
Fungal Endophytes: Discovering What Lies within Some of Canada’s Oldest and Most Resilient Grapevines
by
 , , , and
, , , and
Shawkat Ali
1,*,†,
A. Harrison Wright
1,†,
Joey B. Tanney
2,
Justin B. Renaud
3 and
Mark W. Sumarah
3 1
Agriculture and Agri-Food Canada, Kentville Research and Development Centre, 32 Main St., Kentville, NS B4N 1J5, Canada
2
Natural Resources Canada, Pacific Forestry Centre, 506 Burnside Road West, Victoria, BC V8Z 1M5, Canada
3
Agriculture and Agri-Food Canada, London Research and Development Centre, 1391 Sandford St., London, ON N5V 4T3, Canada
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
J. Fungi 2024, 10(2), 105; https://doi.org/10.3390/jof10020105
Submission received: 7 December 2023
/
Revised: 10 January 2024
/
Accepted: 17 January 2024
/
Published: 26 January 2024
(This article belongs to the Special Issue Fungal Endophytes in Agriculture)
Abstract
:Plant diseases and pests reduce crop yields, accounting for global crop losses of 30% to 50%. In conventional agricultural production systems, these losses are typically controlled by applying chemical pesticides. However, public pressure is mounting to curtail agrochemical use. In this context, employing beneficial endophytic microorganisms is an increasingly attractive alternative to the use of conventional chemical pesticides in agriculture. A multitude of fungal endophytes are naturally present in plants, producing enzymes, small peptides, and secondary metabolites due to their bioactivity, which can protect hosts from pathogens, pests, and abiotic stresses. The use of beneficial endophytic microorganisms in agriculture is an increasingly attractive alternative to conventional pesticides. The aim of this study was to characterize fungal endophytes isolated from apparently healthy, feral wine grapes in eastern Canada that have grown without agrochemical inputs for decades. Host plants ranged from unknown seedlings to long-lost cultivars not widely propagated since the 1800s. HPLC-MS was used to identify unique endophyte-derived chemical compounds in the host plants, while dual-culture competition assays showed a range in endophytes’ ability to suppress the mycelial growth of Botrytis, which is typically controlled in viticulture with pesticides. Twelve of the most promising fungal endophytes isolated were identified using multilocus sequencing and morphology, while DNA barcoding was employed to identify some of their host vines. These fungal endophyte isolates, which consisted of both known and putative novel strains, belonged to seven genera in six families and five orders of Ascomycota. Exploring the fungal endophytes in these specimens may yield clues to the vines’ survival and lead to the discovery of novel biocontrol agents.
1. Introduction
When Leif Eriksson and his crew of Icelandic Norsemen first travelled to North America (c. 1000 AD), they called it Vinland, purportedly for the wild grapes that grew in abundance there [1,2]. The people indigenous to what is now referred to as the Canadian Maritimes were of course familiar with these wild grapes, which, in the form of fresh fruit, made up a small, seasonal part of their diet [3]. When a second wave of both English and French European explorers and settlers began to arrive in the early 1600s, several contemporary accounts note their interest in the wild Vitis species that they encountered [4]. Furthermore, a handful of optimistic European settlers, possibly inspired by their observations of the local flora, established and tended early small-scale plantings in the Canadian Maritimes, including some of the first vines to be cultivated in Canada [4].
Although favourable microclimates for grape growing exist in the Maritimes, and the local grape industry is currently undergoing rapid expansion and growth [5], likely due in part to recent climate warming, the region’s climate does not immediately seem auspicious for viticulture. In fact, commercial viticulture in the region was deemed untenable by early researchers [6]. While the maritime influence results in relatively mild winters, the region also experiences high humidity and heavy precipitation and has a shorter growing season and fewer growing degree days (GDDs) than other important grape-growing regions in Canada. In spite of this, a genuine wine grape industry was born and now thrives there, with the first commercial vineyards springing up in the 1970s and 1980s [4]. However, remnants of much earlier plantings—with both North American and European lineages, some dating back centuries—still persist in many places throughout the Maritimes today. So, given the long history of both indigenous and European grapes in the Maritimes, which is on the edge of where wine grapes are traditionally grown, what microbial communities might be living inside some of these specimens that have stood the test of time?
Complex and diverse microbial communities (microbiomes) exist inside all plants. Although the relationship between these microbes and their hosts ranges from mutualism to pathogenicity, most of these associations are in the little-known middle portion of this spectrum, according to the current understanding [7,8]. Some of these relationships benefit both the endophyte and its plant host, and possibly co-evolved over time [9]. The means by which these endophytes may improve plant health or defend a host plant from pathogens can take various forms, such as competition for space, the production of bioactive metabolites, including antifungal agents, mycoparasitism, or the promotion of plant growth via hormones or an upregulation of natural defense mechanisms [10,11,12,13].
Unfortunately, for the vast majority of the history of modern horticulture, the relationship between the plants we grow and these microorganisms has been largely ignored. Instead, gains in productivity have been achieved by fulfilling plants’ growth requirements using synthetic fertilizers and eliminating factors harmful to growth, such as pests and disease, which account for global losses in the range of 30% to 50%, through the use of pesticides. This over-simplified approach ignores and often disrupts the beneficial natural roles played by many microorganisms. In recent times, with a greater understanding of the adverse effects of agrochemicals on the environment and animal and human health and the resulting tightening of residue restrictions, public pressure is mounting to reduce the use of synthetic fertilizers and pesticides in agriculture. This change is encouraging both governments and private companies to pursue cleaner alternative technologies, including the use of beneficial endophytes as potential growth promoters and biological control agents in commercial plant production [9,14,15,16,17,18].
While other research groups have previously explored the use of endophytic fungi in wild grapes as potential biocontrol agents [19,20], our study is unique in its focus on wine grape cultivars growing ferally in a region probably quite unlike that where most of their evolution took place. The main objective of this study was to isolate, identify, and characterize the fungal endophytes in vines (especially those with some European ancestry) that have managed to persevere and even thrive in the cool, wet climate of the Maritimes for many decades or even centuries. We, therefore, isolated fungal endophytes from the leaves of the oldest vines and identified them through multilocus sequencing and morphology. We also investigated the chemicals that these endophytes produce in order to understand their potential roles as biocontrol agents. While the focus of this research was endophyte pre-screening, exploring the fungal endophytes found in these specimens could yield clues about their resiliency, as well as lead to the discovery of novel growth-promoting and biocontrol agents as part of future research.
2. Materials and Methods
2.1. Collection of Grape Samples and Varietal Testing
More than 80 leaf samples were collected from grapevines across the province of Nova Scotia, Canada. Candidates chosen for sampling were unsprayed, uncared for, and relatively free from disease. Promising specimens were found in locations ranging from woods and the edge of meadows (often long-lost farmsteads) hosting wild and feral grapevines to historic properties containing unruly patches of grapevines of unknown origin. Varietal testing was performed using DNA barcoding (Foundation Plant Services, UC Davis, CA, USA) on vines from a small subsection of the locations thought to be the most likely to contain early named cultivars (e.g., specimens growing near early European settlement sites). Four Canadian sites, and the vines found on them, were chosen for DNA barcoding. They included (1) feral vines in the woods surrounding Bear River, Nova Scotia, Canada a town founded in the early 1600s and rumoured to have early grape plantings; (2) a vine located in the Annapolis Royal Historic Gardens (Annapolis Royal, NS) sourced from feral vines in Bear River, Nova Scotia, and transplanted to its present location in 1980; (3) a vine growing around the old foundations of a former farmstead in Miller Point Peace Park outside Lunenburg, Nova Scotia, a UNESCO World Heritage Site. This vine was supposedly a ‘Diana’ and approximately 170 years old, according to a nonagenarian living in the area who said that, as a young boy, he had harvested fruit from it, as had his father and grandfather (personal communication), and (4) the French Mission in Poplar Grove, Nova Scotia, purportedly built in 1699 and believed to be the oldest standing building in Canada east of Quebec. Grapevines grow all over this historic property and locals believe that early monks used the grapes at this site to make communal wine.
2.2. Leaf Sterilization and Isolation of Foliar Fungal Endophytes
The sterilization and isolation procedure was modified from that described in [21]. Five healthy leaves were randomly chosen from each vine sampled. For each leaf sample, the petiole was removed, and the blade was bisected by a transverse cut. Three 1 cm2 pieces were then cut out of the basal portion of each leaf and retained; the remainder of the leaf was discarded. The five petioles and 15 leaf pieces, representing one sample, were placed inside a tea infuser. In a sterile laminar flow hood, the infuser containing the sample was immersed in a beaker containing 75% ethanol, swirled for one minute, and then transferred to a beaker containing 1 L of 6% sodium hypochlorite solution and 0.05% TWEEN 80, which was swirled intermittently for 7.5 min. The infuser was then transferred to a second ethanol beaker and swirled for an additional minute before being rinsed in a series of three beakers containing sterile ultrapure water (Barnstead™ Nanopure™ D11971, Van Nuys, CA, USA). Without removing them from the sterile environment, the leaf pieces were transferred to a sterilized paper towel. Interior ~0.5 cm2 pieces were removed from each section; the two ends were cut off each petiole, and the remaining piece was cut into three equal pieces of approximately ~0.5 to 1 cm in length, making 30 pieces per sample. To test sterilization efficacy, three pieces of the petiole and three pieces of the blade were pressed onto plates containing half-strength potato dextrose agar (PDA) for five minutes on each side; the plates were then sealed with parafilm and checked after two weeks to confirm the absence of microbial growth. For each sample, three pieces of each tissue type were then transferred to a plate of half-strength PDA for a total of five plates of each tissue type. The plates were then sealed with parafilm and stored in the dark at room temperature. The cut ends and edges were checked for endophyte growth every few days. To obtain pure fungal isolates, the mycelium growing out from these pieces was transferred to fresh PDA plates; in some cases, a second or third sub-culturing of the isolates was performed. For the purpose of this study, we focused exclusively on slow-growing fungal endophytes, i.e., those that took at least six days to emerge from the plated plant tissue sample. The isolated endophytes were given a sample ID based on the order in which the site/field sample was collected and the order in which the endophyte emerged from the sample after a minimum of six days (e.g., En01-2 would be the second endophyte to emerge, after a minimum of 6 d, from the first site/sample collected). For samples that produced only one endophyte, the second number was not used (e.g., En60).
2.3. Culturing and Morphological Observations
To induce in vitro sporulation, strains were inoculated on several media, including 2% malt extract (MEA) (20 g Bacto malt extract, Difco Laboratories, Sparks, MD, USA; 15 g agar, EMD Chemicals Inc., Gibbstown, NJ, USA; 1 L distilled water); cornmeal agar (CA) (Difco Laboratories, Detroit, MI, USA); potato dextrose agar (PDA) (Difco Laboratories, Detroit, MI, USA); oatmeal agar (OA) (extract of 30 g/L boiled oatmeal, 15 g agar, 1 L distilled water); V8 juice agar (V8A) (200 mL Campbell’s V8 juice, 15 g agar, 2 g CaCO3, 800 mL distilled water); and water agar (WA) (20 g agar, 1 L tap water), with or without the addition of sterile filter paper or autoclaved leaves of Gaultheria shallon, Hedera helix, Ilex aquifolium, or Rubus armeniacus [22]. Cultures were incubated at 20 °C in the dark or with a 12 h:12 h fluorescent light cycle, or near a window under ambient conditions.
Micromorphological characters were visualized, described, and measured from living material mounted in deionized or tap water using a Leica DM4 B light microscope (Leica Microsystems CMS GmbH, Wetzlar, Germany). Hand sections of perithecia and pycnidia were obtained using a double-edge safety razor blade. Micrographs were captured using a Leica DFC450 C camera (Leica Microsystems CMS GmbH, Wetzlar, Germany) with Leica LAS X 33.7.1 software. Colonies were observed and photographed using a Leica M165 C stereomicroscope with a TL5000 Ergo light base (Leica Microsystems Ltd., Singapore) and a Leica DMC5400 camera. Photographic plates were assembled using Adobe Photoshop CC 2019 v.23.0.1 (Adobe Systems, San Jose, CA, USA).
2.4. DNA Extraction, Amplification, Sequencing, and Analysis
For DNA extraction, a sterilized scalpel was used to scrape 10–15 mg of mycelium from 5–10-day-old cultures grown on PDA medium at 22 °C. The mycelium was then macerated with steel beads in a 2 mL Eppendorf tube using a Qiagen TissueLyser II (Qiagen, Hilden, Germany) for 2 × 60 s cycles at 30 beats per second. Total genomic DNA was isolated using an E.Z.N.A.® SP Fungal DNA Mini Kit (Omega Bio-Tek, Norcross, GA, USA), following the manufacturer’s instructions. The partial β-tubulin (TUB2), internal transcribed spacer (ITS), translation elongation factor 1-alpha (TEF-1α), RNA polymerase second largest subunit (RPB2), partial actin (ACT), chitin synthase I (CHS-1), and LSU genes were amplified by PCR using the primer pairs shown in Table S1. The PCR amplification was carried out in a 50 μL reaction mixture using the same PCR conditions as previously described [23,24]. PCR-amplified products for all genes were run on 1% agarose in a 1X Tris-borate-EDTA running buffer to confirm the size and amplification of the single band. The PCR-amplified products were then cleaned with an ExoSAP-IT™ PCR Product Cleanup kit (Applied Biosystems, Waltham, MA, USA), following the manufacturer’s protocol. Sequencing was carried out at the Genome Quebec Innovation Centre and Eurofins Genomics using ABI BigDye 3 Terminator Cycle sequencing chemistry (Applied Biosystem’s 3730xl DNA Analyzer technology).
Sequence contigs were assembled and trimmed using Geneious Prime 2019 v.2019.0.4 (Biomatters, Auckland, New Zealand). Using BLASTn (Nucleotide BLAST: Search nucleotide databases using a nucleotide query (nih.gov), https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&BLAST_SPEC=GeoBlast&PAGE_TYPE=BlastSearch, accessed on 10 August 2023), the ITS sequences obtained were compared with those in the National Centre for Biotechnology Information (NCBI) GenBank database, and strains were tentatively identified against available ITS sequences in the NCBI GenBank. Following these identifications, additional taxon-specific secondary barcodes were sequenced and analyzed (Table 1 and Table S1). Phylogenetic analyses were performed for each taxon using the appropriate gene sequence datasets populated with sequences based on NCBI BLAST queries. Sequences were aligned using MAFFT (Multiple Alignment using Fast Fourier Transform, https://mafft.cbrc.jp/alignment/server/index.html, accessed on 10 August 2023) [25] and then manually verified and adjusted if required. Phylogenetic trees were constructed with maximum likelihood (ML) using IQ-TREE v1.6.11 [26]. The best model for each partition was selected using ModelFinder [27], which performed 1000 ultrafast bootstraps [28], 1000 SH-aLRT branch tests, and an approximate Bayes test [29]. The best-fit substitution models based on the Bayesian information criterion (BIC), selected outgroup taxa, and other information on each analysis are presented in Table 1. Phylogenetic trees were built with Geneious (Geneious Prime 2019 v.2019.0.4 (Biomatters, Auckland, New Zealand) and edited with Adobe Illustrator CC 2019 v.23.0.1.
2.5. Screening of Endophyte Bioactivity against Botrytis
The isolated fungal endophyte strains were screened for their bioactivity against the common grape pathogen Botrytis cinerea using dual-culture competition assays [10]. Briefly, a 4 mm agar plug containing mycelia was removed from both a freshly grown pure culture of Botrytis cinerea and the endophyte isolate of interest and transferred to a new 9 cm PDA plate using an aseptic technique. The plugs were kept 7 cm apart from each other and 1 cm away from the edge of the plate. The starting time of the co-culture was based on the growth rate of endophytes and pathogens. In instances where the endophyte grew slowly compared to Botrytis, the endophyte was incubated on the plate for 3–5 days before the pathogen was added; before the endophyte reached the middle of the plate, the same plate was inoculated with Botrytis. The experiment was carried out in triplicate and repeated three times. As a control, a solitary plug of either Botrytis cinerea or the endophyte strain was placed on PDA plates to ensure the pathogen grew normally in the absence of the endophytes, and the endophyte grew normally in the absence of Botrytis. The plates were incubated in the dark at 22 °C and the growth of both endophyte and pathogen were observed daily. Before the two cultures touched each other, the growth of Botrytis on each plate was measured towards the pathogen (P) and was oriented vertically (VP) and oriented horizontally towards both sides of the plate (HP). The percent growth inhibition index (GII) was calculated as (HP-VP)/HP × 100 [76]. Only endophytes that showed some ability to inhibit Botrytis growth (GII > 5% based on averaging alone) were characterized further. An ANOVA was later performed on this group to test for significance, while a post hoc Tukey multiple means comparison was used to determine if specific averages differed significantly [77].
2.6. Metabolite Screening by High-Resolution HPLC-MS
Extracts from fungal endophytes and Botrytis were analyzed using a Q-Exactive Quadrupole-Orbitrap mass spectrometer (MS) (Thermo Scientific, Waltham, MA, USA) coupled with an Agilent 1290 ultra-high-performance liquid chromatography (HPLC) system. Metabolites were resolved using a Zorbax Eclipse Plus RRHD C18 column (2.1 × 50 mm, 1.8 μm; Agilent Technologies, Santa Clara, CA, USA) maintained at 35 °C. The mobile phase consisted of (A) water with 0.1% formic acid and (B) acetonitrile with 0.1% formic acid (Optima grade, Fisher Scientific, Branchburg, NJ, USA). The gradient consisted of 0% B for 30 s before increasing to 100% over 3 min. The mobile phase B was held at 100% for 2 min before returning to 0% over 30 s. Heated electrospray ionization was performed in positive mode using the following settings: capillary voltage, 3.9 kV; capillary temperature, 400 °C; sheath gas, 19 units; auxiliary gas, 8 units; probe heater temperature, 450 °C; and S-Lens RF level, 45.00. MS data were acquired using untargeted data-dependent acquisition (DDA), which included a full MS scan at a 70,000 resolution with a scan range of 106.7–1600 m/z, an automatic gain control target of 3 × 106, and a maximum injection time of 250 ms. The five most intense ions in each full scan were selected for tandem mass spectrometry (MS/MS) analysis using a 1.2-Da isolation window and were analyzed under the following conditions: resolution, 17,500; automatic gain control target, 1 × 106; max injection time, 64 ms; normalized collision energy, 35%; intensity threshold, 1.5 × 105; and dynamic exclusion, 5 s.
3. Results
3.1. Grape Site/DNA Barcoding Results
The DNA barcoding results for the grapevines sampled from the four most promising (i.e., historically significant) sites were as follows: (1) surrounding woods, Bear River, NS, Canada: most likely a seedling from an unknown mix of North American species and V. vinifera ancestry; (2) Annapolis Royal Botanical Gardens, Annapolis Royal, NS: ‘Isabella’; (3) abandoned farmstead, Miller Point Peace Park, Lunenburg, NS: a seedling made up of entirely native North American species; and (4) French Mission, Poplar Grove, NS: ‘Clinton.’
3.2. Isolation and Identification of Endophyte Strains
A total of 12 fungal endophyte strains met the criteria for further characterization: (a) they were collected from a historically significant site; (b) they did not emerge from the explant material for at least six days; and (c) they showed some antagonism towards Botrytis based on averaging (GII > 5%). On the basis of morphology, GenBank queries, and subsequent phylogenetic analyses, nine of the twelve strains were confidently identified as species (Table 1). Six strains belonged to the genus Diaporthe (Diaporthaceae, Diaporthales), including four strains of D. eres (formally D. vacuae; En25-6, En26-4, En26-5, En61-3), which make up part of a strongly supported clade containing the former D. vacuae (CAA823) and D. cotoneastri (CBS 439.82) types in the TUB2-EF1-ITS phylogeny (Figure 1). En20-4 was not conclusively identified and was placed basally in a strongly supported clade that includes the D. angelicae (CBS 111592), D. cucurbitae (DAOM 42078), and D. gulyae (BRIP 54025) types. The remaining strain (En01-1) is a putative novel species basal to the D. gulyae clade and a clade including the D. arctii (DP0492) and D. neoarctii (CBS 109490) types. The En25-6 (D. eres) strain formed condiomata that produced alpha conidia on water agar amended with Ilex aquifolium leaves (Figure 2A–D); En20-4 produced alpha and beta conidia on OA (Figure 2E–H); and En01-1 produced both alpha and gamma conidia and fertile perithecia on WA amended with Rubus armeniacus leaves (Figure 2I–N). En61-1 was identified as Gnomoniopsis paraclavulata (Diaporthaceae, Diaporthales) based on its sequence (ITS and RPB2) similarity with the type specimen (BPI 877448) and the morphology of the conidiomata and the conidia formed on WA amended with R. armeniacus leaves (Figure 3I–K and Figure 4C).
En25-3 is Colletotrichum fioriniae (Glomerellaceae, Glomerellales), with 100% similarity to the ITS and CHS-1 sequences and the ex-type specimen of C. fioriniae (CBS 128517) (Figure 5A). Conidiomata production was stimulated on WA with the addition of R. armeniacus leaves (Figure 3G–H). En61-2 was identified as Neophaeomoniella niveniae (Celotheliaceae, Phaeomoniellales) based on the similarity to the ITS and EF1a sequences with the ex-type N. niveniae strain (CBS131316) and the comparison of its conidiomata and yeast-like conidia, which were abundant when grown on V8A (Figure 3A–F and Figure 4B). En25-1 was identified as Nemania aureolutea (Xylariaceae, Xylariales;) based on its 100% similarity between its ITS sequence and the authenticated strain MAR101219 and by its orange colony, a character distinguishing N. aureolutea from the closely related N. aenea [61] (Figure 5B and Figure 6A).
En60 is an unidentified Ramularia sp. (Mycosphaerellaceae, Mycosphaerellales), which shares a 99% similarity with the ITS sequences of the ex-types of R. heraclei (CBS 108969) and R. weberiana (CBS 136.23) and authenticated cultures of R. alangiicola (CPC 10299), R. gei (CBS 113977), R. interstitiales (CBS 120.68), R. ligustrina (CBS 379.52), R. pratensis (CBS 122105), and R. rumicicola (CPC 11295). The concatenated ACT-ITS-RPB2 phylogeny placed En60 in a strongly supported clade with the ex-types of R. heraclei (CBS 108969) and R. lamii var. lamii (CBS 108970) (Figure 4D and Figure 6B–F). En26-1 was identified as Sphaerulina amelanchieris (Mycosphaerellaceae, Mycosphaerellales) based on its close sequence similarity (99% EF1, ITS) to the ex-type strain CBS 135100 (Figure 4A and Figure 6G–H).
3.3. Screening of Antimicrobial Activity of Endophytic Fungi against Botrytis
According to the dual-culture competition assays performed on the twelve selected isolates thought to be potentially active against Sh, only five of the twelve isolates initially screened showed statistically significant antagonistic activity. Values ranged from a GII of 27% (En61-3) to 35% (En26-5), while no inhibition (GII = 0%) was observed in the control (Figure 7 and Figure S1). All five of the significantly antagonistic strains belonged to the genus Diaporthe, with four out of the five identified as D. eres.
3.4. Extrolite Screening of Bioactive strains
The full complement of ionizable compounds in the extracts was visualized using principal component analysis (PCA) (Figure S2). As expected, the four strains of D. eres were grouped together; however, no significant clusters of the other strains were observed. Specifically, the other Diaporthe species (corresponding to isolates En20-4 and En01-1) did not strongly group with D. eres. C. fioriniae (En25-3) was an outlier (Figure S2): the strain’s unique extrolite profile that is responsible for its outlier position on the PCA plot was driven by unknown compounds (Table S2). Four of the extrolites, with the formulas C18H34O5, C21H34O4, C11H20O6, and C11H18O5, are likely structurally similar according to their respective MS/MS spectra (Figure S3c–f). Searching through published microbial compound databases [78] did not yield any likely candidates for these compounds, thus making them attractive targets for isolation and characterization. Strain En25-3 also had higher levels of carnitine, a common quaternary ammonium compound, than the other tested strains.
En61-1, like N. niveniae, produced several known, but unique, secondary metabolites (Figure S4 and Table S2). A number of linear dipeptides were detected as products of this strain, notably Ser-Val, Leu-Val, Ile-Thr, Ile-Ile, Ile-Pro, and Ile-Phe, although the positions of the amino acids are only putative. The series of unique extrolites produced by the strain G. paracalvulata was limited compared to the other tested strains and showed some degree of overlap with N. niveniae. Two metabolites detected in this strain have the formulas C20H21NO6 and C25H29NO8, which, based on their respective MS/MS product spectra, are likely structurally similar (Figure S5). The similarity of extrolite production in G. paracalvulata and N. niveniae is shown by their close proximity in Figure S2.
Six of the strains tested for metabolite profiling were identified as belonging to the order Diaporthales (Figure 1 and Table 1). In agreement with the phylogeny established, the four strains identified as D. eres were found to produce a similar complement of metabolites and, therefore, are clustered together in the PCA plot (Figure S2). The En20-4 and En1-1 strains did not strongly group with each other, nor did the D. eres strains (Figures S2 and S6). Some metabolites were found to be common across most of the tested Diaporthe strains, notably the osmoprotectant proline betaine. Other metabolites detected in the D. eres strains include tryptamine and phenylethylamine. Additionally, a number of related metabolites detected in D. eres include C18H35NO2, C18H33NO2, C18H35NO, and C18H33NO (Table S2).
4. Discussion
The biological control of plant diseases through the use of antagonistic endophytic microorganisms is an alternative approach for reducing or eliminating the use of chemical pesticides in agriculture. Fungal endophytes belong to a taxonomically and metabolically diverse group of organisms that colonize different plant niches without causing any harm to the host plants. Their efficacy in controlling various plant pathogens has been investigated by a number of authors [79,80,81,82].
4.1. Feral Endophyte Host Identification
The results of DNA barcoding showed that although some vines had purely North American ancestry, others were a mix of North American and European lineages. It is no surprise that some of the tissue samples sent for DNA barcoding were found to be from unnamed seedlings. Feral grapes growing in the woods or on the edges of fields and roadways have become commonplace in the region, and are likely natural crosses distributed by birds and animals. This is most likely the case for the vine from the woods surrounding Bear River, NS, which was shown to be a mix of Vitis vinifera and North American species. However, one unexpected finding was the discovery that the old vine located on a long-abandoned homestead outside the historic town of Lunenburg, NS—widely known for nearly two centuries and believed to be ‘Diana’—is actually unnamed and a product of exclusively native American species. Native North American species are not widespread in Nova Scotia, even though they are abundant in the neighbouring province of New Brunswick. Lastly, our search yielded two named heritage cultivars: ‘Clinton,’ found at the French Mission in Poplar Grove, NS, and ‘Isabelle,’ relocated to the Annapolis Royal Historic Gardens, Annapolis Royal, NS from a specimen found growing wild over 40 years ago in Bear River, NS. Both cultivars are of historical importance and have likely been growing in Nova Scotia for well over a century. ‘Clinton’ is purely North American in its lineage, and thought to be primarily derived from Vitis riparia, but with some Vitis labrusca traits. It came to prominence in the 1830s and is widely cited as the first cultivated grape of Vitis riparia lineage. It was popular in the 19th century because of its high vigour, hardiness, and fruitfulness, and its tolerance to Phylloxera. This cultivar was once a favourite in both North America and France but is now rarely found, although it continues to be influential due to its frequent use in the past by grape breeders and its role as a founding parent of many important cultivars [83]. ‘Isabella’ is the product of a natural cross between Vitis labrusca and Vitis vinifera that was later discovered and named. The cultivar was introduced in 1816 and was the mainstay of the grape industry for the next half-century in New England and the neighbouring North Atlantic region, with extensive plantings also found in Europe. However, the once-popular cultivar eventually fell out of favour. A factor in its downfall may have been the widely held belief that the source of the Great French Wine Blight, a phenomenon that devastated the French wine industry, was Phylloxera brought from North America to Europe on the roots of the ‘Isabella’ vine [83].
4.2. Botrytis Suppression
In this study, five strains of Diaporthe successfully inhibited the growth of Botrytis in dual-culture competition assays (Figure 7). Botrytis is a major grapevine pathogen, reducing yield through bunch rot and premature cluster drop, and lowering table grape quality through postharvest fruit rot. Several spray applications of a chemical fungicide are typically used annually to control Botrytis rot, which not only increases production costs but also raises a range of the environmental issues associated with agrochemical use. Although some biological control agents have been registered for the management of Botrytis rot in grapevines, they are not very effective in completely eliminating the disease. It has been reported that species of the genus Diaporthe can alter the lifestyle between phytopathogens and endophytes on the same host or between different hosts [84,85].
In future work, it would be interesting to test Diaporthe isolates from this study in the in planta inoculation of grapevine and then challenge half of the plants with Botrytis and other common grapevine pathogens. Plants inoculated with Diaporthe but not challenged with pathogens will show whether these strains are pathogenic to grapevines. D. eres isolated from Prunus dulcis has been reported to have significant antifungal activity against three fungal pathogens: Trichothecium roseum, Fusarium avenaceum, and Alternaria alternata [86]. Polonio, et al. [87] observed that D. citri exhibited antifungal activity against F. solani and Didymella bryoniae, as well as antibacterial activity against Staphylococcus aureus. In addition, Diaporthe species isolated from the ornamental plant Pachystachys lutea were able to inhibit the growth of F. oxysporum and Colletotrichum sp. [88]. In a recent study, Verma, et al. [89] reported that D. melonis and D. longicolla suppressed the growth of Corynespora cassiicola and F. solani in dual-culture competition assays. Phomopsis oblonga (current name P. valeta) serves as a natural biocontrol agent for Dutch elm disease by acting as a feeding deterrent for elm bark beetles [90]. Our previous studies have shown that D. maritima isolated from Picea produces antiinsectant and antifungal metabolites [49]. Recently, D. miriciae, an endophytic fungus isolated from tropical medicinal plants, has been shown to produce cytochalasins, which have antifungal and antagonistic activity against plant pathogens [82].
4.3. Endophyte Diversity and Ecology
The twelve fungal endophyte strains isolated in this study represent seven genera in six families and five orders in the phylum Ascomycota. Six of the twelve strains are classified as Diaporthe, a speciose genus consisting of endophytes, saprotrophs, and pathogens that colonize a wide range of host plants [35]. This genus has more than 800 described species, and more than 950 species in its asexual state (formerly Phomopsis) [91]. Two of the isolated strains (En20-4 and En01-1) appear to represent putative novel species, based on their phylogenetic distinction from described species with sequences available in GenBank. Four strains are closely related to D. vacuae, a recently described species in the D. eres complex that has been associated with dieback and twig blight of Vaccinium corymbosum [54]. These strains also had similar extrolite profiles (Figure S2). Recent work resulted in D. vacuae and other related species in the D. eres complex being synonymized with D. eres [92]. En20-4. species are commonly associated with diseases of grapevine worldwide, notably Phomopsis cane and leaf spot, which are attributed to D. ampelina (=D. viticola), and symptoms such as stem and branch dieback, perennial cankers, vascular discoloration, and rachis necrosis [93,94,95,96]. Phomopsis cane and leaf spot can affect most parts of the grapevine, including the flowers and berries, and its potential to cause large crop losses often leads to the application of fungicides and other control measures [97]. Although D. eres causes minor disease in a broad range of hosts, it can also result in serious canker disease in grapevines, apples, and blueberries [54]. For example, strains of D. eres isolated from symptomatic apple rootstocks in the same region as this study (Annapolis Valley, NS) caused necrosis, cankers, and eventually death in young apple plants within four weeks [24]. Other Diaporthe species show varying degrees of virulence in grapevines; for example, the closely related D. ampelina, D. hispaniae, and D. hungariae are highly virulent, while other species, such as D. bohemiae, appear to be avirulent endophytes. Overall, most Diaporthe species, including D. eres, tested in grapevine pathogenicity experiments show some degree of virulence [93,98,99]. Reveglia, et al. [100] demonstrated that a D. eres strain isolated from symptomatic grapevine wood produced phytotoxic secondary metabolites; one phytotoxin, nectriapyrone, was identified in several of our Diaporthe strains. Guarnaccia, Groenewald, Woodhall, Armengol, Cinelli, Eichmeier, Ezra, Fontaine, Gramaje and Gutierrez-Aguirregabiria [39] reported that D. eres was the Diaporthe species most commonly isolated in grapevines sampled in eight countries. Taken together, the fact that Diaporthe represents the majority of endophytes isolated in this study is not unexpected. Its presence does not necessarily reflect the overall health of the sampled grapevines, given its frequency of occurrence on asymptomatic grapevines and potential host resistance [101]. However, it is notable that Kernaghan, Mayerhofer and Griffin [20] did not report any Diaporthe endophytes in wild and hybrid Vitis leaves sampled from wild grapes and vineyards in eastern Canada. Although pathogenicity experiments were not conducted in this study, the virulence of En20-4 and En01-1 should be assessed in the future, especially considering the identification of the phytotoxin nectriapyrone.
Although Gnomoniopsis paraclavulata (En61-1) was isolated from grapevine leaves, this species is best known from Quercus spp. in the US and Europe, where it is found in the stems, wood, asymptomatic leaves, overwintering leaves, leaf litter, and acorns [56,102,103,104]. The most dominant species isolated from asymptomatic and diseased stems of Quercus robur in Poland included G. paraclavulata (and, incidentally, D. eres and C. fioriniae) [62,105]. Subsequent pathogenicity tests on young Q. robur seedlings demonstrated that G. paraclavulata was the most pathogenic species tested, causing dieback symptoms and small lesions on stems. Costa, et al. [106] also identified G. paraclavulata in symptomatic and declining Q. suber in Portugal, while Tosi, et al. [107] implicated G. paraclavulata in chestnut bud and shoot blight (Castanea sativa). Given G. paraclavulata’s apparently narrow association with Quercus, or Fagaceae in general, its isolation from grape leaves is somewhat unexpected, and its interaction with this host is unknown.
N. niveniae (En61-2) is related to Phaeomoniella chlamydospora and is believed to be one of the primary causal agents of Petri disease and esca, two serious grapevine trunk diseases. Other Phaeomoniellales taxa have recently been described as a result of spore-trapping efforts in vineyards but do not appear to be pathogenic [33]. Similarly, Neophaeomoniella spp. has not been implicated in grapevine disease, although N. niveniae has been identified from symptomatic plants, albeit in low relative abundance [108]. Interestingly, N. niveniae was isolated from wild olive trees (Olea europaea subsp. cuspidata) in South Africa and was found to be present in 7.1% of the trees sampled; two strains exhibited low-to-zero virulence and intermediate virulence, respectively, when inoculated on shoots of ‘Frantoio’ olive trees (O. europaea L. subsp. europaea) in South Africa [67,109]. N. niveniae was first described from leaves of Nivenia stokoei in South Africa, where it was isolated from leaf spots, but the causality of this was not tested [66]. Our isolation of N. niveniae is likely the first record in Canada, and its interactions with grapevines are unknown. Interestingly, among the tested strains, En61-2 produced a number of dipeptides, all of which contain Ile or Leu. Although fungi are known to produce a wide range of cyclic dipeptides [110], all the ones reported here are linear and have unknown bioactivity.
Species of Colletotrichum have been implicated in various diseases of agricultural crops and other plants, such as bitter rot of pears and apples [111,112], anthracnose of apples [113], fruit rot of cranberries and blueberries [31], anthracnose fruit rot of raspberries [114], leaf spot disease of walnuts [115], anthracnose of eggplants [116], seedling blight of poison ivy (Toxicodendron radicans) [117], and leaf blight of Mahonia aquifolium [118]. Bitter rot of apples, for example, is considered a major disease of apples in warm and humid regions [104]. On the other hand, several species in this genus are capable of forming symbiotic and mutualistic relationships with plant species [119,120]. C. fioriniae was isolated as a leaf endophyte from apple trees in orchards and from various plant species growing nearby, including angiosperm trees, wild grapes, and others, leading the authors to hypothesize that the species’ main ecological role was indeed that of a leaf endophyte [104]. In the same study, the C. fioriniae strains isolated were also found to be pathogenic in apple fruit. Recently, C. fioriniae was reported as the causal agent of grapevine anthracnose in New York [121], and it is also associated with grapevine ripe rot [122]. C. fioriniae not only has been widely reported as an endophyte and pathogen in a broad range of hosts but is also cited as an entomopathogen of the elongate hemlock scale (Fiorinia externa), the origin of its specific epithet [123,124,125]. Therefore, C. fioriniae may act as a benign or potentially beneficial foliar endophyte in some plant hosts, while also reducing the quality of and accelerating decay in fruit. C. gloeosponoides has also been reported to cause epizootics of the scale insect Orthezia praelonga, a major pest of citrus in Brazil [124].
The family Xylariaceae contains a large number of commonly reported endophytes in a very broad range of host plants, including bryophytes, liverworts, angiosperms, and conifers, and even occurs in the form of endolichenic fungi [126]. These fungi are not only ubiquitous endophytes but also remarkably prolific producers of bioactive natural products [63,127]. The genus Nemania consists of approximately 80 named species (www.indexfungorum.org, accessed on 20 November 2023), including species characterized as endophytes producing described natural products [65,128,129,130,131]. N. serpens and Nemania spp. have occasionally been isolated as endophytes of cultivated and wild grapes [128,132]. We isolated N. aureolutea (En25-1), a rarely observed species that is closely related to the common N. aenea. Our strain produced the characteristic slow-growing orange colonies on PDA, as described by Fournier, Lechat and Ribes Ripoll [61] (Figure 3A). Little is known about N. aureolutea because it is rarely collected, with a small number of reports from Europe, the continental US, and Hawaii, where it grows in the wood, and sometimes on the outer bark, of Acer, Corylus, Populus, Quercus, and Salix; most collections appear to be from Quercus [61,133]. Our strain may be the first report from Canada; however, N. aureolutea is likely more common than perceived, and some specimens identified as N. aenea are actually N. aureolutea [134].
The genus Sphaerulina comprises more than 200 named species (www.indexfungorum.com, accessed on 20 November 2023), with some causing leaf spots on various plant species; notably, S. musiva is responsible for one of the most damaging diseases of poplar in northeastern and north-central North America [135]. S. amelanchier (En26-1) was originally described from Amelanchier sp. leaf litter in the Netherlands and has also been isolated from leaves of unidentified species of Betula, Castanea, and Quercus [72,136]. Given the infrequent reports, little is known about the ecology of S. amelanchier. A related species, S. vaccinii, causes a common leaf spot and stem canker disease in lowbush blueberry (Vaccinium angustifolium) in eastern Canada and the US [137,138,139]. We also isolated another Mycosphaerellaceae species, a strain (En60) of an unknown Ramularia species that is a sister species to R. heraclei and R. lamii var. lamii. Ramularia are also well-known and common associates of leaf spot diseases of varying significance in a broad range of plant hosts. For example, Ramularia leaf spots of sugar beet and barley, caused by R. beticola and R. collo-cygni, respectively, are serious emerging disease [140,141]. Species of Ramularia have been reported as endophytes of grapevines [20], while a Ramularia sp. isolated from Rumex gmelini Turcz has been noted to produce the bioactive secondary metabolite chrysophanol [142]. According to the results of the phylogenetic analysis and the lack of similarity to available sequences, Ramularia sp. (En60) is possibly a novel species.
In general, the host preferences of the endophyte species that we isolated from grapevines are either demonstrably broad (e.g., Colletotrichum fioriniae, D. eres), suggestive of a broader host range than currently known (e.g., G. paraclavulata, Nemania aureolata, Neophaeomoniella niveniae), or are unknown due to possible novelty (e.g., Diaporthe spp. En20-4 and En01-1, Ramularia sp.). Many of these endophytes produce slimy, bright yellow-orange masses of conidia that are presumably spread via rain splash and water runoff and potentially vectored by insects, including pollinators [33,143,144]. Endophytes in the Xylariaceae family (e.g., Nemania aureolata) may be transmitted horizontally to new leaves via dry, airborne conidia and ascospores. While grapevines are probably a secondary host for N. aureolata, endophytes occurring on leaves as foliar endophytes may colonize primary hosts (i.e., host substrate that supports sexual reproduction, such as hardwood trees) through direct contact (viaphytism) [145]. Therefore, the senescent and overwintered leaves and wood of both grapevines and other distantly related plant hosts may serve as inoculum for new infections. Vertical transmission of endophytes is also possible in some cases; for example, as demonstrated or suggested in Diaporthe, Ramularia, and G. paraclavulata [103,146,147].
Some grapevine endophytes are potentially opportunistic pathogens, such as Diaporthe spp. and C. fioriniae, which may affect vines, leaves, and/or fruit. Other species may be commensal or mutualistic, such as N. aureolata or even C. fioriniae, an endophyte that may protect against insect herbivory and, as demonstrated in other Colletotrichum species, provide protection from plant pathogens [31,148,149,150]. Interactions between endophytes and their hosts span the mutualist–pathogen continuum and depend on the host’s status at the given moment [151]. Consequently, the overall effect of endophyte colonization on host fitness may be difficult to generalize; for example, C. fioriniae could benefit its host by protecting leaves against insect herbivory, but its colonization of fruit could reduce seed dispersal by altering signals that attract animal seed dispersers, i.e., by reducing palatability and altering nutrient content [152]. Conversely, the fungal infection of fruit may actually increase dispersal; for example, by increasing levels of attractive volatiles, in which case an anthropocentric view of fruit diseases may lead to the misinterpretation of a fungus’ effects on its host’s fitness in natural systems [153].
In conclusion, this work was successful in identifying a number of promising endophytes, as well as the metabolites they produce, in grapevines with a mix of European and North American ancestry growing ferally in a challenging environment. However, this work is only an initial step towards the ultimate goal: the discovery of novel growth-promoting and biocontrol agents. While a number of the isolates found in this study showed promise in their suppression of Botrytis using ex situ competition assays, the development of molecular markers followed by in situ trials is needed. In future work, inoculation trials should be performed on healthy grapevines to ensure prospective endophytes are not pathogenic and to assess their growth promotion potential relative to control plants. Finally, once confirmed as an endophyte, control and inoculated host plants should be challenged with Botrytis and other common grapevine pathogens to assess their in planta biocontrol capabilities in a controlled environment followed by field trials. Several endophytes have been shown to alter plant hormones or metabolites, and their effect on the pathogen and disease suppression in situ could be very different from the effect of endophytes on pathogen suppression in ex situ competition assays. Also, the inoculated endophyte will have an effect on the microbiome of the plants, which could indirectly affect pathogen and disease suppression, so inoculating the plants with these endophytes in the field trials will help to fully understand this interaction. Future work may include expanding sampling efforts of both the host and isolated taxa while also including fungi from the phylloplane.
5. Significance Statement
Leaf samples collected from over 80 feral grapevine sites were used to isolate a range of fungal endophytes. Endophyte identification, competition assays against a known pathogen, and metabolite screening were used as an initial step towards understanding how these microbes may be utilized as biocontrol agents in commercial plantings.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/jof10020105/s1, Figure S1. Inhibition of Botrytis cinerea growth by grape fungal endophytes in the dual-culture plate method after 7 days of incubation at 22 °C; images on the left are from above the plate, while those on the right are from below; in all the images, the endophyte is the lower microbe and Botrytis is the upper microbe. (A) Gnomoniopsis paraclavulata; (B) Diaporthe sp. En 1-1; and (C) Diaporthe sp. 26-4; (D) Botrytis cinerea control with no endophyte. Figure S2. Principal component analysis derived from ESI+ features extracted with XCMS. Diaporthe species show some degree of clustering, while En25-3 (Colletotrichum fioriniae) is distinct from all other tested strains. Figure S3. Tandem mass spectrometry (MS/MS) spectra of unique unknown extrolites detected for Colletotrichum fioriniae (En25-3). Figure S4. Tandem mass spectrometry (MS/MS) spectra of unique extrolites detected for Neophaeomoniella niveniae (En61-2). This isolate produced notable dipeptides, which were clearly defined by known amino acid immonium ions. Figure S5. Tandem mass spectrometry (MS/MS) spectra of unique unknown extrolites detected for Gnomoniopsis paraclavulata (En61-1). Figure S6. Tandem mass spectrometry (MS/MS) spectra of unique unknown extrolites detected for Diaporthe aff. gulyae (En20-4). Figure S7. Tandem mass spectrometry (MS/MS) spectra of unique unknown extrolites detected for Diaporthe sp. (En01-1). Table S1. Sequences of forward and reverse primers of seven gene loci used in PCR amplification and sequencing for identifying fungal endophytes isolated from grapevine. Table S2. Screening of extrolites produced by fungal endophyte strains to detect bioactive compounds.
Author Contributions
Conceptualization, S.A., A.H.W. and M.W.S.; Methodology, S.A., A.H.W., J.B.T., M.W.S. and J.B.R.; Formal analysis, S.A., A.H.W., J.B.T. and M.W.S.; Investigation, S.A., A.H.W. and M.W.S.; Resources, S.A., A.H.W., J.B.T. and M.W.S.; Data curation, S.A., A.H.W., J.B.T., J.B.R. and M.W.S.; Writing—original draft, S.A., A.H.W. and J.B.T.; Writing—review & editing, S.A., A.H.W., J.B.T., J.B.R. and M.W.S.; Project administration, S.A.; A.H.W. and M.W.S.; Funding acquisition, S.A., A.H.W. and M.W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Agriculture and Agri-Food Canada A-base projects (J-001742 and J-002358).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We wish to thank the property owners who allowed us to collect leaf samples from their vines. We would also like to thank Xiaotang Yang and summer students Rachel Pomerleau, Christianne Hagerman, Phylicity Byers, and Iain Thomas, who assisted with the isolation of the endophytes and in the preparation of samples for DNA sequencing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lawson, G. On the northern limit of wild grape vines. Proc. Nova Scotian Inst. Sci. 1884, 6, 1883–1886. [Google Scholar]
- Wallace, B. L’Anse Aux Meadows, Leif Eriksson’s Home in Vinland. J. North Atl. 2009, 2, 114–125. [Google Scholar] [CrossRef]
- Speck, F.G.; Dexter, R.W. Utilization of animals and plants by the Malecite Indians of New Brunswick. J. Wash. Acad. Sci. 1952, 42, 1–7. [Google Scholar]
- Murray, R.A.; Wright, B. The Tangled Vine: Winegrowing in Nova Scotia; Blue Frog: Bridgewater, NS, Canada, 2004. [Google Scholar]
- Frank, R.; Eyler, R. The Economic Impact of the Wine and Grape Industry in Canada 2015; Canadian Vintners Association, Winery and Grower Alliance of Ontario British Columbia, Wine Institute Winery Association of Nova Scotia: Halifax, NS, Canada, 2017. [Google Scholar]
- Bishop, R.; Craig, D.; MacEachern, C. Observations on the Performance of Grape Cultivars in Nova Scotia1. HortScience 1970, 5, 154–156. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Courty, P.E.; Oger, P. Plant symbionts are engineers of the plant-associated microbiome. Trends Plant Sci. 2019, 24, 905–916. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- De Vries, S.; von Dahlen, J.K.; Schnake, A.; Ginschel, S.; Schulz, B.; Rose, L.E. Broad-spectrum inhibition of Phytophthora infestans by fungal endophytes. FEMS Microbiol. Ecol. 2018, 94, fiy037. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Kohl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological Control of Plant Diseases: The European Situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Grabka, R.; d’Entremont, T.W.; Adams, S.J.; Walker, A.K.; Tanney, J.B.; Abbasi, P.A.; Ali, S. Fungal endophytes and their role in agricultural plant protection against pests and pathogens. Plants 2022, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Afolabi, O.G.; Hussain, M.; Qasim, M.; Wang, L. Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: An insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol. Res. 2018, 217, 34–50. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Latz, M.A.; Jensen, B.; Collinge, D.B.; Jørgensen, H.J. Endophytic fungi as biocontrol agents: Elucidating mechanisms in disease suppression. Plant Ecol. Divers. 2018, 11, 555–567. [Google Scholar] [CrossRef]
- Omomowo, O.I.; Babalola, O.O. Bacterial and fungal endophytes: Tiny giants with immense beneficial potential for plant growth and sustainable agricultural productivity. Microorganisms 2019, 7, 481. [Google Scholar] [CrossRef]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- Brum, M.; Araújo, W.L.; Maki, C.S.; Azevedo, J.L.D. Endophytic fungi from Vitis labrusca L.(‘Niagara Rosada’) and its potential for the biological control of Fusarium oxysporum. Genet. Mol. Res. GMR 2012, 11, 4187–4197. [Google Scholar] [CrossRef] [PubMed]
- Kernaghan, G.; Mayerhofer, M.; Griffin, A. Fungal endophytes of wild and hybrid Vitis leaves and their potential for vineyard biocontrol. Can. J. Microbiol. 2017, 63, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Ginn, F.M. Endophytic Fungi in Vaccinium macrocarpon (Cranberry) and Vaccinium angustifolium (Blueberry); The University of New Brunswick: Fredericton, NS, Canada, 2001. [Google Scholar]
- Tanney, J.B.; Di Stefano, J.; Miller, J.D.; McMullin, D.R. Natural products from the Picea foliar endophytes Niesslia endophytica sp. nov. and Strasseria geniculata. Mycol. Prog. 2023, 22, 17. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Ali, S.; Renderos, W.; Naeem, H.A.; Papadopoulos, Y. First report of Alternaria alternata causing leaf spot and blight symptoms on alfalfa in Canada. Can. J. Plant Pathol. 2018, 40, 451–455. [Google Scholar] [CrossRef]
- Ali, S.; Renderos, W.; Bevis, E.; Hebb, J.; Abbasi, P.A. Diaporthe eres causes stem cankers and death of young apple rootstocks in Canada. Can. J. Plant Pathol. 2020, 42, 218–227. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.; Shivas, R.; Burgess, T.; Decock, C.; Dreyer, L.; Granke, L.; Guest, D.; Hardy, G.S.; Hausbeck, M. Fungal Planet description sheets: 107–127. Persoonia-Mol. Phylogeny Evol. Fungi 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.; Woudenberg, J.; Crous, P. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Polizzi, G.; Crous, P.W. High species diversity in Colletotrichum associated with citrus diseases in Europe. Persoonia-Mol. Phylogeny Evol. Fungi 2017, 39, 32–50. [Google Scholar] [CrossRef]
- Kraus, C.; Damm, U.; Bien, S.; Voegele, R.; Fischer, M. New species of Phaeomoniellales from a German vineyard and their potential threat to grapevine (Vitis vinifera) health. Fungal Syst. Evol. 2020, 6, 139–155. [Google Scholar] [CrossRef]
- Damm, U.; Woudenberg, J.; Cannon, P.; Crous, P. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45. [Google Scholar]
- Gomes, R.; Glienke, C.; Videira, S.; Lombard, L.; Groenewald, J.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia-Mol. Phylogeny Evol. Fungi 2013, 31, 1–41. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. The Diaporthe sojae species complex: Phylogenetic re-assessment of pathogens associated with soybean, cucurbits and other field crops. Fungal Biol. 2015, 119, 383–407. [Google Scholar] [CrossRef]
- Du, Z.; Fan, X.-L.; Hyde, K.D.; Yang, Q.; Liang, Y.-M.; Tian, C.-M. Phylogeny and morphology reveal two new species of Diaporthe from Betula spp. in China. Phytotaxa 2016, 269, 90–102. [Google Scholar] [CrossRef]
- Gao, Y.; Su, Y.; Sun, W.; Cai, L. Diaporthe species occurring on Lithocarpus glabra in China, with descriptions of five new species. Fungal Biol. 2015, 119, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia-Mol. Phylogeny Evol. Fungi 2018, 40, 135–153. [Google Scholar] [CrossRef]
- Thompson, S.; Tan, Y.P.; Shivas, R.G.; Neate, S.M.; Morin, L.; Bissett, A.; Aitken, E. Green and brown bridges between weeds and crops reveal novel Diaporthe species in Australia. Persoonia-Mol. Phylogeny Evol. Fungi 2015, 35, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jiang, N.; Tian, C.-M. Three new Diaporthe species from Shaanxi province, China. MycoKeys 2020, 67, 1. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Correia, V.G.; Phillips, A.J. Primers for mating-type diagnosis in Diaporthe and Phomopsis: Their use in teleomorph induction in vitro and biological species definition. Fungal Biol. 2010, 114, 255–270. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; Crous, P.W.; McKenzie, E.H.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Dissanayake, A.J.; Li, X.; Liu, M.; Wanasinghe, D.N.; Xu, J.; Zhao, W.; Zhang, W.; Zhou, Y.; Hyde, K.D. High genetic diversity and species complexity of Diaporthe associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef]
- Thompson, S.; Tan, Y.; Young, A.; Neate, S.; Aitken, E.; Shivas, R. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia-Mol. Phylogeny Evol. Fungi 2011, 27, 80–89. [Google Scholar] [CrossRef]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, A.; Phillips, A.; Hyde, K.; Yan, J.; Li, X. The current status of species in Diaporthe. Mycosphere 2017, 8, 1106–1156. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Gareth Jones, E.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Zhou, H.; Hou, C.-L. Three new species of Diaporthe from China based on morphological characters and DNA sequence data analyses. Phytotaxa 2019, 422, 157–174. [Google Scholar] [CrossRef]
- Feng, X.; Chen, J.; Wang, G.; Cao, T.; Zheng, Y.; Zhang, C. Diaporthe sinensis, a new fungus from Amaranthus sp. in China. Phytotaxa 2019, 425, 259–268. [Google Scholar] [CrossRef]
- Hilário, S.; Amaral, I.A.; Gonçalves, M.F.; Lopes, A.; Santos, L.; Alves, A. Diaporthe species associated with twig blight and dieback of Vaccinium corymbosum in Portugal, with description of four new species. Mycologia 2020, 112, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Luangsa-Ard, J.; Wingfield, M.; Carnegie, A.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.; Baseia, I.; Cano-Lira, J. Fungal Planet description sheets: 785–867. Persoonia-Mol. Phylogeny Evol. Fungi 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Sogonov, M.V.; Castlebury, L.; Rossman, A.; Mejía, L.C.; White, J. Leaf-inhabiting genera of the Gnomoniaceae, Diaporthales. Stud. Mycol. 2008, 62, 1–77. [Google Scholar] [CrossRef] [PubMed]
- Mejía, L.C.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. Phylogenetic placement and taxonomic review of the genus Cryptosporella and its synonyms Ophiovalsa and Winterella (Gnomoniaceae, Diaporthales). Mycol. Res. 2008, 112, 23–35. [Google Scholar] [CrossRef]
- Sánchez-Ballesteros, J.; González, V.; Salazar, O.; Acero, J.; Portal, M.A.; Julián, M.; Rubio, V.; Bills, G.F.; Polishook, J.D.; Platas, G. Phylogenetic study of Hypoxylon and related genera based on ribosomal ITS sequences. Mycologia 2000, 92, 964–977. [Google Scholar] [CrossRef]
- Newcombe, G.; Shipunov, A.; Eigenbrode, S.; Raghavendra, A.K.; Ding, H.; Anderson, C.L.; Menjivar, R.; Crawford, M.; Schwarzländer, M. Endophytes influence protection and growth of an invasive plant. Commun. Integr. Biol. 2009, 2, 29–31. [Google Scholar] [CrossRef]
- Joan, T.; Chapela, I.H.; Bruns, T.; Taylor, J. The Phyloecology of Hypaxylan sensu lata. Master’s Thesis, University of California, Berkeley, CA, USA, 2001. [Google Scholar]
- Fournier, J.; Lechat, C.; Ribes Ripoll, M. Record of Nemania aureolutea (Xylariaceae) from the southernmost region of Spain. Ascomycete.org 2020, 12, 221–226. [Google Scholar]
- U’Ren, J.M.; Arnold, A.E. Diversity, taxonomic composition, and functional aspects of fungal communities in living, senesced, and fallen leaves at five sites across North America. PeerJ 2016, 4, e2768. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Læssøe, T.; Chlebicki, A.; Lechat, C.; Flessa, F.; Rambold, G.; Peršoh, D. Chemotaxonomic and phylogenetic studies of Thamnomyces (Xylariaceae). Mycoscience 2010, 51, 189–207. [Google Scholar] [CrossRef]
- Lagarde, A.; Millot, M.; Pinon, A.; Liagre, B.; Girardot, M.; Imbert, C.; Ouk, T.-S.; Jargeat, P.; Mambu, L. Antiproliferative and antibiofilm potentials of endolichenic fungi associated with the lichen Nephroma laevigatum. J. Appl. Microbiol. 2019, 126, 1044–1058. [Google Scholar] [CrossRef]
- Ding, W.; Chen, H.-W.; Wang, M.; Wan, M.; Hu, J.-F.; Li, J. Three new eremophilane sesquiterpenes and one new related derivative from Nemania sp. HDF-Br-5, an endophytic fungus of the endangered conifer Pseudotsuga gaussenii Flous. Phytochem. Lett. 2022, 49, 5–11. [Google Scholar] [CrossRef]
- Crous, P.; Summerell, B.; Shivas, R.; Romberg, M.; Mel’nik, V.; Verkley, G.; Groenewald, J. Fungal Planet description sheets: 92–106. Persoonia Mol. Phylogeny Evol. Fungi 2011, 27, 130–162. [Google Scholar] [CrossRef]
- Spies, C.; Mostert, L.; Carlucci, A.; Moyo, P.; Van Jaarsveld, W.; du Plessis, I.; Van Dyk, M.; Halleen, F. Dieback and decline pathogens of olive trees in South Africa. Persoonia-Mol. Phylogeny Evol. Fungi 2020, 45, 196–220. [Google Scholar] [CrossRef]
- Damm, U.; Fourie, P.; Crous, P.W. Coniochaeta (Lecythophora), Collophora gen. nov. and Phaeomoniella species associated with wood necroses of Prunus trees. Persoonia-Mol. Phylogeny Evol. Fungi 2010, 24, 60–80. [Google Scholar] [CrossRef]
- Ferreira, A.B.M.; Leite, L.G.; Hernandes, J.L.; Harakava, R.; Padovani, C.R.; Bueno, C.J. Colonization of vines by Petri disease fungi, susceptibility of rootstocks to Phaeomoniella chlamydospora and their disinfection. Arq. Inst. Biológico 2018, 85, e0882017. [Google Scholar] [CrossRef]
- Videira, S.; Groenewald, J.Z.; Braun, U.; Shin, H.; Crous, P.W. All that glitters is not Ramularia. Stud. Mycol. 2016, 83, 49–163. [Google Scholar] [CrossRef]
- Videira, S.; Groenewald, J.Z.; Kolecka, A.; van Haren, L.; Boekhout, T.; Crous, P.W. Elucidating the Ramularia eucalypti species complex. Persoonia-Mol. Phylogeny Evol. Fungi 2015, 34, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Verkley, G.; Quaedvlieg, W.; Shin, H.-D.; Crous, P. A new approach to species delimitation in Septoria. Stud. Mycol. 2013, 75, 213–305. [Google Scholar] [CrossRef] [PubMed]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.; Summerell, B.; Carnegie, A.; Burgess, T.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia-Mol. Phylogeny Evol. Fungi 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Ismail, S.I.; Batzer, J.C.; Harrington, T.C.; Crous, P.W.; Lavrov, D.V.; Li, H.; Gleason, M.L. Ancestral state reconstruction infers phytopathogenic origins of sooty blotch and flyspeck fungi on apple. Mycologia 2016, 108, 292–302. [Google Scholar] [CrossRef]
- Crous, P.; Braun, U.; Groenewald, J. Mycosphaerella is polyphyletic. Stud. Mycol. 2007, 58, 1–32. [Google Scholar] [CrossRef]
- Ghildiyal, A.; Pandey, A. Isolation of cold tolerant antifungal strains of Trichoderma sp. from glacial sites of Indian Himalayan region. Res. J. Microbiol. 2008, 3, 559–564. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Laatsch, H. AntiBase: The Natural Compound Identifier; Wiley-Vch Weinheim: Weinheim, Germany, 2017. [Google Scholar]
- Badalyan, S.M.; Garibyan, N.; Innocenti, G. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Phytopathol. Mediterr. 2002, 41, 220–225. [Google Scholar]
- Flores, A.C.; Pamphile, J.A.; Sarragiotto, M.H.; Clemente, E. Production of 3-nitropropionic acid by endophytic fungus Phomopsis longicolla isolated from Trichilia elegans A. JUSS ssp. elegans and evaluation of biological activity. World J. Microbiol. Biotechnol. 2013, 29, 923–932. [Google Scholar] [CrossRef]
- Specian, V.; Sarragiotto, M.H.; Pamphile, J.A.; Clemente, E. Chemical characterization of bioactive compounds from the endophytic fungus Diaporthe helianthi isolated from Luehea divaricata. Braz. J. Microbiol. 2012, 43, 1174–1182. [Google Scholar] [CrossRef]
- Carvalho, C.d.; Ferreira-D’Silva, A.; Wedge, D.; Cantrell, C.; Rosa, L. Antifungal activities of cytochalasins produced by Diaporthe miriciae, an endophytic fungus associated with tropical medicinal plants. Can. J. Microbiol. 2018, 64, 835–843. [Google Scholar] [CrossRef]
- Hedrick, U.P.; Booth, N.O.; Maxwell, J.D.; Taylor, O.M.; Wellington, R. The Grapes of New York; State of New York Department of Agriculture: Albany, NY, USA, 1908. [Google Scholar]
- Sessa, L.; Abreo, E.; Lupo, S. Diversity of fungal latent pathogens and true endophytes associated with fruit trees in Uruguay. J. Phytopathol. 2018, 166, 633–647. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Abramczyk, B.; Marzec-Grządziel, A.; Grządziel, J.; Król, E.; Gałązka, A.; Oleszek, W. Biocontrol Potential and Catabolic Profile of Endophytic Diaporthe eres Strain 1420S from Prunus domestica L. in Poland—A Preliminary Study. Agronomy 2022, 12, 165. [Google Scholar] [CrossRef]
- Polonio, J.; Almeida, T.; Garcia, A.; Mariucci, G.; Azevedo, J.; Rhoden, S.; Pamphile, J. Biotechnological prospecting of foliar endophytic fungi of guaco (Mikania glomerata Spreng.) with antibacterial and antagonistic activity against phytopathogens. Genet. Mol. Res. 2015, 14, 7297–7309. [Google Scholar] [CrossRef]
- Da Silva Ribeiro, A.; Polonio, J.C.; Costa, A.T.; Dos Santos, C.M.; Rhoden, S.A.; Azevedo, J.L.; Pamphile, J.A. Bioprospection of culturable endophytic fungi associated with the ornamental plant Pachystachys lutea. Curr. Microbiol. 2018, 75, 588–596. [Google Scholar] [CrossRef]
- Verma, P.; Hiremani, N.S.; Gawande, S.P.; Sain, S.K.; Nagrale, D.T.; Narkhedkar, N.G.; Prasad, Y. Modulation of plant growth and antioxidative defense system through endophyte biopriming in cotton (Gossypium spp.) and non-host crops. Heliyon 2022, 8, e09487. [Google Scholar] [CrossRef]
- Webber, J.; Gibbs, J. Colonization of elm bark by Phomopsis oblonga. Trans. Br. Mycol. Soc. 1984, 82, 348–352. [Google Scholar] [CrossRef]
- Uecker, F. A World List of Phomopsis Names with Notes on Nomenclature,-Morphology and Biology; Cramer Publishers: Berlin, Germany, 1988. [Google Scholar]
- Hilário, S.; Gonçalves, M.F.; Alves, A. Using genealogical concordance and coalescent-based species delimitation to assess species boundaries in the Diaporthe eres complex. J. Fungi 2021, 7, 507. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, A.J.; Liu, M.; Zhang, W.; Chen, Z.; Udayanga, D.; Chukeatirote, E.; Li, X.; Yan, J.; Hyde, K.D. Morphological and molecular characterisation of Diaporthe species associated with grapevine trunk disease in China. Fungal Biol. 2015, 119, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Reveglia, P.; Pacetti, A.; Masi, M.; Cimmino, A.; Carella, G.; Marchi, G.; Mugnai, L.; Evidente, A. Phytotoxic metabolites produced by Diaporthe eres involved in cane blight of grapevine in Italy. Nat. Prod. Res. 2021, 35, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Makris, G.; Solonos, S.; Christodoulou, M.; Kanetis, L.I. First report of Diaporthe foeniculina associated with grapevine trunk diseases on Vitis vinifera in Cyprus. Plant Dis. 2022, 106, 1294. [Google Scholar] [CrossRef] [PubMed]
- Pine, T. Etiology of the dead-arm disease of Grapevines. Phytopathology 1958, 48, 192–197. [Google Scholar]
- Nita, M.; Ellis, M.; Madden, L. Variation in disease incidence of Phomopsis cane and leaf spot of grape in commercial vineyards in Ohio. Plant Dis. 2008, 92, 1053–1061. [Google Scholar] [CrossRef]
- Cinelli, T.; Mondello, V.; Marchi, G.; Burruano, S.; Alves, A.; Mugnai, L. First report of Diaporthe eres associated with cane blight of grapevine (Vitis vinifera) in Italy. Plant Dis. 2016, 100, 532. [Google Scholar] [CrossRef]
- Lesuthu, P.; Mostert, L.; Spies, C.F.; Moyo, P.; Regnier, T.; Halleen, F. Diaporthe nebulae sp. nov. and first report of D. cynaroidis, D. novem, and D. serafiniae on grapevines in South Africa. Plant Dis. 2019, 103, 808–817. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Cimmino, A.; Evidente, A. Phytotoxic metabolites by nine species of Botryosphaeriaceae involved in grapevine dieback in Australia and identification of those produced by Diplodia mutila, Diplodia seriata, Neofusicoccum australe and Neofusicoccum luteum. Nat. Prod. Res. 2019, 33, 2223–2229. [Google Scholar] [CrossRef]
- Barba, P.; Lillis, J.; Luce, R.S.; Travadon, R.; Osier, M.; Baumgartner, K.; Wilcox, W.F.; Reisch, B.I.; Cadle-Davidson, L. Two dominant loci determine resistance to Phomopsis cane lesions in F 1 families of hybrid grapevines. Theor. Appl. Genet. 2018, 131, 1173–1189. [Google Scholar] [CrossRef] [PubMed]
- Pinna, C.; Linaldeddu, B.T.; Deiana, V.; Maddau, L.; Montecchio, L.; Lentini, A. Plant pathogenic fungi associated with Coraebus florentinus (Coleoptera: Buprestidae) attacks in declining oak forests. Forests 2019, 10, 488. [Google Scholar] [CrossRef]
- Fort, T.; Pauvert, C.; Zanne, A.E.; Ovaskainen, O.; Caignard, T.; Barret, M.; Compant, S.; Hampe, A.; Delzon, S.; Vacher, C. Maternal effects shape the seed mycobiome in Quercus petraea. New Phytol. 2021, 230, 1594–1608. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.L.; Peter, K.A. Quantification of Colletotrichum fioriniae in orchards and deciduous forests indicates it is primarily a leaf endophyte. Phytopathology® 2021, 111, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Jankowiak, R.; Stępniewska, H.; Bilański, P.; Taerum, S.J. Fungi as potential factors limiting natural regeneration of pedunculate oak (Quercus robur) in mixed-species forest stands in Poland. Plant Pathol. 2022, 71, 805–817. [Google Scholar] [CrossRef]
- Costa, D.; Tavares, R.M.; Baptista, P.; Lino-Neto, T. Cork oak endophytic fungi as potential biocontrol agents against Biscogniauxia mediterranea and Diplodia corticola. J. Fungi 2020, 6, 287. [Google Scholar] [CrossRef] [PubMed]
- Tosi, L.; Beccari, G.; Rondoni, G.; Covarelli, L.; Ricci, C. Natural occurrence of Fusarium proliferatum on chestnut in Italy and its potential entomopathogenicity against the Asian chestnut gall wasp Dryocosmus kuriphilus. J. Pest Sci. 2015, 88, 369–381. [Google Scholar] [CrossRef]
- Patanita, M.; Albuquerque, A.; Campos, M.D.; Materatski, P.; Varanda, C.M.; Ribeiro, J.A.; Félix, M.d.R. Metagenomic assessment unravels fungal microbiota associated to grapevine trunk diseases. Horticulturae 2022, 8, 288. [Google Scholar] [CrossRef]
- Van Dyk, M.; Spies, C.F.; Mostert, L.; van der Rijst, M.; du Plessis, I.L.; Moyo, P.; van Jaarsveld, W.J.; Halleen, F. Pathogenicity testing of fungal isolates associated with olive trunk diseases in South Africa. Plant Dis. 2021, 105, 4060–4073. [Google Scholar] [CrossRef] [PubMed]
- Bushman, T.J.; Cunneely, Q.; Ciesla, L. Extraction, isolation, and Biological activity of natural cyclic dipeptides. Stud. Nat. Prod. Chem. 2023, 78, 75–99. [Google Scholar]
- Ivic, D.; Voncina, D.; Sever, Z.; Simon, S.; Pejic, I. Identification of Colletotrichum species causing bitter rot of apple and pear in Croatia. J. Phytopathol. 2013, 161, 284–286. [Google Scholar] [CrossRef]
- Munir, M.; Amsden, B.; Dixon, E.; Vaillancourt, L.; Gauthier, N.W. Characterization of Colletotrichum species causing bitter rot of apple in Kentucky orchards. Plant Dis. 2016, 100, 2194–2203. [Google Scholar] [CrossRef]
- Kim, C.H.; Hassan, O.; Chang, T. Diversity, pathogenicity, and fungicide sensitivity of Colletotrichum species associated with apple anthracnose in South Korea. Plant Dis. 2020, 104, 2866–2874. [Google Scholar] [CrossRef]
- Schoeneberg, A.; Hu, M.-J. First report of Anthracnose fruit rot caused by Colletotrichum fioriniae on red raspberry (Rubus idaeus) in the mid-atlantic region of the United States. Plant Dis. 2020, 104, 1855. [Google Scholar] [CrossRef]
- Luongo, L.; Galli, M.; Garaguso, I.; Petrucci, M.; Vitale, S. First Report of Colletotrichum fioriniae and C. nymphaeae as Causal Agents of Anthracnose on Walnut in Italy. Plant Dis. 2022, 106, 327. [Google Scholar] [CrossRef]
- Xu, S.-J.; Aktaruzzaman, M.; Kim, B.S.; Kim, J.; Shin, H.-D. First report of anthracnose caused by Colletotrichum fioriniae on eggplant fruits in Korea. Plant Dis. 2018, 102, 2642. [Google Scholar] [CrossRef]
- Kasson, M.; Pollok, J.; Benhase, E.; Jelesko, J. First report of seedling blight of eastern poison ivy (Toxicodendron radicans) by Colletotrichum fioriniae in Virginia. Plant Dis. 2014, 98, 995. [Google Scholar] [CrossRef]
- Garibaldi, A.; Bertetti, D.; Matić, S.; Luongo, I.; Guarnaccia, V.; Gullino, M. First report of leaf blight caused by Colletotrichum fioriniae on Mahonia aquifolium in Italy. Plant Dis. 2020, 104, 983. [Google Scholar] [CrossRef]
- Redman, R.S.; Dunigan, D.D.; Rodriguez, R.J. Fungal symbiosis from mutualism to parasitism: Who controls the outcome, host or invader? New Phytol. 2001, 151, 705–716. [Google Scholar] [CrossRef]
- Gonzaga, L.; Costa, L.; Santos, T.; Araújo, E.; Queiroz, M. Endophytic fungi from the genus Colletotrichum are abundant in the Phaseolus vulgaris and have high genetic diversity. J. Appl. Microbiol. 2015, 118, 485–496. [Google Scholar] [CrossRef]
- Nigar, Q.; Cadle-Davidson, L.; Gadoury, D.M.; Hassan, M.u. First report of Colletotrichum fioriniae causing grapevine anthracnose in New York. Plant Dis. 2023, 107, 223. [Google Scholar] [CrossRef]
- Cosseboom, S.D.; Hu, M. Predicting ripe rot of grape, caused by Colletotrichum fioriniae, with leaf wetness, temperature, and the crop growth stage. PhytoFrontiers™ 2023, 3, 303–313. [Google Scholar] [CrossRef]
- Marcelino, J.; Giordano, R.; Gouli, S.; Gouli, V.; Parker, B.L.; Skinner, M.; TeBeest, D.; Cesnik, R. Colletotrichum acutatum var. fioriniae (teleomorph: Glomerella acutata var. fioriniae var. nov.) infection of a scale insect. Mycologia 2008, 100, 353–374. [Google Scholar] [CrossRef]
- Marcelino, J.A.; Gouli, S.; Parker, B.L.; Skinner, M.; Schwarzberg, L.; Giordano, R. Host plant associations of an entomopathogenic variety of the fungus, Colletotrichum acutatum, recovered from the elongate hemlock scale, Fiorinia externa. J. Insect Sci. 2009, 9, 25. [Google Scholar] [CrossRef]
- González, J.B.; Lambert, C.A.; Foley, A.M.; Hajek, A.E. First report of Colletotrichum fioriniae infections in brown marmorated stink bugs, Halyomorpha halys. J. Invertebr. Pathol. 2023, 199, 107939. [Google Scholar] [CrossRef]
- Ibrahim, A.; Tanney, J.B.; Fei, F.; Seifert, K.A.; Cutler, G.C.; Capretta, A.; Miller, J.D.; Sumarah, M.W. Metabolomic-guided discovery of cyclic nonribosomal peptides from Xylaria ellisii sp. nov., a leaf and stem endophyte of Vaccinium angustifolium. Sci. Rep. 2020, 10, 4599. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Sørensen, D.; Jenkins, H.A.; Ejim, L.; Capretta, A.; Sumarah, M.W. Epoxynemanione A, nemanifuranones A–F, and nemanilactones A–C, from Nemania serpens, an endophytic fungus isolated from Riesling grapevines. Phytochemistry 2017, 140, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.P.; Araujo, A.R.; Batista, J.M., Jr.; Cardoso, C.L.; Seidl, C.; Vilela, A.F.; Domingos, H.V.; Costa-Lotufo, L.V.; Andersen, R.J.; Silva, D.H. Botryane terpenoids produced by Nemania bipapillata, an endophytic fungus isolated from red alga Asparagopsis taxiformis-Falkenbergia stage. Sci. Rep. 2019, 9, 12318. [Google Scholar] [CrossRef]
- Tibpromma, S.; Zhang, L.; Karunarathna, S.C.; Du, T.-Y.; Phukhamsakda, C.; Rachakunta, M.; Suwannarach, N.; Xu, J.; Mortimer, P.E.; Wang, Y.-H. Volatile constituents of endophytic fungi isolated from Aquilaria sinensis with descriptions of two new species of Nemania. Life 2021, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Bilański, P.; Kowalski, T. Fungal endophytes in Fraxinus excelsior petioles and their in vitro antagonistic potential against the ash dieback pathogen Hymenoscyphus fraxineus. Microbiol. Res. 2022, 257, 126961. [Google Scholar] [CrossRef] [PubMed]
- Aleynova, O.A.; Nityagovsky, N.N.; Kiselev, K.V. Biodiversity of endophytic bacteria and fungi of wild grapes Vitis amurensis Rupr. In Proceedings of the BIO Web of Conferences, Virtual, 23–24 November 2021; p. 05001. [Google Scholar]
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. III. Cryptosphaeria, Eutypa and Eutypella (Diatrypaceae). Fungal Divers. 2006, 22, 243–254. [Google Scholar]
- Zíbarová, L.; Kout, J. Xylariaceous pyrenomycetes from Bohemia: Species of Biscogniauxia and Hypoxylon new to the Czech Republic, and notes on other rare species. Czech Mycol. 2017, 69, 77–108. [Google Scholar] [CrossRef]
- Herath, P.; Beauseigle, S.; Dhillon, B.; Ojeda, D.I.; Bilodeau, G.; Isabel, N.; Gros-Louis, M.-C.; Kope, H.; Zeglen, S.; Hamelin, R.C. Anthropogenic signature in the incidence and distribution of an emerging pathogen of poplars. Biol. Invasions 2016, 18, 1147–1161. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.; Shin, H.-D.; Barreto, R.; Alfenas, A.; Swart, W.; Groenewald, J.; Crous, P.W. Sizing up septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef]
- Ali, S.; Hildebrand, P.D.; Renderos, W.E.; Abbasi, P.A. Identification and Characterization of Sphaerulina vaccinii sp. nov. as the Cause of Leaf Spot and Stem Canker in Lowbush Blueberry and Its Epidemiology. Phytopathology 2021, 111, 1560–1570. [Google Scholar] [CrossRef]
- Gumbrewicz, R.; Calderwood, L. Comparison of wood mulch particle sizes for wild blueberry management in a changing climate. Int. J. Fruit Sci. 2022, 22, 551–567. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Hildebrand, P.D.; Ali, S.; Moreau, D.L.; Renderos, W.E. Effect of RH, Temperature, Light, and Plant Age on Infection of Lowbush Blueberry by Sphaerulina vaccinii. Plant Dis. 2022, 106, 297–303. [Google Scholar] [CrossRef]
- Thach, T.; Munk, L.; Hansen, A.L.; Jørgensen, L.N. Disease variation and chemical control of Ramularia leaf spot in sugar beet. Crop Prot. 2013, 51, 68–76. [Google Scholar] [CrossRef]
- McGrann, G.R.; Havis, N.D. Ramularia leaf spot: A newly important threat to barley production. Outlooks Pest Manag. 2017, 28, 65–69. [Google Scholar] [CrossRef]
- Ding, C.-H.; Wang, Q.-B.; Guo, S.; Wang, Z.-Y. The improvement of bioactive secondary metabolites accumulation in Rumex gmelini Turcz through co-culture with endophytic fungi. Braz. J. Microbiol. 2018, 49, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Gasparoto, M.; Lourenço, S.; Tanaka, F.A.O.; Spósito, M.B.; Marchini, L.C.; Silva Junior, G.; Amorim, L. Honeybees can spread Colletotrichum acutatum and C. gloeosporioides among citrus plants. Plant Pathol. 2017, 66, 777–782. [Google Scholar] [CrossRef]
- González-Domínguez, E.; Caffi, T.; Languasco, L.; Latinovic, N.; Latinovic, J.; Rossi, V. Dynamics of Diaporthe ampelina conidia released from grape canes that overwintered in the vineyard. Plant Dis. 2021, 105, 3092–3100. [Google Scholar] [CrossRef]
- Nelson, A.; Vandegrift, R.; Carroll, G.C.; Roy, B.A. Double lives: Transfer of fungal endophytes from leaves to woody substrates. PeerJ 2020, 8, e9341. [Google Scholar] [CrossRef]
- McGrann, G.R.; Andongabo, A.; Sjökvist, E.; Trivedi, U.; Dussart, F.; Kaczmarek, M.; Mackenzie, A.; Fountaine, J.M.; Taylor, J.M.; Paterson, L.J. The genome of the emerging barley pathogen Ramularia collo-cygni. BMC Genom. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Batzer, J.C.; Mueller, D.S. Soybean fungal endophytes Alternaria and Diaporthe spp. are differentially impacted by fungicide application. Plant Dis. 2020, 104, 52–59. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Nelson, J.M.; Hauser, D.A.; Hinson, R.; Shaw, A.J. A novel experimental system using the liverwort Marchantia polymorpha and its fungal endophytes reveals diverse and context-dependent effects. New Phytol. 2018, 218, 1217–1232. [Google Scholar] [CrossRef]
- Sadoral, J.P.; Cumagun, C.J.R. Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora. Microbiol. Res. 2021, 12, 528–538. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.H. Why fruits rot, seeds mold, and meat spoils. Am. Nat. 1977, 111, 691–713. [Google Scholar] [CrossRef]
- Peris, J.E.; Rodríguez, A.; Peña, L.; Fedriani, J.M. Fungal infestation boosts fruit aroma and fruit removal by mammals and birds. Sci. Rep. 2017, 7, 5646. [Google Scholar] [CrossRef]
Figure 1.
IQ-TREE maximum likelihood consensus tree inferred from the combined TUB2, TEF1-α, and ITS sequence alignment for Diaporthe. Strains representing species formally in the D. eres species complex are now synonymized as D. eres. Strains originating from this study are highlighted by an orange rectangular box. Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0), and a hyphen (-) indicates support lower than the significant values listed for each branch test. The tree is rooted to Diaporthella corylina (CBS 121124). The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.
Figure 1.
IQ-TREE maximum likelihood consensus tree inferred from the combined TUB2, TEF1-α, and ITS sequence alignment for Diaporthe. Strains representing species formally in the D. eres species complex are now synonymized as D. eres. Strains originating from this study are highlighted by an orange rectangular box. Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0), and a hyphen (-) indicates support lower than the significant values listed for each branch test. The tree is rooted to Diaporthella corylina (CBS 121124). The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.

Figure 2.
Morphology of Diaporthe spp. (A–D) D. eres (En25-6) on WA with Ilex aquifolium leaf. (A) Conidiomata; (B) conidioma with exuded droplet containing alpha conidia (on I. aquifolium leaf); (C) conidiogenous cells and alpha conidia; (D) alpha conidia; (E–H) D. aff. gulyae (En20-4); (E) conidiomata with conidial droplets; (F) conidiogenous cells; (G) alpha conidia; (H) alpha and beta conidia; (I–N) Diaporthe sp. (En01-1) on WA with Rubus armeniacus leaf; (I) perithecia on leaf surface; (J) whole perithecium; (K) ascospores; (L) apex of perithecial neck; (M) alpha conidia (top row) and gamma conidia (bottom row); (N) conidiogenous cells. Scale bars: (A,E,I) = 1 mm; (B) = 500 μm; (C,D,F–H,K–N) = 10 μm; (J) = 100 μm.
Figure 2.
Morphology of Diaporthe spp. (A–D) D. eres (En25-6) on WA with Ilex aquifolium leaf. (A) Conidiomata; (B) conidioma with exuded droplet containing alpha conidia (on I. aquifolium leaf); (C) conidiogenous cells and alpha conidia; (D) alpha conidia; (E–H) D. aff. gulyae (En20-4); (E) conidiomata with conidial droplets; (F) conidiogenous cells; (G) alpha conidia; (H) alpha and beta conidia; (I–N) Diaporthe sp. (En01-1) on WA with Rubus armeniacus leaf; (I) perithecia on leaf surface; (J) whole perithecium; (K) ascospores; (L) apex of perithecial neck; (M) alpha conidia (top row) and gamma conidia (bottom row); (N) conidiogenous cells. Scale bars: (A,E,I) = 1 mm; (B) = 500 μm; (C,D,F–H,K–N) = 10 μm; (J) = 100 μm.

Figure 3.
Morphology of Neophaeomoniella niveniae, Colletotrichum fioriniae, and Gnomoniopsis paraclavulata. (A–F) N. niveniae (En61-2); (A) two-week-old colony on V8A. (B,C) intercalary conidiogenous cells and conidia on oatmeal agar (OA); (D) slimy mass of conidia on hypha (OA); (E) conidiomata (OA); (F) conidia (OA); (G,H) C. fioriniae (En25-3) on water agar with Rubus armeniacus leaf; (G) conidiomata with salmon-colored masses of conidia on leaf veins; (H) conidia; (I–K) G. paraclavulata (En61-1) on WA with Rubus armeniacus leaf; (I) conidiomata with orange masses of conidia; (J) conidiogenous cells and conidia; (K) conidia. Scale bars: (A) = 1 mm; (B–D,F,H,J,K) = 10 μm; (E) = 100 μm. (G,I) = 1 mm.
Figure 3.
Morphology of Neophaeomoniella niveniae, Colletotrichum fioriniae, and Gnomoniopsis paraclavulata. (A–F) N. niveniae (En61-2); (A) two-week-old colony on V8A. (B,C) intercalary conidiogenous cells and conidia on oatmeal agar (OA); (D) slimy mass of conidia on hypha (OA); (E) conidiomata (OA); (F) conidia (OA); (G,H) C. fioriniae (En25-3) on water agar with Rubus armeniacus leaf; (G) conidiomata with salmon-colored masses of conidia on leaf veins; (H) conidia; (I–K) G. paraclavulata (En61-1) on WA with Rubus armeniacus leaf; (I) conidiomata with orange masses of conidia; (J) conidiogenous cells and conidia; (K) conidia. Scale bars: (A) = 1 mm; (B–D,F,H,J,K) = 10 μm; (E) = 100 μm. (G,I) = 1 mm.

Figure 4.
IQ-TREE maximum-likelihood consensus trees, with the strains identified in the study highlighted by orange rectangular boxes. (A) IQ-TREE maximum likelihood consensus tree inferred from the TEF1-α sequence alignment for Sphaerulina, rooted to Cercospora chrysanthemoides (CPC 20529), and strain En26-1 originating from this study is highlighted by an orange rectangular box. The tree is rooted to Cercospora chrysanthemoides (CPC 20529). (B) IQ-TREE maximum likelihood consensus tree inferred from the combined ITS and TEF1-α sequence alignment for Neophaeomoniella. The strain (En61-2) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Phaeomoniella chlamydospora (IBVD01). (C) IQ-TREE maximum likelihood consensus tree inferred from the combined ITS and RPB2 sequence alignment for Gnomoniopsis. The strain (En61-1) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Melanconis alni (AR 3748). (D) IQ-TREE maximum likelihood consensus tree inferred from the combined ACT, ITS, and RPB2 sequence alignment for Ramularia. The strain (En60) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Zymoseptoria halophila (CBS 128854). Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0) and a hyphen (-) indicates support lower than the significant values listed for each branch test. The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.
Figure 4.
IQ-TREE maximum-likelihood consensus trees, with the strains identified in the study highlighted by orange rectangular boxes. (A) IQ-TREE maximum likelihood consensus tree inferred from the TEF1-α sequence alignment for Sphaerulina, rooted to Cercospora chrysanthemoides (CPC 20529), and strain En26-1 originating from this study is highlighted by an orange rectangular box. The tree is rooted to Cercospora chrysanthemoides (CPC 20529). (B) IQ-TREE maximum likelihood consensus tree inferred from the combined ITS and TEF1-α sequence alignment for Neophaeomoniella. The strain (En61-2) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Phaeomoniella chlamydospora (IBVD01). (C) IQ-TREE maximum likelihood consensus tree inferred from the combined ITS and RPB2 sequence alignment for Gnomoniopsis. The strain (En61-1) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Melanconis alni (AR 3748). (D) IQ-TREE maximum likelihood consensus tree inferred from the combined ACT, ITS, and RPB2 sequence alignment for Ramularia. The strain (En60) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Zymoseptoria halophila (CBS 128854). Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0) and a hyphen (-) indicates support lower than the significant values listed for each branch test. The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.

Figure 5.
IQ-TREE maximum likelihood consensus trees (A) IQ-TREE maximum likelihood consensus tree inferred from the CHS-1 sequence alignment for Colletotrichum. The strain (En25-3) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Colletotrichum spaethianum (CBS 167.49). (B) IQ-TREE maximum likelihood consensus tree inferred from the ITS sequence alignment for Nemania. The strain (En25-1) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Xylaria longipes (CBS 347.37). Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0), and a hyphen (-) indicates support lower than the significant values listed for each branch test. The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.
Figure 5.
IQ-TREE maximum likelihood consensus trees (A) IQ-TREE maximum likelihood consensus tree inferred from the CHS-1 sequence alignment for Colletotrichum. The strain (En25-3) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Colletotrichum spaethianum (CBS 167.49). (B) IQ-TREE maximum likelihood consensus tree inferred from the ITS sequence alignment for Nemania. The strain (En25-1) originating from this study is highlighted by an orange rectangular box. The tree is rooted to Xylaria longipes (CBS 347.37). Support values at the nodes correspond to SH-aLRT (≥80%), aBayes (≥0.95), and ultrafast bootstrap (≥95%) support values; an asterisk (*) indicates full support (100% or 1.0), and a hyphen (-) indicates support lower than the significant values listed for each branch test. The scale bar shows the expected number of nucleotide substitutions per site. Ex-type and ex-epitype strains are indicated by the symbol T.

Figure 6.
Morphology of Nemania aureolutea, Ramularia sp., and Sphaerulina amelanchier. (A) Nemania aureolutea (En25-1) on PDA in the first two plates show the front and reverse of two-week-old colonies and, the last two plates show the front and reverse of four-week-old colonies. (B–F) Ramularia sp. (En60); (B) two-week-old colony on V8A; (C) conidiomata on WA with Ilex aquifolium leaf; (D–F) conidiophores, conidiogenous cells, and conidia; (G) Sphaerulina amelanchier En26-1 on MEA; (H) conidia. Scale bars: (A,B) = 1 cm; (C) = 500 μm; (D–F,H) = 10 μm.
Figure 6.
Morphology of Nemania aureolutea, Ramularia sp., and Sphaerulina amelanchier. (A) Nemania aureolutea (En25-1) on PDA in the first two plates show the front and reverse of two-week-old colonies and, the last two plates show the front and reverse of four-week-old colonies. (B–F) Ramularia sp. (En60); (B) two-week-old colony on V8A; (C) conidiomata on WA with Ilex aquifolium leaf; (D–F) conidiophores, conidiogenous cells, and conidia; (G) Sphaerulina amelanchier En26-1 on MEA; (H) conidia. Scale bars: (A,B) = 1 cm; (C) = 500 μm; (D–F,H) = 10 μm.

Figure 7.
A box and whisker plot showing the percent growth inhibition index (GII) of Botrytis cinerea by itself (red), which acted as a control, and by different grape fungal endophytes isolated from ancient wine grapes. The percent GII was calculated as (HP-VP)/HP × 100, where HP is the mean of horizontal growth (mean radius) of the pathogen and VP is the vertical growth of the pathogen (mean radius) towards the endophyte. Values with shared letter groupings are not significantly different, as measured via a post hoc Tukey multiple means comparison.
Figure 7.
A box and whisker plot showing the percent growth inhibition index (GII) of Botrytis cinerea by itself (red), which acted as a control, and by different grape fungal endophytes isolated from ancient wine grapes. The percent GII was calculated as (HP-VP)/HP × 100, where HP is the mean of horizontal growth (mean radius) of the pathogen and VP is the vertical growth of the pathogen (mean radius) towards the endophyte. Values with shared letter groupings are not significantly different, as measured via a post hoc Tukey multiple means comparison.


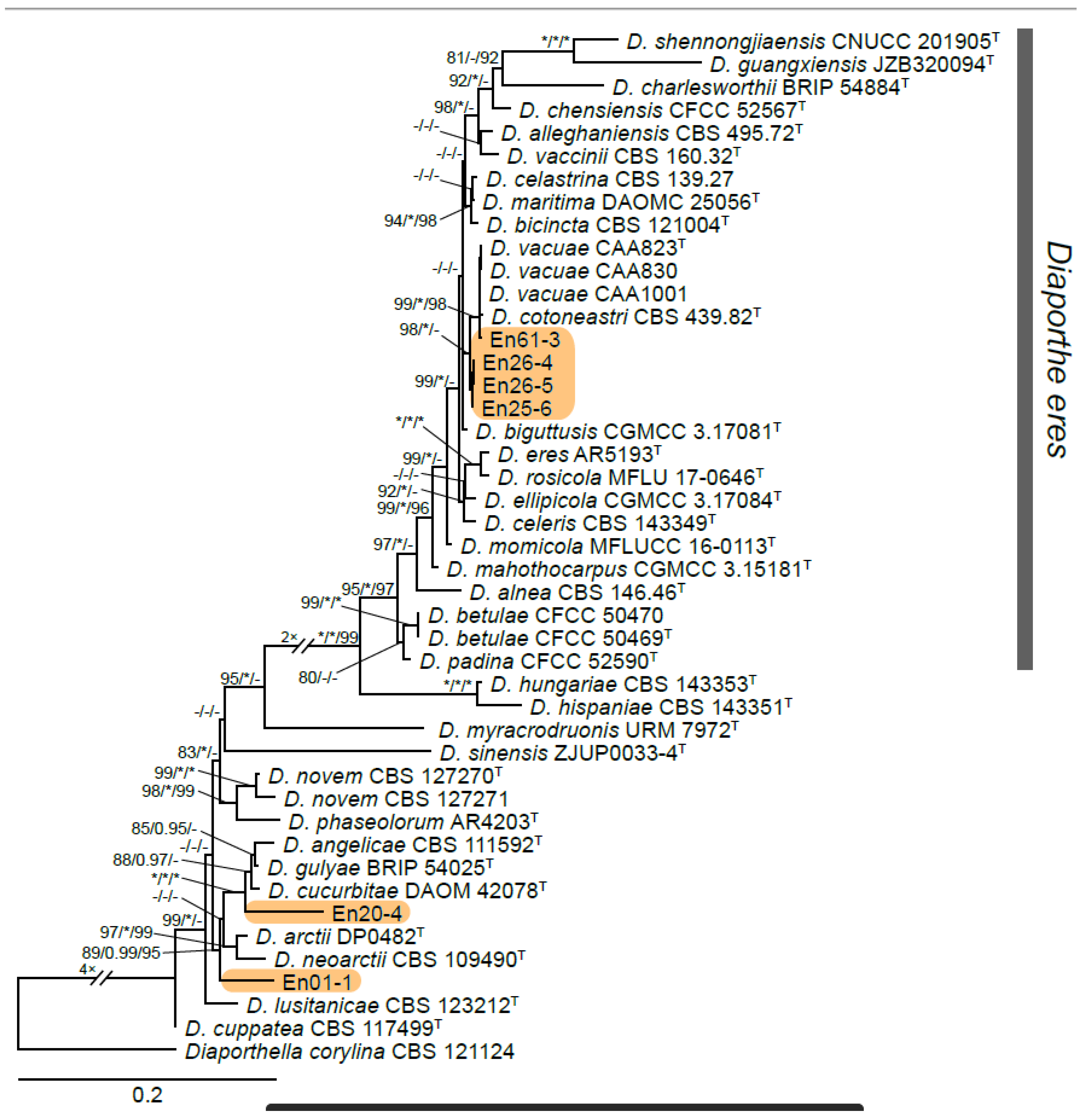{kind=link}
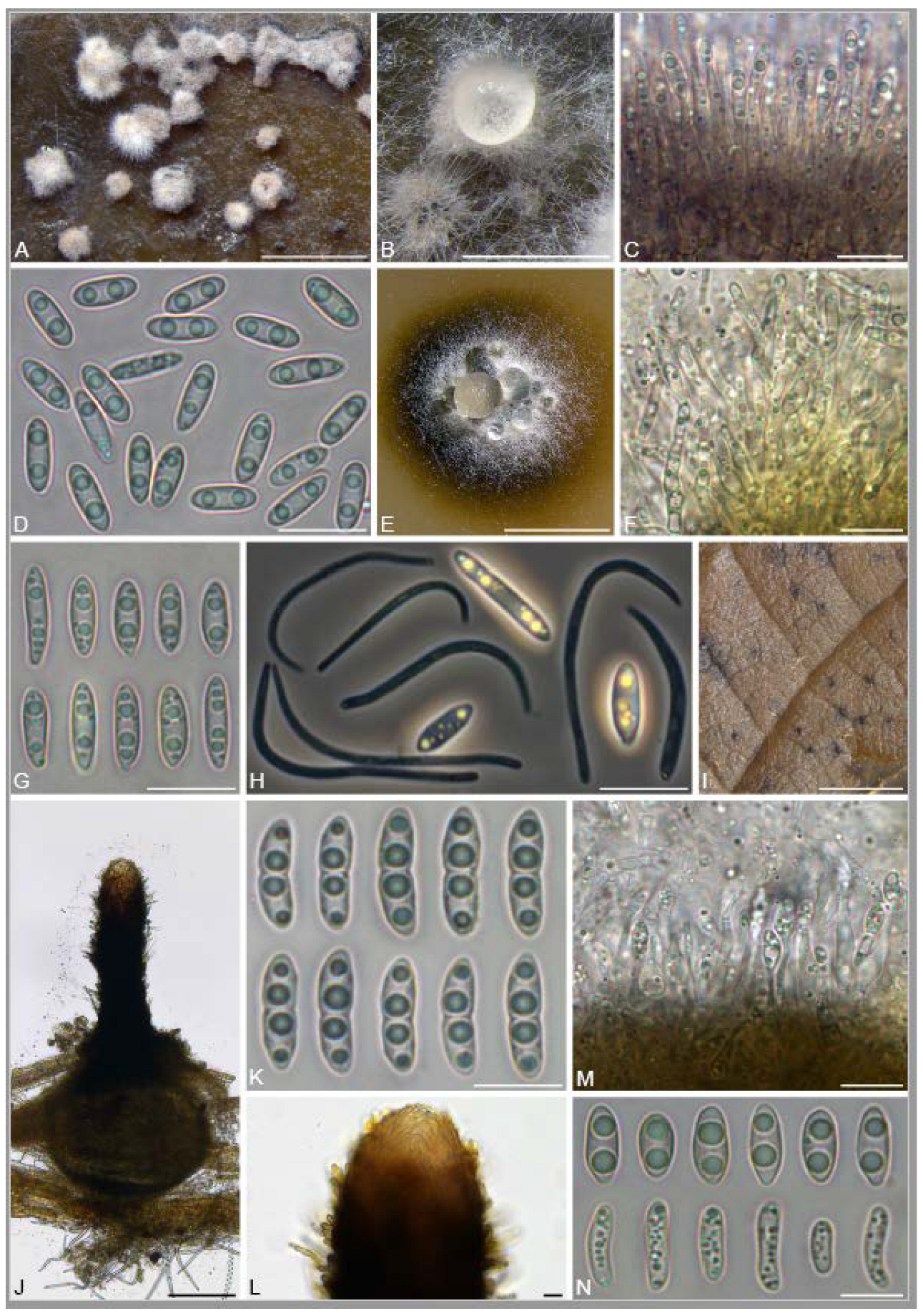{kind=link}
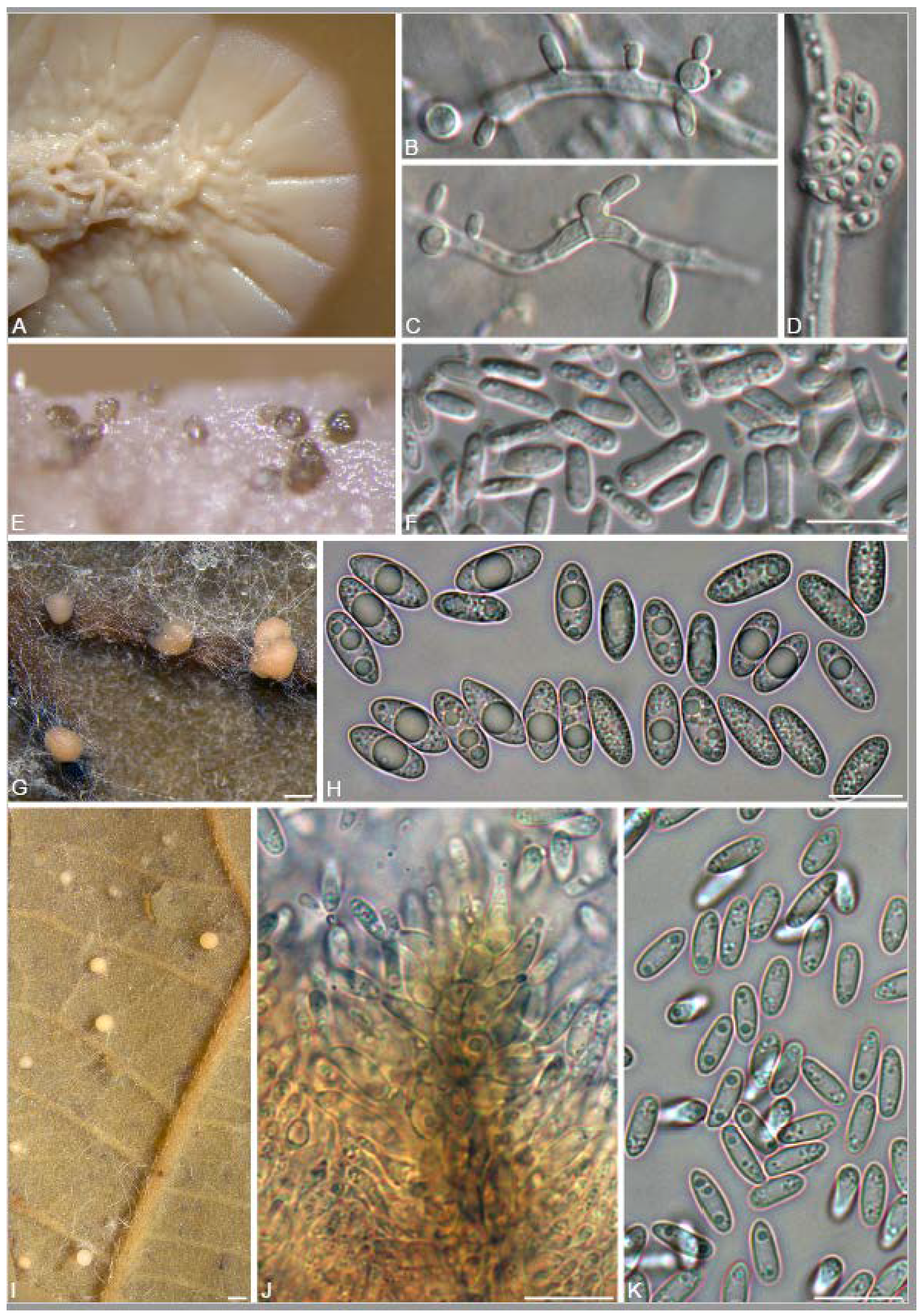{kind=link}
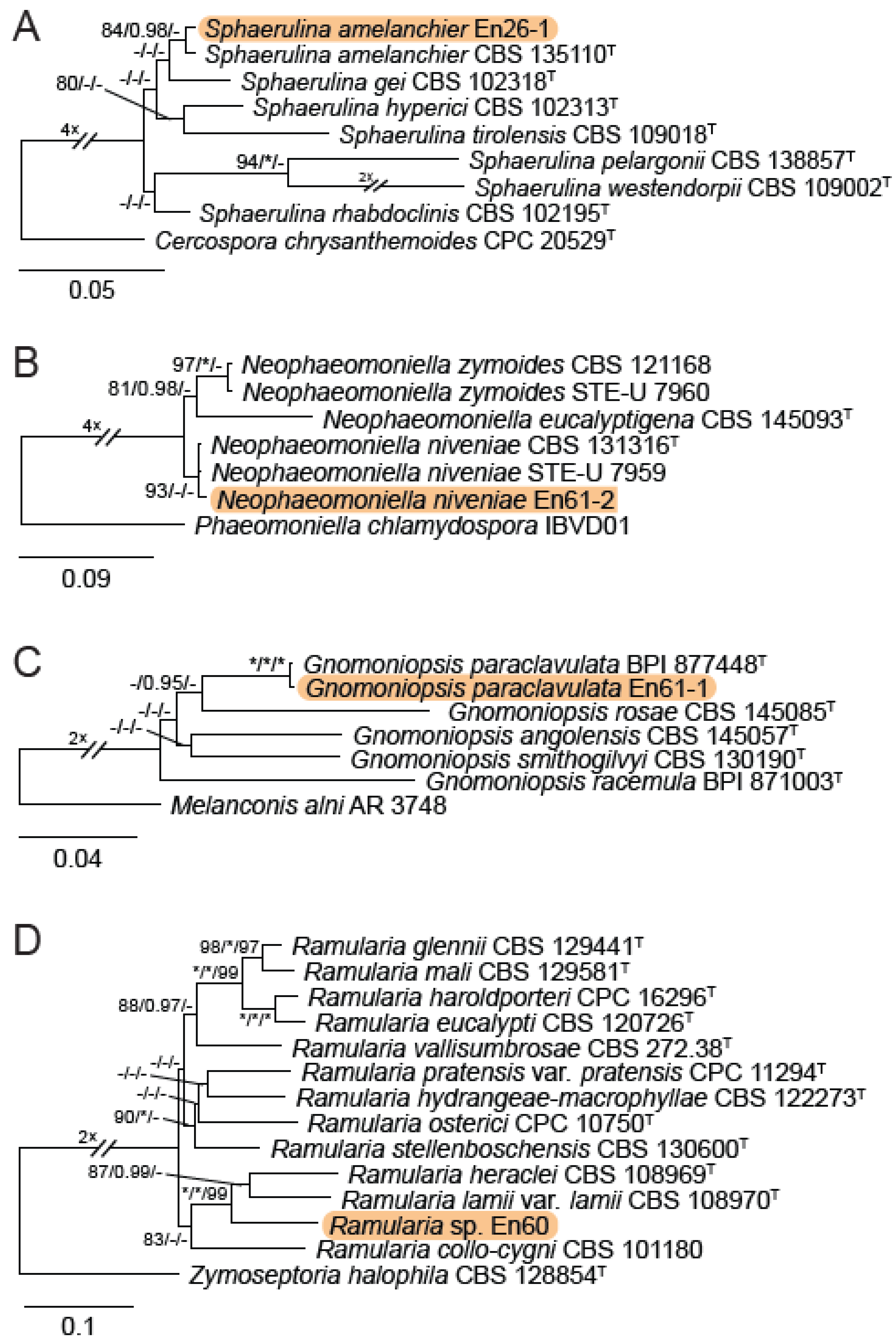{kind=link}
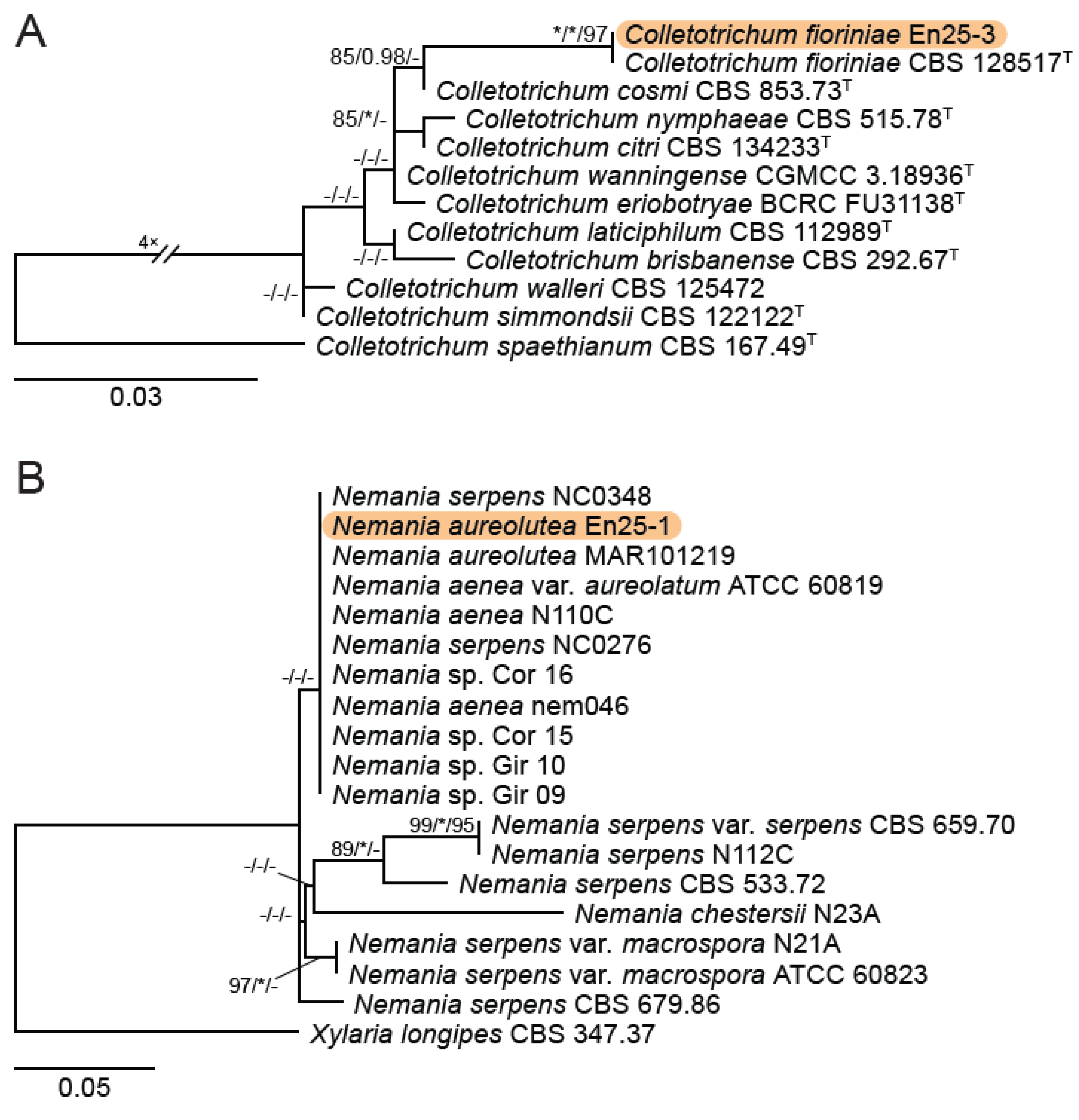{kind=link}
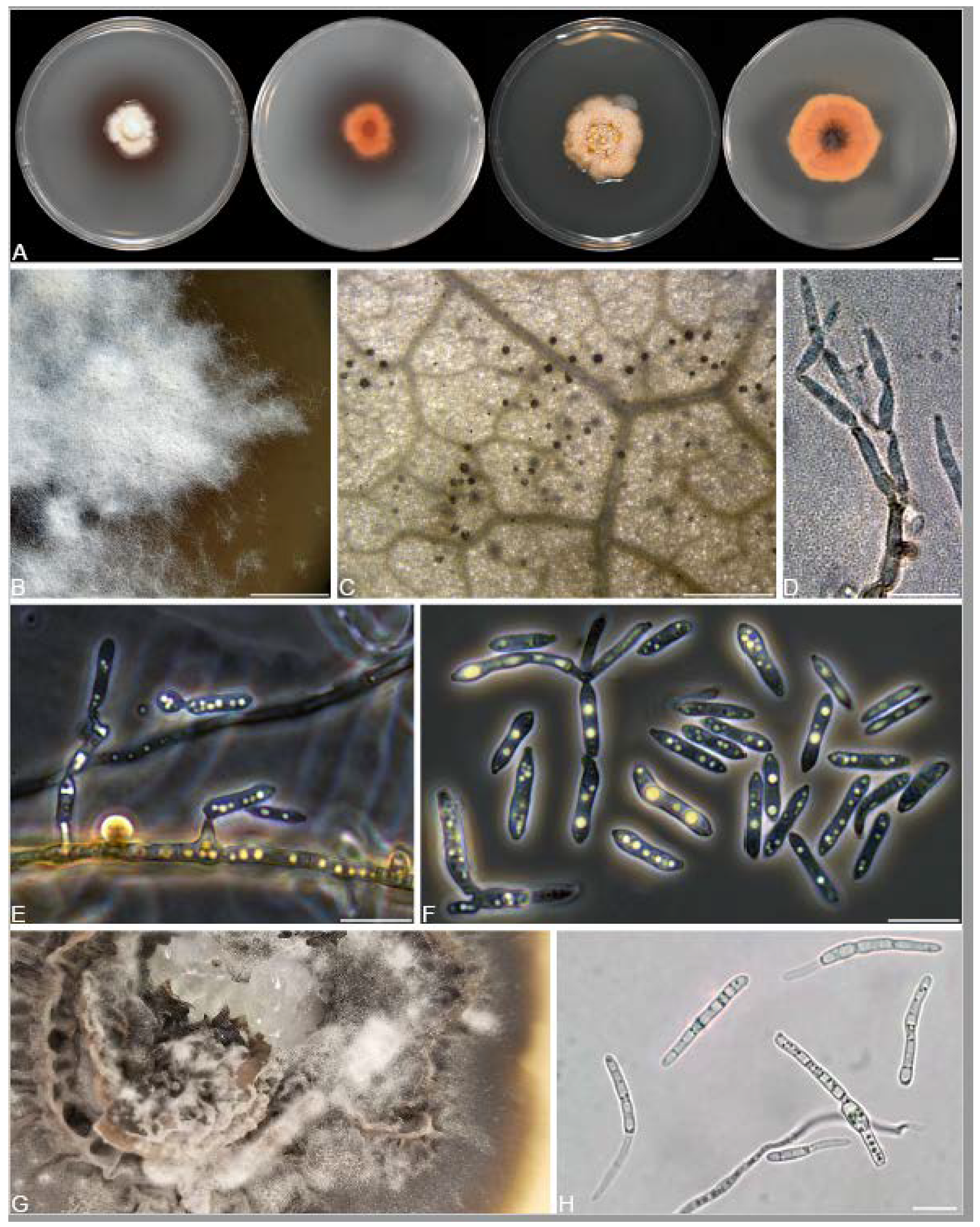{kind=link}
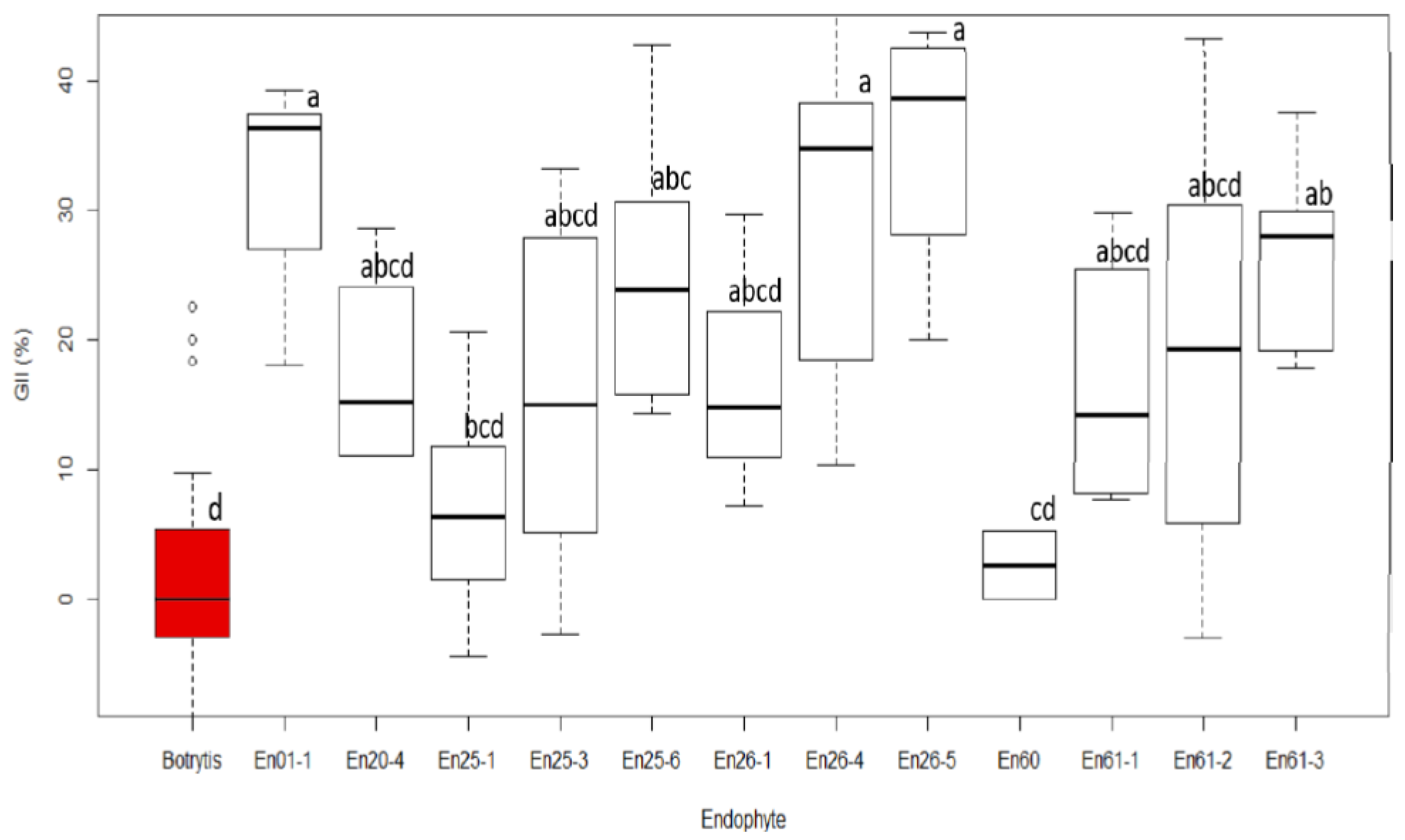{kind=link}
Table 1.
GenBank accession phylogenesis.
| Species | Isolate | Host | Location | GenBank Accession No. | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TUB2 | ITS | TEF1-α | RPB2 | ACT | CHS-1 | LSU | |||||
| Cercospora chrysanthemoides | CPC 20529 | Chrysanthemoides monilifera | South Africa | — | — | KC005813 | — | — | — | — | [30] |
| Colletotrichum brisbanense | CBS 292.67 | Capsicum annuum | Australia | — | — | — | — | — | JQ948952 | — | [31] |
| Colletotrichum citri | CBS 134233 | Citrus aurantifolia | China | — | — | — | — | — | KY856138 | — | [32] |
| Colletotrichum cosmi | CBS 853.73 | Cosmos sp. | Netherlands | — | — | — | — | — | JQ948935 | — | [31] |
| Colletotrichum eriobotryae | BCRC FU31138 | Eriobotrya japonica | Taiwan | — | — | — | — | — | MN191653 | — | [33] |
| Colletotrichum fioriniae | En25-3 | Vitis sp. ‘Clinton’ | Canada | — | MZ127182 | — | OK431476 | — | OK380951 | This study. | |
| Colletotrichum fioriniae | CBS 128517 | Fiorinia externa | USA | — | — | — | — | — | JQ948953 | — | [31] |
| Colletotrichum laticiphilum | CBS 112989 | Hevea brasiliensis | India | — | — | — | — | — | JQ948950 | — | [31] |
| Colletotrichum nymphaeae | CBS 515.78 | Nymphaea alba | Netherlands | — | — | — | — | — | JQ948858 | — | [31] |
| Colletotrichum simmondsii | CBS 122122 | Carica papaya | Australia | — | — | — | — | — | JQ948937 | — | [31] |
| Colletotrichum spaethianum | CBS 167.49 | Hosta sieboldiana | Germany | — | — | — | — | — | GU228297 | — | [34] |
| Colletotrichum walleri | CBS 125472 | Coffea sp. | Vietnam | — | — | — | — | — | JQ948936 | — | [31] |
| Colletotrichum wanningense | CGMCC 3.18936 | Hevea brasiliensis | China | — | — | — | — | — | MZ352012 | — | Liu et al. (Direct Submission) |
| Diaporthe aff. gulyae | En20-4 | Vitis sp. unknown | Canada | OK383388 | MZ127185 | — | — | — | OK380954 | This study | |
| Diaporthe alleghaniensis | CBS 495.72 | Betula alleghaniensis | Canada | KC843228 | NR_103696 | KC343733 | — | — | — | — | [35] |
| Diaporthe alnea | CBS 146.46 | Alnus sp. | Unknown | KC343976 | NR_147525 | KC343734 | — | — | — | — | [35] |
| Diaporthe angelicae | CBS 111592 | Heracleum sphondylium | Austria | KC343995 | KC343027 | KC343753 | — | — | — | — | [35] |
| Diaporthe arctii | DP0482 | Arctium lappa | Austria | KJ610891 | KJ590736 | KJ590776 | — | — | — | — | [36] |
| Diaporthe betulae | CFCC 50470 | Betula platyphylla | China | KT733021 | KT732951 | KT733017 | — | — | — | — | [37]) |
| Diaporthe betulae | CFCC 50469 | Betula platyphylla | China | KT733020 | NR_147578 | KT733016 | — | — | — | — | [37] |
| Diaporthe bicincta | CBS 121004 | Juglans sp. | USA | KC344102 | NR_147526 | KC343860 | — | — | — | — | [35] |
| Diaporthe biguttusis | CGMCC 3.17081 | Lithocarpus glaber | China | KF576306 | NR_147533 | KF576257 | — | — | — | — | [38] |
| Diaporthe celastrina | CBS 139.27 | Celastrus scandens | Unknown | KC344015 | NR_152457 | KC343773 | — | — | — | — | [35] |
| Diaporthe celeris | CBS 143349 | Vitis vinifera | UK | MG281190 | MG281017 | MG281538 | — | — | — | — | [39] |
| Diaporthe charlesworthii | BRIP 54884m | Rapistrum rugosum | Australia | KJ197268 | NR_147538 | KJ197250 | — | — | — | — | [40] |
| Diaporthe chensiensis | CFCC 52567 | Abies chensiensis | China | MH121584 | NR_165876 | MH121544 | — | — | — | — | [41] |
| Diaporthe cotoneastri | CBS 439.82 | Cotoneaster sp. | UK | JX275437 | MH861511 | GQ250341 | — | — | — | — | [42,43,44] |
| Diaporthe cucurbitae | DAOM 42078 | Cucumis sp. | Canada | KP118848 | KM453210 | KM453211 | — | — | — | — | [36] |
| Diaporthe cuppatea | CBS 117499 | Aspalathus linearis | South Africa | KC344025 | MH863021 | KC343783 | — | — | — | — | [35,42] |
| Diaporthe ellipicola | CGMCC 3.17084 | Lithocarpus glaber | China | KF576294 | NR_147531 | KF576245 | — | — | — | — | [38] |
| Diaporthe eres | En61-3 | Vitis sp. ‘Isabella’ | Canada | OK383392 | MZ127189 | — | — | — | OK380958 | This study | |
| Diaporthe eres | En26-4 | Vitis sp. ‘Clinton’ | Canada | OK383390 | MZ127187 | — | — | — | OK380956 | This study | |
| Diaporthe eres | En26-5 | Vitis sp. ‘Clinton’ | Canada | OK383391 | MZ127188 | — | — | — | OK380957 | This study | |
| Diaporthe eres | En25-6 | Vitis sp. ‘Clinton’ | Canada | OK383389 | MZ127186 | — | — | — | MZ127186 | This study | |
| Diaporthe eres | AR5193 | Ulmus sp. | Germany | KJ420799 | KJ210529 | KJ210550 | — | — | — | — | [45] |
| Diaporthe guangxiensis | JZB320094 | Vitis vinifera | China | MK500168 | MK335772 | MK523566 | — | — | — | — | [46] |
| Diaporthe gulyae | BRIP 54025 | Helianthus annuus | Australia | KJ197271 | NR_111615 | JN645803 | — | — | — | — | [40,47,48] |
| Diaporthe hispaniae | CBS 143351 | Vitis vinifera | Spain | MG281296 | MG281123 | MG281644 | — | — | — | — | [39] |
| Diaporthe hungariae | CBS 143353 | Vitis vinifera | Hungary | MG281299 | MG281126 | MG281647 | — | — | — | — | [39] |
| Diaporthe lusitanicae | CBS 123212 | Foeniculum vulgare | Portugal | KC344104 | MH863279 | KC343862 | — | — | — | — | [35,42], |
| Diaporthe mahothocarpus | CGMCC 3.15181 | Lithocarpus glabra | China | KF576312 | KC153096 | KC153087 | — | — | — | — | |
| Diaporthe maritima | DAOMC 25056 | Picea rubens | Canada | KU574615 | NR_152463 | KU552023 | — | — | — | — | [49] |
| Diaporthe momicola | MFLUCC 16-0113 | Prunus persica | China | KU557587 | NR_172386 | KU557631 | — | — | — | — | [50] |
| Diaporthe myracrodruonis | URM 7972 | Astronium urundeuva | Brazil | MK205291 | NR_163320 | MK213408 | — | — | — | — | [15] |
| Diaporthe neoarctii | CBS 109490 | Ambrosia trifida | USA | KC344113 | NR_111854 | KC343871 | — | — | — | — | [35,47] |
| Diaporthe novem | CBS 127270 | Glycine max | Croatia | KC344124 | MH864503 | KC343882 | — | — | — | — | [35,42] |
| Diaporthe novem | CBS 127271 | Glycine max | Croatia | KC344125 | MH864504 | KC343883 | — | — | — | — | [35,42] |
| Diaporthe padina | CFCC 52590 | Prunus padus | China | MH121604 | NR_165879 | MH121567 | — | — | — | — | [41] |
| Diaporthe phaseolorum | AR4203 | Phaseolus vulgaris | USA | KJ610893 | KJ590738 | KJ590739 | — | — | — | — | [36] |
| Diaporthe rosicola | MFLU 17-0646 | Rosa sp. | UK | MG843877 | NR_157515 | MG829270 | — | — | — | — | [51] |
| Diaporthe shennongjiaensis | CNUCC 201905 | Juglans regia | China | MN227012 | MN216229 | MN224672 | — | — | — | — | [52] |
| Diaporthe sinensis | ZJUP0033-4 | Amaranthus sp. | China | MK660447 | MK637451 | MK660449 | — | — | — | — | [53] |
| Diaporthe sp. | En01-1 | Vitis riparia | Canada | OK383387 | MZ127184 | — | — | — | OK380953 | This study | |
| Diaporthe vaccinii | CBS 160.32 | Oxycoccus macrocarpos | USA | KC344196 | NR_103701 | KC343954 | — | — | — | — | [35] |
| Diaporthe vacuae | CAA829 | Vaccinium corymbosum | Portugal | MK837928 | MK792306 | MK828077 | — | — | — | — | [54] |
| Diaporthe vacuae | CAA830 | Vaccinium corymbosum | Portugal | MK837931 | MK792309 | MK828080 | — | — | — | — | [54]) |
| Diaporthe vacuae | CAA1001 | Quercus suber | Portugal | MT309458 | MT237172 | MT309432 | — | — | — | — | [54] |
| Diaporthella corylina | CBS 121124 | Corylus sp. | China | KC343972 | KC343004 | KC343730 | — | — | — | — | [35] |
| Gnomoniopsis angolensis | CBS 145057 | Unknown plant sp. | Angola | — | MK047428 | — | MK047539 | — | — | — | [55] |
| Gnomoniopsis paraclavulata | BPI 877448 | Carpinus caroliniana | USA | — | EU254839 | — | EU219248 | — | — | [56] | |
| Gnomoniopsis paraclavulata | En61-1 | Vitis sp. ‘Isabella’ | Canada | OK383385 | MZ127179 | — | OK431474 | — | — | OK380949 | This study. |
| Gnomoniopsis racemula | BPI 871003 | Epilobium angustifolium | USA | — | EU254841 | — | EU219241 | — | — | — | [56] |
| Gnomoniopsis rosae | CBS 145085 | Rosa sp. | New Zealand | — | NR_161142 | — | MK047547 | — | — | — | [55] |
| Gnomoniopsis smithogilvyi | CBS 130190 | Castanea sp. | Australia | — | MH865607 | — | JQ910648 | — | — | — | [30,42] |
| Melanconis alni | AR 3748 | Alnus viridis | Austria | — | EU199195 | — | EU199153 | — | — | — | [57] |
| Nemania aenea | N110C | Unknown | Unknown | — | AJ390427 | — | — | — | — | — | [58] |
| Nemania aenea | nem046 | Centaurea stoebe | USA | — | EF589887 | — | — | — | — | — | [59] |
| Nemania aenea var. aureolatum | ATCC 60819 | Quercus sp. | Switzerland | — | AF201704 | — | — | — | — | — | [60] |
| Nemania aureolutea | En25-1 | Vitis sp. ‘Clinton’ | Canada | OK383386 | MZ127183 | — | OK431477 | OK431472 | — | OK380952 | This study |
| Nemania aureolutea | MAR101219 | Quercus canariensis | Spain | — | MW136058 | — | — | — | — | — | [61] |
| Nemania chestersii | N23A | Unknown | Unknown | — | AJ390430 | — | — | — | — | — | [58] |
| Nemania serpens | NC0348 | Lecanora oreinoides | USA | — | JQ761380 | — | — | — | — | — | [62] |
| Nemania serpens | NC0276 | Diploschistes rampoddensis | USA | — | JQ761314 | — | — | — | — | — | [62] |
| Nemania serpens | N112C | Unknown | Unknown | — | AJ390436 | — | — | — | — | — | [58] |
| Nemania serpens | CBS 533.72 | Corylus avellana | Netherlands | — | FN428829 | — | — | — | — | — | [63] |
| Nemania serpens | CBS 679.86 | Betula sp. | Switzerland | — | KU683765 | — | — | — | — | — | [62] |
| Nemania serpens var. macrospora | N21A | Unknown | Unknown | — | AJ390433 | — | — | — | — | — | [58] |
| Nemania serpens var. macrospora | ATCC 60823 | Unknown | Unknown | — | AF201707 | — | — | — | — | — | [60] |
| Nemania serpens var. serpens | CBS 659.70 | Soil | Canada | — | MH859890 | — | — | — | — | — | [42] |
| Nemania sp. | Cor 16 | Nephroma laevigatum | France | — | MG916993 | — | — | — | — | — | [64] |
| Nemania sp. | Cor 15 | Nephroma laevigatum | France | — | MG916992 | — | — | — | — | — | [64] |
| Nemania sp. | Gir 10 | Nephroma laevigatum | France | — | MG917014 | — | — | — | — | — | [64] |
| Nemania sp. | Gir 9 | Nephroma laevigatum | France | — | MG917013 | — | — | — | — | — | [64] |
| Neophaeomoniella eucalyptigena | CBS 145093 | Eucalyptus pilularis | Australia | — | NR_161148 | MK047569 | — | — | — | — | [55] |
| Neophaeomoniella niveniae | CBS 131316 | Nivenia stokoei | South Africa | — | JQ044435 | MN861682 | — | — | — | — | [65,66] |
| Neophaeomoniella niveniae | STE-U 7959 | Olea europaea subsp. cuspidata | South Africa | — | MT791053 | MT787396 | — | — | — | — | [67] |
| Neophaeomoniella niveniae | En61-2 | Vitis sp. ‘Isabella’ | Canada | OK383384 | MZ127178 | — | — | — | OK380948 | This study | |
| Neophaeomoniella zymoides | CBS 121168 | Prunus salicina | South Africa | — | GQ154600 | MN861679 | — | — | — | — | [65,68] |
| Neophaeomoniella zymoides | STE-U 7960 | Olea europaea subsp. cuspidata | South Africa | — | MT791054 | MT787397 | — | — | — | — | [67] |
| Phaeomoniella chlamydospora | IBVD01 | Vitis vinifera | Brazil | — | KP213118 | KP213113 | — | — | — | — | [69] |
| Ramularia collo-cygni | CBS 101180 | Hordeum vulgare | Austria | — | NR_154944 | — | KX288543 | KX287666 | — | — | [70] |
| Ramularia eucalypti | CBS 120726 | Corymbia grandifolia | Italy | — | KJ504792 | — | KJ504663 | KF253635 | — | — | [71,72] |
| Ramularia glennii | CBS 129441 | Homo sapiens | Netherlands | — | MH865235 | — | KJ504433 | KJ504640 | — | — | [42,71] |
| Ramularia haroldporteri | CPC 16296 | Unidentified plant | South Africa | — | NR_154911 | — | KJ504637 | KJ504430 | — | — | [71] |
| Ramularia heraclei | CBS 108969 | Heracleum sphondylium | Netherlands | — | NR_154948 | — | KX288578 | KX287702 | — | — | [70] |
| Ramularia hydrangeae-macrophyllae | CBS 122273 | Hydrangea macrophylla | New Zealand | — | NR_145125 | — | KX288592 | KX287716 | — | — | [70] |
| Ramularia lamii var. lamii | CBS 108970 | Lamium album | Netherlands | — | NR_154949 | — | KX288620 | KX287744 | — | — | [70] |
| Ramularia mali | CBS 129581 | Malus sp. | Italy | — | NR_156582 | — | KJ504649 | MH876894 | — | — | [42,71] |
| Ramularia osterici | CPC 10750 | Ostericum koreanum | South Korea | — | NR_154950 | — | KX288642 | KX287765 | — | — | [70] |
| Ramularia pratensis var. pratensis | CPC 11294 | Rumex crispus | South Korea | — | EU019284.2 | — | KT216537 | KF903599 | — | — | [73,74,75] |
| Ramularia sp. | En60-1 | Vitis sp. ‘Marechal foch’ | Canada | OK383393 | MZ127180 | — | OK431478 | OK431473 | — | OK380959 | This study |
| Ramularia stellenboschensis | CBS 130600 | Protea sp. | South Africa | — | NR_145101 | — | KX288676 | KX287798 | — | — | [42,70] |
| Ramularia vallisumbrosae | CBS 272.38 | Narcissus ‘Golden Spur’ | UK | — | NR_154953 | — | KX288698 | KX288699 | — | — | [70] |
| Sphaerulina amelanchier | En26-1 | Vitis sp. ‘Clinton’ | Canada | — | MZ127181 | OK431475 | — | — | OK380950 | This study. | |
| Sphaerulina amelanchier | CBS 135110 | Amelanchier sp. | Netherlands | — | — | KF253543 | — | — | — | — | [72] |
| Sphaerulina gei | CBS 102318 | Geum urbanum | Netherlands | — | — | KF253560 | — | — | — | — | [72] |
| Sphaerulina hyperici | CBS 102313 | Hypericum sp. | Netherlands | — | — | KF253563 | — | — | — | — | [72] |
| Sphaerulina pelargonii | CBS 138857 | Pelargonium sp. | South Africa | — | — | KP004506 | — | — | — | — | [72] |
| Sphaerulina rhabdoclinis | CBS 102195 | Pseudotsuga menziesii | Germany | — | — | KF253578 | — | — | — | — | [72] |
| Sphaerulina tirolensis | CBS 109018 | Rubus idaeus | Austria | — | — | KF253585 | — | — | — | — | [72] |
| Sphaerulina westendorpii | CBS 109002 | Rubus sp. | Netherlands | — | — | KF253588 | — | — | — | — | [72] |
| Xylaria longipes | CBS 347.37 | Unknown | Unknown | — | MH855925 | — | — | — | — | — | [42] |
| Zymoseptoria halophila | CBS 128854 | Hordeum vulgare | Iran | — | MH865126 | — | KX348110 | KF253946 | — | — | [30,42,72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ali, S.; Wright, A.H.; Tanney, J.B.; Renaud, J.B.; Sumarah, M.W. Fungal Endophytes: Discovering What Lies within Some of Canada’s Oldest and Most Resilient Grapevines. J. Fungi 2024, 10, 105. https://doi.org/10.3390/jof10020105
AMA Style
Ali S, Wright AH, Tanney JB, Renaud JB, Sumarah MW. Fungal Endophytes: Discovering What Lies within Some of Canada’s Oldest and Most Resilient Grapevines. Journal of Fungi. 2024; 10(2):105. https://doi.org/10.3390/jof10020105
Chicago/Turabian StyleAli, Shawkat, A. Harrison Wright, Joey B. Tanney, Justin B. Renaud, and Mark W. Sumarah. 2024. "Fungal Endophytes: Discovering What Lies within Some of Canada’s Oldest and Most Resilient Grapevines" Journal of Fungi 10, no. 2: 105. https://doi.org/10.3390/jof10020105
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.