

Grapevine Trunk Diseases in Greece: Disease Incidence and Fungi Involved in Discrete Geographical Zones and Varieties
 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Surveys and Disease Incidence Assessment
2.2. Sampling and Fungal Isolation
2.3. Morphological and Cultural Characterization of Isolates
2.4. DNA Extraction, PCR Amplification and Sequencing
2.5. Identification and Characterization of Isolates
2.6. Correlation between Meteorological Data and GTDs Pathogens’ Frequency
2.7. Pathogenicity Tests
2.8. Statistical Analysis
3. Results
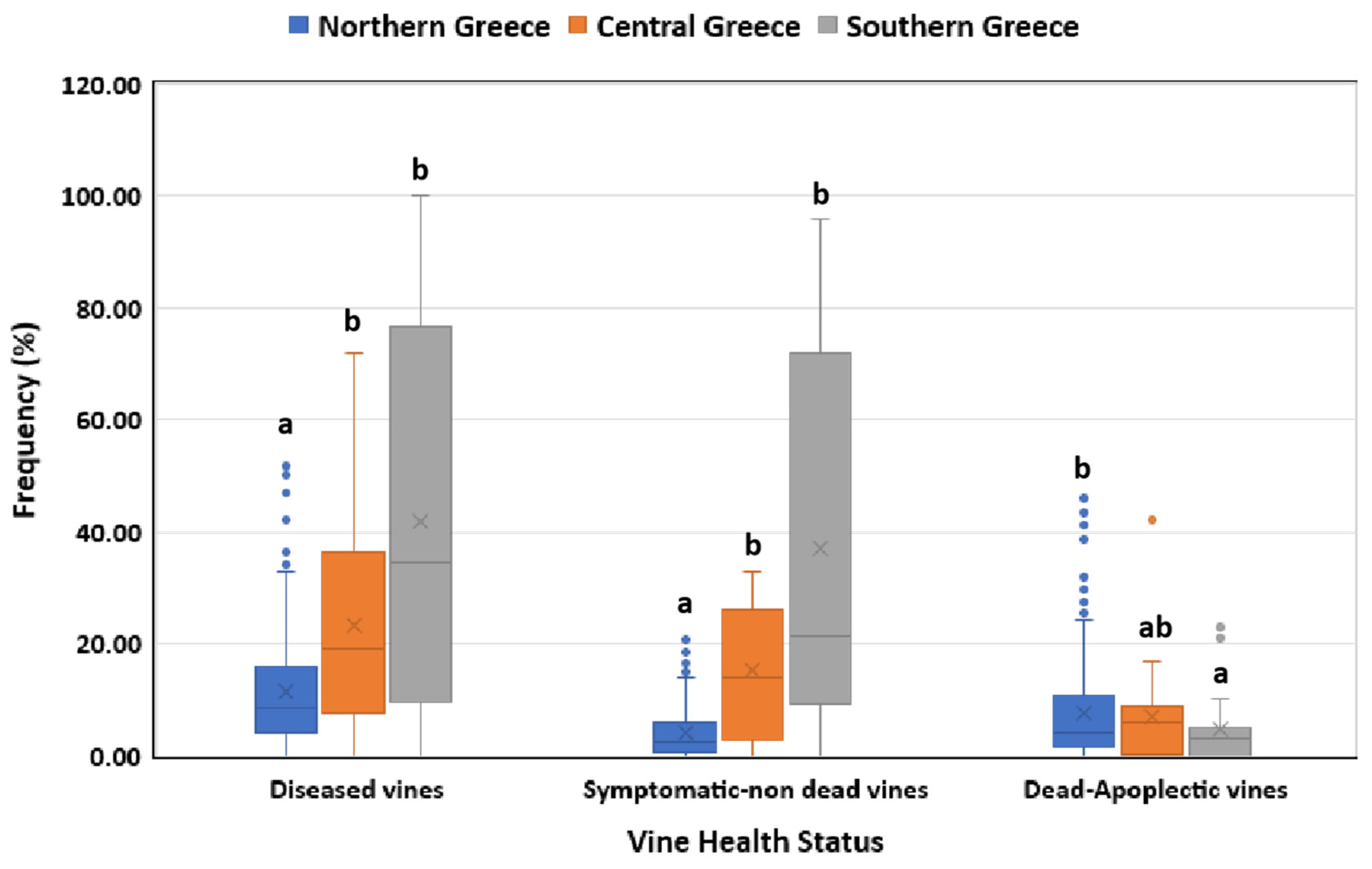
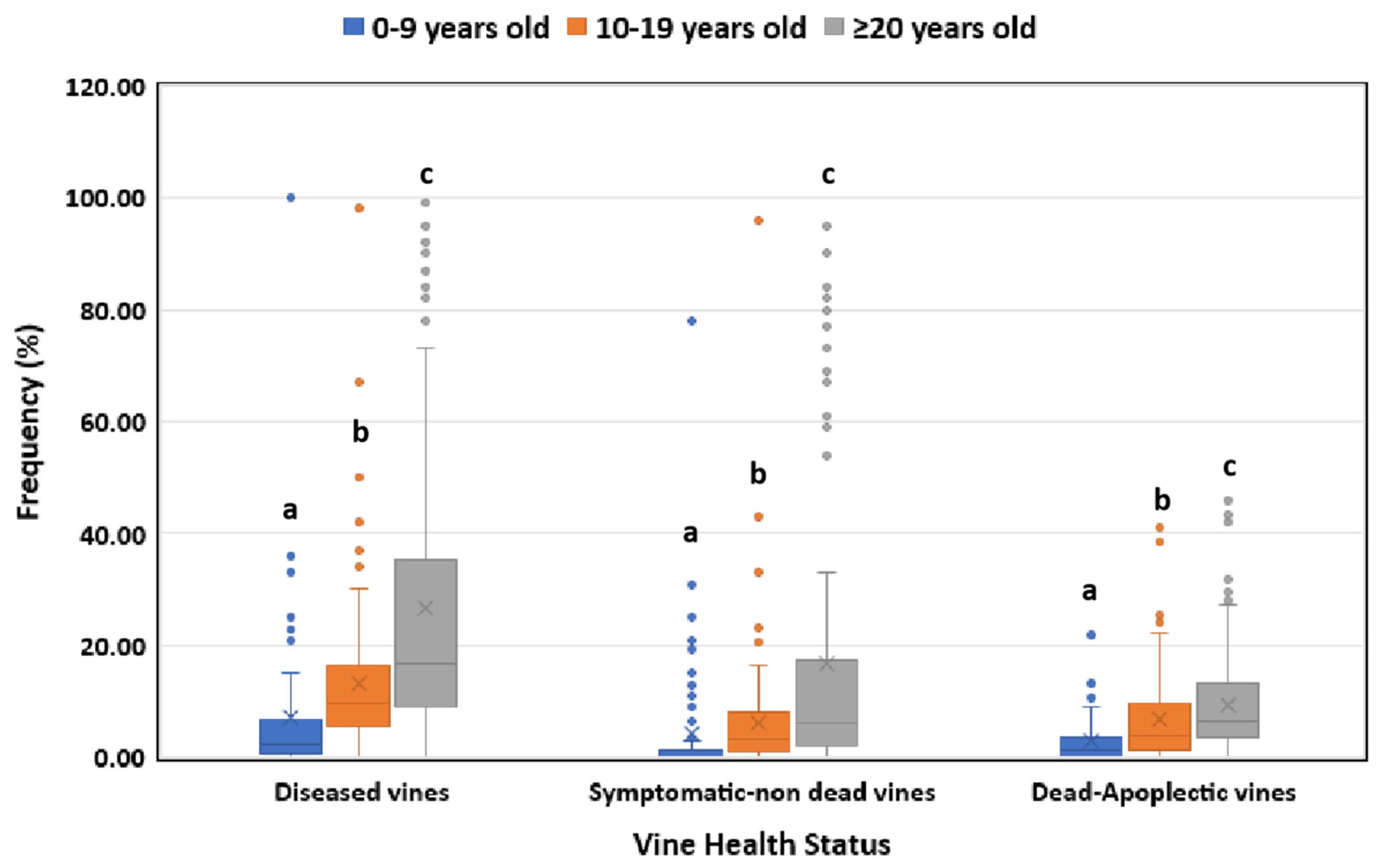
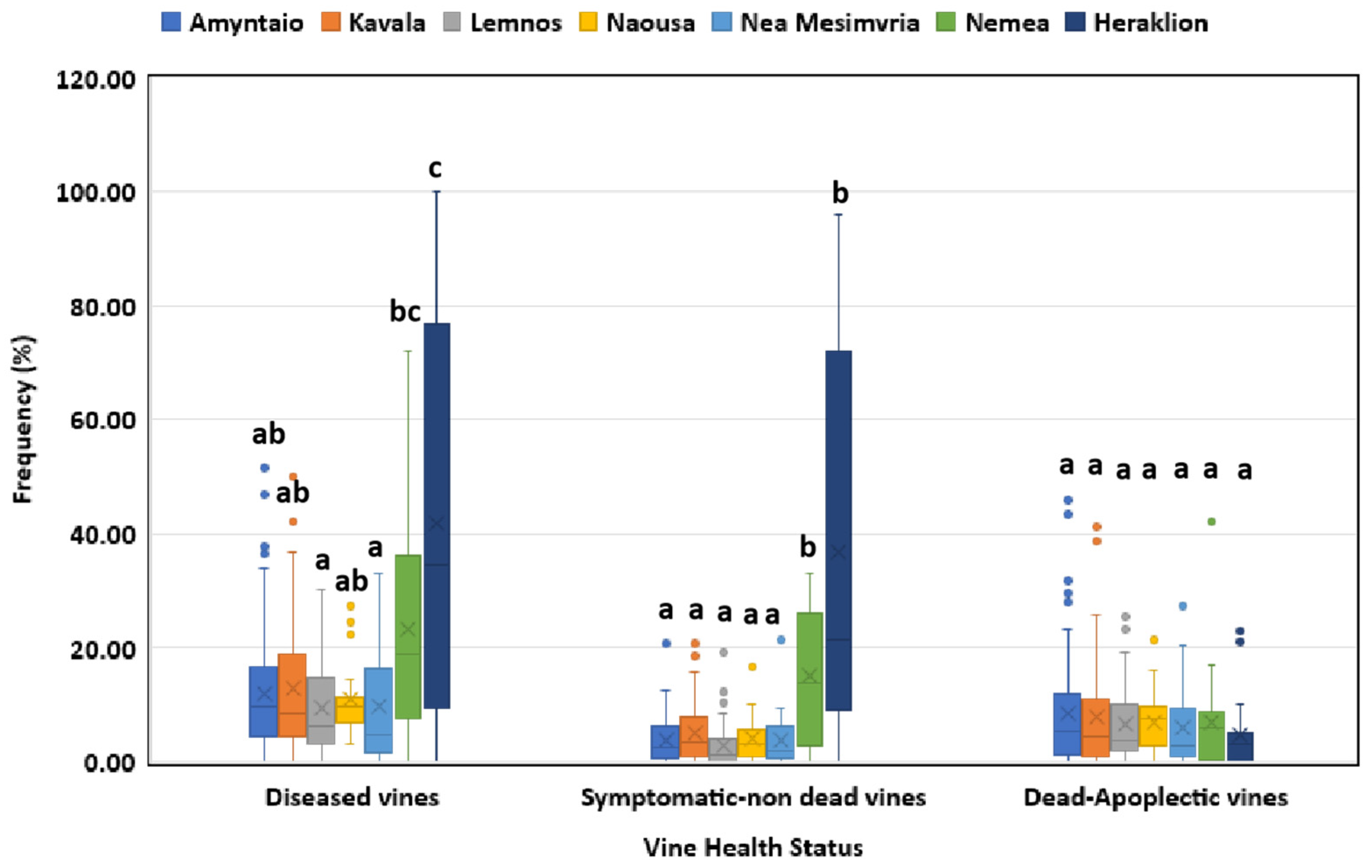
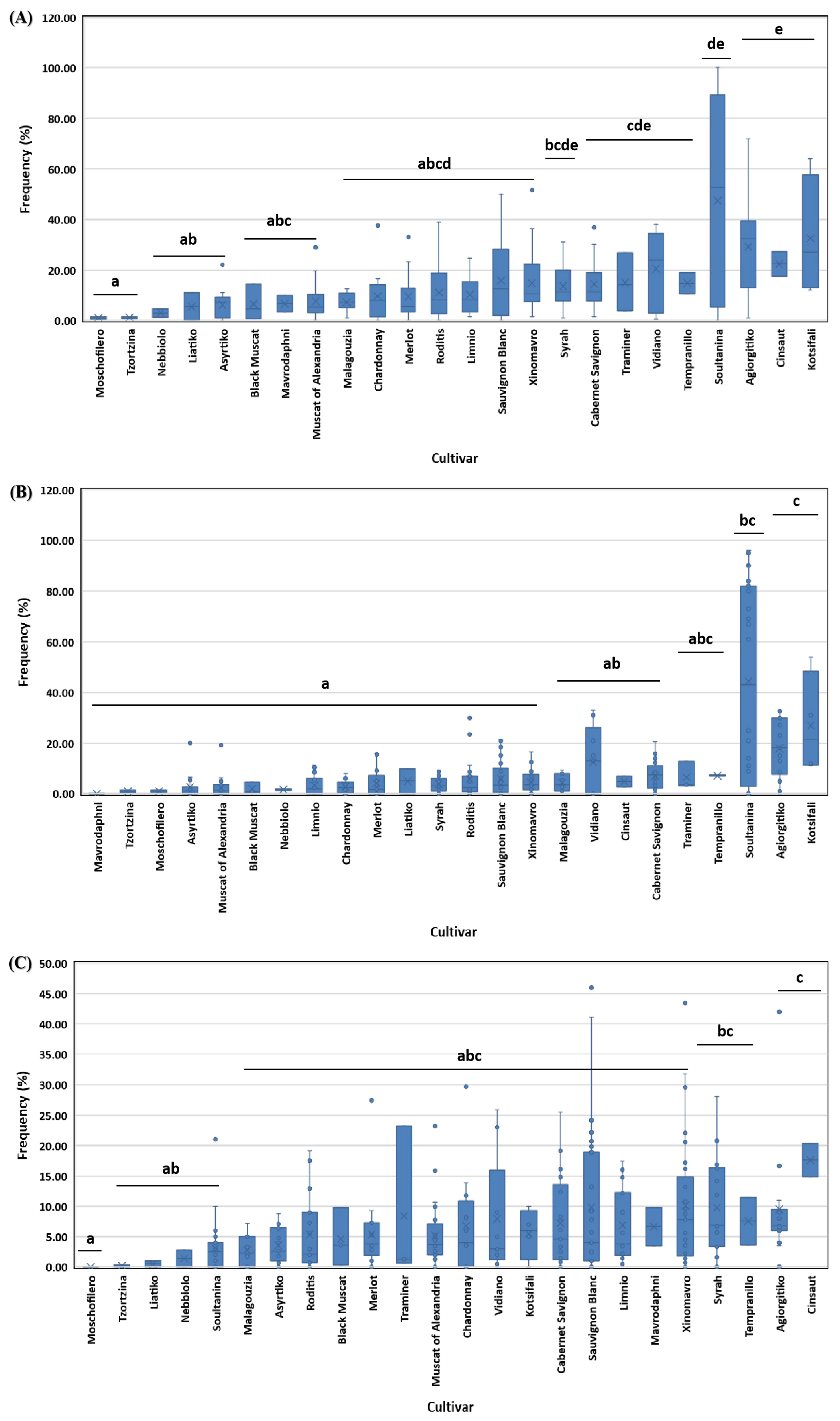
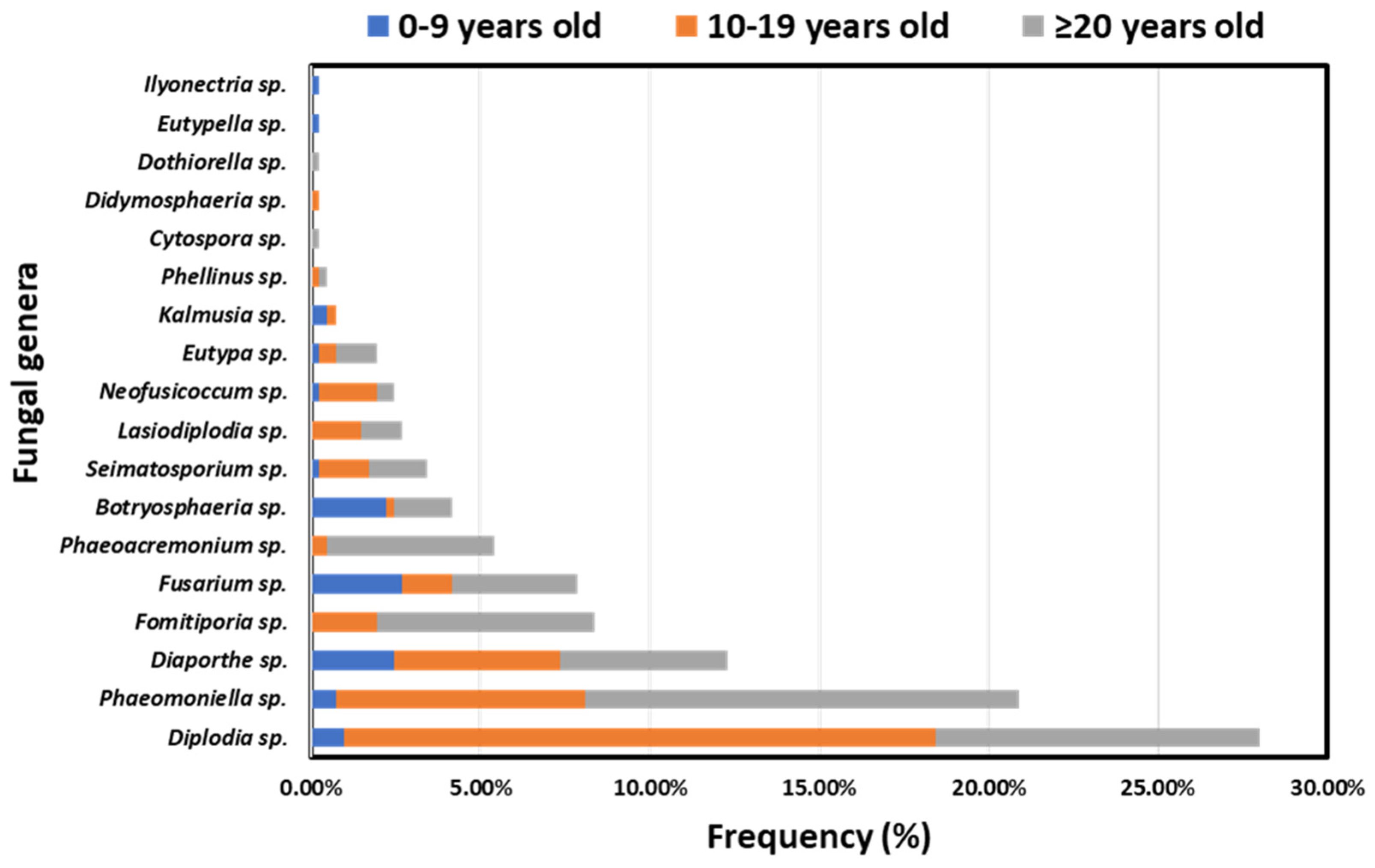
3.1. Disease Incidence in Respect to Geographical Region, Vine Age and Cultivar
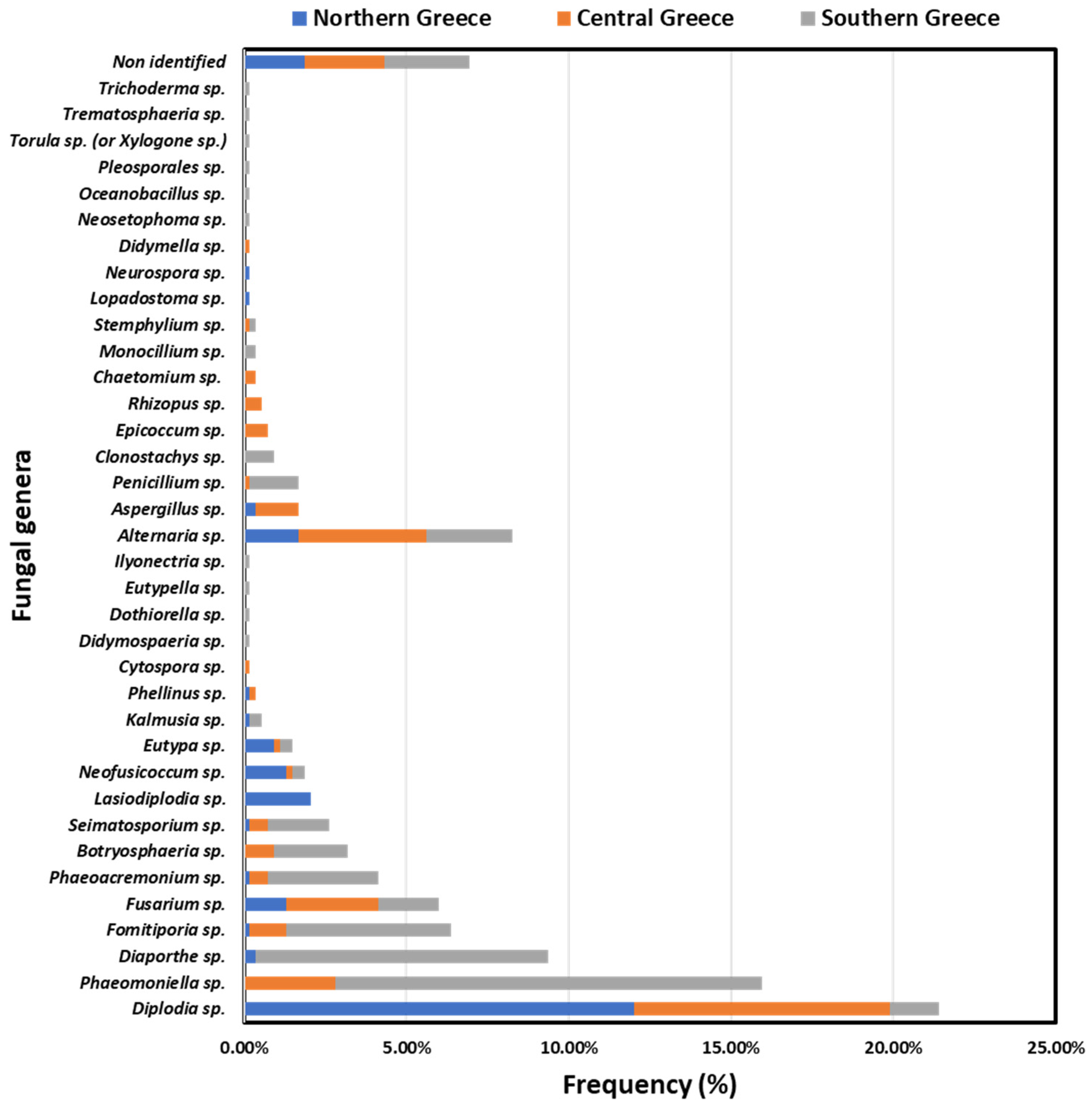
3.2. Fungal Genera Frequencies
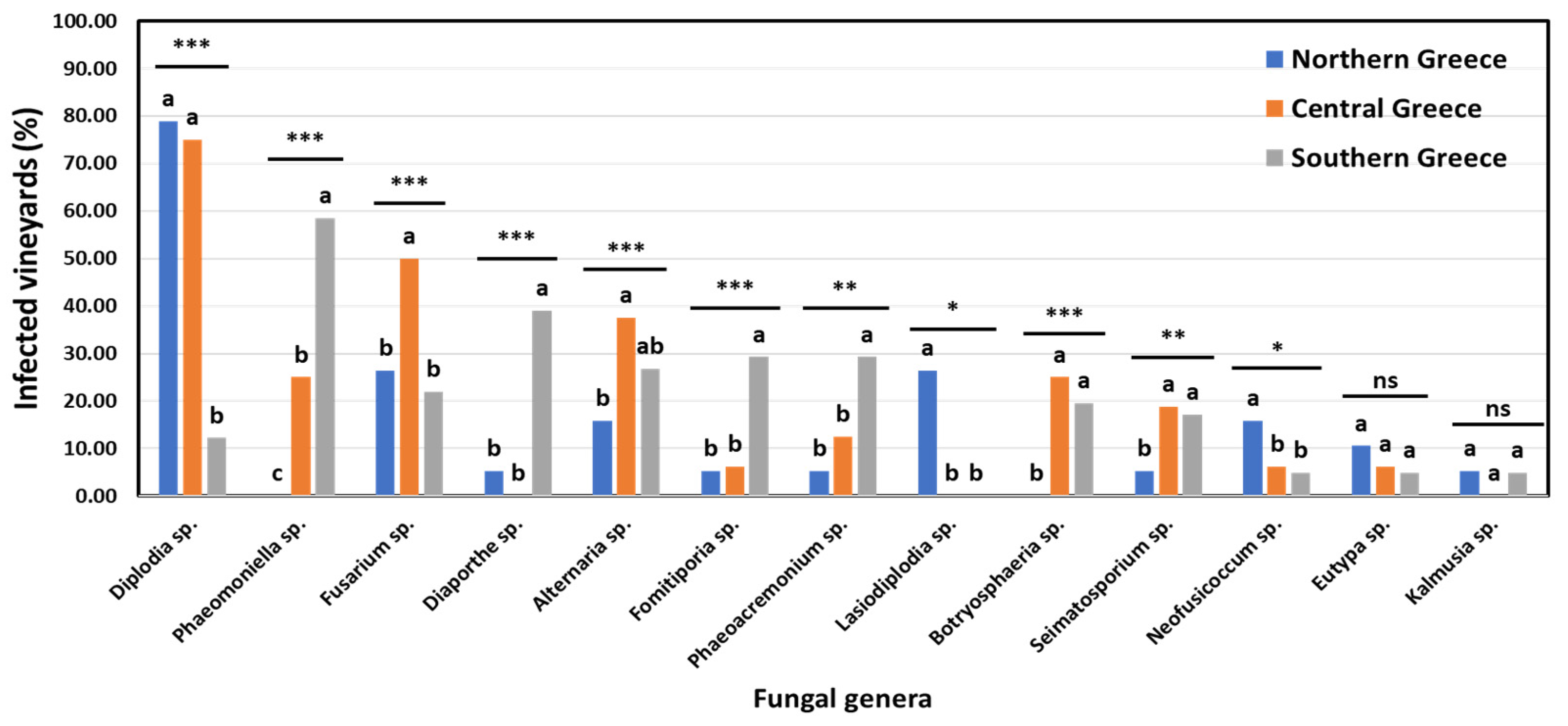
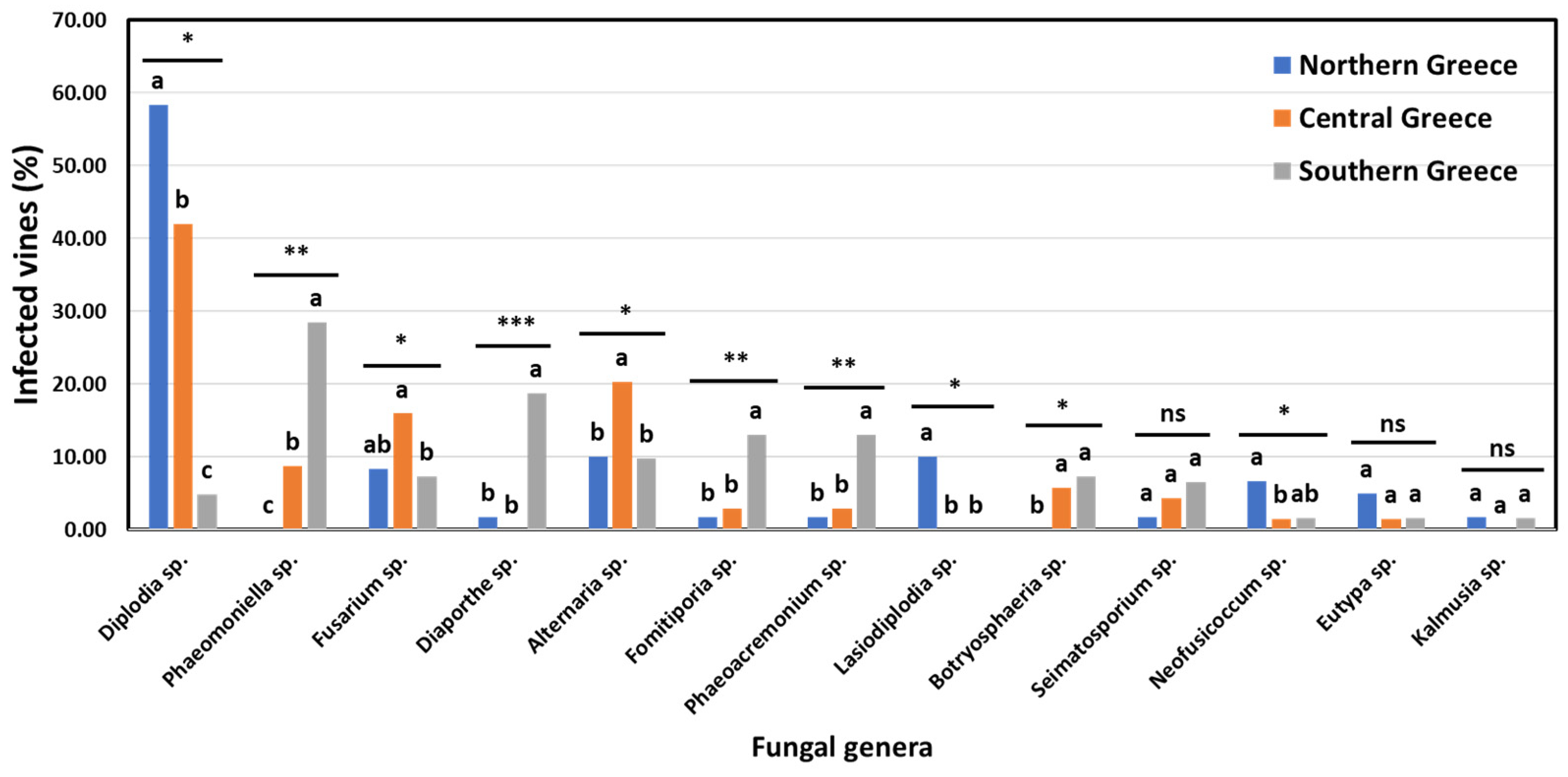
3.3. Incidence of GTD-Associated Fungal Genera in Vineyards and Vines in Discrete Viticultural Zones
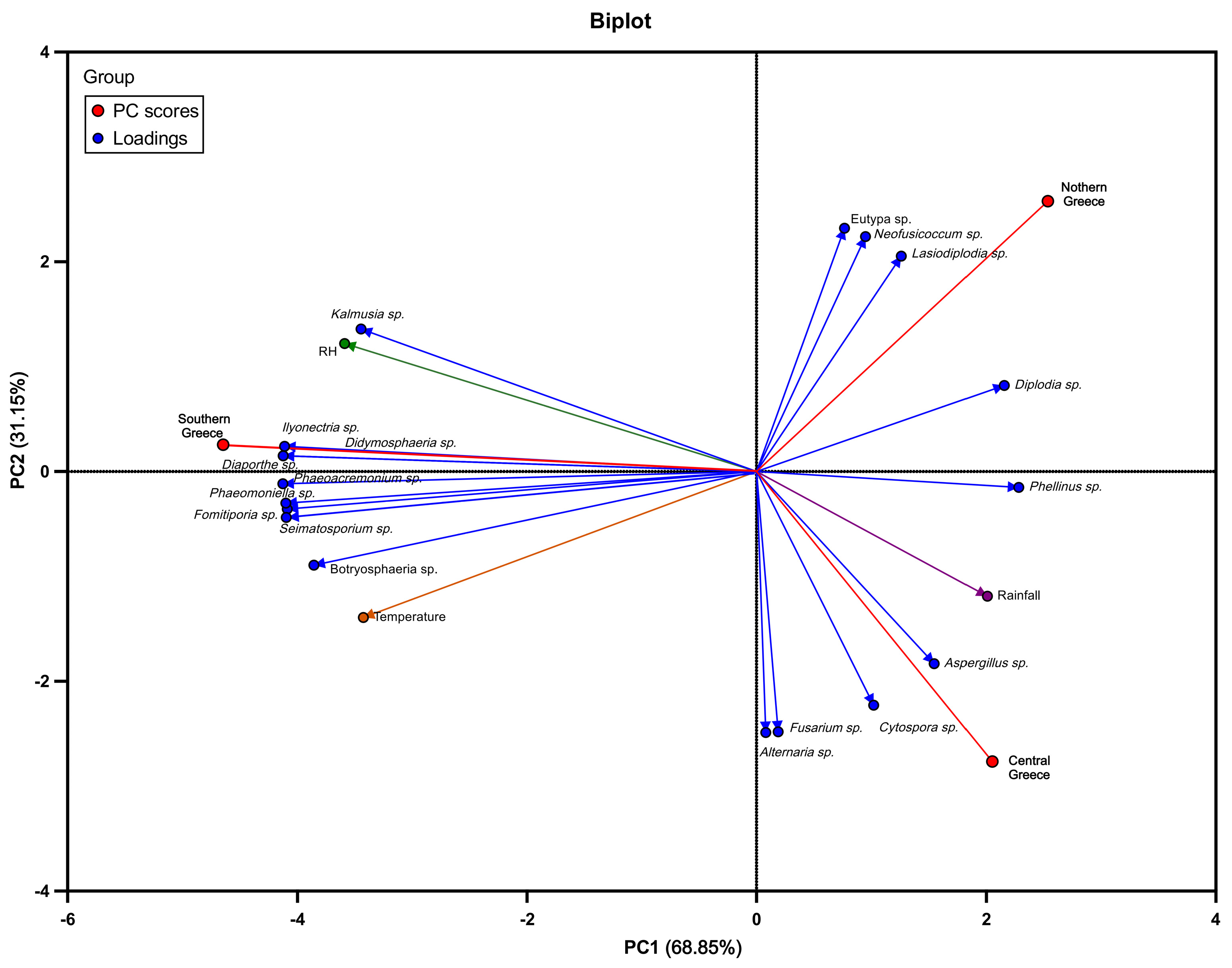
3.4. Correlation between Meteorological Data and GTDs Pathogens’ Frequency
3.5. Morphological, Physiological, and Molecular Identification of Lesser-Known Fungal Species
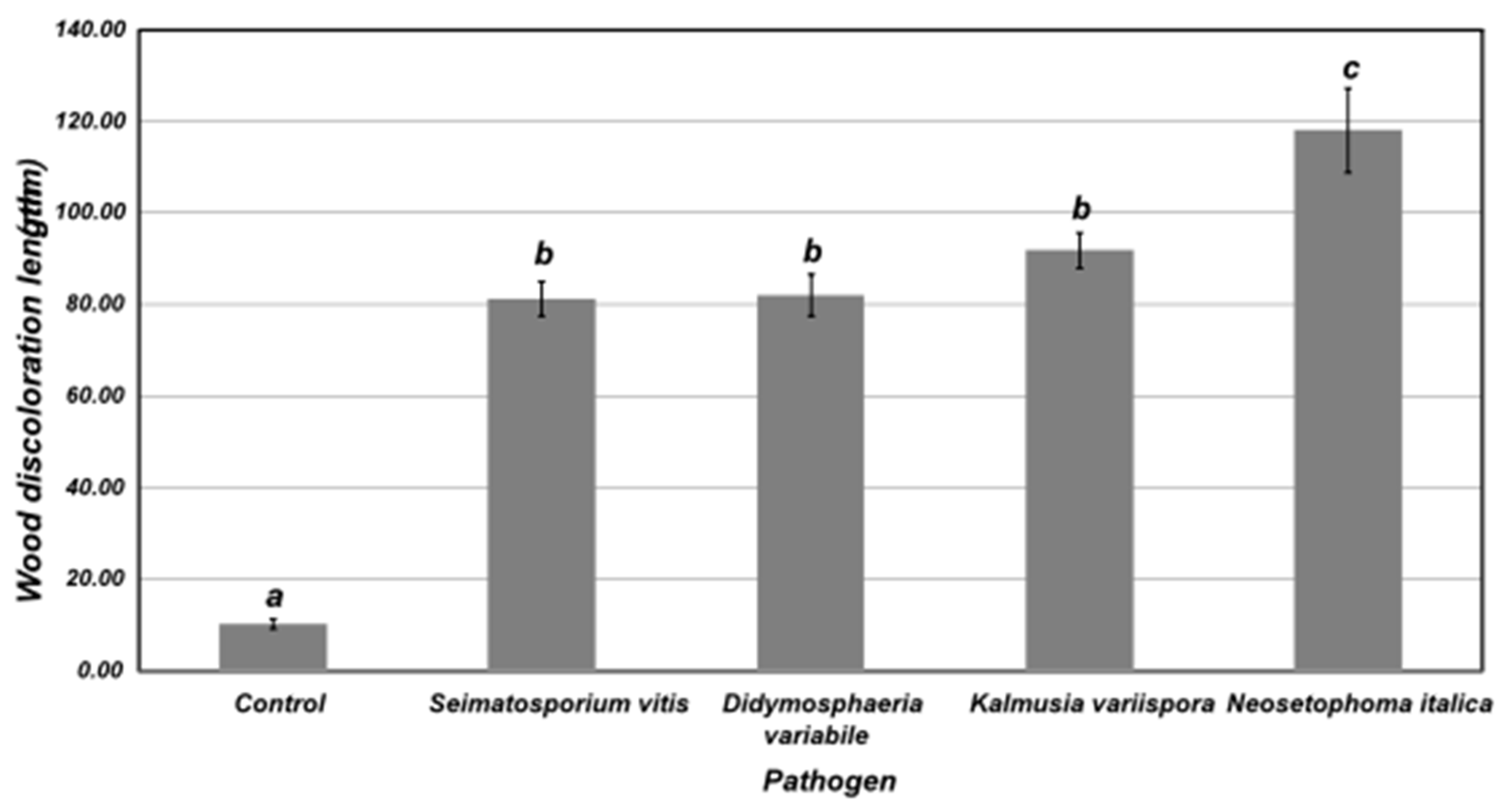
3.6. Pathogenicity of Fungal Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OIV. Distribution of the World’s Grapevine Varieties; International Organisation of Vine and Wine: Dijon, France, 2017. [Google Scholar]
- OIV. Country Statistics; International Organisation of Vine and Wine: Dijon, France, 2021. [Google Scholar]
- FAOSTAT. Statistics “Crops and Livestock Products”; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021; Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 September 2023).
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases: A Review; OIV Publications: Dijon, France, 2016. [Google Scholar]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine Trunk Diseases: Complex and Still Poorly Understood. Plant Pathol. 2012, 62, 243–265. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and Brown Wood-Streaking: Two Old and Elusive Diseases of Grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases with Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Kraus, C.; Markakis, E.; Alves, E.; Armengol, J.; Eichmeier, A.; Compant, S.; Gramaje, D. Fungal Trunk Diseases of Fruit Trees in Europe: Pathogens, Spread and Future Directions. Phytopathol. Mediterr. 2022, 61, 563–599. [Google Scholar] [CrossRef]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Urbez-Torres, J.R. Grapevine Trunk Disease Fungi: Their Roles as Latent Pathogens and Stress Factors That Favour Disease Development and Symptom Expression. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed]
- Songy, A.; Fernandez, O.; Clement, C.; Larignon, P.; Fontaine, F. Grapevine Trunk Diseases under Thermal and Water Stresses. Planta 2019, 249, 1655–1679. [Google Scholar] [CrossRef] [PubMed]
- Markakis, E.A.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Evaluation of Greek Grapevine Cultivars for Resistance to Phaeomoniella Chlamydospora. Eur. J. Plant Pathol. 2017, 149, 277–283. [Google Scholar] [CrossRef]
- Markakis, E.A.; Ligoxigakis, E.K.; Roussos, P.A.; Sergentani, C.; Kavroulakis, N.; Roditakis, E.; Koubouris, G. Differential Susceptibility Responses of Greek Olive Cultivars to Fomitiporia Mediterranea. Eur. J. Plant Pathol. 2018, 153, 1055–1066. [Google Scholar] [CrossRef]
- Bekris, F.; Vasileiadis, S.; Papadopoulou, E.; Samaras, A.; Testempasis, S.; Gkizi, D.; Tavlaki, G.; Tzima, A.; Paplomatas, E.; Markakis, E.; et al. Correction To: Grapevine Wood Microbiome Analysis Identifies Key Fungal Pathogens and Potential Interactions with the Bacterial Community Implicated in Grapevine Trunk Disease Appearance. Environ. Microbiome 2021, 17, 23. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gautier, A.; Laval, V.; Compant, S.; Maurer, W.; Sessitsch, A.; Lebrun, M.; Rey, P. Major Changes in Grapevine Wood Microbiota Are Associated with the Onset of Esca, a Devastating Trunk Disease. Environ. Microbiol. 2020, 22, 5189–5206. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Zhang, W.; Zhang, J.; Wang, H.; Peng, J.; Wang, X.; Yan, J. Belowground Microbiota Analysis Indicates That Fusarium Spp. Exacerbate Grapevine Trunk Disease. Environ. Microbiome 2023, 18, 29. [Google Scholar] [CrossRef] [PubMed]
- Martín, L.; García-García, B.; Alguacil, M.d.M. Interactions of the Fungal Community in the Complex Patho-System of Esca, a Grapevine Trunk Disease. Int. J. Mol. Sci. 2022, 23, 14726. [Google Scholar] [CrossRef] [PubMed]
- Patanita, M.; Albuquerque, A.; Campos, M.D.; Materatski, P.; Varanda, C.M.R.; Ribeiro, J.A.; Félix, M.d.R. Metagenomic Assessment Unravels Fungal Microbiota Associated to Grapevine Trunk Diseases. Horticulturae 2022, 8, 288. [Google Scholar] [CrossRef]
- Bustamante, M.I.; Elfar, K.; Smith, R.J.; Bettiga, L.J.; Tian, T.; Torres, G.A.; Eskalen, A. First Report of Fusarium Annulatum Associated with Young Vine Decline in California. Plant Dis. 2022, 106, 2752. [Google Scholar] [CrossRef]
- Kanetis, L.; Taliadoros, D.; Makris, G.; Christoforou, M. A Novel Seimatosporium and Other Sporocadaceae Species Associated with Grapevine Trunk Diseases in Cyprus. Plants 2022, 11, 2733. [Google Scholar] [CrossRef]
- Raimondo, M.L.; Carlucci, A.; Ciccarone, C.; Sadallah, A.; Lops, F. Identification and Pathogenicity of Lignicolous Fungi Associated with Grapevine Trunk Diseases in Southern Italy. Phytopathol. Mediterr. 2019, 58, 639–662. [Google Scholar]
- Rumbos, I.; Rumbou, A. Fungi Associated with Esca and Young Grapevine Decline in Greece. Phytopathol. Mediterr. 2001, 40, S330–S335. [Google Scholar] [CrossRef]
- Cary, J.W.; Ehrlich, K.C.; Beltz, S.B.; Harris-Coward, P.; Klich, M.A. Characterization of the Aspergillus Ochraceoroseus Aflatoxin/Sterigmatocystin Biosynthetic Gene Cluster. Mycologia 2009, 101, 352–362. [Google Scholar] [CrossRef]
- Bala, K.; Robideau, G.P.; Désaulniers, N.; de Cock, A.W.A.M.; Lévesque, C.A. Taxonomy, DNA Barcoding and Phylogeny of Three New Species of Pythium from Canada. Pers. Mol. Phylogeny Evol. Fungi 2010, 25, 22–31. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium Are Nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and Phylogeny of Gliocladium Analysed from Nuclear Large Subunit Ribosomal DNA Sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Fussler, L.; Kobes, N.; Bertrand, F.; Maumy, M.; Grosman, J.; Savary, S. A Characterization of Grapevine Trunk Diseases in France from Data Generated by the National Grapevine Wood Diseases Survey. Phytopathology 2008, 98, 571–579. [Google Scholar] [CrossRef]
- Markakis, E.A.; Kavroulakis, N.; Ntougias, S.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Characterization of Fungi Associated with Wood Decay of Tree Species and Grapevine in Greece. Plant Dis. 2017, 101, 1929–1940. [Google Scholar] [CrossRef]
- Hernandez, M.; KC, A. A Systematic Survey on Prevalence of Grapevine Trunk Disease Pathogens in Oregon Vineyards. Plant Dis. 2022, 107, 1355–1364. [Google Scholar] [CrossRef]
- White, C.-L.; Halleen, F.; Mostert, L. Symptoms and Fungi Associated with Esca in South African Vineyards. Phytopathol. Mediterr. 2011, 50, S236–S246. [Google Scholar]
- Moretti, S.; Pacetti, A.; Pierron, R.; Kassemeyer, H.-H.; Fischer, M.; Péros, J.-P.; Pérez-González, G.; Bieler, E.; Schilling, M.; Di Marco, S.; et al. Fomitiporia Mediterranea M. Fisch., the Historical Esca Agent: A Comprehensive Review on the Main Grapevine Wood Rot Agent in Europe. Phytopathol. Mediterr. 2021, 60, 351–379. [Google Scholar] [CrossRef]
- Urbez-Torres, J.R.; Peduto, F.; Striegler, R.K.; Urrea-Romero, K.E.; Rupe, J.C.; Cartwright, R.D.; Gubler, W.D. Characterization of Fungal Pathogens Associated with Grapevine Trunk Diseases in Arkansas and Missouri. Fungal Divers. 2011, 52, 169–189. [Google Scholar] [CrossRef]
- Larach, A.; Vega-Celedón, P.; Salgado, E.; Salinas, A.; Riquelme, N.; Castillo-Novales, D.; Sanhueza, P.; Seeger, M.; Besoain, X. Higher Virulence of Diplodia Seriata Isolates on Vines of Cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue. Plants 2023, 12, 2984. [Google Scholar] [CrossRef] [PubMed]
- Cobos, R.; Ibañez, A.; Diez-Galán, A.; Calvo-Peña, C.; Ghoreshizadeh, S.; Coque, J.J.R. The Grapevine Microbiome to the Rescue: Implications for the Biocontrol of Trunk Diseases. Plants 2022, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Del Frari, G.; Cabral, A.; Nascimento, T.; Boavida Ferreira, R.; Oliveira, H. Epicoccum Layuense a Potential Biological Control Agent of Esca-Associated Fungi in Grapevine. PLoS ONE 2019, 14, e0213273. [Google Scholar] [CrossRef] [PubMed]
- Michailides, T.J.; Peacock, W.; Christensen, P.; Morgan, D.P.; Felts, D. First Report of Aspergillus Vine Canker of Table Grapes Caused by Aspergillus Niger. Plant Dis. 2002, 86, 75. [Google Scholar] [CrossRef] [PubMed]
- Silva-Valderrama, I.; Toapanta, D.; de los Angeles Miccono, M.; Lolas, M.; Díaz, G.A.; Cantu, D.; Castro, A. Biocontrol Potential of Grapevine Endophytic and Rhizospheric Fungi against Trunk Pathogens. Front. Microbiol. 2021, 11, 614620. [Google Scholar] [CrossRef]
- Vitale, A.; Cirvilleri, G.; Panebianco, A.; Epifani, F.; Perrone, G.; Polizzi, G. Molecular Characterisation and Pathogenicity of Aspergillus Sect. Nigri Causing Aspergillus Vine Canker of Table Grapes in Italy. Eur. J. Plant Pathol. 2011, 132, 483–487. [Google Scholar] [CrossRef]
- Ntasiou, P.; Samaras, A.; Karaoglanidis, G. Apple Fruit Core Rot Agents in Greece and Control with Succinate Dehydrogenase Inhibitor Fungicides. Plant Dis. 2021, 105, 3072–3081. [Google Scholar] [CrossRef]
- Ligoxigakis, E.K.; Papaioannou, I.A.; Markakis, E.A.; Typas, M.A. First Report of Leaf Spot of Phoenix Theophrasti Caused by Paraconiothyrium Variabile in Greece. Plant Dis. 2013, 97, 1250. [Google Scholar] [CrossRef]
- Liu, J.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal Diversity Notes 1–110: Taxonomic and Phylogenetic Contributions to Fungal Species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Phookamsak, R.; Liu, J.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.A.; Thambugala, K.M.; Dai, D.-Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viticultural Zone | Region | Number of Vineyards | Vineyard Age (Years) | ||
|---|---|---|---|---|---|
| <10 | 10–19 | ≥20 | |||
| Northern Greece | Amyntaio | 78 | 10 | 39 | 29 |
| Kavala | 60 | 8 | 27 | 25 | |
| Lemnos | 51 | 21 | 14 | 16 | |
| Naousa | 20 | 0 | 17 | 3 | |
| Nea Mesimvria | 25 | 9 | 7 | 9 | |
| Central Greece | Nemea | 28 | 3 | 8 | 17 |
| Southern Greece | Heraklion | 48 | 19 | 4 | 25 |
| Total | 310 | 70 | 116 | 124 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Testempasis, S.I.; Markakis, E.A.; Tavlaki, G.I.; Soultatos, S.K.; Tsoukas, C.; Gkizi, D.; Tzima, A.K.; Paplomatas, E.; Karaoglanidis, G.S. Grapevine Trunk Diseases in Greece: Disease Incidence and Fungi Involved in Discrete Geographical Zones and Varieties. J. Fungi 2024, 10, 2. https://doi.org/10.3390/jof10010002
Testempasis SI, Markakis EA, Tavlaki GI, Soultatos SK, Tsoukas C, Gkizi D, Tzima AK, Paplomatas E, Karaoglanidis GS. Grapevine Trunk Diseases in Greece: Disease Incidence and Fungi Involved in Discrete Geographical Zones and Varieties. Journal of Fungi. 2024; 10(1):2. https://doi.org/10.3390/jof10010002
Chicago/Turabian StyleTestempasis, Stefanos I., Emmanouil A. Markakis, Georgia I. Tavlaki, Stefanos K. Soultatos, Christos Tsoukas, Danai Gkizi, Aliki K. Tzima, Epameinondas Paplomatas, and Georgios S. Karaoglanidis. 2024. "Grapevine Trunk Diseases in Greece: Disease Incidence and Fungi Involved in Discrete Geographical Zones and Varieties" Journal of Fungi 10, no. 1: 2. https://doi.org/10.3390/jof10010002