Trends and Limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The Resistance of Quinoa (Chenopodium quinoa Willd.) to Adverse Abiotic Factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Oses, R.; Acuña-Rodríguez, I.S.; Antognoni, F.; Martinez-Mosqueira, E.A.; Coulibaly, A.; Canahua-Murillo, A.; Pinto, M.; Zurita-Silva, A.; et al. Quinoa biodiversity and sustainability for food security under climate change. A review. Agron. Sustain. Dev. 2014, 34, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Bazile, D.; Pulvento, C.; Verniau, A.; Al-Nusairi, M.S.; Ba, D.; Breidy, J.; Hassan, L.; Mohammed, M.I.; Mambetov, O.; Otambekova, M.; et al. Worldwide Evaluations of Quinoa: Preliminary Results from Post International Year of Quinoa FAO Projects in Nine Countries. Front. Plant Sci. 2016, 7, 850. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S.; Jacobsen, S.E.; Echegaray, J.; Ames, T. Impact of downy mildew on the yield of quinoa. CIP Program Rep. 2000, 2000, 397–401. [Google Scholar]
- Danielsen, S.; Munk, L. Evaluation of disease assessment methods in quinoa for their ability to predict yield loss caused by downy mildew. Crop. Protection 2004, 23, 219–228. [Google Scholar] [CrossRef]
- Benlhabib, O.; Boujartani, N.; Maughan, P.J.; Jacobsen, S.E.; Jellen, E.N. Elevated Genetic Diversity in an F2:6 Population of Quinoa (Chenopodium quinoa) Developed through an Inter-ecotype Cross. Front. Plant Sci. 2016, 7, 1222. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S.; Bonifacio, A.; Ames, T. Diseases of Quinoa (Chenopodium quinoa). Food Rev. Int. 2003, 19, 43–59. [Google Scholar] [CrossRef]
- Bazile, D.; Bertero, D.; Nieto, C. State of the Art of Quinoa in the World in 2013; FAO: Santiago, Chile; CIRAD: Montpellier, France, 2014; p. 724. [Google Scholar]
- Choi, Y.-J.; Danielsen, S.; Lübeck, M.; Hong, S.-B.; Delhey, R.; Shin, H.-D. Morphological and molecular characterization of the causal agent of downy mildew on quinoa (Chenopodium quinoa). Mycopathologia 2010, 169, 403–412. [Google Scholar] [CrossRef]
- Mhada, M.; Ezzahiri, B.; Benlhabib, O. Assessment of downy mildew resistance (Peronospora farinosa) in a quinoa (Chenopodium quinoa Willd.) germplasm. Int. J. Agri. Bio. Sci. Eng. 2014, 8, 77–80. [Google Scholar]
- Testen, A.L.; McKemy, J.M.; Backman, P.A. First Report of Quinoa Downy Mildew Caused by Peronospora variabilis in the United States. Plant Dis. 2012, 96, 146. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Choi, I.Y.; Kim, J.S.; Shin, H.D. First Report of Quinoa Downy Mildew Caused by Peronospora variabilis in Republic of Korea. Plant Dis. 2014, 98, 1003. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, S.; Jacobsen, S.E.; Hockenhull, J. First Report of Downy Mildew of Quinoa Caused by Peronospora farinosa f. sp. chenopodii in Denmark. Plant Dis. 2002, 86, 1175. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhargava, A.; Shukla, S.; Singh, H.B.; Ohri, D. Screening of exotic Chenopodium quinoa accessions for downy mildew resistance under mid-eastern conditions of India. Crop Prot. 2006, 25, 879–889. [Google Scholar] [CrossRef]
- Testen, A.L.; del Mar Jiménez-Gasco, M.; Ochoa, J.B.; Backman, P.A. Molecular Detection of Peronospora variabilis in Quinoa Seed and Phylogeny of the Quinoa Downy Mildew Pathogen in South America and the United States. Phytopathology 2014, 104, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielsen, S.; Mercado, V.H.; Ames, T.; Munk, T. Seed transmission of downy mildew (Peronospora farinosa f. sp. chenopodii) in quinoa and effect of relative humidity on seedling infection. Seed Sci. Technol. 2004, 32, 91–98. [Google Scholar] [CrossRef]
- Thines, M.; Choi, Y.-J. Evolution, Diversity, and Taxonomy of the Peronosporaceae, with Focus on the Genus Peronospora. Phytopathology 2016, 106, 6–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baiswar, P.; Chandra, S.; Kumar, R.; Ngachan, S.V. Peronospora variabilis on Chenopodium murale in India. Australas. Plant Dis. Notes 2010, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S.; Ames, T. Mildew (Peronospora farinosa) of Quinoa (Chenopodium Quinua Willd) in the Andean Region: Practical Manual for the Study of the Disease and the Pathogen; International Potato Center: Lima, Peru, 2000; p. 32. [Google Scholar]
- Julio, G.; Nayra, L.; Amalia, V.; Magne, J.; Ada, A.; Jaime, L.T.; Alejandro, B. Quinoa from Valley (Chenopodium quinoa Willd.): Valuable source of genetic resistance to powdery mildew (Peronospora farinosa Willd.). J. Selva Andin. Res. Soc. 2012, 3, 27–44. [Google Scholar]
- Calle-Sillo, L.; del Castillo, C.; Vargas, A.; Bonifacio, A. Asessment of commercial characteristics in red quinua (Chenopodium quinoa Willd.) in K’iphak’iphani, Province Ingavi—La Paz. Rev. De Investig. E Innovación Agropecu. Y De Recur. Nat. 2016, 3, 207–213. [Google Scholar]
- Ochoa, J.; Frinking, H.D.; Jacobs, T. Postulation of virulence groups and resistance factors in the quinoa/downy mildew pathosystem using material from Ecuador. Plant Pathol. 1999, 48, 425–430. [Google Scholar] [CrossRef]
- Kitz, L. Evaluation of Downy Mildew (Peronospora farinosa F. Sp. Chenonopoii) Resistance among Quinoa Genotypes and Investigation of P. farinosa Growth Using Scanning Electron Microscopy; Brigham Young University: Provo, UT, USA, 2008. [Google Scholar]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, C.; Chen, A.; Xiao, L.; Muller, H.M.; Ache, P.; Haberer, G.; Zhang, M.; Jia, W.; Deng, P.; Huang, R.; et al. A high-quality genome assembly of quinoa provides insights into the molecular basis of salt bladder-based salinity tolerance and the exceptional nutritional value. Cell Res. 2017, 27, 1327–1340. [Google Scholar] [CrossRef] [Green Version]
- Yasui, Y.; Hirakawa, H.; Oikawa, T.; Toyoshima, M.; Matsuzaki, C.; Ueno, M.; Mizuno, N.; Nagatoshi, Y.; Imamura, T.; Miyago, M. Draft genome sequence of an inbred line of Chenopodium quinoa, an allotetraploid crop with great environmental adaptability and outstanding nutritional properties. DNA Res. 2016, 23, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Derevnina, L.; Chin-Wo-Reyes, S.; Martin, F.; Wood, K.; Froenicke, L.; Spring, O.; Michelmore, R. Genome Sequence and Architecture of the Tobacco Downy Mildew Pathogen Peronospora tabacina. Mol. Plant Microbe Interact. 2015, 28, 1198–1215. [Google Scholar] [CrossRef] [Green Version]
- Baxter, L.; Tripathy, S.; Ishaque, N.; Boot, N.; Cabral, A.; Kemen, E.; Thines, M.; Ah-Fong, A.; Anderson, R.; Badejoko, W.; et al. Signatures of Adaptation to Obligate Biotrophy in the Hyaloperonospora arabidopsidis Genome. Science 2010, 330, 1549–1551. [Google Scholar] [CrossRef] [Green Version]
- Asai, S.; Rallapalli, G.; Piquerez, S.J.; Caillaud, M.C.; Furzer, O.J.; Ishaque, N.; Wirthmueller, L.; Fabro, G.; Shirasu, K.; Jones, J.D. Expression profiling during Arabidopsis/Downy mildew interaction reveals a highly-expressed effector that attenuates responses to salicylic acid. PLoS Pathog. 2014, 10, e1004443. [Google Scholar] [CrossRef]
- Wallström, S.V.; Aidemark, M.; Escobar, M.A.; Rasmusson, A.G. An alternatively spliced domain of the NDC1 NAD(P)H dehydrogenase gene strongly influences the expression of the ACTIN2 reference gene in Arabidopsis thaliana. Plant Sci. 2012, 183, 190–196. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.M.; Liu, W.C.; Lu, Y.T. CATALASE2 Coordinates SA-Mediated Repression of Both Auxin Accumulation and JA Biosynthesis in Plant Defenses. Cell Host Microbe 2017, 21, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Iwai, T.; Honkura, R.; Ugaki, M.; Ohshima, M.; Ohashi, Y. Four chitinase cDNAs from Chenopodium amaranticolor. Plant Biotechnol. 1997, 14, 85–86. [Google Scholar] [CrossRef]
- Rollano-Peñaloza, O.M.; Mollinedo, P.A.; Widell, S.; Rasmusson, A.G. Transcriptomic Analysis of Quinoa Reveals a Group of Germin-Like Proteins Induced by Trichoderma. Front. Fungal Biol. 2021, 2, 768648. [Google Scholar] [CrossRef]
- Waterfield, W.; Sisler, H. Effect of propiconazole on growth and sterol biosynthesis by Sclerotium rolfsii. Neth. J. Plant Pathol. 1989, 95, 187–195. [Google Scholar] [CrossRef]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol biosynthesis in oomycete pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Takeno, K. Stress-induced flowering: The third category of flowering response. J. Exp. Bot. 2016, 67, 4925–4934. [Google Scholar] [CrossRef] [Green Version]
- Korves, T.M.; Bergelson, J. A developmental response to pathogen infection in Arabidopsis. Plant Physiol. 2003, 133, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, W.; Thabet, M. Variation in downy mildew (Peronospora variabilis Gäum) resistance of some quinoa (Chenopodium quinoa Willd) cultivars under Egyptian conditions. Middle East J. Agric. Res. 2018, 7, 671–682. [Google Scholar]
- Pieterse, C.M.J.; Leon-Reyes, A.; van der Ent, S.; van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Holub, E.B. Natural history of Arabidopsis thaliana and oomycete symbioses. Eur. J. Plant Pathol. 2008, 122, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Mittelberger, C.; Yalcinkaya, H.; Pichler, C.; Gasser, J.; Scherzer, G.; Erhart, T.; Schumacher, S.; Holzner, B.; Janik, K.; Robatscher, P.; et al. Pathogen-Induced Leaf Chlorosis: Products of Chlorophyll Breakdown Found in Degreened Leaves of Phytoplasma-Infected Apple (Malus × domestica Borkh.) and Apricot (Prunus armeniaca L.) Trees Relate to the Pheophorbide a Oxygenase/Phyllobilin Pathway. J. Agric. Food Chem. 2017, 65, 2651–2660. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [Green Version]
- Canto, T.; Palukaitis, P. The Hypersensitive Response to Cucumber Mosaic Virus in Chenopodium amaranticolor Requires Virus Movement Outside the Initially Infected Cell. Virology 1999, 265, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Rich-Griffin, C.; Eichmann, R.; Reitz, M.U.; Hermann, S.; Woolley-Allen, K.; Brown, P.E.; Wiwatdirekkul, K.; Esteban, E.; Pasha, A.; Kogel, K.-H.; et al. Regulation of Cell Type-Specific Immunity Networks in Arabidopsis Roots. Plant Cell 2020, 32, 2742–2762. [Google Scholar] [CrossRef]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Diezel, C.; Somssich, I.E. Arabidopsis WRKY33 is a key transcriptional regulator of hormonal and metabolic responses toward Botrytis cinerea infection. Plant Physiol. 2012, 159, 266–285. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, A.; Casais, C.; Ichimura, K.; Shirasu, K. HSP90 interacts with RAR1 and SGT1 and is essential for RPS2-mediated disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 11777–11782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, J.-Y.; Ahn, G.; Kim, J.Y.; Kang, S.B.; Kim, M.R.; Su’udi, M.; Kim, W.-Y.; Son, D. Structural and functional differences of cytosolic 90-kDa heat-shock proteins (Hsp90s) in Arabidopsis thaliana. Plant Physiol. Biochem. 2013, 70, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [Green Version]
- Bertini, L.; Proietti, S.; Aleandri, M.P.; Mondello, F.; Sandini, S.; Caporale, C.; Caruso, C. Modular structure of HEL protein from Arabidopsis reveals new potential functions for PR-4 proteins. Biol. Chem. 2012, 393, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Huang, X.; Zhu, C.; Zhang, X.; Li, X.; Yang, S. Arabidopsis HSP90 protein modulates RPP4-mediated temperature-dependent cell death and defense responses. New Phytol. 2014, 202, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B.; Beynon, J.L.; Crute, I.R. Phenotypic and genotypic characterization of interactions between isolates of Peronospora parasitica and accessions of Arabidopsis thaliana. Mol. Plant Microbe Interact. 1994, 7, 223–239. [Google Scholar] [CrossRef]
- Rojas, W.; Pinto, M.; Alanoca, C.; Gomez Pando, L.; Leon-Lobos, P.; Alercia, A.; Bazile, D.; Bertero, H.D.; Nieto, C. Quinoa genetic resources and ex situ conservation. In State of the Art Report on Quinoa around the World in 2013; FAO & CIRAD; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; Chapter 1.5; pp. 56–82. [Google Scholar]
- Koroch, A.R.; Villani, T.S.; Pyne, R.M.; Simon, J.E. Rapid Staining Method to Detect and Identify Downy Mildew ( Peronospora belbahrii) in Basil. Appl. Plant Sci. 2013, 1, 1300032. [Google Scholar] [CrossRef] [PubMed]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H.a.a. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 1–9. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A Molecular Phylogeny of Phytophthora and Related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Liang, Y.; Urano, D.; Liao, K.L.; Hedrick, T.L.; Gao, Y.; Jones, A.M. A nondestructive method to estimate the chlorophyll content of Arabidopsis seedlings. Plant Methods 2017, 13, 26. [Google Scholar] [CrossRef]
- Hudspeth, D.S.; Stenger, D.; Hudspeth, M.E. A Cox2 phylogenetic hypothesis for the downy mildews and white rusts. Fungal Divers. 2003, 13, 47–57. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Marshall, O.J. PerlPrimer: Cross-platform, graphical primer design for standard, bisulphite and real-time PCR. Bioinformatics 2004, 20, 2471–2472. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, e1–e4. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Thaliana Gene Code | C. Quinoa Gene | Molecular Function | C. Quinoa Gene Code a | Primer Match b % | Primer Sequence | Product Size | PCR eff. c | |
|---|---|---|---|---|---|---|---|---|
| Fw | Rv | |||||||
| AT3G18780 AtACT2 | CqACT2A | Structural, Reference gene | AUR62014374 | 100 | 100 | ‘5-TACCACAGGTATCGTGCTTGACTC-3’ ‘5-GATCACGTCCGGCAAGATCC-3’ | 113 bp | 1.975 |
| CqACT2B | AUR62019116 | 92 | 100 | |||||
| CqACT2C | AUR62014579 | 54 | 33 | |||||
| CqACT2D | AUR62039382 | 50 | 29 | |||||
| AT2G28390 AtMON1 | CqMON1A | Vacuolar fusion protein, Reference gene | AUR62020295 | 100 | 100 | ‘5-AAGGATCATCTGACCATAAAGC-3’ | 145 bp | 2.057 |
| CqMON1B | AUR62037705 | 100 | 100 | ‘5-TCGTGTCAAGTTAGTTCGGG-3’ | ||||
| AT2G38470 AtWRKY33 | CqWRKY33A | Transcription factor | AUR62006298 | 100 | 100 | ‘5-TCCTTTACACCTGAGACATCCT-3’ | 126 bp | 1.953 |
| CqWRKY33B | AUR62026343 | 95 | 96 | ‘5-ACTGTTCTGTTACCATACCCTGAC-3’ | ||||
| AT5G52640 AtHSP90 | CqHSP83A | Molecular chaperone | AUR62031424 | 100 | 100 | ‘5-ATTCGGTGTTGGTTTCTACTC-3’ | 199 bp | 1.917 |
| CqHSP83B | AUR62021118 | 81 | 100 | ‘5-CCAAGTATTCCAACTGATCTTCC-3’ | ||||
| AT3G04720 AtPR4 | CqPR4A | Fungal growth inhibitor | AUR62004957 | 100 | 100 | ‘5-GGCAACGTACAATAACTATAACCC-3’ ‘5-TGCCATGTTTGTTACCCTGAG-3’ | 192 bp | 2.017 |
| CqPR4B | AUR62001001 | 100 | 71 | |||||
| CqPR4C | AUR62004958 | 100 | 48 | |||||
| AT3G54420 AtEP3 | CqEP3A | Chitinase | AUR62031317 | 100 | 100 | ‘5-CCTTCTTTGCTCATGTCACCC-3’ ‘5-CTGCTCCATAGTTGTAGTTCCA-3’ | 164 bp | 1.969 |
| CqEP3B | AUR62031318 | 100 | 100 | |||||
| CqEP3C | AUR62027403 | 91 | 100 | |||||
| CqEP3D | AUR62031322 | 91 | 100 | |||||
| AT4G35090 AtCAT2 | CqCAT2A | Catalase | AUR62040809 | 100 | 100 | ‘5-CCAGGAGTGAGATATAGATCATGGG-3’ | 145 bp | 1.995 |
| CqCAT2B | AUR62036848 | 100 | 91 | ‘5-CCCAAAGATTTATCCGCCTGAG-3’ | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rollano-Peñaloza, O.M.; Palma-Encinas, V.; Widell, S.; Mollinedo, P.; Rasmusson, A.G. The Disease Progression and Molecular Defense Response in Chenopodium Quinoa Infected with Peronospora Variabilis, the Causal Agent of Quinoa Downy Mildew. Plants 2022, 11, 2946. https://doi.org/10.3390/plants11212946
Rollano-Peñaloza OM, Palma-Encinas V, Widell S, Mollinedo P, Rasmusson AG. The Disease Progression and Molecular Defense Response in Chenopodium Quinoa Infected with Peronospora Variabilis, the Causal Agent of Quinoa Downy Mildew. Plants. 2022; 11(21):2946. https://doi.org/10.3390/plants11212946
Chicago/Turabian StyleRollano-Peñaloza, Oscar M., Valeria Palma-Encinas, Susanne Widell, Patricia Mollinedo, and Allan G. Rasmusson. 2022. "The Disease Progression and Molecular Defense Response in Chenopodium Quinoa Infected with Peronospora Variabilis, the Causal Agent of Quinoa Downy Mildew" Plants 11, no. 21: 2946. https://doi.org/10.3390/plants11212946