
Comparative Assessment of Habitat Suitability and Niche Overlap of Three Cytospora Species in China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Occurrence Records
2.2. Environmental Factor Variables
2.3. Model Simulation, Optimization, and Evaluation
2.4. Assessment of Current and Future Distribution Areas
2.5. Niche Overlap Analysis
3. Results
3.1. Model Performance and Variable Contribution
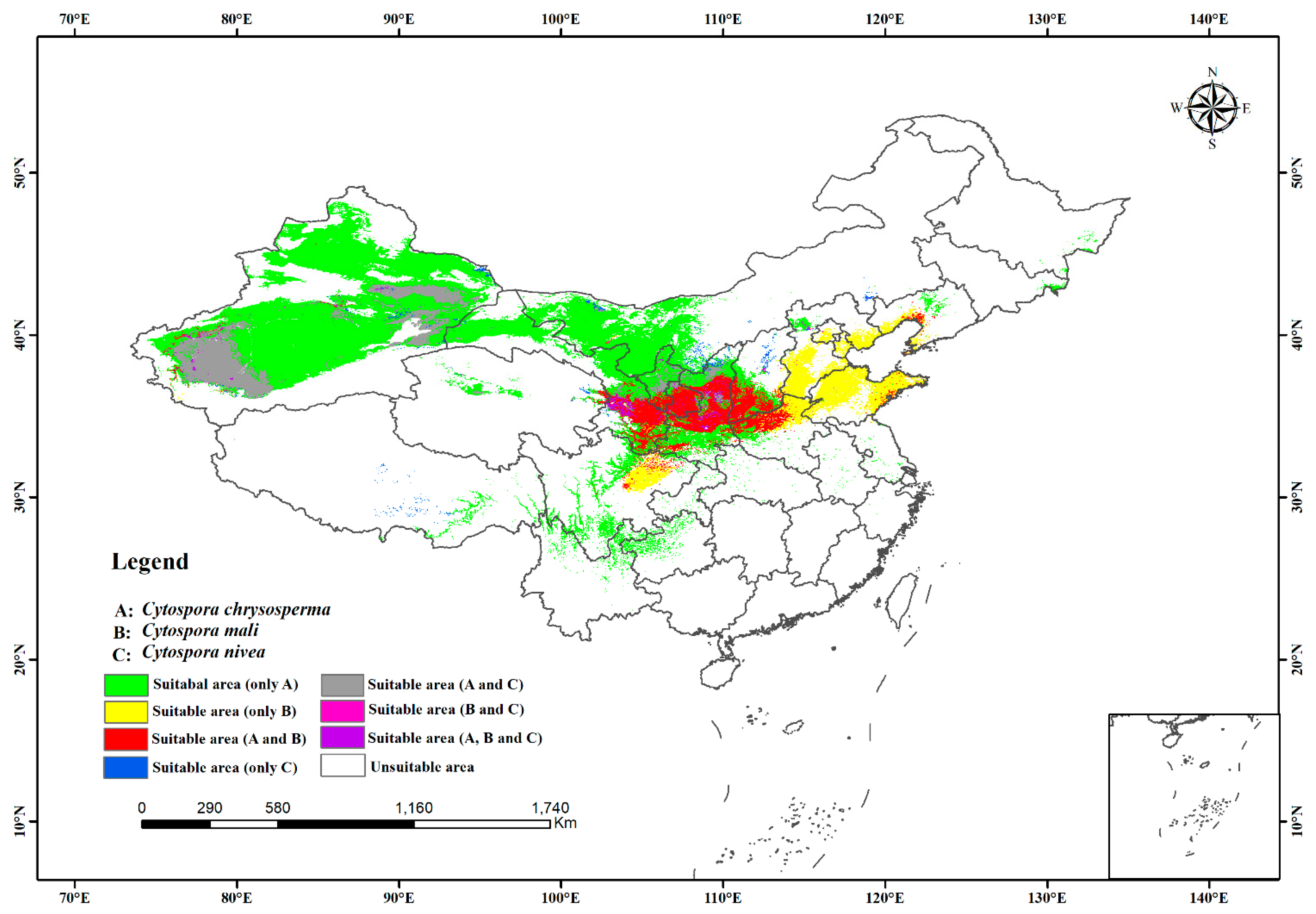
3.2. Current Distribution Regions of the Three Species
3.3. Future Distribution Changes of the Three Species
3.4. Comparisons of Overlapping Areas, Distribution Centroids, and Ecological Niches for the Three Studied Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Change 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Guo, Y.L.; Li, X.; Zhao, Z.F.; Nawaz, Z. Predicting the impacts of climate change, soils and vegetation types on the geographic distribution of Polyporus umbellatus in China. Sci. Total Environ. 2019, 648, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Zhang, W.; Peng, Y.; Zhao, F.; Chang, X.Q.; Xing, K.; Zhu, L.; Ma, G.; Yang, H.P.; Rudolf, V.H.W. Climate warming promotes pesticide resistance through expanding overwintering range of a global pest. Nat. Commun. 2021, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.X.; Guillera-Arroita, G.; May, T.W.; Lahoz-Monfort, J.J.; Elith, J. Using Species Distribution Models For Fungi. Fungal Biol. Rev. 2020, 34, 74–88. [Google Scholar] [CrossRef]
- Yan, C.C.; Hao, H.T.; Wang, Z.; Sha, S.S.; Zhang, Y.W.; Wang, Q.P.; Kang, Z.S.; Huang, L.L.; Wang, L.; Feng, H.Z. Prediction of Suitable Habitat Distribution of Cryptosphaeria pullmanensis in the World and China under Climate Change. J. Fungi 2023, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.C.; Wingfield, M.J.; Common, R.; Roux, H. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Stud Mycol. Stud. Mycol. 2004, 52, 1–144. [Google Scholar]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Persoonia 2020, 45, 1–45. [Google Scholar] [CrossRef]
- Palavouzis, S.C.; Tzamos, S.; Paplomatas, E.; Thomidis, T. First report of cytospora punicae isolated from pomegranate plants with symptom of collar rot in northern Greece. J. Plant Pathol. 2015, 97, 216. [Google Scholar]
- Du, Q.; Zhao, S.F.; Wu, C.L.; Kong, L.; Zhang, P. Root Rot of Chinese Jujube (Ziziphus jujuba) Caused by Cytospora sacculus in China. Plant Dis. 2015, 97, 1661. [Google Scholar] [CrossRef]
- Guyon, J.; Hoffman, J. Survey of aspen disease in the intermountain region. USDA For. Serv. For. Health Prot. Ogden Utah. 2011. Available online: https://digitalcommons.usu.edu/aspen_bib/7040/ (accessed on 1 December 2023).
- Worrall, J.J.; Rehfeldt, G.E.; Hamann, A.; Hogg, E.H.; Marchetti, S.B.; Michaelian, M.; Gray, L.K. Recent declines of Populus tremuloides in North America linked to climate. For. Ecol. Manag. 2013, 299, 35–51. [Google Scholar] [CrossRef]
- Rong, M. Taxonomy and Phylogeny of Cytospora in Xinjiang, Northwest of China; Beijing Forestry University: Beijing, China, 2017. [Google Scholar]
- Fan, X.L.; Tian, C.M.; Yang, Q.; Liang, Y.M.; You, C.J.; Zhang, Y.B. Cytospora from Salix in northern China. Mycotaxon 2014, 129, 303–315. [Google Scholar] [CrossRef]
- Xinlei, F. Phylogeny and Taxonomy of Cytospora in Yellow River Region of China; Beijing Forestry University: Beijing, China, 2016. [Google Scholar]
- Shasha, X. The Occurrence, Pathogenic Structural Composition and Genetic Diversity of Walnut Canker Disease in Xinjiang; Tarim University: Alar, China, 2022. [Google Scholar]
- Zhe, W. Pathogen Population Composition of Fragrant Pear Valsa Canker in Fragrant Pear Trees and Transcriptome Analysis of Dominant Species Cytospora pyri; Tarim University: Alar, China, 2023. [Google Scholar]
- Sha, S. Population Structure Analysis of Apple Valsa Canker Fungus and Evaluation of the Biocontrol Potential of Bacillus velezensis SY01 Surrounding the Tarim Basin; Tarim University: Alar, China, 2023. [Google Scholar]
- Yinming, L. Study of Ascomycetous Species on Branches Disease from Desert Plants in Xinjiang; Xinjiang Agricultural University: Urumqi, China, 2017. [Google Scholar]
- Jia, W.; Ma, Q.; Su, J.; Li, Z.; Liu, Y.; Song, P. Pathogen Identification of Poplar Stem Rot in Hohhot of InnerMongolia. J. Northwest For. Univ. 2022, 37, 210–215+237. [Google Scholar]
- Wang, Y.; Xie, S.; Cao, J.; Zhao, H.; Yin, X.; Guo, Y.; Xu, C.; Guo, L.; Wu, H.; Zhang, M. Lasiodiplodia regiae sp. nov.: A new species causing canker and dieback of fruit trees in China. Phytopathology 2023, 113, 1210–1221. [Google Scholar] [CrossRef]
- Ayse, K.; Semra, K.; Gulsan, T.S.; Esin, S.; Takashi, S.; Gerard, A.; Gerard, V.; Richard, S. Fulminating fungal sinusitis caused by Valsa sordida, a plant pathogen, in a patient immunocompromised by acute myeloid leukemia. Med. Mycol. 2006, 44, 531–539. [Google Scholar] [CrossRef]
- Spielman, L.J. A monograph of Valsa on hardwoods in North America. Can. J. Bot. 1985, 63, 1355–1378. [Google Scholar] [CrossRef]
- Yawei, Z. The Species Distribution, Biological Characteristics and Pathogenicity of Cytospora on Populus and Salix in Xinjiang; Xinjiang Agricultural University: Urumqi, China, 2022. [Google Scholar]
- Xu, W.; Sun, H.; Jin, J.; Cheng, J. Predicting the potential distribution of apple canker pathogen (Valsa mali) in China under climate change. Forests 2020, 11, 1126. [Google Scholar] [CrossRef]
- Ke, X.; Huang, L.; Han, Q.; Gao, X.; Kang, Z. Histological and cytological investigations of the infection and colonization of apple bark by Valsa mali var. mali. Australas. Plant Pathol. 2013, 42, 85–93. [Google Scholar] [CrossRef]
- Kamiri, L.K. Epidemiology of Cytospora Canker Caused in Colorado Blue Spruce by Valsa kunzei. Phytopathology 1981, 71, 941. [Google Scholar] [CrossRef]
- Wang, S.; Hu, T.; Wang, Y.; Luo, Y.; Michailides, T.J.; Cao, K. New understanding on infection processes of Valsa canker of apple in China. Eur. J. Plant Pathol. 2016, 146, 531–540. [Google Scholar] [CrossRef]
- Fan, X.; Hyde, K.; Liu, M.; Tian, C. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 119, 310–319. [Google Scholar] [CrossRef]
- Kepley, J.B.; Reeves, F.B.; Jacobi, W.R.; Adams, G.C. Species associated with cytospora canker on Populus tremuloides. Mycotaxon 2015, 130, 783–805. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- McKay, A. Pathogens in a warming world. Nat. Ecol. Evol. 2023, 7, 2. [Google Scholar] [CrossRef]
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. Forest pathogens with higher damage potential due to climate change in Europe. Can. J. Plant Pathol. 2008, 30, 177–195. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Gu, W. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, K.; Ma, M.; Cui, Y.; Xu, J.; Chen, W.; Yang, R.; Wu, C.; Gong, G. Occurrence Regionalization of Kiwifruit Brown Spot in Sichuan. J. Fungi 2023, 9, 899. [Google Scholar] [CrossRef]
- Andrew, C.; Diez, J.; James, T.Y.; Kauserud, H. Fungarium specimens: A largely untapped source in global change biology and beyond. Philos. Trans. R. Soc. B Biol. Sci. 2018, 374, 20170392. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Hausser, J.; Chessel, D.; Perrin, N. cological-niche factor analysis: How to compute habitat-suitability maps without absence data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Pattison, R.R.; Mack, R.N. Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: Evaluating CLIMEX predictions with field trials. Glob. Change Biol. 2008, 14, 813–826. [Google Scholar] [CrossRef]
- Bell, J.F. Tree-based methods. In Machine Learning Methods for Ecological Applications; Springer: Boston, MA, USA, 1999; pp. 89–105. [Google Scholar]
- Yee, T.W.; Mitchell, N.D. Generalized additive models in plant ecology. J. Veg. Sci. 1991, 2, 587–602. [Google Scholar] [CrossRef]
- Bai, R.; Wang, J.; Li, N. Climate change increases the suitable area and suitability degree of rubber tree powdery mildew in China. Ind. Crops Prod. 2022, 189, 115888. [Google Scholar] [CrossRef]
- Zhao, H.X.; Xian, X.Q.; Yang, N.W.; Zhang, Y.J.; Liu, H.; Wan, F.H.; Guo, J.Y.; Liu, W.X. Insights from the biogeographic approach for biocontrol of invasive alien pests: Estimating the ecological niche overlap of three egg parasitoids against Spodoptera frugiperda in China. Sci. Total Environ. 2023, 862, 13. [Google Scholar] [CrossRef]
- Ajene, I.J.; Khamis, F.; van Asch, B.; Pietersen, G.; Rasowo, B.A.; Ekesi, S.; Mohammed, S. Habitat suitability and distribution potential of Liberibacter species (“Candidatus Liberibacter asiaticus“ and “Candidatus Liberibacter africanus“) associated with citrus greening disease. Divers. Distrib. 2020, 26, 575–588. [Google Scholar] [CrossRef]
- Ding, W.C.; Li, H.Y.; Wen, J.B. Response of the invasive plant Ailanthus altissima (Mill.) Swingle and its two important natural enemies (Eucryptorrhynchus scrobiculatus (Motschulsky) and E. brandti (Harold)) to climate change. Ecol. Indic. 2022, 143, 14. [Google Scholar] [CrossRef]
- Cohen, S.D. Estimating the Climate Niche of Sclerotinia sclerotiorum Using Maximum Entropy Modeling. J. Fungi 2023, 9, 892. [Google Scholar] [CrossRef]
- Ejaz, M.R.; Jaoua, S.; Ahmadi, M.; Shabani, F. An examination of how climate change could affect the future spread of Fusarium spp. around the world, using correlative models to model the changes. Environ. Technol. Innov. 2023, 31, 15. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M.J.E. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Krechemer, F.S. Prevention is better than cure: Integrating habitat suitability and invasion threat to assess global biological invasion risk by insect pests under climate change. Pest Manag. Sci. 2021, 77, 4510–4520. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Lu, Y.; Wu, T.; Li, Y.; Yang, B. Mitigation of the double ITCZ syndrome in BCC-CSM2-MR through improving parameterizations of boundary-layer turbulence and shallow convection. Geosci. Model Dev. 2021, 14, 5183–5204. [Google Scholar] [CrossRef]
- Zhou, Y.R.; Lu, X.; Zhang, G.F. Potentially differential impacts on niche overlap between Chinese endangered Zelkova schneideriana and its associated tree species under climate change. Front. Ecol. Evol. 2023, 11. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. Peerj 2017, 5, e3093. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L.J.P. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Hosni, E.M.; Nasser, M.G.; Al-Ashaal, S.A.; Rady, M.H.; Kenawy, M.A.J.S.R. Modeling current and future global distribution of Chrysomya bezziana under changing climate. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Hu, X.; Wu, F.; Guo, W.; Liu, N.J.S.S.S. Identification of potential cultivation region for Santalum album in China by the MaxEnt ecologic niche model. Sci. Silvae Sin. 2014, 50, 27–33. [Google Scholar]
- Rezaei, S.; Mohammadi, A.; Shadloo, S.; Ranaie, M.; Wan, H.Y. Climate change induces habitat shifts and overlaps among carnivores in an arid and semi-arid ecosystem. Ecol. Inform. 2023, 77, 9. [Google Scholar] [CrossRef]
- Eskildsen, A.; le Roux, P.C.; Heikkinen, R.K.; Hoye, T.T.; Kissling, W.D.; Pöyry, J.; Wisz, M.S.; Luoto, M. Testing species distribution models across space and time: High latitude butterflies and recent warming. Glob. Ecol. Biogeogr. 2013, 22, 1293–1303. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, H.; Yue, J.; Xie, T.; Xu, Y.; Tian, Y.J.F. Predicting shifts in the suitable climatic distribution of walnut (Juglans regia L.) in China: Maximum entropy model paves the way to forest management. Forests. 2018, 9, 103. [Google Scholar] [CrossRef]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Hyseni, C.; Garrick, R.C. Ecological Drivers of Species Distributions and Niche Overlap for Three Subterranean Termite Species in the Southern Appalachian Mountains, USA. Insects 2019, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Rödder, D.; Engler, J.O. Quantitative metrics of overlaps in Grinnellian niches: Advances and possible drawbacks. Glob. Ecol. Biogeogr. 2011, 20, 915–927. [Google Scholar] [CrossRef]
- Yan, C.; Hao, H.; Feng, H.; Wang, Z.; Sha, S.; Li, M.; Wang, L.; Kang, Z. Whole genome sequence of Cryptosphaeria pullmanensis, an important pathogenic fungus potentially threatening crop and forestry production. Genomics 2023, 115, 110576. [Google Scholar] [CrossRef] [PubMed]
- Moya, W.; Jacome, G.; Yoo, C. Past, current, and future trends of red spiny lobster based on PCA with MaxEnt model in Galapagos Islands, Ecuador. Ecol. Evol. 2017, 7, 4881–4890. [Google Scholar] [CrossRef]
- Ye, X.; Zhang, M.; Yang, Q.; Ye, L.; Liu, Y.; Zhang, G.; Chen, S.; Lai, W.; Wen, G.; Zheng, S. Prediction of suitable distribution of a critically endangered plant Glyptostrobus pensilis. Forests 2022, 13, 257. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Zhang, J.; Nielsen, S.E.; Chen, Y.; Georges, D.; Qin, Y.; Wang, S.S.; Svenning, J.C.; Thuiller, W. Extinction risk of North American seed plants elevated by climate and land-use change. J. Appl. Ecol. 2017, 54, 303–312. [Google Scholar] [CrossRef]
- Yu, F.Y.; Wang, T.J.; Groen, T.A.; Skidmore, A.K.; Yang, X.F.; Ma, K.P.; Wu, Z.F. Climate and land use changes will degrade the distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef]
- Chen, C.; Li, B.H.; Dong, X.L.; Wang, C.X.; Lian, S.; Liang, W.X. Effects of Temperature, Humidity, and Wound Age on Valsa mali Infection of Apple Shoot Pruning Wounds. Plant Dis. 2016, 100, 2394–2401. [Google Scholar] [CrossRef]
- Min, X.J.; Zang, Y.X.; Sun, W.; Ma, J.Y. Contrasting water sources and water-use efficiency in coexisting desert plants in two saline-sodic soils in northwest China. Plant Biol. 2019, 21, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.R. Biological Characteristics of the Pathogens from Different Host Plants Caused Canker and Screening Fungicide against Pear Valsa; Huazhong Agriculture University: Wuhan, China, 2015. [Google Scholar]
- Zhao, Q.; Li, R.; Gao, Y.Y.; Yao, Q.; Guo, X.Q.; Wang, W.G. Modeling impacts of climate change on the geographic distribution of medicinal plant Fritillaria cirrhosa D. Don. Plant Biosyst. 2018, 152, 349–355. [Google Scholar] [CrossRef]
- Ljubicic, I.; Varga, F.; Bogdanovic, S.; Sklepic, L.; Britvec, M.; Temunovic, M. Comparative assessment of habitat suitability and niche overlap of three medicinal and melliferous Satureja L. species (Lamiaceae) from the eastern Adriatic region: Exploring potential for cultivation. Ecol. Inform. 2023, 76, 13. [Google Scholar] [CrossRef]
- Wang, W.T.; Gao, S.Y.; Wang, S.F. Predictive studies of potential invasive areas for four poisonous weeds in Gansu grassland. Acta Ecol. Sin. 2019, 39, 5301–5307. [Google Scholar]
- Silvertown, J.W. The distribution of plants in limestone pavement-tests of species interaction and niche separation against null hypotheses. J. Ecol. 1983, 71, 819–828. [Google Scholar] [CrossRef]
- Abolmaali, S.M.R.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. chrysosperma | C. mali | C. nivea | |
|---|---|---|---|
| n | 374 | 164 | 166 |
| RM | 0.4 | 1.5 | 1.2 |
| FC | LQP | LQH | QT |
| ntrain | 280 | 123 | 124 |
| ntest | 94 | 41 | 42 |
| Train. AUC | 0.9186 | 0.9892 | 0.9654 |
| avg.test.AUC | 0.921 | 0.987 | 0.946 |
| AICc | 10,614.12 | 3921.17 | 4564 |
| delta.AICc | 0 | 0 | 0 |
| TSS | 0.797 | 0.898 | 0.802 |
| Climate Scenario | Period | C. chrysosperma | C. mali | C. nivea | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Centroid Coordinates | Direction | Migration Distance (between Two Adjacent Decades)/km | Centroid Coordinates | Direction | Migration Distance (between Two Adjacent Decades)/km | Centroid Coordinates | Direction | Migration Distance (between Two Adjacent Decades)/km | |||||
| Longitude/°E | Latitude/°N | Longitude/°E | Latitude/°N | Longitude/°E | Latitude/°N | ||||||||
| Current | 1970–2000 | 95.167233 | 38.975466 | 111.386901 | 36.032607 | 88.912738 | 38.976171 | ||||||
| SSP126 | 2021–2040/2030s | 96.384066 | 39.277208 | Northeast | 121.2 | 112.219521 | 36.523203 | Northeast | 94.9 | 88.8893 | 39.451271 | North | 47.3 |
| 2041–2060/2050s | 95.031757 | 39.852458 | North | 87.9 | 109.422709 | 37.353043 | Northwest | 233.1 | 89.254026 | 39.630005 | Northeast | 72.8 | |
| 2061–2080/2070s | 93.884674 | 39.677991 | Northwest | 142 | 111.562266 | 36.889047 | North | 89.9 | 89.63857 | 39.039926 | East | 70.3 | |
| 2081–2100/2090s | 94.108974 | 39.599974 | Northwest | 119.5 | 112.715643 | 37.086877 | Northeast | 167.9 | 88.766862 | 39.619537 | North | 65.4 | |
| SSP370 | 2021–2040/2030s | 94.694191 | 39.439458 | Northwest | 64.9 | 111.993312 | 36.533684 | Northeast | 78.1 | 87.694677 | 39.260652 | Northwest | 120.9 |
| 2041–2060/2050s | 96.251491 | 39.65637 | Northeast | 124.5 | 110.404936 | 36.987467 | Northwest | 136.6 | 90.421173 | 39.58158 | Northeast | 157.5 | |
| 2061–2080/2070s | 96.823543 | 40.015276 | Northeast | 190.1 | 110.302057 | 38.467943 | Northwest | 269.6 | 92.634879 | 39.851333 | Northeast | 369.5 | |
| 2081–2100/2090s | 95.217266 | 40.065931 | North | 108 | 109.974273 | 39.039433 | Northwest | 334.7 | 92.541241 | 39.471202 | Northeast | 353.5 | |
| SSP585 | 2021–2040/2030s | 96.086851 | 39.460776 | Northeast | 101 | 111.449265 | 36.712647 | North | 70.4 | 92.446198 | 39.424921 | Northeast | 343.8 |
| 2041–2060/2050s | 95.797745 | 39.988979 | Northeast | 117.3 | 109.659796 | 37.813257 | Northwest | 247.1 | 88.884588 | 39.648996 | North | 66.8 | |
| 2061–2080/2070s | 94.599344 | 39.759687 | Northwest | 95.1 | 111.468502 | 38.464054 | North | 248.2 | 90.324758 | 39.656755 | Northeast | 152.1 | |
| 2081–2100/2090s | 93.923118 | 39.618821 | Northwest | 136 | 111.381883 | 39.413539 | North | 342.8 | 90.021173 | 39.851333 | Northeast | 137.7 | |
| Species | Niche Overlap (D) | Niche Overlap (I) | Range Overlap | Niche Breadth | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C. chrysosperma | C. mali | C. nivea | C. chrysosperma | C. mali | C. nivea | C. chrysosperma | C. mali | C. nivea | ||
| C. chrysosperma | 1 | 0.6093 | 0.6848 | 1 | 0.8730 | 0.8956 | 1 | 0.5285 | 0.9246 | 0.49 |
| C. mali | - | 1 | 0.5488 | - | 1 | 0.8342 | - | 1 | 0.0912 | 0.31 |
| C. nivea | - | - | 1 | - | - | 1 | - | - | 1 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, C.; Hao, H.; Sha, S.; Wang, Z.; Huang, L.; Kang, Z.; Wang, L.; Feng, H. Comparative Assessment of Habitat Suitability and Niche Overlap of Three Cytospora Species in China. J. Fungi 2024, 10, 38. https://doi.org/10.3390/jof10010038
Yan C, Hao H, Sha S, Wang Z, Huang L, Kang Z, Wang L, Feng H. Comparative Assessment of Habitat Suitability and Niche Overlap of Three Cytospora Species in China. Journal of Fungi. 2024; 10(1):38. https://doi.org/10.3390/jof10010038
Chicago/Turabian StyleYan, Chengcai, Haiting Hao, Shuaishuai Sha, Zhe Wang, Lili Huang, Zhensheng Kang, Lan Wang, and Hongzu Feng. 2024. "Comparative Assessment of Habitat Suitability and Niche Overlap of Three Cytospora Species in China" Journal of Fungi 10, no. 1: 38. https://doi.org/10.3390/jof10010038