
First Report of Clonostachys rosea as a Mycoparasite on Sclerotinia sclerotiorum Causing Head Rot of Cabbage in India

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
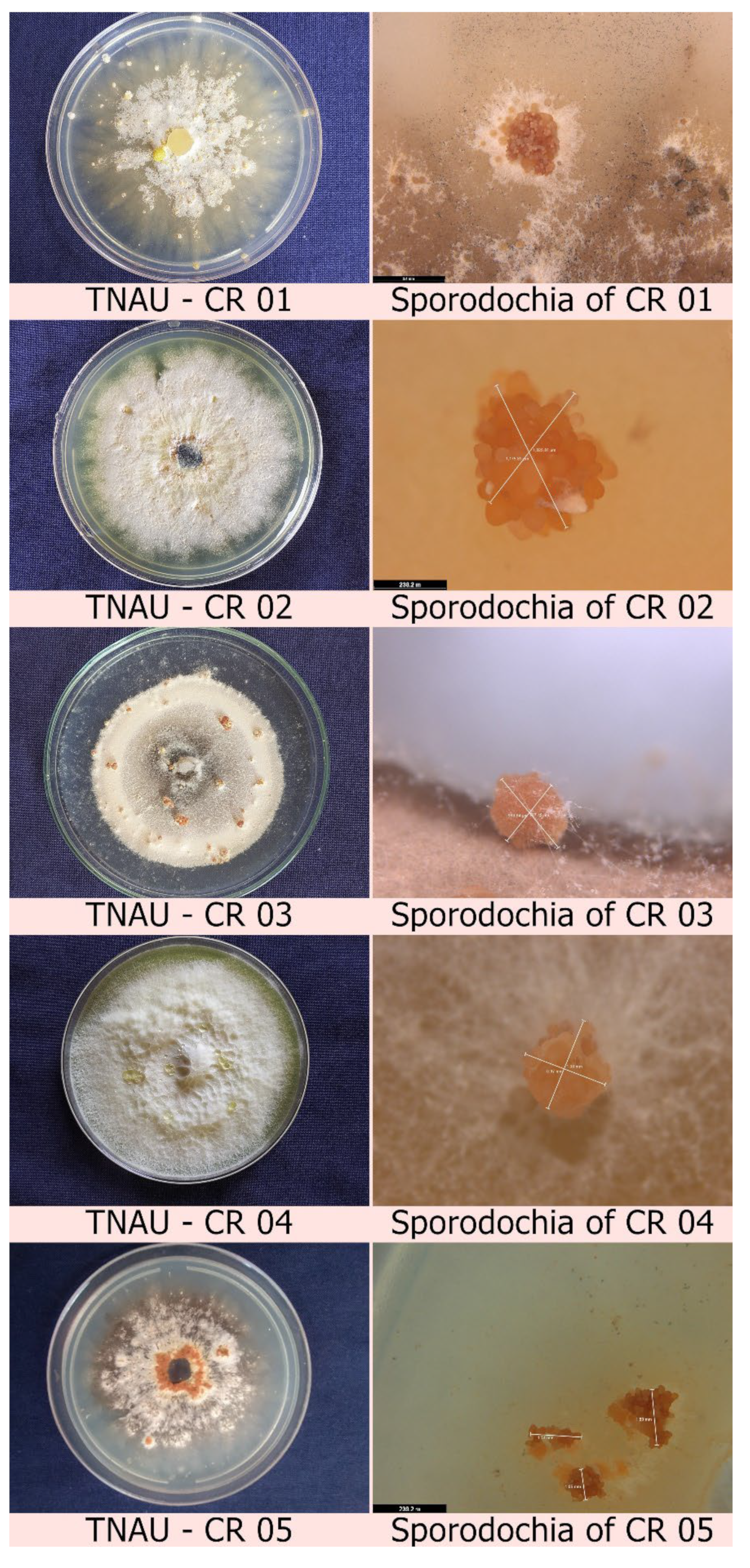
2.1. Isolation of Mycoparasitic Fungi from the Infected Sclerotia of Cabbage
2.2. Molecular Characterization of Clonostachys sp. Isolates
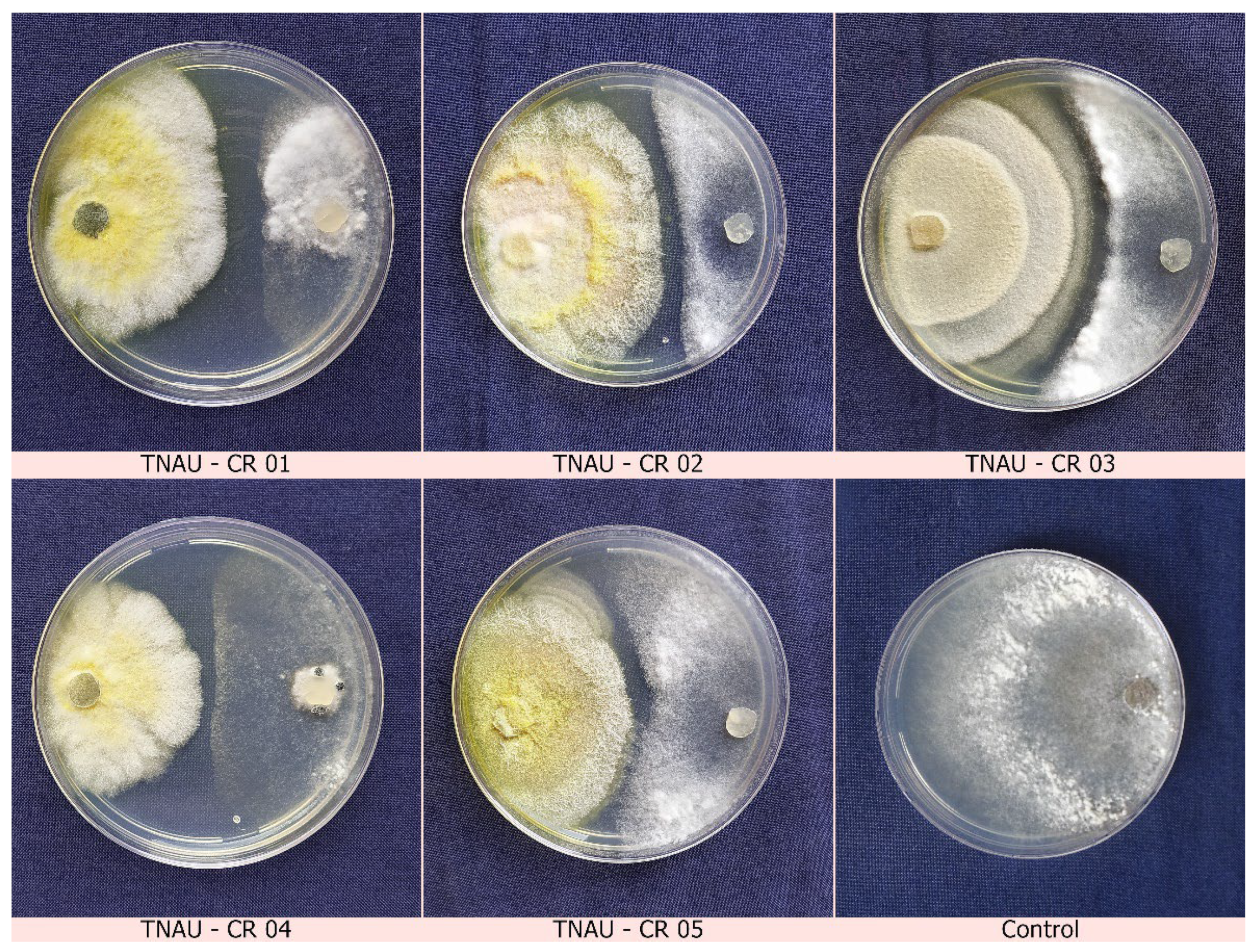
2.3. Dual Culture Assays
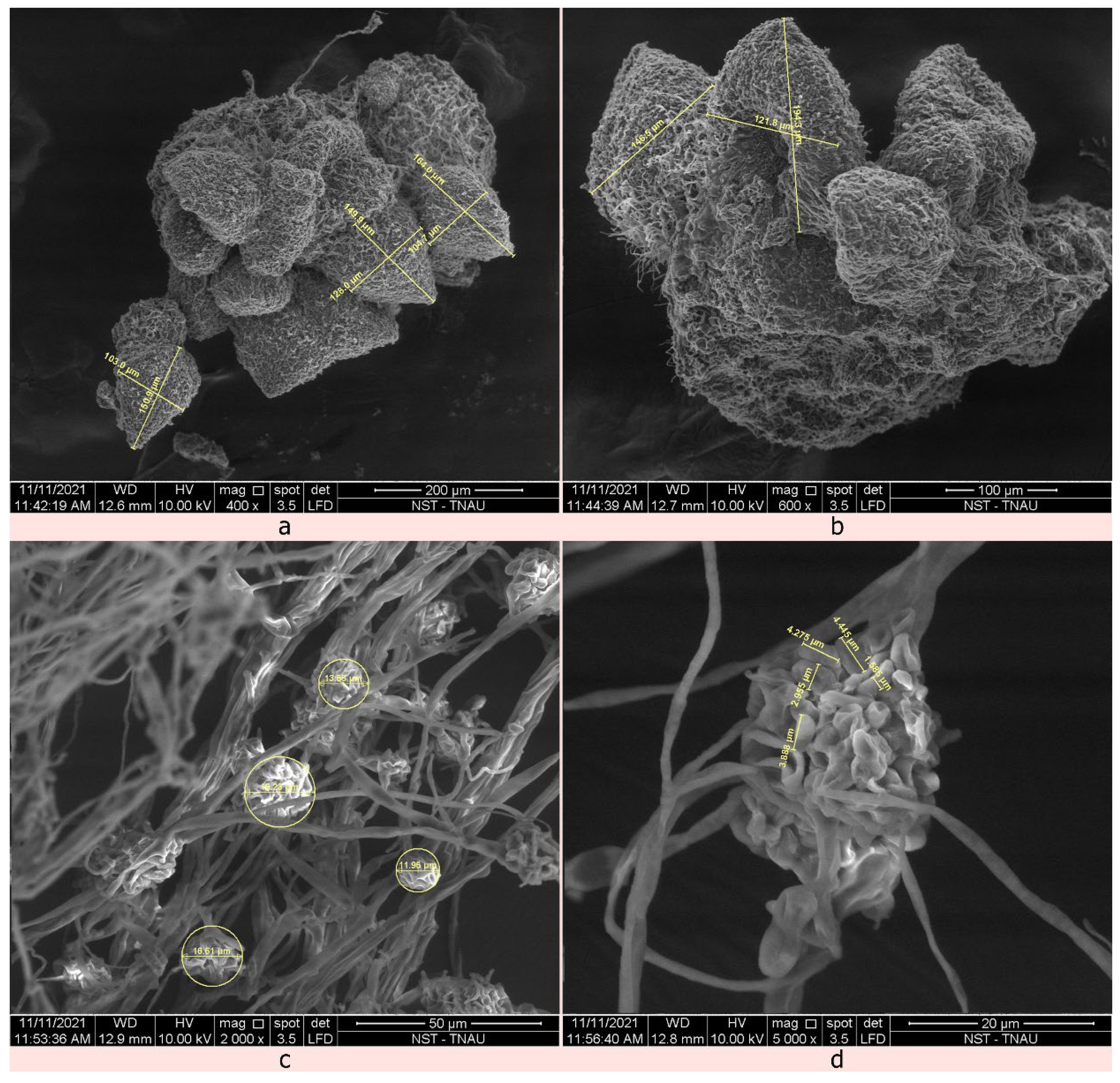
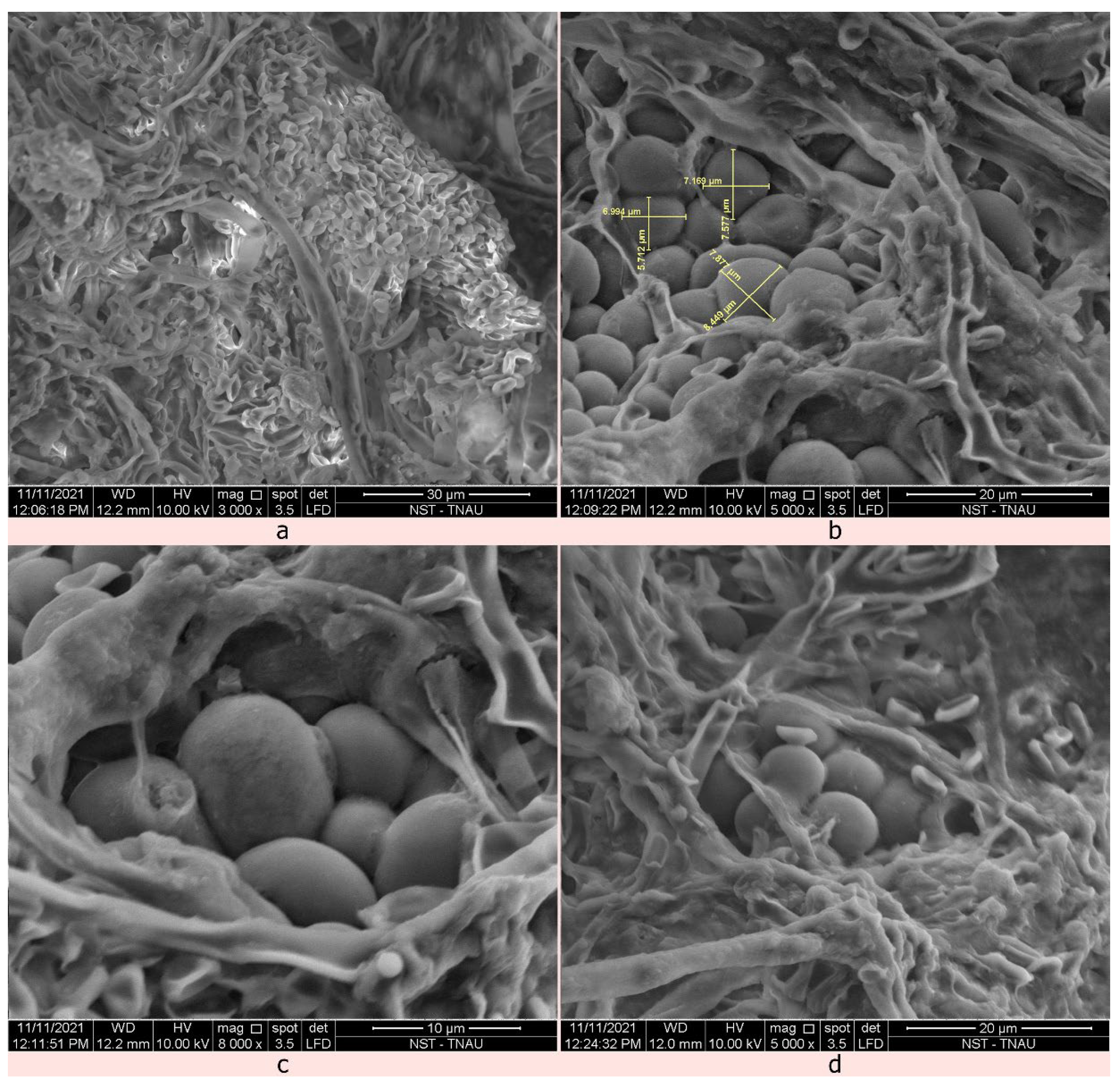
2.4. Identification of the Mycoparasitic Growth of C. rosea against S. sclerotiorum Using Scanning Electron Microscope
2.5. Metabolite Fingerprinting of the Volatile/Non-Volatile Bioactive Compounds Extracted from the Ethyl Acetate Fractions of C. rosea TNAU-CR 02
2.6. Effects of the Crude Antibiotics of C. rosea TNAU-CR 02 against S. sclerotiorum
2.7. The Effect of C. rosea TNAU-CR 02 Liquid Bioformulation for the Management of Cabbage Head Rot Pathogens under Greenhouse Conditions
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Fungal Isolation
4.2. Morphological and Molecular Characterization of Mycoparasitic Fungal Isolates from Infected Sclerotia
4.3. Sequencing and Phylogenetic Analysis
4.4. Dual Culture Assays
4.5. Preparation of Conidiospore Suspension of Effective Mycoparasitic Fungal Isolates
4.6. Scanning Electron Microscopy Analysis
4.7. GC-MS Analysis of the Volatile Organic Compounds and Non-Volatile Organic Compounds Extracted from the Ethyl Acetate Fractions of C. rosea TNAU-CR 02
4.8. Bioassays of the Antifungal Activities of C. rosea against S. sclerotiorum
4.9. Development of a Liquid Formulation of the C. rosea TNAU-CR 02 Isolate
4.10. Fungal and Bacterial Isolates
4.11. Pathogen Mass Multiplication
4.12. The Efficacy of the Biocontrol Agents on the Severity of Cabbage Head Rot Pathogens under Greenhouse Conditions
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Purdy, L.H. Sclerotinia sclerotiorum: History, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology 1979, 69, 875–880. [Google Scholar] [CrossRef]
- Willetts, H.; Wong, J. The biology of Sclerotinia sclerotiorum, S. trifoliorum, and S. minor with emphasis on specific nomenclature. Bot. Rev. 1980, 46, 101–165. [Google Scholar] [CrossRef]
- Pressete, C.G.; Giannini, L.S.V.; de Paula, D.A.C.; do Carmo, M.A.V.; Assis, D.M.; Santos, M.F.C.; Machado, J.d.C.; Marques, M.J.; Soares, M.G.; Azevedo, L. Sclerotinia Sclerotiorum (White Mold): Cytotoxic, Mutagenic, and Antimalarial Effects In Vivo and In Vitro. J. Food Sci. 2019, 84, 3866–3875. [Google Scholar] [CrossRef]
- Wu, B.; Subbarao, K.; Liu, Y.-B. Comparative survival of sclerotia of Sclerotinia minor and S. sclerotiorum. Phytopathology 2008, 98, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Blad, B.; Steadman, J.; Weiss, A. Canopy structure and irrigation influence white mold disease and microclimate of dry edible beans. Phytopathology 1978, 68, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- Rotem, J.; Palti, J. Irrigation and plant diseases. Annu. Rev. Phytopathol. 1969, 7, 267–288. [Google Scholar] [CrossRef]
- Sankar, P.; Jeyarajan, R. Biological control of sesamum root rot by seed treatment with Trichoderma spp. and Bacillus subtilis. Indian J. Mycol. Plant Pathol. 1996, 26, 217–220. [Google Scholar]
- Barrière, V.; Lecompte, F.; Nicot, P.C.; Maisonneuve, B.; Tchamitchian, M.; Lescourret, F. Lettuce cropping with less pesticides. A review. Agron. Sustain. Dev. 2014, 34, 175–198. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Mousa, K.M.; ELnahas, M.O.; Daba, G.M. Fungi against insects and contrariwise as biological control models. Egypt. J. Biol. Pest Control 2021, 31, 13. [Google Scholar] [CrossRef]
- Schroers, H.-J.; Samuels, G.J.; Seifert, K.A.; Gams, W. Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 1999, 91, 365–385. [Google Scholar] [CrossRef]
- Schroers, H.-J. A monograph of bionectria ascomycota, hypocreales, bionectriaceae and its Clonostachys anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Zhai, M.-M.; Qi, F.-M.; Li, J.; Jiang, C.-X.; Hou, Y.; Shi, Y.-P.; Di, D.-L.; Zhang, J.-W.; Wu, Q.-X. Isolation of secondary metabolites from the soil-derived fungus Clonostachys rosea YRS-06, a biological control agent, and evaluation of antibacterial activity. J. Agric. Food Chem. 2016, 64, 2298–2306. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, R.; Mendoza-de-Gives, P.; Aguilar-Marcelino, L.; López-Arellano, M.E.; Gamboa-Angulo, M.; Hanako Rosas-Saito, G.; Reyes-Estébanez, M.; Guadalupe García-Rubio, V. In vitro lethal activity of the nematophagous fungus Clonostachys rosea (Ascomycota: Hypocreales) against nematodes of five different taxa. BioMed Res. Int. 2018, 2018, 3501827. [Google Scholar] [CrossRef] [Green Version]
- Keyser, C.A.; Jensen, B.; Meyling, N.V. Dual effects of Metarhizium spp. and Clonostachys rosea against an insect and a seed-borne pathogen in wheat. Pest Manag. Sci. 2016, 72, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Rybczyńska-Tkaczyk, K.; Korniłłowicz-Kowalska, T. Activities of Versatile Peroxidase in Cultures of Clonostachys rosea f. catenulata and Clonostachys rosea f. rosea during Biotransformation of Alkali Lignin. J. AOAC Int. 2018, 101, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Sutton, J.C.; Li, D.-W.; Peng, G.; Yu, H.; Zhang, P.; Valdebenito-Sanhueza, R. Gliocladium roseum a versatile adversary of Botrytis cinerea in crops. Plant Dis. 1997, 81, 316–328. [Google Scholar] [CrossRef] [Green Version]
- Mueller, J.; Sinclair, J.B. Occurrence and role of Gliocladium roseum in field-grown soybeans in Illinois. Trans. Br. Mycol. Soc. 1986, 86, 677–680. [Google Scholar] [CrossRef]
- Muvea, A.M.; Meyhöfer, R.; Subramanian, S.; Poehling, H.-M.; Ekesi, S.; Maniania, N.K. Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS ONE 2014, 9, e108242. [Google Scholar] [CrossRef] [Green Version]
- Samsudin, N.I.P.; Rodriguez, A.; Medina, A.; Magan, N. Efficacy of fungal and bacterial antagonists for controlling growth, FUM1 gene expression and fumonisin B1 production by Fusarium verticillioides on maize cobs of different ripening stages. Int. J. Food Microbiol. 2017, 246, 72–79. [Google Scholar] [CrossRef]
- Sun, Z.-B.; Sun, M.-H.; Zhou, M.; Li, S.-D. Transformation of the endochitinase gene Chi67-1 in Clonostachys rosea 67-1 increases its biocontrol activity against Sclerotinia Sclerotiorum. AMB Express 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-Y.; Li, S.-D.; Sun, M.-H. Transaldolase gene Tal67 enhances the biocontrol activity of Clonostachys rosea 67-1 against Sclerotinia sclerotiorum. Biochem. Biophys. Res. Commun. 2016, 474, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Jiang, N.; Hasan, R.; Chen, Y.; Sun, M.; Li, S. Cell wall biogenesis protein phosphatase CrSsd1 is required for conidiation, cell wall integrity, and mycoparasitism in Clonostachys rosea. Front. Microbiol. 2020, 11, 1640. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.-B.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, M.-H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.-B.; Wang, Q.; Sun, M.-H.; Li, S.-D. The heat shock protein 70 gene is involved for colony morphology, sporulation and mycoparasitism of Clonostachys rosea. FEMS Microbiol. Lett. 2019, 366, fnz188. [Google Scholar] [CrossRef]
- Rodriguez, M.A.; Cabrera, G.; Gozzo, F.; Eberlin, M.; Godeas, A. Clonostachys rosea BAFC3874 as a Sclerotinia sclerotiorum antagonist: Mechanisms involved and potential as a biocontrol agent. J. Appl. Microbiol. 2011, 110, 1177–1186. [Google Scholar] [CrossRef]
- Keinath, A.; Fravel, D.; Papavizas, G. Potential of Gliocladium roseum for biocontrol of Verticillium dahliae. Phytopathology 1991, 81, 644–648. [Google Scholar] [CrossRef]
- James, T.; Sutton, J. Biological control of botrytis leaf blight of onion by Gliocladium roseum applied as sprays and with fabric applicators. Eur. J. Plant Pathol. 1996, 102, 265–275. [Google Scholar] [CrossRef]
- Abdullah, M.T.; Ali, N.Y.; Suleman, P. Biological control of Sclerotinia sclerotiorum (Lib.) de Bary with Trichoderma harzianum and Bacillus amyloliquefaciens. Crop Prot. 2008, 27, 1354–1359. [Google Scholar] [CrossRef]
- Hadi, M.; Sorkhi, R.; Kashefi, B.; Sobhanipur, A.; Shamsi, M.H. Investigation antifungal activity of some medicinal plant extracts on growth and spore germination of Penicillium digitatum Sacc. in vitro. Med. East J. Sci. Res 2013, 17, 1701–1708. [Google Scholar]
- Ravindran, D.R.; Bharathithasan, M.; Ramaiah, P.; Rasat, M.S.M.; Rajendran, D.; Srikumar, S.; Ishak, I.H.; Said, A.R.; Ravi, R.; Mohd Amin, M.F. Chemical composition and larvicidal activity of flower extracts from Clitoria ternatea against Aedes (Diptera: Culicidae). J. Chem. 2020, 2020, 3837207. [Google Scholar] [CrossRef]
- Lucie, A.T.; Dogo, S.; Béranger, L.D.P.; Florent, B.O.S.; Talla, G.M.; Anna, T.; Salomon, N.; Kandioura, N.; Mbacké, S.; Jean-Laurent, S.-M. Chemical characterization and insecticidal activity of ethyl acetate and dichloromethane extracts of Drypetes gossweileri against Sitophilus zeamais, Tribolium castaneum and Rhyzopertha dominica. J. Life Sci. 2013, 7, 1030. [Google Scholar]
- Gohilapriya, M.; Auxcilia, J.; Hemaprabha, M.A.R.K. Profiling of phytoconstituents in papaya varieties and wild genotypes as a omic breeding approach for Papaya Ring Spot Virus resistance (PRSV). Pharma Innov. J. 2021, 10, 976–982. [Google Scholar]
- Monteiro, V.N.; do Nascimento Silva, R.; Steindorff, A.S.; Costa, F.T.; Noronha, E.F.; Ricart, C.A.O.; de Sousa, M.V.; Vainstein, M.H.; Ulhoa, C.J. New insights in Trichoderma harzianum antagonism of fungal plant pathogens by secreted protein analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Shelke, D.B.; Tayade, S.; Gawande, P.; Sonawane, H.B. GC-MS analysis and antioxidant potential of wild underutilized medicinally important legume, velvet bean (Mucuna pruriens L. DC.). Not. Sci. Biol. 2022, 14, 11098. [Google Scholar]
- Jangir, M.; Pathak, R.; Sharma, S.; Sharma, S. Biocontrol mechanisms of Bacillus sp., isolated from tomato rhizosphere, against Fusarium oxysporum f. sp. lycopersici. Biol. Control 2018, 123, 60–70. [Google Scholar] [CrossRef]
- Majeed, F.A.; Munir, H.; Rashid, R.; Zubair, M.T. Antimicrobial, cytotoxicity, mutagenicity and anti-epileptic potential of ethanol extracts of a multipurpose medicinal plant Dalbergia sissoo. Biocatal. Agric. Biotechnol. 2019, 19, 101155. [Google Scholar] [CrossRef]
- Khan, I.H.; Javaid, A. In vitro biocontrol potential of Trichoderma pseudokoningii against Macrophomina phaseolina. Int. J. Agric. Biol. 2020, 24, 730–736. [Google Scholar]
- Senthamizh Selvan, N.; Isaiah, S. GC-MS Analysis and Antibacterial Activity of different Solvent Extracts of Shuteria involucrata. J. Phytopharm. 2018, 7, 116–120. [Google Scholar]
- Pourakbar, L.; Moghaddam, S.S.; Enshasy, H.A.E.; Sayyed, R. Antifungal activity of the extract of a macroalgae, Gracilariopsis persica, against four plant pathogenic fungi. Plants 2021, 10, 1781. [Google Scholar] [CrossRef]
- Akshaya, S.; Krishnamoorthy, A.; Nakkeeran, S.; Poornima, K.; Sivakumar, U. Inhibitory potential of ethyl acetate extract from mushrooms against root-knot nematode (Meloidogyne incognita). J. Entomol. Zool. Stud. 2021, 9, 528–534. [Google Scholar]
- Li, L.; Liu, Q.; Xue, H.; Bi, Y.; Raza, H.; Zhang, R.; Carelle, J.K.; Peng, H.; Long, H.; Prusky, D. Acetylsalicylic acid (ASA) suppressed Fusarium rot development and neosolaniol (NEO) accumulation by activating phenylpropane metabolism in muskmelon fruit. Eur. J. Plant Pathol. 2022, 163, 625–639. [Google Scholar] [CrossRef]
- Khan, I.H.; Javaid, A. Comparative antifungal potential of stem extracts of four quinoa varieties against Macrophomina phaseolina. Int. J. Agric. Biol. 2020, 24, 441–446. [Google Scholar]
- Mohamad, O.A.; Li, L.; Ma, J.-B.; Hatab, S.; Xu, L.; Guo, J.-W.; Rasulov, B.A.; Liu, Y.-H.; Hedlund, B.P.; Li, W.-J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [Green Version]
- Reghmit, A.; Benzina-tihar, F.; López Escudero, F.J.; Halouane-Sahir, F.; Oukali, Z.; Bensmail, S.; Ghozali, N. Trichoderma spp. isolates from the rhizosphere of healthy olive trees in northern Algeria and their biocontrol potentials against the olive wilt pathogen, Verticillium dahliae. Org. Agric. 2021, 11, 639–657. [Google Scholar] [CrossRef]
- Kachhadia, R.; Kapadia, C.; Singh, S.; Gandhi, K.; Jajda, H.; Alfarraj, S.; Ansari, M.J.; Danish, S.; Datta, R. Quorum Sensing Inhibitory and Quenching Activity of Bacillus cereus RC1 Extracts on Soft Rot-Causing Bacteria Lelliottia amnigena. ACS Omega 2022, 7, 25291–25308. [Google Scholar] [CrossRef]
- Sangeetha, C.; Krishnamoorthy, A. Production of Volatile Organic Compounds of Fusarium oxysporum f. sp. lycopersici on Coinoculation with the Metabolites of Chaetomium globosum. Madras Agric. J. 2021, 108, 1. [Google Scholar]
- Mohammed, A.A.; Ahmed, F.A.; Younus, A.S.; Kareem, A.A.; Salman, A.M. Molecular identification of two entomopathogenic fungus Clonostachys rosea strains and their efficacy against two aphid species in Iraq. J. Genet. Eng. Biotechnol. 2022, 20, 67. [Google Scholar] [CrossRef]
- Gromadzka, K.; Chelkowski, J.; Popiel, D.; Kachlicki, P.; Kostecki, M.; Golinski, P. Solid substrate bioassay to evaluate the effect of Trichoderma and Clonostachys on the production of zearalenone by Fusarium species. World Mycotoxin J. 2009, 2, 45–52. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, X.; Asemoloye, M.D.; Wang, Y.; Gargano, M.L.; Xue, H.-J.; Pecoraro, L. First record of Clonostachys rosea as entomopathogenic fungus of Coleoptera in China. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 1240–1246. [Google Scholar] [CrossRef]
- Mejía, L.C.; Rojas, E.I.; Maynard, Z.; Van Bael, S.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52 (Suppl. S1), 487–511. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J.; Budge, S. Screening for sclerotial mycoparasites of Sclerotinia sclerotiorum. Mycol. Res. 1990, 94, 607–612. [Google Scholar] [CrossRef]
- Jones, E.; Stewart, A. Selection of mycoparasites of sclerotia of Sclerotinia sclerotiorum isolated from New Zealand soils. New Zealand J. Crop Hortic. Sci. 2000, 28, 105–114. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Indumathi, T.; Nakkeeran, S. Trichoderma asperellum (NVTA2) as a potential antagonist for the management of stem rot in carnation under protected cultivation. Biol. Control 2017, 113, 58–64. [Google Scholar] [CrossRef]
- Rangaswami, G.; Mahadevan, A. Diseases of Crop Plants in India, 4th ed.; Prentice Hall of India Pvt Ltd.: New Delhi, India, 1998. [Google Scholar]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Sambrook, H.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Whipps, J.; Magan, N. Effects of nutrient status and water potential of media on fungal growth and antagonist-pathogen interactions [Sclerotinia sclerotiorum, Botrytis cinerea, Rhizoctonia solani; Coniothyrium minitans, Gliocladium roseum]. Bull. OEPP 1987, 17, 581–591. [Google Scholar] [CrossRef]
- Jing, L.; Navi, S.S.; Yang, X. Effects of colonisation by different strains of Coniothyrium minitans on the viability of sclerotia of Sclerotinia sclerotiorum. Biocontrol Sci. Technol. 2015, 25, 460–474. [Google Scholar] [CrossRef]
- Cawoy, H.; Debois, D.; Franzil, L.; De Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Fagbohun, O.F.; Olawoye, B.; Ademakinwa, A.N.; Oriyomi, O.V.; Fagbohun, O.S.; Fadare, O.A.; Msagati, T.A. UHPLC/GC-TOF-MS metabolomics, MTT assay, and molecular docking studies reveal physostigmine as a new anticancer agent from the ethyl acetate and butanol fractions of Kigelia africana (Lam.) Benth. fruit extracts. Biomed. Chromatogr. 2021, 35, e4979. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, Y.; Qin, J.; Qu, X.; Liu, J.; Li, X.; Pan, H. Antifungal metabolites produced by Chaetomium globosum No. 04, an endophytic fungus isolated from Ginkgo biloba. Indian J. Microbiol. 2013, 53, 175–180. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl.No. | Isolate | Percent Inhibition over Control * |
|---|---|---|
| 1 | TNAU-CR 01 | 70.37 c (57.02) |
| 2 | TNAU-CR 02 | 79.63 a (63.17) |
| 3 | TNAU-CR 03 | 74.07 b (59.39) |
| 4 | TNAU-CR 04 | 52.22 e (46.27) |
| 5 | TNAU-CR 05 | 58.14 d (49.68) |
| Sl.NO | Compound | Retention Time | Peak Area Percentage | Probability | Activity | References |
|---|---|---|---|---|---|---|
| 1 | Dihydro-3-(2H)-thiophenone | 3.123 | 0.277 | 11.2 | Antifungal activity | [30] |
| 2 | dl-Glyceraldehyde dimer | 3.324 | 1.774 | 59.3 | Insecticidal property | [31] |
| 3 | Propanedioic acid, phenyl- | 8.301 | 0.553 | 19.5 | Insecticidal property | [32] |
| 4 | Butanamide, 2-hydroxy-N,2,3,3-tetramethyl | 10.827 | 0.155 | 9.8 | Antiviral activity | [33] |
| 5 | à-Methyl 4-O-methyl-D-mannoside | 11.237 | 0.392 | 29.9 | Antifungal activity | [34] |
| 6 | 4-Methyl(trimethylene)silyloxyoctane | 12.187 | 0.188 | 15.8 | Antioxidant and antimicrobial activity | [35] |
| 7 | 1-Tetradecanamine, N, N-dimethyl- | 17.034 | 0.406 | 60.2 | Antifungal activity | [36] |
| 8 | Pilocarpine | 20.67 | 0.149 | 37.6 | Mutagenicity, cytotoxicity, and antimicrobial activity | [37] |
| 9 | Hexadecanoic acid, 2-hydroxy-1-(hydroxymethyl)ethyl ester | 21.281 | 1.356 | 52.3 | Antifungal activity | [38] |
| 10 | Pentadecanoic acid, 13-methyl-, methyl ester | 21.541 | 0.165 | 24.5 | Anti-bacterial activity | [39] |
| 11 | 9-Hexadecenoic acid | 21.751 | 0.49 | 9.9 | Antifungal activity | [40] |
| 12 | Octadecane, 3-ethyl-5-(2-ethylbutyl)- | 21.936 | 0.169 | 10.6 | Nematicidal property | [41] |
| 13 | Erucic acid | 24.047 | 1.088 | 11.2 | Antifungal activity | [42] |
| 14 | 8,11-Octadecadienoic acid, methyl ester | 24.697 | 0.178 | 9.1 | Antifungal activity | [43] |
| 15 | 9-Octadecenoic acid, methyl ester, (E)- | 24.822 | 0.289 | 7 | Antifungal activity | [44] |
| 16 | 13-Docosenamide, (Z)- | 26.358 | 4.829 | 72.4 | Antifungal activity | [45] |
| 17 | cis-11-Eicosenamide | 28.354 | 0.138 | 4.5 | Antibacterial activity | [46] |
| 18 | 17-Pentatriacontene | 28.554 | 0.262 | 10.9 | Antifungal activity | [47] |
| T. No. | Type of Treatments | Disease Incidence (Per Cent Disease) | ||
|---|---|---|---|---|
| 30 DAT | 45 DAT | 60 DAT | ||
| T1 | Foliar application of C. rosea isolate TNAU-CR 02 at 5 mL/litre on 30, 45 and 60 DAT | 5.9 b (14.05) | 8.3 b (16.73) | 15.1 b (22.86) |
| T2 | Foliar application of Trichoderma asperellum (TRI 15) at 5 mL/litre on 30, 45 and 60 DAT | 6.4 c (14.64) | 9.1 c (17.55) | 17.9 c (25.02) |
| T3 | Foliar application of Bacillus subtilis (Bbv 57) at 5 mL/litre on 30, 45 and 60 DAT | 6.9 bc (15.22) | 9.7 b (18.14) | 17.1 b (24.41) |
| T4 | Tebuconazole + trifloxystrobin at 1.5 g/litre on 30, 45 and 60 DAT | 0.0 a (2.86) | 2.2 a (8.52) | 5.9 a (14.05) |
| T5 | Inoculated control | 9.5 d (17.95) | 16.5 d (23.96) | 22.8 d (28.52) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venkatesan, R.M.; Muthusamy, K.; Iruthayasamy, J.; Prithiviraj, B.; Kumaresan, P.V.; Lakshmanan, P.; Perianadar, I.V. First Report of Clonostachys rosea as a Mycoparasite on Sclerotinia sclerotiorum Causing Head Rot of Cabbage in India. Plants 2023, 12, 199. https://doi.org/10.3390/plants12010199
Venkatesan RM, Muthusamy K, Iruthayasamy J, Prithiviraj B, Kumaresan PV, Lakshmanan P, Perianadar IV. First Report of Clonostachys rosea as a Mycoparasite on Sclerotinia sclerotiorum Causing Head Rot of Cabbage in India. Plants. 2023; 12(1):199. https://doi.org/10.3390/plants12010199
Chicago/Turabian StyleVenkatesan, Ruppavalli M., Karthikeyan Muthusamy, Johnson Iruthayasamy, Balakrishnan Prithiviraj, Parthiban V. Kumaresan, Pugalendhi Lakshmanan, and Irene Vethamoni Perianadar. 2023. "First Report of Clonostachys rosea as a Mycoparasite on Sclerotinia sclerotiorum Causing Head Rot of Cabbage in India" Plants 12, no. 1: 199. https://doi.org/10.3390/plants12010199