
Phylogenetic and Taxonomic Analyses of Five New Wood-Inhabiting Fungi of Botryobasidium, Coltricia and Coltriciella (Basidiomycota) from China
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Herbarium Specimen Preparation
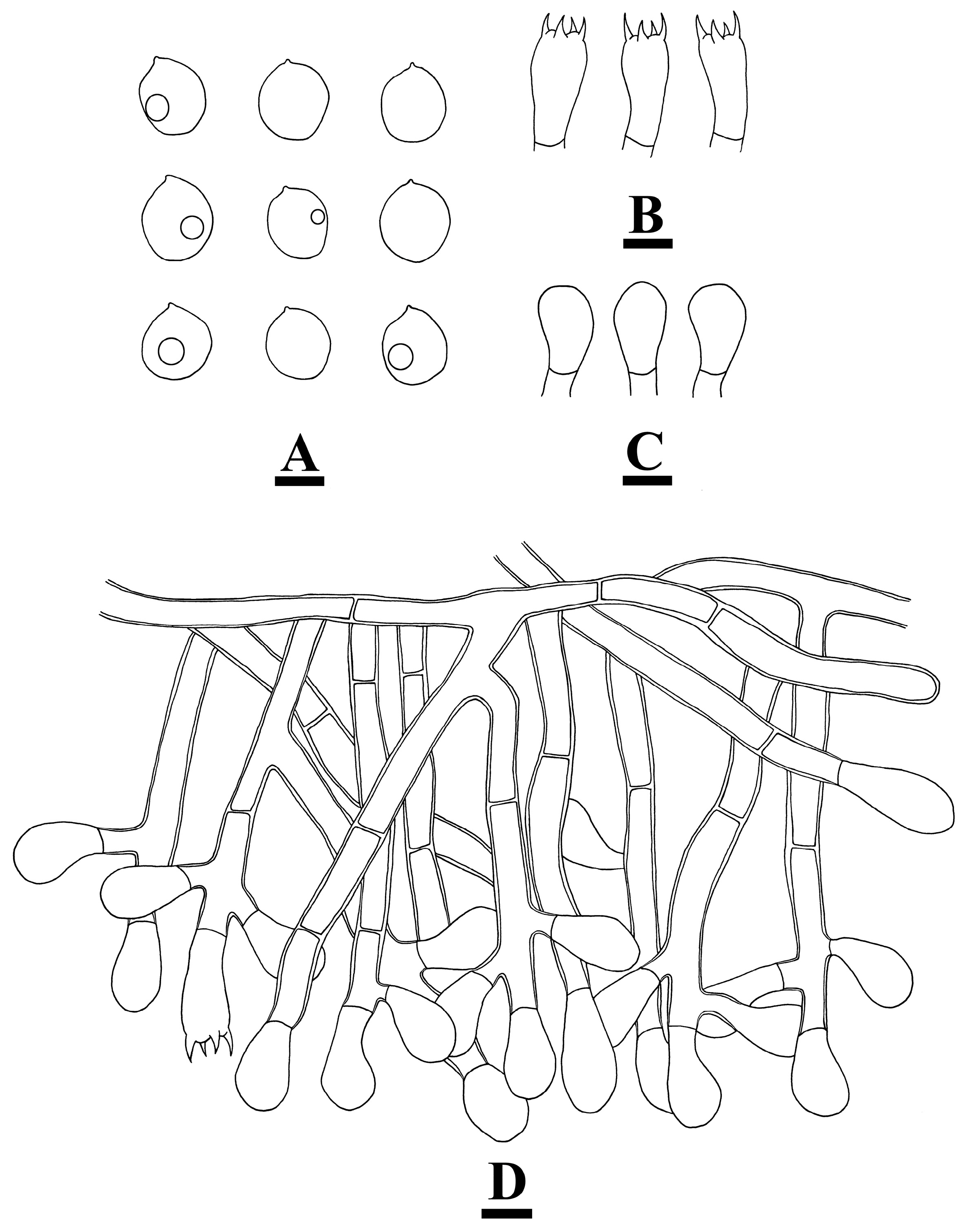
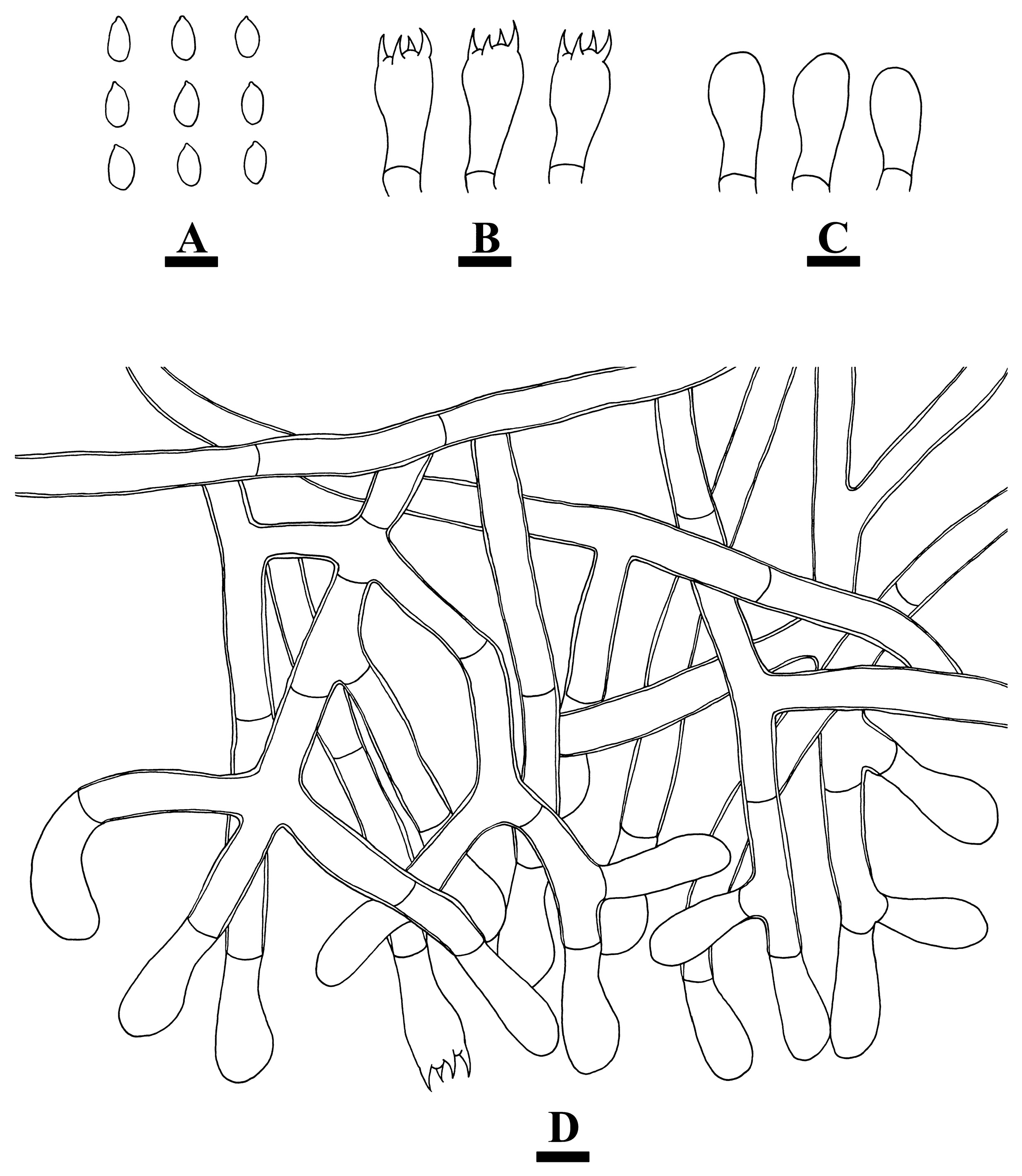
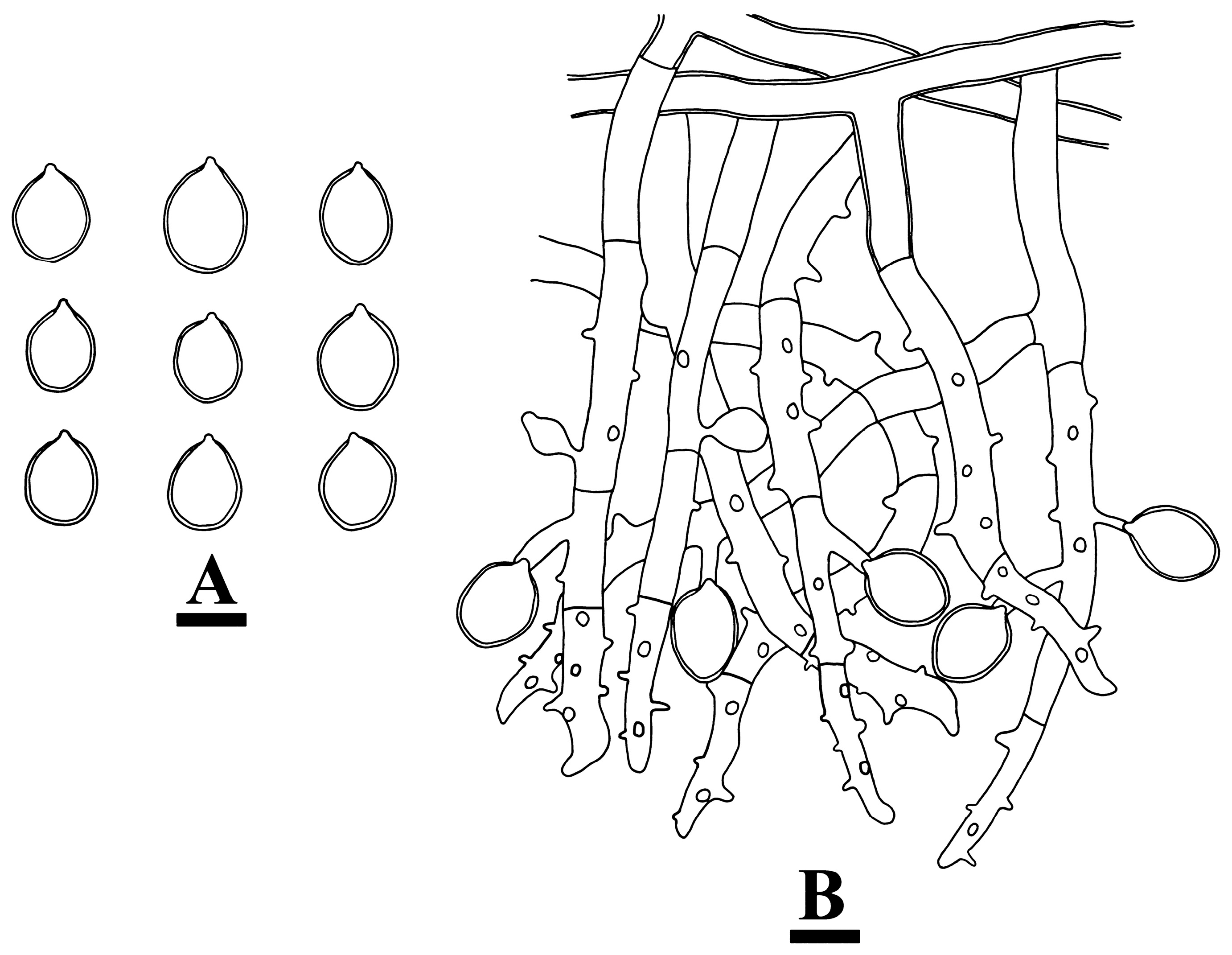
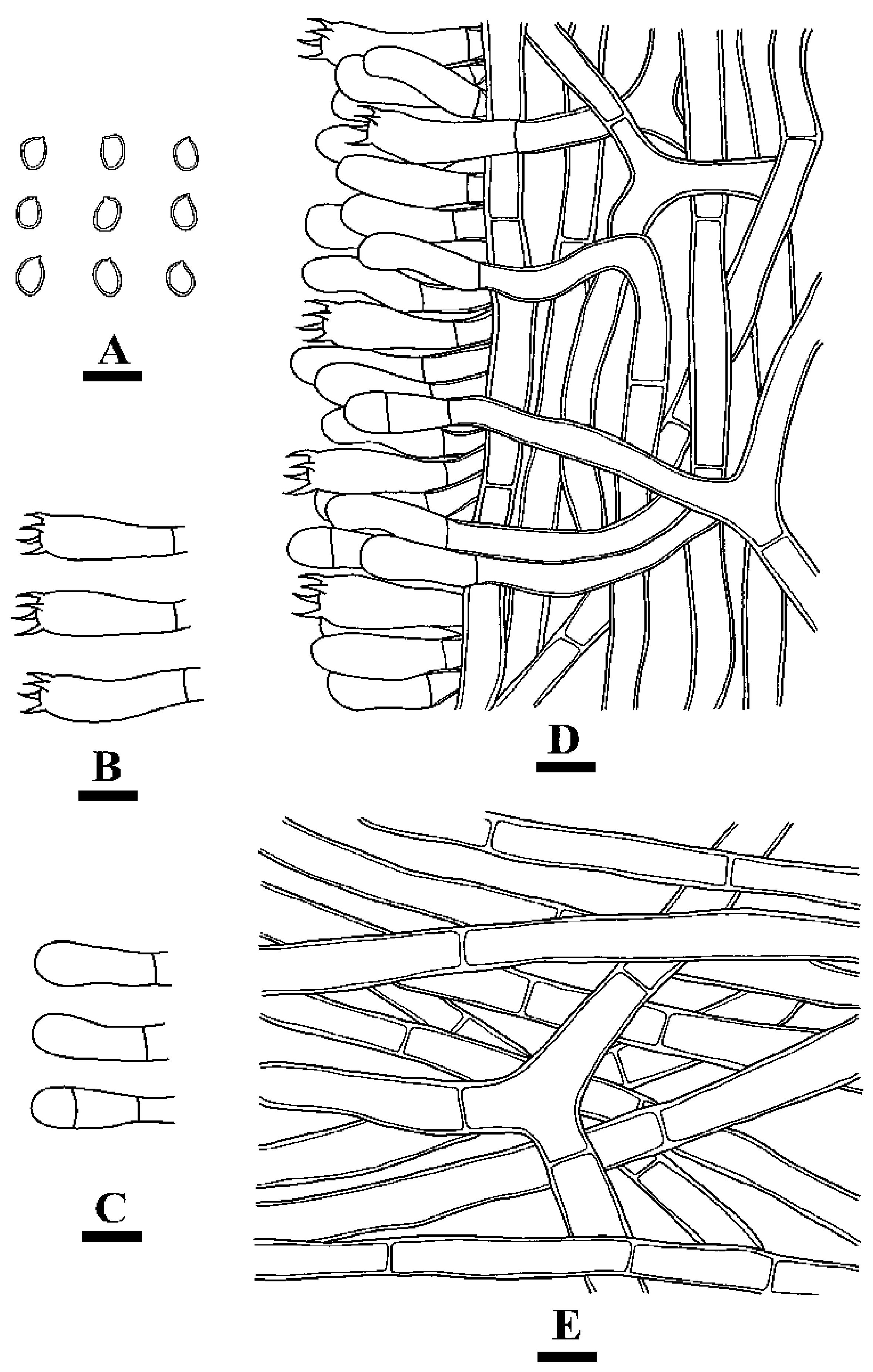
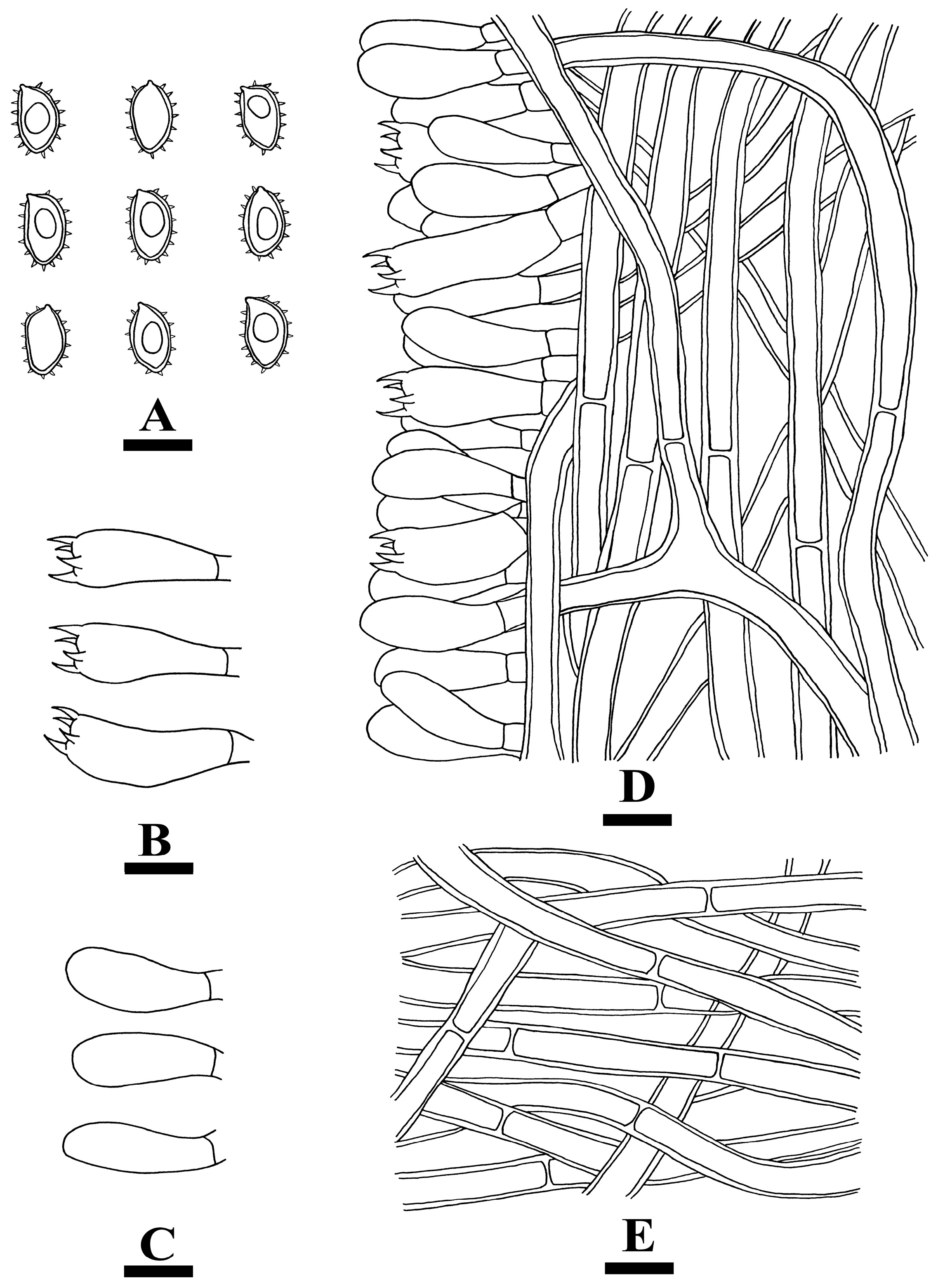
2.2. Morphology
2.3. DNA Extraction and Sequencing
2.4. Phylogenetic Analyses
3. Results
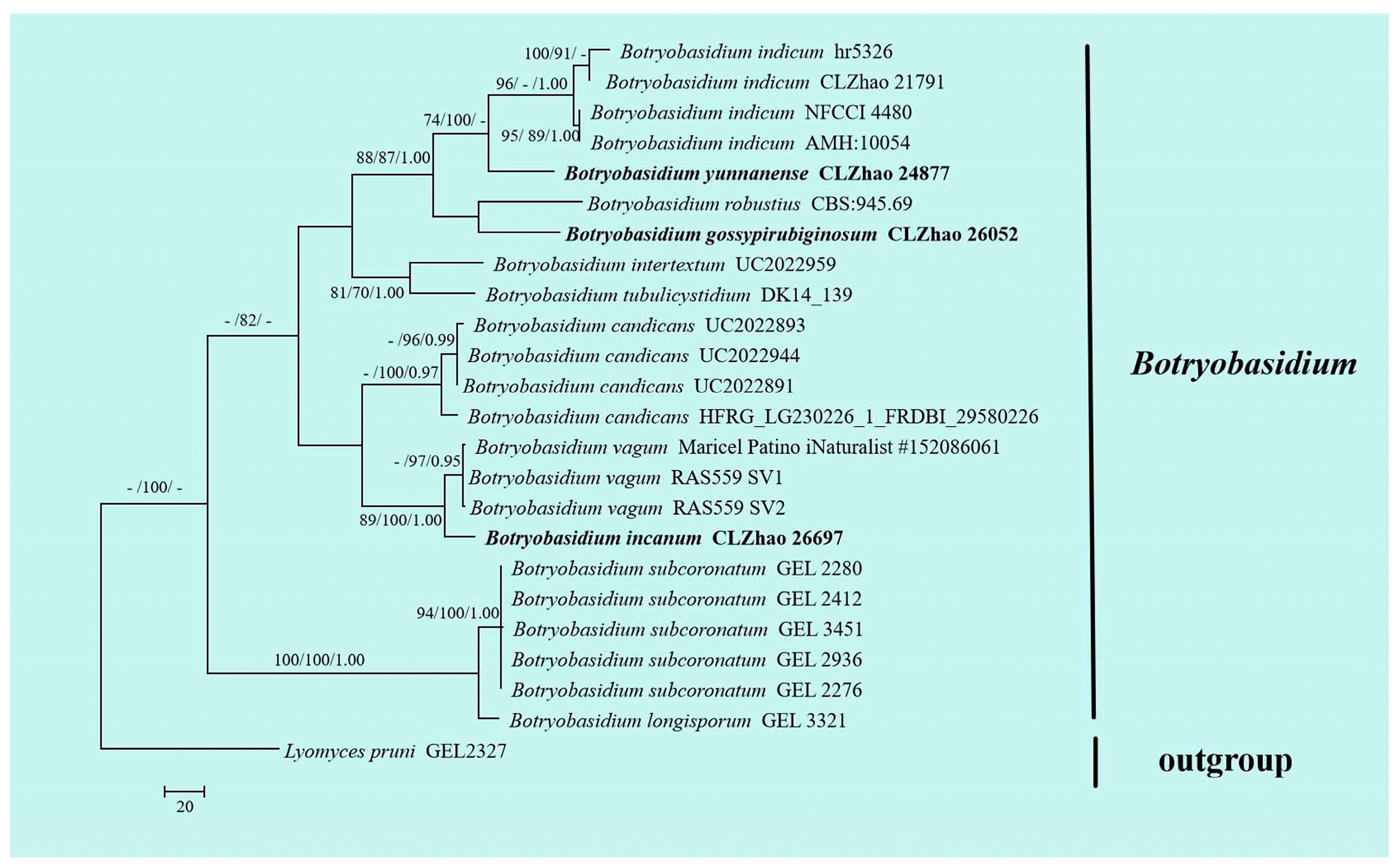
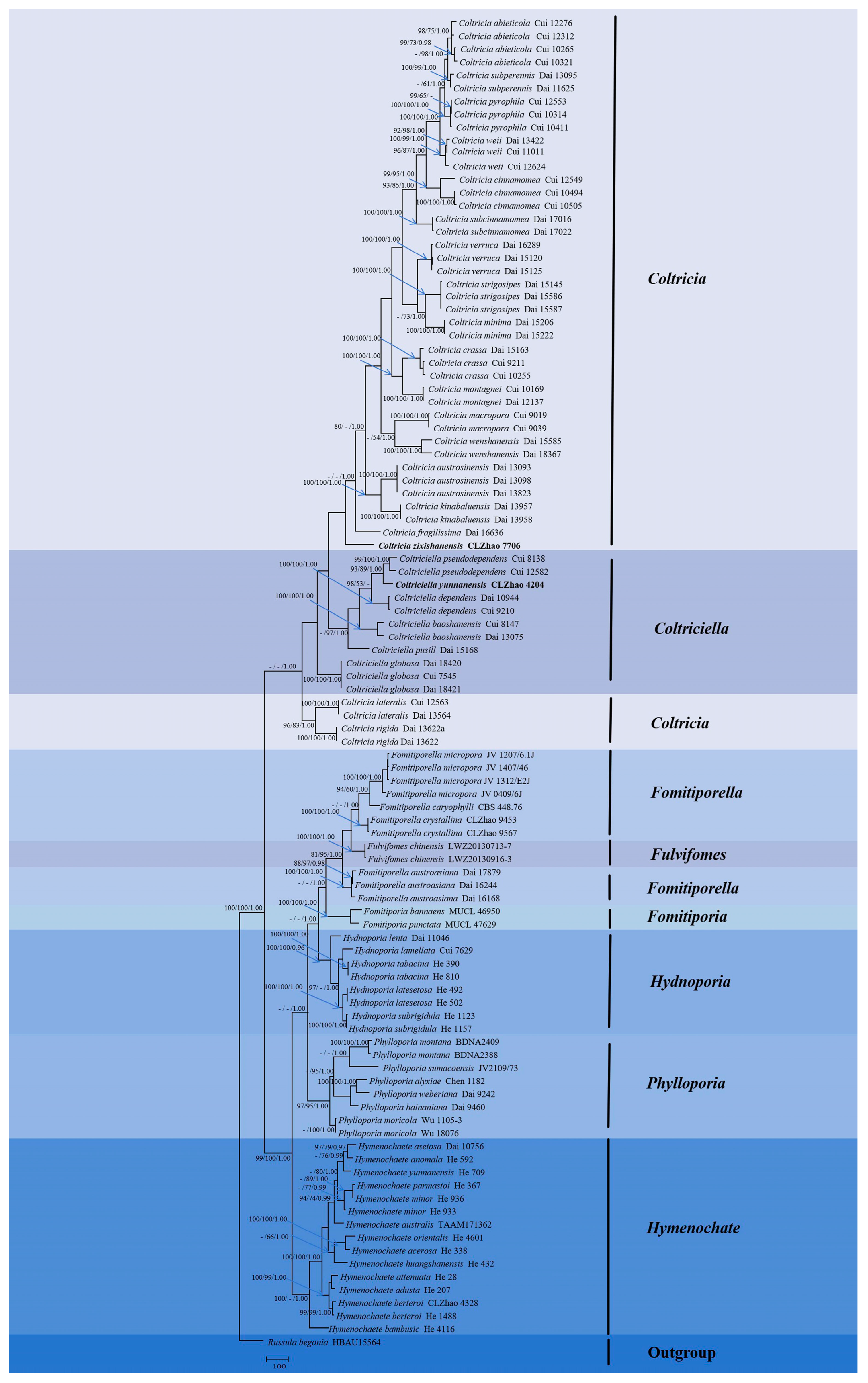
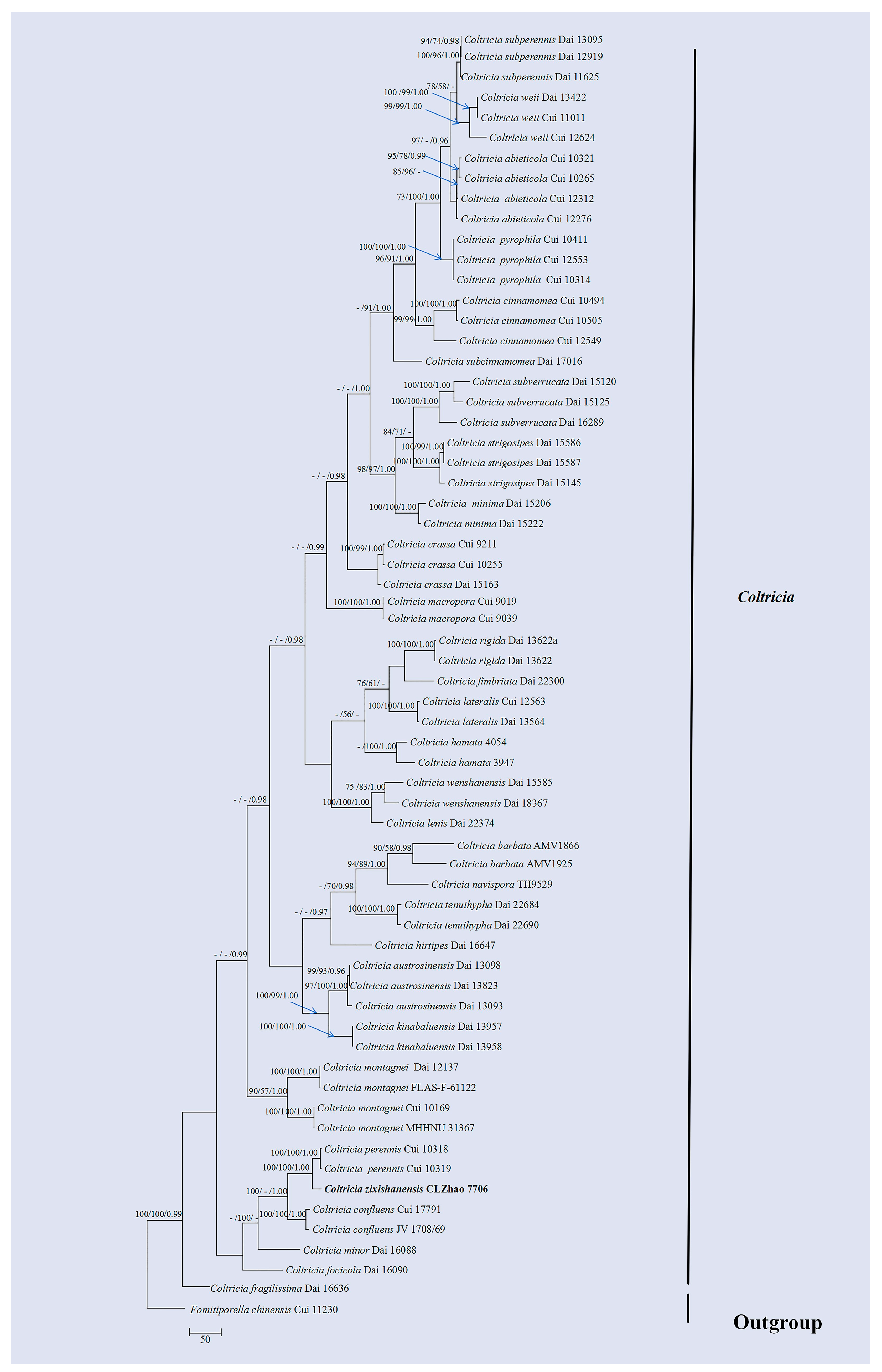
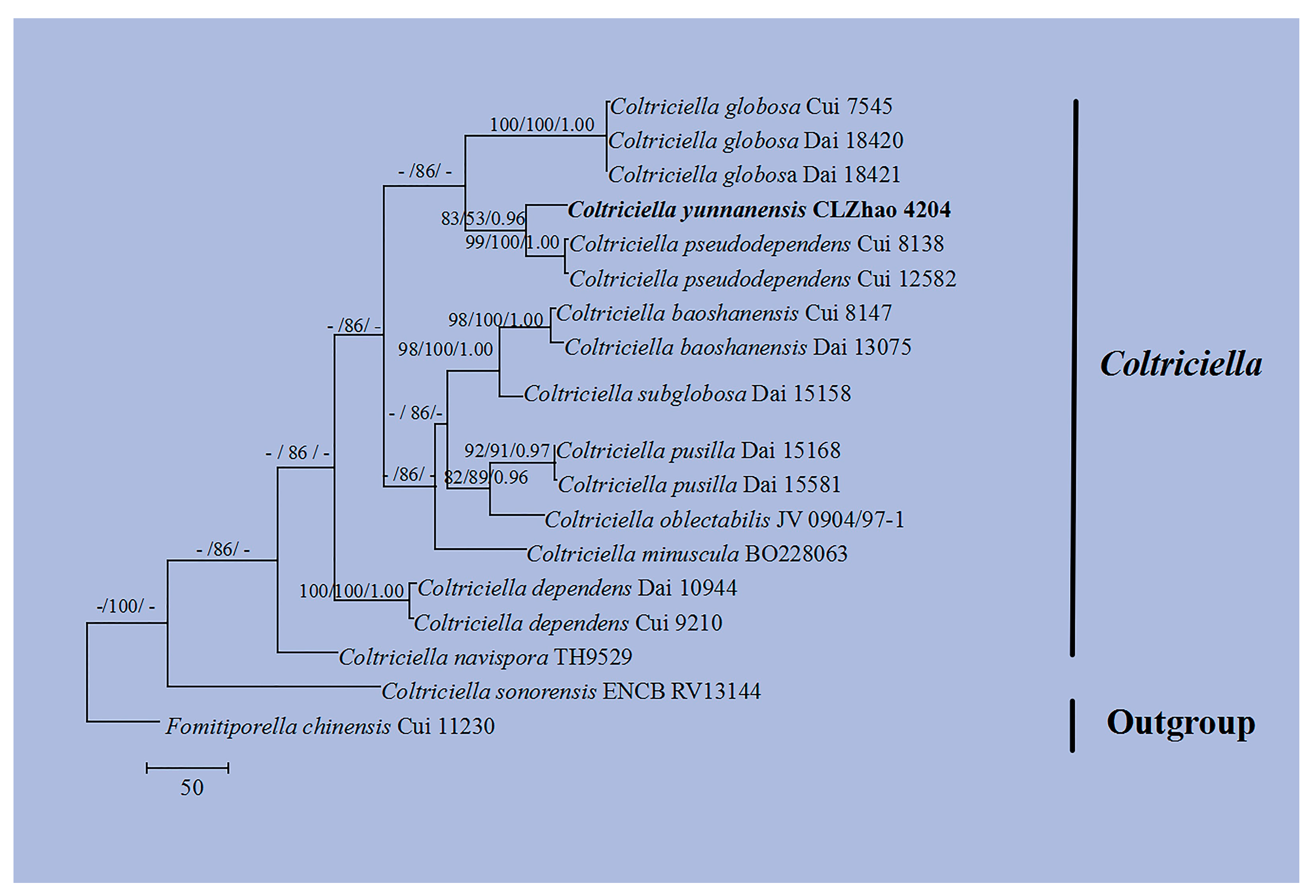
3.1. Molecular Phylogeny
3.2. Taxonomy










4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Gareth Jones, E.B.; Maharachchikumbura, S.S.N.; Raspe, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal divers notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Ponce, Á.; Salerni, E.; D’Aguanno, M.N.; Perini, C. Wood-decay fungi fructifying in mediterranean deciduous oak forests: A community composition, richness and productivity study. Forests 2023, 14, 1326. [Google Scholar] [CrossRef]
- Fukasawa, Y. Ecological impacts of fungal wood decay types: A review of current knowledge and future research directions. Ecol. Res. 2021, 36, 910–931. [Google Scholar] [CrossRef]
- Bujoczek, L.; Bujoczek, M. Factors influencing the diversity of deadwood, a crucial microhabitat for many rare and endangered saproxylic organisms. Ecol. Indic. 2022, 142, 109197. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjón, S.P. Fungi Europaei 12: Corticiaceae s.l.; Edizioni Candusso: Salamanca, Spain, 2010; pp. 1–1007. [Google Scholar]
- Bernicchia, A.; Langer, G.; Gorjón, S.P. Botryobasidium sassofratinoense sp. nov. (Cantharellales, Basidiomycota) from Italy. Mycotaxon 2010, 111, 403–409. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; Porter, T.M.; Margaritescu, S.; Weiss, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Langer, G.; Langer, E. Haplotrichum parmastii sp. nov. collected in Costa Rica. Folia Cryptogam. Est. 1998, 33, 63–69. [Google Scholar]
- Dai, Y.C.; Yuan, H.S.; Cui, B.K. Coltricia (Basidiomycota, Hymenochaetaceae) in China. Sydowia 2010, 62, 11–21. [Google Scholar]
- Valenzuela, R.; Raymundo, T.; Cifuentes, J.; Esqueda, M.; Amalfi, M.; Decock, C. Coltriciella sonorensis sp. nov. (Basidiomycota, Hymenochaetales) from Mexico: Evidence from morphology and DNA sequence data. Mycol. Prog. 2012, 11, 181–189. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, Y.C.; Wu, F.; Liu, X.Y.; Maurice, S.; Krutovsky, K.V.; Pavlov, I.N.; Lindner, D.L.; Martin, F.M.; Yuan, Y. Insights into the ecological diversification of the Hymenochaetales based on comparative genomics and phylogenomics with an emphasis on Coltricia. Genome Biol. Evol. 2023, 15, evad136. [Google Scholar] [CrossRef]
- Susan, D.; Retnowati, A.; Sukarno, N. Coltriciella minuscula sp. nov., a new species of poroid fungus on Pinus merkusii from an Indonesian tropical forest. Mycoscience 2017, 59, 49–53. [Google Scholar] [CrossRef]
- Ryvarden, L. Genera of polypores. Nomenclature and taxonomy. Synop. Fungorum 1991, 5, 1–363. [Google Scholar]
- Ryvarden, L. Neotropical polypores. Part 1: Introduction, Ganodermataceae & Hymenochaetaceae. Synop. Fungorum 2004, 19, 1–229. [Google Scholar]
- Wagner, T.; Fischer, M. Proceedings towards a natural classification of the worldwide taxa Phellinus s.l. and Inonotus s.l., and phylogenetic relationships of allied genera. Mycologia 2002, 94, 998–1016. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Suvi, T.; Beaver, K.; Saar, I. Ectomycorrhizas of Coltricia and Coltriciella (Hymenochaetales, Basidiomycota) on Caesalpiniaceae, Dipterocarpaceae and Myrtaceae in Seychelles. Mycol. Prog. 2007, 6, 101–107. [Google Scholar] [CrossRef]
- Larsson, K.H.; Parmasto, E.; Fischer, M.; Langer, E.; Nakasone, K.K.; Redhead, S.A. Hymenochaetales: A molecular phylogeny for the hymenochaetoid clade. Mycologia 2006, 98, 926–936. [Google Scholar] [CrossRef]
- Bian, L.S.; Dai, Y.C. Molecular phylogeny and morphology reveal two new species of Coltricia (Hymenochaetaceae Basidiomycota) from China. Mycol. Prog. 2020, 19, 657–666. [Google Scholar] [CrossRef]
- Murrill, W.A. “The Polyporaceae of North America-VII. The Genera Hexagona, Grifola, Romellia, Coltricia and Coltriciella”. Bull. Torrey Bot. 1904, 31, 325–348. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. The Ganish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996; pp. 1–6. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Fukami, T.; Dickie, I.A.; Paula Wilkie, J.; Paulus, B.C.; Park, D.; Roberts, A.; Buchanan, P.K.; Allen, R.B. Assembly history dictates ecosystem functioning: Evidence from wood decomposer communities. Ecol. Lett. 2010, 13, 675–684. [Google Scholar] [CrossRef]
- Bian, L.S.; Dai, Y.C. Morphological and molecular evidence for three new species of Coltricia (Hymenochaetaceae, Basidiomycota) from southern China. Mycologia 2017, 109, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wu, F.; Dai, Y.C.; Vlasák, J. Two new species of Phylloporia (Hymenochaetales) from the Neotropics. MycoKeys 2022, 90, 71–83. [Google Scholar] [CrossRef]
- Bian, L.S.; Wu, F.; Dai, Y.-C. Two new species of Coltricia (Hymenochaetales) from southern China based on morphological and molecular characters. Mycol. Prog. 2016, 15, 15–27. [Google Scholar] [CrossRef]
- Ji, H.X.; Fan, L. Fomitiporella crystallina sp. nov. (Basidiomycota, Hymenochaetales) from China. BDJ 2023, 11, e95945. [Google Scholar] [CrossRef]
- Bian, L.S.; Zhou, M.; Yu, J. Three new Coltricia (Hymenochaetaceae, Basidiomycota) species from China based on morphological characters and molecular evidence. Mycol. Prog. 2022, 21, 45. [Google Scholar] [CrossRef]
- He, S.H.; Dai, Y.C. Taxonomy and phylogeny of Hymenochaete and allied genera of Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 2012, 56, 77–93. [Google Scholar] [CrossRef]
- Spirin, V.; Runnel, K.; Poldmaa, K. Studies in the bark-dwelling species of Hymenochaete (Hymenochaetales, Basidiomycota) reveal three new species. Mycologie 2015, 36, 167–176. [Google Scholar] [CrossRef]
- Nie, T.; Tian, Y.; Liu, S.L.; Yang, J.; He, S.H. Species of Hymenochaete (Hymenochaetales, Basidiomycota) on bamboos from East Asia, with descriptions of two new species. MycoKeys 2017, 20, 51–65. [Google Scholar] [CrossRef]
- Li, G.J.; Deng, C.Y.; Wei, T.Z.; Li, S.M. Four new species of Russula subsection sardoninae from China. J. Fungi 2023, 9, 199. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinf. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4. 0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling High-Impact Science for Phylogenetics Researchers with Limited Resources. In Proceedings of the XSEDE’12: 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the eXtreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; Volume 39, pp. 1–8. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Upp-sala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Dai, Y.C.; Li, H.J. Type studies on Coltricia and Coltriciella described by E. J. H. Corner from Southeast Asia. Mycoscience 2012, 53, 337–346. [Google Scholar] [CrossRef]
- Valenzuela, R.; Raymundo, T.; Decock, C.; Lara Chávez, M.B.N.; Luna-Vega, I.; Garcia-Sandoval, R. Coltriciella multipileata (Agaricomycetes, Hymenochaetaceae), a new species from Mexico, related to ectomycorrhizal lineages. Phytotaxa 2020, 475, 79–90. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal divers notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–24. [Google Scholar] [CrossRef]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martinez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef]
- Bian, L.S.; Dai, Y.C. Coltriciella globosa and C. pseudodependens spp. nov. (Hymenochaetales) from southern China based on morphological and molecular characters. Mycoscience 2015, 56, 190–197. [Google Scholar] [CrossRef]
- Dai, Y.C.; Wei, Y.L.; Zhou, L.W. Polypore richness along an elevational gradient: A case study in Changbaishan Nature Reserve, Northeastern China. Fungal Ecol. 2015, 13, 226–228. [Google Scholar] [CrossRef]
- Bai, M.; Zhou, L.W.; Tong, Y.J.; Yin, F. Risk assessment and warning system for strategic biological resources in China. Innov. Life 2023, 1, 100004. [Google Scholar] [CrossRef]
- Liu, X.Y.; Liu, S.L.; Wei, H.W.; Wang, X.W.; Yu, J.; Shen, S.; Zhou, L.W. Preliminary species diversity and community phylogenetics of wood-inhabiting basidiomycetous fungi in the Dabie Mountains, Central China reveal unexpected richness. IMA Fungus 2023, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, Q.Q.; Li, Q.; Yang, J.W.; Cha, L.J.; Cheng, L.; Yang, S.Q.; Zhao, C.L.; Zhou, H.M. Molecular systematics and taxonomic analyses of three new wood-inhabiting fungi of Hyphoderma (Hyphodermataceae, Basidiomycota). J. Fungi 2023, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.X.; Zhao, T.J.; Zhao, C.L. Morphological characters and phylogenetic analyses reveal two new species of Peniophorella from Southern China. Mycol. Prog. 2020, 19, 397–404. [Google Scholar] [CrossRef]
- Wang, D.Q.; Zhao, C.L. Morphological and phylogenetic evidence for recognition of two new species of Phanerochaete from East Asia. J. Fungi 2021, 7, 1063. [Google Scholar] [CrossRef]
- Zhao, C.L.; Qu, M.H.; Huang, R.X.; Karunarathna, S.C. Multi-Gene Phylogeny and taxonomy of the wood-rotting fungal genus Phlebia sensu lato (Polyporales, Basidiomycota). J. Fungi 2023, 9, 320. [Google Scholar] [CrossRef]
- Shen, S.; Liu, S.L.; Zhou, L.W. Taxonomy of Hyphodermella: A case study to show that simple phylogenies cannot always accurately place species in appropriate genera. IMA Fungus 2023, 14, 11. [Google Scholar] [CrossRef]
- Ma, X.; Huang, R.X.; Zhang, Y.; Zhao, C.L. Hyphoderma fissuratum and H. mopanshanense spp. nov. (Polyporales) from southern China. Mycoscience 2021, 62, 36–41. [Google Scholar] [CrossRef]
- Guan, Q.X.; Li, Y.F.; Zhao, C.L. Morphological and phylogenetic evidence for recognition of two new species of Hyphoderma (Basidiomycota) from southern China, with a key to all Chinese Hyphoderma. MycoKeys 2021, 83, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.Y.; Guan, Q.X.; Luo, K.Y.; Zhao, C.L. Morphological and molecular identification of three new resupinate species of Hyphoderma (Hyphodermataceae, Agaricomycetes) from East Asia. Phytotaxa 2023, 599, 1–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Sample No. | GenBank Accession No. | References | |
|---|---|---|---|---|
| ITS | nLSU | |||
| Botrybasidium candicans | UC2022891 | KP814227 | [23] | |
| Botrybasidium candicans | UC2022944 | KP814546 | [23] | |
| Botrybasidium candicans | HFRC_LG230226_1_FRDBI_29580226 | OR896129 | Unpublished | |
| Botrybasidium candicans | UC2022893 | KP814200 | [23] | |
| Botrybasidium gossypirubiginosum | CLZhao 26052 | OR668924 | OR708665 | Present study |
| Botrybasidium incanum | CLZhao 26697 | OR668923 | OR708664 | Present study |
| Botrybasidium indicum | NFCCI 4480 | NR171230 | Unpublished | |
| Botrybasidium indicum | AMH:10054 | MK391496 | MK391493 | Unpublished |
| Botrybasidium indicum | hr5326 | OP806032 | Unpublished | |
| Botrybasidium indicum | CLZhao 21791 | ON406471 | Unpublished | |
| Botrybasidium intertextum | UC2022959 | KP814540 | Unpublished | |
| Botrybasidium longisporum | GEL 3321 | AJ389797 | Unpublished | |
| Botrybasidium robustius | CBS:945.69 | MH859491 | MH871272 | [24] |
| Botrybasidium subcoronatum | GEL 2276 | AJ389807 | Unpublished | |
| Botrybasidium subcoronatum | GEL 2280 | AJ389806 | Unpublished | |
| Botrybasidium subcoronatum | GEL 2412 | AJ389788 | [24] | |
| Botrybasidium subcoronatum | GEL 3451 | AJ389796 | [24] | |
| Botrybasidium subcoronatum | GEL 2936 | AJ389809 | [24] | |
| Botrybasidium tubulicystidium | DK14139 | OL436769 | Unpublished | |
| Botrybasidium vagum | RAS559 SV1 | OR471091 | [18] | |
| Botrybasidium vagum | RAS559 SV2 | OR471092 | Unpublished | |
| Botrybasidium vagum | Maricel Patino iNaturalist#152086061 | OR680661 | Unpublished | |
| Botrybasidium yunnanense | CLZhao 24877 | OR668925 | OR708666 | Present study |
| Coltricia abieticola | Cui 10265 | KX364784 | KX364803 | [25] |
| Coltricia abieticola | Cui 10321 | KX364785 | KX364804 | [25] |
| Coltricia abieticola | Cui 12276 | KU360673 | KU360643 | [25] |
| Coltricia abieticola | Cui 12312 | KU360674 | KU360644 | [25] |
| Coltricia austrosinensis | Dai 13093 | KU360670 | KU360640 | [25] |
| Coltricia austrosinensis | Dai 13098 | KU360671 | KU360640 | [25] |
| Coltricia austrosinensis | Dai 13823 | KU360672 | KU360642 | [25] |
| Coltricia barbata | AMV1866 | KT724137 | Unpublished | |
| Coltricia barbata | AMV1925 | KT724136 | Unpublished | |
| Coltricia cinnamomea | Cui 10494 | KU360675 | KJ000217 | [25] |
| Coltricia cinnamomea | Cui 10505 | KU360676 | KU360645 | [25] |
| Coltricia cinnamomea | Cui 12549 | KY693728 | KY693742 | [25] |
| Coltricia crassa | Cui 9211 | KU360677 | KU360646 | [25] |
| Coltricia crassa | Cui 10255 | KU360678 | KU360647 | [25] |
| Coltricia crassa | Dai 15163 | KU360679 | KU360648 | [25] |
| Coltricia confluens | Cui 17791 | ON567327 | Unpublished | |
| Coltricia confluens | JV 1708/69 | ON567325 | Unpublished | |
| Coltricia fimbriata | Dai 22300 | NR182965 | [26] | |
| Coltricia fragilissima | Dai 16636 | KY693733 | KY693749 | [25] |
| Coltricia focicola | Dai 16090 | KX364786 | [26] | |
| Coltricia hamata | 4054 | MZ484546 | [27] | |
| Coltricia hamata | 3947 | MZ484545 | [27] | |
| Coltricia hirtipes | Dai 16647 | KY693734 | KU360649 | [25] |
| Coltricia kinabaluensis | Dai 13957 | KX364787 | KX364806 | [25] |
| Coltricia kinabaluensis | Dai 13958 | KX364788 | KX364807 | [25] |
| Coltricia lateralis | Cui 12563 | KX364789 | KX364808 | [25] |
| Coltricia lateralis | Dai 13564 | KX364790 | KX364809 | [25] |
| Coltricia lenis | Dai 22374 | OL691609 | KJ000220 | [26] |
| Coltricia macropora | Cui 9019 | KU360680 | KJ000221 | [25] |
| Coltricia macropora | Cui 9039 | KU360681 | KU360649 | [25] |
| Coltricia minima | Dai 15206 | KU360682 | KU360650 | [25] |
| Coltricia minima | Dai 15222 | KU360683 | KJ000220 | [25] |
| Coltricia minor | Dai 16088 | KU360684 | [28] | |
| Coltricia montagnei | Cui 10169 | KU360685 | KU360652 | [25] |
| Coltricia montagnei | Dai 12137 | KX364810 | [25] | |
| Coltricia montagnei | MHHNU 31367 | MK182316 | Unpublished | |
| Coltricia montagnei | FLAS-F-61122 | MH399864 | Unpublished | |
| Coltricia navispora | TH9529 | KT339262 | Unpublished | |
| Coltricia perennis | Cui 10318 | KU360686 | KU360650 | [25] |
| Coltricia perennis | Cui 10319 | KU360687 | KU360652 | [25] |
| Coltricia perennis | Cui 10318 | KU360686 | [28] | |
| Coltricia perennis | Cui 10319 | KU360687 | [28] | |
| Coltricia pyrophila | Cui 10314 | KU360689 | KU360655 | [25] |
| Coltricia pyrophila | Cui 10411 | KU360690 | KU360656 | [25] |
| Coltricia pyrophila | Cui 12553 | KX364792 | KX364812 | [25] |
| Coltricia rigida | Dai 13622 | KX364793 | KX364813 | [25] |
| Coltricia rigida | Dai 13622a | KX364794 | KX364814 | [25] |
| Coltricia strigosipes | Dai 15145 | KX364795 | KX364815 | [25] |
| Coltricia strigosipes | Dai 15586 | KU360692 | KU360658 | [25] |
| Coltricia strigosipes | Dai 15587 | KU360693 | KU360659 | [25] |
| Coltricia subcinnamomea | Dai 17016 | KY693740 | KY693755 | [25] |
| Coltricia subcinnamomea | Dai 17022 | KY693756 | [25] | |
| Coltricia subperennis | Dai 11625 | KY693735 | KY693753 | [25] |
| Coltricia subperennis | Dai 13095 | KY693736 | KY693754 | [25] |
| Coltricia subperennis | Dai 12919 | MT174242 | [25] | |
| Coltricia tenuihypha | Dai 22684 | OL691610 | [26] | |
| Coltricia tenuihypha | Dai 22690 | OL691611 | [26] | |
| Coltricia verrucata | Dai 15120 | KU360694 | KU360660 | [25] |
| Coltricia verrucata | Dai 15125 | KU360695 | KU360661 | [25] |
| Coltricia verrucata | Dai 16289 | KU360696 | KU360662 | [25] |
| Coltricia weii | Cui 11011 | KU360698 | KU360664 | [25] |
| Coltricia weii | Cui 12624 | KX364796 | KX364816 | [25] |
| Coltricia weii | Dai 13422 | KX364797 | KX364817 | [25] |
| Coltricia wenshanensis | Dai 15585 | KX364798 | KX364818 | [25] |
| Coltricia wenshanensis | Dai 18367 | MT174244 | MT174237 | [25] |
| Coltricia zixishanensis | CLZhao 7706 | OR668922 | OR708662 | Present study |
| Coltriciella baoshanensis | Cui 8147 | KX364799 | KX364819 | [25] |
| Coltriciella baoshanensis | Dai 13075 | KX364800 | KX364820 | [25] |
| Coltriciella dependens | Dai 10944 | KY693737 | KY693757 | [25] |
| Coltriciella dependens | Cui 9210 | KY693738 | KY693758 | [25] |
| Coltriciella globosa | Cui 7545 | KJ540930 | KJ000226 | [25] |
| Coltriciella globosa | Dai 18420 | MT174245 | MT174238 | [25] |
| Coltriciella globosa | Dai 18421 | MT174246 | MT174239 | [25] |
| Coltriciella minuscula | BO228063 | KX086684 | Unpublished | |
| Coltriciella navispora | TH9529 | KT339262 | Unpublished | |
| Coltriciella oblectabilis | JV 0904/97-1 | ON567332 | Unpublished | |
| Coltriciella pseudodependens | Cui 8138 | KJ540931 | KJ000227 | [25] |
| Coltriciella pseudodependens | Cui 12582 | KX364801 | KX364821 | [25] |
| Coltriciella pusilla | Dai 15581 | KY693739 | [25] | |
| Coltriciella pusilla | Dai 15168 | KU360701 | KU36066 | [25] |
| Coltriciella sonorensis | ENCB RV13144 | HQ439179 | Unpublished | |
| Coltriciella subglobosa | Dai 15158 | KU360702 | [25] | |
| Coltriciella yunnanensis | CLZhao 4204 | OR668921 | OR708662 | Present study |
| Fomitiporella austroasiana | Dai 16244 | MG657328 | MG657320 | [29] |
| Fomitiporella austroasiana | Dai 16168 | MG657329 | MG657321 | [29] |
| Fomitiporella austroasiana | Dai 17879 | MG657330 | MG657324 | [29] |
| Fomitiporella caryophylli | CBS 448.76 | AY558611 | AY059021 | [29] |
| Fomitiporella chinensis | Cui 11230 | KX181309 | [30] | |
| Fomitiporella crystallina | CLZhao 9453 | ON493552 | ON493576 | [29] |
| Fomitiporella crystallina | CLZhao 9567 | ON493553 | ON493577 | [29] |
| Fomitiporella micropora | JV 1312/E2J | KX181294 | KX181333 | [29] |
| Fomitiporella micropora | JV 1407/46 | KX181295 | KX181332 | [29] |
| Fomitiporella micropora | JV 0409/6J | KX181296 | KX181331 | [29] |
| Fomitiporella micropora | JV 1207/6.1J | KX181297 | KX181330 | [29] |
| Fomitiporia bannaensis | MUCL 46950 | GU461943 | EF429218 | [31] |
| Fomitiporia punctata | MUCL 47629 | GU461950 | GU461982 | [31] |
| Fulvifomes chinensis | LWZ20130713-7 | KJ787817 | KJ787808 | [29] |
| Fulvifomes chinensis | LWZ20130916-3 | KJ787818 | KJ787809 | [29] |
| Hymenochaete acerosa | He 338 | JQ279543 | JQ279657 | [31] |
| Hymenochaete adusta | He 207 | JQ279523 | KU975497 | [31] |
| Hymenochaete anomala | He 592 | JQ279566 | JQ279650 | [31] |
| Hymenochaete asetosa | Dai 10756 | JQ279559 | JQ279642 | [31] |
| Hymenochaete attenuata | He 28 | JQ279526 | JQ279633 | [31] |
| Hymenochaete australis | TAAM171362 | KM017414 | [31] | |
| Hymenochaete bambusicola | He 4116 | KY425674 | NG060687 | [32] |
| Hymenochaete berteroi | CLZhao 4328 | OM959409 | OM967405 | [28] |
| Hymenochaete berteroi | He 1488 | KU975459 | KU975498 | [31] |
| Hymenochaete huangshanensis | He 432 | NR120041 | NG060638 | Unpublished |
| Hymenochaete minor | He 933 | NR120044 | JQ279654 | Unpublished |
| Hymenochaete minor | He 936 | JQ279556 | [31] | |
| Hymenochaete orientalis | He 4601 | KY425677 | NG060688 | [32] |
| Hymenochaete parmastoi | He 367 | NR120102 | [29] | |
| Hymenochaete yunnanensis | He 709 | JQ279571 | Unpublished | |
| Hydnoporia lamellata | Cui 7629 | JQ279603 | JQ279617 | [30] |
| Hydnoporia latesetosa | He 492 | JQ716404 | JQ716411 | [30] |
| Hydnoporia latesetosa | He 502 | JQ716405 | JQ716410 | [30] |
| Hydnoporia lenta | Dai 11046 | JQ279616 | JQ279628 | [30] |
| Hydnoporia subrigidula | He 1123 | JQ716402 | JQ716408 | [30] |
| Hydnoporia subrigidula | He 1157 | JQ716403 | JQ716409 | [30] |
| Hydnoporia tabacina | He 390 | JQ279610 | JQ279625 | [30] |
| Hydnoporia tabacina | He 810 | JQ279611 | JQ279626 | [30] |
| Lyomyces pruni | GEL2327 | DQ340312 | [31] | |
| Phylloporia alyxiae | Chen 1182 | LC528152 | LC514407 | [33] |
| Phylloporia hainaniana | Dai9460 | JF712928 | [27] | |
| Phylloporia montana | BDNA2409 | MH151177 | MG738811 | [27] |
| Phylloporia montana | BDNA2388 | MH151176 | MG738810 | [27] |
| Phylloporia moricola | Wu 1105-3 | LC514413 | [27] | |
| Phylloporia moricola | Wu 18076 | LC589619 | [27] | |
| Phylloporia sumacoensis | JV2109/73 | ON129552 | ON006468 | [27] |
| Phylloporia weberiana | Dai9242 | LC528151 | JF712936 | [27] |
| Russula begonia | HBAU15564 | MZ573252 | OQ077072 | [33] |
| Lyomyces pruni | GEL2327 | DQ340312 | [31] | |
| Phylloporia alyxiae | Chen 1182 | LC528152 | LC514407 | [33] |
| Phylloporia hainaniana | Dai9460 | JF712928 | [27] | |
| Phylloporia montana | BDNA2409 | MH151177 | MG738811 | [27] |
| Phylloporia montana | BDNA2388 | MH151176 | MG738810 | [27] |
| Phylloporia moricola | Wu 1105-3 | LC514413 | [27] | |
| Phylloporia moricola | Wu 18076 | LC589619 | [27] | |
| Phylloporia sumacoensis | JV2109/73 | ON129552 | ON006468 | [27] |
| Phylloporia weberiana | Dai9242 | LC528151 | JF712936 | [27] |
| Russula begonia | HBAU15564 | MZ573252 | OQ077072 | [33] |
| Lyomyces pruni | GEL2327 | DQ340312 | [31] | |
| Phylloporia alyxiae | Chen 1182 | LC528152 | LC514407 | [33] |
| Phylloporia hainaniana | Dai9460 | JF712928 | [27] | |
| Phylloporia montana | BDNA2409 | MH151177 | MG738811 | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Q.; Jiang, Q.; Yang, X.; Yang, J.; Zhao, C.; Zhao, J. Phylogenetic and Taxonomic Analyses of Five New Wood-Inhabiting Fungi of Botryobasidium, Coltricia and Coltriciella (Basidiomycota) from China. J. Fungi 2024, 10, 205. https://doi.org/10.3390/jof10030205
Zhou Q, Jiang Q, Yang X, Yang J, Zhao C, Zhao J. Phylogenetic and Taxonomic Analyses of Five New Wood-Inhabiting Fungi of Botryobasidium, Coltricia and Coltriciella (Basidiomycota) from China. Journal of Fungi. 2024; 10(3):205. https://doi.org/10.3390/jof10030205
Chicago/Turabian StyleZhou, Qian, Qianquan Jiang, Xin Yang, Jiawei Yang, Changlin Zhao, and Jian Zhao. 2024. "Phylogenetic and Taxonomic Analyses of Five New Wood-Inhabiting Fungi of Botryobasidium, Coltricia and Coltriciella (Basidiomycota) from China" Journal of Fungi 10, no. 3: 205. https://doi.org/10.3390/jof10030205