
When Morphology and Biogeography Approximate Nuclear ITS but Conflict with Plastid Phylogeny: Phylogeography of the Lotus dorycnium Species Complex (Leguminosae)

 ,
,  and
and
Abstract
:1. Introduction
- Lotus dorycnium ssp. herbaceus (Vill.) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Dorycnium herbaceum Vill. 1779, Prosp. Hist. Pl. Dauphine: 44; Synonym: Dorycnium intermedium Ledeb.) (Figure 1). Distribution range: East Mediterranean, Balkan Peninsula, extending westwards to Italy and southeastern France, northwards to Germany, Poland, Czech Republic, Slovakia, and Transcarpathian Ukraine, eastwards to the Northern part of Asia Minor, the Crimea, the Caucasus, and Transcaucasia, and to the Western part of the Caspian region [11,12,13]. Dorycnium intermedium has been described from the Crimea as a species close to D. herbaceum, but differing from the latter mainly in a patent (not appressed) pubescence on the calyx [14]. Rikli [9] believed that D. intermedium and D. herbaceum do not differ either morphologically or geographically, but Steinberg [15] considered D. intermedium the easternmost race of D. herbaceum.
- Lotus dorycnium ssp. gracilis (Jord.) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Dorycnium gracile Jord. 1846, Obs. Pl. Crit. 3: 70; Synonyms: Dorycnium herbaceum ssp. gracile (Jord.) Nyman; Dorycnium pentaphyllum ssp. gracile (Jord.) Rouy; Dorycnium jordanii Loret & Barrandon; Lotus jordanii (Loret & Barrandon) Coulot, Rabaute & J.-M. Tison) (Figure 2). Distribution range: West Mediterranean: France, Spain, Balearic Islands, Algeria [9,11,13].
- Lotus dorycnium ssp. germanicus (Gremli) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Dorycnium jordanii subsp. germanicum Gremli 1889, Excursionsfl. Schweiz, Ed. 6.: 496; Synonyms: Lotus germanicus (Gremli) Peruzzi; Dorycnium pentaphyllum ssp. germanicum (Gremli) Gams; Dorycnium germanicum (Gremli) Rouy) (Figure 3). Distribution range: the largest continuous part: much of the former Yugoslavia, Albania, Northern Greece, W part of Bulgaria and southwestern Romania; minor part: Eastern Alps, Eastern Switzerland, Bavaria, and the Pannonian region [12].
- Lotus dorycnium ssp. dorycnium (Synonyms: Dorycnium pentaphyllum ssp. pentaphyllum; Dorycnium pentaphyllum ssp. suffruticosum Bonnier & Layens; Dorycnium pentaphyllum ssp. transmontaum Franco) (Figure 4). Distribution range: Portugal and West Mediterranean (Spain, France, Italy, Algeria, Tunisia) [11,13].
- Lotus dorycnium ssp. anatolicus (Boiss. & Heldr.) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Dorycnium anatolicum Boiss. & Heldr. 1849, Diagn. Pl. Orient. ser. 1, 9: 31; Synonym: Dorycnium pentaphyllum ssp. anatolicum (Boiss. & Heldr.) Gams) (Figure 5A,B). Distribution range: Asiatic Turkey, Syria, Lebanon [11,13].
- Lotus dorycnium ssp. haussknechtii (Boiss.) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Dorycnium haussknechtii Boiss. 1872, Fl. Orient. 2: 163; Synonym: Dorycnium pentaphyllum ssp. haussknechtii (Boiss.) Gams) (Figure 5C–E). Distribution range: Asiatic Turkey, Syria, Lebanon, Bulgaria [11,13,16].
- Lotus dorycnium ssp. fulgurans (Porta) Kramina & D.D. Sokoloff, comb. nov. (Basionym: Anthyllis fulgurans Porta 1887, Nuovo Giorn. Bot. Ital. 19: 303; Synonyms: Dorycnium fulgurans (Porta) Lassen; Dorycnium pentaphyllum subsp. fulgurans (Porta) Cardona, Lorens & Sierra) (Figure 6). Restricted to Balearic Islands. According to morphological [17,18] and molecular data [7], it is a member of the L. dorycnium complex; we have therefore included it in the study.
2. Results
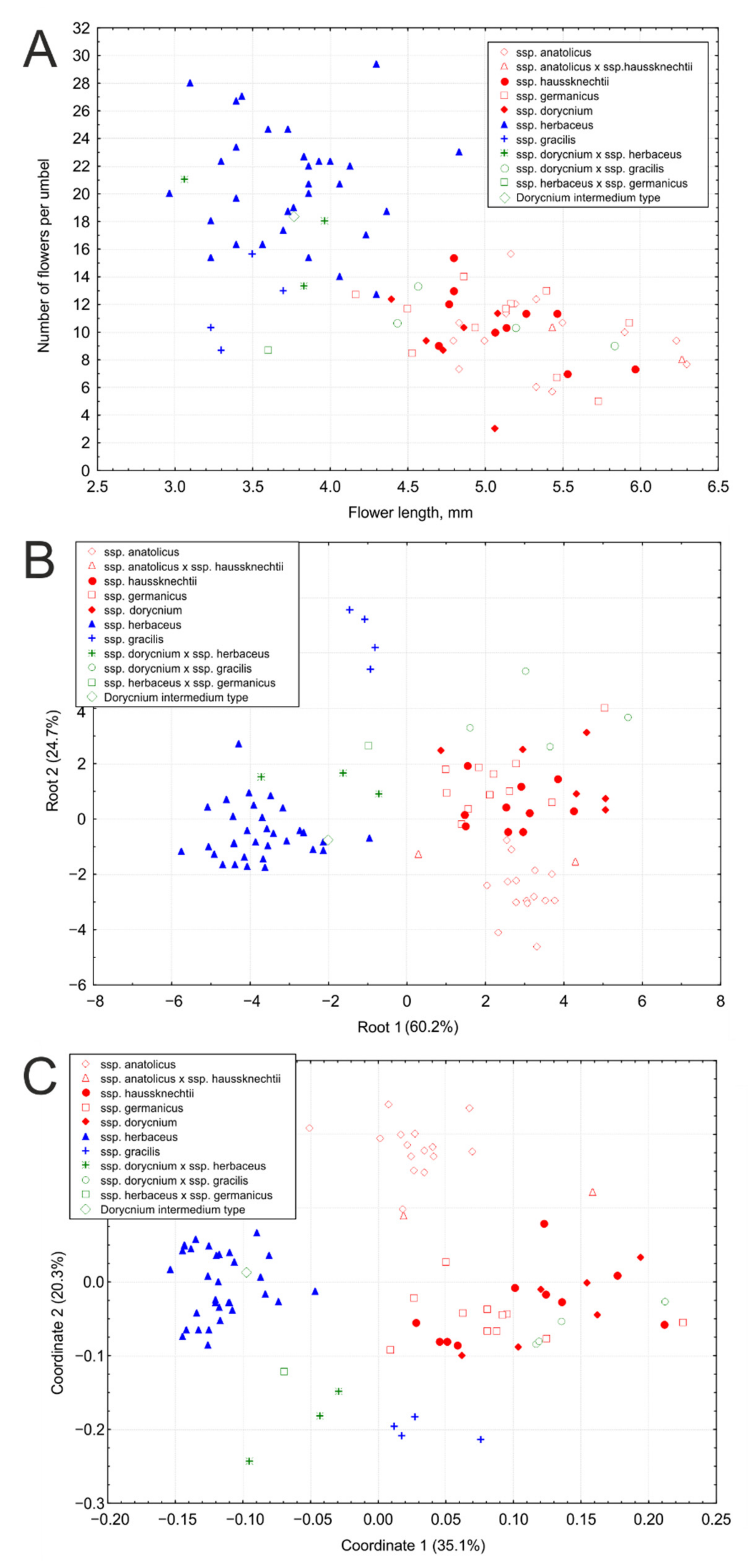
2.1. Morphometric Analyses
- Blue group: umbels with numerous (usually more than 13) small (usually shorter than 4.5 mm) flowers; this group includes ssp. herbaceus and ssp. gracilis.
- Red group: umbels with generally less numerous, but larger flowers; this group includes ssp. dorycnium, ssp. germanicus, ssp. anatolicus, and. ssp. haussknechtii.
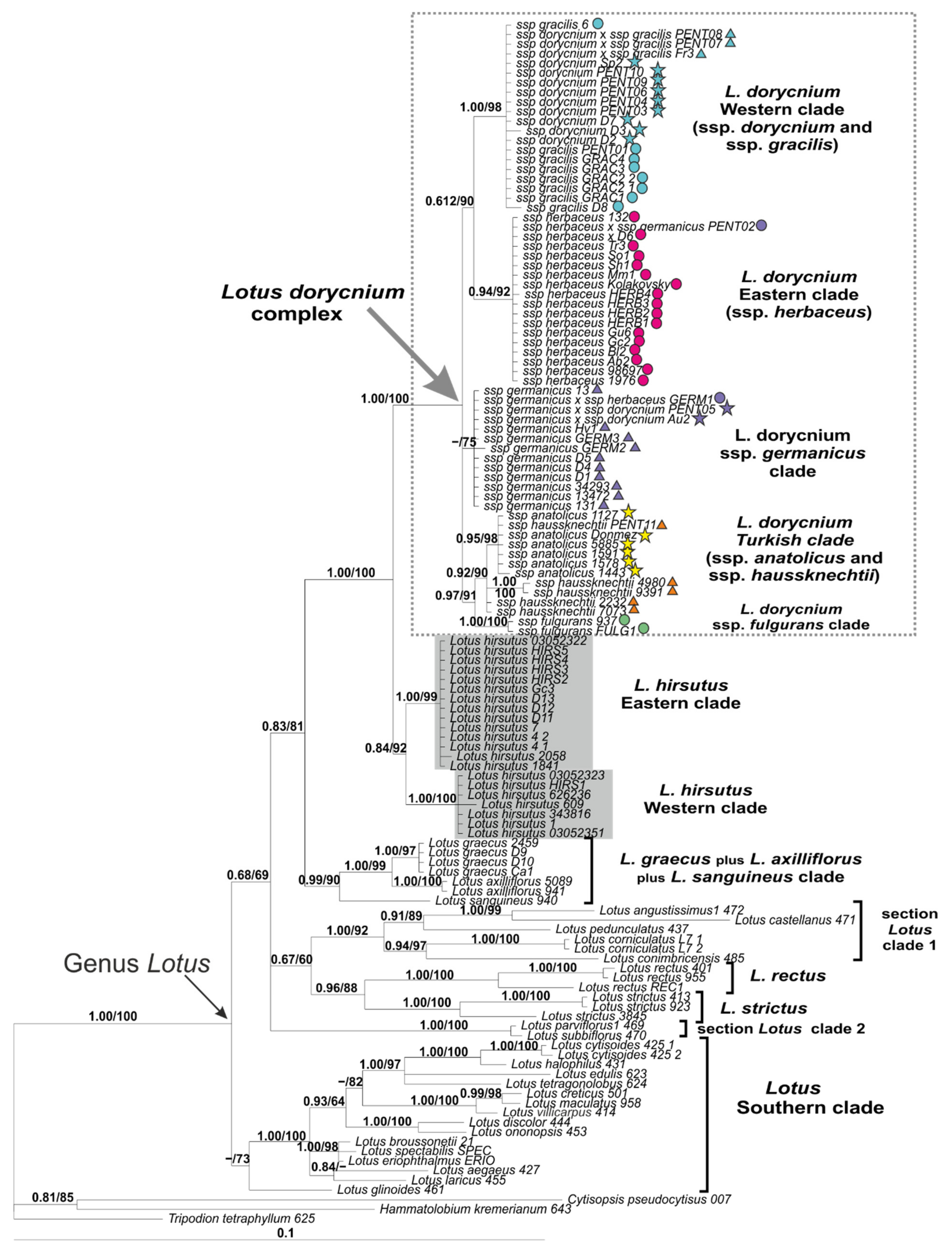
2.2. Phylogenetic Analysis of nrDNA ITS1-2 Dataset
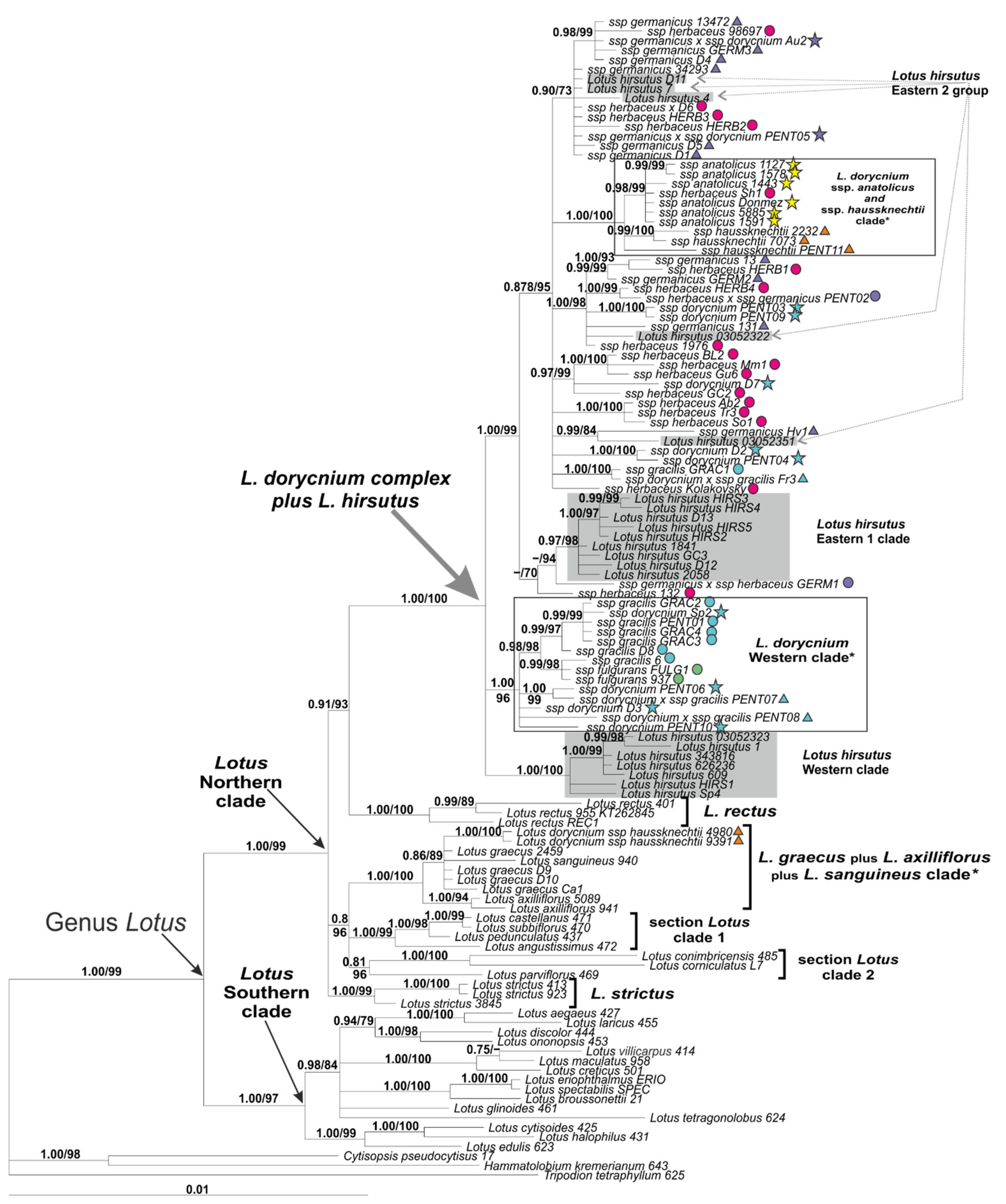
2.3. Phylogenetic Analysis of the Plastid DNA Dataset
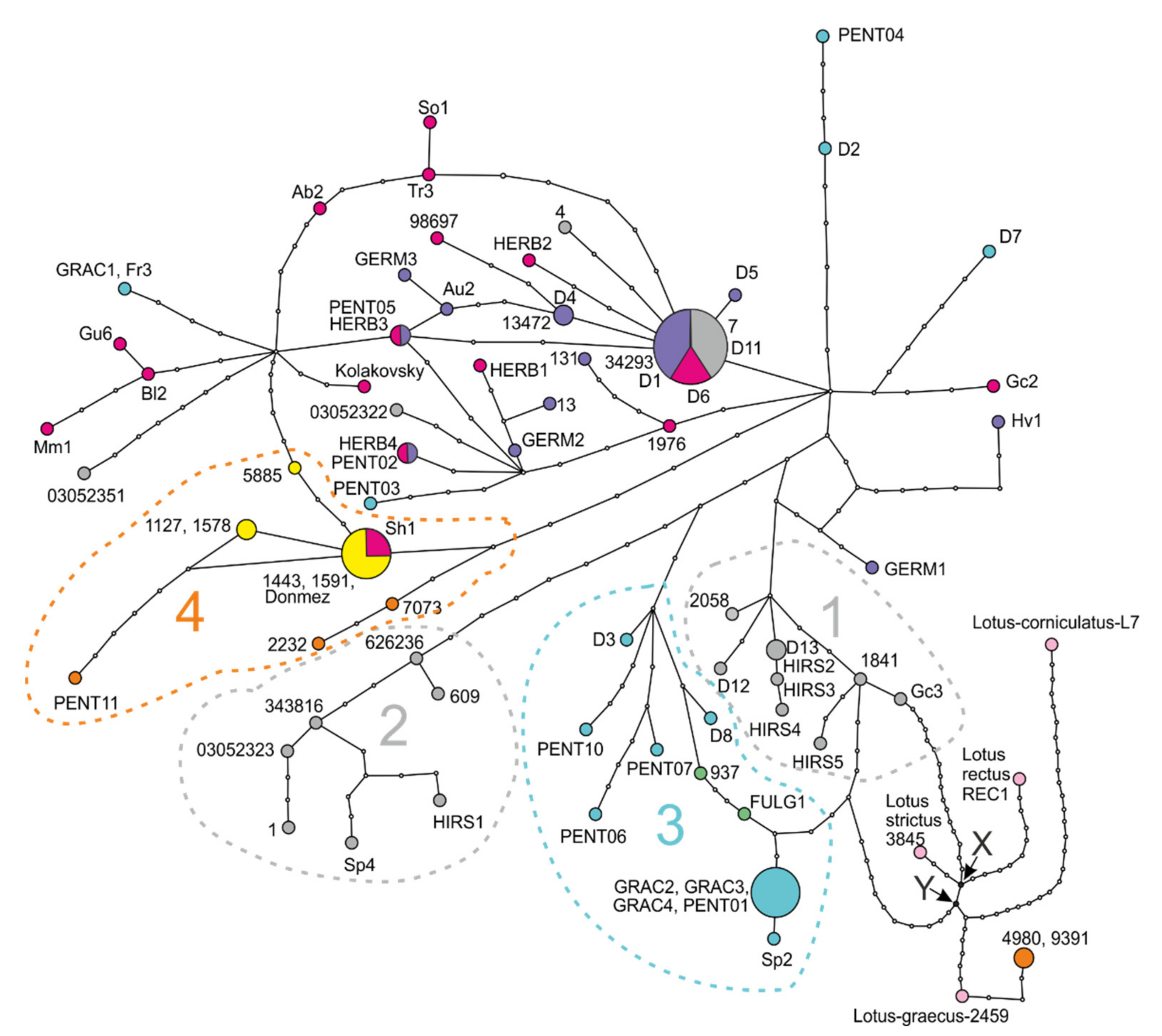
2.4. Haplotype Network Based on the Concatenated Plastid DNA Dataset
3. Discussion
3.1. Taxonomic Identification of Specimens
3.2. Phylogenetic Placement of Lotus dorycnium s.l. and Relationships among Its Subspecies
3.3. Phylogeography of the Lotus Dorycnium Complex
4. Materials and Methods
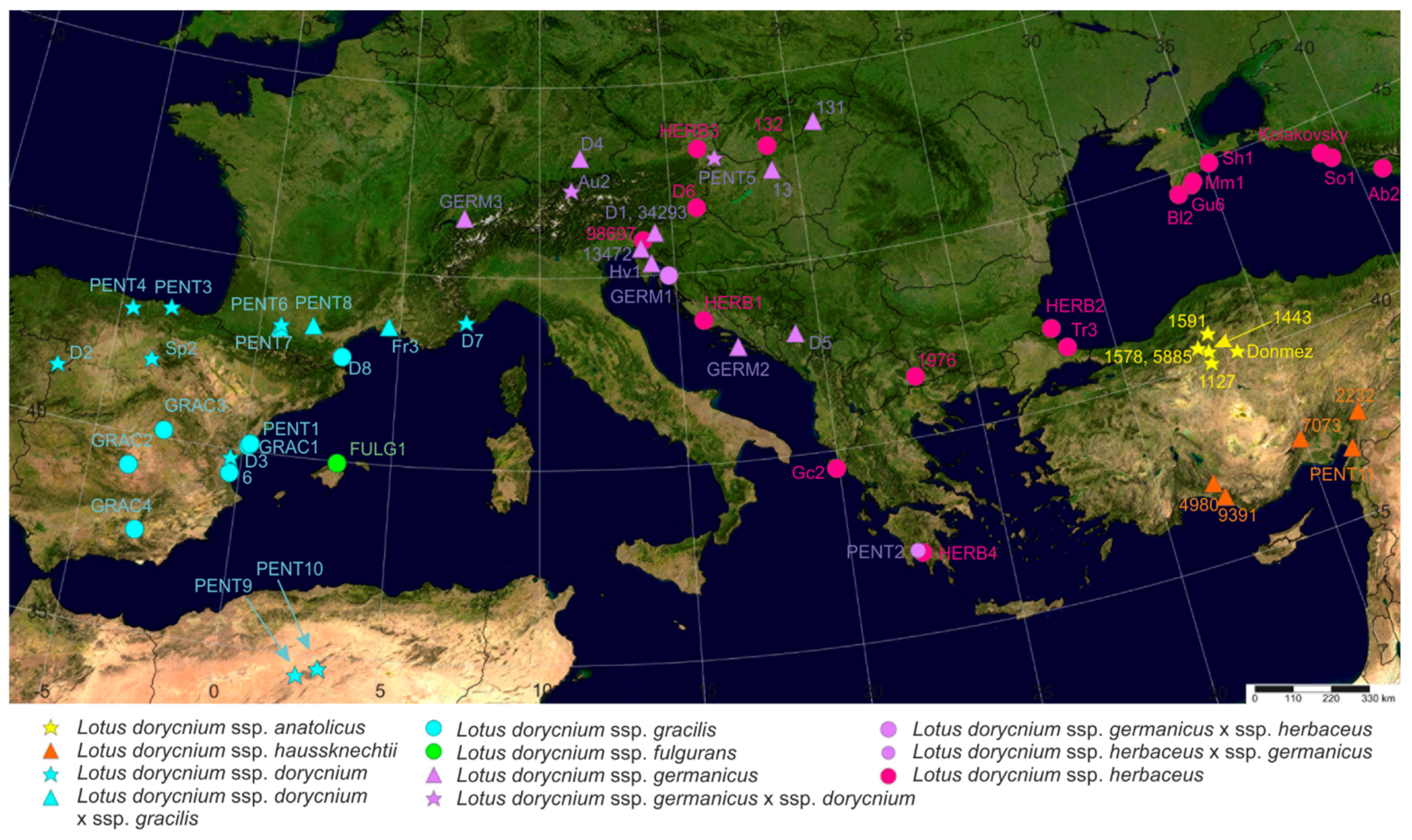
4.1. Plant Material
4.2. DNA Extraction, Amplification, and Sequencing
4.3. Phylogenetic Analyses
4.4. Morphometric Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Ball, P.W. Dorycnium Miller. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 172–173. [Google Scholar]
- Ball, P.W.; Chrtková-Žertová, A. Lotus L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 173–176. [Google Scholar]
- Reichenbach, L. Flora Germanica Excursoria; Carolum Cnobloch: Leipzig, Germany, 1832; Volume 2, pp. 505–507, 515–517. [Google Scholar]
- Gillett, J.B. Lotus in Africa south of the Sahara (excluding Cape Verde Islands and Socotra) and its distinction from Dorycnium. Kew Bull. 1958, 13, 361–381. [Google Scholar] [CrossRef]
- Lassen, P. Acmispon sect. Simpeteria, Acmispon roudairei, Dorycnium strictum, Lotus benoistii. Willdenovia 1986, 16, 107–112. [Google Scholar]
- Sokoloff, D.D. On taxonomy and phylogeny of the tribe Loteae DC. (Leguminosae). Byulleten’ Mosk. Obs. Ispyt. Prirody. Otd. Biol. 2003, 108, 35–48. (In Russian) [Google Scholar]
- Kramina, T.E.; Degtjareva, G.V.; Samigullin, T.H.; Valiejo-Roman, C.M.; Kirkbride, J.H., Jr.; Volis, S.; Deng, T.; Sokoloff, D.D. Phylogeny of Lotus (Leguminosae: Loteae): Partial incongruence between nrITS, nrETS and plastid markers and biogeographic implications. Taxon 2016, 65, 997–1018. [Google Scholar] [CrossRef]
- Kramina, T.; Lysova, M.; Samigullin, T.; Schanzer, I.; Özbek, M.; Sokoloff, D. Phylogenetic placement and phylogeography of large-flowered Lotus species (Leguminosae) formerly classified in Dorycnium: Evidence of pre-pleistocene differentiation of western and eastern intraspecific groups. Plants 2021, 10, 260. [Google Scholar] [CrossRef]
- Rikli, M. Die Gattung Dorycnium. Bot. Jahrb. 1901, 31, 314–404. [Google Scholar]
- Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Valiejo-Roman, C.M.; Antonov, A.S. Phylogeny of the genus Lotus (Leguminosae, Loteae): Evidence from nrITS sequences and morphology. Can. J. Bot. 2006, 84, 813–830. [Google Scholar] [CrossRef]
- Greuter, W.; Burdet, H.M.; Long, G. Med-Checklist; Dicotyledones (Lauraceae-Rhamnaceae); Conservatoire et Jardin botaniques de la Ville de Genève: Geneva, Switzerland, 1989; Volume 4, Available online: http://ww2.bgbm.org/mcl/ (accessed on 6 December 2021).
- Slavík, B. A plant-geographical study of the genus Dorycnium Mill. (Fabaceae) in the Czech Republic. Folia Geobot. Phytotaxon. 1995, 30, 291–314. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Home Page. 2021. Available online: https://www.gbif.org (accessed on 6 December 2021).
- von Ledebour, C.F. Index Seminum Horti Academici Dorpatensis; Russian Empire: Moscow, Russia, 1820; pp. 1–18. [Google Scholar]
- Steinberg, E.I. Dorycnium L. In Flora SSSR [Flora of the USSR]; Publishers of Academy of Sciences of USSR: Moscow, Russia, 1945; Volume 11, pp. 281–284. (In Russian) [Google Scholar]
- Marinov, Y.; Stoyanov, S. First record of Dorycnium haussknechtii (Fabaceae) for the Balkans. Phytol. Balc. 2019, 25, 63–68. [Google Scholar]
- Conesa, M.A.; Mus, M.; Rosselló, J.A. Lotus x minoricensis (Fabaceae), a new hybrid from the Balearic Islands. Flora Montiberica 2006, 30, 25–27. [Google Scholar]
- Conesa, M.A.; Mus, M.; Rosselló, J.A. Who threatens who? Natural hybridization between Lotus dorycnium and the island endemic Lotus fulgurans (Fabaceae). Biol. J. Linn. Soc. 2010, 101, 1–12. [Google Scholar] [CrossRef]
- Demiriz, H. Dorycnium Miller. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Cambridge University Press: Cambridge, UK, 1970; Volume 3, pp. 512–518. [Google Scholar]
- Díaz Lifante, Z. Dorycnium Mill. In Flora Iberica; Real Jardín Botaníco: Madrid, Spain, 2000; Volume 7, pp. 812–823. [Google Scholar]
- Lysova, M.V.; Meschersky, I.G.; Kramina, T.E. On the morphological variability and phylogenetic relationships of Crimean and Caucasian members of Lotus section Dorycnium. Turczaninowia 2019, 22, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Harpending, H.C.; Batzer, M.A.; Gurven, M.; Jorde, L.B.; Rogers, A.R.; Sherry, S.T. Genetic traces of ancient demography. Proc. Natl. Acad. Sci. USA 1998, 95, 1961–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, P. Stirpium in insulis Balearium anno 1885 collectarum enumeration. Nuovo G Bot. Ital. 1887, 19, 276–338. [Google Scholar]
- Lassen, P. Dorycnium fulgurans, a neglected species from the Balearic Islands. Bot. Not. 1979, 132, 357–358. [Google Scholar]
- Hewitt, G.M. Mediterranean peninsulas: The evolution of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–147. [Google Scholar] [CrossRef]
- Feliner, G.N. Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 265–278. [Google Scholar] [CrossRef]
- Degtjareva, G.V.; Valiejo-Roman, C.M.; Samigullin, T.H.; Guara-Requena, M.; Sokoloff, D.D. Phylogenetics of Anthyllis (Leguminosae: Papilionoideae: Loteae): Partial incongruence between nuclear and plastid markers, a long branch problem and implications for morphological evolution. Mol. Phylogenetics Evol. 2012, 62, 693–707. [Google Scholar] [CrossRef]
- Cardona, M.A.; Contandriopoulos, J.; Sierra Ràfols, E. Ètude biosystèmatique d’Anthyllis hystrix de Minorque et d’A. hermanniae de la Mèditerranèe orientale et central. Orsis 1986, 2, 5–25. [Google Scholar]
- Cardona, M.A. Contribution a l’etude cytotaxonomique de la flora des Baleares, I. Acta Phytotaxon. Barcinonensia 1973, 14, 1–20. [Google Scholar]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- Mort, M.E.; Archibald, J.K.; Randle, C.P.; Levsen, N.D.; O’leary, T.R.; Topalov, K.; Wiegand, C.M.; Crawford, D.J. Inferring phylogeny at low taxonomic levels: Utility of rapidly evolving cpDNA and nuclear ITS loci. Am. J. Bot. 2007, 94, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Jakob, S.S.; Blattner, F.R. A chloroplast genealogy of Hordeum (Poaceae): Long-term persisting haplotypes, incomplete lineage sorting, regional extinction, and the consequences for phylogenetic inference. Mol. Biol. Evol. 2006, 23, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Zozomová-Lihová, J.; Marhold, K.; Španiel, S. Taxonomy and evolutionary history of Alyssum montanum (Brassicaceae) and related taxa in Southwestern Europe and Morocco: Diversification driven by polyploidy, geographic and ecological isolation. Taxon 2014, 63, 562–591. [Google Scholar] [CrossRef] [Green Version]
- Kramina, T.E.; Meschersky, I.G.; Degtjareva, G.V.; Samigullin, T.H.; Belokon, Y.S.; Schanzer, I.A. Genetic variation in the Lotus corniculatus complex (Fabaceae) in northern Eurasia as inferred from nuclear microsatellites and plastid trnL-trnF sequences. Bot. J. Linn. Soc. 2018, 188, 87–116. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. 2010. Available online: http://www.simplemappr.net (accessed on 12 January 2021).
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Wen, J.; Zimmer, E. Phylogeny and biogeography of Panax L. (the ginseng genus, Araliaceae): Inferences from ITS sequences of nuclear ribosomal DNA. Mol. Phylogenetics Evol. 1996, 6, 167–177. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and diverse origins of the polyploid species. Syst. Bot. 2003, 28, 723–737. [Google Scholar]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Oxelman, B.; Lidén, M.; Berglund, D. Chloroplast rps16 intron phylogeny of the tribe Sileneae (Caryophyllaceae). Plant Syst. Evol. 1997, 206, 393–410. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, M.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Ali, R.H.; Bark, M.; Miró, J.; Muhammad, S.A.; Sjöstrand, J.; Zubair, S.M.; Abbas, R.M.; Arvestad, L. VMCMC: A graphical and statistical analysis tool for Markov chain Monte Carlo traces. BMC Bioinform. 2017, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- STATISTICA (Data Analysis Software System), Version 7.1; StatSoft Inc.: Tulsa, OK, USA, 2006.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 6 December 2021).
- Bateman, R.M.; Rudall, P.J.; Murphy, A.R.M.; Cowan, R.S.; Devey, D.S.; Peréz-Escobar, O.A. Whole plastomes are not enough: Phylogenomic and morphometric exploration at multiple demographic levels of the bee orchid clade Ophrys sect. Sphegodes. J. Exp. Bot. 2021, 72, 654–681. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | ssp. herbaceus | ssp. gracilis | ssp. fulgurans | ssp. dorycnium | ssp. germanicus | ssp. anatolicus | ssp. haussknechtii |
|---|---|---|---|---|---|---|---|
| Life history | Perennial herb, suffrutescent plant or small shrub | Perennial herb | Thorny shrub. Thorns are formed at the ends of the shoots | Perennial herb, suffrutescent plant or small shrub | |||
| Stem length | 20–65 cm | 30–80 cm | up to 100 cm | 10–50 cm | 10–50 cm | 20–35 cm | 30–60 cm |
| Pubescence | sparse with patent long and somewhat curved hairs | appressed, ±sericeous | appressed, sericeous | appressed | appressed | dense with subpatent long hairs | dense sericeous with appressed short hairs |
| Leaflet shape | oblong-obovate | linear-oblanceolate to linear | obovate-spathulate | linear-oblanceolate | oblong-obovate | oblong-obovate | oblong-obovate |
| Leaflet size | 4–20 × 2–6 mm | 10–20 × 2–4 mm | 3.5–7.5 × 1.2–2.3 mm | 6–12 × 2–3 mm | (8-)10–20 × 2–4 mm | 6–18 × 1–4 mm | 8–20 × 1.5–5 mm |
| Number of flowers per umbel | 12–30 | 12–20 | 1–4 | 5–15 | 4–15 | ||
| Peduncles | long | short, up to 0.5 mm | long | ||||
| Flower length | 3–5 mm | 3.2–5.5 mm | 4–6(-7) mm | ||||
| Pedicels | as long as or longer than calyx tube | usually longer than calyx tube | usually shorter than calyx tube | ||||
| Calyx teeth | 1/3–1/2 (2/3) length of tube | as long as tube | shorter than tube | 1/2–3/4 length of tube | |||
| Sample Code: VOUCHER information (Herbarium Code); Coordinates. Underlined Sample Codes Indicate That the Sample Was Included in the Morphometric Analysis. | ITS | trnL-F | rps16 | psbA-trnH |
|---|---|---|---|---|
| Lotus dorycnium ssp. anatolicus | ||||
| 1127: Turkey, Çubuk II Barajı, 28.VI.1982, F. Demircioğlu 1127 (GAZI); 40.0045 N, 32.9329 E | OL688389 * | OL697810 * | OL988837 * | OL753485 * |
| 1443: Turkey, A4 Ankara, Çubuk, Ovacık-Saraycık Köyleri Hallayik pinaiimuk, 03.VIII.1992, E. Dundar 1443 (GAZI); 40.3047 N, 32.9616 E | OL688390 * | OL697811 * | OL988838 * | OL753486 * |
| 1578: Turkey, A4 Ankara: Kızılcahamam, Soğuksu Milli Parki, Kaya Tepe civari, 10.VI.1990, O. Eyuboglu 1578 (GAZI); 40.4686 N, 32.6302 E | OL688391 * | OL697812 * | OL988839 * | OL753487 * |
| 1591: Turkey, A4 Çankırı, Atkaracalar, Dumanli Dagi, 09.VII.1992, Ahmet Duran 1591 (GAZI); 40.7589 N, 33.1222 E | OL688392 * | OL697813 * | OL988840 * | OL753488 * |
| 5885: Turkey, A4 Ankara: Kızılcahamam, 01.VIII.1991, M. Vural 5885 (GAZI); 40.4703 N, 32.6509 E | OL688393 * | OL697814 * | OL988841 * | OL753489 * |
| Donmez: Turkey, A4 Kırıkkale: Koҫubaba kasabasi, bağlar yöresi, bozkır, 16.VI.1990, A. A. Dönmez s.n. (GAZI); 40.0835 N, 33.8789 E | OL688394 * | OL697815 * | OL988842 * | OL753490 * |
| Lotus dorycnium ssp. dorycnium | ||||
| D2: Portugal, prov. Trás-os-Montos, Mogadouro, 25.V.1988, R. Auriault 14166 (H 1657556); 41.3384 N, 6.7202 W | KT250860 | MK751661 | KT262882 | KT262812 |
| D3: Spain, Valencia, Algar, 18.IV.1995, J. Riera, J. Güemes & E. Estrelles 17073 (H); 39.7808 N, 0.3679 W | KT250862 | MK751662 | KT262884 | KT262814 |
| D7: France, Alpes-Maritimes, Blausasc, 14.V.1977, A. Charpin & P. Hainard 9350 (H 1456976); 43.8050 N, 7.3634 E | KT250861 | MK751660 | KT262883 | KT262813 |
| PENT03: Spain, Cantabria, Brazomar, en Castro Urdiales, 02.VI.1996, E. Loriente s.n. (MA 658686); 43.3745 N, 3.2123 W | OL688395 * | OL697816 * | OL988854 * | OL753502 * |
| PENT04: Spain, Cantabria, Valle de Bedoya, Cillorigo, 31.VII.1986, E. Loriente s.n. (MA 658693); 43.1795 N, 4.5659 W | OL688396 * | OL697817 * | OL988855 * | OL753503 * |
| PENT06: France, MIDI-Pyrenees, Haute-Garonne, Puymarium, 11.IX.1992, F.I. van Nek 1042 (P 00851078); 43.3667 N, 0.7500 E | OL688397 * | OL697818 * | OL988856 * | OL753504 * |
| PENT09: Algeria, Saharan Atlas, near the village Zenina, road to Charet, 10.VII.1968, V.P. Bochantsev 758 (LE); 34.4585 N, 2.5300 E | OL688398 * | OL697819 * | OL988857 * | OL753505 * |
| PENT10: Algeria, Saharan Atlas, W of Djelfa, 22.I.1968, L.E. Rodin et al. 167 (LE); 34.6729 N, 3.1851 E | OL688399 * | OL697820 * | OL988858 * | OL753506 * |
| Sp2: Spain, Burgos, on the road from Lerma to Barriosuso, 18 km from Lerma, 07.VI.2018, T. Kramina & L. Koppel s.n. (MW); 41.9687 N, 3.5742 W | OL688400 * | OL697821 * | OL988859 * | OL753507 * |
| Lotus dorycnium ssp. dorycnium × L. d. ssp. gracilis | ||||
| Fr3: France, Arles, Camargue, bord de canal entre Gageron et Villeneuve, 06.X.1978, J. & A. Raynal 20915 (P 03031137); 43.5932 N, 4.5945 E | OL688401 * | OL697822 * | OL988860 * | OL753508 * |
| PENT07: France, MIDI-Pyrenees, Haute-Garonne, road between Peguikhan and Mondilhan, near Busquet-Bas, 01.VI.1993, F.I. van Nek 1622 (P 00851079); 43.3000 N, 0.7000 E | OL688402 * | OL697823 * | OL988861 * | OL753509 * |
| PENT08: France, Haute Garonne, Observation Point at St. Felix on N622 to Revel, 04.VII.1980, Verdcourt & Wilmot-Dear 5358 (P 03615698); 43.4483 N, 1.8905 E | OL688403 * | OL697824 * | OL988862 * | OL753510 * |
| Lotus dorycnium ssp. haussknechtii | ||||
| 2232: Turkey, C6 Kahramanmaraş: Engizeh Dağı, Aksu mahallesi sevresi, 1100 m, 05.VII.1986, H. Duman & F. Karaveliogulcar 2232 (GAZI); 37.5718 N, 36.9198 E | OL688420 * | OL697840 * | OL988849 * | OL753497 * |
| 4980: Turkey, C3 Antalya: Manavgat-Akseki 10 km, 28.VI.1993, H. Duman & F. Karavelioğulları 4980 (GAZI); 37.0465 N, 31.7903 E | OL688421 * | OL697841 * | OL988850 * | OL753498 * |
| 7073: Turkey, C5 Adana: Pozantı, 1570 m, 17.VII.1995, Z. Aytaç & V.N. Adıgüzel 7073 (GAZI); 37.4276 N, 34.8768 E | OL688422 * | OL697842 * | OL988851 * | OL753499 * |
| 9391: Turkey, C3 Antalya, Türkbaş Yaylası-Mahmut Seydi Köyü arası, maki, 10.VI.1993, Tuna Ekim 9391 (GAZI); 36.6344 N, 32.0242 E | OL688423 * | OL697843 * | OL988852 * | OL753500 * |
| PENT11: Syria, Mont Amanus, region d’Hasan, VII.1908, M. Haradjian s.n. (LE); 36.7508 N, 36.3330 E | OL688424 * | OL697844 * | OL988853 * | OL753501 * |
| Lotus dorycnium ssp. fulgurans | ||||
| 937: United Kingdom, Cultivated at Royal Botanic Gardens, Kew, 2010: origin Spain, Balearic Is. | KT250865 | MF314954 | KT262887 | KT262817 |
| FULG1: Spain, Cabo de Formentor, Baleares, Mallorca, 23.V.1977, P. Auquier, J. Duvigneaud 77 E 388 (P 03062315); 39.9502 N, 3.1955 E | OL688434 * | OL697856 * | OL988863 * | OL753512 * |
| Lotus dorycnium ssp. germanicus | ||||
| 13: Hungary, Budapest, Sashegy, steppefied meadow on the slope, 23.X.2005, I. Illarionova 13 (LE); 47.4820 N, 19.0189 E | OL688404 * | OL697825 * | OL988864 * | OL753513 * |
| 131: Slovakia austro-orientalis, distr. Roznava: in declivi cartiensi australi infra arcem Turnansky hrad prope vicum Turna n. Bodvou, c.350 m, 19.VII.1962, J. Ujcik 131 (LE); 48.6016 N, 20.8765 E | OL688405 * | OL697826 * | OL988865 * | OL753514 * |
| 13472: Italie: Collines au Nord de Rupingrande 350–400 m, 30.V.1976, M.T. Misset 13472 (LE); 45.7215 N, 13.7875 E | OL688406 * | OL697827 * | OL988866 * | OL753515 * |
| 34293: Slovenija, Polhograjsko hribovje: In pratis prope Govejek supra vicum Medvode. Solo dolom. 800 m, 19.VI.1973, D. Trpin & T. Wraber 9852/3 (LE); 46.1162 N, 14.3457 E | OL688407 * | OL697828 * | OL988867 * | OL753516 * |
| D1: Slovenia, Polhograjsko Hribovje, prope Govejek, supra vicum Medvode, 19.VI.1973, D. Trpin & T. Wraber 9852/3 (H 1081128); 46.1159 N, 14.3459 E | KT250868 | MK751666 | KT262889 | KT262819 |
| D4: Germany, Bayern, Oberbayerische Hochebene, n.München, 06.VII.1991, H. Kalheber 91-0625 (H 1662801); 48.1374 N, 11.5823 E | KT250869 | MK751667 | KT262890 | KT262820 |
| D5: Montenegro, 40 km NNE of Nikšic, Žabljak, P. Uotila 10652 (H 1033157); 43.1558 N, 19.1235 E | KT250870 | MK751668 | KT262891 | KT262821 |
| GERM2: Croatia, Kneža, 06.VI.1981, unknown s.n. (ZA); 42.9712 N, 17.0518 E | OL688408 * | OL697829 * | OL988868 * | OL753517 * |
| GERM3: Switzerland, Vaud, Réserve de Pupplinge (Borière) [Borière, Pas de la Borière, Alpes Friburgeoises, commune probable: Grandvillard (8 km E)], VII. 1981, M. Bournérias 240985 (P 03615699); 46.5362 N, 7.1481 E | OL688409 * | OL697830 * | OL988869 * | OL753518 * |
| Hv1: Croatia, Istarskaya Zhupanya, 16.V.2016, I. Schanzer, N. Stepanova, A. Fedorova s.n. (MW); 45.3222 N, 14.1517 E | OL688410 * | OL697831 * | OL988870 * | OL753519 * |
| Lotus dorycnium ssp. germanicus × L. d. ssp. dorycnium | ||||
| PENT05: Austria, Northern Burgenland, W-shore of Lake Neusiedl, 18.VI.2007, T. Barta s.n. (P 00851081); 47.9506 N, 16.8390 E | OL688412 * | OL697833 * | OL988872 * | OL753521 * |
| Au2: Austria, Tirol, Inntal: Zirler Berg NW Zirl, 820 m; Hange nape der Strasse, 05.VIII.1980, D. Podlech 34422 (P 03020864); 47.2848 N, 11.2249 E | OL688411 * | OL697832 * | OL988871 * | OL753520 * |
| Lotus dorycnium ssp. germanicus × L. d. ssp. herbaceus | ||||
| GERM1: Croatia, island Krk, in the port Baška nova, 10.VII.1981, B. Korica s.n. (ZA); 44.9709 N, 14.7628 E | OL688413 * | OL697834 * | OL988873 * | OL753522 * |
| Lotus dorycnium ssp. gracilis | ||||
| 6: Spain, Valecnia: El Saler south of Valencia, 14.VIII.1965, S.A. Renvoize 340 (LE); 39.3825 N, 0.3331 W | OL688414 * | OL697835 * | OL988843 * | OL753491 * |
| D8: France, dép. Pyrénées-Orientales, Canet, 02.VII.1981, J. Lambinon, R. Renard & L. Smeets 81/287 (H 1542915); 42.7041 N, 3.0223 E | KT250859 | MK751682 | KT262881 | KT262811 |
| GRAC1: Spain, Castellon, Cabanes, P.N. Prat de Cabanes-Torreblanca, 06.IX.2005, S. Fos 50/05 (MA 774818); 40.1374 N, 0.1651 E | OL688415 * | OL697836 * | OL988844 * | OL753492 * |
| GRAC2: Spain, Ciudad Real, Daimiel, Tablas de Daimiel, Isla de Algeciras, 21.VII.1992, S. Cirujano s.n. (MA 552216); 39.1628 N, 3.6818 W | OL688416 *OL688417 * | OL697837 * | OL988845 * | OL753493 * |
| GRAC3: Spain, Cuenca, Garcinarro, hacia Huete, pr. Cerros de Mudarra, 810 m, 10.VII.2004, V.J. Arán & M.J. Tohá 5930 (MA 732859); 40.2004 N, 2.7225 W | OL688418 * | OL697838 * | OL988846 * | OL753494 * |
| GRAC4: Spain, Granada, Villanueva de las Torres, 789 m, 08.VII.2008, A. Amor et al. 4/7 (MA 838410); 37.5577 N, 3.0888 W | OL688419 * | OL697839 * | OL988847 * | OL753495 * |
| PENT01: Spain, Castellón, Torreblanca (la Plana Alta), pr. Torrenostra, 30.VII.2012, V.J. Arán 8084 (MA 877350); 40.1946 N, 0.2245 E | MN545714 | MN553697 | OL988848 * | OL753496 * |
| Lotus dorycnium ssp. herbaceus | ||||
| 132: Slovakia meridionalis: in declivibus prope vicum Horné Turovce haud procul ab oppido Sahy, 24.VI.1958, J. Nitka 132 (LE); 48.1214 N, 18.9418 E | OL688425 * | OL697845 * | OL988874 * | OL753525 * |
| 1976: Bulgaria, m. Ograzden: in lapidosis herbosis ad pagum Nikudin, distr. Sandanski, 26.VII.1976, V. Vakov s.n. (LE); 41.5661 N, 23.0443 E | OL688426 * | OL697846 * | OL988875 * | OL753526 * |
| 98697: Slovenia, Primorsko: In graminosis fruticosis inter Lokavec prope Ajdovscina et Predmeja, cca 490–500 m, 14.VII.1980, M. Palma & D. Trpin 49/3 (LE); 45.9237 N, 13.8850 E | OL688427 * | OL697847 * | OL988876 * | OL753527 * |
| Ab2: Abkhazia, Sukhumi highway, roadside, 07.VI.2019, M. Lysova & S. Polevova Ab2 (MW); 43.0835 N, 40.8876 E | OL688428 * | OL697848 * | OL988877 * | OL753528 * |
| BL2: Crimea, Laspi, 02.VI.2015, C. Fomichev BL2 (MW); 44.4157 N, 33.7098 E | MN545735 | OL697849 * | OL988878 * | OL753529 * |
| D6: Austria, Steirisches Hügelland, Steiermark, Umgebung von Radkersburg, 7.VII.1976, H.Mayrhofer & H.Teppner s.n. (H 1216503); 46.6897 N, 15.9886 E | KT250882 | MK751681 | KT262898 | KT262828 |
| Gu6: Crimea, Gurzuf, 22.XI.2016, T.E. Kramina Gu6 (MW); 44.5452 N, 34.2647 E | MN545717 | OL697850 * | OL988879 * | OL753530 * |
| HERB1: Croatia, Šibenik, 15.VI.1997, M. Milović s.n. (ZA); 43.7376 N, 15.9094 E | OL688429 * | OL697851 * | OL988880 * | OL753531 * |
| HERB2: Bulgaria, Burgas prov., Bay Cilistar, 03.VII.2017, D. Lyskov s.n. (MW); 42.0232 N, 28.0045 E | OL688430 * | OL697852 * | OL988881 * | OL753532 * |
| HERB3: Austria, Wien, 14 Bezirk, Hohe Wand-Wiese bei Vordenhainbach, 280–360 m, 26.VII.2004, Thomas Barta 2004-317 (MA 759930); 48.2298 N, 16.2025 E | OL688431 * | OL697853 * | OL988882 * | OL753533 * |
| HERB4: Greece, Peloponnesus, Nom. Messinia, Ep. Kalamata, Taijetos Pass between Tripi and Artemisio, 1200–1350 m, 10.VI.1997, G. Kamari et al. s.n. (MA 871895); 37.0667 N, 22.2667 E | OL688432 * | OL697854 * | OL988883 * | OL753534 * |
| Kolakovsky: Russia, Krasnodar Krai, Tuapse, 14.VI.1957, A. Kolakovsky s.n. (LE); 44.0931 N, 39.0885 E | OL688433 * | OL697855 * | OL988884 * | OL753535 * |
| Mm1: Crimea, Malyy Mayak, 12.VI.2017, T.E. Kramina & O.V. Yurtseva Mm1 (MW); 44.6120 N, 34.3594 E | MN545721 | MN553698 | OL988885 * | OL753536 * |
| Sh1: Crimea, Shchebetovka, 15.VI.2017, T.E. Kramina & O.V. Yurtseva Sh1 (MW); 44.9338 N, 35.1458 E | MN545728 | MN553701 | OL988886 * | OL753537 * |
| So1: Krasodar krai, Sochi, between Volkonskaya and Soloniki, 05.VI.2017, M.V. Kuturova So1 (MW); 43.8736 N, 39.3772 E | MN545730 | MN553702 | OL988887 * | OL753538 * |
| Tr3: Turkey, Istanbul, 5 km N of Karacaköy, 26.V.2019, M. Lysova & T. Kramina Tr3 (MW); 41.4503 N, 28.3836 E | OL620157 * | OL624881 * | OL753482 * | OL753539 * |
| Lotus dorycnium ssp. herbaceus × L. d. ssp. germanicus | ||||
| Gc2: Greece, Kerkyra, Benitses, 25.VIII.2018, D.D. Sokoloff Gc2 (MW); 39.5378 N, 19.9117 E | OL620158 * | OL624882 * | OL753483 * | OL753541 * |
| PENT02: Greece, Peloponnesus, Nom. Messinia, Ep. Kalamata, 6–8 km NE of Ano Amfia, 14.VI.1995, G. Kamari et al. 2514 (MA 871946); 37.1389 N, 22.1250 E | MN545713 | MN553696 | OL988888 * | OL753540 * |
| Lotus dorycnium Complex | Lotus hirsutus | ||||
|---|---|---|---|---|---|
| Western Group | Eastern Group | Turkish Group * | Western Group | Eastern Group | |
| Number of sequences | 19 | 30 | 9 | 8 | 13 |
| Number of haplotypes | 14 | 25 | 6 | 8 | 10 |
| Number of sites | 1993 | 1999 | 1985 | 1990 | 1986 |
| Invariable sites | 1924 | 1887 | 1965 | 1944 | 1954 |
| Variable (polymorphic) sites: | 47 (2.36%) | 64 (3.2%) | 12 (0.6%) | 33 (1.66%) | 21 (1.06%) |
| Singleton variable sites | 16 | 42 | 7 | 27 | 11 |
| Parsimony informative sites | 31 (1.56%) | 22 (1.1%) | 5 (0.25%) | 6 (0.3%) | 10 (0.5%) |
| Haplotype diversity Hd | 0.953 | 0.986 | 0.889 | 1.000 | 0.949 |
| Nucleotide diversity Pi | 0.00571 | 0.00432 | 0.00191 | 0.00482 | 0.00305 |
| Mismatch distribution | Multimodal | Unimodal | Multimodal | Multimodal | Multimodal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kramina, T.E.; Lysova, M.V.; Samigullin, T.H.; Özbek, M.U.; Sokoloff, D.D. When Morphology and Biogeography Approximate Nuclear ITS but Conflict with Plastid Phylogeny: Phylogeography of the Lotus dorycnium Species Complex (Leguminosae). Plants 2022, 11, 410. https://doi.org/10.3390/plants11030410
Kramina TE, Lysova MV, Samigullin TH, Özbek MU, Sokoloff DD. When Morphology and Biogeography Approximate Nuclear ITS but Conflict with Plastid Phylogeny: Phylogeography of the Lotus dorycnium Species Complex (Leguminosae). Plants. 2022; 11(3):410. https://doi.org/10.3390/plants11030410
Chicago/Turabian StyleKramina, Tatiana E., Maya V. Lysova, Tahir H. Samigullin, Mehmet U. Özbek, and Dmitry D. Sokoloff. 2022. "When Morphology and Biogeography Approximate Nuclear ITS but Conflict with Plastid Phylogeny: Phylogeography of the Lotus dorycnium Species Complex (Leguminosae)" Plants 11, no. 3: 410. https://doi.org/10.3390/plants11030410