
Phylogenomics of Southern European Taxa in the Ranunculus auricomus Species Complex: The Apple Doesn’t Fall Far from the Tree
 ,
,  ,
,
Abstract
:
1. Introduction
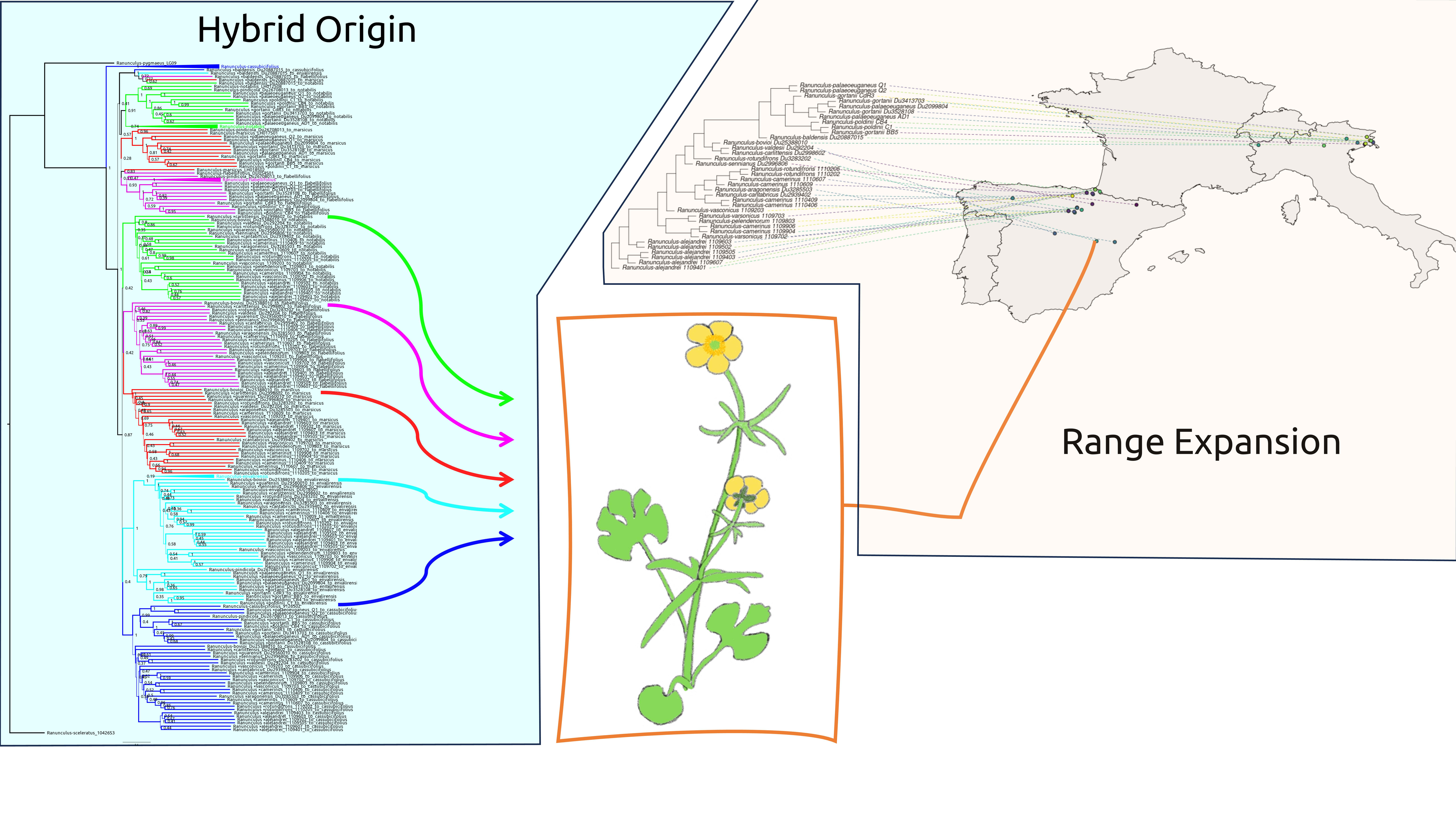
2. Results
2.1. Somatic Ploidy and Reproductive Mode
2.1.1. Somatic Ploidy
2.1.2. Reproductive Mode
2.2. Coalescent-Based Species Tree Estimation
2.3. Phasing of Suspected Nothotaxa
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Somatic Ploidy and Reproductive Mode Determination
4.3. DNA Extraction and Target Enrichment
4.4. Phylogenomic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savolainen, V.; Chase, M.W. A Decade of Progress in Plant Molecular Phylogenetics. Trends Genet. 2003, 19, 717–724. [Google Scholar] [CrossRef] [PubMed]
- The Angiosperm Phylogeny Group. An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Stull, G.W.; Pham, K.K.; Soltis, P.S.; Soltis, D.E. Deep Reticulation: The Long Legacy of Hybridization in Vascular Plant Evolution. Plant J. 2023, 114, 743–766. [Google Scholar] [CrossRef] [PubMed]
- Ott, T.; Schall, M.; Vogt, R.; Oberprieler, C. The Warps and Wefts of a Polyploidy Complex: Integrative Species Delimitation of the Diploid Leucanthemum (Compositae, Anthemideae) Representatives. Plants 2022, 11, 1878. [Google Scholar] [CrossRef] [PubMed]
- Folk, R.A.; Visger, C.J.; Soltis, P.S.; Soltis, D.E.; Guralnick, R.P. Geographic Range Dynamics Drove Ancient Hybridization in a Lineage of Angiosperms. Am. Nat. 2018, 192, 171–187. [Google Scholar] [CrossRef]
- Dayrat, B. Towards Integrative Taxonomy: INTEGRATIVE TAXONOMY. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef]
- Melichárková, A.; Šlenker, M.; Zozomová-Lihová, J.; Skokanová, K.; Šingliarová, B.; Kačmárová, T.; Caboňová, M.; Kempa, M.; Šrámková, G.; Mandáková, T.; et al. So Closely Related and Yet So Different: Strong Contrasts Between the Evolutionary Histories of Species of the Cardamine Pratensis Polyploid Complex in Central Europe. Front. Plant Sci. 2020, 11, 588856. [Google Scholar] [CrossRef]
- Abukrees, F.; Kozlowski, G.; Meade, C. Characterization of Diverse Ploidy in the Arctic-alpine Arenaria ciliata Species Complex (Caryophyllaceae) Using Shoot Meristem Staining and Flow Cytometry Analysis of Archived Frozen Tissue. Plant Species Biol. 2018, 33, 144–152. [Google Scholar] [CrossRef]
- Lo, E.Y.Y.; Stefanović, S.; Dickinson, T.A. Geographical Parthenogenesis in Pacific Northwest Hawthorns (Crataegus; Rosaceae). Botany 2013, 91, 107–116. [Google Scholar] [CrossRef]
- Rutherford, S. Insights into Speciation and Species Delimitation of Closely Related Eucalypts Using an Interdisciplinary Approach. Aust. Syst. Bot. 2020, 33, 110. [Google Scholar] [CrossRef]
- Zozomová-Lihová, J.; Melichárková, A.; Svitok, M.; Španiel, S. Pleistocene Range Disruption and Postglacial Expansion with Secondary Contacts Explain the Genetic and Cytotype Structure in the Western Balkan Endemic Alyssum austrodalmaticum (Brassicaceae). Plant Syst. Evol. 2020, 306, 47. [Google Scholar] [CrossRef]
- Hörandl, E. Novel Approaches for Species Concepts and Delimitation in Polyploids and Hybrids. Plants 2022, 11, 204. [Google Scholar] [CrossRef]
- Oxelman, B.; Brysting, A.K.; Jones, G.R.; Marcussen, T.; Oberprieler, C.; Pfeil, B.E. Phylogenetics of Allopolyploids. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 543–557. [Google Scholar] [CrossRef]
- Freudenstein, J.V.; Broe, M.B.; Folk, R.A.; Sinn, B.T. Biodiversity and the Species Concept—Lineages Are Not Enough. Syst. Biol. 2016, 66, 644–656. [Google Scholar] [CrossRef]
- Hörandl, E. The Classification of Asexual Organisms: Old Myths, New Facts, and a Novel Pluralistic Approach. Taxon 2018, 67, 1066–1081. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and Speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef]
- Hörandl, E.; Temsch, E.M. Introgression of Apomixis into Sexual Species Is Inhibited by Mentor Effects and Ploidy Barriers in the Ranunculus auricomus Complex. Ann. Bot. 2009, 104, 81–89. [Google Scholar] [CrossRef]
- Hühn, P.; Dillenberger, M.S.; Krause, S.; Kadereit, J.W. Polyploid Hybrid Speciation in the Calcarata Species Complex of Viola Section Melanium (Violaceae): Relating Hybrid Species to Parent Species Distribution and Ecology. Bot. J. Linn. Soc. 2023, 201, 309–328. [Google Scholar] [CrossRef]
- Landis, J.B.; Soltis, D.E.; Li, Z.; Marx, H.E.; Barker, M.S.; Tank, D.C.; Soltis, P.S. Impact of Whole-genome Duplication Events on Diversification Rates in Angiosperms. Am. J. Bot. 2018, 105, 348–363. [Google Scholar] [CrossRef]
- Levin, D.A. Polyploidy and Novelty in Flowering Plants. Am. Nat. 1983, 122, 1–25. [Google Scholar] [CrossRef]
- Schneider, D.J.; Levin, R.A.; Miller, J.S. Reproductive Isolation between Diploid and Tetraploid Individuals in Mixed-cytotype Populations of Lycium austral. Am. J. Bot. 2023, 110, e16133. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Schiavinato, M.; Bodrug-Schepers, A.; Dohm, J.C.; Himmelbauer, H. Subgenome Evolution in Allotetraploid Plants. Plant J. 2021, 106, 672–688. [Google Scholar] [CrossRef] [PubMed]
- Asker, S.; Jerling, L. Apomixis in Plants; CRC Press: Boca Raton, FL, USA, 1992; ISBN 978-0-8493-4545-6. [Google Scholar]
- Grant, V. Plant Speciation; Columbia University Press: Columbia, WA, USA, 1981; ISBN 978-0-231-88811-0. [Google Scholar]
- Bierzychudek, P. Patterns in Plant Parthenogenesis. Experientia 1985, 41, 1255–1264. [Google Scholar] [CrossRef]
- Hörandl, E. Geographical Parthenogenesis in Alpine and Arctic Plants. Plants 2023, 12, 844. [Google Scholar] [CrossRef]
- Karbstein, K.; Rahmsdorf, E.; Tomasello, S.; Hodač, L.; Hörandl, E. Breeding System of Diploid Sexuals within the Ranunculus auricomus Complex and Its Role in a Geographical Parthenogenesis Scenario. Ecol. Evol. 2020, 10, 14435–14450. [Google Scholar] [CrossRef]
- Hörandl, E. Species Concepts in Agamic Complexes: Applications in the Ranunculus auricomus Complex and General Perspectives. Folia Geobot. 1998, 33, 335–348. [Google Scholar] [CrossRef]
- Hörandl, E. Evolutionary Implications of Self-Compatibility and Reproductive Fitness in the Apomictic Ranunculus auricomus Polyploid Complex (Ranunculaceae). Int. J. Plant Sci. 2008, 169, 1219–1228. [Google Scholar] [CrossRef]
- Paule, J.; Dunkel, F.G.; Schmidt, M.; Gregor, T. Climatic Differentiation in Polyploid Apomictic Ranunculus auricomus Complex in Europe. BMC Ecol. 2018, 18, 16. [Google Scholar] [CrossRef]
- Nogler, G.A. Genetics of Apospory in Apomictic Ranunculus auricomus. V: Conclusion. Bot. Helvetica 1984, 94, 411–422. [Google Scholar]
- Hojsgaard, D.; Greilhuber, J.; Pellino, M.; Paun, O.; Sharbel, T.F.; Hörandl, E. Emergence of Apospory and Bypass of Meiosis via Apomixis after Sexual Hybridisation and Polyploidisation. New Phytol. 2014, 204, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Barke, B.H.; Daubert, M.; Hörandl, E. Establishment of Apomixis in Diploid F2 Hybrids and Inheritance of Apospory From F1 to F2 Hybrids of the Ranunculus auricomus Complex. Front. Plant Sci. 2018, 9, 1111. [Google Scholar] [CrossRef]
- Jonsell, B. Ranunculaceae. In Flora Nordica; Bergius Foundation: Stockholm, Sweden, 2001; Volume 2, pp. 227–334. ISBN 9171900373. [Google Scholar]
- Fagerström, L.; Kvist, G. Vier Neue Arktische Und Subarktische Ranunculus auricomus-Sippen. Ann. Bot. Fenn. 1983, 20, 237–243. [Google Scholar]
- Parker, C. Floristic Inventory of Selected Sites in the Nulato Hills, Western Alaska; Bureau of Land Management-Anchorage Field Office: Anchorage, AK, USA, 1999. [Google Scholar]
- Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 8. Nymphaeaceae to Ranunculaceae; Jalas, J., Suominen, J., Eds.; The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helsinki, Finland, 1989; ISBN 951-9108-07-6. [Google Scholar]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Dunkel, F.G.; Daubert, M.; Hörandl, E. Phylogenomics Supported by Geometric Morphometrics Reveals Delimitation of Sexual Species within the Polyploid Apomictic Ranunculus auricomus Complex (Ranunculaceae). Taxon 2020, 69, 1191–1220. [Google Scholar] [CrossRef]
- Marklund, G. Der Ranunculus Auricomus-Komplex in Finnland, Volume 1; Tilgman: Helsinki, Finland, 1961. [Google Scholar]
- Marklund, G. Der Ranunculus Auricomus-Komplex in Finnland, Volume 2; Tilgman: Helsinki, Finland, 1961. [Google Scholar]
- Ericsson, S. The Microspecies of the Ranunculus auricomus Complex Treated at the Species Level. Ann. Bot. Fenn. 1992, 29, 123–158. [Google Scholar]
- Raab-Straube, E. Ranunculaceae. In Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity; Botanic Garden and Botanical Museum Berlin-Dahlem: Berlin, Germany, 2015. [Google Scholar]
- Tomasello, S.; Karbstein, K.; Hodač, L.; Paetzold, C.; Hörandl, E. Phylogenomics Unravels Quaternary Vicariance and Allopatric Speciation Patterns in Temperate-montane Plant Species: A Case Study on the Ranunculus auricomus Species Complex. Mol. Ecol. 2020, 29, 2031–2049. [Google Scholar] [CrossRef]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Lorberg, E.; Daubert, M.; Hörandl, E. Moving beyond Assumptions: Polyploidy and Environmental Effects Explain a Geographical Parthenogenesis Scenario in European Plants. Mol. Ecol. 2021, 30, 2659–2675. [Google Scholar] [CrossRef] [PubMed]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Wagner, N.; Marinček, P.; Barke, B.H.; Paetzold, C.; Hörandl, E. Untying Gordian Knots: Unraveling Reticulate Polyploid Plant Evolution by Genomic Data Using the Large Ranunculus auricomus Species Complex. New Phytol. 2022, 235, 2081–2098. [Google Scholar] [CrossRef]
- Dunkel, F.G. Contribution to the Knowledge of the Ranunculus auricomus Complex (Ranunculaceae) in Spain. Stapfia 2021, 112, 5–59. [Google Scholar]
- Dunkel, F.G. The Ranunculus auricomus L. Complex (Ranunculaceae) in Central and Southern Italy, with Additions to North Italian Taxa. Webbia 2011, 66, 165–193. [Google Scholar] [CrossRef]
- Dunkel, F.G. Ranunculus pindicola sp. Nov., the Only Species of the R. Auricomus Complex (Ranunculaceae) in Greece. Willdenowia 2015, 45, 223–230. [Google Scholar] [CrossRef]
- Ulum, F.B.; Costa Castro, C.; Hörandl, E. Ploidy-Dependent Effects of Light Stress on the Mode of Reproduction in the Ranunculus auricomus Complex (Ranunculaceae). Front. Plant Sci. 2020, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Klatt, S.; Schinkel, C.C.F.; Kirchheimer, B.; Dullinger, S.; Hörandl, E. Effects of Cold Treatments on Fitness and Mode of Reproduction in the Diploid and Polyploid Alpine Plant Ranunculus Kuepferi (Ranunculaceae). Ann. Bot. 2018, 121, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Dunkel, F.G. The Ranunculus auricomus L. Complex (Ranunculaceae) in Northern Italy. Webbia 2010, 65, 179–227. [Google Scholar] [CrossRef]
- Hodač, L.; Karbstein, K.; Tomasello, S.; Wäldchen, J.; Bradican, J.P.; Hörandl, E. Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex. Biology 2023, 12, 418. [Google Scholar] [CrossRef]
- Feliner, G.N. Southern European Glacial Refugia: A Tale of Tales. Taxon 2011, 60, 365–372. [Google Scholar] [CrossRef]
- Petit, R.J.; Aguinagalde, I.; De Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial Refugia: Hotspots But Not Melting Pots of Genetic Diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef]
- Pellino, M.; Hojsgaard, D.; Schmutzer, T.; Scholz, U.; Hörandl, E.; Vogel, H.; Sharbel, T.F. Asexual Genome Evolution in the Apomictic Ranunculus auricomus Complex: Examining the Effects of Hybridization and Mutation Accumulation. Mol. Ecol. 2013, 22, 5908–5921. [Google Scholar] [CrossRef]
- Marklund, G.; Rousi, A. Outlines of Evolution in the Pseudogamous Ranunculus auricomus Group in Finland. Evolution 1961, 15, 510–522. [Google Scholar] [CrossRef]
- Paun, O.; Greilhuber, J.; Temsch, E.M.; Hörandl, E. Patterns, Sources and Ecological Implications of Clonal Diversity in Apomictic Ranunculus Carpaticola (Ranunculus auricomus Complex, Ranunculaceae). Mol. Ecol. 2006, 15, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Matzk, F.; Meister, A.; Schubert, I. An Efficient Screen for Reproductive Pathways Using Mature Seeds of Monocots and Dicots: Screen for the Pathway of Reproduction. Plant J. 2000, 21, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Fér, T.; Schmickl, R.E. HybPhyloMaker: Target Enrichment Data Analysis From Raw Reads to Species Trees. Evol. Bioinform. 2018, 14, 117693431774261. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.G.; Gardner, E.M.; Liu, Y.; Medina, R.; Goffinet, B.; Shaw, A.J.; Zerega, N.J.C.; Wickett, N.J. HybPiper: Extracting Coding Sequence and Introns for Phylogenetics from High-throughput Sequencing Reads Using Target Enrichment. Appl. Plant Sci. 2016, 4, 1600016. [Google Scholar] [CrossRef] [PubMed]
- Tange, O. Gnu Parallel 2018; Zenodo: Geneva, Switzerland, 2018. [Google Scholar]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Zhang, C.; Rabiee, M.; Sayyari, E.; Mirarab, S. ASTRAL-III: Polynomial Time Species Tree Reconstruction from Partially Resolved Gene Trees. BMC Bionform. 2018, 19, 153. [Google Scholar] [CrossRef]
- Nauheimer, L.; Weigner, N.; Joyce, E.; Crayn, D.; Clarke, C.; Nargar, K. HybPhaser: A Workflow for the Detection and Phasing of Hybrids in Target Capture Data Sets. Appl. Plant Sci. 2021, 9, aps3.11441. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab.(LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Revell, L. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population ID | Country | Number of Individuals | Number of Seeds | Percent Apomictic | Percent Sexual | Individuals Sexual |
|---|---|---|---|---|---|---|
| 17842 | BA | 1 | 5 | 100 | 0 | 0 |
| Q | IT | 3 | 28 | 100 | 0 | 0 |
| AD | IT | 2 | 19 | 100 | 0 | 0 |
| BB | IT | 3 | 29 | 100 | 0 | 0 |
| BosBo | IT | 3 | 26 | 100 | 0 | 0 |
| CdR | IT | 3 | 27 | 100 | 0 | 0 |
| C | IT | 3 | 27 | 100 | 0 | 0 |
| 11092 | ES | 5 | 25 | 100 | 0 | 0 |
| 11094 | ES | 6 | 24 | 91.30 | 8.70 | 2:11094_5, 33% Sexual; 11094_4, 25% Sexual |
| 11096 | ES | 2 | 10 | 100 | 0 | 0 |
| 11097 | ES | 2 | 11 | 100 | 0 | 0 |
| 11098 | ES | 2 | 19 | 100 | 0 | 0 |
| 11099 | ES | 3 | 25 | 100 | 0 | 0 |
| 11102 | ES | 3 | 34 | 100 | 0 | 0 |
| 11104 | ES | 4 | 32 | 100 | 0 | 0 |
| 11105 | ES | 1 | 10 | 100 | 0 | 0 |
| 11106 | ES | 3 | 25 | 100 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradican, J.P.; Tomasello, S.; Boscutti, F.; Karbstein, K.; Hörandl, E. Phylogenomics of Southern European Taxa in the Ranunculus auricomus Species Complex: The Apple Doesn’t Fall Far from the Tree. Plants 2023, 12, 3664. https://doi.org/10.3390/plants12213664
Bradican JP, Tomasello S, Boscutti F, Karbstein K, Hörandl E. Phylogenomics of Southern European Taxa in the Ranunculus auricomus Species Complex: The Apple Doesn’t Fall Far from the Tree. Plants. 2023; 12(21):3664. https://doi.org/10.3390/plants12213664
Chicago/Turabian StyleBradican, John Paul, Salvatore Tomasello, Francesco Boscutti, Kevin Karbstein, and Elvira Hörandl. 2023. "Phylogenomics of Southern European Taxa in the Ranunculus auricomus Species Complex: The Apple Doesn’t Fall Far from the Tree" Plants 12, no. 21: 3664. https://doi.org/10.3390/plants12213664