
Defense Mechanisms Induced by Celery Seed Essential Oil against Powdery Mildew Incited by Podosphaera fusca in Cucumber
 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Celery Seeds and Essential Oil Extraction
2.2. Chemical Analysis of Essential Oil from Celery Seeds (CSEO)
2.3. Effectiveness of CSEO in Controlling Cucumber Powdery Mildew
2.4. Effects of Treatment with CSEO on the Physiology of Cucumber Seedlings and Induction of Defense Mechanisms against Powdery Mildew
2.4.1. Experimental Design
2.4.2. Determination of Chlorophyll, Phenolic, and Flavonoid Compounds in Cucumber Leaves
2.4.3. Effect of CSEO on Activity of Defense Enzymes in Cucumber Leaves
2.4.4. Gene Expression of β-1,3-Glucanase, Chitinase, and Phenylalanine Ammonia-Lyase in Cucumber Leaves
RNA Extraction
cDNA Synthesis
Quantitative Real-Time PCR Assay
2.5. Statistical Analysis
3. Results
3.1. Chemical Composition of CSEO
3.2. Efficacy of CSEO in Controlling Cucumber Powdery Mildew
3.3. Effects of the Treatment with CSEO on the Physiology of Cucumber Seedlings and Its Ability to Induce Defense Mechanisms against Powdery Mildew
3.3.1. Chlorophyll Content in Cucumber Leaves
3.3.2. Content of Phenolic Compounds in Cucumber Leaves
3.3.3. Content of Flavonoid Compounds in Cucumber Leaves
3.3.4. Effect of CSEO on the Activity of Defense Enzymes against Cucumber Powdery Mildew
β-1,3-Glucanase
Chitinase
Phenylalanine Ammonia-Lyase
Peroxidase
Polyphenol Oxidase
3.3.5. Gene Expression in Cucumber Leaves
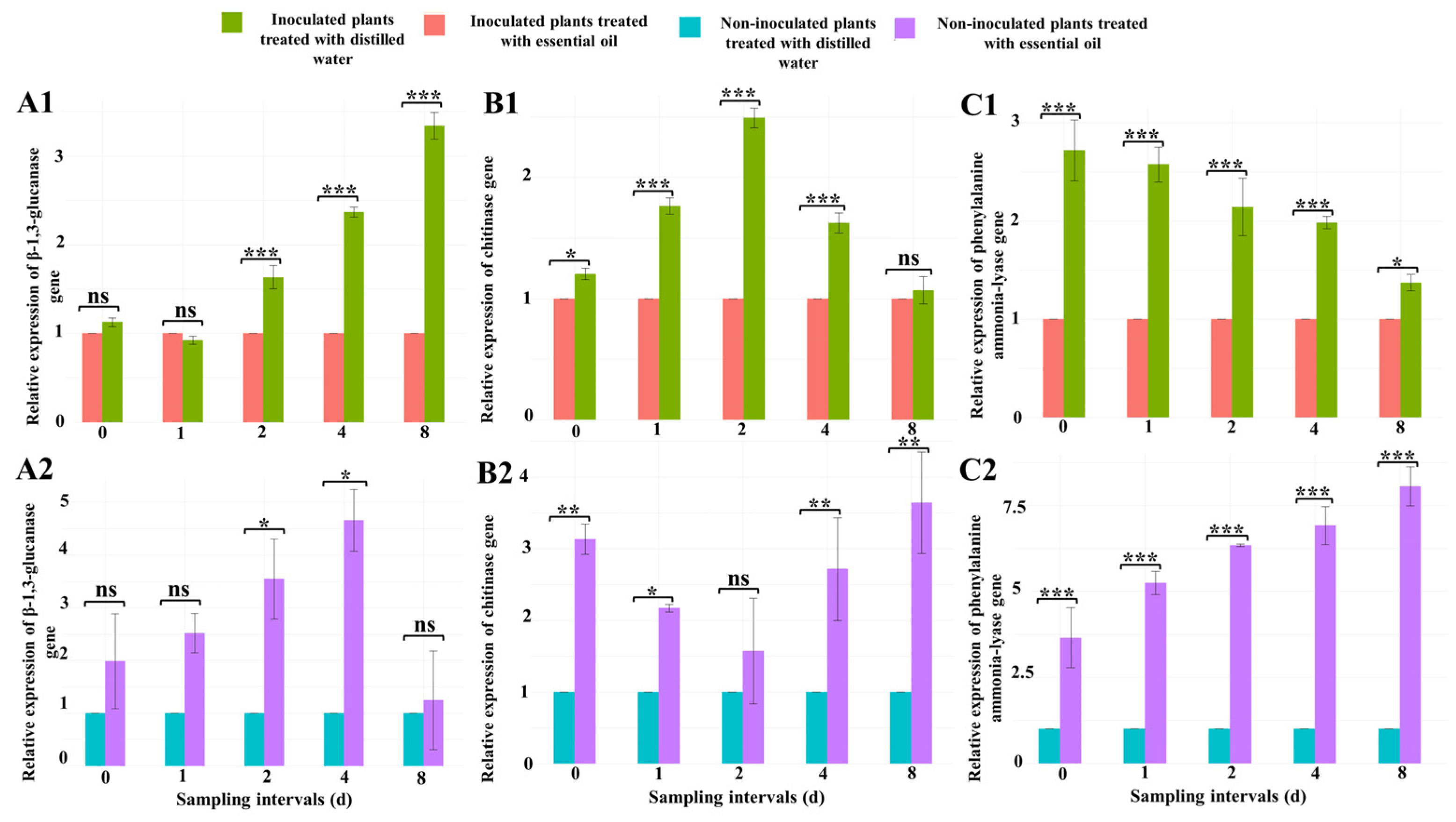
β-1,3-Glucanase Gene
Chitinase Gene
Phenylalanine Ammonia-Lyase Gene
3.4. Correlation between the β-1,3-Glucanase, Chitinase, and Phenylalanine Ammonia-Lyase Enzyme Activities and the Expression of the Corresponding Genes
3.5. Heatmap of Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mostafa, Y.S.; Hashem, M.; Alshehri, A.M.; Alamri, S.; Eid, E.M.; Ziedan, E.S.H.; Alrumman, S.A. Effective management of cucumber powdery mildew with essential oils. Agriculture 2021, 11, 1177. [Google Scholar] [CrossRef]
- Pérez-García, A.; Romero, D.; Fernández-Ortuño, D.; López-Ruiz, F.; De Vicente, A.; Torés, J.A. The powdery mildew fungus Podosphaera fusca (synonym Podosphaera xanthii), a constant threat to cucurbits. Mol. Plant Pathol. 2009, 10, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, E.A.; Abd-Elsyed, M.H.; Ebrahiem, A.M. Biological control of cucumber powdery mildew (Podosphaera xanthii) (Castagne) under greenhouse conditions. Egypt. J. Biol. Pest Control 2020, 30, 65. [Google Scholar] [CrossRef]
- Aqleem, A. A report of powdery mildews on cucumbers in village nomal, Gilgit Baltistan (GB)–Pakistan. Clin. Biotechnol. Microbiol. 2017, 1, 99–104. [Google Scholar]
- Wahul, S.M.; Jagtap, G.P.; Rewale, K.A.; Bhosale, R.P. Survey on powdery mildew of cucumber in Aurangabad and Jalna districts, India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1618–1624. [Google Scholar] [CrossRef]
- Abd-Elsayed, M.H.F.; Sarhan, E.A.D.; Ebrahiem, A.M.Y. Management of cucumber powdery mildew disease caused by Podosphaera xanthii by some fungicides and resistance inducing chemicals. Curr. Sci. Int. 2019, 8, 820–828. [Google Scholar] [CrossRef]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Biorgan. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef]
- Hüter, O.F. Use of natural products in the crop protection industry. Phytochem. Rev. 2011, 10, 185–194. [Google Scholar] [CrossRef]
- Loiseleur, O. Natural products in the discovery of agrochemicals. Chimia 2017, 71, 810. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal activity of bioactive metabolites produced by Trichoderma asperellum and Trichoderma atroviride in liquid medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, N.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 19, 1002–1017. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Kurt, Ş.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Sturchio, E.; Donnarumma, L.; Annesi, T.; Milano, F.; Casorri, L.; Masciarelli, E.; Zanellato, M.; Meconi, C.; Boccia, P. Essential oils: An alternative approach to management of powdery mildew diseases. Phytopathol. Mediterr. 2014, 53, 385–395. [Google Scholar] [CrossRef]
- Cattò, C.; de Vincenti, L.; Borgonovo, G.; Bassoli, A.; Marai, S.; Villa, F.; Cappitelli, F.; Saracchi, M. Sub-lethal concentrations of Perilla frutescens essential oils affect phytopathogenic fungal biofilms. J. Environ. Manag. 2019, 245, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Parikh, L.; Agindotan, B.O.; Burrows, M.E. Antifungal activity of plant-derived essential oils on pathogens of pulse crops. Plant Dis. 2021, 105, 1692–1701. [Google Scholar] [CrossRef]
- Kalleli, F.; Ghassen, A.B.I.D.; Salem, I.B.; Boughalleb-m’hamdi, N.; M’hamdi, M. Essential oil from fennel seeds (Foeniculum vulgare) reduces Fusarium wilt of tomato (Solanum lycopersicon). Phytopathol. Mediterr. 2020, 59, 63–76. [Google Scholar]
- Yuan, S.; Li, W.; Li, Q.; Wang, L.; Cao, J.; Jiang, W. Defense responses, induced by p-coumaric acid and methyl p-coumarate, of jujube (Ziziphus jujuba Mill.) fruit against black spot rot caused by Alternaria alternata. J. Agric. Food Chem. 2019, 67, 2801–2810. [Google Scholar] [CrossRef]
- La Spada, F.; Stracquadanio, C.; Riolo, M.; Pane, A.; Cacciola, S.O. Trichoderma counteracts the challenge of Phytophthora nicotianae infections on tomato by modulating plant defense mechanisms and the expression of crinkler, necrosis-inducing Phytophthora protein 1, and cellulose-binding elicitor lectin pathogenic effectors. Front. Plant Sci. 2020, 11, 583539. [Google Scholar] [CrossRef]
- Khalil, A.; Nawaz, H.; Ghania, J.B.; Rehman, R.; Nadeem, F. Value added products, chemical constituents and medicinal uses of celery (Apium graveolens L.)—A review. Int. J. Chem. Biochem. Sci. 2015, 8, 40–48. [Google Scholar]
- Ojala, T.; Remes, S.; Haansuu, P.; Vuorela, H.; Hiltunen, R.; Haahtela, K.; Vuorela, P. Antimicrobial activity of some coumarin containing herbal plants growing in Finland. J. Ethnopharmacol. 2000, 73, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Sirous, A.; Jamali Zavareh, A. An investigation into the effectiveness of extracts of three plants for the control of cucumber powdery mildew. Plant Prot. 2014, 37, 47–56. [Google Scholar]
- Khamesipour, F.; Razavi, S.M.; Hejazi, S.H.; Ghanadian, S.M. In vitro and in vivo anti-toxoplasma activity of Dracocephalum kotschyi essential oil. Food Sci. Nutr. 2021, 9, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Cook, T.A.; Inman, A.J.; Shin, H.D. The taxonomy of powdery mildew fungi. In The Powdery Mildews. A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; APS Press: Saint Paul, MN, USA, 2002; pp. 13–55. [Google Scholar]
- Rodriguez-Tudela, J.L.; Arendrup, M.C.; Arikan, S.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.W.; Donnelly, J.P.; et al. EUCAST definitive document E. DEF 9.1: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. Clin. Microbiol. Infect. 2008, 9, 1–13. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Maizura, M.; Aminah, A.; Wan Aida, W.M. Total phenolic content and antioxidant activity of kesum (Polygonum minus), ginger (Zingiber officinale) and turmeric (Curcuma longa) extract. Int. Food Res. J. 2011, 18, 526–531. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Yedidia, I.; Benhamou, N.; Kapulnik, Y.; Chet, I. Induction and accumulation of PR proteins activity during early stages of root colonization by the mycoparasite Trichoderma harzianum strain T-203. Plant Physiol. Biochem. 2000, 38, 863–873. [Google Scholar] [CrossRef]
- Khan, M.F.; Umar, U.U.D. Application of a robust microplate assay to determine induced β-1, 3-glucanase and chitinase activity in the cotton plant. Biotechniques 2021, 70, 202–208. [Google Scholar] [CrossRef]
- Goldson, A.; Lam, M.; Scaman, C.H.; Clemens, S.; Kermode, A. Screening of phenylalanine ammonia lyase in plant tissues, and retention of activity during dehydration. J. Sci. Food Agric. 2008, 88, 619–625. [Google Scholar] [CrossRef]
- Monsur, M.B.; Ivy, N.A.; Haque, M.M.; Hasanuzzaman, M.; El Sabagh, A.; Rohman, M.M. Oxidative stress tolerance mechanism in rice under salinity. Phyton 2020, 89, 497–517. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed]
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank (accessed on 14 September 2023).
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 135, 365–386. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lucero, M.E.; Fredrickson, E.L.; Estell, R.E.; Morrison, A.A.; Richman, D.B. Volatile composition of Gutierrezia sarothrae (broom snakeweed) as determined by steam distillation and solid phase microextraction. J. Essent. Oil Res. 2006, 18, 121–125. [Google Scholar] [CrossRef]
- Adams, R.P.; Elizondo, M.S.G.; Elizondo, M.G.; Slinkman, E. DNA fingerprinting and terpenoid analysis of Juniperus blancoi var. huehuentensis (Cupressaceae), a new subalpine variety from Durango, Mexico. Biochem. Syst. Ecol. 2006, 34, 205–211. [Google Scholar] [CrossRef]
- Fan, G.; Lu, W.; Yao, X.; Zhang, Y.; Wang, K.; Pan, S. Effect of fermentation on free and bound volatile compounds of orange juice. Flavour Fragr. J. 2009, 24, 219–225. [Google Scholar] [CrossRef]
- Velickovic, D.T.; Randjelovic, N.V.; Ristic, M.S.; Smelcerovic, A.A.; Velickovic, A.S. Chemical composition and antimicrobial action of the ethanol extracts of Salvia pratensis L., Salvia glutinosa L. and Salvia aethiopis L. J. Serb. Chem. Soc. 2002, 67, 639–646. [Google Scholar] [CrossRef]
- Andriamaharavo, N.R. Retention Data. NIST Mass Spectrometry Data Center, 2014. National Institute of Standards and Technology. US Department of Commerce. NIST Chemistry Webook, SRD 69. Available online: https://webbook.nist.gov/cgi/cbook.cgi?Author=Andriamaharavo%2C+N.R.&Units=SI&Mask=2000 (accessed on 14 September 2023).
- Miyazawa, M.; Kawata, J. Identification of the key aroma compounds in dried roots of Rubia cordifolia. J. Oleo Sci. 2006, 55, 37–39. [Google Scholar] [CrossRef]
- Saroglou, V.; Dorizas, N.; Kypriotakis, Z.; Skaltsa, H.D. Analysis of the essential oil composition of eight Anthemis species from Greece. J. Chromatogr. A 2006, 1104, 313–322. [Google Scholar] [CrossRef]
- Pitarokili, D.; Couladis, M.; Petsikos-Panayotarou, N.; Tzakou, O. Composition and antifungal activity on soil-borne pathogens of the essential oil of Salvia sclarea from Greece. J. Agric. Food Chem. 2002, 50, 6688–6691. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.; Tzakou, O.; Couladis, M. Essential oil composition of Turkish herbal tea (Salvia aucheri Bentham var. canescens Boiss. & Heldr.). Flavour Fragr. J. 2003, 18, 325–327. [Google Scholar] [CrossRef]
- Asuming, W.A.; Beauchamp, P.S.; Descalzo, J.T.; Dev, B.C.; Dev, V.; Frost, S.; Ma, C.W. Essential oil composition of four Lomatium Raf. species and their chemotaxonomy. Biochem. Syst. Ecol. 2005, 33, 17–26. [Google Scholar] [CrossRef]
- Elgamal, N.G.; Khalil, M.S. First report of powdery mildew caused by Podosphaera xanthii on Luffa cylindrica in Egypt and its control. J. Plant Prot. Res. 2020, 60, 311–319. [Google Scholar] [CrossRef]
- Sowbhagya, H.B. Chemistry, technology, and nutraceutical functions of celery (Apium graveolens L.): An overview. Crit. Rev. Food Sci. Nutr. 2014, 54, 389–398. [Google Scholar] [CrossRef]
- Dąbrowska, J.A.; Kunicka-Styczyńska, A.; Śmigielski, K.B. Biological, chemical, and aroma profiles of essential oil from waste celery seeds (Apium graveolens L.). J. Essent. Oil Res. 2020, 32, 308–315. [Google Scholar] [CrossRef]
- Nirmala, M.J.; Durai, L.; Gopakumar, V.; Nagarajan, R. Preparation of celery essential oil-based nanoemulsion by ultrasonication and evaluation of its potential anticancer and antibacterial activity. Int. J. Nanomed. 2020, 15, 7651–7666. [Google Scholar] [CrossRef]
- Han, X.B.; Zhao, J.; Cao, J.M.; Zhang, C.S. Essential oil of Chrysanthemum indicum L.: Potential biocontrol agent against plant pathogen Phytophthora nicotianae. Environ. Sci. Pollut. Res. 2019, 26, 7013–7023. [Google Scholar] [CrossRef]
- Zorga, J.; Kunicka-Styczyńska, A.; Gruska, R.; Śmigielski, K. Ultrasound-assisted hydrodistillation of essential oil from celery seeds (Apium graveolens L.) and its biological and aroma profiles. Molecules 2020, 25, 5322. [Google Scholar] [CrossRef]
- Kong, Q.; Zhou, L.; Wang, X.; Luo, S.; Li, J.; Xiao, H.; Zhang, X.; Feng, S.; Chen, T.; Yuan, M.; et al. Chemical composition and allelopathic effect of essential oil of Litsea pungens. Agronomy 2021, 11, 1115. [Google Scholar] [CrossRef]
- Moharam, M.H.A. Induction of defence-related biochemical changes in okra leaves to powdery mildew disease by several plant-derived agents. Arch. Phytopathol. Plant Prot. 2013, 46, 1667–1682. [Google Scholar] [CrossRef]
- Oubihi, A.; Ballaoui, F.Z.; Imtara, H.; Jaber, H.; Ettouil, A.; Haida, S.; Ouhssine, M.; Noman, O.M.; Mothana, R.A.; Tarayrah, M.; et al. Phytochemical compounds, acute toxicity, anti-Inflammatory and antioxidant activities of Thymus leptobotrys Murb essential oil. Molecules 2023, 28, 1355. [Google Scholar] [CrossRef] [PubMed]
- Singha, I.M.; Kakoty, Y.; Unni, B.G.; Kalita, M.C.; Das, J.; Naglot, A.; Wann, S.B.; Singh, L. Control of Fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici using leaf extract of Piper betle L.: A preliminary study. World J. Microbiol. Biotechnol. 2011, 27, 2583–2589. [Google Scholar] [CrossRef]
- Khaleil, M.M.; Alnoman, M.M.; Elrazik, E.S.A.; Zagloul, H.; Khalil, A.M.A. Essential oil of Foeniculum vulgare Mill. As a green fungicide and defense-inducing agent against fusarium root rot disease in Vicia faba L. Biology 2021, 10, 696. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Chen, Q.; Bu, Y.; Luo, R.; Hao, S.; Zhang, J.; Tian, J.; Yao, Y. Flavonoid accumulation plays an important role in the rust resistance of Malus plant leaves. Front. Recent Sci. 2017, 8, 1286. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.A.; Hafez, E.E.; Al-Kolaibe, A.M.; Abdel Razik, E.S.S.; Abd-Ellatif, S.; Ibrahim, A.A.; Kabeil, S.S.A.; Elshafie, H.S. Biochemical characterization, antifungal activity, and relative gene expression of two mentha essential oils controlling Fusarium oxysporum, the causal agent of Lycopersicon esculentum root rot. Plants 2022, 11, 189. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against gray mold and Fusarium wilt. Plant Physiol. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove essential oil as an alternative approach to control postharvest blue mold caused by Penicillium italicum in citrus fruit. Biomolecules 2019, 9, 197. [Google Scholar] [CrossRef]
- Kaur, S.; Samota, M.K.; Choudhary, M.; Choudhary, M.; Pandey, A.K.; Sharma, A.; Thakur, J. How do plants defend themselves against pathogens-Biochemical mechanisms and genetic interventions. Physiol. Mol. Biol. Plants 2022, 28, 485–504. [Google Scholar] [CrossRef]
- Saltos-Rezabala, L.A.; Silveira, P.R.D.; Tavares, D.G.; Moreira, S.I.; Magalhães, T.A.; Botelho, D.M.D.S.; Alves, E. Thyme essential oil reduces disease severity and induces resistance against Alternaria linariae in tomato plants. Horticulturae 2022, 8, 919. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, F.; Zhou, W.; Zhang, P.; Fan, Y.J. Application of osthol induces a resistance response against powdery mildew in pumpkin leaves. Int. J. Mol. Sci. 2007, 8, 1001–1012. [Google Scholar] [CrossRef]
- Amri, R.; Font i Forcada, C.; Giménez, R.; Pina, A.; Moreno, M.Á. Biochemical characterization and differential expression of PAL genes associated with “translocated” peach/plum graft-incompatibility. Front. Plant Sci. 2021, 12, 622578. [Google Scholar] [CrossRef] [PubMed]
- Bill, M.; Sivakumar, D.; Beukes, M.; Korsten, L. Expression of pathogenesis-related (PR) genes in avocados fumigated with thyme oil vapours and control of anthracnose. Food Biochem. 2016, 194, 938–943. [Google Scholar] [CrossRef]
- Han, C.; Shao, H.; Zhou, S.; Mei, Y.; Cheng, Z.; Huang, L.; Lv, G. Chemical composition and phytotoxicity of essential oil from invasive plant, Ambrosia artemisiifolia L. Ecotoxicol. Environ. Saf. 2021, 211, 111879. [Google Scholar] [CrossRef]
- Banani, H.; Olivieri, L.; Santoro, K.; Garibaldi, A.; Gullino, M.L.; Spadaro, D. Thyme and savory essential oil efficacy and induction of resistance against Botrytis cinerea through priming of defense responses in apple. Foods 2018, 7, 11. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Molecular Formula | CAS | RI a | RI b | Rt c (Min) | TIC d (%) | Identification Method | References |
|---|---|---|---|---|---|---|---|---|
| β-Pinen | C10H16 | 000127-91-3 | 989 | 981 | 4.27 | 1.72 | EI Mass, RI | [37] |
| β-Myrcene | C10H16 | 000123-35-3 | 996 | 991 | 4.58 | 1.07 | EI Mass, RI | [38] |
| d-Limonene | C10H16 | 000138-86-3 | 1037 | 1035 | 5.77 | 42.74 | EI Mass, RI | [39] |
| Pentylbenzene | C11H16 | 000538-68-1 | 1157 | 1158 | 10.03 | 2.38 | EI Mass, RI | [40] |
| Pentanophenone | C11H14O | 001009-14-9 | 1350 | 1359 | 18.44 | 1.48 | EI Mass, RI | [41] |
| β-Selinene | C15H24 | 017066-67-0 | 1476 | 1479 | 23.86 | 11.91 | EI Mass, RI | [42] |
| α-Selinene | C15H24 | 000473-13-2 | 1481 | 1485 | 24.1 | 1.7 | EI Mass, RI | [43] |
| Caryophyllene oxide | C15H24O | 001139-30-6 | 1568 | 1566 | 27.57 | 1.46 | EI Mass, RI | [44] |
| β-Selinenol | C15H26O | 000473-15-4 | 1638 | 1638 | 30.28 | 1.74 | EI Mass, RI | [45] |
| 3-Butylphthalide | C12H14O2 | 6066-49-5 | 1658 | 1658 | 31.08 | 14.42 | EI Mass, RI | [46] |
| Mandelic acid | C10H12O3 | 000774-40-3 | 1731 | - e | 33.72 | 10.92 | EI Mass | f |
| Aromatics | 29.20 | |||||||
| Monoterpenoids: Non-oxygenated | 45.53 | |||||||
| Monoterpenoids: Oxygenated | - g | |||||||
| HSesquiterpenoids | 16.81 | |||||||
| Minor compounds less than 1% | 8.46 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soleimani, H.; Mostowfizadeh-Ghalamfarsa, R.; Ghanadian, M.; Karami, A.; Cacciola, S.O. Defense Mechanisms Induced by Celery Seed Essential Oil against Powdery Mildew Incited by Podosphaera fusca in Cucumber. J. Fungi 2024, 10, 17. https://doi.org/10.3390/jof10010017
Soleimani H, Mostowfizadeh-Ghalamfarsa R, Ghanadian M, Karami A, Cacciola SO. Defense Mechanisms Induced by Celery Seed Essential Oil against Powdery Mildew Incited by Podosphaera fusca in Cucumber. Journal of Fungi. 2024; 10(1):17. https://doi.org/10.3390/jof10010017
Chicago/Turabian StyleSoleimani, Hajar, Reza Mostowfizadeh-Ghalamfarsa, Mustafa Ghanadian, Akbar Karami, and Santa Olga Cacciola. 2024. "Defense Mechanisms Induced by Celery Seed Essential Oil against Powdery Mildew Incited by Podosphaera fusca in Cucumber" Journal of Fungi 10, no. 1: 17. https://doi.org/10.3390/jof10010017