
Community Structure and Diversity of Endophytic Fungi in Cultivated Polygala crotalarioides at Two Different Growth Stages Based on Culture-Independent and Culture-Based Methods

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Samples
2.2. DNA Extraction, Amplification of ITS rDNA Region and High-Throughput Sequencing
2.3. Sequence Processing and Data Analysis for HTS
2.4. Isolation and Identification of Endophytic Fungi with Culture-Based Method
2.5. AChE-Inhibitory-Activity Screening of Isolated Endophytic Fungi
3. Results
3.1. Surface-Disinfection Efficiency
3.2. Analysis of Endophytic Fungal Community Structures with HTS
3.2.1. Overview of Sample Sequences
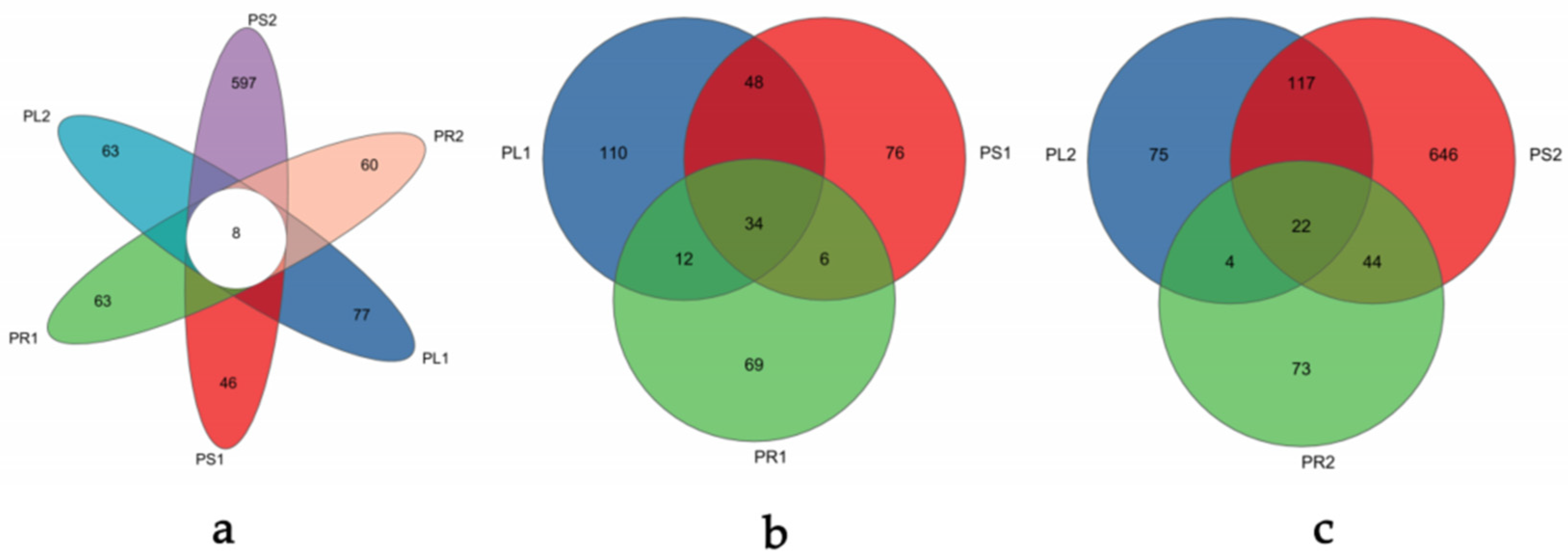
3.2.2. Analysis of Operational Taxonomic Units (OTUs)
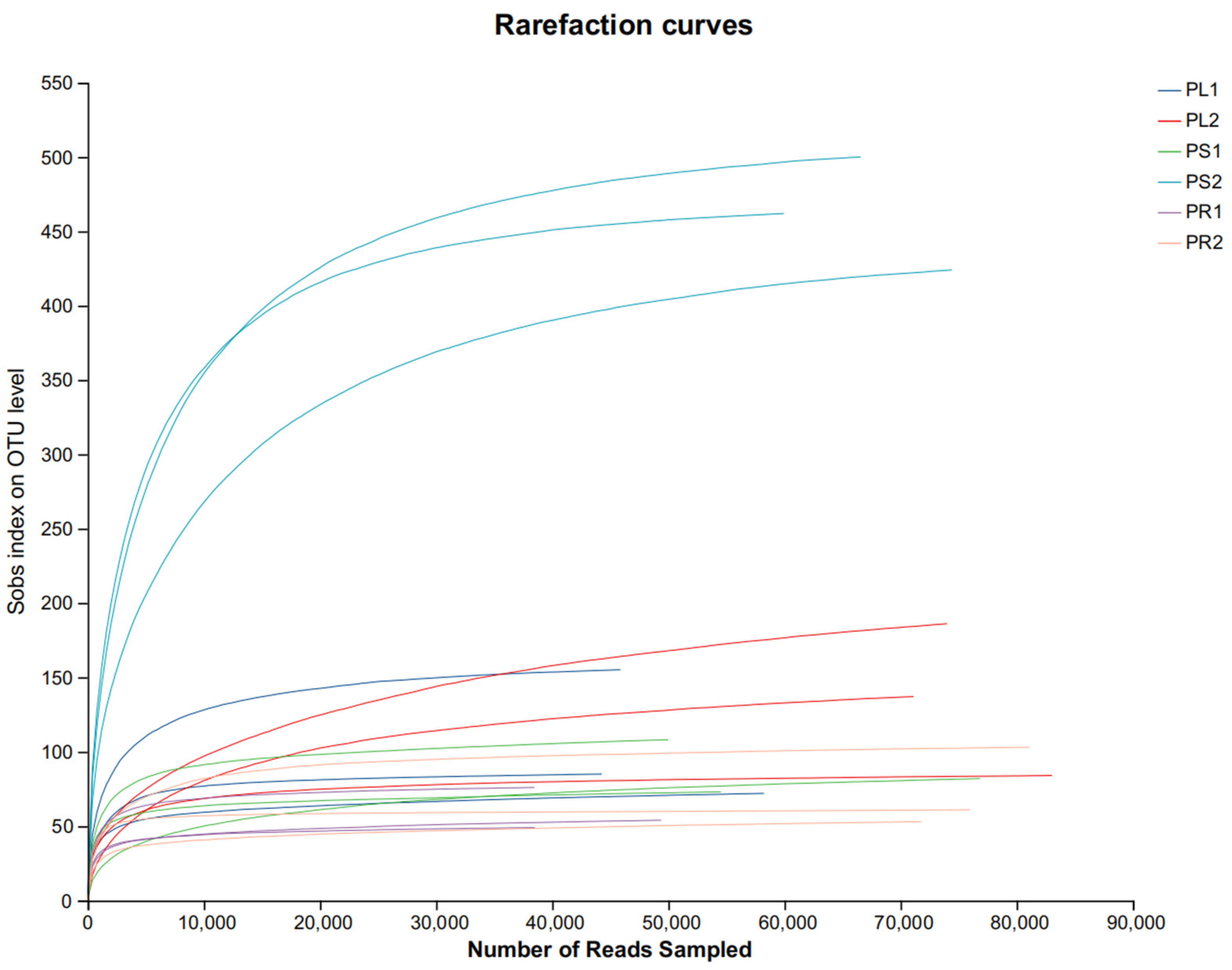
3.2.3. Alpha Diversity Analysis of the Endophytic Fungal Communities
3.2.4. Beta Diversity Analysis of the Endophytic Fungal Communities
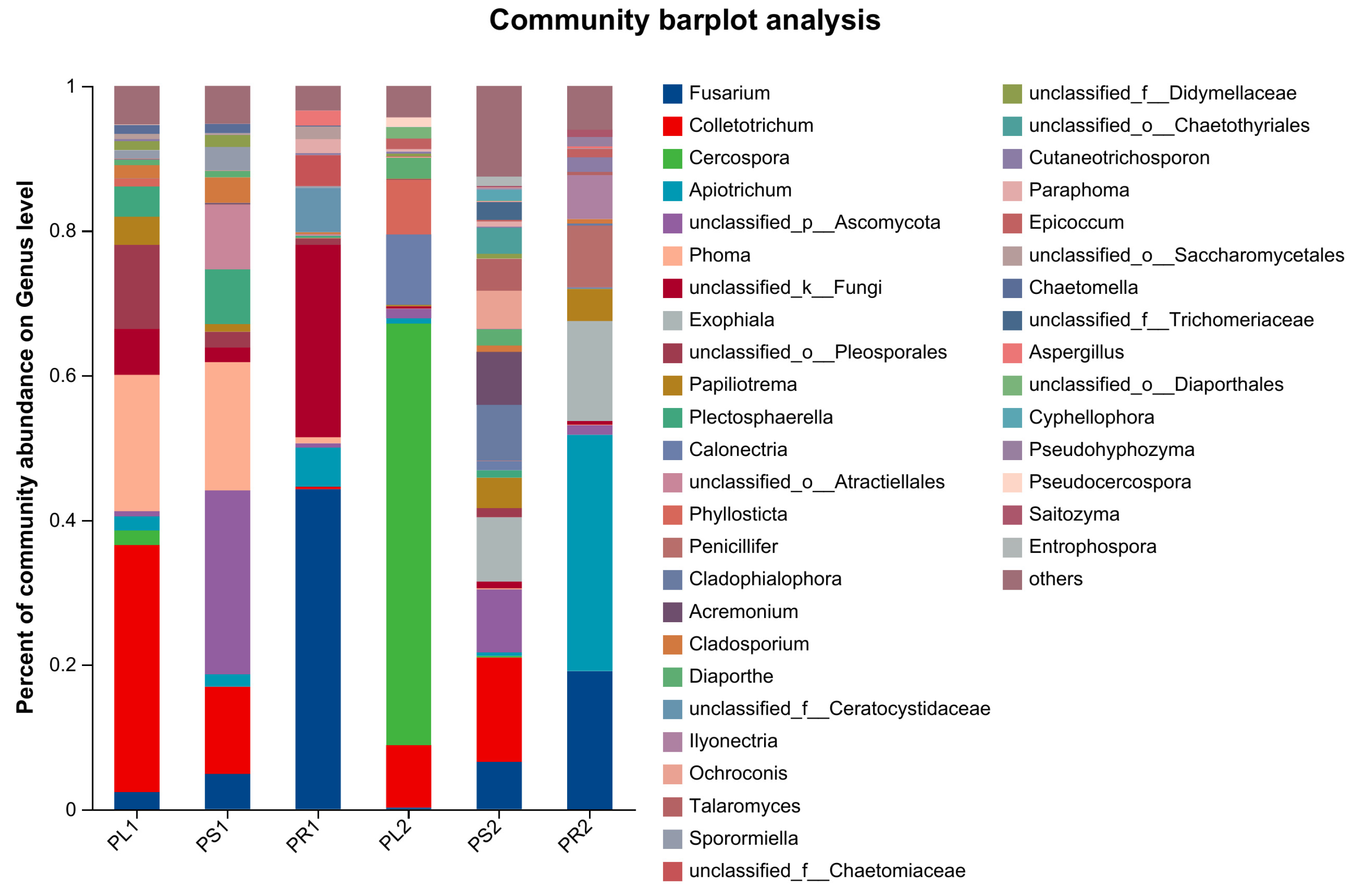
3.2.5. Composition of Endophytic Fungal Communities
3.3. Analysis of Culturable Endophytic Fungus Community Structure
3.4. AChE-Inhibiting Activity of Isolated Endophytic Fungi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: San Diego, CA, USA, 2008; pp. 419–457. [Google Scholar]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Dos Santos, R.; Morais-Urano, R.P.; Marçal, R.M.; Silva, G.H.; Santos, M.F.C. Acetylcholinesterase and butyrylcholinesterase inhibition by nectriapyrone and tryptophol isolated from endophytic fungus Phomopsis sp. Nat. Prod. Res. 2022, 36, 4153–4158. [Google Scholar] [CrossRef]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef]
- Fan, M.; Chen, X.; Luo, X.; Zhang, H.; Liu, Y.; Zhang, Y.; Wu, J.; Zhao, C.; Zhao, P. Diversity of endophytic fungi from the leaves of Vaccinium dunalianum. Lett. Appl. Microbiol. 2020, 71, 479–489. [Google Scholar] [CrossRef]
- Qin, X.; Xu, J.; An, X.; Yang, J.; Wang, Y.; Dou, M.; Wang, M.; Huang, J.; Fu, Y. Insight of endophytic fungi promoting the growth and development of woody plants. Crit. Rev. Biotechnol. 2024, 44, 78–99. [Google Scholar] [CrossRef]
- Ripa, F.A.; Cao, W.D.; Tong, S.; Sun, J.G. Assessment of plant growth promoting and abiotic stress tolerance properties of wheat endophytic fungi. Biomed. Res. Int. 2019, 2019, 6105865. [Google Scholar] [CrossRef]
- Du, Y.C.; Kong, L.J.; Cao, L.S.; Zhang, W.; Zhu, Q.; Ma, C.Y.; Sun, K.; Dai, C.C. Endophytic fungus Phomopsis liquidambaris enhances Fe absorption in peanuts by reducing hydrogen peroxide. Front. Plant Sci. 2022, 13, 872242. [Google Scholar] [CrossRef]
- Akram, S.; Ahmed, A.; He, P.; He, P.; Liu, Y.; Wu, Y.; Munir, S.; He, Y. Uniting the role of endophytic fungi against plant pathogens and their interaction. J. Fungi 2023, 9, 72. [Google Scholar] [CrossRef]
- Lacerda, I.; Polonio, J.C.; Golias, H.C. Endophytic fungi as a source of antiviral compounds—A review. Chem. Biodivers. 2022, 19, e202100971. [Google Scholar] [CrossRef]
- Zheng, H.; Qiao, M.; Xu, J.; Yu, Z. Culture-based and culture-independent assessments of endophytic fungal diversity in aquatic plants in Southwest China. Front. Fungal Biol. 2021, 2, 692549. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Miao, L.; Li, H.; Lin, A.; Song, F.; Zhang, P. Illumina-based analysis yields new insights into the diversity and composition of endophytic fungi in cultivated Huperzia serrata. PLoS ONE 2020, 15, e0242258. [Google Scholar] [CrossRef]
- Yan, K.; Pei, Z.; Meng, L.; Zheng, Y.; Wang, L.; Feng, R.; Li, Q.; Liu, Y.; Zhao, X.; Wei, Q.; et al. Determination of community structure and diversity of seed-vectored endophytic fungi in Alpinia zerumbet. Front. Microbiol. 2022, 13, 814864. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Ju, M.; Yan, S.; Zhang, Q.; Gu, P. The endophytic fungi diversity, community structure, and ecological function prediction of Sophora alopecuroides in Ningxia, China. Microorganisms 2022, 10, 2099. [Google Scholar] [CrossRef]
- Lu, Z.; Ma, Y.; Xiao, L.; Yang, H.; Zhu, D. Diversity of endophytic fungi in Huperzia serrata and their acetylcholinesterase inhibitory activity. Sustainability 2021, 13, 12073. [Google Scholar] [CrossRef]
- Johnston, P.R.; Park, D.; Smissen, R.D. Comparing diversity of fungi from living leaves using culturing and high-throughput environmental sequencing. Mycologia 2017, 109, 643–654. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Prakathi, P.; Sivakumar, T.; Thiruvengadam, M.; Jayaprakash, B.; Baskar, V.; Rebezov, M.; Derkho, M.; Zengin, G.; Shariati, M.A. Bioactive components and health potential of endophytic micro-fungal diversity in medicinal plants. Antibiotics 2022, 11, 1533. [Google Scholar] [CrossRef]
- Zhang, L.; Yong, Y.-Y.; Deng, L.; Wang, J.; Law, B.Y.-K.; Hu, M.-L.; Wu, J.-M.; Yu, L.; Wong, V.K.-W.; Yu, C.-L.; et al. Therapeutic potential of Polygala saponins in neurological diseases. Phytomedicine 2023, 108, 154483. [Google Scholar] [CrossRef]
- Ma, J.; Wang, J.; Li, C.; Liu, J.; Chen, C.; Hua, Y. The complete chloroplast genome characteristics of Polygala crotalarioides Buch.-Ham. ex DC. (Polygalaceae) from Yunnan, China. Mitochondrial DNA B Resour. 2021, 6, 2838–2840. [Google Scholar] [CrossRef]
- Hua, Y.; Chen, C.X.; Zhou, J. Two new triterpenoid saponins from Polygala crotalarioides. Chin. Chem. Lett. 2010, 21, 1107–1110. [Google Scholar] [CrossRef]
- Deng, X.; Zhao, S.; Liu, X.; Han, L.; Wang, R.; Hao, H.; Jiao, Y.; Han, S.; Bai, C. Polygala tenuifolia: A source for anti-Alzheimer’s disease drugs. Pharm. Biol. 2020, 58, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-H.; Zhang, S.-Y.; Guo, Q.; Chai, X.-Y.; Jiang, Y.; Tu, P.-F. Chemical investigation of the roots of Polygala sibirica L. Chin. J. Nat. Med. 2014, 12, 225–228. [Google Scholar] [CrossRef]
- Zhao, X.; Cui, Y.; Wu, P.; Zhao, P.; Zhou, Q.; Zhang, Z.; Wang, Y.; Zhang, X. Polygalae Radix: A review of its traditional uses, phytochemistry, pharmacology, toxicology, and pharmacokinetics. Fitoterapia 2020, 147, 104759. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Pharmacological potential of fungal endophytes associated with medicinal plants: A review. J. Fungi 2021, 7, 147. [Google Scholar] [CrossRef]
- Gómez, O.C.; Luiz, J.H.H. Endophytic fungi isolated from medicinal plants: Future prospects of bioactive natural products from Tabebuia/Handroanthus endophytes. Appl. Microbiol. Biot. 2018, 102, 9105–9119. [Google Scholar] [CrossRef]
- Mattson, M.P. Pathways towards and away from Alzheimer’s disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef]
- Akıncıoğlu, H.; Gülçin, İ. Potent Acetylcholinesterase Inhibitors: Potential Drugs for Alzheimer’s Disease. Mini-Rev. Med. Chem. 2020, 20, 703–715. [Google Scholar] [CrossRef]
- Alrouji, M.; DasGupta, D.; Ashraf, G.M.; Bilgrami, A.L.; Alhumaydhi, F.A.; Al Abdulmonem, W.; Shahwan, M.; Alsayari, A.; Atiya, A.; Shamsi, A. Inhibition of microtubule affinity regulating kinase 4 by an acetylcholinesterase inhibitor, Huperzine A: Computational and experimental approaches. Int. J. Biol. Macromol. 2023, 235, 123831. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Pant, A.; Vasundhara, M. Endophytic fungi: A potential source for drugs against central nervous system disorders. Braz. J. Microbiol. 2023, 54, 1479–1499. [Google Scholar] [CrossRef] [PubMed]
- Vig, R.; Bhadra, F.; Gupta, S.K.; Sairam, K.; Vasundhara, M. Neuroprotective effects of quercetin produced by an endophytic fungus Nigrospora oryzae isolated from Tinospora cordifolia. J. Appl. Microbiol. 2022, 132, 365–380. [Google Scholar] [CrossRef]
- Hamed, A.A.; El-Shiekh, R.A.; Mohamed, O.G.; Aboutabl, E.A.; Fathy, F.I.; Fawzy, G.A.; Al-Taweel, A.M.; Elsayed, T.R.; Tripathi, A.; Al-Karmalawy, A.A. Cholinesterase inhibitors from an endophytic fungus Aspergillus niveus fv-er401: Metabolomics, isolation and molecular docking. Molecules 2023, 28, 2559. [Google Scholar] [CrossRef]
- Ren, F.; Dong, W.; Yan, D.H. Organs, cultivars, soil, and fruit properties affect structure of endophytic mycobiota of Pinggu peach trees. Microorganisms 2019, 7, 922. [Google Scholar] [CrossRef]
- Thi Minh Le, T.; Thi Hong Hoang, A.; Thi Bich Le, T.; Thi Bich Vo, T.; Van Quyen, D.; Hoang Chu, H. Isolation of endophytic fungi and screening of Huperzine A-producing fungus from Huperzia serrata in Vietnam. Sci. Rep. 2019, 9, 16152. [Google Scholar] [CrossRef]
- Kadiyala, M.; Ponnusankar, S.; Elango, K. Screening of siddha medicinal plants for its in-vitro acetylcholinesterase and butyrylcholineseerase inhibitory activity. Pharmacogn. Mag. 2014, 10, S294–S298. [Google Scholar]
- Ancheeva, E.; Daletos, G.; Proksch, P. Bioactive secondary metabolites from endophytic fungi. Curr. Med. Chem. 2020, 27, 1836–1854. [Google Scholar] [CrossRef]
- Tiwari, P.; Bae, H. Endophytic fungi: Key insights, emerging prospects, and challenges in natural product drug discovery. Microorganisms 2022, 10, 360. [Google Scholar] [CrossRef]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on endophytic fungi related to secondary metabolites and biotransformation applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological activities of some new secondary metabolites isolated from endophytic fungi: A review study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Nascimento, C.C.F.; Barbosa, R.d.N.; da Silva, D.C.V.; Svedese, V.M.; Silva-Nogueira, E.B.; Gomes, B.S.; Paiva, L.M.; Souza-Motta, C.M. Endophytic fungi from medicinal plant Bauhinia forficata: Diversity and biotechnological potential. Braz. J. Microbiol. 2015, 46, 49–57. [Google Scholar] [CrossRef]
- Strobel, G.; Stierle, A.; Stierle, D.; Hess, W.M. Taxomyces andreanae, a proposed new taxon for a bulbilliferous hyphomycete associated with Pacific yew (Taxus brevifolia). Mycotaxon 1993, 47, 71–80. [Google Scholar]
- Wang, Y.; Zhang, Y.; Cong, H.; Li, C.; Wu, J.; Li, L.; Jiang, J.; Cao, X. Cultivable endophyte resources in medicinal plants and effects on hosts. Life 2023, 13, 1695. [Google Scholar] [CrossRef]
- Sang, X.; Yang, M.; Su, J. Research on endophytic fungi for producing huperzine A on a large-scale. Crit. Rev. Microbiol. 2020, 46, 654–664. [Google Scholar] [CrossRef]
- Cao, D.; Sun, P.; Bhowmick, S.; Wei, Y.; Guo, B.; Wei, Y.; Mur, L.A.J.; Sun, Z. Secondary metabolites of endophytic fungi isolated from Huperzia serrata. Fitoterapia 2021, 155, 104970. [Google Scholar] [CrossRef]
- Zheng, Y.K.; Miao, C.P.; Chen, H.H.; Huang, F.F.; Xia, Y.M.; Chen, Y.W.; Zhao, L.X. Endophytic fungi harbored in Panax notoginseng: Diversity and potential as biological control agents against host plant pathogens of root-rot disease. J. Ginseng Res. 2017, 41, 353–360. [Google Scholar] [CrossRef]
- Cheng, X.; Wei, Z.; Pu, S.; Xiang, M.; Yan, A.; Zhang, Y.; Wang, X. Diversity of endophytic fungi of Paeonia lactiflora Pallas and screening for fungal paeoniflorin producers. FEMS Microbiol. Lett. 2018, 365, fny263. [Google Scholar] [CrossRef]
- Li, L.; Li, Y.Q.; Fu, Y.S.; Zhang, H.; Alkhalifah, D.H.M.; Salam, N.; Hozzein, W.N.; Asem, M.D.; Li, W.J. Nesterenkonia endophytica sp. nov. isolated from roots of Glycyrrhiza uralensis. Int. J. Syst. Evol. Microbiol. 2018, 68, 2659–2663. [Google Scholar] [CrossRef]
- Ju, M.; Zhang, Q.; Wang, R.; Yan, S.; Li, Z.; Li, P.; Gu, P. Correlation in endophytic fungi community diversity and bioactive compounds of Sophora alopecuroides. Front. Microbiol. 2022, 13, 955647. [Google Scholar] [CrossRef]
- Niu, L.; Rustamova, N.; Ning, H.; Paerhati, P.; Lu, C.; Yili, A. Diversity and biological activities of endophytic fungi from the flowers of the medicinal plant Vernonia anthelmintica. Int. J. Mol. Sci. 2022, 23, 11935. [Google Scholar] [CrossRef]
- de Paula Nogueira Cruz, F.; Ferreira de Paula, A.; Nogueira, C.T.; Marques de Andrade, P.H.; Borges, L.M.; Lacava, P.T.; Baratella da Cunha Camargo, I.L.; Anibal, F.F.; Paiva de Sousa, C. Discovery of a novel lineage Burkholderia cepacia ST 1870 endophytically isolated from medicinal Polygala paniculata which shows potent in vitro antileishmanial and antimicrobial effects. Int. J. Microbiol. 2021, 2021, 6618559. [Google Scholar] [CrossRef]
- Pawle, G.; Singh, S.K. Antioxidant potential of endophytic fungus Colletotrichum species isolated from Polygala elongata. Int. J. Pharm. Biol. Sci. 2014, 5, B313–B319. [Google Scholar]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A. Endophytic fungal communities and their biotechnological implications for agro-environmental sustainability. Folia Microbiol. 2022, 67, 203–232. [Google Scholar] [CrossRef]
- Parmar, S.; Li, Q.; Wu, Y.; Li, X.; Yan, J.; Sharma, V.K.; Wei, Y.; Li, H. Endophytic fungal community of Dysphania ambrosioides from two heavy metal-contaminated sites: Evaluated by culture-dependent and culture-independent approaches. Microb. Biotechnol. 2018, 11, 1170–1183. [Google Scholar] [CrossRef]
- Dos Reis, J.B.A.; Lorenzi, A.S.; do Vale, H.M.M. Methods used for the study of endophytic fungi: A review on methodologies and challenges, and associated tips. Arch. Microbiol. 2022, 204, 675. [Google Scholar] [CrossRef]
- Kohout, P.; Sykorova, Z.; Ctvrtlikova, M.; Rydlova, J.; Suda, J.; Vohnik, M.; Sudova, R. Surprising spectra of root-associated fungi in submerged aquatic plants. FEMS Microbiol. Ecol. 2012, 80, 216–235. [Google Scholar] [CrossRef]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Shi, D.; Ren, L. Diversity and antimicrobial activity of endophytic fungi isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef]
- Duan, X.; Xu, F.; Qin, D.; Gao, T.; Shen, W.; Zuo, S.; Yu, B.; Xu, J.; Peng, Y.; Dong, J. Diversity and bioactivities of fungal endophytes from Distylium chinense, a rare waterlogging tolerant plant endemic to the Three Gorges Reservoir. BMC Microbiol. 2019, 19, 278. [Google Scholar] [CrossRef]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef]
- Park, Y.H.; Kim, Y.C.; Park, S.U.; Lim, H.S.; Kim, J.B.; Cho, B.K.; Bae, H. Age-dependent distribution of fungal endophytes in Panax ginseng Roots cultivated in Korea. J. Ginseng Res. 2012, 36, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Chen, D.; Wang, Y.P.; Ehmet, N.; Ma, J.; Sun, K. Analysis of endophyte diversity of Gentiana officinalis among different tissue types and ages and their association with four medicinal secondary metabolites. PeerJ 2022, 10, e13949. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, Z.; Song, L.; Fu, W.; Liu, L. Anti-Alzheimer’s natural products derived from plant endophytic fungi. Molecules 2023, 28, 2259. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | PL1 | PS1 | PR1 | PL2 | PS2 | PR2 |
|---|---|---|---|---|---|---|
| shannon | 2.191 ± 0.63 b | 2.11 ± 0.91 b | 1.71 ± 0.61 b | 1.51 ± 0.71 b | 3.61 ± 0.31 a | 1.91 ± 0.51 b |
| simpson | 0.266 ± 0.16 a | 0.31 ± 0.21 a | 0.41 ± 0.21 a | 0.51 ± 0.31 a | 0.11 ± 0.01 a | 0.31 ± 0.21 a |
| chao | 108.7 ± 42.46 bc | 99.71 ± 15.61 bc | 62.11 ± 14.71 c | 153.41 ± 65.91 b | 474.61 ± 36.41 a | 79.71 ± 26.41 bc |
| ace | 112.7 ± 40.18 bc | 99.11 ± 16.41 bc | 63.51 ± 14.41 c | 154.01 ± 69.01 b | 473.91 ± 35.01 a | 83.71 ± 22.21 bc |
| coverage | 0.9998 ± 0.00004 | 0.9998 ± 0.00004 | 0.9999 ± 0.0001 | 0.9997 ± 0.00003 | 0.9999 ± 0.0001 | 0.9999 ± 0.00005 |
| Group Label | Number of Common Genera | Top 5 Genera and Their Relative Abundance |
|---|---|---|
| PL1:PS1:PR1 | 29 | Fusarium (19.14%) |
| Colletotrichum (17.31%) | ||
| Phoma (13.91%) | ||
| Plectosphaerella (4.47%) | ||
| Apiotrichum (3.35%) | ||
| PL2:PS2:PR2 | 27 | Apiotrichum (28.24%) |
| Fusarium (21.56%) | ||
| Calonectria (9.37%) | ||
| Papiliotrema (7.37%) | ||
| Acremonium (6.13%) | ||
| PL1:PL2 | 43 | Cercospora (34.57%) |
| Colletotrichum (24.54%) | ||
| Phoma (10.81%) | ||
| Phyllosticta (4.96%) | ||
| Diaporthe (2.09%) | ||
| PS1:PS2 | 65 | Colletotrichum (15.84%) |
| Phoma (10.71%) | ||
| Fusarium (6.82%) | ||
| Plectosphaerella (5.13%) | ||
| Cladophialophora (4.64%) | ||
| PR1:PR2 | 24 | Fusarium (43.62%) |
| Apiotrichum (26.24%) | ||
| Papiliotrema (3.12%) | ||
| Aspergillus (1.61%) | ||
| Cutaneotrichosporon (1.60%) |
| Source | Isolates | Closest Relatives in NCBI (Accession No.) | GenBank No. | Identity% | AChE Inhibition% |
|---|---|---|---|---|---|
| 1-year-old leaf | PCAM003 | Fusarium tricinctum (MK962346) | OR592306 | 100.00 | NA |
| PCAM010 | Phialophora mustea (MK102700) | OR592307 | 99.83 | 54.02 ± 1.42 | |
| PCAM033 | Didymella bellidis (MN274965) | OR592308 | 100.00 | NA | |
| PCAM034 | Phialophora sp. (MT576439) | OR592309 | 100.00 | 22.87 ± 3.16 | |
| PCAM073 | Pezicula chiangraiensis (KU310621) | OR592310 | 100.00 | 14.10 ± 4.01 | |
| PCAM074 | Fusarium tricinctum (MK962346) | OR592311 | 100.00 | NA | |
| PCAM078 | Didymella bellidis (MN274965) | OR592312 | 100.00 | NA | |
| PCAM087 | Talaromyces funiculosus (MT367866) | OR592313 | 100.00 | NA | |
| PCAM091 | Didymella bellidis (MN274965) | OR592314 | 100.00 | NA | |
| PCAM095 | Coniochaeta velutina (MK656231) | OR592315 | 100.00 | NA | |
| 1-year-old stem | PCAP001 | Phialophora mustea (MK102700) | OR592316 | 100.00 | NA |
| PCAP003 | Fusarium reticulatum (MT601889) | OR592317 | 99.81 | 11.05 ± 0.40 | |
| PCAP012 | Geotrichum candidum (KT921188) | OR592318 | 100.00 | NA | |
| PCAP033 | Fusarium flocciferum (MG386078) | OR592319 | 100.00 | NA | |
| PCAP036 | Juxtiphoma sp. (MK100167) | OR592320 | 100.00 | 23.28 ± 0.99 | |
| PCAP041 | Alternaria alternata (OQ248210) | OR592321 | 100.00 | 19.37 ± 0.51 | |
| PCAP093 | Pezicula sporulosa (MW487233) | OR592322 | 99.06 | NA | |
| PCAP096 | Fusarium reticulatum (MT601889) | OR592323 | 99.81 | 14.54 ± 1.95 | |
| PCAR020 | Coniochaeta velutina (MK656231) | OR592324 | 100.00 | NA | |
| 1-year-old root | PCBM001 | Botrytis cinerea (KR002909) | OR592325 | 99.62 | NA |
| PCBM022 | Colletotrichum dematium (MT446146) | OR592326 | 100.00 | NA | |
| PCBM027 | Diaporthe nobilis (KX866924) | OR592327 | 98.91 | 65.92 ± 2.29 | |
| PCBM035 | Botrytis cinerea (MT573470) | OR592328 | 100.00 | NA | |
| PCBP027 | Talaromyces muroii (KU744629) | OR592329 | 100.00 | NA | |
| PCBR013 | Colletotrichum acutatum (MN856423) | OR592330 | 100.00 | NA | |
| PCBR020 | Cladosporium sp. (OQ248224) | OR592331 | 100.00 | NA | |
| 2-year-old leaf | LM17 | Alternaria alternata (OW986459) | OR597909 | 100.00 | 24.03 ± 0.78 |
| LM35 | Leptosphaerulina arachidicola (MK555325) | OR597910 | 100.00 | NA | |
| LM80 | Alternaria blumeae (MN612548) | OR597911 | 100.00 | NA | |
| LP01 | Penicillium brefeldianum (MH864250) | OR597912 | 100.00 | NA | |
| LP24 | Colletotrichum horii (MT568591) | OR597913 | 100.00 | 11.97 ± 2.19 | |
| LP34 | Nigrospora chinensis (MK371770) | OR597914 | 100.00 | NA | |
| LP41 | Fusarium oxysporum (MT530243) | OR597915 | 100.00 | 59.13 ± 0.83 | |
| LP851 | Penicillium brefeldianum MH864250 | OR597916 | 100.00 | NA | |
| LP87 | Pyronema omphalodes (MK886722) | OR597917 | 100.00 | NA | |
| LR101 | Nigrospora rubi (NR_153470) | OR597918 | 100.00 | 44.73 ± 1.93 | |
| LR43 | Nigrospora chinensis (MK371770) | OR597919 | 100.00 | NA | |
| LR51 | Colletotrichum gloeosporioides (MT568599) | OR597920 | 100.00 | 13.96 ± 0.68 | |
| LR62 | Nigrospora oryzae (MW411591) | OR597921 | 100.00 | NA | |
| LR87 | Colletotrichum boninense (MT568597) | OR597922 | 100.00 | NA | |
| 2-year-old stem | SM04 | Schizophyllum commune (MK647986) | OR597926 | 100.00 | NA |
| SM21 | Nigrospora chinensis (MN341444) | OR597927 | 100.00 | NA | |
| SM40 | Nigrospora rubi (MN486553) | OR597928 | 100.00 | 45.77 ± 0.82 | |
| SM79 | Pestalotiopsis kenyana (MN341553) | OR597929 | 100.00 | NA | |
| SM81 | Phoma herbarum (MT420621) | OR597930 | 100.00 | 89.88 ± 2.79 | |
| SR58 | Chaetomium cochliodes (MT520580) | OR597931 | 100.00 | NA | |
| SR60 | Fusarium oxysporum (MT530243) | OR597932 | 100.00 | 57.88 ± 1.85 | |
| 2-year-old root | RM06 | Schizophyllum commune (MT601949) | OR597923 | 100.00 | NA |
| RM07 | Epicoccum nigrum (MN947593) | OR597924 | 100.00 | NA | |
| RR09 | Aspergillus sp. (LN898690) | OR597925 | 100.00 | NA | |
| FM25 | Colletotrichum horii (MT568591) | OR597904 | 100.00 | 10.80 ± 0.92 | |
| FM83 | Phyllosticta sorghina (MK762588) | OR597905 | 100.00 | NA | |
| FP76 | Diaporthe unshiuensis (MN816431) | OR597906 | 100.00 | 34.25 ± 2.09 | |
| FR89 | Alternaria alternata (OW986459) | OR597907 | 100.00 | 14.57 ± 1.22 | |
| FR92 | Periconia pseudobyssoides (MN944517) | OR597908 | 100.00 | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, K.; Xiong, Y.; Liu, Y.; Fan, X.; Zhu, R.; Hu, Z.; Li, C.; Hua, Y. Community Structure and Diversity of Endophytic Fungi in Cultivated Polygala crotalarioides at Two Different Growth Stages Based on Culture-Independent and Culture-Based Methods. J. Fungi 2024, 10, 195. https://doi.org/10.3390/jof10030195
Shen K, Xiong Y, Liu Y, Fan X, Zhu R, Hu Z, Li C, Hua Y. Community Structure and Diversity of Endophytic Fungi in Cultivated Polygala crotalarioides at Two Different Growth Stages Based on Culture-Independent and Culture-Based Methods. Journal of Fungi. 2024; 10(3):195. https://doi.org/10.3390/jof10030195
Chicago/Turabian StyleShen, Kaize, Yu Xiong, Yanfang Liu, Xingwang Fan, Rui Zhu, Zumao Hu, Congying Li, and Yan Hua. 2024. "Community Structure and Diversity of Endophytic Fungi in Cultivated Polygala crotalarioides at Two Different Growth Stages Based on Culture-Independent and Culture-Based Methods" Journal of Fungi 10, no. 3: 195. https://doi.org/10.3390/jof10030195