
Production of Non-Volatile Metabolites from Sooty Molds and Their Bio-Functionalities

 , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. DNA Extraction, Amplification and Sequencing
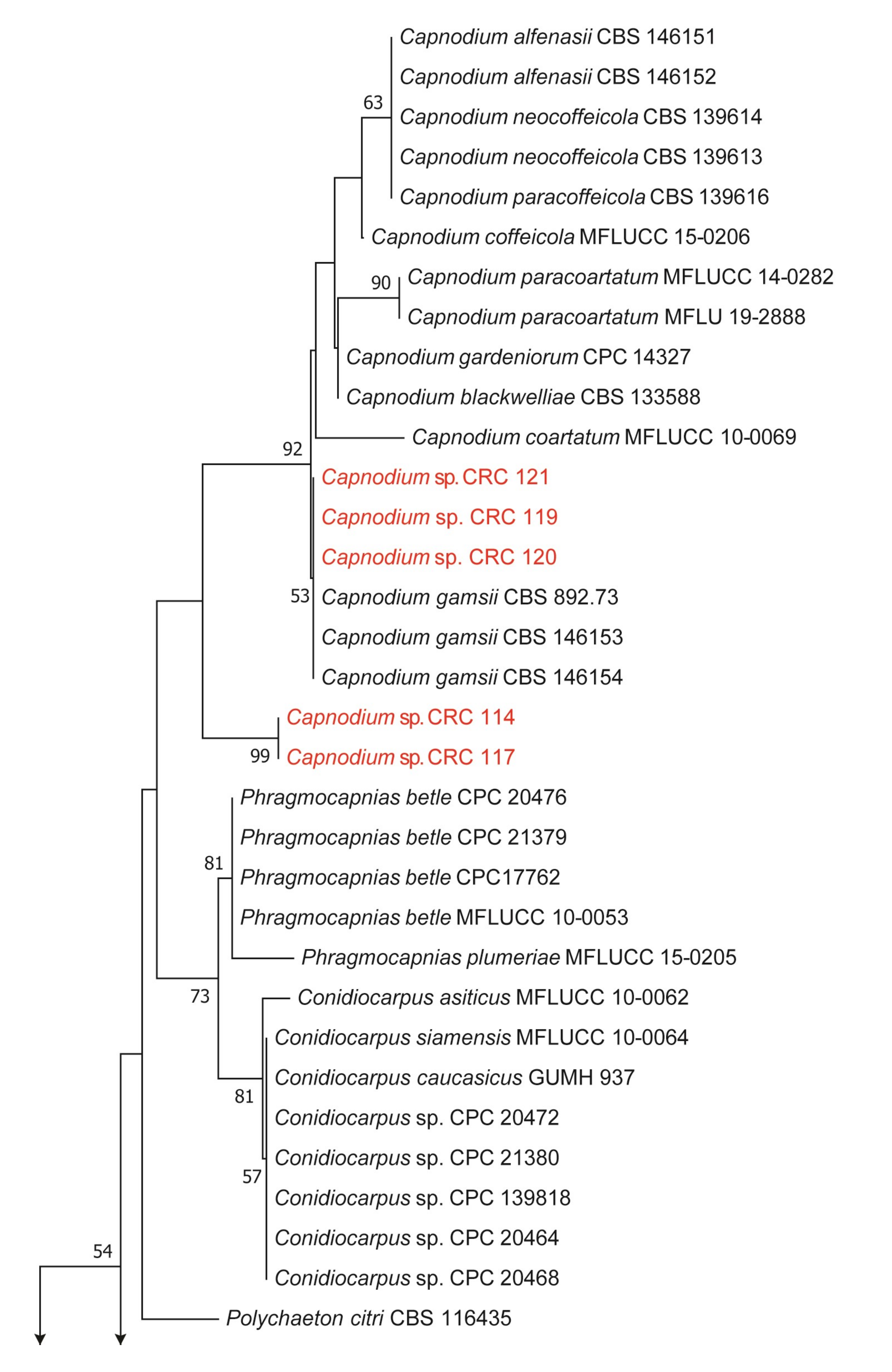
2.3. Phylogenetic Analysis
2.4. Preparation of Fungal Extract
2.5. Protein Analysis
2.6. Quantitative Polyphenol Analysis
2.6.1. Total Phenolic Content
2.6.2. Total Flavonoid Content
2.7. Antioxidant Activities
2.7.1. DPPH• Radical Scavenging Activity
2.7.2. ABTS + Radical Cation Decolorization Assay
2.8. Antifungal Activities
2.8.1. Microorganisms
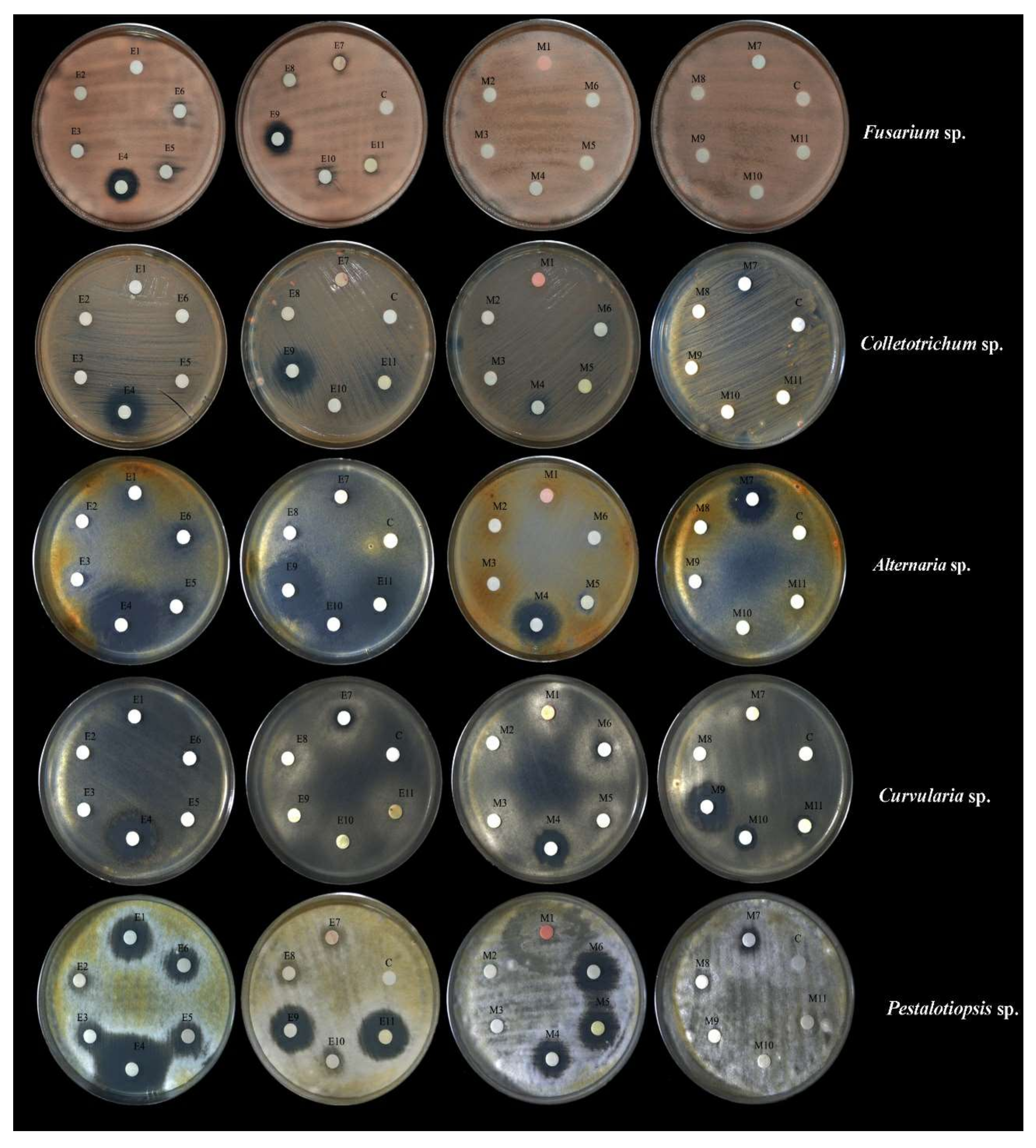
2.8.2. Determination of Antifungal Activity
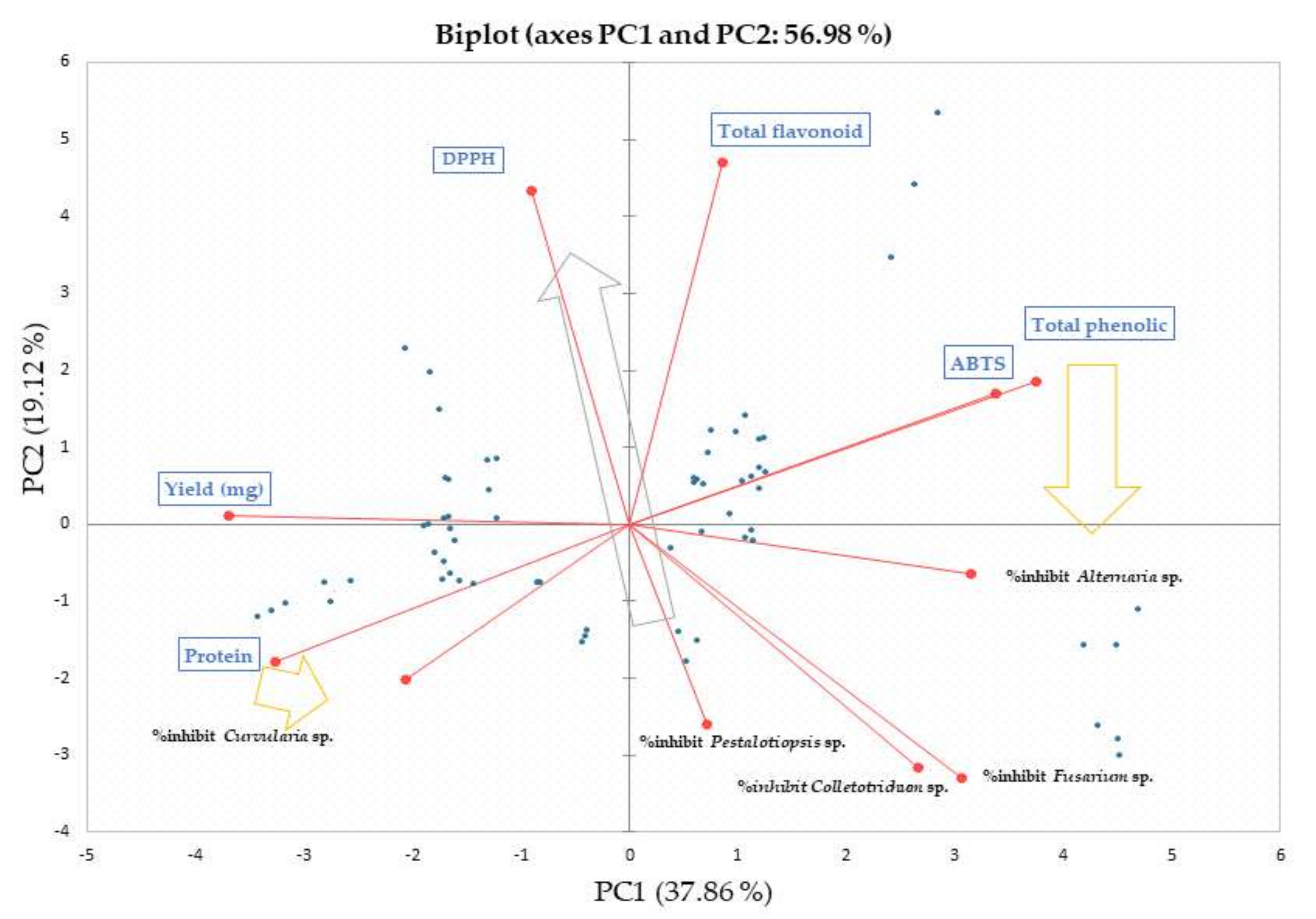
2.8.3. Chemometric and Statistical Analyses
3. Results
3.1. Characteristics and Phylogenetic Analyses
3.2. Quantitative Analysis of the Metabolites
3.3. Biological Activities
3.3.1. Antioxidant Assay
3.3.2. Antifungal Activities
3.4. Chemometric Relations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Callan, B.; Carris, L.M. Fungi on living plant substrata, including fruits. In Biodiversity of Fungi; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Academic Press: Burlington, CA, USA, 2004; pp. 105–126. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef]
- Chomnunti, P.; Schoch, C.L.; Aguirre-Hudson, B.; Ko-Ko, T.W.; Hongsanan, S.; Jones, E.B.G.; Kodsueb, R.; Phookamsak, R.; Chukeatirote, E.; Bahkali, A.H.; et al. Capnodiaceae. Fungal Divers. 2011, 51, 103–134. [Google Scholar] [CrossRef]
- Flessa, F.; Harjes, J.; Cáceres, M.E.S.; Rambold, G. Comparative analyses of sooty mould communities from Brazil and Central Europe. Mycol. Prog. 2021, 20, 869–887. [Google Scholar] [CrossRef]
- Chomnunti, P.; Bhat, D.J.; Jones, E.B.G.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Trichomeriaceae, a new sooty mould family of Chaetothyriales. Fungal Divers. 2012, 56, 63–76. [Google Scholar] [CrossRef]
- Reynolds, D.R. Capnodiaceous sooty mold phylogeny. Can. J. Bot. 1998, 76, 2125–2130. [Google Scholar] [CrossRef]
- Winka, K.; Eriksson, O.E.; Bång, Å. Molecular evidence for recognizing the Chaetothyriales. Mycologia 2018, 90, 822–830. [Google Scholar] [CrossRef]
- Hongsanan, S.; Tian, Q.; Hyde, K.; Chomnunti, P. Two new species of sooty moulds, Capnodium coffeicola and Conidiocarpus plumeriae in Capnodiaceae. Mycosp 2015, 6, 814–824. [Google Scholar] [CrossRef]
- Wisetkomolmat, J.; Inta, A.; Krongchai, C.; Kittiwachana, S.; Jantanasakulwong, K.; Rachtanapun, P.; Sommano, S.R. Ethnochemometric of plants traditionally utilised as local detergents in the forest dependent culture. Saudi J. Biol. Sci. 2021, 28, 2858–2866. [Google Scholar] [CrossRef] [PubMed]
- Castaño, C.; Alday, J.; Lindahl, B.; de Aragón, M.J.; Miguel, S.D.; Colinas, C.; Parladé, J.; Pera, J.; Bonet, J. Lack of thinning effects over inter-annual changes in soil fungal community and diversity in a Mediterranean pine forest. For. Ecol. Manag. 2018, 424, 420–427. [Google Scholar] [CrossRef]
- McGuire, K.L.; Bent, E.; Borneman, J.; Majumder, A.; Allison, S.D.; Tresederi, K.K. Functional diversity in resource use by fungi. Ecology 2010, 91, 2324–2332. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Kenerley, C.M. Secondary metabolism in Trichoderma--a genomic perspective. Microbiology 2012, 158, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusztahelyi, T.; Holb, I.; Pócsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef] [Green Version]
- Renshaw, J.C.; Robson, G.D.; Trinci, A.P.J.; Wiebe, M.G.; Livens, F.R.; Collison, D.; Taylor, R.J. Fungal siderophores: Structures, functions and applications. Mycol. Res. 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Caicedo, N.H.; Davalos, A.F.; Puente, P.A.; Rodríguez, A.Y.; Caicedo, P.A. Antioxidant activity of exo-metabolites produced by Fusarium oxysporum: An endophytic fungus isolated from leaves of Otoba gracilipes. Microbiologyopen 2019, 8, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kaushik, N.; Kumar, S.; Kaushik, N. Metabolites of endophytic fungi as novel source of biofungicide: A review. Phytochem Rev. 2013, 11, 507–522. [Google Scholar] [CrossRef]
- Ribeiro, B.; Valentão, P.; Baptista, P.; Seabra, R.M.; Andrade, P.B. Phenolic compounds, organic acids profiles and antioxidative properties of beefsteak fungus (Fistulina hepatica). Food Chem. Toxicol. 2007, 45, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Yu, J.-H.; Zhu, K.; Wang, Y.; Cheng, Z.-Q.; Jiang, C.-S.; Dai, J.-G.; Wu, J.; Zhang, H. Phenolic bisabolane sesquiterpenoids from a Thai mangrove endophytic fungus, Aspergillus sp. xy02. Fitoterapia 2018, 127, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Senwanna, C.; Wanasinghe, D.; Bulgakov, T.; Wang, Y.; Bhat, D.J.; Tang, A.; Mortimer, P.; Xu, J.; Hyde, K.D.; Phookamsak, R. Towards a natural classification of Dothidotthia and Thyrostroma in Dothidotthiaceae (Pleosporineae, Pleosporales). Mycosphere 2019, 10, 701–738. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Amplifications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Haituk, S.; Suwannarach, N.; Hongsanan, S.; Senwanna, C.; Cheewangkoon, R. New epiphytic sooty molds: Alloscorias syngonii (Readerielliopsidaceae) from Thailand. Phytotaxa 2021, 507, 271–282. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Haituk, S.; Pakdeeniti, P.; Al-Sadi, A.M.; Hongsanan, S.; Chomnunti, P.; Cheewangkoon, R. Phaeosaccardinula coffeicola and Trichomerium chiangmaiensis, two new species of Chaetothyriales (Eurotiomycetes) from Thailand. Mycosphere 2018, 9, 769–778. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumla, J.; Suwannarach, N.; Tanruean, K.; Lumyong, S. Comparative Evaluation of Chemical Composition, Phenolic Compounds, and Antioxidant and Antimicrobial Activities of Tropical Black Bolete Mushroom Using Different Preservation Methods. Foods 2021, 10, 781. [Google Scholar] [CrossRef] [PubMed]
- Sangta, J.; Wongkaew, M.; Tangpao, T.; Withee, P.; Haituk, S.; Arjin, C.; Sringarm, K.; Hongsibsong, S.; Sutan, K.; Pusadee, T.; et al. Recovery of Polyphenolic Fraction from Arabica Coffee Pulp and Its Antifungal Applications. Plants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Adedapo, A.A.; Jimoh, F.O.; Koduru, S.; Afolayan, A.J.; Masika, P.J. Antibacterial and antioxidant properties of the methanol extracts of the leaves and stems of Calpurnia aurea. BMC Complement. Altern. Med. 2008, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; de Hoog, G.S.; Selbmann, L. Extremotolerant rock inhabiting black fungi from Italian monumental sites. Fungal Divers. 2015, 76, 75–96. [Google Scholar] [CrossRef]
- Hyde, K.; Silva, N.; Abeywickrama, P.; Phookamsak, R.; Jeewon, R.; Jayawardena, R.; Boonmee, S.; Manawasinghe, I.; Tennakoon, D.; Wanasinghe, D.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Hongsanan, S.; Tian, Q.; Hyde, K.D.; Hu, D.M. The asexual morph of Trichomerium gloeosporum. Mycosphere 2016, 7, 1473–1479. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Spigno, G.; Tramelli, L.; De Faveri, D.M. Effects of extraction time, temperature and solvent on concentration and antioxidant activity of grape marc phenolics. J. Food Eng. 2007, 81, 200–208. [Google Scholar] [CrossRef]
- Selim, K.; Elkhateeb, W.; Tawila, A.; El-Beih, A.; Abdel-Rahman, T.; El Diwany, A.; Ahmed, E. Antiviral and Antioxidant Potential of Fungal Endophytes of Egyptian Medicinal Plants. Fermentation 2018, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Pintać, D.; Majkić, T.; Torović, L.; Orčić, D.; Beara, I.; Simin, N.; Mimica–Dukić, N.; Lesjak, M. Solvent selection for efficient extraction of bioactive compounds from grape pomace. Ind. Crops Prod. 2018, 111, 379–390. [Google Scholar] [CrossRef]
- Gonçalves, T.D.; Viana, L.B.B.; Lopes, F.R.; Ferreira Duarte, W.; Gomes Cardoso, P. Antioxidant activity and phenolic compounds of the extract from pigment-producing fungi isolated from Brazilian caves. Biocatal. Agric. 2018, 16, 148–154. [Google Scholar] [CrossRef]
- Silva, J.; Fill, T.; Ferreira-Silva, B.; Rodrigues-Fo, E. Diclavatol and tetronic acids from Penicillium griseoroseum. Nat. Prod. Res. 2012, 27, 9–16. [Google Scholar] [CrossRef]
- Wösten, H.A.; Moukha, S.M.; Sietsma, J.H.; Wessels, J.G. Localization of growth and secretion of proteins in Aspergillus niger. J. Gen. Microbiol. 1991, 137, 2017–2023. [Google Scholar] [CrossRef] [Green Version]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal Agents in Agriculture: Friends and Foes of Public Health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Kingsley, K.L.; Bergen, M.S.; Kowalski, K.P.; White, J.F. Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis. Microorganisms 2018, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Prom, L. The Effects of Fusarium thapsinum, Curvularia lunata, and Their Combination on Sorghum Germination and Seed Mycoflora. J. New Seeds 2004, 6, 39–49. [Google Scholar] [CrossRef]
- Suksathan, R.; Rachkeeree, A.; Puangpradab, R.; Kantadoung, K.; Sommano, S.R. Phytochemical and nutritional compositions and antioxidants properties of wild edible flowers as sources of new tea formulations. NFS J. 2021, 24, 15–25. [Google Scholar] [CrossRef]
- Wongkaew, M.; Tinpovong, B.; Sringarm, K.; Leksawasdi, N.; Jantanasakulwong, K.; Rachtanapun, P.; Hanmoungjai, P.; Sommano, S.R. Crude Pectic Oligosaccharide Recovery from Thai Chok Anan Mango Peel Using Pectinolytic Enzyme Hydrolysis. Foods 2021, 10, 627. [Google Scholar] [CrossRef] [PubMed]
- Negreiros, C.P.L.; Silva, E.O.; Chagas-Paula, D.A.; Hortolan, L.J.H.; Ikegaki, M. Importance and Implications of the Production of Phenolic Secondary Metabolites by Endophytic Fungi: A Mini-Review. Mini Rev. Med. Chem. 2016, 16, 259–271. [Google Scholar] [CrossRef]
- Mousa, W.K.; Raizada, M.N. The diversity of anti-microbial secondary metabolites produced by fungal endophytes: An interdisciplinary perspective. Front. Microbiol. 2013, 4, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanichamy, P.; Krishnamoorthy, G.; Kannan, S.; Marudhamuthu, M. Bioactive potential of secondary metabolites derived from medicinal plant endophytes. Egypt. J. Basic Appl. Sci. 2018, 5, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Shull, T.E.; Kurepa, J.; Miller, R.D.; Martinez-Ochoa, N.; Smalle, J.A. Inhibition of Fusarium oxysporum f. sp. nicotianae Growth by Phenylpropanoid Pathway Intermediates. Plant Pathol. J. 2020, 36, 637–642. [Google Scholar] [CrossRef]
- Steinkellner, S.; Mammerler, R. Effect of flavonoids on the development of Fusarium oxysporum f. sp. lycopersici. J. Plant Interact. 2007, 2, 17–23. [Google Scholar] [CrossRef]
- Baker, S.E. Grand Challenges in Fungal Biotechnology. Front. Fungal Biol. 2020, 1, 626551. [Google Scholar] [CrossRef]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganendren, R.; Widmer, F.; Singhal, V.; Wilson, C.; Sorrell, T.; Wright, L. In vitro antifungal activities of inhibitors of phospholipases from the fungal pathogen Cryptococcus neoformans. Antimicrob. Agents Chemother. 2004, 48, 1561–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, B.; Nagpure, A.; Gupta, R.K. Fungal cell-wall lytic enzymes, antifungal metabolite(s) production, and characterization from Streptomyces exfoliatus MT9 for controlling fruit-rotting fungi. J. Basic Microbiol. 2014, 54, 1295–1309. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Host | Collected Location | GenBank Accession Number | Morphological Description | |
|---|---|---|---|---|---|
| LSU | ITS | ||||
| Leptoxyphium sp. CRC 98 | Durian (Durio ziberhinus) | Chanthaburi, Thailand | MZ725032 | MZ725022 | Conidiomata synnematous, simple, 27–43 × 28–45 μm. Conidia ellipsoidal, hyaline, aseptate and guttulate, 4.1–6.7 × 2.1–2.7 μm |
| Leptoxyphium sp. CRC 101 | Sugar Apple (Annona squamosal) | Chiang Rai, Thailand | MZ725033 | MZ725023 | Conidiomata synnematous, simple, 27.5–43 × 28.5–46 μm. Conidia ellipsoidal, hyaline, aseptate and guttulate, 4.1–6.8 × 2–2.6 μm |
| Leptoxyphium sp. CRC 102 | Bird of paradise (Strelitzia reginae) | Chiang Mai, Thailand | MZ725034 | MZ725024 | Conidiomata synnematous, simple, 27–43 × 28–45 μm. Conidia ellipsoidal, hyaline, aseptate and guttulate, 4–6.7 × 2.2–2.8 μm |
| Leptoxyphium sp. CRC 114 | Globe amaranth (Gomphrena globosa) | Chiang Mai, Thailand | MZ725035 | MZ725025 | Conidiomata synnematous, simple, 28–44 × 28.5–46 μm. Conidia ellipsoidal, hyaline, aseptate and guttulate, 4.1–6.7 × 2.1–2.7 μm |
| Leptoxyphium sp. CRC 110 | Queen’s crepe-myrtle (Lagerstroemia speciosa) | Chiang Mai, Thailand | MZ725036 | MZ725026 | Conidiomata synnematous, simple, 27–43 × 28–45 μm. Conidia ellipsoidal, hyaline, aseptate and guttulate, 4.1–6.7 × 2.1–2.7 μm |
| Capnodium sp. CRC 108 | Guava (Psidium guajava) | Songkhla, Thailand | MZ725037 | - | Conidiomata synnematous, simple or branched, 25–45 × 16.5–30 μm. Conidia oblong to ellipsoid, hyaline, aseptate, 4.5–6.5 × 1.8–2.7 μm |
| Capnodium sp. CRC 117 | Sapodilla (Manilkara zapota) | Songkhla, Thailand | MZ725038 | - | Conidiomata synnematous, simple or branched, 24–43 × 17–29 μm. Conidia oblong to ellipsoid, hyaline, aseptate, 4.5–7 × 2–2.9 μm |
| Capnodium sp. CRC 119 | Areca palm (Dypsis lutescens) | Chiang Mai, Thailand | MZ725039 | MZ725027 | Conidiomata synnematous, simple or branched, 23–44 × 16–29 μm. Conidia oblong to ellipsoid, hyaline, aseptate, 4.5–6.5 × 1.7–2.5 μm |
| Capnodium sp. CRC 120 | Orchid (Dendrobium hybrid) | Chiang Mai, Thailand | MZ725040 | MZ725028 | Conidiomata synnematous, simple or branched, 23–44 × 16–29 μm. Conidia oblong to ellipsoid, hyaline, aseptate, 4–5.5 × 1.9–2.4 μm |
| Capnodium sp. CRC 121 | Queen’s crepe-myrtle (Lagerstroemia speciosa) | Chiang Mai, Thailand | MZ725041 | MZ725029 | Conidiomata synnematous, simple or branched, 23–42 × 16–28 μm. Conidia oblong to ellipsoid, hyaline, aseptate, 4–6 × 1.8–2.5 μm |
| Trichomerium deniqulatum CRC 141 | White meranti (Shorea roxburghii) | Chiang Mai, Thailand | MZ373260 | MZ373260 | lateral arms from 1–2 globose to subglobose, Conidial arms 1–3, rarely 4–5 septate, hyaline to pale brown, 12–19 × 2.8–3.5 μm |
| No. | Isolate | Yield (mg) | TPC (mg GAE/g) | TFC (mg CE/g) | DPPH Assay (%) | ABTS Assay (%) | Total Protein (mg/mL) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | ||
| 1 | CRC 98 | 16.63 ± 0.03 h | 34.30 ± 0.12 c | 0.92 ± 0.02 bc | 0.20 ± 0.004 a | 0.08 ± 0.009 a | 0.12 ± 0.002 abc | 55.31 ± 1.33 f | 69.26 ± 0.07 d | 98.33 ± 2.00 c | 5.45 ± 5.45 a | 0.13 ± 0.001 b | 3.13 ± 0.15 b |
| 2 | CRC 101 | 7.50 ± 0.00 e | 43.067 ± 0.03 d | 1.49 ± 0.09 d | 0.14 ± 0.01 a | 0.21 ± 0.04 ab | 0.08 ± 0.01 ab | 57.26 ± 4.80 f | 50.46 ± 7.05 bc | 98.21 ± 0.00 c | 34.43 ± 2.64 bc | 0.09 ± 0.006 a | 1.83 ± 0.08 a |
| 3 | CRC 102 | 5.83 ± 0.03 c | 52.13 ± 0.09 f | 1.40 ± 0.08 d | 0.18 ± 0.01 a | 0.10 ± 0.06 ab | 0.28 ± 0.04 c | 43.90 ± 2.39 e | 48.71 ± 1.93 abc | 97.42 ± 0.47 c | 51.81 ± 4.50 c | 0.14 ± 0.001 b | 6.22 ± 0.47 d |
| 4 | CRC 108 | 3.90 ± 0.06 a | 23.20 ± 0.06 b | 1.59 ± 0.05 de | 0.20 ± 0.008 a | n/a | 0.17 ± 0.03 abc | 29.41 ± 4.88 bc | 52.09 ± 0.86 c | 92.50 ± 2.20 c | n/a | 0.08 ± 0.0002 a | 1.71 ± 0.13 a |
| 5 | CRC 110 | 6.30 ± 0.12 d | 62.13 ± 0.07 h | 1.03 ± 0.01 c | 0.19 ± 0.003 a | 0.10 ± 0.06 ab | 0.16 ± 0.08 abc | 39.14 ± 3.69 de | 43.16 ± 0.24 abc | 99.11 ± 0.52 c | 6.18 ± 3.88 a | 0.10 ± 0.002 a | 5.14 ± 0.24 c |
| 6 | CRC 114 | 8.13 ± 0.09 f | 58.73 ± 0.03 g | 0.73 ± 0.04 ab | 0.19 ± 0.001 a | 0.16 ± 0.02 ab | 0.23 ± 0.02 bc | 28.63 ± 0.51 bc | 52.11 ± 1.07 c | 92.22 ± 1.37 c | 18.64 ± 18.64 ab | 0.21 ± 0.006 c | 1.18 ± 0.008 a |
| 7 | CRC 117 | 4.03 ± 0.03 a | 74.13 ± 0.03 i | 1.36 ± 0.08 d | 0.44 ± 0.05 b | 0.001 ± 0.002 a | 0.71 ± 0.03 d | 10.49 ± 4.03 a | 77.77 ± 10.04 d | n/a | n/a | 0.16 ± 0.008 c | 1.40 ± 0.03 a |
| 8 | CRC 119 | 9.13 ± 0.09 g | 44.13 ± 0.03 e | 0.68 ± 0.02 ab | 0.18 ± 0.002 a | 0.61 ± 0.17 b | 0.17 ± 0.01 abc | 36.08 ± 0.32 cde | 36.86 ± 2.74 a | 92.17 ± 0.68 c | n/a | 0.14 ± 0.049 b | 2.99 ± 0.10 b |
| 9 | CRC 120 | 4.53 ± 0.03 b | 11.7 ± 0.12 a | 1.83 ± 0.22 e | 0.15 ± 0.004 a | 0.31 ± 0.10 ab | 0.05 ± 0.002 a | 34.72 ± 0.14 cd | 38.03 ± 1.41 ab | 92.05 ± 1.49 c | n/a | 0.14 ± 0.006 b | 3.39 ± 0.11 b |
| 10 | CRC 121 | 7.53 ± 0.09 e | 62.13 ± 0.09 h | 0.50 ± 0.02 a | 0.17 ± 0.01 a | 0.14 ± 0.08 ab | 0.14 ± 0.001 abc | 20.89 ± 1.25 b | 38.19 ± 0.46 ab | 53.07 ± 16.76 b | 0.58 ± 0.58 a | 0.3 2± 0.001 e | 5.92 ± 0.95 d |
| 11 | CRC 141 | 4.53 ± 0.09 b | 43.20 ± 0.06 d | 2.45 ± 0.01 f | 0.14 ± 0.002 a | 2.03 ± 0.49 c | 0.22 ± 0.02 bc | 80.44 ± 1.13 g | 38.94 ± 1.39 ab | 87.95 ± 0.88 c | 10.63 ± 1.08 a | 0.28 ± 0.001 d | 5.94 ± 0.38 d |
| Isolate (I) | * | * | * | * | * | * | |||||||
| solvent type (S) | * | * | ns | * | * | * | |||||||
| I*S | * | * | * | * | * | * | |||||||
| No. | Isolate | Inhibition of Fusarium sp. (CRC 161) | Inhibition of Colletotrichum sp. (CRC 149) | Inhibition of Pestalotiopsis sp. (CRC 151) | Inhibition of Alternaria sp. (CRC 152) | Inhibition of Curvularia sp. (CRC 162) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | EtOAc | MeOH | ||
| 1 | CRC 98 | - | - | - | - | 18.33 ± 0.09 g | 14.73 ± 0.33 d | - | - | - | 7.20 ± 0.05 b |
| 2 | CRC 101 | - | - | - | - | 11.03 ± 0.03 d | - | - | - | - | - |
| 3 | CRC 102 | - | - | - | - | 14.20 ± 0.058 e | 24.73 ± 0.33 f | - | - | - | - |
| 4 | CRC 108 | 16.5 ± 0.05 d | - | 13.60 ± 0.06 b | - | 28.93 ± 0.12 g | 16.33 ± 0.33 e | 26.13 ± 0.03 f | 19.50 ± 0.10 c | - | 13.90 ± 0.11 d |
| 5 | CRC 110 | - | - | - | - | - | 9.20 ± 0.05 a | 9.90 ± 0.05 c | - | - | 12.06 ± 0.88 c |
| 6 | CRC 114 | - | - | - | - | 17.60 ± 0.10 f | - | 9.20 ± 0.057 b | - | - | - |
| 7 | CRC 117 | 6.73 ± 0.03 b | - | - | - | 7.93 ± 0.08 b | - | - | - | 9.73 ± 0.08 b | - |
| 8 | CRC 119 | - | - | - | - | 8.80 ± 0.17 c | - | - | - | - | - |
| 9 | CRC 120 | 14.63 ± 0.03 c | - | 17.37 ± 0.07 c | 11.80 ± 0.58 b | - | 11.93 ± 0.33 b | 26.93 ± 0.05 g | - | - | - |
| 10 | CRC 121 | - | - | - | - | - | - | 13.3 ± 0.06 d | - | - | 20.60 ± 0.57 e |
| 11 | CRC 141 | - | - | - | - | - | 14.43 ± 0.33 c | 22.26 ± 0.06 e | 9.90 ± 0.10 b | - | - |
| Isolate (I) | * | * | * | * | * | ||||||
| solvent type (S) | * | * | * | * | * | ||||||
| I*S | * | * | * | * | * | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haituk, S.; Withee, P.; Sangta, J.; Senwanna, C.; Khamsaw, P.; Karunarathna, A.; Hongsibsong, S.; Sringarm, K.; Prasad, S.K.; Sommano, S.R.; et al. Production of Non-Volatile Metabolites from Sooty Molds and Their Bio-Functionalities. Processes 2022, 10, 329. https://doi.org/10.3390/pr10020329
Haituk S, Withee P, Sangta J, Senwanna C, Khamsaw P, Karunarathna A, Hongsibsong S, Sringarm K, Prasad SK, Sommano SR, et al. Production of Non-Volatile Metabolites from Sooty Molds and Their Bio-Functionalities. Processes. 2022; 10(2):329. https://doi.org/10.3390/pr10020329
Chicago/Turabian StyleHaituk, Sukanya, Patchareeya Withee, Jiraporn Sangta, Chanokned Senwanna, Pattarapol Khamsaw, Anuruddha Karunarathna, Surat Hongsibsong, Korawan Sringarm, Shashanka K. Prasad, Sarana Rose Sommano, and et al. 2022. "Production of Non-Volatile Metabolites from Sooty Molds and Their Bio-Functionalities" Processes 10, no. 2: 329. https://doi.org/10.3390/pr10020329