
The Lotus angustissumus Group (Fabaceae): Can Phylogenetic Patterns Be Accommodated by a Taxonomic Concept?

1
Department of Higher Plants, Biological Faculty, Lomonosov Moscow State University, GSP-1, Leninskie Gory, 119234 Moscow, Russia
2
A.N. Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, GSP-1, Leninskie Gory, 119991 Moscow, Russia
*
Author to whom correspondence should be addressed.
Plants 2024, 13(1), 101; https://doi.org/10.3390/plants13010101
Submission received: 2 December 2023
/
Revised: 25 December 2023
/
Accepted: 26 December 2023
/
Published: 28 December 2023
(This article belongs to the Section Plant Systematics, Taxonomy, Nomenclature and Classification)
Abstract
:The Lotus angustissimus group represents an example of obvious contradictions between the molecular and morphological data. This group includes from six to eight mostly annual species of Lotus section Lotus, with the center of species diversity in the Mediterranean. We performed a phylogenetic analysis of the genus Lotus with an expanded representation of all known species of the L. angustissimus group using both nuclear (nrITS) and a set of plastid DNA markers and compared the results with traditional taxonomy of this group. Our results demonstrated a non-monophyletic nature of the study group. In addition, the nrITS and plastid dataset tree topologies were incongruent with each other in both Bayesian and ML analyses. We revealed very complex phylogenetic relationships among the species of the group. Important results obtained in this study are: (1) genetic and geographical differentiation in the latitudinal direction (between L. angustissimus and L. praetermissus) and in the longitudinal direction among closely related accessions identified as L. castellanus, L. lourdes-santiagoi and L. palustris; (2) close genetic links between the Anatolian endemic L. macrotrichus and L. praetermissus from Eastern Europe and Central Asia; and (3) the hybrid nature of L. subbiflorus with L. parviflorus as its presumed male parent species. These results are discussed in the context of morphology, biogeography and taxonomy.
1. Introduction
Molecular phylogenetic studies have radically changed the traditional taxonomy of many groups of flowering plants, including the Fabaceae family (e.g., [1]). In some cases, taxa isolated by classical taxonomy methods have been confirmed using molecular phylogenetic approaches. In other cases, there were obvious contradictions between the molecular and morphological data, and one such example is the Lotus angustissimus group. This is a relatively small group of mostly annual Lotus species of the Lotus section, which includes, according to various points of view, from six to eight species and has a center of species diversity in the Mediterranean.
1.1. Taxonomic Composition of the Lotus angustissimus Group
The species Lotus angustissimus L. was described by Linnaeus [2]. Heyn included it and a number of morphologically similar species in the Lotus angustissimus group [3]. She showed the importance of keel beak and fruit length features for the delimitation of species in this group. Heyn [3] separated species of the group into two subgroups: with a straight keel beak (L. angustissimus, L. macrotrichus Boiss., L. suaveolens Pers. and L. parviflorus Desf.) and a curved keel beak (L. subbiflorus Lag. and L. palustris Willd.).
Kuprianova [4] described a species Lotus praetermissus Kuprian. distributed in Eastern Europe and Central Asia, which differs from L. angustissimus in solitary erect stems with spreading branches, wider and shorter legumes, and dark brown seeds. Greuter et al. [5], in the checklist of Mediterranean flora, distinguished the following species: L. angustissimus (incl. L. praetermissus), L. divaricatus Boiss. (syn. L. macrotrichus), L. parviflorus, and L. palustris aggr., which includes L. palustris, L. castellanus Boiss. & Reut. and L. hispidus Desf. ex DC. (syn. L. suaveolens, L. subbiflorus).
Kramina conducted a taxonomic revision of the Lotus angustissimus group [6]. She demonstrated that the priority name for the species with a long straight keel beak and middle-sized pods is L. subbiflorus [7], not L. suaveolens [8]. She also argued why the name L. hispidus Desf. cannot be used for this taxon and demonstrated that the name L. castellanus Boiss. & Reut. has a priority for the species with a long, curved keel beak. She discovered a new significant character, i.e., ovary and pod pilosity along the ventral suture, which distinguishes L. castellanus from L. palustris. In L. castellanus, fruits and ovaries are hairy along the ventral suture and in L. palustris, they are glabrous or sometimes with a few trichomes [6]. Multivariate morphological study of the L. angustissimus group from the same work [6] demonstrated the following results: (1) L. praetermissus is not distinguished from L. angustissimus by main diagnostic features (except those of seeds); (2) L. macrotrichus is close to L. angustissimus and L. praetermissus; (3) there are transitional morphological forms between L. castellanus and L. palustris; (4) analysis carried out on a large set of morphological features led to the division of the dataset into four groups—a. L. parviflorus, b. L. subbiflorus, c. L. angustissimus + L. praetermissus + L. macrotrichus, and d. L. castellanus + L. palustris—but for a reduced set of characteristics, including only diagnostic ones, and connections were found between L. subbiflorus and groups a and d.
Pina and Valdés described a new species in the L. angustissimus group, Lotus lourdes-santiagoi Pina & Valdés, close to L. castellanus but differing by hairy standard petals. They also clarified a set of diagnostic characteristics for distinguishing species in the group [9].
In the present paper we consider the Lotus angustissimus group according to Pina and Valdés [9], including seven species: L. angustissimus L. (incl. L. praetermissus Kuprian.), L. macrotrichus Boiss., L. subbiflorus Lag., L. parviflorus Desf., L. castellanus Boiss. & Reut., L. lourdes-santiagoi Pina & Valdés, and L. palustris Willd. Important morphological characteristics are presented in Figure 1. Main diagnostic characteristics of species of the Lotus angustissimus group [3,4,6,9] are summarized in Table 1.
1.2. Chromosome Numbers in the Lotus angustissimus Group
Traditionally, the Lotus angustissimus group was included in Lotus section Lotus, which is characterized by main chromosome number x = 6, in contrast to the other sections of the genus with main chromosome number x = 7 [10]. According to literature data, species of L. angustissimus are diploids with 2n = 2x = 12 (L. castellanus, L. parviflorus, L. praetermissus), tetraploids with 2n = 4x = 24 (L. subbiflorus) or have two ploidy levels 2n = 12, 24 (L. angustissimus) or variable chromosome numbers 2n = 12, 14, 24 (L. macrotrichus, L. palustris) [3,10,11,12,13]. The number 2n = 14 indicated for the species of L. angustissimus group is doubtful. In general, the chromosome numbers published for species of the L. angustissimus group should be treated with caution due to the difficulties in determining the species in this group and the different taxonomic views of the authors.
1.3. Geographical Distribution of Lotus angustissimus Group
Data on the geographical distribution of the species of the Lotus angustissimus group are summarized from several sources [3,6,14]. Lotus angustissimus is distributed in the Mediterranean, Europe (except the northern part), and the Middle East. The distribution area of Lotus praetermissus is in Eastern Europe and Central Asia (Kazakhstan, South Ural and South Siberia). Lotus macrotrichus is endemic to Western Anatolia and adjacent islands. Of a pair of closely related species, Lotus castellanus is common in the Western Mediterranean, and Lotus palustris is sporadically found throughout the Mediterranean and in Western Europe, although the exact range of the latter is unknown due to poorly defined morphological boundaries between the two species. Lotus lourdes-santiagoi is endemic to Andalusia (southwestern Spain). The main distribution area of Lotus parviflorus and Lotus subbiflorus is the Western Mediterranean and Macaronesia. Lotus subbiflorus also was rarely recorded from the Eastern Mediterranean and southwest England.
1.4. Previous Phylogenetic Studies of Lotus angustissimus Group
Early molecular phylogenetic study of the tribe Loteae investigated phylogenetic relationships among several species of the L. angustissimus group by nrITS markers [15]. In this work, the group was not monophyletic: L. angustissimus was clustered with L. parviflorus in a common clade, and L. palustris with L. corniculatus in another unrelated clade. L. subbiflorus took a separate position. The results obtained by Allan et al. [15] were not confirmed by subsequent studies.
Degtjareva et al. [16] analyzed nrITS markers in a representative sample of Lotus species and revealed the separation of the section Lotus into two highly supported clades. The first clade included L. subbiflorus, L. parviflorus and L. conimbricensis, and the second all the other studied species of the section, including L. praetermissus and L. angustissimus, as well as representatives of the groups L. pedunculatus and L. corniculatus. Interestingly, these two clades were not sister to each other on the phylogenetic tree [16]. Subsequent phylogenetic studies of the genus Lotus revealed that L. conimbricensis is not related to any species of the Lotus angustissimus group [17,18].
Phylogenetic study of a representative set of Lotus species based on nuclear (nrITS, nrETS) and plastid (psbA-trnH, rps16) DNA markers revealed subdivision of the genus Lotus into the so-called Southern clade and one or more (according to different markers) Northern clades [17]. The Southern clade includes the majority of sections of the genus, and the Northern one only three sections (i.e., Lotus, Dorycnium and Bonjeanea). The study demonstrated a non-monophyletic pattern of the section Lotus [17]. The species of the Lotus angustissimus group belong to the Northern clade (by plastid markers) or are distributed among the northern clades (by nuclear markers). None of the markers confirmed the monophyly of the Lotus angustissimus group as understood by Heyn [3]. Close relationships were demonstrated between L. subbiflorus and L. parviflorus and between L. castellanus and L. palustris by ITS marker, and between L. subbiflorus and L. castellanus and between L. angustissimus and L. praetermissus by plastid markers. The plastid dataset also clearly showed a close relationship between L. angustissimus and L. pedunculatus groups. However, an insufficiently presented sample of species of the Lotus angustissimus group does not allow the authors to draw confident conclusions about the phylogenetic relationships of the species of the group [17]. The species L. macrotrichus and L. lourdes-santiagoi have not been studied by molecular methods, and their phylogenetic position was unknown until now.
The present study was aimed at conducting a phylogenetic analysis of the genus Lotus with an expanded representation of all currently known species of the Lotus angustissimus group using both nuclear (ITS1-5.8S-ITS2), and plastid DNA markers and comparison of the results obtained with the data of the traditional taxonomy of this group.
2. Results
2.1. Taxonomic Identification of Specimens Based on Morphology
The vast majority of samples were unambiguously identified as representatives of the following species based on morphological data: L. angustissimus (ANG1–ANG5, ANG7–ANG11, ANG13, ANG15, ZLA, 472), L. praetermissus (TH4-TH7, STN6, POY, 458, 473), L. macrotrichus (819), L. castellanus (CAS1–CAS2, 417, 471), L. lourdes-santiagoi (LS1-LS2), L. palustris (PAL3–PAL5, 426, 869, 875), L. subbiflorus (457, 470, 475, 476, SUBB1, TH2), L. parviflorus (PRV1–PRV5, 469). The specimen TH1 from Israel (MW 0740247) possesses no reproductive structures (flowers, fruits). It was previously identified as Lotus palustris Willd.; however reexamination of this specimen led to its reidentification as Lotus rectus L. Voucher information is presented in Table 2. Two specimens from Spain and Portugal formerly identified as L. palustris (PAL1 and PAL2) were intermediate between L. castellanus and L. palustris (Table 3). Three specimens (ANG12, ANG14, and ANG17) were intermediate between L. angustissimus and L. subbiflorus in their morphological characteristics and could not be unambiguously determined (Table 4).
2.2. Phylogenetic Analysis of nrITS Dataset
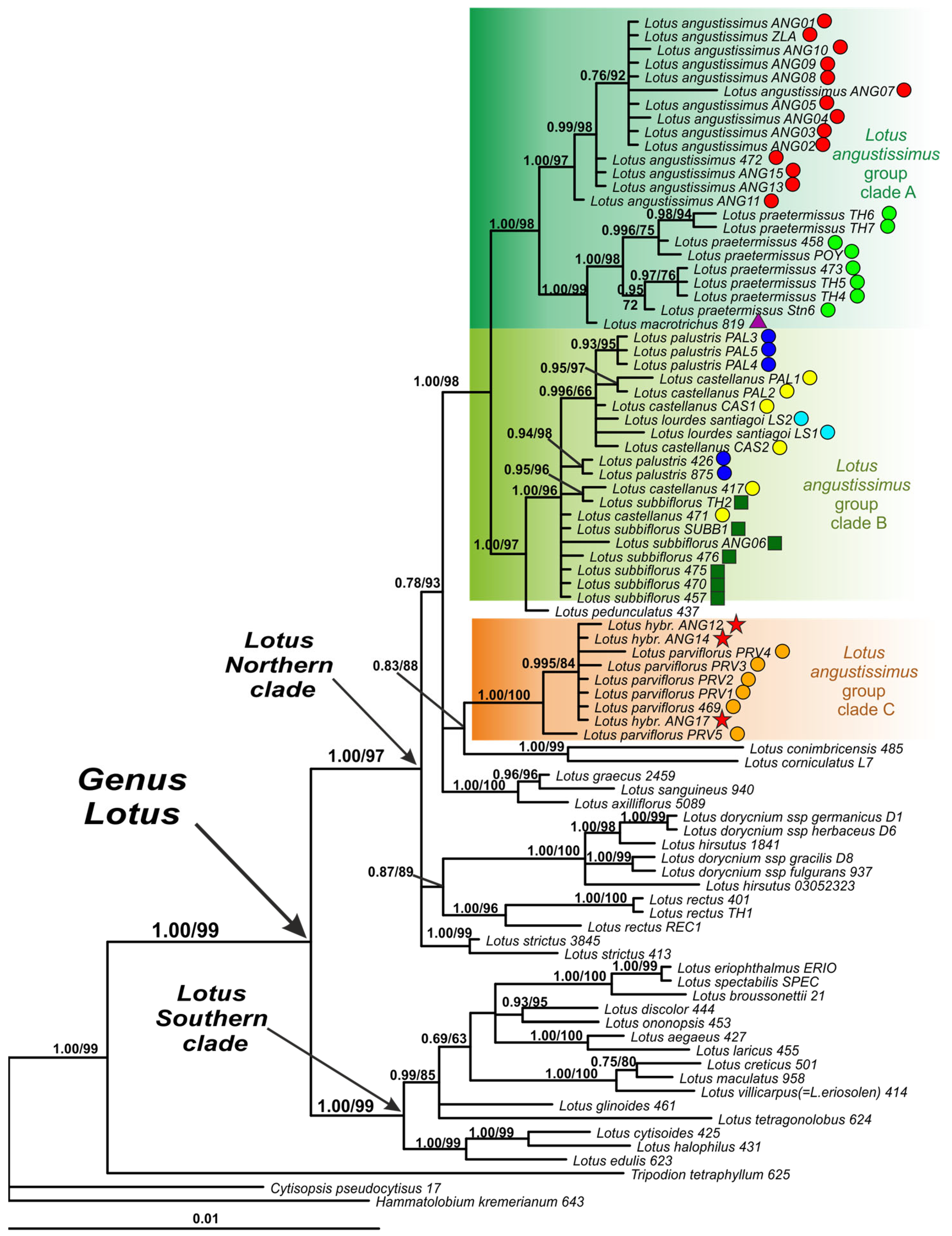
On the nrITS phylogenetic tree, the genus Lotus forms a well-supported clade with several subclades (Figure 2). Among them, a Southern clade, several northern clades (i.e., Lotus Northern clades I, II, III and IV) and a separate branch of Lotus glinoides are observed (Figure 2). Species of the Lotus angustissimus group are clustered within two highly supported clades, Lotus angustissimus group clade 1 and Lotus angustissimus group clade 2, which are not related to each other. Clade 1 is a part of the Lotus Northern clade I, and clade 2 corresponds to the Lotus Northern clade III (Figure 2).
Within clade 1, L. praetermissus and L. angustissimus form well-supported species-specific subclades. Lotus macrotrichus is clustered with L. praetermissus taking a basal position in the common L. praetermissus plus L. macrotrichus clade. Lotus palustris, L. castellanus and L. lourdes-santiagoi form another clade, where specimens of the latter two species are mixed in a separate smaller subclade, and specimens of L. palustris specimens occupy a basal position.
Clade 2 includes a mixture of studied specimens of L. parviflorus and L. subbiflorus, as well as three specimens with uncertain taxonomic identification (ANG12, ANG14, and ANG17). In the specimen ANG17, two variants of nrITS were found, differing in position 483 of the alignment (T in clone 1/C in clone 2).
Phylogenetic analysis of the nrITS dataset confirmed that the specimen TH1 belongs to the species Lotus rectus L.
2.3. Phylogenetic Analysis of the Plastid DNA Dataset
On the phylogenetic tree constructed by the plastid DNA dataset, the genus Lotus is also well defined and subdivided into two large highly supported clades: the Northern clade and the Southern clade (Figure 3). In addition, the Northern clade on the phylogenetic tree, constructed by plastid data, unites all members of Northern clades I, II, III and IV, identified on the tree constructed by nrITS. Lotus glinoides is included in the Southern clade.
Representatives of the Lotus angustissimus group occupy positions in three highly supported subclades within the Northern clade, that is, in Clades A, B and C. The composition of none of these three subclades is identical to that of either clade 1 or clade 2 from the nrITS tree.
Clade A on the plastid DNA phylogenetic tree corresponds to a part of clade 1 on the nrITS tree and includes three well separated species-specific groups consisting of specimens of L. angustissimus, L. praetermissus and L. macrotrichus, respectively.
Clade C on the plastid DNA phylogenetic tree corresponds to a part of clade 2 on the nrITS tree and includes L. parviflorus and three specimens of uncertain taxonomic identification (ANG12, ANG14, and ANG17).
Clade B of the plastid DNA phylogenetic tree partially corresponds to clade 1 and clade 2 of the nrITS tree and includes L. palustris, L. castellanus and L. lourdes-santiagoi together with L. subbiflorus. Within clade B, the species L. palustris, L. castellanus, L. lourdes-santiagoi and L. subbiflorus are in an unresolved position. It is worth noting that the sister position in relation to clade B is occupied by Lotus pedunculatus Cav., which is not a member of the Lotus angustissimus group, but represents a separate group of the Lotus section. Clades A and (B + L. pedunculatus) together form a larger, well-supported clade, while clade C occupies a position away from them.
Phylogenetic analysis of the plastid dataset confirmed the belonging of the TH1 sample to the species Lotus rectus L.
2.4. Geographic Patterns Observed in the Lotus angustissimus Group
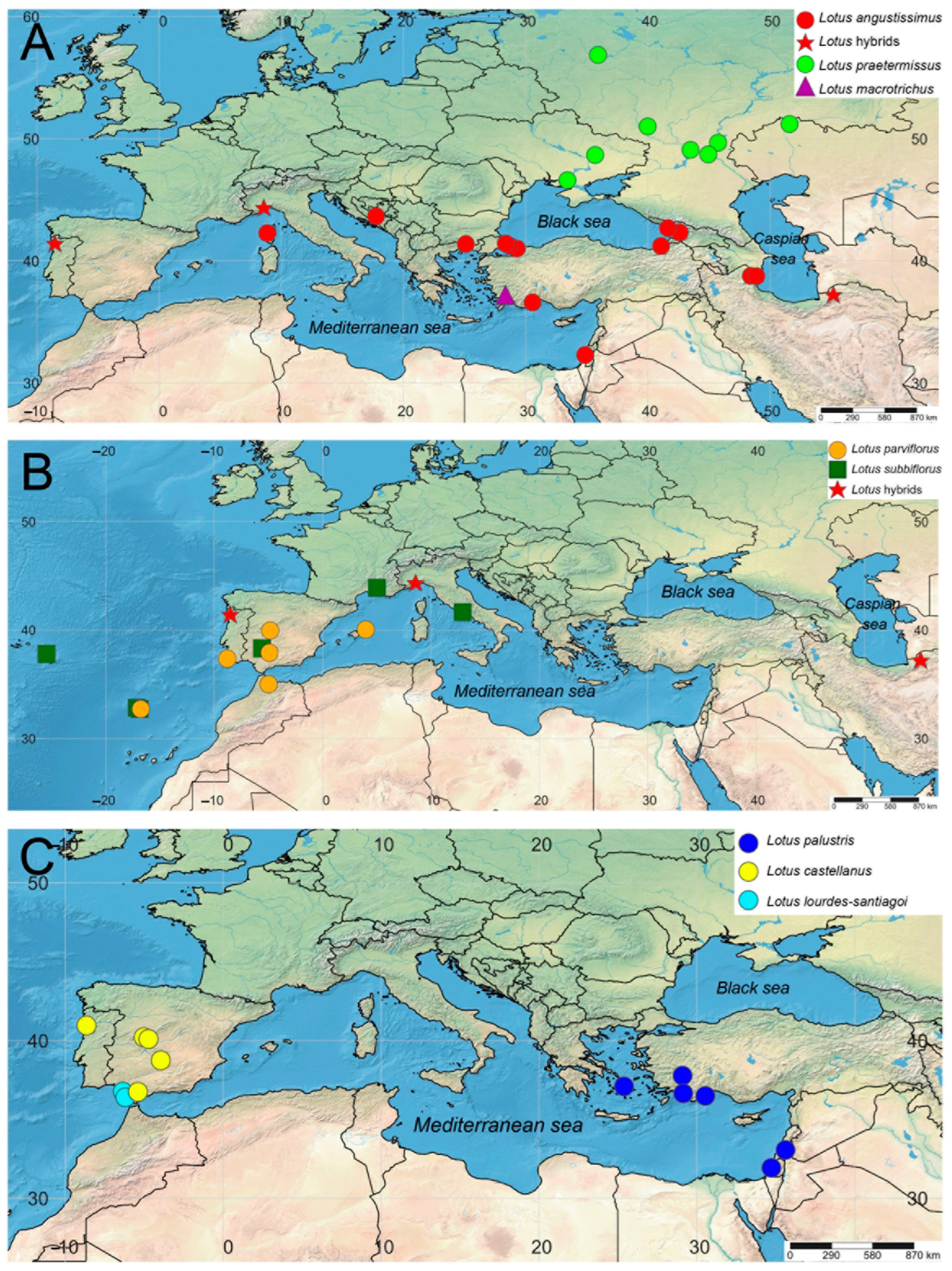
Both nrITS and studied plastid markers confirm the geographical differentiation within the pair of the closely related species Lotus angustissimus and L. praetermissus. The studied specimens of L. angustissimus occupy the southern part of the common range of these two species, mainly around the Mediterranean and Black Seas and in the southern part of the Caspian Sea, and specimens of L. praetermissus are distributed in the northern part, namely in the center and southeast of the European part of Russia, Ukraine and western Kazakhstan (Figure 2, Figure 3 and Figure 4A). The only studied specimen L. macrotrichus from SW Turkey is clustered together with L. praetermissus in both phylogenetic reconstructions, though geographically it is closer to L. angustissimus (Figure 2, Figure 3 and Figure 4A).
Geographic pattern was also observed within the complex of Lotus castellanus, L. lourdes-santiagoi and L. palustris on the nrITS phylogenetic tree. All specimens of L. palustris from the Eastern Mediterranean are unresolved at the base of the common clade of the complex, whereas all specimens from the Western Mediterranean, including L. castellanus and L. lourdes-santiagoi, formed a highly supported subclade. Two specimens of L. castellanus from Spain and Portugal formerly classified within L. palustris (PAL1 and PAL2), but having an intermediate morphologic pattern between typical L. palustris and L. castellanus, were also members of this western subclade (Figure 2, Figure 3 and Figure 4C). In the phylogenetic tree constructed using plastid markers, no phylogenetic or geographical structure was observed in this complex, since these markers do not allow for resolving the relationships in this group of species (Figure 3).
3. Discussion
Here, we present the first molecular phylogenetic study that includes all known up-to-date species of the Lotus angustissimus group, analyzed by both nuclear and plastid DNA markers. Despite comparatively small datasets studied for each species, we tried to cover different parts of their ranges, and the results obtained allowed us to make several important conclusions concerning phylogenetic relationships, geography and taxonomy in this group.
The phylogenetic reconstructions obtained in this study confirmed the non-monophyletic nature of the Lotus angustissimus group, which was previously assumed in earlier studies of the genus Lotus [16,17,18]. However, all representatives of this group are connected in a complex way by common molecular features. The Lotus angustissimus group represents a good example of the discrepancy between nuclear and plastid phylogeny, which may indicate different trends in the evolution of nuclear and plastid lines and the presence of traces of network evolution.
3.1. Lotus angustissimus, L. praetermissus and L. macrotrichus
The results of this study by both nuclear and plastid markers clearly support the separation of L. praetermissus from L. angustissimus. This point of view is shared by many authors of regional floras of the territories of the former USSR, Eastern Europe and Central Asia [19,20,21,22,23,24]. The opposite concept was expressed by Heyn [3], some Western European authors [5,25,26], and botanists of the Legume Phylogeny Working Group [27] who did not support the segregation of L. praetermissus from L. angustissimus. Detailed morphometric study [6] did not reveal a clear difference between the two species; however, some important characteristics (e.g., seed surface color) were not included in the analysis due to the absence of ripe seeds in the majority of studied specimens. One of the most prominent morphological features distinguishing typical L. praetermissus from typical L. angustissimus is a specific growth pattern: L. praetermissus is usually a small annual plant with a prominent erect main shoot branched in the upper half, whereas L. angustissimus is a more branched annual or perennial plant with ascending shoots. However, a number of transitional forms are observed between the two extreme morphological variants. Moreover, when the plants of L. praetermissus were grown from seeds in the greenhouse with sufficient water supply, they had a habitus more resembling that of L. angustissimus (Kramina, unpublished data).
The genetic separation of L. praetermissus and L. angustissimus is in good agreement with their geographical distribution, and the range of L. praetermissus is confined to more northern and eastern regions. As a whole, the results obtained in this work can serve as an argument in favor of the allocation of two taxa in the rank of independent species. This contradicts the conclusions made earlier only on morphological grounds [6]. Apparently, the problem deserves further study with the involvement of analysis of population variability at both the morphological and molecular levels.
The only studied specimen of the local endemic of Western Anatolia, Lotus macrotrichus, is related to Lotus praetermissus based on both nuclear and plastid DNA markers. These results are more or less consistent with morphometric studies [6], where a specimen of L. macrotrichus was morphologically close to L. praetermissus and Turkish specimens of L. angustissimus in many characteristics, but differed from them by the shape of upper leaflets, larger flowers, styles, and fruits. Sister phylogenetic relationships between L. macrotrichus and L. praetermissus do not contradict the recognition of the former as a separate species, but a more comprehensive study is needed for a balanced decision.
3.2. Lotus castellanus, L. lourdes-santiagoi and L. palustris
All three species share a common morphological feature, namely, a keel with a curved keel beak. Their close relationship is confirmed by the ITS nuclear marker. Our study demonstrated that L. palustris is paraphyletic in relation to L. castellanus and L. lourdes-santiagoi by the nrITS marker, but plastid data cannot resolve the relationships among these three species.
The molecular phylogenetic results obtained in the present study do not allow support for the isolation of L. lourdes-santiagoi from L. castellanus. These two species can be clearly distinguished from each other, but only by one morphological feature—the presence of pubescence on a standard petal. This character is very interesting because it illustrates the phenomenon of the occurrence of morphological features of a neighboring taxon in this taxon. The pubescent back surface of standard petal is known in Lotus section Pedrosia (Lowe) Christ. (in seven species, incl. Lotus chazaliei H. Boissieu, L. loweanus Webb & Berthel., and others) and section Rhyncholotus (Monod) D.D. Sokoloff (all members), as well as in Cytisopsis ahmedii (Batt. & Pit.) Lassen, Tripodion tetraphyllum (L.) Fourr., and Hammatolobium lotoides Fenzl from the tribe Loteae. Within the species Hammatolobium lotoides, some plants have a pubescent standard petal, whereas the others have a glabrous one [28]. A similar variability pattern was observed within four species of Lotus section Pedrosia [29]. In the Lotus section Lotus, to which representatives of the L. angustissimus group belong, L. lourdes-santiagoi is the only example of a plant with a pubescent standard petal.
The results of nrITS phylogenetic analysis indicate that the genetic data for the three species have a clear geographical structure: samples from the Western Mediterranean (typical L. castellanus, atypical samples from Spain and Portugal previously identified as L. palustris, and L. lourdes-santiagoi) have a number of molecular synapomorphies that distinguish them from samples distributed in the Eastern Mediterranean (typical L. palustris). Such a pattern can be interpreted in at least two ways: (1) all studied samples belong to the same species; and (2) samples from the Western Mediterranean belong to L. castellanus (or to L. castellanus and L. lourdes-santiagoi) and those from the Eastern Mediterranean should be treated as L. palustris. Perhaps the use of other more rapidly evolving molecular markers could be successful in this case.
The nrITS data suggest that Lotus palustris from the Eastern Mediterranean is closer to a hypothetical genetic variant ancestral to the pair of species L. castellanus and L. palustris, which, however, is not confirmed by the analysis of plastid data. The studied plastid data did not allow resolution of the relationships within the group of species L. castellanus, L. lourdes-santiagoi, L. palustris and L. subbiflorus, which together form clade B in the phylogenetic tree (Figure 3). One of the reasons for this phylogenetic pattern may be the slow rate of mutation of plastid DNA sequences, which, combined with incomplete lineage sorting, may lead to a lack of isolation of plastid sequences found in each of the four species. The second possible cause of such a pattern may be hybridization events that may have occurred in the evolution of this group of four species, either in recent times or in the more distant past. Two of the species forming clade B, L. palustris and L. subbiflorus, are known as possible tetraploids [3,10,11], which can be considered an argument in favor of the second opinion.
3.3. Lotus subbiflorus
Our study showed that Lotus subbiflorus is genetically close to L. parviflorus based on nuclear ITS data and to the complex L. castellanus–L. lourdes-santiagoi–L. palustris by a set of plastid markers. Such a genetic pattern may indicate the hybrid nature of Lotus subbiflorus. This assumption is consistent with the data on the tetraploid chromosome number of this species, 2n = 24 [3,10], and the cytogenetic data obtained by Ferreira and Pedrosa-Harand [11]. Ferreira and Pedrosa-Harand [11], using the FISH method, demonstrated the presence of Ljcen1 signals in only one set of L. subbiflorus chromosomes, which allowed them to assume an allotetraploid origin of this species. According to the results of our study, the diploid L. parviflorus may be the presumed male parent species of L. subbiflorus, and a representative of L. castellanus–L. lourdes-santiagoi–L. palustris complex may be its putative female parent species. Data on morphology and geography tentatively suggest the participation of L. castellanus as a second parent species of L. subbiflorus. L. castellanus is similar to L. subbiflorus in long keel beak and middle-sized fruit (ca. 1.5–3 times as long as calyx), but they differ from each other in the keel beak shape, which is straight in L. subbiflorus and curved in L. castellanus. The curve of the keel tip is sometimes not very noticeable, so from time to time the two species are confused. Sympatric geographical distribution of L. castellanus and L. subbiflorus in the Western Mediterranean does not contradict the assumption of the participation of L. castellanus as a second parent species of L. subbiflorus either.
The phylogenetic position of three specimens, ANG12 from Portugal, ANG14 from Italy and ANG17 from Iran, raises questions. According to their morphological characteristics, these specimens occupy a more or less intermediate position between L. angustissimus and L. subbiflorus. By nrITS marker, they are clustered within a clade of L. parviflorus + L. subbiflorus, and by a plastid dataset, they are grouped with L. parviflorus. We checked these results by conducting two separate DNA isolations from ANG12 and ANG17 and several repetitions of each PCR reaction for all three specimens. The result was the same. As such, in this case, we have a real discrepancy between morphological identification and molecular data. The results of molecular studies imply the participation of L. parviflorus in the formation of the three individual plants, but we did not observe any morphological features of this species in the mentioned individuals, except for a straight keel beak, which is common for several species of the group. The first reason for such variability may be the hybrid nature of the three specimens with hidden morphological features of one parent species. One of the facts in favor of such a conclusion, which can be made for the sample ANG17, is the polymorphism of nrITS. However, this character does not allow identification of the putative parent species, as it was demonstrated for other cases within the genus Lotus (e.g., [30]). The second reason is incomplete lineage sorting, which is in good agreement with the geographical location of mentioned specimens in glacial refugia of the Mediterranean basin [31] and the southern Caspian region [32]. As such, the taxonomic status of the specimens ANG12, ANG14 and ANG17 remains uncertain. We prefer to identify them as Lotus spp. or Lotus hybrids. This problem needs further study using an expanded set of molecular markers, as well as various methods of analysis.
3.4. Lotus parviflorus
Heyn [3] considered Lotus parviflorus to be the least problematic species within the Lotus angustissimus group due to a set of clear diagnostic features, such as a long straight keel beak and a short fruit not exceeding the calyx. The results obtained in this work confirmed the isolated position of the clade Lotus parviflorus + L. subbiflorus within the genus Lotus in the nrITS phylogenetic trees, which was found in previous studies on a limited dataset [17,18]. In the phylogenetic reconstructions made in this study using the plastid DNA dataset, L. parviflorus forms a separate clade, without L. subbiflorus. The inclusion of three specimens with uncertain taxonomic identification in this clade was discussed above. Thus, there is clear evidence of a genetic relationship between L. parviflorus and only one representative of the Lotus angustissimus group, L. subbiflorus, which is consistent with morphological data [6]. We are not inclined to combine these two species into one, since the hybrid nature of L. subbiflorus is shown in the present and previous studies.
4. Materials and Methods
4.1. Plant Material
The molecular study involved 88 specimens, including 53 specimens of Lotus angustissimus group, 32 specimens of other Lotus species representing all main sections of the genus, and 3 specimens of genera Cytisopsis, Hammatolobium and Tripodion, closely related to Lotus. Samples for molecular studies were taken from herbarium specimens stored in herbaria ANK, B, BM, GAZI, LE, MA, MHA, MW, and NSW. Voucher information and GenBank accession numbers are presented in Table 2 and Appendix A. Geographical distribution of specimens included in molecular analyses is presented on a map (Figure 4) prepared using SimpleMappr online software (https://www.simplemappr.net, accessed on 2 December 2023) [33]. All the studied specimens of the Lotus angustissimus group were taxonomically identified by morphological features using descriptions and identification keys from the main literary sources [3,6,9].
4.2. DNA Extraction, Amplification and Sequencing
DNA was extracted from herbarium specimens (ca. 20 mg of leaf tissue) with a NucleoSpin Plant II kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions or using the CTAB method [34]. The nrDNA ITS and plastid DNA regions psbA-trnH intergenic spacer, trnL-trnF intergenic spacer (IGS) and trnL intron, and rps16 intron were selected for the analysis because of their utility in Lotus and high variability [17,35,36]. The sequences of the nrITS were amplified with primers NNC-18S10, C26A [37], ITS2 and ITS3 [38]. The amplification of the psbA-trnH spacer was conducted using primers trnH2 [39] and psbAF [40]. The sequences of the trnL-trnF region of plastid DNA were amplified using standard primers “c”, “d”, “e” and “f” [41], and the sequences of rps16 intron using primers rpsF, rpsR2 [42], Lot-rps16-F and Lot-rps16-intR [35]. PCRs were performed in a 0.02 mL mixture containing 10–20 ng DNA, 3.2 pmol of each primer and MasDDTaqMIX (Dialat LTD, Moscow, Russia) containing 0.2 mM of each dNTP, 1.5 mM MgCl2, and 1.5 units of SmarTaqDNA polymerase. Amplification of the nrITS region and all plastid DNA regions was performed under the following conditions: hold 94 °C, 3 min; 94 °C, 30 s; 57 °C, 40 s; 72 °C, 60 s; repeat 30 cycles; extend 72 °C, 3 min. Amplification of the same regions in the samples taken from old herbarium specimens was performed under the following program: hold 94 °C, 1 min; 94 °C, 30 s; 57 °C, 40 s; 60 °C, 1 min 20 s; repeat 35 cycles; 57 °C, 40 s; 60 °C, 1 min 20 s; repeat 2 cycles.
PCR products were purified using a Cleanup Mini kit (Evrogen, Moscow, Russia) and then used as a template in sequencing reactions with the ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit v. 3.1. Sequencing was performed on the 3730 DNA Analyzer (Life Technologies, Carlsbad, CA, USA) in the Syntol company (Moscow, Russia). Forward and reverse strands of all samples were sequenced. For the majority of samples, the polymorphism of ITS within one specimen was detected by direct sequencing (without cloning), by the presence of double peaks on electropherogram. ITS sequences of the specimen ANG17 were cloned in the E. coli vector and then sequenced in the Evrogen Joint Stock Company (Moscow, Russia).
The sequences were aligned using MAFFT version 7.215 [43,44] and then adjusted manually in BioEdit version 7.2.5 [45]. The matrices of psbA-trnH spacer, trnL-F and rps16 intron plastid DNA regions were concatenated into a single matrix. Gap-rich and ambiguous positions were excluded from the analyses. The aligned data matrices are presented in Supplementary Materials Datasets S1 and S2.
4.3. Phylogenetic Analyses
Phylogenetic analyses were performed separately for the nrITS dataset and a concatenated plastid DNA dataset using both maximum likelihood and Bayesian inference methods.
Maximum likelihood analyses were performed in IQ-tree version 1.6.12 [46], and internal branch support was assessed using the ultrafast bootstrap [47] with 10,000 resamplings. The GTR + F + G4 model of nucleotide substitutions for plastid data and the SYM + G4 model for nrITS were selected as the most appropriate based on the Bayesian information criterion in the built-in ModelFinder utility [48].
Bayesian inference was performed using MrBayes v. 3.2.6 [49] considering the optimal model of nucleotide substitutions selected by AICc in PAUP version 4.0a [50] for each marker: SYM + Γ (symmetrical model with substitution rate heterogeneity) for nrITS, and GTR + Γ for plastid data. The Bayesian analysis used four independent runs of 25 million generations and four chains sampling every 1000th generation. Non-convergence assessment and burn-in estimation was carried out in VMCMC ver. 1.0.1 [51]. The first two million generations were discarded as burn-in, and the remaining trees from both runs were combined in a 50% majority-rule consensus tree.
5. Conclusions
The results of the present phylogenetic study of the Lotus angustissimus group together with a representative set of Lotus species, using nrITS and a set of three plastid DNA regions, clearly demonstrated a non-monophyletic nature of the study group. The nrITS and plastid dataset tree topologies were incongruent in both Bayesian and ML analyses.
Our data demonstrated geographical differentiation between genetic variants within a pair of species Lotus angustissimus L. and L. praetermissus Kuprian., and the southern variants corresponded to the first and the northern ones to the second of the species. The phylogenetic position of the Anatolian endemic Lotus macrotrichus Boiss. has been determined for the first time, which turned out to be close to L. praetermissus, but separated from the latter by a number of mutations both in nrITS and plastid sequences. The position of Lotus subbiflorus Lag. in phylogenetic trees confirms its hybrid nature and implies that its presumed male parent is L. parviflorus Desf. The present phylogenetic results do not allow us to support the isolation of L. lourdes-santiagoi from L. castellanus, which is consistent with the very weak morphological difference between these taxa. The nrITS data indicate a clear geographical structure within the L. castellanus–L. lourdes-santiagoi–L. palustris complex, where the samples of the Western Mediterranean differ from those of the Eastern Mediterranean.
The present study has revealed several trends in genetic variability within the Lotus angustissimus group. In order to assess the limits of this variability and the degree of genetic isolation of individual species, it is necessary to expand the study both with respect to the sample (including the population level) and with respect to the studied genetic markers.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/plants13010101/s1. Dataset S1: nrITS dataset. Dataset S2: Combined plastid dataset (trnL-F, rps16 and psbA-trnH).
Author Contributions
T.E.K. contributed to the field collection of samples, all the laboratory work, including DNA extraction, PCR, sequencing, analyses of the data, figure preparation, and manuscript writing and editing. T.R.H. contributed to all the laboratory work, including DNA extraction, PCR, and sequencing. T.H.S. contributed to phylogenetic analyses of the data and to manuscript writing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Russian Science Foundation, project 23-24-00052.
Data Availability Statement
Data are contained within the article or Supplementary Materials.
Acknowledgments
We are grateful to Dmitry Geltman, Vladimir Dorofeev, and Larisa Orlova for facilitating access to collections of the Herbarium of the Komarov Botanical Institute (LE), to Alexei Seregin, Natalia Reshetnikova, Nina Stepanova, Maria Nosova and Ivan Schanzer for kind assistance during the use of herbarium collections at Herbaria MW and MHA; the curators of the Herbaria ANK, B, BM, GAZI, LE, MA, MHA, MW, and NSW for permission for destructive sampling; Alexandra Milko for her kind help in figure preparation; and Dmitry Sokoloff, Galina Degtjareva, and Olga Yurtseva for helpful discussion. We thank the Ministry of Science and Higher Education of the Russian Federation for its support of the Centre of Collective Use Herbarium MBG RAS.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Taxa, sample code, locality, and voucher information (herbarium code) of Lotus, Cytisopsis, Hammatolobium and Tripodion species used in molecular analyses. GenBank accession numbers of sequences taken from GenBank are given for the four markers ITS, trnL-F, rps16, and psbA-trnH.
Cytisopsis pseudocytisus (Boiss.) Fertig: 7, Turkey, C1, Muğla, Datça, Knidos, 29–31.V.1995, A.P. Khokhryakov and M.T. Mazurenko s.n. (MHA), AY325282, MK751647, HM468299, HM468259; Hammatolobium kremerianum (Coss.) C.Muell.: 643, Morocco, Podlech 51378 (MHA), KT250926, MK751648, KT262933, KT262863; Lotus aegaeus Boiss.: 427, Turkey, C3, Antalya Korkuteli, Termessos, Büyükkumluca, Çakıllı geçidi, 04.VI.1995, A.P.Khokhryakov and M.T.Mazurenko 1135 (MHA), DQ160276, MK751649, KT262865, KT262794; L. axilliflorus (Hub.-Mor.) D.D. Sokoloff: 5089, Turkey, C2 Burdur, Yeşilova, Salda gölü, 12.VIII.1993, H. Duman, Z. Aytac and Dönmez 5089 (GAZI), MW412842, MW470873, MW498319, OL753484; Lotus broussonetii Choisy ex Ser.: 21, Cultivated at Royal Botanic Gardens, Kew: introduced from Canary Is., DQ160278, MK751653, KT262872, KT262802; Lotus conimbricensis Brot.: 485, Spain, Badajoz, Almendral, 27.IV.1966, Segura Zubizarreta 960 (Z), FJ411114, MF158231, KT262874, KT262804; Lotus corniculatus L., L7, Russia, Moscow prov., Lutzino, 03.VII.2008, Kramina 74-7 (MW), JF784200 and JF784201, MW470874, KT262876, KT262806; Lotus creticus L.: 501, Cultivated in Australia from seeds collected in Azores Is., Sandral SA39213 (MW), FJ938296, OL697808, KT262877, KT262807; Lotus cytisoides L.: 425, Cyprus, Seregin and Sokoloff 280 (MW), DQ166241, DQ160280, OL697809, KT262878, KT262808; Lotus discolor E. Mey: 444, Cameroon, S. Lisowski B-3330 (BR), DQ160288, MK751659, KT262880, KT262810; Lotus edulis L.: 623, Cyprus, 10 km to W from Limassol, 13.III.2004, Seregin and Sokoloff A-280 (MW), KT250863, MK751663, KT262885, KT262815; Lotus eriophthalmus Webb and Berthel.: ERIO, Spain, Tenerife, Cultivated at Botany Dept. of University of La Laguna, 11.V.1984, A. Gharpin M. del Asco 185745 (MA 318437), MW412843, MW470875, MW498320, OL753511; Lotus dorycnium ssp. fulgurans (Porta) Kramina and D.D. Sokoloff: 937, United Kingdom, Cultivated at Royal Botanic Gardens, Kew, 2010: origin Spain, Balearic Is., KT250865, MF314954, KT262887, KT262817; Lotus dorycnium ssp. germanicus (Gremli) Kramina and D.D. Sokoloff: D1, Slovenia, Polhograjsko Hribovje, prope Govejek, supra vicum Medvode, 19.VI.1973, D. Trpin and T. Wraber 9852/3 (H 1081128), KT250868, MK751666, KT262889, KT262819; Lotus dorycnium ssp. gracilis (Jord.) Kramina and D.D. Sokoloff: D8, France, dép. Pyrénées-Orientales, Canet, 02.VII.1981, J. Lambinon, R. Renard and L. Smeets 81/287 (H 1542915), KT250859, MK751682, KT262881, KT262811; Lotus dorycnium ssp. herbaceus (Vill.) Kramina and D.D. Sokoloff: D6, Austria, Steirisches Hügelland, Steiermark, Umgebung von Radkersburg, 7.VII.1976, H. Mayrhofer and H. Teppner s.n. (H 1216503), KT250882, MK751681, KT262898, KT262828; Lotus glinoides Del.: 461, Egypt, 7.V.1962, Bochantsev s.n. (LE), DQ166220, MK751677, KT262892, KT262822; Lotus graecus L.: 2459, Turkey, B3 Kütahya, Dumlupınar, Gökdağ, Akdene mevkii, 22.VII.1983, M. Vural, F. Maluen 2459 (GAZI), MW412844, MW470876, MW498321, OL753523; Lotus halophilus Boiss. and Spruner: 431, Greece, Karpathos, Pigadia, 19.IV.1984, Th.Raus 9307 (MHA), KT250879, MK751680, KT262896, KT262826; Lotus hirsutus L.: 03052323, Spain, Prov. Teruel, Mosqueruella, 24.V.1992, C. Fabregat and S. López s.n. (P 03052323), MW412854, MW470889, MW498338, OL753543; 1841, Turkey, C2 Muğla, Marmaris, Bağli Tepe Livari, 27.VI.1997, H. Sağban 1841 (GAZI), MW412857, MW470893, MW498342, OL753546; Lotus laricus Rech.f., Aellen and Esfand.: 455, Abu Dhabi, Abu Dhabi Island, Al Mushrif Palaca, 04.V.1982, R.A. Western 275 (E), DQ166233, MK751687, KT262906, KT262836; Lotus maculatus Breitf.: 958, Canary Is. (cult.), Tenerif. Municipio de la Orotava, Puerto de la Cruz, 14.IV.2000, H. Väre 10894 and H. Kaipiainen (H 1702795), KT250890, MK751688, KT262907, KT262837; Lotus ononopsis Balf. f.: 453, Yemen, Muqadrihon Pass, c. 10 km SW of Hadiboh, 26.I.1990, A.G. Miller et al. 10097 (E), DQ166219, MK751690, KT262909, KT262839; Lotus pedunculatus Cav.: 437, Spain, Soria, Santa Inés, 18.VII.1972, Segura Zubizarreta s.n. (LE), DQ166222, MF158224, KT262910, KT262840; Lotus rectus L.: 401, Lebanon, on the bank of the Nahr el Kalb, 05.VI.1959, T.D.Maitland 401 (LE), MW412874, MW470909, MW498361, OL753561; Lotus sanguineus (Vural) D.D. Sokoloff: 940, Turkey, C4 Konya, 00.00.1981, M. Vural 1976 (E), KT250904, MN553710, KT262916, KT262846; Lotus spectabilis Choisy ex Ser.: SPEC, Spain, Tenerife, Güimar, 00.VIII.1977, A. Santos-Ricardo 5124 (MA 839030), MW412881, MW470917, MW498370, OL753563; Lotus strictus Fisch. and C.A. Mey.: 3845, Turkey, B4, Tuz gölü, Aksaray-Eşmekaya sazlığı, 13.VII.1997, M. Aydoğdu 3845 (ANK), MW412882, MW470918, MW498371, OL753564; 413, Russia, Altai Krai, Mikhaylovsky distr. 18.IX.2003, Korolyuk s.n. (MW), DQ160286, MF158210, KT262923, KT262853; Lotus tetragonolobus L.: 624, Cyprus, to E from Limassol, Amathus, 08.III.2004, A. Seregin et al. A-110 (MW), HM468334, MK751696, KT262927, KT262857; Lotus villicarpus Andr. (syn. L. eriosolen (Maire) Mader et Podlech): 414, Morocco, prov. Ourzazate, 06.IV.1995, D. Podlech 52619 (M), DQ160281, MK751664, KT262886, KT262816; Tripodion tetraphyllum (L.) Fourr.: 625, Cyprus, 7.5 km to N from Limassol, 11.III.2004, A. Seregin and D. Sokoloff A-240 (MW), HM468340, MK751698, HM468314, HM468274.
References
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.R.; Barbosa Pinto, R.; Boatwright, J.S.; Borges, L.M.; Brown, G.K.; Bruneau, A.; et al. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny: The Legume Phylogeny Working Group (LPWG). Taxon 2017, 66, 44–77. [Google Scholar] [CrossRef]
- Linnaeus, C. Species Plantarum; Holmiae: Stockholm, Sweden, 1753; Tomus 2. [Google Scholar]
- Heyn, C.C. Studies in Lotus. III. The L. angustissimus group. Isr. J. Bot. 1970, 19, 271–292. [Google Scholar]
- Kuprianova, L.A. New annual Lotus. Bot. Mat. Herb. Bot. Inst. 1937, 7, 37–39. (In Russian) [Google Scholar]
- Greuter, W.; Burdet, H.M.; Long, G. Med-Checklist; Conservatoire et Jardin Botaniques de la Ville de Genève: Geneva, Switzerland, 1989; Volume 4, Available online: http://ww2.bgbm.org/mcl/ (accessed on 2 December 2023).
- Kramina, T.E. A contribution to the taxonomic revision of the Lotus angustissimus complex (Leguminosae, Loteae). Wulfenia 2006, 13, 57–92. [Google Scholar]
- Lagasca, M. Caractéres diferenciales de once especies nuevas de plantas y de otras dos poco conocidas. Varied. Cienc. Lit. Art. 1805, 2, 212–217. [Google Scholar]
- Persoon, C.H. Synopsis Plantarum; Treuttel: Paris, France, 1807; Volume 2, p. 354. [Google Scholar]
- Pina, F.J.; Valdés, B. A New Species of Lotus (Leguminosae, Loteae) from the L. angustissimus (sect. Lotus) Complex. Syst. Bot. 2009, 34, 709–714. [Google Scholar] [CrossRef]
- Grant, W.F. A chromosome atlas and interspecific-intergeneric index for Lotus and Tetragonolobus (Fabaceae). Can. J. Bot. 1995, 73, 1787–1809. [Google Scholar] [CrossRef]
- Ferreira, J.; Pedrosa-Harand, A. Lotus cytogenetics. In The Lotus japonicus Genome, Compendium of Plant Genomes; Tabata, S., Stougaard, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 9–20. [Google Scholar] [CrossRef]
- Tanaka, H.; Chotekajorn, A.; Kai, S.; Ishigaki, G.; Hashiguchi, M.; Akashi, R. Determination of Genome Size, Chromosome Number, and Genetic Variation Using Inter-Simple Sequence Repeat Markers in Lotus spp. Cytologia 2016, 81, 95–102. [Google Scholar] [CrossRef]
- An’kova, T.V.; Korolyuk, E.A. IAPT/IOPB chromosome data 26. Taxon 2017, 66, 1487–1488. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Home Page. 2023. Available online: https://www.gbif.org (accessed on 2 December 2023).
- Allan, G.J.; Zimmer, E.A.; Wagner, W.L.; Sokoloff, D.D. Molecular phylogenetic analyses of tribe Loteae (Leguminosae): Implications for classification and biogeography. In Advances in Legume Systematics; Klitgaard, B.B., Bruneau, A., Eds.; Royal Botanic Gardens: Kew, UK, 2003; Part 10; pp. 371–393. [Google Scholar]
- Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Valiejo-Roman, C.M.; Antonov, A.S. Phylogeny of the genus Lotus (Leguminosae, Loteae): Evidence from nrITS sequences and morphology. Can. J. Bot. 2006, 84, 813–830. [Google Scholar] [CrossRef]
- Kramina, T.E.; Degtjareva, G.V.; Samigullin, T.H.; Valiejo-Roman, C.M.; Kirkbride, J.H., Jr.; Volis, S.; Deng, T.; Sokoloff, D.D. Phylogeny of Lotus (Leguminosae: Loteae): Partial incongruence between nrITS, nrETS and plastid markers and biogeographic implications. Taxon 2016, 65, 997–1018. [Google Scholar] [CrossRef]
- Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Sandral, G.; Valiejo-Roman, C.M. New data on nrITS phylogeny of Lotus (Leguminosae, Loteae). Wulfenia 2008, 15, 35–49. [Google Scholar]
- Kuprianova, L.A. Lotus L. In Flora SSSR; Komarov, V.L., Ed.; Akad. Nauk SSSR: Moscow, Russia, 1945; Volume 11, pp. 284–297. (In Russian) [Google Scholar]
- Grossheim, A.A. Flora Kavkaza, 2nd ed.; Akad. Nauk SSSR: Moscow, Russia, 1952; Volume 5. (In Russian) [Google Scholar]
- Tulyaganova, M. Lotus L. In Opredelitel’ Rasteniy Sredney Azii; Kamelin, R., Kovalevskaja, S.S., Nabijev, M.M., Eds.; Izd-vo Fan: Tashkent, Uzbekistan, 1981; Volume 6, pp. 48–50. (In Russian) [Google Scholar]
- Miniaev, N.A.; Ulle, Z.G. Lotus L. In Flora Europejskoj Chasti SSSR; Fedorov, A.A., Ed.; Nauka: Leningrad, Russia, 1987; Volume 6, pp. 103–115. (In Russian) [Google Scholar]
- Kurbatsky, V.I. Lotus L. In Flora Sibiriae; Malyshev, L.I., Polozhy, A.V., Eds.; VO Nauka: Novosibirsk, Russia, 1994; Volume 9, pp. 168–170. (In Russian) [Google Scholar]
- Yakovlev, G.P.; Sytin, A.K.; Roskov, Y.R. Legumes of Northern Eurasia: A Checklist; Royal Botanic Gardens Kew: Kew, UK, 1996; pp. 432–440. [Google Scholar]
- Lock, J.M. Legumes of Africa: A Check-List; Royal Botanic Gardens Kew: Kew, UK, 1989. [Google Scholar]
- Lock, J.M.; Simpson, K. Legumes of West Asia: A Check-List; Royal Botanic Gardens Kew: Kew, UK, 1991. [Google Scholar]
- Andrella, G.C.; Atahuachi Burgos, M.; Bagnatori Sartori, Â.L.; Balan, A.; Bandyopadhyay, S.; Barbosa Pinto, R.; Barrett, R.; Boatwright, J.S.; Borges, L.M.; Legume Phylogeny Working Group (LPWG); et al. The World Checklist of Vascular Plants (WCVP): Fabaceae; 2023v.4; Govaerts, R., Ed.; Royal Botanic Gardens Kew: Kew, UK, 2023. [Google Scholar] [CrossRef]
- Sokoloff, D.D. Morphology and Classification of the Tribe Loteae DC. of the Family Leguminosae. Ph.D. Thesis, Higher Plants Department, Moscow State University, Moscow, Russia, 2003. (In Russian). [Google Scholar]
- Sandral, G.; Remizowa, M.V.; Sokoloff, D.D. A taxonomic survey of Lotus section Pedrosia (Leguminosae, Loteae). Wulfenia 2006, 13, 97–192. [Google Scholar]
- Sandral, G.; Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Hughes, S.; Valiejo-Roman, C.M. Are Lotus creticus and Lotus cytisoides (Leguminosae) closely related species? Evidence from nuclear ribosomal ITS sequence data. Genet. Resour. Crop Evol. 2010, 57, 501–514. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Tarkhnishvili, D.; Gavashelishvili, A.; Mumladze, L. Palaeoclimatic models help to understand current distribution of Caucasian forest species. Biol. J. Linn. Soc. 2012, 105, 231–248. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. 2010. Available online: http://www.simplemappr.net (accessed on 25 December 2023).
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Kramina, T.; Lysova, M.; Samigullin, T.; Schanzer, I.; Özbek, M.; Sokoloff, D. Phylogenetic placement and phylogeography of large-flowered Lotus species (Leguminosae) formerly classified in Dorycnium: Evidence of pre-pleistocene differentiation of western and eastern intraspecific groups. Plants 2021, 10, 260. [Google Scholar] [CrossRef]
- Kramina, T.E.; Lysova, M.V.; Samigullin, T.H.; Özbek, M.U.; Sokoloff, D.D. When Morphology and Biogeography Approximate Nuclear ITS but Conflict with Plastid Phylogeny: Phylogeography of the Lotus dorycnium Species Complex (Leguminosae). Plants 2022, 11, 410. [Google Scholar] [CrossRef]
- Wen, J.; Zimmer, E. Phylogeny and biogeography of Panax L. (the ginseng genus, Araliaceae): Inferences from ITS sequences of nuclear ribosomal DNA. Mol. Phylogenet. Evol. 1996, 6, 167–177. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and diverse origins of the polyploid species. Syst. Bot. 2003, 28, 723–737. [Google Scholar]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Oxelman, B.; Lidén, M.; Berglund, D. Chloroplast rps16 intron phylogeny of the tribe Sileneae (Caryophyllaceae). Plant Syst. Evol. 1997, 206, 393–410. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accu-rate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, M.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, OR, USA, 2003. [Google Scholar]
- Ali, R.H.; Bark, M.; Miró, J.; Muhammad, S.A.; Sjöstrand, J.; Zubair, S.M.; Abbas, R.M.; Arvestad, L. VMCMC: A graphical and statistical analysis tool for Markov chain Monte Carlo traces. BMC Bioinform. 2017, 18, 97. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphological characteristics of species of the Lotus angustissimus group. (A–C) Lotus praetermissus (herbarium specimen MG1: European Russia, Volgograg region, T. Kramina, MW), (A) umbel at anthesis, (B,C) ripe fruits; (D) Lotus macrotrichus (herbarium specimen 819: Turkey, H. and E. Walter 375, B), unripe fruits and keel petal; (E–G) Lotus angustissimus, (herbarium specimen: Azerbaijan, Lerik, V. Egorov, MHA 0246632), (E) flower, (F) ripe fruit, (G) unripe fruit; (H–J) Lotus lourdes-santiagoi (herbarium specimen LS1: Spain, Cádiz, S. Silvestre et B. Valdés 2548, MA), (H) flower, (I) pubescence on standard petal, (J) unripe fruit pubescent along the ventral suture; (K,L) Lotus subbiflorus (herbarium specimen 470: Italy, M. Iberite 15222, MHA), (K) umbel at anthesis, (L) fruits; (M–O) Lotus castellanus (herbarium specimens: Spain, prov. Ávila, Mombeltrán, A. Segura-Zubizarreta 38.109 and 38.111 (sample 471), MHA), (M) flower, (N) ripe fruit, (O) unripe fruit pubescent along the ventral suture; (P,Q) Lotus parviflorus (herbarium specimen 469: Spain, Toledo, A. Segura Zubizarreta 34.567, MHA), (P) umbel at anthesis, (Q) unripe fruit; (R–T) Lotus palustris (herbarium specimens: Israel, Philistaean Plain, 14.7.1926, M. Zohary, and 22.06.1958, M. Zohary and I. Amdursky, MHA), (R) umbel at anthesis, (S) unripe fruit, (T) ripe fruit.
Figure 1.
Morphological characteristics of species of the Lotus angustissimus group. (A–C) Lotus praetermissus (herbarium specimen MG1: European Russia, Volgograg region, T. Kramina, MW), (A) umbel at anthesis, (B,C) ripe fruits; (D) Lotus macrotrichus (herbarium specimen 819: Turkey, H. and E. Walter 375, B), unripe fruits and keel petal; (E–G) Lotus angustissimus, (herbarium specimen: Azerbaijan, Lerik, V. Egorov, MHA 0246632), (E) flower, (F) ripe fruit, (G) unripe fruit; (H–J) Lotus lourdes-santiagoi (herbarium specimen LS1: Spain, Cádiz, S. Silvestre et B. Valdés 2548, MA), (H) flower, (I) pubescence on standard petal, (J) unripe fruit pubescent along the ventral suture; (K,L) Lotus subbiflorus (herbarium specimen 470: Italy, M. Iberite 15222, MHA), (K) umbel at anthesis, (L) fruits; (M–O) Lotus castellanus (herbarium specimens: Spain, prov. Ávila, Mombeltrán, A. Segura-Zubizarreta 38.109 and 38.111 (sample 471), MHA), (M) flower, (N) ripe fruit, (O) unripe fruit pubescent along the ventral suture; (P,Q) Lotus parviflorus (herbarium specimen 469: Spain, Toledo, A. Segura Zubizarreta 34.567, MHA), (P) umbel at anthesis, (Q) unripe fruit; (R–T) Lotus palustris (herbarium specimens: Israel, Philistaean Plain, 14.7.1926, M. Zohary, and 22.06.1958, M. Zohary and I. Amdursky, MHA), (R) umbel at anthesis, (S) unripe fruit, (T) ripe fruit.

Figure 2.
Phylogenetic relationships in Lotus with expanded representation of the L. angustissimus group inferred from Bayesian analysis of the nrITS dataset. Branch lengths are proportional to the number of expected nucleotide substitutions: scale bar corresponds to 0.1 substitutions per site. Numbers above branches are posterior probabilities. Numbers below branches or after slashes are bootstrap support values found in maximum likelihood (ML) analysis of the same dataset (values equal or more than 0.6/60% shown). The colored symbols after the names of the samples indicate belonging to different species or hybrids. See Table 2 and Appendix A for voucher information.
Figure 2.
Phylogenetic relationships in Lotus with expanded representation of the L. angustissimus group inferred from Bayesian analysis of the nrITS dataset. Branch lengths are proportional to the number of expected nucleotide substitutions: scale bar corresponds to 0.1 substitutions per site. Numbers above branches are posterior probabilities. Numbers below branches or after slashes are bootstrap support values found in maximum likelihood (ML) analysis of the same dataset (values equal or more than 0.6/60% shown). The colored symbols after the names of the samples indicate belonging to different species or hybrids. See Table 2 and Appendix A for voucher information.

Figure 3.
Phylogenetic relationships in Lotus with expanded representation of the L. angustissimus group inferred from Bayesian analysis of the plastid DNA dataset. Branch lengths are proportional to the number of expected nucleotide substitution: scale bar corresponds to 0.01 substitutions per site. Numbers above branches are posterior probabilities. Numbers below branches or after slashes are bootstrap support values found in ML analysis of the same dataset (values equal or more than 0.6/60% shown). The colored symbols after the names of the samples indicate belonging to different species or hybrids. See Table 2 and Appendix A for voucher information.
Figure 3.
Phylogenetic relationships in Lotus with expanded representation of the L. angustissimus group inferred from Bayesian analysis of the plastid DNA dataset. Branch lengths are proportional to the number of expected nucleotide substitution: scale bar corresponds to 0.01 substitutions per site. Numbers above branches are posterior probabilities. Numbers below branches or after slashes are bootstrap support values found in ML analysis of the same dataset (values equal or more than 0.6/60% shown). The colored symbols after the names of the samples indicate belonging to different species or hybrids. See Table 2 and Appendix A for voucher information.

Figure 4.
Geographical localities of specimens of the Lotus angustissimus group studied here using molecular methods: (A) L. angustissimus, L. praetermissus, L. macrotrichus, and Lotus hybrids, (B) L. parviflorus, L. subbiflorus, and Lotus hybrids, (C) L. palustris, L. castellanus, and L. lourdes-santiagoi. See Table 2 and Appendix A for voucher information.
Figure 4.
Geographical localities of specimens of the Lotus angustissimus group studied here using molecular methods: (A) L. angustissimus, L. praetermissus, L. macrotrichus, and Lotus hybrids, (B) L. parviflorus, L. subbiflorus, and Lotus hybrids, (C) L. palustris, L. castellanus, and L. lourdes-santiagoi. See Table 2 and Appendix A for voucher information.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Morphological characteristics of species of the Lotus angustissimus group.
| Species | L. angustissimus | L. praetermissus | L. macrotrichus | L. subbiflorus | L. parviflorus | L. castellanus | L. lourdes-santiagoi | L. palustris |
|---|---|---|---|---|---|---|---|---|
| Keel beak | Straight, short | Straight, middle-sized | Straight, long | Curved | ||||
| Fruit length | Long ((2) 3–9 times as long as calyx) | Middle-sized (1.5–3.5 times as long as calyx) | Short (± as long as calyx) | Middle-sized (1.2–3.3 times as long as calyx) | Middle-sized or long (1.6–4.5 times as long as calyx) | |||
| Ovary pilosity | glabrous | Hairy at least in the upper half | Hairy in the upper third or fourth or glabrous | |||||
| Style length | 2.3–3.3 mm | 2.5–3 mm | 5.5–6.5 mm | 2.9–4.4 mm | 3.2–3.9 mm | 3.8–4.9 mm | 3.8–4.9 mm | 3.2–5.1 mm |
| Standard back | glabrous | hairy | glabrous | |||||
| Density of indumentum | low | medium or high | high | medium | high | low | medium | high |
| Life form | annual or perennial | annual | annual | annual, biennial or perennial | annual | annual or perennial | annual | annual |
| Chromosome numbers (2n) | 12, 24 | 12 | 12, 14 (?), 24 | 24 | 12 | 12 | ? | 12, 14 (?), 24 |
Table 2.
Taxa, sample code, voucher information and GenBank accession numbers of Lotus angustissimus group specimens used in molecular analyses. Herbarium codes according to Index Herbariorum. New sequences indicated by an asterisk. Accessions numbers without an asterisk were taken from GenBank.
Table 2.
Taxa, sample code, voucher information and GenBank accession numbers of Lotus angustissimus group specimens used in molecular analyses. Herbarium codes according to Index Herbariorum. New sequences indicated by an asterisk. Accessions numbers without an asterisk were taken from GenBank.
| Sample Code: Voucher Information (Herbarium Code); Coordinates | ITS | trnl-F | rps16 | psbA-trnH |
|---|---|---|---|---|
| Lotus angustissimus L. | ||||
| 472: Australia, Norfolk Island, Collins Head Road, 14.10.1999, B.M. Waterhouse BMW 5510 (NSW); 29.042 S, 167.969 E | DQ166243 | MF158217 | KT262868 | KT262798 |
| ANG01: Turkey, C3 Antalya Kemer, Faselis koyu, 23.06.1978, H. Pesmen 4035 et al. (GAZI); 36.602 N, 30.5601 E | OR789491 * | OR825410 * | OR825408 * | OR920533 * |
| ANG02: Turkey, A2(A) Istanbul, Pasaköy, Omerli Baraj, 30.06.2001, E. Yurdakulol 3729 (ANK); 41.0163 N, 29.2758 E | OR789492 * | OR825411 * | OR825409 * | OR920534 * |
| ANG03: Turkey, A8 Rize: Ardeşen-Findikli, 23.05.1981, A. Güner 3591 (ANK); 41.2201 N, 41.0686 E | OR794120 * | OR825412 * | OR842897 * | OR920535 * |
| ANG04: Turkey, Istanbul, 2 km W of Istanbul Airport, 26.05.2019, T. Kramina, M. Lysova s.n. (MW); 41.2444 N, 28.6667 E | OR794121 * | OR825413 * | OR842898 * | OR920536 * |
| ANG05: Turkey, Istanbul, near Karacaköy, 26.05.2019, T. Kramina, M. Lysova s.n. (MW); 41.4147 N, 28.3722 E | OR804011 * | OR863133 * | OR863092 * | OR920537 * |
| ANG07: Georgia, Tsqaltubo near Kutaisi, 28.10.1935, V. Komarov s.n. (LE); 42.3271 N, 42.6003 E | OR804012 * | OR863134 * | OR863093 * | OR920538 * |
| ANG08: Azerbaijan, N of Lerik, 25.07.1963, A.V. Bobrov, N.N. Tzvelev 1010 (LE); 38.7734 N, 48.4153 E | OR804013 * | OR863135 * | OR863094 * | OR920539 * |
| ANG09: Abkhasia, Gal’sky distr., near Achigvara, 23.06.1989, A. Dolmatova, V. Dorofeev 2656 (LE); 42.683 N, 41.6366 E | OR804014 * | OR863136 * | OR863095 * | OR920539 * |
| ANG10: Azerbaijan, S of the mouth of Lenkoranchay river, 10.07.1963, A.E. Bobrov, N.N. Tzvelev 27 (LE); 38.7505 N, 48.8692 E | OR804015 * | OR863137 * | OR863096 * | OR920541 * |
| ANG11: Israel, Sharon Plain, Netanya, 29.04.1964, C. Heyn s.n. (MHA); 32.3085 N, 34.8595 E | OR804016 * | OR863138 * | OR863097 * | OR920542 * |
| ANG13: France, Corsica, Forêt d’Aïtone bei Evisa, 25.07.1932, Paul Aellen 175 (LE); 42.2765 N, 8.8491 E | OR804017 * | OR863139 * | OR863098 * | OR920543 * |
| ANG15: Hercegovina, supra vicum Jablanica, 07.1933, Sillinger et Deyl s.n. (LE); 43.6605 N, 17.7573 E | OR804018 * | OR863140 * | OR863099 * | OR920544 * |
| ZLA: Bulgaria, M. Rhodope Orientalis, prope urbem Zlatograd, 23.07.1954, N. Stojanov et al. s.n. (MW); 41.383 N, 25.1 E | OR804019 * | MF158219 | OR863100 * | OR920545 * |
| Lotus praetermissus Kuprian. | ||||
| 458: Russia, Volgograd region, Bykovo distr., 1 km to N of Krasnoseletz, Moguta, 20.07.1993, T.E. Kramina s.n. (MW); 49.6726 N, 45.7392 E | DQ166227 | MF158223 | KT262912 | KT262842 |
| 473: Ukraine, Kherson region, 15–18 km to the E from the village Novaya Mayachka, 05.06.1984, N.N. Tzvelev et al. 1630 (MW); 46.607 N, 33.4123 E | DQ168370 | MF158218 | OR863119 * | OR920564 * |
| POY: W. Kazakhstan, Uralskaya Prov., near Poyma railway station, 17.08.1952, S.A. Nikitin s.n. (MW); 51.2 N, 51.5833 E | OR804033 * | MF158220 | OR863120 * | OR920565 * |
| Stn6: Russia, Volgograd region, Sredneakhtubinsky distr., Standartnyy village, 20.06.2008, T. Kramina Stn6 (MW); 48.7169 N, 44.9406 E | OR804034 * | MF158221 | OR863121 * | OR920566 * |
| TH4: Russia, Volgograd region, Upper Golubaya river, 21.06.1939, P. Smirnov s.n. (MW); 49.0875 N; 43.4809 E | OR804035 * | OR863156 * | OR863122 * | OR920567 * |
| TH5: Russia, Tver region, Tverskoy district, 03.10.1988, A.A. Notov s.n. (MW); 56.8596 N, 35.9119 E | OR804036 * | OR863157 * | OR863123 * | OR920568 * |
| TH6: Russia, Voronezh region, village Varvarinki, 28.06.1948, S.V. Golitsyn s.n. (MW); 51.0046 N, 39.9776 E | OR804037 * | OR863158 * | OR863124 * | OR920569 * |
| TH7: Ukraine, Dnepropetrovsk region, Novomoskovsky district, Kocherezhki village, 26.06.1982, Yu. Alekseev s.n. (MW); 48.6688 N, 35.6808 E | OR804038 * | OR863159 * | OR863125 * | OR920570 * |
| Lotus macrotrichus Boiss. | ||||
| 819: Turkey, Anatolien, Tahir Muğla nach Yatağan, 12.06.1955, H. und E. Walter 375 (B); 37.2250 N, 28.3416 E | OR804026 * | OR863147 * | OR863108 * | OR920553 * |
| Lotus lourdes-santiagoi Pina and Valdés | ||||
| LS1: Spain, Cádiz, Sanlúcar de Barrameda, Pinar de Algaida. 09.07.1968, S. Silvestre and B. Valdés 2548 (MA); 36.7737 N, 6.3556 W | OR804024 * | OR863145 * | OR863106 * | OR920551 * |
| LS2: Spain, Cádiz, Municipio de Chiclana de la Frontera, Estero La Isleta, 18.05.2013, P. Barbera and A. Quintanar 749 PB (MA); 36.4122 N, 6.1897 W | OR804025 * | OR863146 * | OR863107 * | OR920552 * |
| Lotus palustris Willd. | ||||
| 426: Turkey, Antalya, near village Tekirova, 17.10.1999, S.R. Majorov s.n. (MW); 36.5010 N, 30.5255 E | DQ160275 | OR863148 * | OR863109 * | OR920554 * |
| 869: Greece, Naxos, 24.10.1991, N. Böhling 582 (B); 37.1027 N, 25.3804 E | KT250896 | - | - | - |
| 875: Israel, 22.06.1958, M. Zohary and I. Amdursky s.n. (B); 31.9331 N, 34.7048 E | KT250897 | OR863149 * | OR863110 * | OR920555 * |
| PAL3: Turkey, Denizli suburb, ruderal place, 22.07.1999, V.D. Bochkin s.n. (MHA); 37.785 N, 29.0837 E | OR804027 * | OR863150 * | OR863111 * | OR920556 * |
| PAL4: Turkey, C2 Muğla, Fethiye, J. Akman 15120 (ANK); 36.6606 N, 29.126 E | OR804028 * | OR863151 * | OR863112 * | OR920557 * |
| PAL5: Flora Palaestine, Hula Plain, W. of Gonen Marsh, 24.05.1963, M. Zohary and U. Plitman 62457 (LE); 33.0699 N, 35.5998 E | OR804029 * | OR863152 * | OR863113 * | OR920558 * |
| Lotus castellanus Boiss. and Reut. | ||||
| 417: Portugal, Beira Litoral, 12.07.1977, Malato-Beliz and J.A. Guerra 13585 (MW); 40.9634 N, 8.6477 W | DQ160272 | MF158216 | OR863101 * | OR920546 * |
| 471: Spain, prov. Ávila, Mombeltrán, 01.07.1990, A. Segura Zubizarreta 38.111 (MHA); 40.2169 N, 5.0298 W | DQ166238 | MF158215 | KT262873 | KT262803 |
| CAS1: Spain, Ciudad Real: Argamasilla de Calatrava, 26.10.2001, M. Bellet et al. RGC431 (MA); 38.7524 N, 3.9446 W | OR804020 * | OR863141 * | OR863102 * | OR920547 * |
| CAS2: Spain, Toledo, Hinojosa del Monte, 15.07.1977, A. Segura Zubizarreta 15.112 (LE); 40.1043 N, 4.7225 W | OR804021 * | OR863142 * | OR863103 * | OR920548 * |
| PAL1: Spain, Cádiz: Algodonales. Rio guadalete. Puerto de la Nava., 21.07.1983, A. Aparicio s.n. (MA); 36.7562 N, 5.3915 W | OR804022 * | OR863143 * | OR863104 * | OR920549 * |
| PAL2: Portugal, Beira Litoral: Barrinha de Esmoriz, 12.07.1977, Malato-Beliz and J.A. Guerra 13585 (MA); 40.9634 N, 8.6477 W | OR804023 * | OR863144 * | OR863105 * | OR920550 * |
| Lotus parviflorus Desf. | ||||
| 469: Spain, Toledo, Loc. Talavera de Ca Reina: Gamonal, 09.05.1987, A. Segura Zubizarreta 34.567 (MHA); 39.9567 N, 4.8409 W | DQ166230 | MF314955 | MW498357 | OL753560 |
| PRV1: Morocco: c. 10 km W of Bab Berred along road to Bab Taza, 28.05.2002, S.L. Jury et al. 19385 (MA); 35.0193 N, 4.9857 W | MN545738 | MN553708 | OR863114 * | OR920559 * |
| PRV2: Portugal, Algarve, Rogil, 20.04.2011, T. Buira, J. Calvo and L. Mauro TB1529 (MA); 37.3633 N, 8.8006 W | MN545739 | MN553709 | OR863115 * | OR920560 * |
| PRV3: Spain, Islas Baleares, Menorca, 01.05.2013, J.L. Fernandez-Alonso, JFA30449 (MA); 40.0372 N, 3.9783 E | OR804030 * | OR863153 * | OR863116 * | OR920561 * |
| PRV4: Spain, Córdoba, La Jarosa, alt. 450 m, 03.05.1977, A. Segura Zubizarreta 14.988 (LE); 37.9067 N, 4.9103 W | OR804031 * | OR863154 * | OR863117 * | OR920562 * |
| PRV5: Madeira, Machico, 04.04.1985, J.R. Press s.n. (BM); 32.7314 N, 16.791 W | OR804032 * | OR863155 * | OR863118 * | OR920563 * |
| Lotus subbiflorus Lag. | ||||
| 457: Australia, New South Wales, 15.03.1992, P.G. Kodela et al. 163 (NSW); 35.1537 S, 150.7002 E | DQ166237 | MF158214 | OR863127 * | OR920572 * |
| 470: Italy, prov. Latina, Lazio, Pianura Pontina, 15.06.1991, M. Iberite 15222 (MHA); 41.6420 N, 12.9153 E | DQ166231 | MF158212 | KT262925 | KT262855 |
| 475: France, dep. Vaucluse, Avignon, 20.06.1988, G. Dutartre 570 (MHA); 43.8896 N, 5.0116 E | DQ168369 | OR863161 * | OR863128 * | OR920573 * |
| 476: Spain, Soria, Granja de Torrehermosa (Badajoz), 17.05.1987, A. Segura Zubizarreta 34.566 (MHA); 38.3099 N, 5.597 W | OR913548 * | OR863162 * | OR863129 * | OR920574 * |
| ANG06: Madeira, Ribeira de Janela to Seixal, 17.06.1985, J.R. Press s.n. (BM); 32.8469 N, 17.156 W | OR804040 * | OR863163 * | OR863130 * | OR920575 * |
| SUBB1: Madeira, Levada do Furado, 23.10.1984, M.J. Short 117 (BM); 32.735 N, 16.8861 W | OR913549 * | OR863164 * | OR863131 * | OR920576 * |
| TH2: Azores, São Miguel Island, 04.04.2013, A. Seregin, I. Seregina s.n. (MW); 37.799 N, 25.4854 W | OR804041 * | OR863165 * | OR863132 * | OR920577 * |
| Lotus hybrids | ||||
| ANG12: Portugal, Mouquim, 08.06.1973, A. Fernandes et al. 12403 (MHA); 41.4350 N, 8.5274 W | OR913550 * | OR920527 * | OR920524 * | OR920530 * |
| ANG14: Italy, Liguria occid., Varazze, 18.06.1908, L.G. Gresino s.n. (LE); 44.3654 N, 8.5721 E | OR913551 * | OR920528 * | OR920525 * | OR920531 * |
| ANG17: Iran, Astrabad [Gorgan] prov., S of Gumbet [Gonbad-e Kavus], 01.05.1914, A. Michelson s.n. (LE); 37.2475 N, 55.1705 E | OR913552 *, OR913553 * | OR920529 * | OR920526 * | OR920532 * |
| Lotus rectus L. | ||||
| TH1: Israel, Hula Plain, 29.01.2014, A. Seregin et al. A903 (MW); 33.0688 N, 35.597 E | OR804039 * | OR863160 * | OR863126 * | OR920571 * |
Table 3.
Diagnostic morphological characteristics of Lotus castellanus, L. palustris and intermediate specimens. Median and lower–upper quartile values (in brackets) are given for each quantitative character for “pure” species (taken from the previous study [6]). Observed values are given for the specimens PAL1 and PAL2. Character states for each character are indicated as follows: C—like in L. castellanus, P—like in L. palustris, C/P—intermediate, ?—unknown.
Table 3.
Diagnostic morphological characteristics of Lotus castellanus, L. palustris and intermediate specimens. Median and lower–upper quartile values (in brackets) are given for each quantitative character for “pure” species (taken from the previous study [6]). Observed values are given for the specimens PAL1 and PAL2. Character states for each character are indicated as follows: C—like in L. castellanus, P—like in L. palustris, C/P—intermediate, ?—unknown.
| Lotus castellanus | PAL1 Spain (MA) | PAL2 Portugal (MA) | Lotus palustris | |
|---|---|---|---|---|
| Density of indumentum | Low | High | Medium | High |
| C | P | C/P | P | |
| Ovary pilosity along the ventral suture | In the upper 1/2 | glabrous | Unknown | In the upper 1/3–1/4 or glabrous |
| C | P | ? | P | |
| Style length, mm | 4.45 (4.3–4.7) | 3.5 | 4.5 | 4.1 (3.8–4.4) |
| C | P | C | P | |
| Fruit length, mm | 9.5 (7.2–13) | 24 | No fruits | 17 (14–18) |
| C | P | ? | P | |
| Fruit-calyx index | 1.98 (1.5–2.5) | 2.6 | No fruits | 2.81 (2.6 –3.1) |
| C | P | ? | P | |
| Terminal leaflet length in mm | 9.55 (7.8–11.5) | 7.65 (7.5; 7.8) | 10.8 (9.7; 12) | 14.1 (12–17.4) |
| C | C | C/P | P | |
| Taxonomic identification based on morphology | L. castellanus | Intermediate individual closer to L. palustris | Intermediate individual closer to L. castellanus | L. palustris |
Table 4.
Diagnostic morphological characteristics of Lotus angustissimus, L. subbiflorus and intermediate specimens. Median and lower–upper quartile values (in brackets) are given for each quantitative character for “pure” species (taken from the previous study [6]). Observed values are given for the specimens ANG12, ANG14 and ANG17. Character states for each character are indicated as follows: A—like in L. angustissimusus, S—like in L. subbiflorus, A/S—intermediate.
Table 4.
Diagnostic morphological characteristics of Lotus angustissimus, L. subbiflorus and intermediate specimens. Median and lower–upper quartile values (in brackets) are given for each quantitative character for “pure” species (taken from the previous study [6]). Observed values are given for the specimens ANG12, ANG14 and ANG17. Character states for each character are indicated as follows: A—like in L. angustissimusus, S—like in L. subbiflorus, A/S—intermediate.
| Lotus angustissimus | ANG14 Italy (LE) | ANG17 Iran (LE) | ANG12 Portugal (MHA) | Lotus subbiflorus | |
|---|---|---|---|---|---|
| Keel length, mm | 3.8 (3.6–4.1) | 4.0 | 4.4 | 4.1 | 4.4 (4–4.6) |
| A | A/S | S | A/S | S | |
| Keel beak length, mm | 1.3 (1.2–1.8) | 1.6 | 2.1 | 2.1 | 2.3 (2.2–2.5) |
| A | A | S | S | S | |
| Number of flowers in umbel | 1.5 (1.5–1.5) | 3 (2; 4) | 1.5 (1; 2) | 2 (1; 3) | 2.5 (2–2.5) |
| A | S | A | A/S | S | |
| Style length, mm | 2.9 (2.7–3.2) | 3.0 (2.8; 3.2) | 3.1 (2.7; 3.5) | 3.6 (3.5; 3.7) | 3.9 (3.5–4) |
| A | A | A | S | S | |
| Number of seeds in pod | 22 (20–26) | 17 | 12 | 13; 14 | 11 (9 –12) |
| A | A/S | S | A/S | S | |
| Fruit length, mm | 20.5 (16–22) | 18 (16; 20) | 16.75 (13.5; 20) | 15 | 11 (11–14) |
| A | A | A | A/S | S | |
| Keel index | 0.36 (0.34–0.43) | 0.41 | 0.48 | 0.59 | 0.55 (0.51–0.58) |
| A | A | A/S | S | S | |
| Fruit-calyx index | 3.45 (3–4.8) | 3 (2.67; 3.33) | 2.7 | 3 | 2.37 (2.3–2.6) |
| A | A | A/S | A | S | |
| Taxonomic identification based on morphology | L. angustissimus | Intermediate individual closer to L. angustissimus | Intermediate between L. subbiflorus and L. angustissimus | Intermediate between L. subbiflorus and L. angustissimus | L. subbiflorus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kramina, T.E.; Hadziev, T.R.; Samigullin, T.H. The Lotus angustissumus Group (Fabaceae): Can Phylogenetic Patterns Be Accommodated by a Taxonomic Concept? Plants 2024, 13, 101. https://doi.org/10.3390/plants13010101
AMA Style
Kramina TE, Hadziev TR, Samigullin TH. The Lotus angustissumus Group (Fabaceae): Can Phylogenetic Patterns Be Accommodated by a Taxonomic Concept? Plants. 2024; 13(1):101. https://doi.org/10.3390/plants13010101
Chicago/Turabian StyleKramina, Tatiana E., Tamerlan R. Hadziev, and Tahir H. Samigullin. 2024. "The Lotus angustissumus Group (Fabaceae): Can Phylogenetic Patterns Be Accommodated by a Taxonomic Concept?" Plants 13, no. 1: 101. https://doi.org/10.3390/plants13010101
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.