
The Alien Plant Species Impact in Rice Crops in Northwestern Italy

Department of Agricultural and Environmental Sciences, University of Milan, Via Celoria 2, I-20133 Milan, Italy
*
Author to whom correspondence should be addressed.
Plants 2023, 12(10), 2012; https://doi.org/10.3390/plants12102012
Submission received: 21 March 2023
/
Revised: 12 May 2023
/
Accepted: 15 May 2023
/
Published: 17 May 2023
(This article belongs to the Special Issue Effects of Invasive Alien Plant Species on Native Plant Diversity and Crop Yield)
Abstract
:Alien species represent one of the causes of biodiversity loss, both in natural and anthropic environments. This study contributes to the assessment of alien species impact on Western Po Plain rice field cultivations, referring to different agricultural management practices and ecological traits. Flora and vegetation were studied (the latter through the phytosociological method), and α-biodiversity was estimated through Shannon and Simpson Indices. Results highlighted a significant floristic contingent depletion and increase in therophyte and alien components, compared to pre-existing studies (1950s); higher α-biodiversity levels in organic farms, compared to conventional farms, but also a higher invasive alien species percentage. The high deterioration of the territorial–landscape context appears to play a major role in shaping these patterns. Some of these alien species are particularly aggressive (e.g., Murdannia keisak), as confirmed by two experimental rice field plots which were left unharvested, continuously flooded, making it possible to assess the competitiveness between weed species. The detected weed vegetation is attributed to the Oryzo sativae-Echinochloetum cruris-galli association, already described for Southern Europe, with two different ecological and floristic variants. Future studies, by including other sites and framing their territorial–landscape context, may further complement this overview on the alien species distribution and behavior in rice fields, hence facilitating their strategic management.
1. Introduction
Alien species invasion is described as one of the causes of biodiversity loss by many scientific studies [1,2,3,4,5,6,7,8,9,10] and reports from world organizations such as the IUCN and UN [11,12,13,14,15]. This not only concerns natural and seminatural habitats, but also anthropic environments [3,9]. Biodiversity loss alters the structure of ecosystems, their functionalities, and leads to a decline in ecosystem services, economic losses and human health issues [16,17,18,19,20].
In Italy, 1628 alien plant taxa are documented (about 16% of total Italian flora) [21,22,23,24,25,26], and can be mainly found in anthropic settings, such as urban and agricultural areas, reforested areas and artificial water courses, where they often become the dominant component of vegetational associations. The Po Plain, a highly urbanized, industrialized and intensively cultivated lowland in Northern Italy, is the most affected Italian region, especially with respect to invasive alien species [9,21,26,27]. Here, alien species tend to constitute monospecific populations or to become predominant in some habitats. They significantly shape plant communities and threaten the autochthonous habitats, especially those of conservation interest [27,28]. Moreover, it has been shown that intensive, highly simplified, agricultural systems and anthropized territories represent a threat for the native flora while favouring alien species invasion [29]. Such territorial traits affect the vast majority of the Po Plain area, with oversized agricultural patches, often matched with anthropic linear infrastructures, not leaving room to natural and seminatural ecosystems.
For Italy, there are no recent specific studies on alien plant species impacts on irrigated crops (including rice crops), except for a study conducted in Sardinia [30], which only focused on floristic and not vegetational traits. The most recent updates on Italian rice fields weed alien species are Viggiani’s reports [31,32]: they provide a generalized overview on the occurring taxa in Italy, but they do not address any specific impact issue. Analysis of weed vegetation in rice fields in the Po Plain (the first Italian and European rice producer region [33]) dates back to more than 40 years ago [34,35,36]; updated and comprehensive floristic–vegetational studies focusing on alien species impacts (also in relation to biodiversity issues) are currently lacking.
In this work, we focused on agricultural settings related to rice cultivation in the Western Po Plain area. Here, the impact of alien species has been increasing over the years [9,37], mainly due to changes in rice cultivation techniques [31]. In Italy, until the early 1960s, rice cultivation was generally based on traditional systems (wet seeding with continuous rice paddies flooding, starting before sowing and ending a few weeks before harvest). Nowadays, traditional practices are uncommonly applied, being almost entirely replaced by rice cultivation systems based on simplified rotation or monoculture, dry seeding with postponed flooding or alternate wetting and drying systems [38,39,40,41]. Traditional cultivars are mostly replaced by short-stature rice varieties managed through conventional rice systems, involving deep soil tillage and the extensive application of chemical herbicides, pesticides and mineral fertilizers. These shifts in agronomic and water management techniques, together with the prolonged chemical weed control, led to changes in rice weed flora and fostered alien species invasion [31,32,37,38,42]. Rice paddy environments often constitute refuge areas for rare species, substituting the role of wetlands and protected habitats, which are currently severely threatened by climate change, changes in agricultural management practices as well as by alien species invasion [42,43,44,45,46,47].
Our study objectives were to assess the alien plant species occurrence and consistency among differently managed rice farming systems (conventional versus organic systems) and to estimate their impact on biodiversity. This study is part of a wider research project where agrobiodiversity traits are assessed trough multi-scale approaches, where the influence of landscape scale processes is also taken into account [48]. Floristic–vegetational studies are also being conducted on other spontaneous phytocoenoses types (such as other crop fields, field banks, field margins, uncultivated areas, hedgerows and tree lines, woody areas, ditches, wetlands). For landscape scale analyses, landscape ecology [49,50,51,52,53] and landscape bionomic [54,55] approaches are adopted, framing the ecological functional patterns influencing field scale biodiversity.
2. Materials and Methods
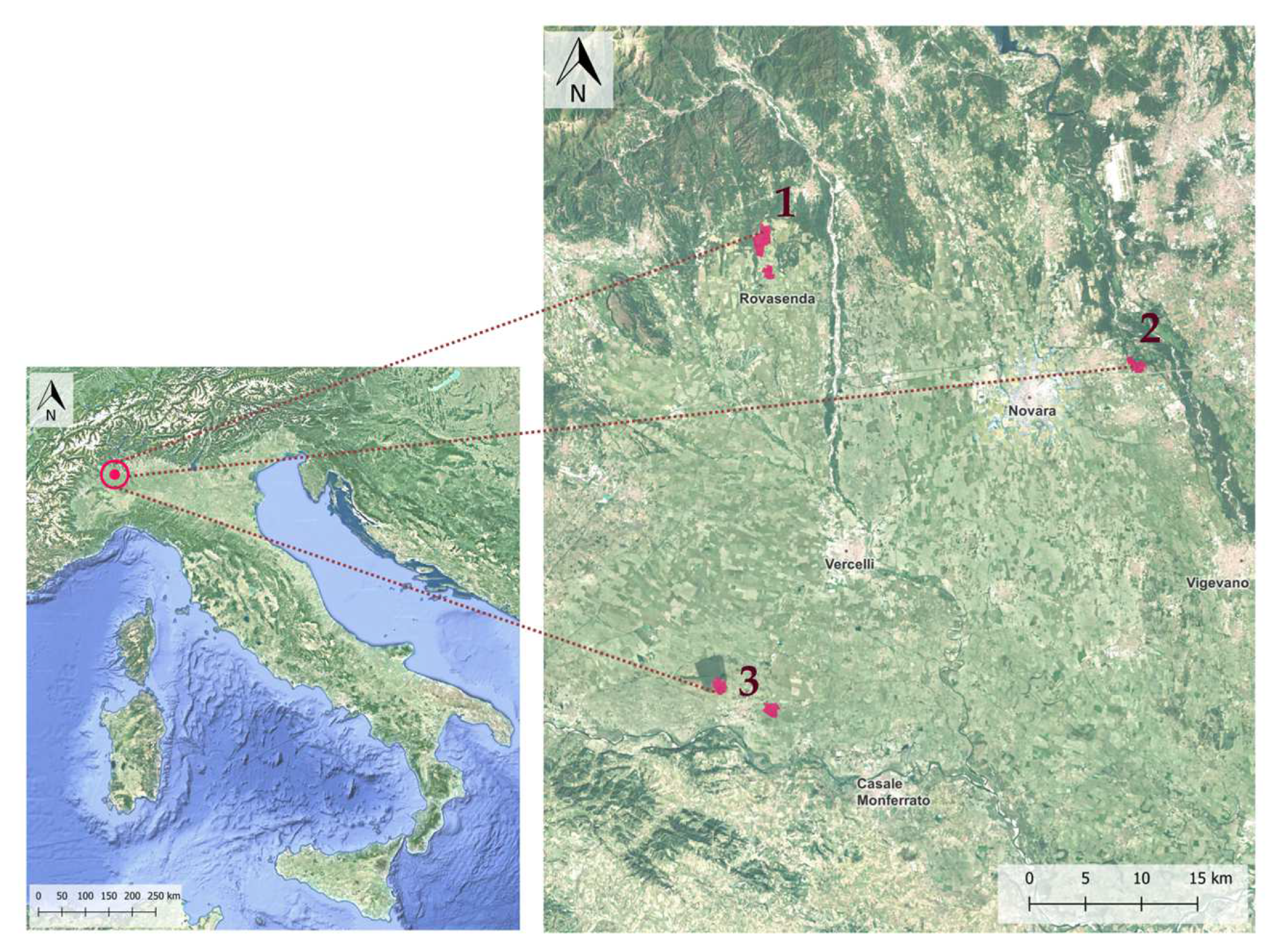
The study area is located in the Western Po Plain (Piedmont region) and consists of 3 sites belonging to the Vercelli and Novara districts (Rovasenda, Romentino and Trino Vercellese municipalities) (Figure 1). In each site, both organic and conventional farms were investigated.
The 3 sites were chosen because of the presence, among the organic farms, of 3 farms applying traditional cultivation practices based on crop diversification: rotations, continuous flooding conditions for rice fields, land races cultivation, presence of in-field linear embankments, maintenance of trenches along the field margins which are kept flooded throughout the year (allowing the persistence of diffused wet habitats for wild fauna and flora), spread hedgerows and tree lines in between fields (Figure 2).
The study area is characterized by a temperate bioclimate, ranging from temperate continental (from upper meso-temperate, upper sub-humid (site 3) to lower supra-temperate, lower humid (site 2)) to temperate oceanic (lower supra-temperate, lower humid (site 1)) according to the Rivas-Martinez Worldwide Bioclimatic Classification System [56,57,58,59,60].
Soils vary from young Entisols and Inceptisols (loamy texture; sub-acid (site 2) and calcareous (site 3) soils) to Riss alluvial terrace Alfisols (clay-silty texture, acid soils (site 1)) [61].
Flora and vegetation studies were carried out over 5 years (2018–2022) during the rice cultivation time period (from sowing to harvesting) among the 3 different sites, representing both conventional and organic rice cultivation systems (Figure 1). Only the species occurring inside the rice paddy were included in the analysis (i.e., excluding rice field banks, field edges and in-field embankments). Scientific nomenclature used is in accordance with the Italian flora system ([62,63,64,65]. We built a floristic list (see Table S1, Supplementary Materials) where each taxon was associated with its reference Raunkiær life form and chorotype [62,63,64,65,66]; ecological indicator values, according to the Ellenberg-Pignatti updated reference values, commonly adopted for the Italian flora [62,63,67,68,69]; and observation site. For life forms, the following abbreviations are applied: P = phanerophyte, Ch = chamaephyte, H = hemicryptophyte, G = geophyte, He/I = hydrophyte/helophyte, T = therophyte; scap = scapose, caesp = caespitose, bulb = bulbose, rhiz = rhizomatose, rept = reptant. The chorotypes of the native species were grouped into 7 biogeographical autochthonous regions: Boreal, Cosmopolitan, Euro-Asiatic, Orophyte South European, Mediterranean, Atlantic, Endemic. The others were classified as alien.
Floristic data were compared with Pomini’s pre-existing studies describing the rice fields weed flora detected in the same area (Vercelli and Novara districts) in the 1950s [70,71]. As in our study, Pomini’s data were collected during weed growing season (rice cultivation time frame). For data comparison, Pomini’s data were cleaned from species that were not detected within the rice paddy; scientific nomenclature was updated to current taxonomy; species were linked to their reference life forms and chorological traits, as made for current survey data. The floristic list obtained from Pomini’s studies also takes into account the previous study results [72,73,74]. Floristic studies following the 1950s have not been considered, because from the 1960s, agricultural practices widely changed (chemical synthesis products deployment) and the agri-environmental context became similar to that at present.
The vegetational study was based on the phytosociological method [75,76,77,78,79]: 40 phytosociological relevés were conducted among both conventional and organic farming systems, within the rice paddy, excluding rice field banks’ phytocoenoses. The relevés were tabulated, ordered and interpreted according to the phytosociological method, allowing us to identify vegetational communities with homogeneous floristic composition and ecology, which were then ascribed to the coherent syntaxonomic categories (associations, and the upper level syntaxa). Cover-abundance phytosociological indices (alpha-numerical Braun-Blanquet scale) were transformed into real numbers according to Van der Maarel [80] (conversion to central values, in order to compute the Specific Coverage Index (SCI) [78,81]). Statistical analyses were carried out using XLSTAT software, principal component analysis (PCA), and cluster analysis (using Euclidean distance as a dissimilarity coefficient). Syntaxa were classified according to the Prodromo della vegetazione d’Italia [82,83,84,85,86,87,88,89]. Quantitative biodiversity estimates were based on the J-Shannon Index [90] and E-Simpson Index [91], which were applied to phytosociological relevés. Four additional relevés were conducted over two years in two experimental plots (located in site 1) where rice was left unharvested and continuous flooding occurred during the whole year.
3. Results and Discussion
3.1. Flora
Floristic analysis (see floristic list in Table S1, Supplementary Materials) detected 38 plant taxa inside the rice paddies. This is a significantly lower number if compared to previous censuses conducted in the Vercelli and Novara districts rice fields during the 1950s, where 141 taxa were detected [70]. The decrease in taxa number is linked to agricultural practice changes and chemical herbicide spread [31,32,37]. Indeed, current flora are dominated by short-cycle herbaceous species (therophytes: 36.84%), followed by geophytes (21.50%), hydrophytes/helophytes (18.42%) and perennial herbaceous species (hemicryptophytes: 18.42%) (Figure 3). In contrast, past flora was dominated by hemicryptophytes (30.5%) and hydrophytes/helophytes (28.37%), followed by geophytes (22.70%) and therophytes (15.60%) [70] (Figure 3). The current therophyte abundance is coherent with the influence of dry seeding, alternate wet and drying irrigation practices, and generally higher soil disturbance, which predominate. Similar patterns were detected in other diachronic studies on irrigated crops. For instance, Covarelli’s study compared corn weed flora historical data (1960s versus 2000s) and reported a significant increase in therophytes and a parallel decrease in hemicryptophytes, while chamephytes almost disappeared [37]. Although these results do not refer to rice crops, they highlight a general trend that can be related to the abandonment of traditional cultivation practices and a shift towards more intensive cultivation techniques.
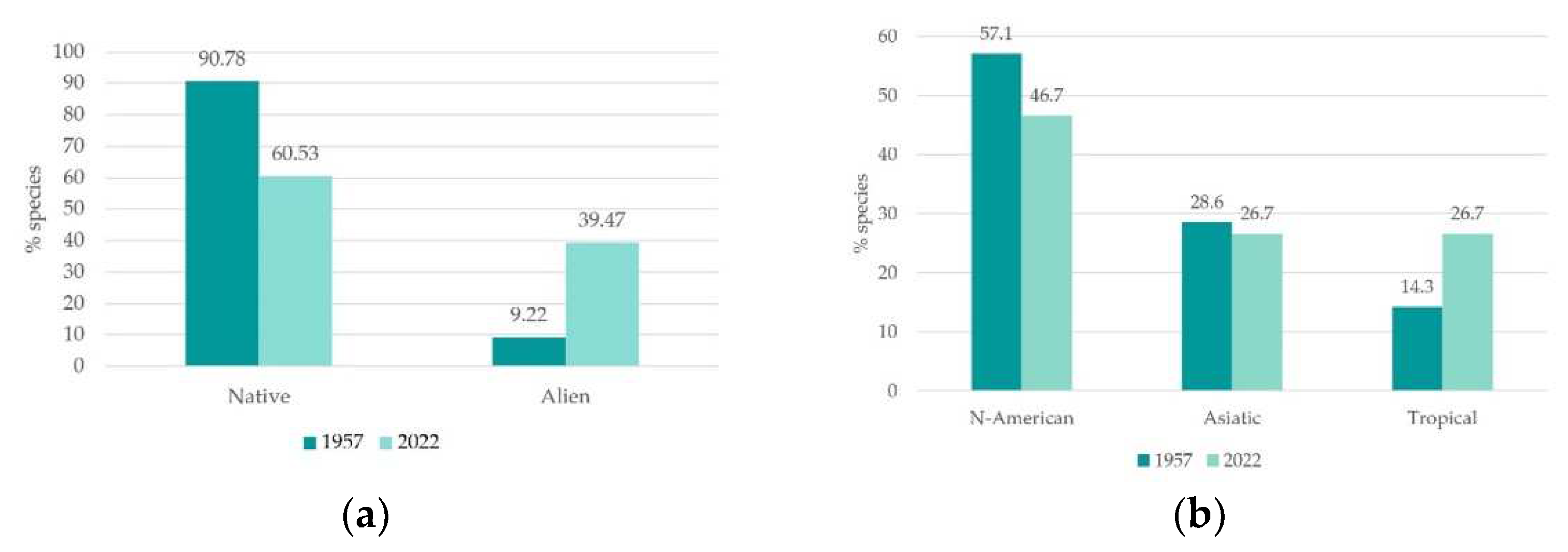
We observed a significant increase in alien species occurrence. Indeed, they represented 39.47% of total flora (of which 25.34% were invasive alien species), while they only represented 9.22% of total rice paddy flora in the 1950s [70] (Figure 4a). This result corresponds with the diachronic trends reported by other studies [9,37,92] and, accordingly to literature and to our study results, the occurrence of alien species poses a severe threat to biodiversity, as discussed hereafter. Compared to the 1950s, the alien species contingent has completely changed. The only invasive archaeophyte inventoried both in the 1950s and in current surveys was Oryza sativa L. var. sylvatica Chiappelli; the other 12 alien species observed in 1950s were not detected in the 3 study sites. The majority of the currently registered alien species are of northern origin: American (46.67%) followed by Asiatic (26.67%) and tropical (26.27%) (Figure 3b). Parallelly, in 1950s the species of American origin dominated (57.14%), followed by Asiatic (28.57%) and tropical (14.29) (Figure 4b). The decrease in North American species and the increase in the tropical ones might likely be a consequence of climate change and rice cultivation technique changes.
From a chorological perspective, the autochthonous species contingent has decreased, if compared to the 1950s (Figure 5). Wide distribution species have increased, to the detriment of the Euro-Asiatic and Boreal ones. In the 1950s, a low occurrence of Mediterranean s.l. and Atlantic species was registered, as well as an endemic species (Isoëtes malinverniana Ces. & De Not.), which all disappeared in current investigations.
Most of current detected species are of wide distribution and trivial, with the exception of Marsilea quadrifolia L., which is classified as an endangered species by the IUCN [47,93]. Many species observed in the 1950s and are now missing, are classified by the IUCN as critically endangered species (Isoëtes malinverniana Ces. & De Not., Pilularia globulifera L.), endangered species (Eleocharis carniolica W.D.J.Koch, Carex buekii Wimm., Typha minima Funk ex Hoppe, Sagittaria sagittifolia L., Myricaria germanica (L.) Desv., Utricularia vulgaris L.), vulnerable species (Salvinia natans (L.) All.) and near threatened species (Utricularia australis R.Br., Zannichellia palustris L.) [3,93,94]. These species also have become very rare in natural wetland environments outside rice fields. These environments, due to anthropogenic and climatic causes, have been greatly reduced and are often polluted. The rice field environment, if properly managed, can serve as an important secondary habitat for the survival of threatened species.
3.2. Vegetation
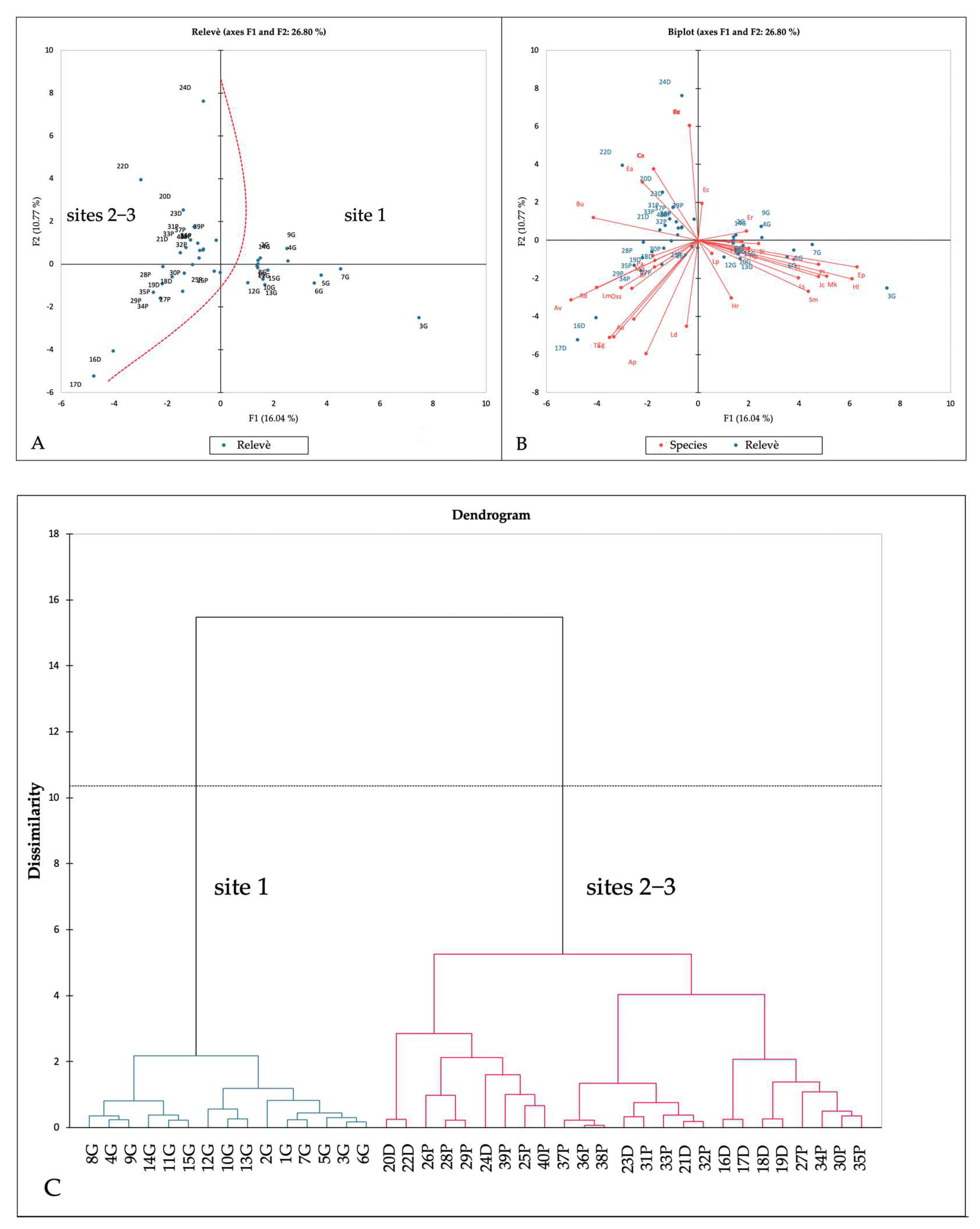
Table 1 reports the phytosociological relevés, ordered and grouped (Table 1, see Appendix A for further details). Cluster analysis and PCA analysis highlighted two distinct groups (Figure 6). The first includes the relevés made at site 1, while the second one includes those made at sites 2–3. These two main clusters are differentiated by groups of species, mainly of alien origin (group 1: Eleocharis flavescens (Poir.) Urb., Heteranthera limosa (Sw.) Willd., Murdannia keisak (Hassk.) Hand.-Mazz. and Juncus conglomeratus L.; group 2: Rotala densiflora (Roth) Koehne, Eleocharis acicularis (L.) Roem. & Schult. And Cyperus glomeratus L.).
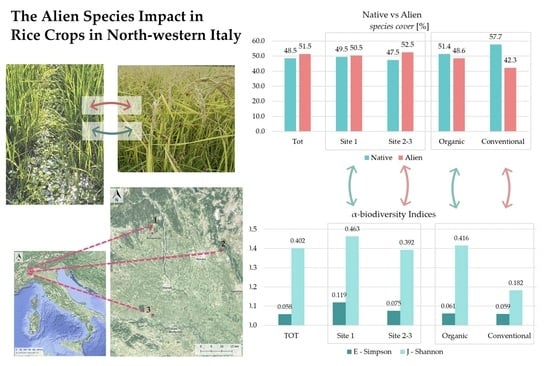
The vegetational study further highlighted alien species impact. Considering alien species coverage through SCI (Figure 7), alien species percentage overcame the native ones’ (51.5% versus 48.5%), especially in sites 2−3 (52.5% versus 47.5%). These impacts have to be linked to the higher anthropization degree of the territorial and landscape context of sites 2–3, which are located in the middle of the Po Plain. Sites 2 and 3 were more similar to each other than they were to site 1, from a biogeographical, floristic and ecological point of view.
Conventional rice fields showed a lower alien species percentage cover compared to the organic ones (Figure 7b). However, conventional fields generally had a lower occurrence and cover of the whole weed contingent: if we consider the mean species number, organic rice fields settled at 9.135, while the conventional ones settled at 5.778 (Table 2).
Periodical observations and surveys of the two experimental plots in site 1 (which were kept under continuous flooding conditions, leaving rice unharvested) showed an increase in alien species occurrence and coverage, to the detriment of the native ones (Table 3, see Appendix A for further details). Particularly, Murdannia keisak (Hassk.) Hand. −Mazz. exhibited greater aggressivity since it outcompeted native and non-native species and became dominant (cover values exceeding 75%). This led to a decrease of the overall species number (from 13 to 6) (Table 3). Murdannia keisak is a perennial geophyte which behaves as an annual species when growing in the rice field, where it easily reproduces via seeds. It also relies on vegetative propagation and tends to form dense and extensive populations, causing lodging of rice plants. These results seem to be in contrast with the results of a previous study conducted in Vercelli district [95], which specifically investigated Murdannia keisak responses to different water managements regimes. In this study, greenhouse experiments and field experiments indicated that continuous flooding conditions to be the best solution for preventing Murdannia keisak spread. Nonetheless, in this study, continuous flooding conditions were always paired to chemical pesticide application, while no pesticides were used in our experimental plots, as our aim was to simulate natural conditions which are likely to occur in wet natural habitats neighboring rice field habitats (acting as species source areas). These results could usefully orient targeted invasive alien species management, as well as other types of interventions aimed at supporting rice field system biodiversity (e.g., wetland restoration).
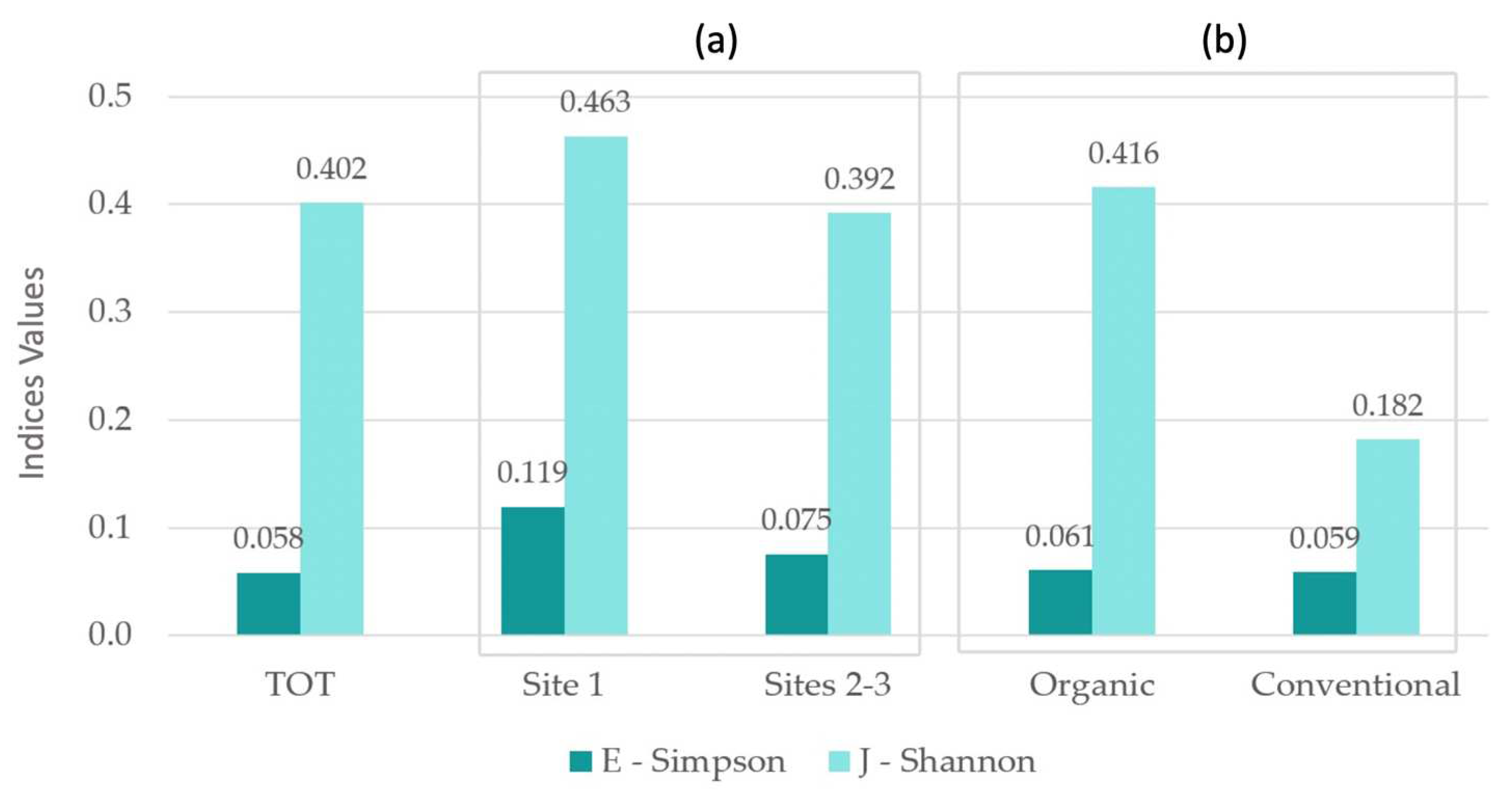
E-Simpson and J−Shannon indices where higher among organic rice fields (respectively, 0.061 versus 0.059 and 0.416 versus 0.182); the highest α-biodiversity values occurred in site 1 (respectively, 0.119 versus 0.075 in site 2−3 and 0.463 versus 0.392 in site 2−3), where the territorial and landscape context showed a higher naturality degree (Table 2; Figure 8).
Our study confirmed a significant alien species impact, even though it did not confirm a generalized direct negative relation between alien species and in-field biodiversity values. Alien species have a qualitative impact on biodiversity, more than the quantitative one. Our results support the following hypothesis: generally, the highly depleted ecological status of the territorial context entails higher vulnerability to the contamination of the floristic contingent by alien species [29,96]; higher biodiversity values can be linked to higher alien species occurrence (see organic field values) (Figure 7 and Figure 8). Where better territorial and landscape conditions occur (site 1), higher biodiversity values are linked to relatively lower alien species occurrence (site 1 versus site 2–3) (Figure 7 and Figure 8). All these results highlight the importance of adopting targeted precautions when addressing biodiversity issues in such depleted contexts. Notably, they highlight the need for multi-scale biodiversity assessment and management also through landscape-scale intervention strategies in order to successfully address biodiversity targets. These interpretations are aligned with Pellegrini’s study results (Northeastern Italy), which highlighted the role of landscape-scale extensive agricultural management in limiting alien plant invasion: even a small percentage of green infrastructure and extensive agricultural land amid the intensive one was proved to reduce alien species occurrence and facilitate native species diversity [29].
The weed species contingent showed various differences between site 1 and sites 2–3 (see Figure 6, Table 1 and Table S1, Supplementary Materials). In site 1, the following species occurred, which were absent in sites 2–3: Eleocharis flavescens (Poir.) Urb., Heteranthera limosa (Sw.) Willd., Murdannia keisak (Hassk.) Hand.−Mazz. and Juncus conglomeratus L. In contrast, the following species only occurred in sites 2 and 3: Rotala densiflora (Roth) Koehne, Eleocharis acicularis (L.) Roem. & Schult. and Cyperus glomeratus L. These floristic differences are due to single species bio-evolutionary and distribution reasons, as well as to ecological reasons. In fact, Ellenberg−Pignatti ecograms [62,63,67,68,69] show comparable climatic conditions between site 1 and site 2–3 (except for slightly higher temperatures at site 1) (Figure 9). Nonetheless, they highlight soil differences: in site 1, soils appeared to be more acid and poorer in adsorbable nitrogen content (Figure 9). Therefore, different soil characteristics may explain the different distribution of weed species in rice fields.
There are no recent phytosociological studies of Italian rice paddy vegetation and its syntaxonomic characterization.The most recent are those related to the Pavia and Vercelli districts [34,35] and Ferrara district [36]. The phytosociological data reported in these studies were used in a study by Carretero [97] concerning European rice field vegetation and reviewing the Oryzetea sativae Miyawaki 1960 class.
The vegetation detected in the study area (Table 1, Figure 6) can be ascribed to the Oryzo sativae−Echinochloetum cruris−galli Soo 1946 ex Ubrizsy 1948 association (an European widely distributed syntaxa [81]), even if depleted; indeed, only Echinochloa crus−galli (L.) P. Beauv. occasionally occurs among the overall character-species set. The association belongs to the Oryzo−Echinochloion oryzoidis O. Bolos & Masclans 1955 alliance, Cypero−Echinochloetalia oryzoidis O. Bolos & Masclans 1955 order, Oryzetea sativae Miyawaki 1960 class; this class includes weed vegetation of rice fields comprising vascular phanerogams and cryptogams, particularly algae. This vegetation might be referred to the thermophilic paspaletosum distichi W. Koch 1954 ex Carretero 1989 sub-association, which was identified for the Iberian Peninsula, Southern France and Italy [97], even though only Lindernia dubia (L.) Pennell was detected among the sub-association differential-species set. A variant with Eleocharis flavescens (Poir.) Urb., Heteranthera limosa (Sw.) Willd., Murdannia keisak (Hassk.) Hand.−Mazz. is identified at site 1, indicating more acidic substrates poor in organic matter (Table 1, Figure 6).
In addition to the characteristic and differential species of the association and the upper syntaxa, companion species are listed in Table 1. Among companion species, we underline the occurrence of species typical of marshy environments ascribed to the Phragmito australis−Magnocaricetea elatae Klika in Klika & Novk 1941 class (such as: Typha latifolia L., T. angustifolia L., Alisma plantago-aquatica L.), that come from the neighboring wet environments (channels, irrigation ditches and ponds). Among the companion species, there were also nitrophilous, hygrophilous pioneer species belonging to the Bidentetea tripartitae Tuxen, Lohmeyer & Preising ex Von Rochow 1951 class (such as: Bidens frondosa L. and Persicaria lapathifolia (L.) Delarbre), that come from the dry environments (field banks, in-field embankments, uncultivated areas).
Further Northwestern Po Plain data collection is needed in order to build a more comprehensive overview of rice paddies’ floristic and vegetational traits, allowing description of new sub-associations able to demonstrate the floristic and ecological characteristics of the rice fields belonging to these Po Plain districts.
4. Conclusions
This study contributes in updating the current knowledge on rice fields’ weed flora and vegetation in Vercelli and Novara Western Po Plain districts. Specifically, it depicts a significant change in rice paddies’ spontaneous weed flora, compared to the 1950s: the overall taxa number has dramatically decreased; therophyte species, tightly linked to higher variability and disturbance of soil conditions, are favored by current cultivation practices, and have become predominant; current flora is depleted also from a chorological point of view, and the alien species proportion has significantly increased. Moreover, species of conservation interest which were detected in the 1950s were not encountered (except for Marsilea quadrifolia L.). This shows how the current rice field environmental and management conditions in the Western Po Plain are undermining the capability of rice field systems to behave as spread wet habitats supporting agrobiodiversity.
Moreover, vegetational studies have shown how organic rice field management sustains higher α-biodiversity values, compared to conventional techniques. Nonetheless, organic farms’ rice field phytocoenoses also host higher invasive alien species amounts. The results suggested that the depleted, overexploited territorial and landscape context might play a major role in enhancing the vulnerability to alien species invasion (differences between site 1 and sites 2–3, located in the middle of the intensively cultivated Po Plain). Murdannia keisak (Hassk.) Hand.−Mazz. appeared to be the most aggressive alien species among permanently flooded areas (such as wetlands), at site 1. Finally, vegetation studies showed how the distribution of rice field weed species (especially alien species) is mainly related to edaphic parameters, especially soil pH and organic matter content. In fact, a variant of the Oryzo sativae−Echinochloetum cruris−galli Soo 1946 ex Ubrizsy 1948 association was identified in site 1, and was related to more acidic and organic matter-poor soils.
Given the recognized growing impacts of alien species invasion, the here-detected patterns between biodiversity values, alien species occurrence, and territorial–landscape context deterioration, urge further floristic–vegetational investigations of Po Plain rice fields, also through multi-scale biodiversity assessment approaches. This could help in identifying targeted precautions for addressing biodiversity issues in Po Plain rice lands. Rice fields, if properly managed, are wet habitats that can act as biodiversity source areas and plant diversity conservation sites, where productive and conservation functions can positively coexist [42,46,47]. Rice fields are also pivotal habitats for faunal conservation: indeed, rice lands often host high conservation interest areas, such as “Natura 2000” European Network sites [98,99]. However, for effective agrobiodiversity support, the influence of the upper scale landscape context should be primarily taken into account [100,101,102]. Indeed, landscape system ecological rehabilitation and diversification can directly and indirectly influence rice field and, more generally, crop field biodiversity and alien species invasion patterns [29,96,100,101,102,103,104,105]. Hence, multi-scale biodiversity assessments are needed for orienting landscape scale management strategies, in order to successfully address biodiversity targets [106]. Our study results would benefit from the integration of landscape scale ecological analyses as well as from the assessment of the out-field spontaneous flora and vegetation, allowing better framing and understanding of the reasons behind the alien species impact patterns.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12102012/s1, Table S1: Floristic List.
Author Contributions
Conceptualization, I.V.; methodology, I.V.; software, I.V. and G.C.; validation, I.V. and G.C.; formal analysis, I.V.; investigation, I.V.; resources, I.V.; data curation, I.V.; writing—original draft preparation, I.V. and G.C.; writing—review and editing, I.V. and G.C.; visualization, I.V.; supervision, I.V.; project administration, I.V.; funding acquisition, I.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table 1: Localities and dates: rel. 1G, 2G Organic Farm Cascina dell’Angelo, Rovasenda (VC) 8 September 2019; rel. 3G, 4G, 5G, 6G, 7G, 8G, 9G Organic Farm Baraggia, Rovasenda (VC) 24 July 2018; rel. 10G Organic Farm Baraggia, Rovasenda (VC) 14 July 2022; rel. 11G, 12G, 13G Organic Farm Cascina Teglio, Rovasenda (VC) 23 July 2019; rel. 14G, 15G Conventional Farm Rovasenda (VC) 14 July 2022; rel. 16D, 17D Organic Farm Dulcamara, Romentino (NO) 23 July 2019; rel. 18D, 19D Organic Farm Dulcamara Romentino (NO) 15 September 2020; rel. 20D, 21D, 22D Organic Farm Romentino (NO) 6 June 2020; rel. 23D, 24D Conventional Farm Santuario del Varallino, Romentino (NO) 6 June 2020; rel. 25P, 26P, 27P, 28P, 29P Organic Farm Priorato, Trino Vercellese (VC) 26 July 2019; rel. 30P, 31P, 32P, 33P, 34P, 35P Organic Farm Priorato, Trino Vercellese (VC) 14 July 2022; rel. 36P, 37P, 38P Conventional Trino Vercellese (VC) 14 July 2022; rel. 39P, 40P Conventional Farm Trino Vercellese (VC) 26 July 2019.
Sporadic species—rel. 3G: Lythrum salicaria L. +, rel. 5G: Hypericum perforatum L. +, rel. 7G: Setaria italica subsp. viridis (L.) Thell. +, rel. 12G: Lolium perenne L. +, rel. 17D: Artemisia vulgaris L. +; rel. 24D: Commelina communis L. +, Erigeron canadensis L. +, Amaranthus retroflexus L. 2.3, rel. 40P: Portulaca nitida (Danin & H.G. Baker) Ricceri & Arrigoni +.
Table 3: Locality and dates: Organic Farm Cascina dell’Angelo, Rovasenda (VC), rel. 1: 13 July 2018, 2: 24 July 2018, rel. 3, 4: 23 July 2019.
References
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Gentili, R.; Schaffner, U.; Martinoli, A.; Citterio, S. Invasive alien species and biodiversity: Impacts and management. Biodiversity 2021, 22, 1–3. [Google Scholar] [CrossRef]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing plants under full national responsibility: Extinction risk and threats in the vascular flora endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Gallardo, B.; Bacher, S.; Bradley, B.; Comín, F.; Gallien, L.; Jeschke, J.; Sorte, C.; Vilà, M. InvasiBES: Understanding and managing the impacts of Invasive alien species on Biodiversity and Ecosystem Services. NeoBiota 2019, 50, 109–122. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Williams, J.W.; Slayback, D.; Stoms, D.M.; Viers, J.H.; Dobson, A.P. Human impacts, plant invasion, and imperiled plant species in California. Ecol. Appl. 2006, 16, 1338–1350. [Google Scholar] [CrossRef]
- Baker, R.; Cannon, R.; Bartlett, P.; Barker, I. Novel strategies for assessing and managing the risks posed by alien species to global crop production and biodiversity. Ann. Appl. Biol. 2005, 146, 177–191. [Google Scholar] [CrossRef]
- Mack, R.; Simberloff, D.; Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F. Biotic Invasions: Causes, Epidemiology, Global Consequences, and Control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Walker, B.; Steffen, W. An Overview of the Implications of Global Change for Natural and Managed Terrestrial Ecosystems. Conserv. Ecol. 1996, 1, 2. [Google Scholar] [CrossRef]
- Celesti-Grapow, L.; Alessandrini, A.; Arrigoni, P.V.; Assini, S.; Banfi, E.; Barni, E.; Bovio, M.; Brundu, G.; Cagiotti, M.R.; Camarda, I.; et al. Non-native flora of Italy: Species distribution and threats. Plant Biosyst. 2010, 144, 12–28. [Google Scholar] [CrossRef]
- Dogra, K.; Sood, S.; Dobhal, P.; Sharma, S. Alien plant invasion and their impact on indigenous species diversity at global scale: A review. J. Ecol. Nat. Environ. 2010, 2, 175–186. [Google Scholar]
- Clout, M.; De Poorter, M. IUCN Guidelines for the Prevention Of Biodiversity Loss Caused By Alien Invasive Species. Aliens 2000, 11, 1–21. [Google Scholar]
- IUCN. Invasive Alien Species and Climate Change. Available online: https://www.iucn.org/sites/default/files/2022-04/ias_and_climate_change_issues_brief_2021.pdf (accessed on 16 May 2023).
- IUCN. Unified Classification of Direct Threats: Version 3.2. Available online: https://nc.iucnredlist.org/redlist/content/attachment_file/dec_2012_guidance_threats_classification_scheme.pdf (accessed on 13 March 2023).
- Convention on Biological Diversity. Available online: https://www.cbd.int/invasive/ (accessed on 14 March 2023).
- Secretariat of the Convention on Biological Diversity. Invasive Alien Species, A Threat To Biodiversity; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2009.
- Levine, J.M.; Vilà, M.; Antonio, C.M.D.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Bai, B.B. Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species. Environ. Entomol. 2008, 37, 277. [Google Scholar] [CrossRef]
- Inghilesi, A.F.; Mazza, G.; Cervo, R.; Gherardi, F.; Sposimo, P.; Tricarico, E.; Zapparoli, M. Alien insects in Italy: Comparing patterns from the regional to European level. J. Insect Sci. 2013, 13, 73. [Google Scholar] [CrossRef]
- Adla, K.; Dejan, K.; Neira, D.; Dragana, Š. Chapter 9—Degradation of ecosystems and loss of ecosystem services. In One Health; Prata, J.C., Ribeiro, A.I., Rocha-Santos, T., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 281–327. [Google Scholar]
- Markandya, A. The Economic Feedbacks of Loss of Biodiversity and Ecosystems Services. OECD Environ. Work. Pap. 2015, 93, 1–26. [Google Scholar] [CrossRef]
- Domina, G. Invasive Aliens in Italy. In Invasive Alien Species: Observations and Issues from Around the World, Volume 3: Issues and Invasions in Europe, First Edition; John Wiley & Sons: New York, NY, USA, 2021; pp. 190–214. [Google Scholar]
- Galasso, G.; Domina, G.; Adorni, M.; Ardenghi, N.; Bonari, G.; Buono, S.; Cancellieri, L.; Chianese, G.; Ferretti, G.; Fiaschi, T.; et al. Notulae to the Italian alien vascular flora: 5. Ital. Bot. 2018, 5, 45–56. [Google Scholar] [CrossRef]
- Galasso, G.; Domina, G.; Alessandrini, A.; Ardenghi, N.; Bacchetta, G.; Ballelli, S.; Bartolucci, F.; Brundu, G.; Buono, S.; Busnardo, G.; et al. Notulae to the Italian alien vascular flora: 6. Ital. Bot. 2018, 6, 65–90. [Google Scholar] [CrossRef]
- Galasso, G.; Domina, G.; Ardenghi, N.; Aristarchi, C.; Bacchetta, G.; Bartolucci, F.; Bonari, G.; Bouvet, D.; Brundu, G.; Buono, S.; et al. Notulae to the Italian alien vascular flora: 7. Ital. Bot. 2019, 7, 157–182. [Google Scholar] [CrossRef]
- Galasso, G.; Domina, G.; Andreatta, S.; Angiolini, C.; Ardenghi, N.; Aristarchi, C.; Arnoul, M.; Azzella, M.; Bacchetta, G.; Bartolucci, F.; et al. Notulae to the Italian alien vascular flora: 8. Ital. Bot. 2019, 8, 63–93. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.; Adorni, M.; Aleffi, M.; Allegrezza, M.; Angiolini, C.; et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociol. 2020, 57, 29–54. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef]
- Pellegrini, E.; Buccheri, M.; Martini, F.; Boscutti, F. Agricultural land use curbs exotic invasion but sustains native plant diversity at intermediate levels. Sci. Rep. 2021, 11, 8385. [Google Scholar] [CrossRef]
- Cossu, T.A.; Camarda, I.; Brundu, G. A catalogue of non-native weeds in irrigated crops in Sardinia (Italy). Webbia 2014, 69, 145–156. [Google Scholar] [CrossRef]
- Viggiani, P.; Tabacchi, M. Piante Infestanti di Risaie e Canali: Botanica e Riconoscimento; Edagricole: Milano, Italy, 2017. [Google Scholar]
- Viggiani, P. Weed flora in Italian rice fields. In Proceedings of the 13th EWRS symposium, Bari, Italy, 19–23 June 2005. [Google Scholar]
- European Commission. agridata.ec.europa.eu. Available online: https://agridata.ec.europa.eu/extensions/DashboardRice/RiceProduction.html (accessed on 1 March 2023).
- Pignatti, S. La vegetazione delle risaie pavesi: Studio fitosociologico. Arch. Bot. Biogeogr. Ital. 1957, 33, 129–193. [Google Scholar]
- Tomaselli, R. Aspetti della vegetazione in risaia da vicenda del Pavese e del Vercellese prima e dopo il diserbo. Arch. Bot. Biogeogra. Ital. 1958, 34, 217–253. [Google Scholar]
- Piccoli, F.; Gerdol, R. Rice-field weed communities in Ferrara Province (northern Italy). Aquat. Bot. 1981, 10, 317–328. [Google Scholar] [CrossRef]
- Covarelli, G. Evoluzione della flora e della vegetazione infestante le principali colture agrarie in Italia. Fitosociologia 2002, 39, 3–13. [Google Scholar]
- Ferrero, A.; Tinarelli, A. Rice Cultivation in the E.U. Ecological Conditions and Agronomical Practices. In Pesticide Risk Assessment in Rice Paddies: Theory and Practice; Capri, E., Karpouzas, D.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Hill, J.H.; Bayer, D.E.; Bocchi, S.; Clampett, W.S. Direct Seeded Rice in the Temperate Climates of Australia, Italy and North America; International Rice Research Institute: Los Baños, Pilippines, 1991; Volume IRRI Monograph, pp. 91–103. [Google Scholar]
- Monaco, S.; Volante, A.; Orasen, G.; Cochrane, N.; Oliver, V.; Price, A.H.; Teh, Y.A.; Martínez-Eixarch, M.; Thomas, C.; Courtois, B.; et al. Effects of the application of a moderate alternate wetting and drying technique on the performance of different European varieties in Northern Italy rice system. Field Crop. Res. 2021, 270, 108220. [Google Scholar] [CrossRef]
- Zampieri, M.; Ceglar, A.; Manfron, G.; Toreti, A.; Duveiller, G.; Romani, M.; Rocca, C.; Scoccimarro, E.; Podrascanin, Z.; Djurdjevic, V. Adaptation and sustainability of water management for rice agriculture in temperate regions: The Italian case-study. Land Degrad. Dev. 2019, 30, 2033–2047. [Google Scholar] [CrossRef]
- Rossi, G.; Tazzari, E.; Abeli, T.; Cauzzi, P.; Ardenghi, N.; Orsenigo, S.; Vagge, I. Activities and Perspectives of Plant Diversity Conservation in Rice Paddies (and Surrounding) of Po River Plain, N-Italy. In Organic Rice Farming and Production Systems: Conference Folder; Università degli Studi di Milano: Milan, Italy, 2015. [Google Scholar]
- Fasola, M.; Ruiz, X. The Value of Rice Fields as Substitutes for Natural Wetlands for Waterbirds in the Mediterranean Region. Colon. Waterbirds 1996, 19, 122. [Google Scholar] [CrossRef]
- Sánchez-Guzmán, J.M.; Morán, R.; Masero, J.A.; Corbacho, C.; Costillo, E.; Villegas, A.; Santiago-Quesada, F. Identifying new buffer areas for conserving waterbirds in the Mediterranean basin: The importance of the rice fields in Extremadura, Spain. Biodivers. Conserv. 2007, 16, 3333–3344. [Google Scholar] [CrossRef]
- Bogliani, G.; Della Rocca, F. Biodiversity and Rice Production in Rice Agro-Ecosystem—The Action Plan—Action E.5; International Rice Field Ecological Network LIFE Project LIFE09 NAT/IT/000093 ECORICE; International Rice Research Institute: Manila, Philippines, 2014. [Google Scholar]
- Orsenigo, S.; Corli, A. Buone Pratiche di Gestione di Risaie e Prati Umidi per la Conservazione di Specie Vegetali di Interesse Comunitario; Università di Pavia: Pavia, Italy, 2022; Volume Manuali del progetto CLOVER: Agroecosistemi e Conservazione in LOmbardia di specie VEgetali Rare di Direttiva Habitat. [Google Scholar]
- Corli, A.; Orsenigo, S.; Gerdol, R.; Bocchi, S.; Smolders, A.P.; Brancaleoni, L.; Caffi, M.T.; Abeli, T.; Rossi, G. Coexistence of rice production and threatened plant species: Testing Marsilea quadrifolia L. in N-Italy. Paddy Water Environ. 2021, 19, 395–400. [Google Scholar] [CrossRef]
- Vagge, I.; Chiaffarelli, G. Validating the Contribution of Nature-Based Farming Solutions (NBFS) to Agrobiodiversity Values through a Multi-Scale Landscape Approach. Agronomy 2023, 13, 233. [Google Scholar] [CrossRef]
- Dramstad, W.E.; Olson, J.D.; Forman, R.T.T. Landscape Ecology Principles in Landscape Architecture and Land Use Planning; Island Press: Washington, DC, USA, 1996. [Google Scholar]
- Dover, J.W.; Bunce, R.G.H. Key Concepts in Landscape Ecology; IALE UK, Coplin Cross Printers Ltd.: Garstang, UK, 1998. [Google Scholar]
- Forman, R.T.T. Land Mosaics: The Ecology of Landscapes and Regions, 1st ed.; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Forman, R.T.T.; Godron, M. Landscape Ecology; J. Wiley and Sons: New York, NY, USA, 1986. [Google Scholar]
- Turner, M.G.; Gardner, R.H. Landscape Ecology in Theory and Practice, Pattern and Process; Springer Verlag: New York, NY, USA, 2015. [Google Scholar]
- Ingegnoli, V. Landscape Bionomics: Biological-Integrated Lanscape Ecology; Springer: Milan, Italy, 2015. [Google Scholar]
- Ingegnoli, V. Landscape Ecology: A Widening Foundation; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Globalbioclimatics. Available online: www.globalbioclimatics.org (accessed on 21 October 2022).
- Rivas-Martínez, S. Global Bioclimatics, Clasificación Bioclimática de la Tierra; CIF: Madrid, Spain, 2004. [Google Scholar]
- Rivas-Martínez, S.; Sáenz, S.; Penas, A. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 634. [Google Scholar]
- Geoportale Piemonte. Available online: www.geoportale.piemonte.it/cms/ (accessed on 10 October 2022).
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017–2019. [Google Scholar]
- Dryades. Available online: http://dryades.units.it/cercapiante/index.php (accessed on 11 October 2022).
- Flora Italiae ActaPlantarum. Available online: www.floraitaliae.actaplantarum.org (accessed on 11 October 2022).
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon press: Oxford, UK, 1934. [Google Scholar]
- Domina, G.; Galasso, G.; Bartolucci, F.; Guarino, R. Ellenberg Indicator Values for the vascular flora alien to Italy. Electronic Supplementary File 1. Flora Mediterr. 2018, 28, 53–61. [Google Scholar] [CrossRef]
- Guarino, R.; Domina, G.; Pignatti, S. Ellenberg’s Indicator values for the Flora of Italy—First update: Pteridophyta, Gymnospermae and Monocotyledoneae. Flora Mediterr. 2012, 22, 197–209. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di bioindicazione delle piante vascolari della flora d’Italia, Bioindicator values of vascular plants of the flora of Italy. Braun-Blanquetia Rev. Geobot. Monogr. 2005, 39, 1–97. [Google Scholar]
- Pomini, L. Saggio di Flora Della Risaia Vercellese-Novarese; Istituto Tecnico Agrario di Vercelli—S.A.V.I.T.: Vercelli, Italy, 1957. [Google Scholar]
- Pomini, L. Piante Vascolari Infestanti la Risaia; Istituto Tecnico Agrario di Vercelli: Vercelli, Italy, 1955; Volume 3. [Google Scholar]
- Ciferri, R.; Giacomini, V.; Poggio, P. La Flora Fanerogamica Delle Risaie Dell’italia Transpadana. Suppl. Agli Atti Dell’istituto Bot. Dell’università Lab. Crittogam. Pavia 1949, 5, 1–26. [Google Scholar]
- Koch, W. Pflanzensoziologische skizzen aus den reisfeld-gebieten des piemont (Po-ebene). Vegetatio 1954, 5/6, 487–493. [Google Scholar] [CrossRef]
- Koch, W. Zur Flora der Oberitalienischen Reisfelder; Buchdruckerei Buchler & Co: Bern, Switzerland, 1952; Volume 62. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde; Springer: Wien, Austria; New York, NY, USA, 1964. [Google Scholar]
- Géhu, J.M. L’analyse symphytosociologique et géosymphytosociologique de l’espace, Théorie et métodologie. Coll. Phytosoc. 1988, XVII, 11–46. [Google Scholar]
- Géhu, J.M.; Rivas-Martínez, S. Notions fondamentales de phytosociologie. Ber. Int. Simp. Int. Ver. Veg. 1981, 31.3, 5–33. [Google Scholar]
- Pirola, A. Elementi di Fitosociologia; CLUEB: Bologna, Italy, 1970. [Google Scholar]
- Rivas-Martínez, S. Nociones sobre Fitosociología, Biogeografía e Bioclimatología. In La Vegetation de España; Universidad de Alcalá de Henares: Madrid, Spain, 1987; pp. 19–45. [Google Scholar]
- van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer: Berlin/Heidelberg, Germany, 1928. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Vagge, I.; Blasi, C. New and validated syntaxa for the checklist of Italian vegetation. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 318–332. [Google Scholar] [CrossRef]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Soriano, P.; Tesei, G.; Blasi, C. New insight on Mediterranean and sub-Mediterranean syntaxa included in the Vegetation Prodrome of Italy. Flora Mediterr. 2015, 25, 77–102. [Google Scholar] [CrossRef]
- Biondi, E.; Casavecchia, S.; Pesaresi, S.; Gangale, C.; Uzunov, D. New syntaxa for the prodrome of Italian vegetation. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 723–727. [Google Scholar] [CrossRef]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Poldini, L.; Sburlino, G.; Vagge, I.; Venanzoni, R. New syntaxonomic contribution to the Vegetation Prodrome of Italy. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2015, 149, 603–615. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 728–814. [Google Scholar] [CrossRef]
- Geoportale Nazionale. Available online: www.pcn.minambiente.it/mattm/ (accessed on 10 October 2022).
- Prodromo Della Vegetazione Italiana. Available online: www.prodromo-vegetazione-italia.org (accessed on 21 October 2022).
- Verde, S.; Assini, S.; Andreis, C. Le Serie di Vegetazione Della Regione Lombardia. Available online: https://air.unimi.it/handle/2434/149481 (accessed on 16 May 2023).
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1949; pp. 1–125. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Ferrero, A.; Vidotto, F. Weeds and weed management in Italian rice fields. In Agro-Economical Traits of Rice Cultivation in Europe and India; Edizioni Mercurio: Vercelli, Italy, 2007; pp. 55–72. [Google Scholar]
- Rossi, G.; Montagnani, C.; Gargano, D.; Peruzzi, L.; Abeli, T.; Ravera, S.; Cogoni, A.; Giuseppe, F.; Magrini, S.; Gennai, M.; et al. Lista Rossa della Flora Italiana. 1. Policy Species e Altre Specie Minacciate; Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Rome, Italy, 2013. [Google Scholar]
- Rossi, G.; Orsenigo, S.; Gargano, D.; Montagnani, C.; Peruzzi, L.; Giuseppe, F.; Abeli, T.; Alessandrini, A.; Astuti, G.; Bacchetta, G.; et al. Lista Rossa IUCN Della Flora Italiana: 2. ENDEMITI e Altre Piante Minacciate; Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Rome, Italy, 2020. [Google Scholar]
- Ferrero, A.; Tesio, F.; Tabacchi, M.; Vidotto, F. The effects of water management, timing and the rate of several herbicides on the growth of Murdannia keisak (Hassk.) Handel-Mazz. Crop Prot. 2012, 38, 53–56. [Google Scholar] [CrossRef]
- O’Reilly-Nugent, A.; Palit, R.; Lopez-Aldana, A.; Medina-Romero, M.; Wandrag, E.; Duncan, R.P. Landscape Effects on the Spread of Invasive Species. Curr. Landsc. Ecol. Rep. 2016, 1, 107–114. [Google Scholar] [CrossRef]
- Carretero, J.L. La vegetacion emergente de los arrozales europeos. Anales de Biologia 1989, 15, 135–141. [Google Scholar]
- European Parliament. Directive 2009/147/EC of the European Parliament and of the Council of 30 November 2009 on the conservation of wild birds. Off. J. Eur. Union 2010, L 20/7, 19. [Google Scholar]
- Council of the European Communities. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 50. [Google Scholar]
- Chiatante, G.; Pellitteri-Rosa, D.; Torretta, E.; Nonnis Marzano, F.; Meriggi, A. Indicators of biodiversity in an intensively cultivated and heavily human modified landscape. Ecol. Indic. 2021, 130, 108060. [Google Scholar] [CrossRef]
- Maskell, L.C.; Botham, M.; Henrys, P.; Jarvis, S.; Maxwell, D.; Robinson, D.A.; Rowland, C.S.; Siriwardena, G.; Smart, S.; Skates, J.; et al. Exploring relationships between land use intensity, habitat heterogeneity and biodiversity to identify and monitor areas of High Nature Value farming. Biol. Conserv. 2019, 231, 30–38. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.-L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Ali, M.P.; Biswas, M.; Clemente-Orta, G.; Kabir, M.M.M.; Datta, J.; Haque, S.S.; Qin, X.; Landis, D.; Kaur, P.; Pittendrigh, B.R.; et al. Landscape diversity influences the arthropod species diversity in the rice field. Front. Environ. Sci. 2022, 10, 15. [Google Scholar] [CrossRef]
- Holland, J.; Fahrig, L. Effect of woody borders on insect density and diversity in crop fields: A landscape-scale analysis. Agric. Ecosyst. Environ. 2000, 78, 115–122. [Google Scholar] [CrossRef]
- Smart, S.M.; Marrs, R.H.; Le Duc, M.G.; Thompson, K.E.N.; Bunce, R.G.H.; Firbank, L.G.; Rossall, M.J. Spatial relationships between intensive land cover and residual plant species diversity in temperate farmed landscapes. J. Appl. Ecol. 2006, 43, 1128–1137. [Google Scholar] [CrossRef]
- Schindler, S.; von Wehrden, H.; Poirazidis, K.; Wrbka, T.; Kati, V. Multiscale performance of landscape metrics as indicators of species richness of plants, insects and vertebrates. Ecol. Indic. 2013, 31, 41–48. [Google Scholar] [CrossRef]
Figure 1.
Study area location in the Western Po Plain region (north of Italy), and survey site location: site 1—Rovasenda, site 2—Romentino, site 3—Trino Vercellese (in red).
Figure 1.
Study area location in the Western Po Plain region (north of Italy), and survey site location: site 1—Rovasenda, site 2—Romentino, site 3—Trino Vercellese (in red).

Figure 2.
(A) A photo of rice fields of the organic farm Cascina dell’Angelo, Rovasenda, site 1; (B,C) an example of a planting scheme (plan (B); section (C)) of trees and shrubs in between fields and shrubs along in-field linear embankments, site 1.
Figure 2.
(A) A photo of rice fields of the organic farm Cascina dell’Angelo, Rovasenda, site 1; (B,C) an example of a planting scheme (plan (B); section (C)) of trees and shrubs in between fields and shrubs along in-field linear embankments, site 1.

Figure 3.
Life forms spectra (% total species) comparison 1950s versus current state. Abbreviations: Ch = chamaephyte, G = geophyte, H = hemicryptophyte, He/I = hydrophyte/helophyte, P = phanerophyte, T = therophyte.
Figure 3.
Life forms spectra (% total species) comparison 1950s versus current state. Abbreviations: Ch = chamaephyte, G = geophyte, H = hemicryptophyte, He/I = hydrophyte/helophyte, P = phanerophyte, T = therophyte.

Figure 4.
(a) Native and alien species proportion (% of total flora), 1950s versus current state floristic data comparison. (b) Alien species origin (% of total flora), 1950s versus current state floristic data comparison.
Figure 4.
(a) Native and alien species proportion (% of total flora), 1950s versus current state floristic data comparison. (b) Alien species origin (% of total flora), 1950s versus current state floristic data comparison.

Figure 5.
Floristic data: total chorotypes proportions (% of total flora) comparison (1950s versus current state). Abbreviations: Cosmop. = cosmopolitan; Orop. S- Europ. = orophyte South European; Medit. s.l. = Mediterranean.
Figure 5.
Floristic data: total chorotypes proportions (% of total flora) comparison (1950s versus current state). Abbreviations: Cosmop. = cosmopolitan; Orop. S- Europ. = orophyte South European; Medit. s.l. = Mediterranean.

Figure 6.
(A,B) PCA analysis results; (C) dendrogram (cluster analysis). See Table 1 for species and relevés code abbreviations.
Figure 6.
(A,B) PCA analysis results; (C) dendrogram (cluster analysis). See Table 1 for species and relevés code abbreviations.

Figure 7.
Native and alien species proportion (cover-abundance weighted % species). Comparison between: total vegetation; site 1 versus sites 2−3 (a); organic rice fields versus the conventional ones (b).
Figure 7.
Native and alien species proportion (cover-abundance weighted % species). Comparison between: total vegetation; site 1 versus sites 2−3 (a); organic rice fields versus the conventional ones (b).

Figure 8.
α-biodiversity calculated on vegetational data. Comparison between: total sites; site 1 and sites 2−3 (a); organic and conventional rice fields (b).
Figure 8.
α-biodiversity calculated on vegetational data. Comparison between: total sites; site 1 and sites 2−3 (a); organic and conventional rice fields (b).

Figure 9.
Ellenberg-Pignatti ecogram comparison (site 1 and sites 2–3), calculated on vegetational data.
Figure 9.
Ellenberg-Pignatti ecogram comparison (site 1 and sites 2–3), calculated on vegetational data.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Phytosociological table reporting the total 40 relevés (for details regarding date, locality, organic/conventional farm and sporadic species, see Appendix A).
Table 1.
Phytosociological table reporting the total 40 relevés (for details regarding date, locality, organic/conventional farm and sporadic species, see Appendix A).
| SITE 1 (G) | SITES 2 (D)−3 (P) | ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8G | 4G | 9G | 14G | 11G | 15G | 12G | 10G | 13G | 2G | 1G | 3G | 7G | 5G | 6G | 20D | 22D | 26P | 28P | 29P | 24D | 39P | 25P | 40P | 37P | 36P | 38P | 23D | 31P | 33P | 21D | 32P | 16D | 17D | 18D | 19D | 27P | 34P | 30P | 35P | ||||
| Coverage (%) | 95 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 95 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 95 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | pres. | freq. | |
| Area (m2) | 70 | 80 | 70 | 85 | 100 | 90 | 90 | 75 | 90 | 90 | 90 | 80 | 70 | 60 | 80 | 100 | 100 | 80 | 90 | 90 | 100 | 100 | 90 | 100 | 100 | 90 | 95 | 100 | 95 | 85 | 100 | 90 | 100 | 100 | 100 | 100 | 80 | 90 | 90 | 100 | |||
| Species number | 8 | 10 | 7 | 10 | 7 | 10 | 9 | 10 | 10 | 7 | 9 | 12 | 8 | 8 | 9 | 9 | 9 | 8 | 9 | 11 | 5 | 4 | 9 | 7 | 4 | 5 | 4 | 3 | 8 | 7 | 7 | 9 | 11 | 14 | 10 | 10 | 10 | 9 | 7 | 8 | |||
| Os Oryza sativa s.l. | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 4.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 4.4 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 4.4 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 40 | V | |
| Char. and diff. species of Oryzo sativae−Echinocletum cruris galli | |||||||||||||||||||||||||||||||||||||||||||
| Ec Echinocloa crus−galli (L.) P. Beauv. | 1.1 | + | 1.2 | +0.2 | 1.2 | + | − | +0.2 | 1.1 | + | 1.2 | + | +0.2 | 1.2 | − | − | 2.3 | 2.2 | 1.2 | + | 1.1 | + | 1.2 | + | + | + | − | − | +0.2 | + | + | + | − | +0.2 | +0.2 | 1.2 | + | + | 1.2 | + | 34 | V | |
| Ld Lindernia dubia (L.) Pennell | − | − | − | − | − | − | + | + | + | − | + | − | − | 1.1 | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2.2 | + | + | + | − | − | − | − | 10 | II | |
| Char. and diff. species of the upper units (Oryzo−Echinochloion oryzoidis alliance, Cypero−Echinochloetalia oryzoidis order, Oryzetea sativae class) | |||||||||||||||||||||||||||||||||||||||||||
| Sm | Schoenoplectiella mucronata (L.) J. Jung & H.K. Choi | 2.3 | 3.4 | 2.3 | 1.2 | 1.2 | 2.2 | 1.1 | 1.2 | 1.1 | 3.3 | +.2 | 2.3 | 3.3 | 2.3 | 3.4 | + | 1.2 | 3.3 | + | +0.2 | − | − | 2.2 | − | − | 1.1 | + | − | + | + | − | +0.2 | + | 1.1 | − | + | +.2 | 1.2 | 1.1 | 2.2 | 33 | IV |
| Hr | Heteranthera reniformis Ruiz & Pav. | − | 1.2 | − | 1.2 | 3.3 | 2.2 | 2.3 | 2.4 | 3.3 | 1.1 | + | 2.3 | − | − | − | − | − | − | − | + | − | − | 1.1 | − | − | − | − | − | + | − | 3.3 | + | − | + | + | + | 2.3 | 2.3 | 1.2 | 1.3 | 22 | III |
| Av | Ammania verticillata (Ard.) Lam. | − | − | − | + | − | − | − | − | − | − | − | − | − | − | − | + | + | + | − | − | − | − | 1.1 | − | + | 1.2 | + | − | + | 3.3 | 2.3 | 1.2 | 2.2 | 4.4 | 1.2 | 2.3 | 2.2 | 2.3 | 2.2 | 3.3 | 20 | III |
| Ep | Eleocharis flavescens (Poir.) Urb. | +0.2 | 1.2 | + | +0.2 | 1.3 | 1.2 | − | +0.2 | 1.2 | 1.1 | +0.2 | 1.2 | 2.3 | 3.4 | 2.4 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 14 | II |
| Hl | Heteranthera limosa (Sw.) Willd. | + | + | − | +0.2 | − | + | 1.2 | 1.3 | − | − | − | 3.4 | 2.3 | 3.4 | 1.2 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 10 | II |
| Mk | Murdannia keisak (Hassk.) Hand.−Mazz. | − | + | − | 1.1 | − | + | − | + | − | +0.2 | + | 3.4 | − | − | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 8 | I |
| Rd | Rotala densiflora (Roth) Koehne | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + | − | + | 1.1 | − | + | + | 1.2 | + | 1.2 | − | 1.1 | 1.1 | 2.3 | 13 | II |
| Ea | Eleocharis acicularis (L.) Roem. & Schult. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | +.2 | − | − | − | − | − | − | − | − | − | − | 1.2 | + | 1.2 | 1.2 | − | − | − | + | − | − | − | − | − | 7 | I |
| Oss | Oryza sativa L. var. sylvatica Chiappelli | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 1.2 | 2.2 | − | − | +0.2 | − | − | − | − | − | − | − | − | − | − | +0.2 | − | − | +0.2 | − | − | − | 5 | I |
| Others species | |||||||||||||||||||||||||||||||||||||||||||
| Pl | Persicaria lapathifolia (L.) Delarbre | 2.3 | + | 1.1 | − | 2.2 | − | 1.1 | + | +0.2 | + | 2.3 | 2.2 | 1.2 | + | 2.3 | 1.1 | − | 1.1 | − | + | − | + | 1.2 | + | − | − | − | − | − | 1.2 | − | 1.2 | − | − | + | − | + | − | − | − | 23 | III |
| Ap | Alisma plantago−aquatica L. | 1.1 | − | − | − | 1.1 | + | + | 1.2 | 1.1 | − | +0.2 | 1.2 | − | − | − | − | − | 1.1 | + | +0.2 | − | − | + | − | − | − | − | − | − | − | 1.2 | + | 4.4 | 2.3 | 2.2 | 1.2 | +0.2 | + | +0.2 | − | 21 | III |
| Bf | Bidens frondosa L. | + | + | − | + | − | + | + | − | + | − | − | − | − | − | + | + | + | + | 2.2 | − | − | + | − | + | − | − | − | − | − | − | − | − | + | + | + | 1.2 | − | − | − | + | 18 | III |
| Bu | Butomus umbellatus L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 1.2 | 3.2 | − | 1.2 | 1.2 | − | − | − | − | − | − | − | + | + | − | +0.2 | 1.2 | 1.1 | + | − | + | − | − | − | − | 11 | II |
| Lm | Lemna minor L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | +0.2 | 2.3 | − | − | +0.3 | 1.1 | − | − | − | − | − | − | − | − | + | − | − | − | + | 1.1 | − | + | 8 | I |
| Ta | Typha angustifolia L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 1.2 | + | − | − | + | + | − | − | 4 | I |
| Jc | Juncus conglomeratus L. | − | − | − | − | − | − | − | − | 1.2 | − | − | 1.1 | 1.1 | − | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 4 | I |
| Cg | Cyperus glomeratus L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | + | − | − | − | − | − | − | − | − | − | − | − | − | +0.2 | 2.3 | − | − | − | − | − | − | 4 | I |
| Mq | Marsilea quadrifolia L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | − | − | − | − | − | 1.2 | − | 2.3 | 3 | I |
| Tl | Typha latifolia L. | − | − | + | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Er | Elymus repens (L.) Gould | − | + | +0.2 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Vb | Veronica beccabunga L. | − | − | − | + | − | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Cs | Cyperus strigosus L. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | 1.2 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Ce | Calamagrostis epigejos (L.) Roth | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | 1.2 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Pa | Phragmites australis (Cav.) Trin. ex Steud. | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 2 | I |
| Sporadic species | − | − | − | − | − | − | 1 | − | − | − | − | 1 | 1 | 1 | − | − | − | − | − | − | 3 | − | − | 1 | − | − | − | − | − | − | − | − | − | 1 | − | − | − | − | − | − | |||
Table 2.
α-biodiversity calculated on vegetational data and average number of taxa. Comparison between: total sites; site 1 and sites 2−3; organic and conventional rice fields.
Table 2.
α-biodiversity calculated on vegetational data and average number of taxa. Comparison between: total sites; site 1 and sites 2−3; organic and conventional rice fields.
| TOT | Site 1 | Sites 2–3 | Organic | Conventional | |
|---|---|---|---|---|---|
| E—Simpson | 0.058 | 0.119 | 0.075 | 0.061 | 0.059 |
| J—Shannon | 0.402 | 0.463 | 0.392 | 0.416 | 0.182 |
| Average taxa number | 8.275 | 8.933 | 7.880 | 9.135 | 5.778 |
Table 3.
Phytosociological table reporting the 4 relevés related to the 2 experimental plots (site 1) (see Appendix A for further details).
Table 3.
Phytosociological table reporting the 4 relevés related to the 2 experimental plots (site 1) (see Appendix A for further details).
| 1 | 2 | 3 | 4 | |
| Coverage (%) | 100 | 100 | 100 | 100 |
| Area (m2) | 90 | 80 | 80 | 80 |
| Species number | 13 | 10 | 7 | 6 |
| Oryza sativa s.l. | 3.3 | 2.3 | 1.1 | + |
| Murdannia keisak (Hassk.) Hand.−Mazz. | 4.5 | 4.4 | 5.5 | 5.5 |
| Schoenoplectiella mucronata (L.) J. Jung & H.K. Choi | 2.3 | 3.4 | 1.1 | 1.1 |
| Heteranthera reniformis Ruiz & Pav. | 3.3 | 1.2 | +0.2 | 1.2 |
| Eleocharis flavescens (Poir.) Urb. | 1.2 | 2.2 | − | − |
| Echinocloa crus−galli (L.) P. Beauv. | − | +0.2 | − | +0.2 |
| Persicaria lapathifolia (L.) Delarbre | 1.1 | 1.1 | + | − |
| Alisma plantago−aquatica L. | 1.1 | 1.2 | − | − |
| Bidens frondosa L. | 1.2 | + | + | − |
| Typha angustifolia L. | + | − | + | 1.1 |
| Typha latifolia L. | 1.1 | + | − | − |
| Polygonum aviculare L. | 1.1 | − | − | − |
| Robinia pseudoacacia L. | (+) | − | − | − |
| Salix alba L. | (+) | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vagge, I.; Chiaffarelli, G. The Alien Plant Species Impact in Rice Crops in Northwestern Italy. Plants 2023, 12, 2012. https://doi.org/10.3390/plants12102012
AMA Style
Vagge I, Chiaffarelli G. The Alien Plant Species Impact in Rice Crops in Northwestern Italy. Plants. 2023; 12(10):2012. https://doi.org/10.3390/plants12102012
Chicago/Turabian StyleVagge, Ilda, and Gemma Chiaffarelli. 2023. "The Alien Plant Species Impact in Rice Crops in Northwestern Italy" Plants 12, no. 10: 2012. https://doi.org/10.3390/plants12102012
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.