

Higher Virulence of Diplodia seriata Isolates on Vines of cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
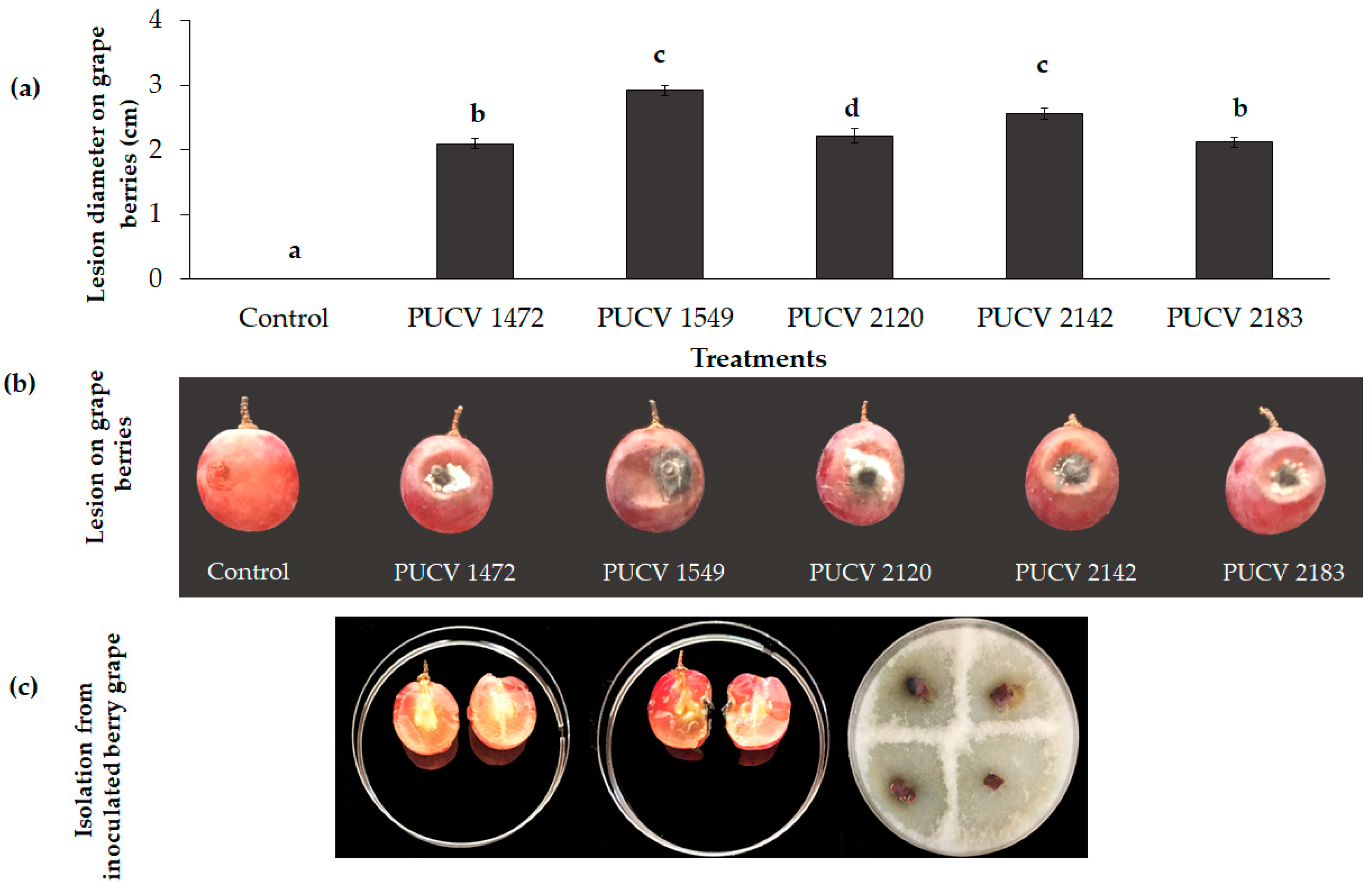
2.1. Pathogenicity Tests on Berry Grapes
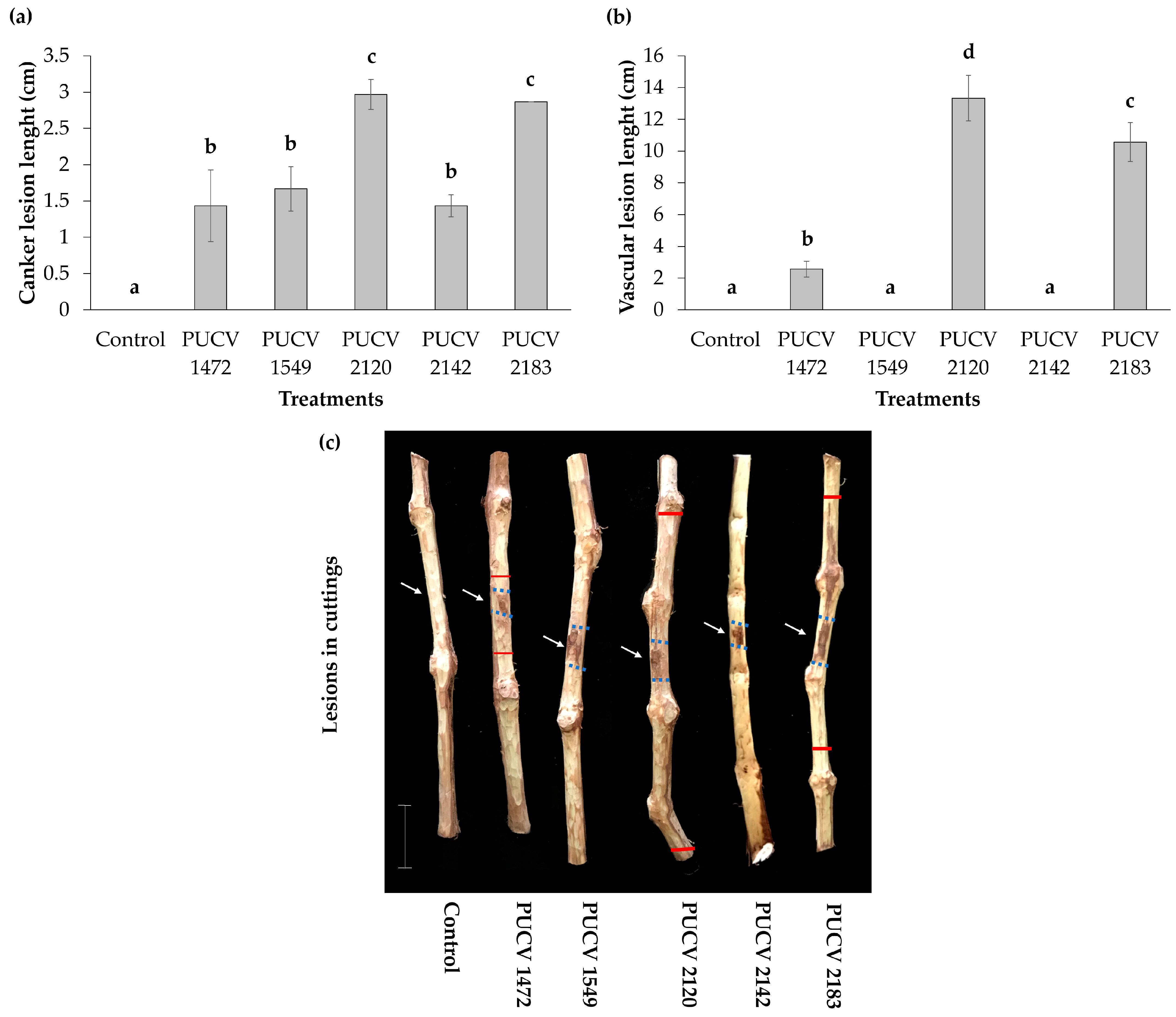
2.2. Pathogenicity Tests in Detached Canes (Activated Isolates)
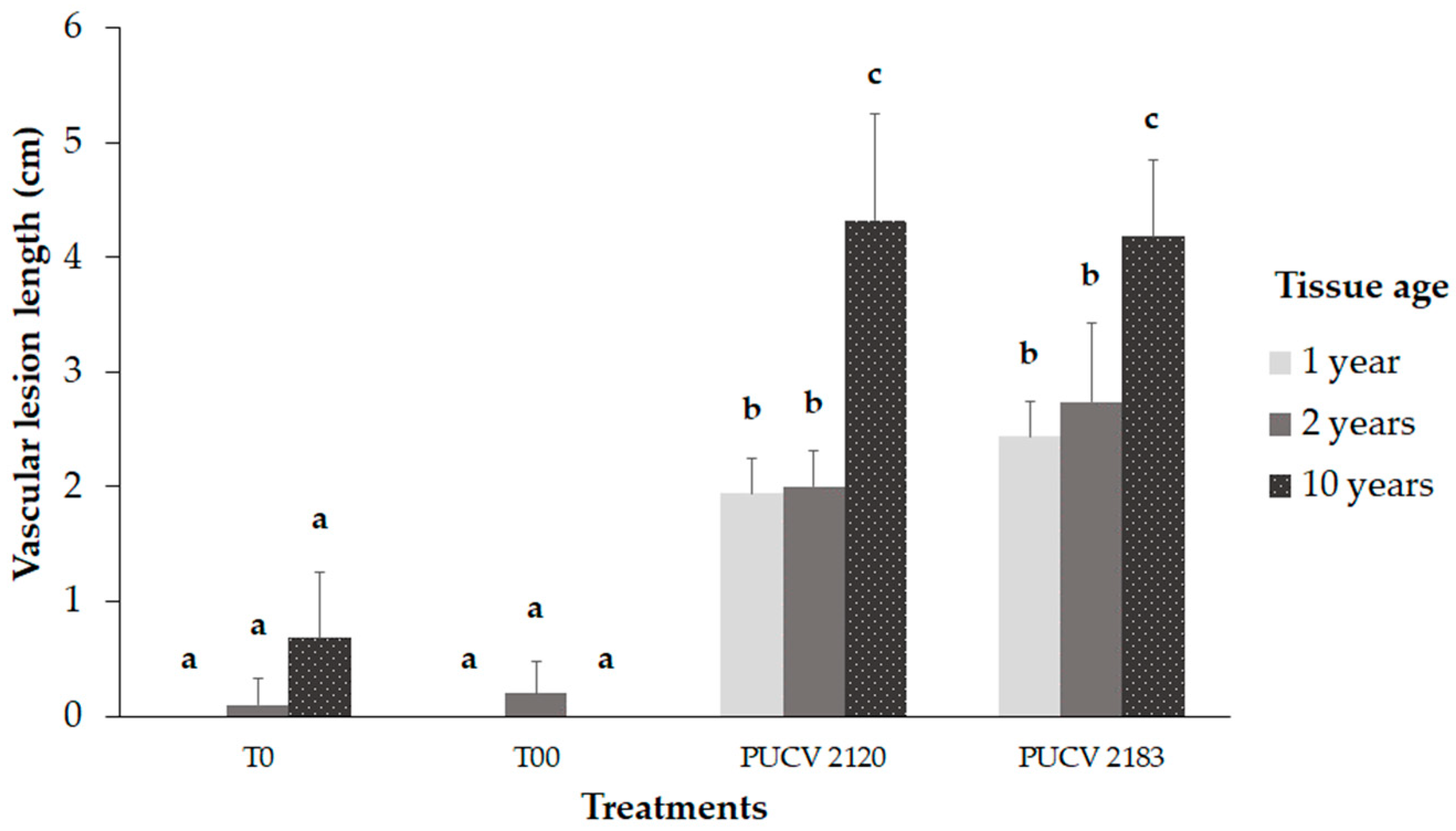
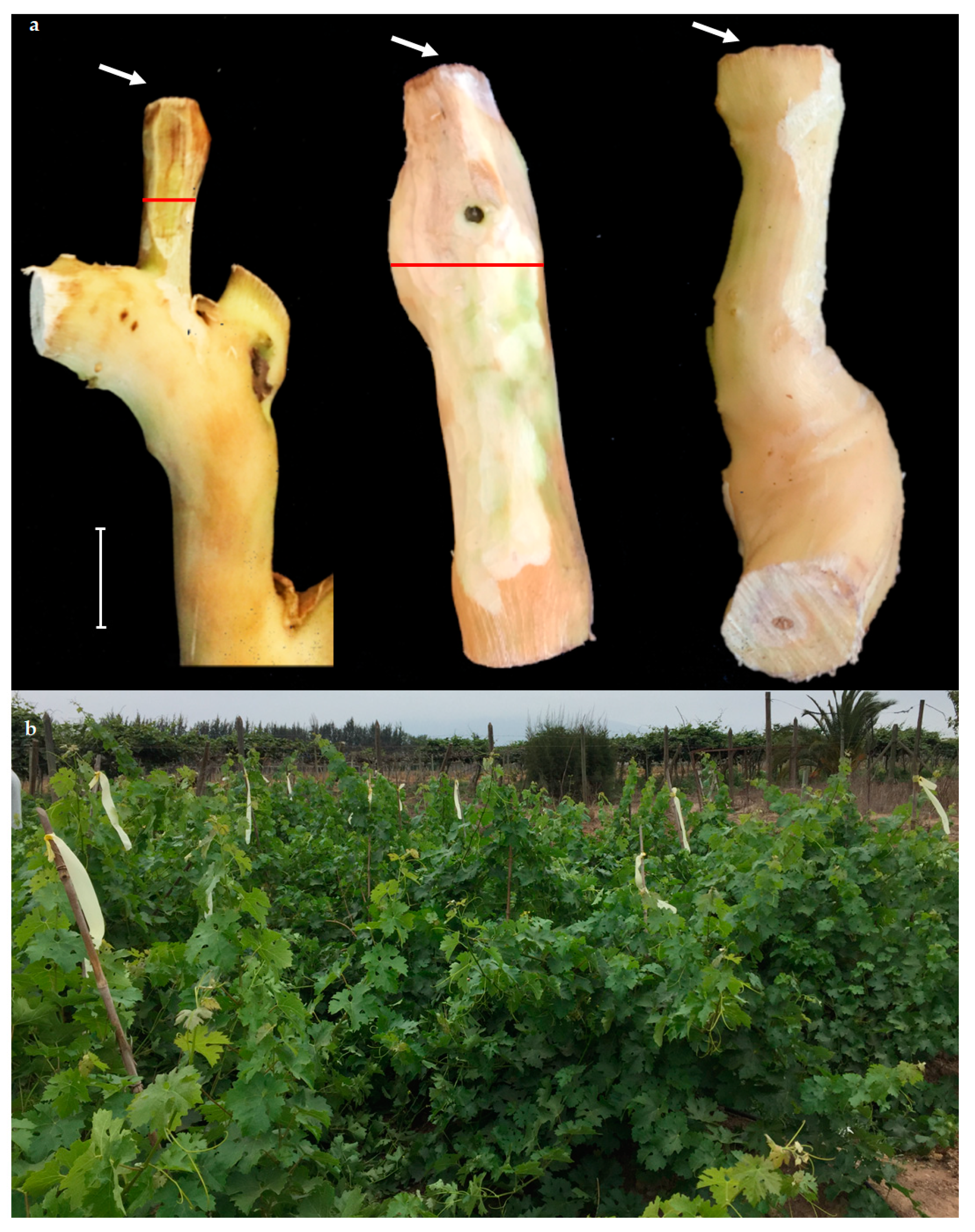
2.3. Pathogenicity Trial in the Field
2.4. Maximum Parsimony Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents, and Culture Media
4.2. Fungal Isolates
4.3. Pathogenicity Tests on Berry Grapes
4.3.1. Fungal Isolates and Inoculation
4.3.2. Damage Assessment and Pathogen Recovery
4.4. Pathogenicity Tests on Detached Canes
Damage Assessment and Pathogen Recovery
4.5. Pathogenicity Trial in the Field
4.5.1. Fungal Isolates and Inoculation
4.5.2. Damage Assessment and Pathogen Recovery
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.; Da Costa, J.; Guerin-Dubrana, L.; et al. Overview of grapevine trunk diseases in France in the 2000s. Phytopathol. Mediterr. 2013, 52, 262–275. [Google Scholar]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Catarina, A.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2016, 114, 707–721. [Google Scholar] [CrossRef]
- Hillis, V.; Lubell, M.; Kaplan, J.; Doll, D.; Baumgartner, K. The role of pest control advisers in preventative management of grapevine trunk diseases. Phytopathology 2016, 106, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.; Travadon, R.; Cooper, M.; Hillis, V.; Lubell, M.; Baumgartner, K. Identifying Economic Hurdles to Early Adoption of Preventative Practices: The Case of Trunk Diseases in California Winegrape Vineyards. Wine Econ. Policy 2016, 5, 127–141. [Google Scholar] [CrossRef]
- Larach, A.; Torres, C.; Riquelme, N.; Valenzuela, M.; Salgado, E.; Seeger, M.; Besoain, X. Yield loss estimation and pathogen identification from (BD) in vineyards of Central Chile over two growing seasons. Phytopathol. Mediterr. 2020, 59, 537–548. [Google Scholar]
- Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.; El Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlli, R.; et al. A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, (BD), and Esca Disease. J. Fungi. 2022, 8, 595. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Gubler, W.D. Susceptibility of grapevine pruning wounds to infection by Lasiodiplodia theobromae and Neofusicoccum parvum. Plant Pathol. 2011, 60, 261–270. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Battany, M.; Bettiga, L.J.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R.; Verdegaal, P.; Gubler, W. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant Dis. 2010, 94, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Besoain, X. Grapevine Trunk Diseases (GTDs): Impact on Table Grapes and Wine Vineyards in Chile. In: Grapes and Wines-Advances in Production, Processing, Analysis and Valorization. InTechOpen 2018, 3, 43–58. [Google Scholar]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and Future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Savocchia, S. A review of Botryosphaeriaceae species associated with grapevine trunk diseases in Australia and New Zealand. Aust. Plant Pathol. 2019, 48, 3–18. [Google Scholar] [CrossRef]
- Rolshausen, P.; Baumgartner, K.; Travador, R.; Funjiyoshi, P.; Pouzoulet, J.; Wilcox, W. Identification of Eutypa spp. Causing Eutypa Dieback o Grapevine in Eastern North America. Plant Dis. 2014, 98, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Burruano, S.; Mondello, V.; Conigliario, G.; Alfonzo, A.; Spagnolo, A.; Mugnai, L. Grapevine decline in Italy caused by Lasiodiplodia theobromae. Phytopathol. Mediterr. 2008, 47, 132–136. [Google Scholar]
- Reis, P.; Magnin-Robert, M.; Nascimento, T.; Spagnolo, A.; Abou-Mansour, E.; Fioretti, C.; Clément, C.; Rego, C.; Fontaine, F. Reproducing (BD) foliar symptoms in a simple model system. Plant Dis. 2016, 100, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Latore, B.; Piontelli, E.; Besoain, X. Botryosphaeriaceae Species affecting table grape vineyards in Chile and cultivar susceptibility. Cien Investig. Agrar. 2012, 39, 445. [Google Scholar] [CrossRef]
- Díaz, G.A.; Auger, J.; Besoain, X.; Bordeu, E.; Latorre, B.A. Prevalence and pathogenicity of fungi associated with grapevine trunk diseases in Chilean vineyards. Cienc. Investig. Agrar. 2013, 40, 327–339. [Google Scholar] [CrossRef]
- Savocchia, S.; Steel, C.C.; Stodart, B.J.; Somers, A. Pathogenicity of Botryosphaeria species isolated from declining grapevines in sub-tropical regions of Eastern Australia. VITIS-J. Grapevine Res. 2007, 46, 27–32. [Google Scholar]
- Burgess, T.; Wingfield, M.J. Quarantine is important in restricting the spread of exotic seed-borne tree pathogens in the southern hemisphere. Int. For. Rev. 2002, 4, 56. [Google Scholar]
- Mohali, S.; Burgess, T.I.; Wingfield, M.J. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. For. Pathol. 2005, 35, 385–396. [Google Scholar] [CrossRef]
- Burgess, T.I.; Barber, P.A.; Mohali, S.; Pegg, G.; De Beer, W.; Wingfield, M.J. Three new Lasiodiplodia spp. from the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 2006, 98, 423–435. [Google Scholar] [CrossRef]
- Kuntzmann, P.; Villaume, S.; Larignon, P.; Bertsch, C. Esca, BDA and Eutypiosis: Foliar symptoms, trunk lesions and fungi observed in diseased vinestocks in two vineyards in Alsace. Vitis 2010, 49, 71–76. [Google Scholar]
- Valencia, D.; Torres, C.; Camps, R.; López, E.; Celis-Diez, J.L.; Besoain, X. Dissemination of Botryosphaeriaceae conidia in vineyards in the semiarid Mediterranean climate of the Valparaíso Region of Chile. Phytopathol. Mediterr. 2015, 54, 394–402. [Google Scholar]
- Burgess, T.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.; Shuttleworth, L.; Daniel, R.; Dann, E.; Parkinson, L.; Dihn, Q.; et al. Current status of the Botryosphaeriaceae in Australia. Aust. Plant Pathol. 2018, 48, 35–44. [Google Scholar] [CrossRef]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Lourenco, I.; Saramago, J.; Mota, M.; Rego, C. Early season symptoms on stem, inflorescences and flowers of grapevine associated with Botryosphaeriaceae species. Plants 2020, 11, 1427. [Google Scholar] [CrossRef]
- Castillo-Pando, M.; Somers, A.; Green, C.D.; Priest, M.; Sriskanthades, M. Fungi associated with dieback of Semillon grapevines in the Hunter Valley of New South Wales. Aust. Plant Pathol. 2001, 30, 59–63. [Google Scholar] [CrossRef]
- Pitt, W.M.; Huang, R.; Steel, C.C.; Savocchia, S. Identification, distribution and current taxonomy of Botryosphaeriaceae species associated with grapevine decline in New South Wales and South Australia. Aust. J. Grape Wine Res. 2010, 16, 258–271. [Google Scholar] [CrossRef]
- Qiu, Y.; Steel, C.C.; Ash, G.J.; Savocchia, S. Survey of Botryosphaeriaceae associated with grapevine decline in the Hunter Valley and Mudgee grape growing regions of New South Wales. Aust. Plant Pathol. 2011, 40, 1–11. [Google Scholar] [CrossRef]
- Taylor, A.; Hardy, G.E.S.J.; Wood, P.; Burgess, T. Identification and pathogenicity of Botryosphaeria species associated with grapevine decline in Western Australia. Aust. Plant Pathol. 2005, 34, 187–195. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Voegel, T.M.; Gubler, W.D. Identification and distribution of Botryosphaeria spp. associated with grapevine cankers in California. Plant Dis. 2006, 90, 1490–1503. [Google Scholar] [CrossRef]
- Larignon, P.; Fulchic, R.; Cere, L.; Dubos, B. Observation on black arm in French vineyards. Phytopathol. Mediterr. 2001, 40, 336–342. [Google Scholar]
- Van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA Phylogeny, Morphology and Pathogenicity of Botryosphaeria species on Grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef] [PubMed]
- Baskarathevan, J.; Jaspers, M.; Jones, E.E.; Ridgway, H.J. Incidence and distribution of botryosphaeriaceous species in New Zealand vineyards. Eur. J. Plant Pathol. 2012, 132, 549–560. [Google Scholar] [CrossRef]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, A.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef]
- Spagnolo, A.; Larignon, P.; Magnin-Robert, M.; Hovasse, A.; Cilindre, C.; Van Dorsselaer, A.; Clément, C.; Schaeffer-Reiss, C.; Fontaine, F. Flowering as the most highly sensitive period of grapevine (Vitis vinifera L. cv Mourvédre) to the (BD) agents Neofusicoccum parvum and Diplodia seriata infection. Int. J. Mol. Sci. 2014, 15, 9644–9669. [Google Scholar] [CrossRef] [PubMed]
- Bénard-Gellon, M.; Farine, S.; Goddard, M.L.; Schmitt, M.; Stempien, E.; Pensec, F.; Laloue, H.; Mazet-Kieffer, F.; Fontaine, F.; Larignon, P.; et al. Toxicity of extracellular proteins from Diplodia seriata and Neofusicoccum parvum involved in grapevine Botryosphaeria dieback. Protoplasma 2015, 252, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Morales-Cruz, A.; Amrine, K.C.H.; Blanco-Ulate, B.; Lawrence, D.P.; Travadon, R.; Rolshausen, R.P.; Baumgartner, K.; Cantu, D. Distinctive expansion of gene families associated with plant cell wall degradation, secondary metabolism, and nutrient uptake in the genomes of grapevine trunk pathogens. BMC Genom. 2015, 16, 469. [Google Scholar] [CrossRef]
- ODEPA. Oficina de Estudios y Políticas Agrarias. Statistics Productive. 2022. Available online: https://www.odepa.cl (accessed on 20 February 2022).
- Luque, J.; Elena, G.; Garcia-Figueres, F.; Reyes, J.; Barrios, G.; Legorburu, F.J. Natural infections of pruning wounds by fungal trunk pathogens in mature grapevines in Catalonia (Northeast Spain). Aust. J. Grape Wine Res. 2014, 20, 134–143. [Google Scholar] [CrossRef]
- Luque, J.; Martos, S.; Aroca, A.; Raposo, R.; Garcia-Figueres, F. Symptoms and fungi associated with declining mature grapevine plants in northeast spain. Plant Pathol. 2009, 91, 381–390. [Google Scholar]
- Bellée, A.; Comont, G.; Nivault, A.; Abou-Mansour, E.; Coppin, C.; Dufour, M.C.; Corio-Costet, M. Life traits of four Botryosphaeriaceae species and molecular responses of different grapevine cultivars or hybrids. Plant Pathol. 2017, 66, 763–776. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Guerrero, J.C.; Guevara, J.; Gubler, W.D. Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of Grapevines in Mexico. Plant Dis. 2008, 92, 519–529. [Google Scholar] [CrossRef]
- Sosnowski, M.R.; Ayres, M.R.; McCarthy, M.G.; Scott, E.S. Winegrape cultivars (Vitis vinifera) vary in susceptibility to the grapevine trunk pathogens Eutypa lata and Diplodia seriata. Aust. J. Grape Wine 2022, 28, 166–174. [Google Scholar] [CrossRef]
- Van de Wouw, A.P.; Howlett, B.J. Fungal pathogenicity genes in the age of “omics”. Mol. Plant Pathol. 2011, 12, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Elena, G.; Garcia-Figueres, F.; Reigada, S.; Luque, J. Intraspecific variation in Diplodia seriata isolates occurring on grapevines in Spain. Plant Pathol. 2015, 64, 680–689. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gerbore, J.; Lecomte, P.; Da Costa, J.P.; Guerin-Dubrana, L.; Rey, P. Analyses of the temporal dynamics of fungal communities colonizing the healthy wood tissues of esca leaf-symptomatic and asymptomatic vines. PLoS ONE 2014, 9, e95928. [Google Scholar] [CrossRef]
- Boddy, L.; Rayner, A. Origins of decay in living deciduous trees: The role of moisture content and a re-appraisal of the expanded concept of tree decay. New Phytol. 1983, 94, 623–641. [Google Scholar] [CrossRef]
- Rey, P.; Lecomte, P.; Guérin-Dubrana, L.; Corio-Costel, F.; Fonteine, F.; Gomes, E.; Goutouly, J.P.; Hofdtetter, V.; Legorburu, X.; De La Rocque, B. Maladies du bois de la vigne situation enjeux et perpective. Phytoma 2011, 640, 22–26. [Google Scholar]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef]
- Andreolli, M.; Lampis, S.; Zapparoli, G.; Angelini, E.; Vallini, G. Diversity of bacterial endophytes in 3- and 15-year-old grapevines of Vitis vinifera cv. Corvina and their potential for plant growth promotion and phytopathogen control. Microbiological 2016, 183, 42–52. [Google Scholar] [CrossRef]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wery, J. Current knowledge on Grapevine Trunk Diseases with complex etiology: A systemic approach. Phytopathol. Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, T.J. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathol 2010, 100, 1340–1351. [Google Scholar] [CrossRef]
- Amalfitano, C.; Evidente, A.; Surico, G.; Tegli, S.; Bertelli, E.; Mugnai, L. Phenols and Stilbene Polyphenols in the Wood of Esca-Diseased Grapevines. Phytopathol. Mediterr. 2000, 39, 178–183. [Google Scholar]
- Del Rio, J.; Gonzalez, A.; Fuster, M.; Botia, J.; Gomez, P.; Frias, V.; Ortuño, A. Tylose formation and changes in phenolic compounds of grape roots infected with Phaeomoniella chlamydospore and Phaeocremonium species. Phytopathol. Mediterr. 2001, 40, 394–399. [Google Scholar]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L.: I. Characterization of secondary metabolites in culture media and host responses to the pathogens in calli. Physiol. Mol. Plant Pathol. 2006, 69, 209–223. [Google Scholar] [CrossRef]
- Martin, N.; Vesentini, D.; Rego, C.; Monteiro, S.; Oliveira, H.; Ferreira, R.B. Phaeomoniella chlamydospora infection induces changes in phenolic compounds content in Vitis vinifera. Phytopathol. Mediterr. 2009, 48, 101–116. [Google Scholar]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Clément, C.; Larignon, P.; Ramirez-Suero, M.; Chong, J.; et al. Differential responses of three grapevine cultivars to (BD). Phytopathol. 2014, 104, 1021–1031. [Google Scholar] [CrossRef]
- Gómez, P.; Báidez, A.G.; Ortuño, A.; Del Río, J.A. Grapevine xylem response to fungi involved in trunk diseases. Ann. Appl. Biol. 2016, 169, 116–124. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Pivovaroff, A.L.; Santiago, L.S.; Rolshausen, P.E. Can vessel dimension explain tolerance toward fungal vascular wilt diseases in woody plants? Lessons from Dutch elm disease and esca disease in grapevine. Front. Plant Sci. 2014, 5, 253. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Cimmino, A.; Evidente, A. Phytotoxic metabolites by nine species of Botryosphaeriaceae involved in grapevine dieback in Australia and identification of those produced by Diplodia mutila, Diplodia seriata, Neofusicoccum australe and Neofusicoccum luteum. Nat. Prod. Res. 2019, 33, 2223–2229. [Google Scholar] [CrossRef]
- Martos, S.; Andolfi, A.; Luque, J.; Mugnai, L.; Surico, G.; Evidente, A. Production of phytotoxic metabolites by five species of Botryosphaeriaceae causing decline on grapevines, with special interest in the species Neofusicoccum luteum and N. parvum. Eur. J. Plant Pathol. 2008, 121, 451–461. [Google Scholar] [CrossRef]
- Zarzycka, H. Effect of culturing Phytophthora infestans on various substrates on expression of its virulence and aggressiveness to potato. Phytopathol. Polonica. 1995, 9, 81–88. [Google Scholar]
- Zarzycka, H. Effect of various methods of maintenance of Phytophthora infestans on its pathogenicity to potato. Plant Breed. Seed Sci. 1996, 40, 31–43. [Google Scholar]
- Balendres, M.A.; De Torres, R.; Dela Cueva, F. Culture storage age and fungal re-isolation from host-tissue influence Colletotrichum spp. virulence to pepper fruits. J. Phytopathol. 2019, 167, 510–515. [Google Scholar] [CrossRef]
- Pearce, R.B. Antimicrobial defenses in the wood of living trees. New Phytol. 1996, 132, 203–233. [Google Scholar] [CrossRef]
- Ramírez, M.; Pérez, L.M.; Montealegre, J.R. Susceptibility of different grapevine (Vitis vinifera L.) cultivars to Diplodia seriata and Diplodia mutila. Cien. Inv. Agric. 2018, 45, 93–98. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Scudiero, E.; Schiavon, M.; Rolshausen, P.E. Xylem Vessel Diameter Affects the Compartmentalization of the Vascular Pathogen Phaeomoniella chlamydospora in Grapevine. Front. Plant Sci. 2017, 21, 1442. [Google Scholar] [CrossRef]
- Amponsah, N.T.; Jones, E.E.; Ridgway, H.J.; Jaspers, M. Infection and disease progression of Neofusicoccum luteum in grapevine plants. Plant Health Manag. An. Integr. Approach 2009, 118. [Google Scholar]
- Úrbez-Torres, J.R.; Bruez, E.; Hurtado, J.; Gubler, W.D. Effect of temperature on conidial germination of Botryosphaeriaceae species infecting grapevines. Plant Dis. 2010, 94, 1476–1484. [Google Scholar] [CrossRef]
- Koichiro, T.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate of Diplodia seriata * | Year of Collection | Locality Origin, Region of Chile | Access No. GenBank | ||
|---|---|---|---|---|---|
| ITS | BT | EF1-α | |||
| PUCV 1472 | 2010 | Palmilla, O’Higgins | KM372581 | KP762454 | - |
| PUCV 1549 | 2010 | Peralillo, O’Higgins | KM580514 | KP762464 | - |
| PUCV 2120 | 2018 | Palmilla, O’Higgins | MT023573 | MT063140 | MT120827 |
| PUCV 2142 | 2018 | Batuco, Maule | MT023574 | MT063141 | MT120827 |
| PUCV 2183 | 2018 | Pencahue, Maule | MT023587 | MT063154 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larach, A.; Vega-Celedón, P.; Salgado, E.; Salinas, A.; Riquelme, N.; Castillo-Novales, D.; Sanhueza, P.; Seeger, M.; Besoain, X. Higher Virulence of Diplodia seriata Isolates on Vines of cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue. Plants 2023, 12, 2984. https://doi.org/10.3390/plants12162984
Larach A, Vega-Celedón P, Salgado E, Salinas A, Riquelme N, Castillo-Novales D, Sanhueza P, Seeger M, Besoain X. Higher Virulence of Diplodia seriata Isolates on Vines of cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue. Plants. 2023; 12(16):2984. https://doi.org/10.3390/plants12162984
Chicago/Turabian StyleLarach, Alejandra, Paulina Vega-Celedón, Eduardo Salgado, Aldo Salinas, Natalia Riquelme, Diyanira Castillo-Novales, Paulina Sanhueza, Michael Seeger, and Ximena Besoain. 2023. "Higher Virulence of Diplodia seriata Isolates on Vines of cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue" Plants 12, no. 16: 2984. https://doi.org/10.3390/plants12162984