
Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China


1
School of Life Science and Technology, Center for Informational Biology, University of Electronic Science and Technology of China, Chengdu 611731, China
2
Institute of Crop Germplasm Resources, Guizhou Academy of Agricultural Sciences, Guiyang 550006, China
3
Guizhou Key Laboratory of Agricultural Biotechnology, Guizhou Academy of Agricultural Sciences, Guiyang 550006, China
4
Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(11), 893; https://doi.org/10.3390/jof7110893
Submission received: 26 August 2021
/
Revised: 11 October 2021
/
Accepted: 18 October 2021
/
Published: 22 October 2021
(This article belongs to the Special Issue Fungal Biodiversity and Ecology 2.0)
Abstract
:Botryosphaeriales is an important order of diverse fungal pathogens, saprobes, and endophytes distributed worldwide. Recent studies of Botryosphaeriales in China have discovered a broad range of species, some of which have not been formerly described. In this study, 60 saprobic isolates were obtained from decaying woody hosts in southwestern China. The isolates were compared with other species using morphological characteristics, and available DNA sequence data was used to infer phylogenetic analyses based on the internal transcribed spacer (ITS), large subunit rRNA gene (LSU), and translation elongation factor 1-α (tef) loci. Three novel species were illustrated and described as Botryobambusa guizhouensis, Sardiniella elliptica, and Sphaeropsis guizhouensis, which belong to rarely identified genera within Botryosphaeriaceae. Botryobambusa guizhouensis is the second species identified from the respective monotypic genus. The previously known species were identified as Aplosporella hesperidica, Barriopsis tectonae, Botryosphaeria dothidea, Diplodia mutila, Di. neojuniperi, Di. pseudoseriata, Di. sapinea, Di. seriata, Dothiorella sarmentorum, Do. yunnana, Lasiodiplodia pseudotheobromae, Neofusicoccum parvum, Sardiniella celtidis, Sa. guizhouensis, and Sphaeropsis citrigena. The results of this study indicate that numerous species of Botryosphaeriales are yet to be revealed in southwestern China.
1. Introduction
The Botryosphaeriales are among the most widespread, common, and important fungal pathogens of woody plants. Many are known to exist as endophytes in healthy plant tissues and also as saprobes in dead tree materials. This fungal order has gone through significant revisions, and several new families, genera, and species have been introduced or synonymized over the last decade, mainly on the basis of combined morphological and multiple gene sequence data [1,2,3,4,5,6,7,8,9]. Schoch et al. [10] introduced the order Botryosphaeriales to accommodate the single family Botryosphaeriaceae. In the “Outline of Ascomycetes” [7], nine families (Aplosporellaceae, Botryosphaeriaceae, Melanopsaceae, Phyllostictaceae, Planistromellaceae, Saccharataceae, Septorioideaceae, Endomelanconiopsidaceae, and Pseudofusicoccaceae) were recognized in Botryosphaeriales. Phillips et al. [3] revised the order and accepted Aplosporellaceae, Botryosphaeriaceae, Melanopsaceae, Phyllostictaceae, Planistromellaceae, and Saccharataceae, while Endomelanconiopsidaceae, Pseudofusicoccaceae, and Septorioideaceae were considered as synonyms of Botryosphaeriaceae, Phyllostictaceae, and Saccharataceae, respectively. We followed this last taxonomical revision in our study.
Presently, the order Botryosphaeriales comprises 33 genera [3,9]. Alanomyces and Aplosporella are the only two genera accepted within the family Aplosporellaceae. The family Botryosphaeriaceae currently comprises 22 genera: Alanphillipsia, Barriopsis, Botryobambusa, Botryosphaeria, Cophinforma, Diplodia, Dothiorella, Endomelanconiopsis, Eutiarosporella, Lasiodiplodia, Macrophomina, Marasasiomyces, Mucoharknessia, Neodeightonia, Neofusicoccum, Neoscytalidium, Oblongocollomyces, Phaeobotryon, Sakireeta, Sardiniella, Sphaeropsis, and Tiarosporella [3,9]. Melanopsaceae accommodates only one genus, Melanops, which was supported by several phylogenetic analyses [3,4,11], while Phyllostictaceae includes two genera, Phyllosticta and Pseudofusicoccum [3,4,11]. Planistromellaceae was revised to accommodate Kellermania and Umthunziomyces [3]. Saccharataceae comprises three genera, Pileospora, Saccharata, and Septorioides [3,4].
Botryosphaeriales species cause blight, canker, dieback, and fruit rots on a variety of woody perennials globally [5,12]. In China, infections related to Botryosphaeriales have been described on numerous hosts such as grapes [13,14,15], Caragana arborescens [16], Cercis chinensis [17], Eucalyptus [12], Chinese hackberry [18], blueberry [19,20], forest trees [21,22], and various other woody hosts. Hence, the aim of this study was to characterize the Botryosphaeriales taxa associated with woody hosts in southwestern China based on morphology, DNA sequence data, and phylogeny.
2. Materials and Methods
2.1. Collection of Specimens, Isolation, Morphology, and Culture Characteristics
From 2017 to 2019, specimens were collected in field investigations of numerous decomposing woody hosts in Fanjing mountain (Tongren District), Forest Park (Chishui District), Huaxi wetland park, Xiaochehe wetland park (Guiyang District), Maolan natural reserve (Libo District), Suiyang broad water nature reserve, and Xingyi Wanfenglin in the Karst region of Guizhou province (Table 1). Samples were placed into ziplock plastic bags, relevant data (location, date, etc.) were documented, and samples were taken into the laboratory.
Morphological observations of conidiomata or ascostromata were carried out using a Motic SMZ 168 series stereomicroscope and photographed using a Nikon E80i microscope-camera system. Tarosoft® Image Framework was used to measure morphological characters as in Liu et al. [23], and images included in figures were processed with Adobe Photoshop cs v. 5. To isolate single spores, the procedure according to Chomnunti et al. [24] was followed. Spores germinated on water agar (WA) for 12–24 h were examined and then transferred to potato dextrose agar (PDA) media (OXOID CM0139). Obtained pure cultures were incubated at 25 °C for two weeks, and colony characteristics and morphology of fungal structures were examined for a total of 60 isolates. According to Rayner [25], colony color was inspected after 5–10 days of progression on PDA at 25 °C. Approximately 20 ascomata/conidiomata, 25 asci, and 50 conidia/ascospores were measured to obtain the mean size/length. Shape, color, and presence or absence of the mucous sheath of conidia/ascospores were also documented.
Herbarium specimens were deposited at the HKAS (Herbarium of Cryptogams, Kunming Institute of Botany Academia Sinica Kunming, China) and GZAAS (Herbarium of Guizhou Academy of Agricultural Sciences, Guiyang, China), while living cultures were deposited in the CGMCC (China General Microbiological Culture Collection Center in Beijing, China) and GZCC (Guizhou Culture Collection in Guiyang, China) (Table 1).
2.2. DNA Extraction and Molecular Based Amplification
About 10 mg of aerial mycelia were scraped from 5 day-old isolates grown on PDA medium at 25 °C for the extraction of total genomic DNA using an Extraction Kit of Biospin Fungus Genomic DNA (BioFlux®, Hangzhou, China) according to the manufacturer’s protocol (Hangzhou, China). For initial species confirmation, the internal transcribed spacer (ITS) region was sequenced for all isolates. The BLAST tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 14 August 2020) was used to compare the resulting sequences with those in GenBank. After confirmation of Botryosphaeriales species, two additional gene regions coding for translation elongation factor 1-α (tef) and large subunit rRNA gene (LSU) were sequenced as in Dissanayake et al. [5]. The primer pairs and amplification conditions for each of the above-mentioned gene regions are provided in Table 2. A Bio-Rad C1000 thermal cycler was used to conduct the PCR reactions. The resulting PCR products were visualized on a 1% agarose gel stained with ethidium bromide under UV light by a Gel DocTM XR Molecular Imager (Bio-Rad, USA). All positive amplicons were sequenced by Shanghai Sangon Biological Engineering Technology and Services Co., Ltd. (Shanghai, China).
2.3. Sequence Alignment and Phylogenetic Analyses
Sequence quality was assured by inspecting the chromatograms using BioEdit v. 5 [29]. Sequences were obtained with both forward and reverse primers, and consensus sequences were obtained using DNAStar v. 5.1 (DNASTAR, Inc.). The sequence data generated in this study have been deposited in GenBank (Table 1).
Reference sequences of ITS, tef, and LSU were retrieved from NCBI GenBank, referring to recent publications [3,4,5,6,9] (Table 3) to conduct phylogenetic analyses. The reference sequences were aligned with the sequences obtained in this study (Table 1) using MAFFT (http://www.ebi.ac.uk/Tools/msa/mafft/, accessed on 22 December 2020) [30], then manually adjusted, and phylogenetic relationships were inferred with maximum likelihood (ML), maximum parsimony (MP), and Bayesian inference (BI) using procedures provided in detail by Dissanayake et al. [31]. An overview phylogenetic tree for the order Botryosphaeriales was constructed using ITS, LSU, and tef sequence data as some families in Botryosphaeriales (except Botryosphaeriaceae) comprise only ITS and LSU sequences. Separate phylogenetic trees of the diverse genera (Botryosphaeria, Diplodia, Dothiorella, Lasiodiplodia, and Neofusicoccum) were constructed by combining ITS and tef sequences.
Alignments generated in this study were submitted to TreeBASE (https://treebase.org/treebase-web/home.html, accessed on 18 August 2021). The submission numbers and reviewer access URL for each alignment are provided in Table 4. Taxonomic novelties were submitted to the Faces of Fungi database [32] and Index fungorum (http://www.indexfungorum.org, accessed on 5 August 2021). New species were established based on the guidelines provided by Jeewon and Hyde [33].
3. Results
3.1. Phylogenetic Analyses
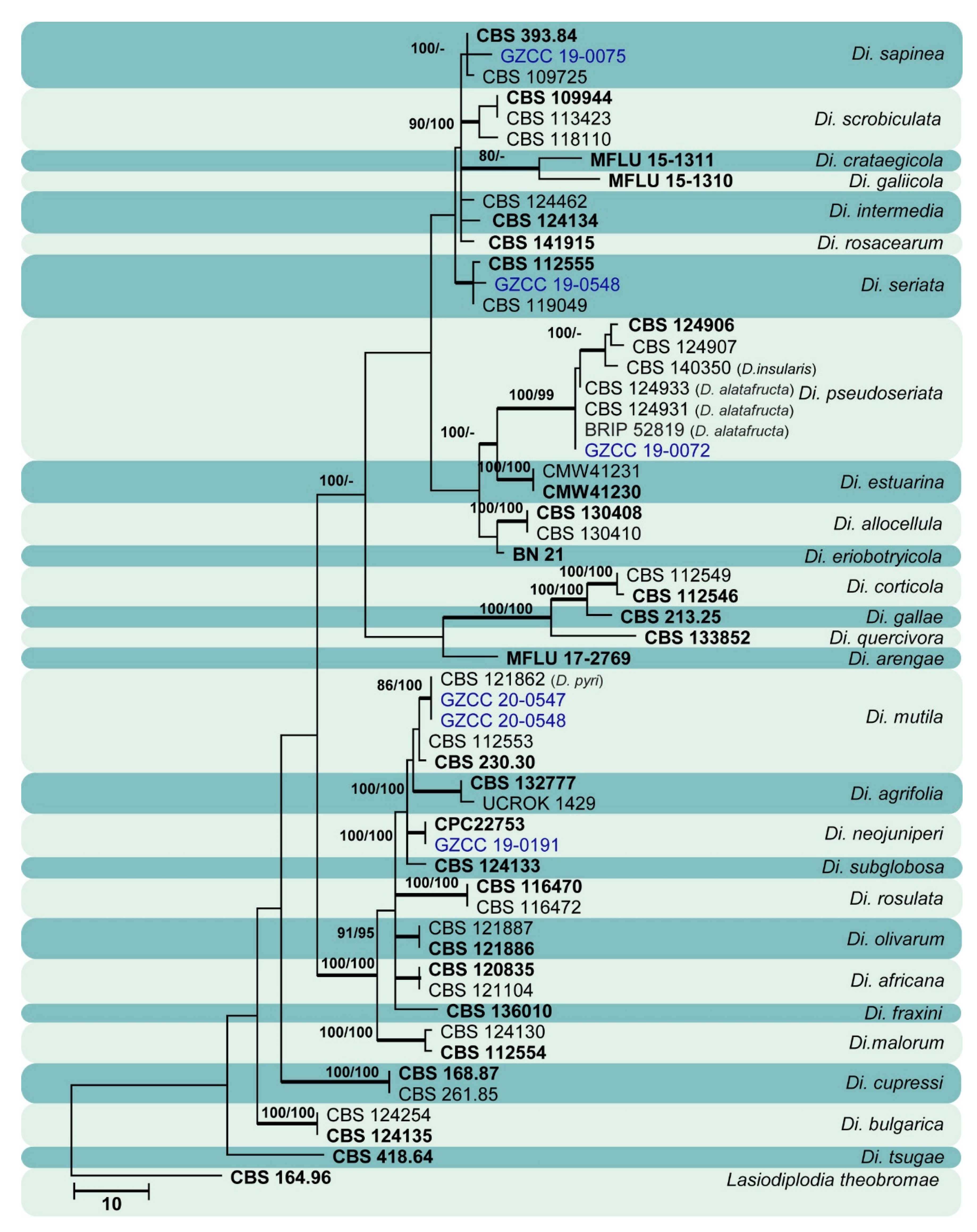
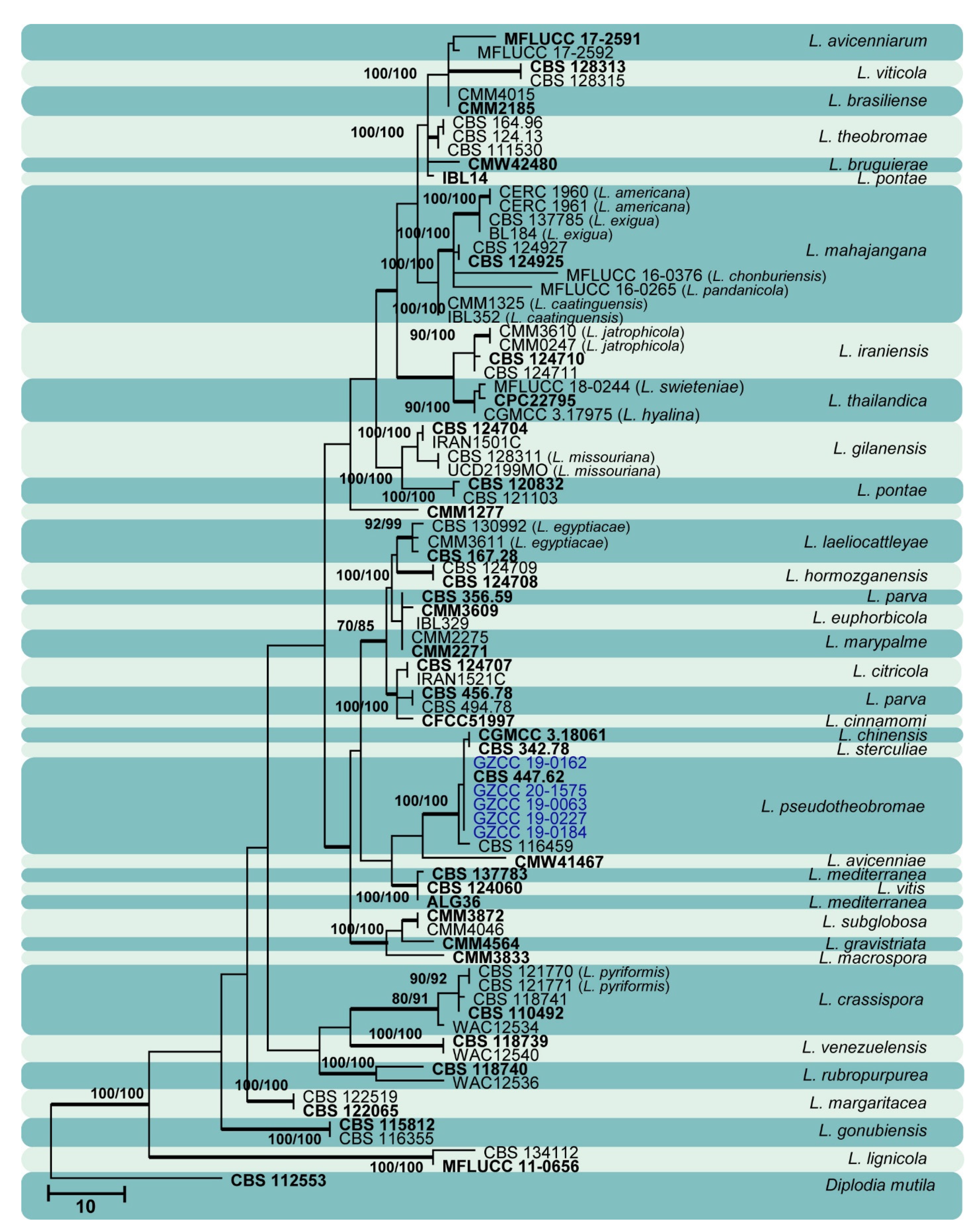
Sixty isolates obtained from various decaying woody hosts in various locations in Guizhou province were primarily recognized by colony characteristics, such as abundant greenish black aerial mycelia on PDA medium. The ITS gene region sequences compared with those in GenBank using the BLAST tool exhibited 95–99% similarity to known Botryosphaeriales species, and these closely related known species were included in the phylogenetic analysis. All details of the alignments (ITS, LSU, tef alignment of the overview phylogenetic tree for the order Botryosphaeriales and ITS and tef alignments for the genera Botryosphaeria, Diplodia, Dothiorella, Lasiodiplodia, and Neofusicoccum) are provided in Table 4. The best-scoring RAxML tree (Figure 1) is presented as the MP and BI methods produced trees with topologies similar to those of ML.
Six different phylogenetic trees were constructed for the 60 isolates obtained in this study. Twelve isolates (20% of total isolates) were treated together in an overview phylogenetic tree and seven of them did not cluster with any known Botryosphaeriales species, thus, three novel species were identified based on the morphological and phylogenetic evidence (Figure 1). In this phylogeny, the isolates obtained in the study were clustered with Aplosporella hesperidica (Figure 2), Barriopsis tectonae (Figure 3), Botryobambusa guizhouensis sp. nov. (Figure 4), Sardiniella celtidis (Figure 5), Sardiniella elliptica sp. nov. (Figure 6 and Figure 7), Sardiniella guizhouensis (Figure 8), Sphaeropsis citrigena, and Sphaeropsis guizhouensis sp. nov (Figure 9).
Twenty-three isolates (38.3% of total isolates) belong to the genus Botryosphaeria, and all of them clustered with B. dothidea (Figure 10). Six isolates (10% of total isolates) belong to the genus Diplodia and were identified as Di. mutila, Di. neojuniperi, Di. pseudoseriata, Di. sapinea, and Di. seriata (Figure 11). Two isolates (3.3% of total isolates) were identified as species of Dothiorella (Do. sarmentorum and Do. yunnana, Figure 12), while five isolates (8.3% of total isolates) belong to the genus Lasiodiplodia (L. pseudotheobromae, Figure 13). All twelve isolates (20% of total isolates) of the genus Neofusicoccum were identified as N. parvum (Figure 14).
3.2. Taxonomy
Aplosporella hesperidica Speg., Anal. Soc. cient. argent. 13: 18 (1882) (Figure 2).
Index Fungorum number: IF218239; Facesoffungi number: FoF07830.
Saprobic on decaying wood. Sexual morph: Not observed. Asexual morph: Conidiomata 220–360 × 420–610 µm ( = 320 × 550 µm, n = 20), solitary, dark brown, immersed to semi-immersed, erumpent, multiloculate, locules separated by pale brown textura prismatica. Ostiole 60–80 µm diam., central. Peridium 75–150 µm (6–10 cell-layers), outer layers composed of dark brown textura angularis, becoming hyaline towards the inner region. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 6–11 × 2.5–3 µm ( = 8 × 2.5 µm, n = 20), holoblastic, hyaline, cylindrical to doliiform, smooth-walled, proliferating percurrently with 1–3 annellations near the apex. Paraphyses 35–95 × 4–8 µm ( = 60 × 5 µm, n = 20), wide at the base, 1–3 µm wide in the upper part, hyaline, smooth-walled, septate, branched below. Conidia 17–25 × 10–18 µm ( = 20 × 12 µm, n = 50), aseptate, initially hyaline, smooth-walled, broadly ellipsoidal to subcylindrical, with rounded ends, becoming dark brown (black in mass), prominently verruculose before discharge from pycnidia.
Culture characteristics: Conidia germinate on WA within 12 h at room temperature. Colonies on PDA after five days at 25 °C become olivaceous to grey-olivaceous in the center, olivaceous-buff to greenish-olivaceous towards the margin. Aerial mycelium appressed, floccose, white to smoke grey. Colonies flat with undulate edge, 38 mm diameter after two days, reaching the edge of the Petri dish within 10 days.
Material examined: China, Guizhou province, Tongren District, Fanjing mountain, on decaying woody host, July 2018, Y. Y. Chen, GZAAS 19-1814, living culture GZCC 19-0095.
Notes: Our sample morphologically lines up with the description of Aplosporella hesperidica provided by Spegazzini [34] as it has erumpent, black conidiomata and brown, smooth-walled, oblong conidia with overlapping spore dimensions of 22–25 × 9–11 µm. It is identical to A. hesperidica based on morphology and phylogeny (Figure 1). This is the first time A. hesperidica has been reported in China.
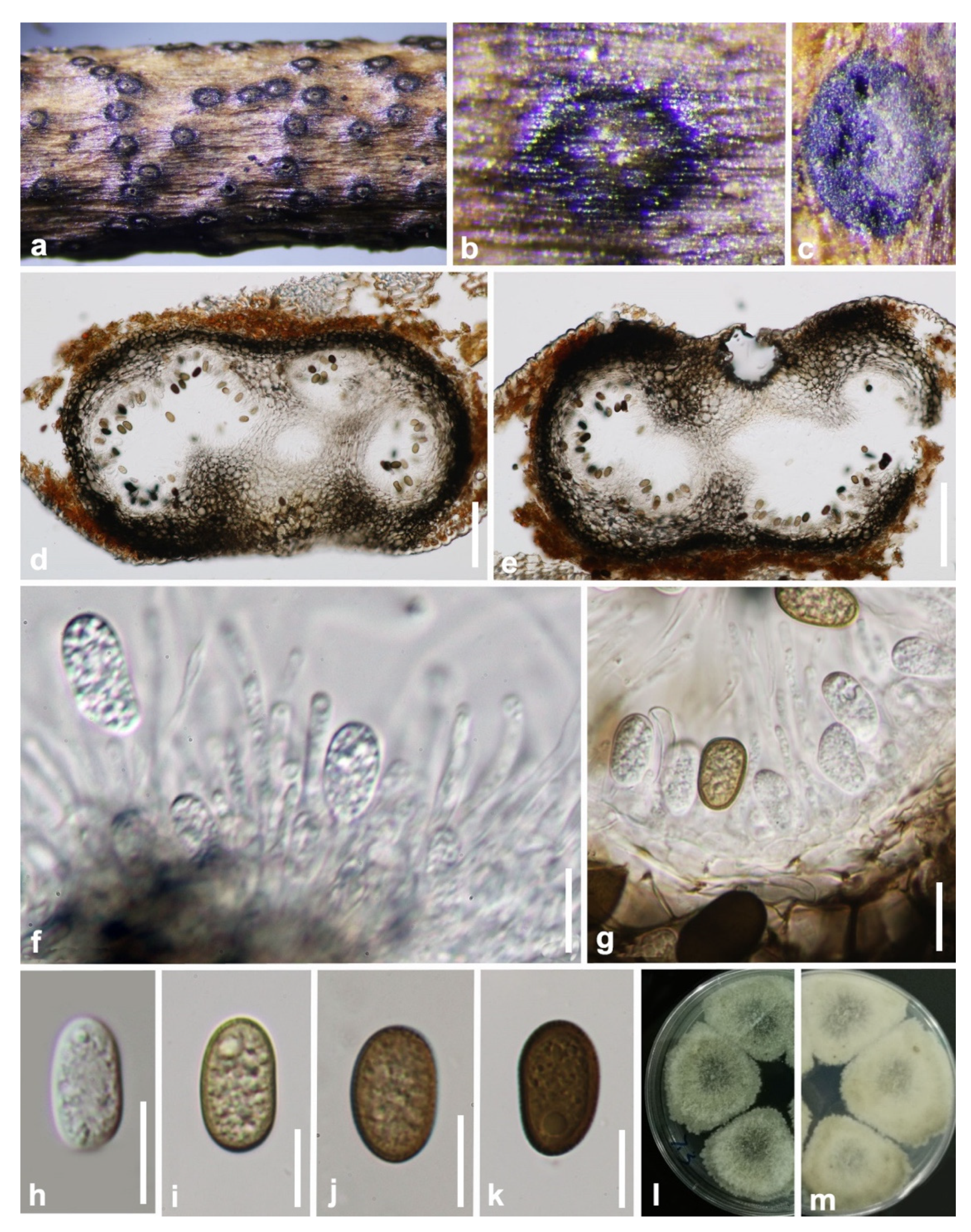
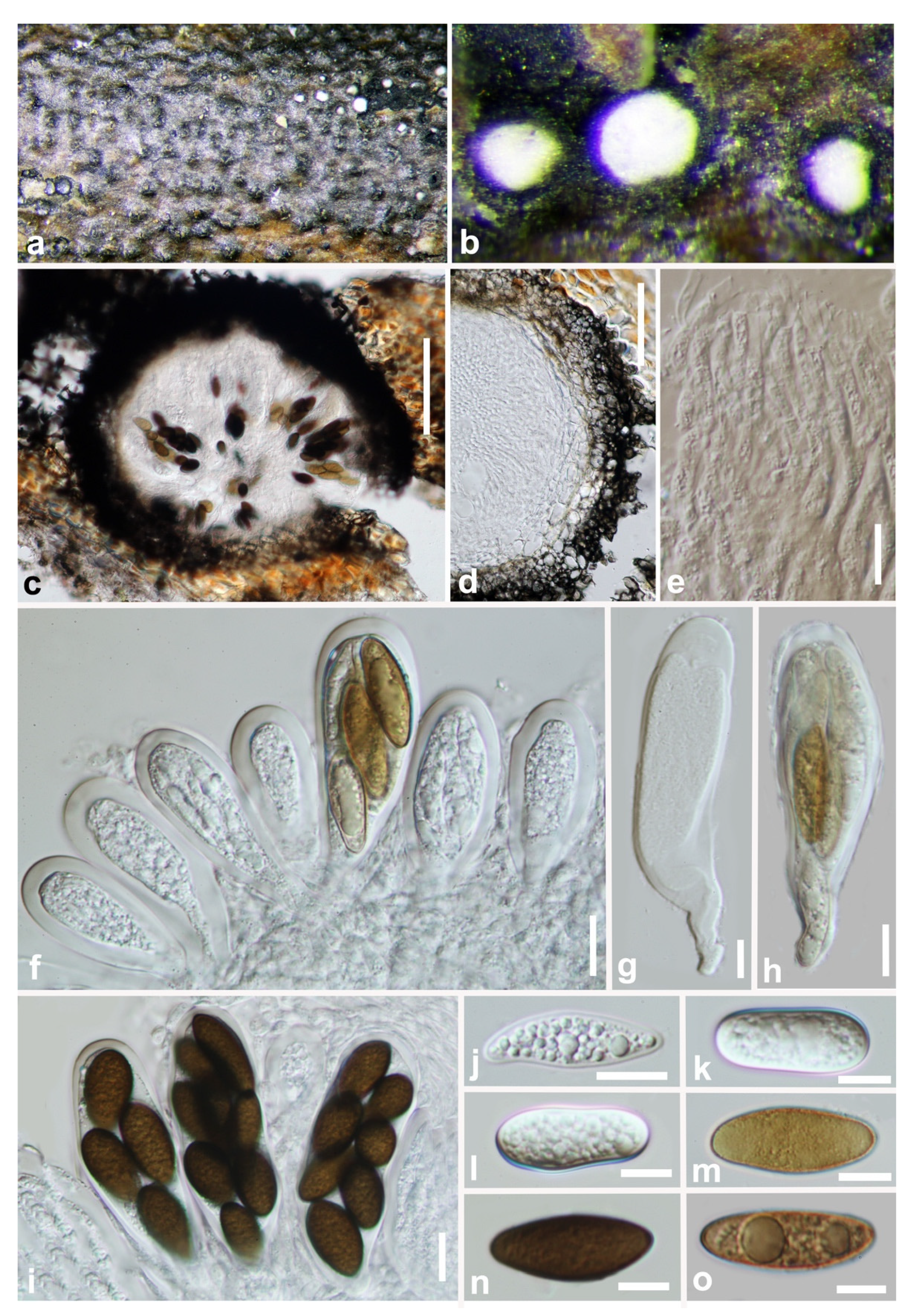
Figure 2.
Aplosporella hesperidica (GZAAS 19-1814). (a–c) Conidiomata on host surface. (d,e) Vertical hand sections of multiloculate conidiomata. (f,g) Developing conidia attached to conidiogenous cells. (h) Hyaline immature conidium. (i–k) Mature brown conidia. (l,m) Five-day-old culture on PDA (OXOID CM0139) from above and below. Scale bars: (d,e) = 100 μm, (f–k) = 10 μm.
Figure 2.
Aplosporella hesperidica (GZAAS 19-1814). (a–c) Conidiomata on host surface. (d,e) Vertical hand sections of multiloculate conidiomata. (f,g) Developing conidia attached to conidiogenous cells. (h) Hyaline immature conidium. (i–k) Mature brown conidia. (l,m) Five-day-old culture on PDA (OXOID CM0139) from above and below. Scale bars: (d,e) = 100 μm, (f–k) = 10 μm.

Barriopsis tectonae Doilom, L.A. Shuttlew., and K.D. Hyde, Phytotaxa 176: 84 (2014) (Figure 3).
Index Fungorum number: IF808202; Facesoffungi number: FoF09644.
Saprobic on decaying wood. Sexual morph: Ascostromata 286–420 × 258–349 μm ( = 350 × 318 µm, n = 20), black, immersed, aggregated or clustered, scattered, multi-loculate, composed of one or up to three globose ascomata in each ascostroma, erumpent through the bark at maturity, discoid to pulvinate or hemispherical, discrete or wide-spreading. Peridium composed of several layers of dark brown-walled cells of textura angularis. Pseudoparaphyses 2.5–3.5 μm wide, hyphae-like, septate, embedded in a gelatinous matrix. Asci 107–154 × 34–41 ( = 129 × 36 µm, n = 25), eight-spored, bitunicate, fissitunicate, clavate to sub-clavate, broad, with a short pedicel and apically rounded with an ocular chamber. Ascospores 31–34 ×14–15 µm ( = 32 × 15 µm, n = 50), biseriate, brown to dark brown, aseptate, ellipsoid-oval, inequilateral, slightly curved, widest in the median, ends rounded, light brown in the center, smooth or verrucose, without a gelatinous sheath. Asexual morph: Not observed.
Culture characteristics: Ascospores germinate on WA within 18 h. Colonies growing on PDA reach 2 cm diameter after five days at 25 °C. Effuse, velvety with entire to slightly undulate edge. Blackish green to black.
Material examined: China, Guizhou province, Libo District, Maolan natural reserve, on decaying woody host, July 2017, Y. Y. Chen, GZAAS 19-1985, living culture GZCC 19-0266.
Notes: In the phylogenetic analysis, an isolate obtained in this study (GZCC 19-0266) was grouped with the ex-type isolate of Barriopsis tectonae (Figure 1) with absolute bootstrap support (ML/MP/BI = 100/100/1.0). This isolate is morphologically similar to Ba. tectonae as of the report by Doilom et al. [35] with erumpent, black ascostromata and overlapping biseriate, brown, aseptate, ellipsoid, thick-walled conidia with overlapping spore dimensions of 29–33 × 13–15 μm ( = 30 × 14 μm, n = 30). It is identical to Ba. tectonae based on morphology and phylogeny (Figure 1). We therefore identify our isolate as Ba. tectonae based on phylogenetic analyses, and the isolate is introduced here as a new locality record from Guizhou province, China. This is the first time Ba. tectonae has been reported in China.
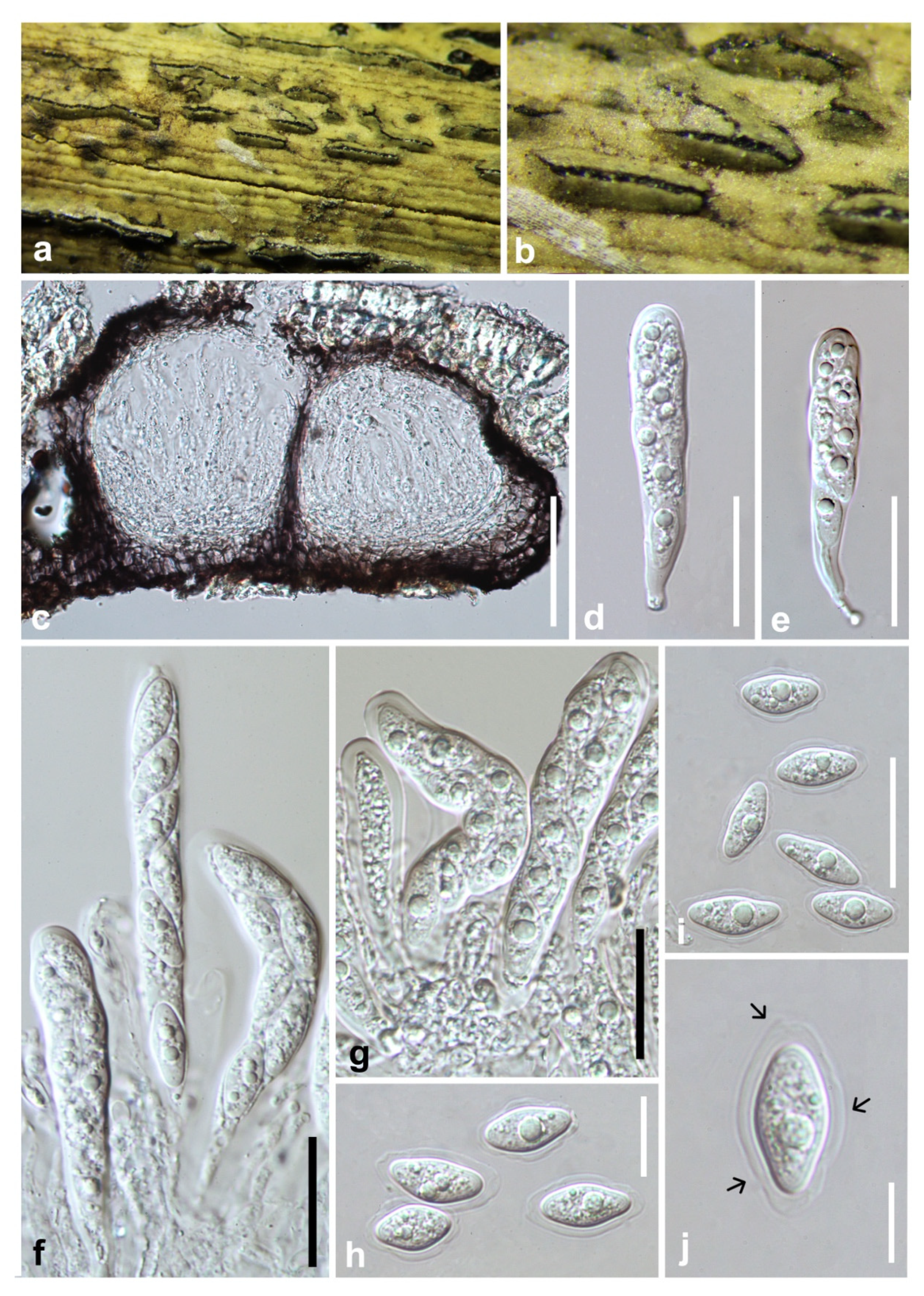
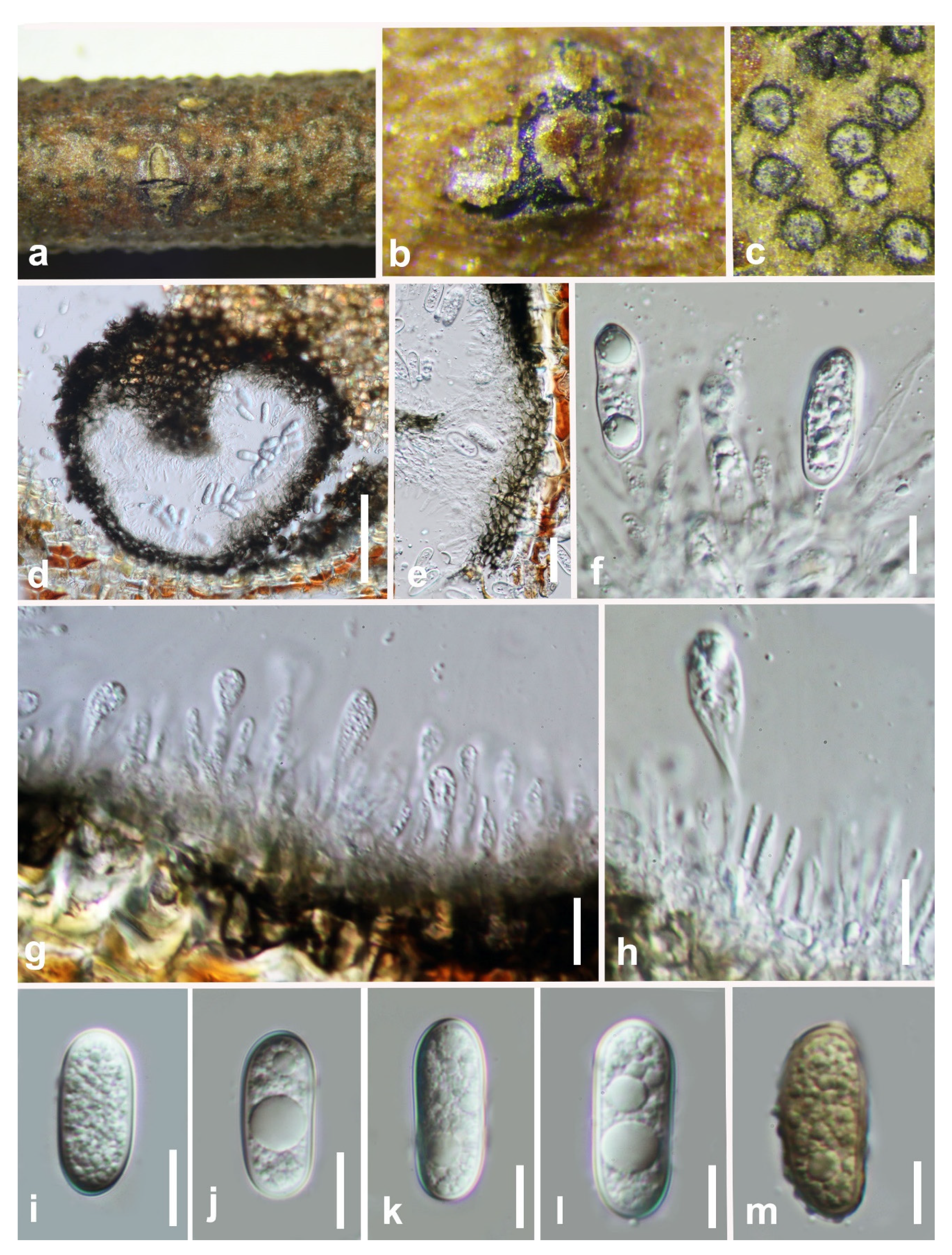
Figure 3.
Barriopsis tectonae (GZAAS 19-1985). (a,b) Appearance of ascomata on wood. (c) Horizontal section of the ascomata. (d,e) Vertical section of ascomata. (f) Peridium. (g,h) Immature asci. (i) Immature and mature asci. (j) Mature asci. (k) Pseudoparaphyses. (l–p). Brown ascospores. Scale bars: (d) = 50 μm, (e) = 100 μm, (f) = 50 μm, (g–j) = 20 μm, (k–p) = 10 μm.
Figure 3.
Barriopsis tectonae (GZAAS 19-1985). (a,b) Appearance of ascomata on wood. (c) Horizontal section of the ascomata. (d,e) Vertical section of ascomata. (f) Peridium. (g,h) Immature asci. (i) Immature and mature asci. (j) Mature asci. (k) Pseudoparaphyses. (l–p). Brown ascospores. Scale bars: (d) = 50 μm, (e) = 100 μm, (f) = 50 μm, (g–j) = 20 μm, (k–p) = 10 μm.

Index Fungorum number: IF558473; Facesoffungi number: FoF09645.
Etymology: Name refers to the location where the fungus was collected, Guizhou, China.
Holotype: HKAS 112600.
Saprobic on a decaying bamboo. Sexual morph: Ascostromata 218–340 × 210–420 μm ( = 275 × 345 μm, n = 20), black, immersed or erumpent, gregarious, uniloculate, locules globose to subglobose, coriaceous. Peridium 36–60 μm, comprises several layers of cells textura angularis, broader at the base, outer layers dark to dark brown and thick-walled, inner layers hyaline and thin-walled. Asci 78–115 × 12–16 μm ( = 94.5 × 14.5 μm, n = 25), eight-spored, bitunicate, fissitunicate, clavate to cylindro-clavate, usually wider at the apex, pedicellate, apically rounded with well-developed ocular chamber. Ascospores 13–22 × 8–11 μm ( = 17.5 × 9 μm, n = 50), uniseriate at the base or irregularly biseriate, hyaline, aseptate, ellipsoidal to obovoid, thick-walled, surrounded by distinctive structured mucilaginous sheath. Asexual morph: Not observed.
Culture characteristics: Ascospores germinate on WA within 24 h. Colonies growing on PDA reach a 5 cm diameter after five days at 25 °C. Fast growing; white in the first few days, become grey to green-black after one week. Reverse grey to black, flattened, fairly dense, surface smooth with crenate edge.
Material examined: China, Guizhou province, Chishui District, Forest Park, on decaying bamboo, July 2019, Y. Y. Chen, 171, (HKAS 112600, holotype); ex-type living culture CGMCC 3.20348; ibid., (GZAAS 20-0718, paratype), living culture GZCC 19-0734.
Notes:Botryobambusa guizhouensis formed a distinct clade with absolute support (ML/MP/BI = 100/100/1.0) and differed from its closely related species Bo. fusicoccum in the concatenated alignment by 7/680 bp in ITS, 4/803 bp in LSU, and 5/479 bp in tef. Morphologically, Bo. guizhouensis differs from Bo. fusicoccum in having longer asci (94.5 × 14.5 μm vs. 60 × 15.5 μm) and larger ascospores (17.5 × 9 μm vs. 11.5 × 6 μm).
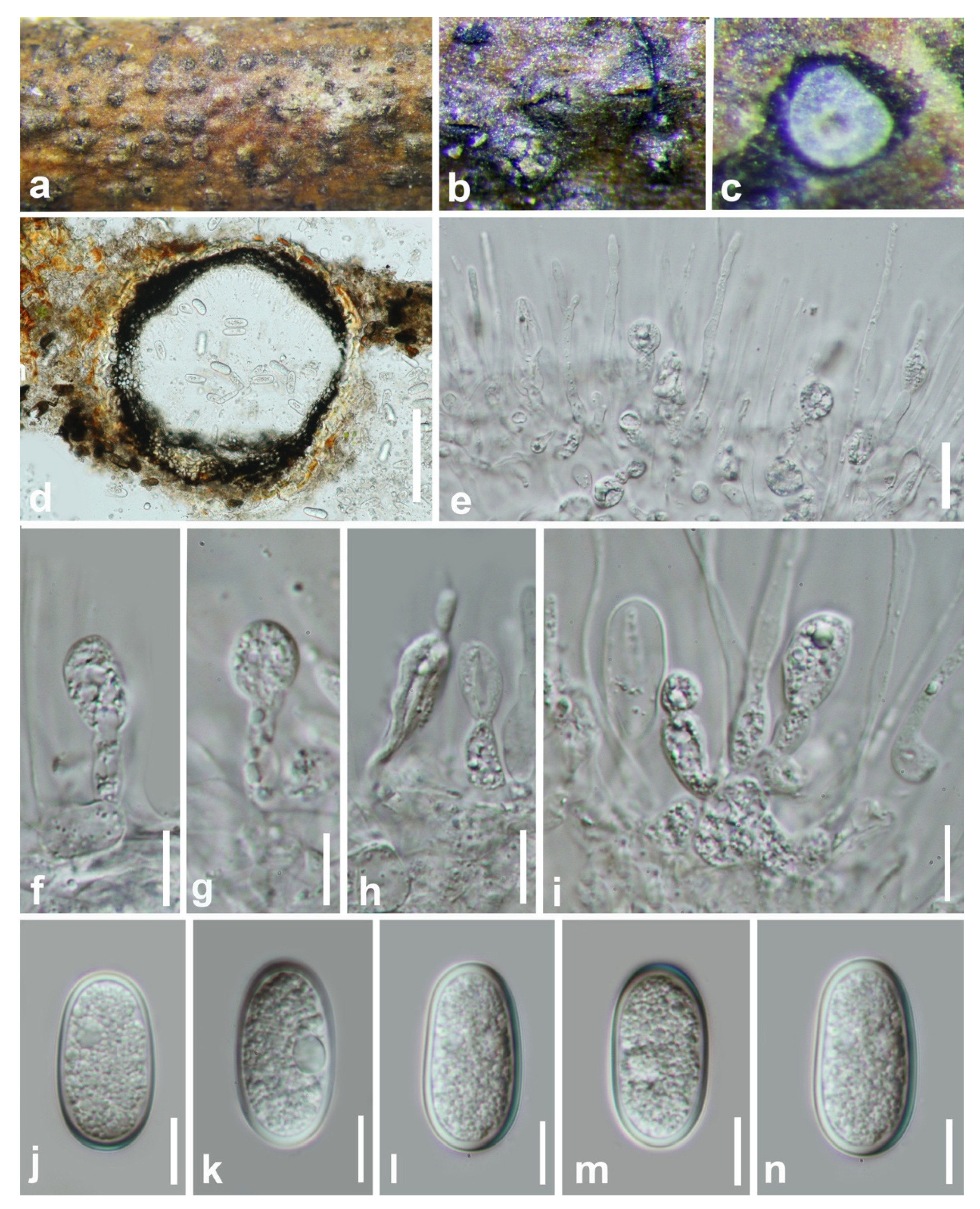
Figure 4.
Botryobambusa guizhouensis (HKAS 112600, holotype). (a,b) Appearance of ascomata on bamboo. (c) Vertical section of ascomata. (d,e) Immature asci. (f,g) Immature and mature asci. (h–j) Hyaline, aseptate ascospores enclosed with mucilaginous sheath. Scale bars: (c) = 50 μm, (d–g) = 100 μm, (h,i) = 20 μm, (j) = 10 μm.
Figure 4.
Botryobambusa guizhouensis (HKAS 112600, holotype). (a,b) Appearance of ascomata on bamboo. (c) Vertical section of ascomata. (d,e) Immature asci. (f,g) Immature and mature asci. (h–j) Hyaline, aseptate ascospores enclosed with mucilaginous sheath. Scale bars: (c) = 50 μm, (d–g) = 100 μm, (h,i) = 20 μm, (j) = 10 μm.

Sardiniella celtidis Dissan., Camporesi and K.D. Hyde, Fungal Divers 87: 12 (2017) (Figure 5).
Index Fungorum number: IF552896; Facesoffungi number: FoF02732.
Saprobic on a decaying host. Sexual morph: Ascostromata 210–300 × 275–340 μm ( = 270 × 310 μm, n = 20), dark brown to black, globose, immersed in the substrate, partially erumpent at maturity, ostiolate. Ostiole circular, central. Peridium 48–72 μm thick, composed of dark brown thick-walled cells of textura angularis, becoming thin-walled and hyaline towards the inner region. Pseudoparaphyses 3–5 μm wide, thin-walled, hyaline. Asci 62–90 × 26–34 μm ( = 78 × 30 μm, n = 20), four- to eight-spored, bitunicate, cylindric-clavate, endotunica thick-walled, with a well-developed ocular chamber. Ascospores 19–27 × 15–18 μm ( = 23 × 16 μm, n = 50), 1-septate, irregularly biseriate, dark brown, oblong to ovate, widest in center, straight, moderately thick-walled, surface smooth. Asexual morph: Not observed.
Culture characters: Ascospores germinate on WA within 24 h. Colonies on PDA reach a 2 cm diameter after five days at 25 °C. Mycelium velvety and moderately fluffy with an irregular margin, surface initially white and later turning dark olivaceous from the middle of the colony and dark grey in reverse.
Material examined: China, Guizhou province, Guiyang city, Xingyi Wanfenglin, on decaying woody host, June 2019, Y.Y. Chen, GZAAS 19-1967, living culture GZCC 19-0248.
Notes: Our sample is phylogenetically identical to Sardiniella celtidis (Figure 1). Only the asexual morph of Sa. celtidis was provided when Hyde et al. [36] introduced this species. Here, we provide the sexual morph of Sa. celtidis.
Figure 5.
Sardiniella celtidis (GZAAS 19-1967). (a,b) Appearance of ascomata on decaying wood. (c) Peridium. (d,e) Vertical section of ascomata. (f) Immature ascus. (g,h) Mature asci. (i) Pseudoparaphyses. (j–l) Brown, one-septate ascospores. (m,n) Five-day-old culture on PDA (OXOID CM0139) from above and below. Scale bars: (c–e) = 100 μm, (f–l) = 20 μm.
Figure 5.
Sardiniella celtidis (GZAAS 19-1967). (a,b) Appearance of ascomata on decaying wood. (c) Peridium. (d,e) Vertical section of ascomata. (f) Immature ascus. (g,h) Mature asci. (i) Pseudoparaphyses. (j–l) Brown, one-septate ascospores. (m,n) Five-day-old culture on PDA (OXOID CM0139) from above and below. Scale bars: (c–e) = 100 μm, (f–l) = 20 μm.

Sardiniella elliptica Y.Y. Chen, A. J. Dissanayake, and Jian K. Liu., sp. nov (Figure 6 and Figure 7).
Index Fungorum number: IF558474; Facesoffungi number: FoF09646.
Etymology: Named referring to the shape of the conidiospores.
Holotype: HKAS 112594.
Saprobic on decaying host. Sexual morph: Ascostromata 280–390 × 295–340 μm ( = 340 × 310 μm, n = 20), dark brown to black, globose, submerged in the substrate, partially erumpent at maturity, ostiolate. Ostiole circular, central. Peridium 30–48 μm thick, composed of dark brown thick-walled cells of textura angularis, becoming thin-walled and hyaline towards the inner region. Pseudoparaphyses 2–3 μm wide, thin-walled, hyaline. Asci 71–93 × 19–24 μm, ( = 86 × 22 μm, n = 25), 4(–8)-spored, bitunicate, cylindric-clavate, endotunica thick-walled, with a well-developed ocular chamber. Ascospores 26–33 × 9–12 μm ( = 29 × 11 μm, n = 50), irregularly biseriate, initially hyaline and becoming dark brown, oblong to ovate, widest in center, straight, moderately thick-walled, surface smooth. Asexual morph: Appearing as subepidermal black spots on the substrate with black margins, with circular sunken perforation through the bark. Conidiomata 190–240 × 274–310 µm ( = 220 × 290 µm, n = 20), pycnidial, immersed, arranged singly or in small groups within the bark, globose to subglobose, dark brown to black, solitary or gregarious. Ostiole central. Peridium 27–35 µm thick, comprising dark brown to hyaline, multi-layered (3–5 layered), thick-walled cells of textura angularis. Conidiogenous cells lining the inner surface of the conidioma, hyaline, short obpyriform to subcylindrical. Conidia 25–32 × 10–13 µm ( = 28 × 12 µm, n = 50), ellipsoid to obovoid, immature conidia hyaline, mature conidia becoming medium to dark brown.
Culture characters: Colonies on PDA reaching a 70 mm diameter after five days at 25 °C. Mycelium velvety and moderately fluffy with an irregular margin. Surface initially white and later turning dark olivaceous from the middle of the colony and dark grey in reverse.
Material examined: China, Guizhou province, Guiyang District, Huaxi wetland park, on decaying woody host, April 2017, Y.Y. Chen, 18-76 (HKAS 112594, holotype); ex-type living culture CGMCC 3.20349; ibid., Libo District, Maolan natural reserve, July 2017, 19-120 (GZAAS 19-1855, paratype), living culture GZCC 19-0262; ibid., Xingyi Wanfenglin, June 2019, 19-96 (GZAAS 19-1838, paratype), living culture GZCC 19-0245.
Notes: Three isolates of Sardiniella elliptica clustered together with Sa. celtidis, Sa. guizhouensis, and Sa. urbana and formed a well-supported clade representing the genus Sardiniella; thus, it can be recognized as a distinct lineage within Sardiniella. Sardiniella elliptica can be distinguished from the above closely related species based on ITS and tef loci for Sa. celtidis (5/680 bp in ITS, 21/479 bp in tef) and Sa. urbana (5/680 bp in ITS, 8/803 bp in LSU, 27/479 bp in tef). In addition, Sa. elliptica can be morphologically distinguished from other known Sardiniella species based on its aseptate mature ascospores.
Figure 6.
Sardiniella elliptica (HKAS 112594, holotype, sexual morph). (a,b) Appearance of ascomata on decaying wood. (c) Vertical section of ascomata. (d) Peridium. (e) Pseudoparaphyses. (f) Immature and mature asci. (g) Immature ascus. (h,i) Mature asci. (j–l) Immature hyaline ascospores. (m–o) Mature, brown, aseptate ascospores. Scale bars: (c) = 100μm, (d) = 50 μm, (e–o) = 10 μm.
Figure 6.
Sardiniella elliptica (HKAS 112594, holotype, sexual morph). (a,b) Appearance of ascomata on decaying wood. (c) Vertical section of ascomata. (d) Peridium. (e) Pseudoparaphyses. (f) Immature and mature asci. (g) Immature ascus. (h,i) Mature asci. (j–l) Immature hyaline ascospores. (m–o) Mature, brown, aseptate ascospores. Scale bars: (c) = 100μm, (d) = 50 μm, (e–o) = 10 μm.

Figure 7.
Sardiniella elliptica (GZAAS 19-1838, asexual morph). (a–c) Conidiomata on host surface. (d) Vertical section of conidiomata. (e) Peridium. (f–h) Conidiogenous cells and developing conidia. (i–l) Immature, hyaline conidia. (m) Mature, brown conidia. Scale bars: (d) = 100 μm, (e) = 20 μm, (f–m) = 10 μm.
Figure 7.
Sardiniella elliptica (GZAAS 19-1838, asexual morph). (a–c) Conidiomata on host surface. (d) Vertical section of conidiomata. (e) Peridium. (f–h) Conidiogenous cells and developing conidia. (i–l) Immature, hyaline conidia. (m) Mature, brown conidia. Scale bars: (d) = 100 μm, (e) = 20 μm, (f–m) = 10 μm.

Sardiniella guizhouensis Y.Y. Chen, and Jian K. Liu, Phytotaxa 508: 190 (2021) (Figure 8).
Index Fungorum number: IF558352; Facesoffungi number: FoF09647.
Saprobic on decaying wood. Sexual morph: Not observed. Asexual morph: Conidiomata 180–245 × 275–395 µm ( = 229 × 330 µm, n = 20), immersed, arranged singly or in small groups within the bark, globose to subglobose, dark brown to black, solitary or gregarious. Ostiole central. Peridium 22–34 µm thick, outer layer composed of pigmented thick-walled cells of textura angularis, inner layer composed of hyaline thin-walled cells of textura angularis (three- to five-layered). Conidiogenous cells 6–11 × 6–7 µm ( = 8.5 × 6.5 μm, n = 25), lining the inner surface of the conidioma, hyaline, short obpyriform to subcylindrical. Conidia 21–28 × 10–14 µm ( = 24.5 × 12.5 μm, n = 50), ellipsoid to obovoid, immature conidia hyaline, mature conidia becoming medium to dark brown.
Culture characteristics: Conidia germinating on WA within 18 h and producing germ tubes from each septum. Colonies growing on PDA, reaching a diameter of 4 cm after five days at 25 °C, effuse, velvety, with entire to slightly undulate edge. The early stage of the white, later green.
Material examined: China, Guizhou province, Libo District, Maolan natural reserve, July 2017, GZAAS 19-1948, living culture GZCC 19-0229.
Notes:Sardiniellaguizhouensis was introduced by Chen et al. [37] with both sexual and asexual morphs. One isolate obtained in this study clustered with the ex-type of Sa. guizhouensis (CGMCC 3.19222) in the phylogenetic analyses of combined ITS, LSU, and tef sequence data (Figure 1). We identified our collection as Sa. guizhouensis based on morphology and phylogeny.
Figure 8.
Sardiniella guizhouensis (GZAAS 19-1809, asexual morph). (a,b) Conidiomata on host surface. (c) Horizontal section of conidiomata. (d) Vertical section of conidiomata. (e–i) Conidiogenous cells and developing conidia. (j–n) Hyaline conidia. Scale bars: (d) = 100 μm, (e–n) = 10 μm.
Figure 8.
Sardiniella guizhouensis (GZAAS 19-1809, asexual morph). (a,b) Conidiomata on host surface. (c) Horizontal section of conidiomata. (d) Vertical section of conidiomata. (e–i) Conidiogenous cells and developing conidia. (j–n) Hyaline conidia. Scale bars: (d) = 100 μm, (e–n) = 10 μm.

Index Fungorum number: IF558475; Facesoffungi number: FoF09648.
Etymology: Name refers to the location where the fungus was collected, Guizhou, China.
Holotype: HKAS 112084.
Saprobic on decaying wood. Sexual morph: Ascostromata 132–185 × 122–165 µm ( = 152 × 145 µm, n = 20), initially immersed under host epidermis, becoming semi-immersed to erumpent, breaking through cracks in bark, gregarious and fused, uniloculate, globose to subglobose, membraneous, ostiolate. Ostiole 43–52 μm high, 30–42 μm wide, central, papillate, pale brown, relatively broad, periphysate. Peridium 28–44 μm wide, broader at the base, comprising several layers of relatively thick-walled, dark brown to black-walled cells arranged in a textura angularis. Pseudoparaphyses hyphae-like, numerous, embedded in a gelatinous matrix. Asci 67–101 × 19–23 µm ( = 89 × 20 μm, n = 25), eight-spored, bitunicate, fissitunicate, clavate to cylindro-clavate, sometimes short pedicellate, mostly long pedicellate, apex rounded with an ocular chamber. Ascospores 20–23 × 7.8–8.3 µm ( = 22 × 8 μm, n = 50), overlapping uniseriate to biseriate, hyaline, aseptate, ellipsoidal to obovoid, slightly wide above the center, minutely guttulate, smooth-walled. Asexual morph: Not observed.
Culture characteristics: Ascospores germinating on PDA within 18 h. Germ tubes produced from both ends of the ascospores. Fast growing; fimbriate, flat or effuse, dense, convex with papillate surface, reaching the edge of the Petri dish after seven days.
Material examined: China, Guizhou province, Libo District, Maolan natural reserve, on decaying woody host, July 2017, Y.Y. Chen, GZAAS4 (HKAS 112084, holotype); ex-type living culture CGMCC 3.20352; ibid., Xingyi Wanfenglin, on decaying woody host, June 2019, Y.Y. Chen, (GZAAS 19-2892, paratype), living culture GZCC 19-0273.
Notes:Sphaeropsis guizhouensis formed a distinct clade (Figure 1) and is phylogenetically distinct from Sp. eucalypticola (MFLUCC 11-0579) in a clade with absolute support (ML/MP/BI = 100/100/1.0). Sphaeropsis guizhouensis can be distinguished from Sp. eucalypticola based on ITS (3/680 bp), LSU (1/803 bp), and tef (25/479 bp). Sphaeropsis guizhouensis differs from Sp. eucalypticola in having smaller asci (89 × 20 μm vs. 106 × 29 μm) and ascospores (22 × 8 μm vs. 30 × 12 μm).
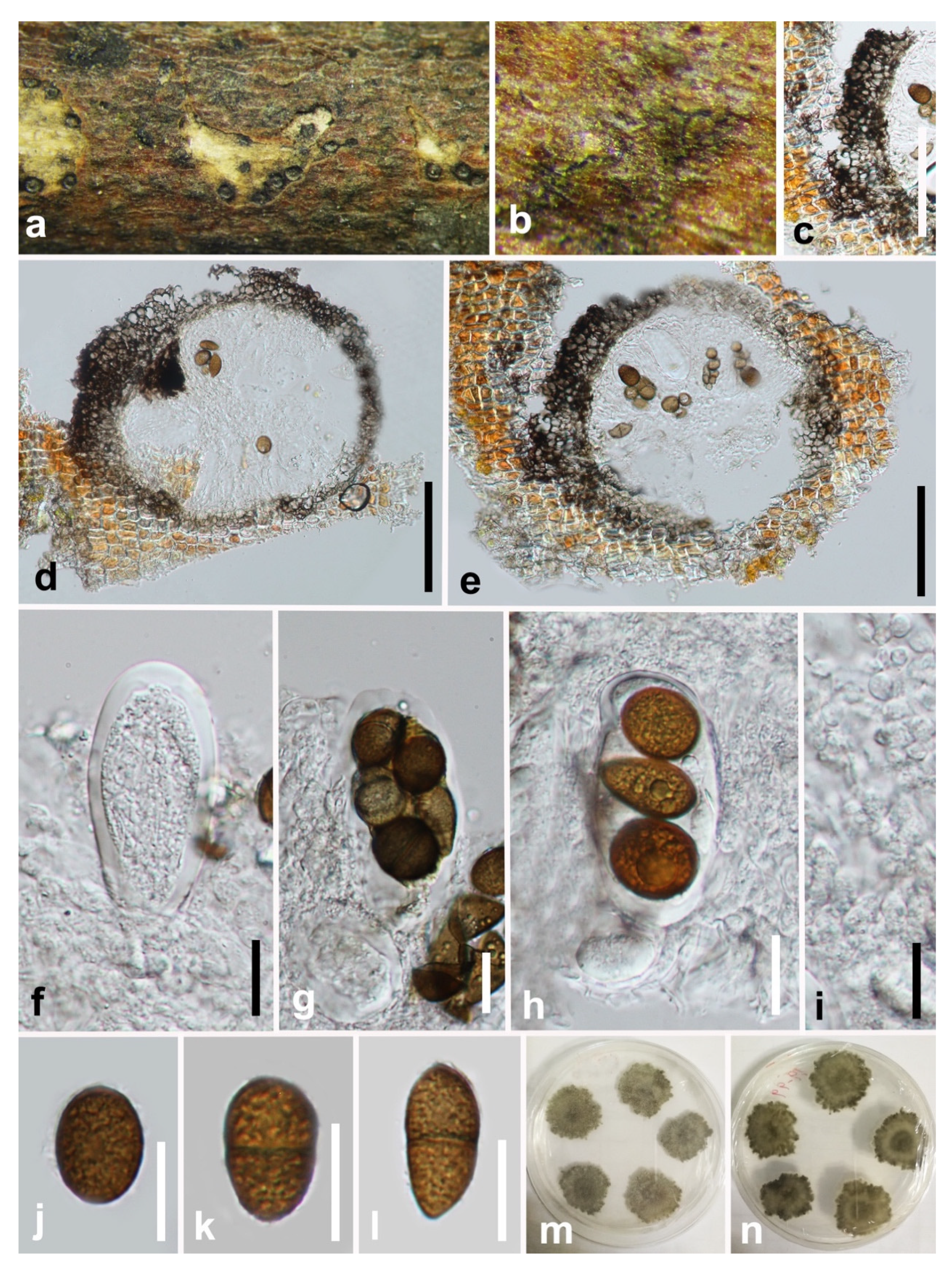
Figure 9.
Sphaeropsis guizhouensis (HKAS 112084, holotype). (a–c) Appearance of ascomata on wood. (d,e) Vertical section of ascomata. (f) Peridium. (g–j) Immature and mature asci. (k) Pseudoparaphyses. (l–p) Hyaline mature ascospores. Scale bars: (d,e) = 50 μm, (f) = 20 μm, (g–p) = 10 μm.
Figure 9.
Sphaeropsis guizhouensis (HKAS 112084, holotype). (a–c) Appearance of ascomata on wood. (d,e) Vertical section of ascomata. (f) Peridium. (g–j) Immature and mature asci. (k) Pseudoparaphyses. (l–p) Hyaline mature ascospores. Scale bars: (d,e) = 50 μm, (f) = 20 μm, (g–p) = 10 μm.

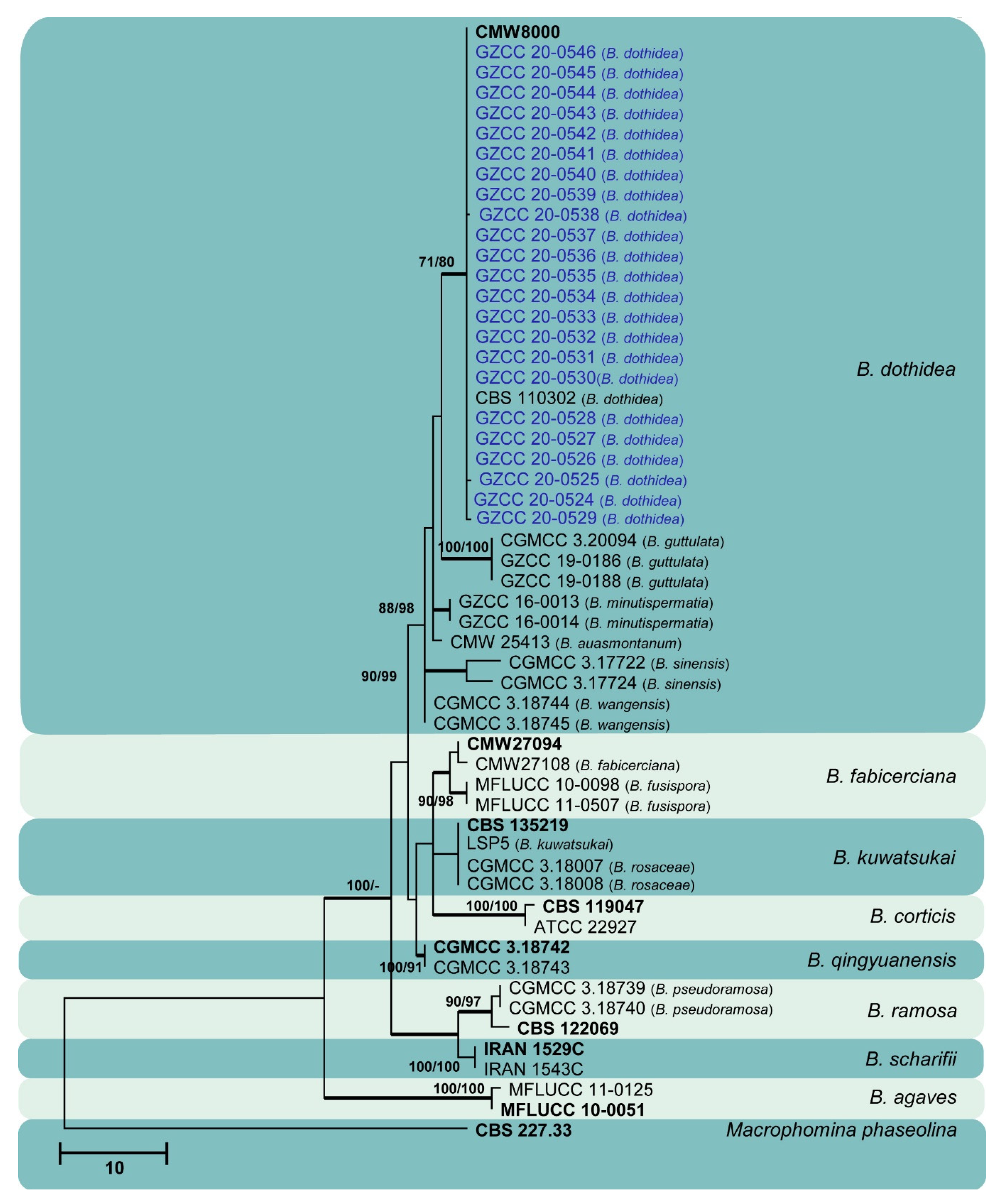
Figure 10.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Botryosphaeria. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Macrophomina phaseolina (CBS 227.33).
Figure 10.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Botryosphaeria. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Macrophomina phaseolina (CBS 227.33).

Figure 11.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Diplodia. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Lasiodiplodia theobromae (CBS 164.96).
Figure 11.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Diplodia. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Lasiodiplodia theobromae (CBS 164.96).

Figure 12.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Dothiorella. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Neofusicoccum parvum (CMW9081).
Figure 12.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Dothiorella. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Neofusicoccum parvum (CMW9081).

Figure 13.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Lasiodiplodia. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Diplodia mutila (CBS 112553).
Figure 13.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Lasiodiplodia. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Diplodia mutila (CBS 112553).

Figure 14.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Neofusicoccum. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Dothiorella viticola (CBS 117009).
Figure 14.
Maximum likelihood tree based on analysis of combined ITS and tef sequences of Neofusicoccum. Bootstrap support values of ML, MP > 75% are shown near the nodes, and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species. Ex-type strains are in bold. The tree is rooted to Dothiorella viticola (CBS 117009).

4. Discussion
This study reports the largest collection of Botryosphaeriales isolates from Guizhou province, China, and is the first attempt to characterize Botryosphaeriales species from various nature reserves in the province. Three new species were described in three rarely observed genera: Botryobambusa (Bo. guizhouensis), Sardiniella (Sa. elliptica), and Sphaeropsis (Sp. guizhouensis). These three genera have few species and have been sporadically isolated worldwide. Apart from these novel species, 15 known species—Aplosporella hesperidica, Barriopsis tectonae, Botryosphaeria dothidea, Diplodia mutila, Di. neojuniperi, Di. pseudoseriata, Di. sapinea, Di. seriata, Dothiorella sarmentorum, Do. yunnana, Lasiodiplodia pseudotheobromae, Neofusicoccum parvum, Sardiniella celtidis, Sa. guizhouensis, and Sphaeropsis citrigena—were isolated and included in their respective phylogenies.
The genus Aplosporella consists of plant pathogens, saprobes, and endophytes. Though 300 epithets of Aplosporella are registered in Index Fungorum (http://www.indexfungorum.org/Names/Names.asp, accessed in 23 August 2021), only 12 species are accepted within this genus (www.botryosphaeriales.org). Aplosporella hesperidica has been reported in several Asian countries [17,22,38,39,40,41,42], but this is the first time it has been identified in China.
Five species are accepted in the genus Barriopsis [35,43,44,45]. Barriopsis tectonae was introduced by Doilom et al. [35] from a dead Tectona grandis branch collected in Thailand. So far, this species has been reported only from Thailand (http://nt.arsgrin.gov/fungaldatabases/, accessed on 25 September 2021), and here we provide a new country report of Ba. tectonae (Figure 3) based on sexual morphological characteristics and molecular evidence.
Botryobambusa is a monotypic genus (www.botryosphaeriales.org). Liu et al. [1] introduced and compared Botryobambusa with other existing genera in Botryosphaeriales. It is thus far only identified from bamboo in Thailand. In this study, two isolates obtained from decaying bamboo in Forest Park, Chishui District in Guizhou province were identified as a novel species (Bo. guizhouensis). The sexual morph of Bo. guizhouensis was distinguished from Bo. fusicoccum by its larger asci and ascospores as well as by molecular phylogeny. Our study shows that the genus can be clearly discriminated from the morphologically similar genus Botryosphaeria by its velvety, hyaline, and sheathed ascospores. Phylogenetically, these two genera are clearly distinct lineages.
Linaldeddu et al. [46] introduced the genus Sardiniella by denoting Sa. urbana as the type species. Hyde et al. [36] introduced the second species in the genus, Sa. celtidis, while Chen et al. [37] introduced the third species, Sa. guizhouensis, reporting a sexual morph of the genus for the first time. In this study, another new species (Sa. elliptica) is described and assigned to the genus with details provided for a previously known species (Sa. celtidis). With morphological and molecular support, here we present the sexual morph report for Sa. elliptica; a newly introduced species in this study (Figure 6 and Figure 7). So far, Sardiniella species are known only from Italy and China (http://nt.arsgrin.gov/fungaldatabases/, accessed on 25 September 2021).
Though more than 630 names exist in Sphaeropsis (Index Fungorum, August 2021), only five species are currently accepted [2,3]. In this study, two isolates obtained from decaying woody hosts in Guizhou province were identified as a novel species, Sp. guizhouensis. The sexual morph of Sp. guizhouensis (Figure 9) is distinguished from the other species in this genus by ascospore dimensions. Another previously known species Sp. citrigena was also isolated and included in the phylogenetic analysis.
Botryosphaeria dothidea (Figure 10) and Neofusicoccum parvum (Figure 14) were the most isolated species in this study, consistent with some prior studies [47,48,49,50,51], which indicates the ability of species in these genera to inhabit a variety of plant species and geographic areas globally. Certain Diplodia species occupy extensive host ranges, such as Di. seriata which has been documented on more than 250 hosts [3,5]. In this study, we isolated five Diplodia species (Figure 11): Di. mutila, Di. neojuniperi, Di. pseudoseriata, Di. sapinea, and Di. seriata. Our study revealed two previously known Dothiorella species (Figure 12), Do. sarmentorum and Do. yunnana, for the first time from Guizhou province. Lasiodiplodia pseudotheobromae is a common and cosmopolitan species on diverse host plants and has been reported from various localities globally. This study revealed five saprobic L. pseudotheobromae isolates (Figure 13) in Guizhou province.
Members of Botryosphaeriales signify a rising risk to agricultural crops and urban and natural forest ecosystems in China. Collecting and identifying Botryosphaeriales isolates from various hosts and locations is required to describe and understand these species. The occurrence and significance of Botryosphaeriales species in various nature reserves has not been investigated at a larger scale so far in Guizhou province. Hence, in this study, we provided a larger collection of Botryosphaeriales isolates and identify them to species level by both morphology and phylogeny. Further studies are needed to explore and gather data on their occurrence, as precise data of the causal agents is essential.
5. Conclusions
We carried out fungal diversity investigations at a large scale in southwestern China and here we provided a report of Botryosphaeriales species isolated from various woody hosts in Guizhou province, China. The identification of 18 Botryosphaeriales species (15 known species and three new species) associated with saprobic woody hosts was revealed.
Author Contributions
Conceptualization, J.-K.L. and Y.-Y.C.; methodology, Y.-Y.C. and A.J.D.; formal analysis, Y.-Y.C. and A.J.D.; resources, Y.-Y.C. and J.-K.L.; data curation, A.J.D., R.C., and J.-K.L.; writing—original draft preparation, A.J.D.; writing—review and editing, A.J.D. and J.-K.L.; supervision, J.-K.L.; project administration, J.-K.L.; funding acquisition, J.-K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by the Joint Fund of the National Natural Science Foundation of China and the Karst Science Research Center of Guizhou province (Grant No. U1812401).
Acknowledgments
We would like to thank Shaun Pennycook (Landcare Research-Manaaki Whenua, New Zealand) for advising on the new fungal names. A.J.D. acknowledges the University of Electronic Science and Technology of China for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, J.K.; Phookamsak, R.; Doilom, M.; Wikee, S.; Li, Y.M. Towards a natural classification of Botryosphaeriales. Fungal Divers. 2012, 57, 149–210. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J.K. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Li, X.H. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, T.; Liu, J.K.; Maharachchikumbura, S.S.N. Outline of Ascomycota-2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Wikee, S.; Lombard, L.; Nakashima, C.; Motohashi, K.; Chukeatirote, E. A phylogenetic re-evaluation of Phyllosticta (Botryosphaeriales). Stud. Mycol. 2013, 76, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.Y.; Xie, Y.; Zhang, W.; Wang, Y.; Liu, J.K. Species of Botryosphaeriaceae involved in grapevine dieback in China. Fungal Divers. 2013, 61, 221–236. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Zhang, W.; Liu, M.; Chukeatirote, E.; Yan, J.Y. Lasiodiplodia pseudotheobromae causes pedicel and peduncle discolouration of grapes in China. Australas. Plant Dis. 2015, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Zhang, W.; Li, X.H.; Zhou, Y.; Hyde, K.D. First report of Neofusicoccum mangiferae associated with grapevine dieback in China. Phytopathol. Mediterr. 2015, 54, 414–419. [Google Scholar]
- Chen, Y.Y.; Dissanayake, A.J.; Liu, Z.Y.; Liu, J.K. Additions to Karst Fungi 4: Botryosphaeria spp. associated with woody hosts in Guizhou province, China including B. guttulata sp. nov. Phytotaxa 2020, 454, 186–202. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Lasiodiplodia chinensis, a new holomorphic species from China. Mycosphere 2017, 8, 521–532. [Google Scholar] [CrossRef]
- Liang, L.; Li, H.; Zhou, L.; Chen, F. Lasiodiplodia pseudotheobromae causes stem canker of Chinese hackberry in China. J. For. Res. 2020, 31, 2571–2580. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Impaprasert, R.; Zhao, J.R.; Xu, S.G.; Wu, M. Stem die-back of highbush blueberries caused by Neofusicoccum parvum in China. New Dis. Rep. 2013, 27, 3. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhang, H.; Zhou, Z.; Hu, T.; Wang, S. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Pan, Y.; Ye, H.; Lu, J. Isolation and identification of Sydowia polyspora and its pathogenicity on Pinus yunnanensis in southwestern China. J. Phytopathol. 2019, 166, 386–395. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Tian, C.M.; Fan, X.L. Studies of botryosphaerialean fungi associated with canker and dieback of tree hosts in Dongling Mountain of China. Phytotaxa 2018, 348, 63–76. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute & British Mycological Society: Kew, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hall, T. Bioedit 7.5.0.3. Department of Microbiology, North Carolina State University. 2006. Available online: http://www.mbio.ncsu.edu/BioEdit/Bioedit.html (accessed on 11 August 2020).
- Katoh, K.; Standley, K. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Spegazzini, C. Fungi argentini additis nonnullis brasiliensibus montevideensibusque. Pugillus quartus (Continuacion). Anal. Soc. Cient. Argent. 1882, 12, 97–117. [Google Scholar]
- Doilom, M.; Shuttleworth, L.; Roux, J.; Chukeatirote, E.; Hyde, K.D. Barriopsis tectonae sp. nov. a new species of Botryosphaeriaceae from Tectona grandis (teak) in Thailand. Phytotaxa 2014, 176, 081–091. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T. Fungal diversity notes 603–708: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Dissanayake, A.J.; Liu, J.K. Additions to Karst Fungi 5: Sardiniella guizhouensis sp. nov. (Botryosphaeriaceae) associated with woody hosts in Guizhou province, China. Phytotaxa 2021, 508, 187–196. [Google Scholar] [CrossRef]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Botryosphaeriaceae species overlap on four unrelated, native South African hosts. Fungal Biol. 2014, 118, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Hyde, K.D.; Liu, J.K.; Liang, Y.M.; Tian, C.M. Multigene phylogeny and morphology reveal Phaeobotryon rhois sp. nov (Botryosphaeriales, Ascomycota). Phytotaxa 2015, 205, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Ekanayaka, A.H.; Dissanayake, A.J.; Jayasiri, S.C.; To-anun, C.; Jones, E.B.G. Aplosporella thailandica; a novel species revealing the sexual-asexual connection in Aplosporellaceae (Botryosphaeriales). Mycosphere 2016, 7, 440–447. [Google Scholar] [CrossRef]
- Du, Z.; Fan, X.L.; Yang, Q.; Hyde, K.D.; Tian, C.M. Aplosporella ginkgonis (Aplosporellaceae, Botryosphaeriales), a new species isolated from Ginkgo biloba in China. Mycosphere 2017, 8, 1246–1252. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Pennycook, S.R.; Johnston, P.R.; Ramaley, A. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Persoonia 2008, 21, 29–55. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Mohammadi, G.E.; Javadi, A.; Shams-Bakhsh, M.; Zare, R. Barriopsis iraniana and Phaeobotryon cupressi: Two new species of the Botryosphaeriaceae from trees in Iran. Persoonia 2009, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Konta, S.; Phillips, A.J.L.; Bahkali, A.H.; Jones, E.B.G.; Eungwanichayapant, D.P. Botryosphaeriaceae from palms in Thailand—Barriopsis archontophoenicis sp. nov, from Archontophoenix alexandrae. Mycosphere 2016, 7, 921–932. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Alves, A.; Phillips, A.J.L. Sardiniella urbana gen. et sp. nov., a new member of the Botryosphaeriaceae isolated from declining Celtis australis trees in Sardinian streetscapes. Mycosphere 2016, 7, 893–905. [Google Scholar] [CrossRef]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Pathol. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Giambra, S.; Piazza, G.; Alves, A.; Mondello, V.; Berbegal, M. Botryosphaeriaceae species associated with diseased loquat trees in Italy and description of Diplodia rosacearum sp. nov. Mycosphere 2016, 7, 978–989. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemlera, M.; Jami, F.; Nagela, J.H. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
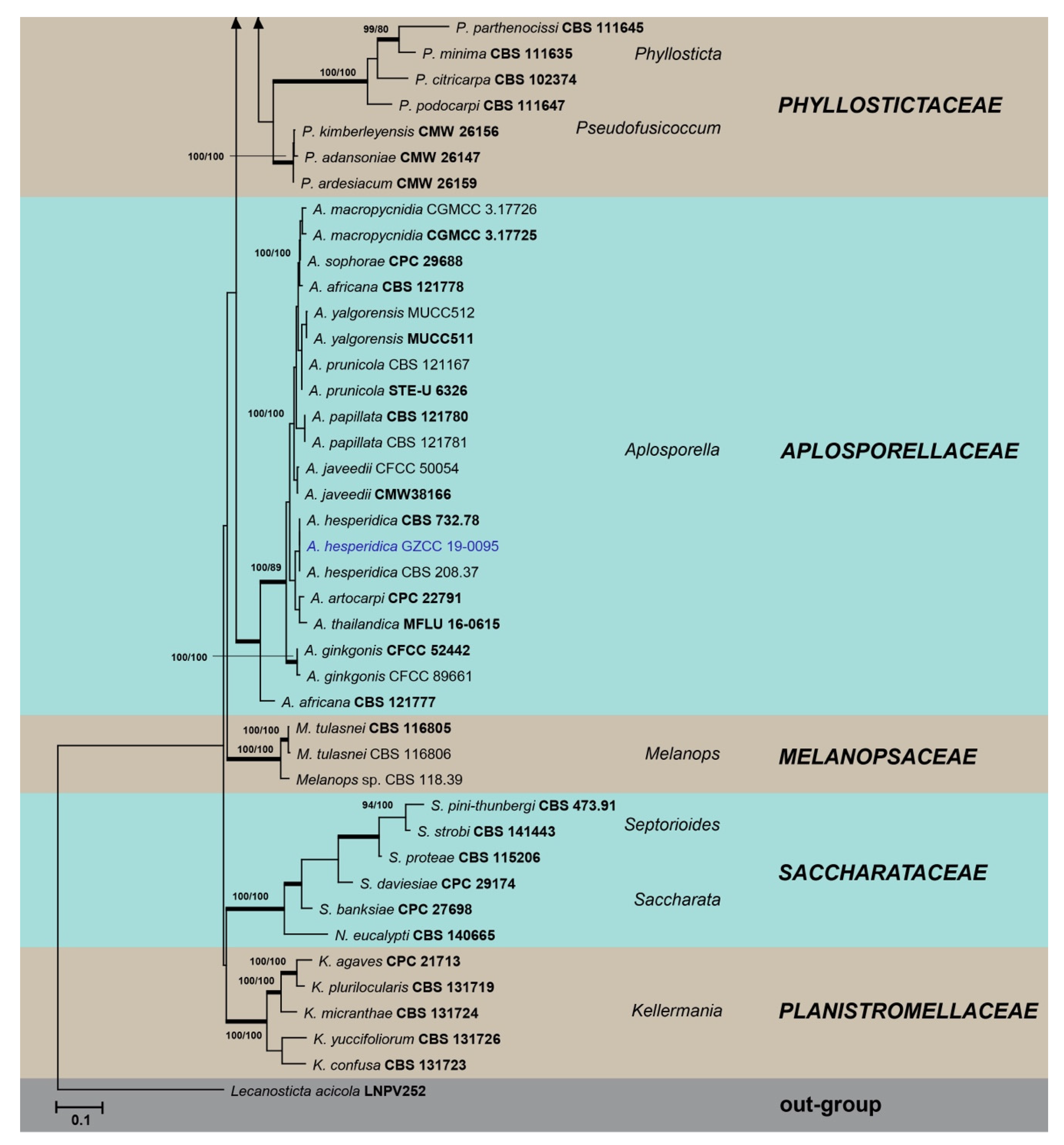
Figure 1.
Maximum likelihood tree based on analysis of combined ITS, LSU, and tef sequences of selected Botryosphaeriales taxa. Bootstrap values of ML, MP >75% are shown near nodes and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species, and novel taxa are in red. Ex-type strains are in bold. The tree is rooted to Lecanosticta acicola (LNPV252). The scale bar represents the expected number of nucleotide substitutions per site.
Figure 1.
Maximum likelihood tree based on analysis of combined ITS, LSU, and tef sequences of selected Botryosphaeriales taxa. Bootstrap values of ML, MP >75% are shown near nodes and branches in bold indicate BI probabilities >0.95. Isolates obtained in this study are in blue for known species, and novel taxa are in red. Ex-type strains are in bold. The tree is rooted to Lecanosticta acicola (LNPV252). The scale bar represents the expected number of nucleotide substitutions per site.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Botryosphaeriales species isolated and characterized in this study. The details of the type species of novel taxa are given in bold. N/A: No sequence available.
Table 1.
Botryosphaeriales species isolated and characterized in this study. The details of the type species of novel taxa are given in bold. N/A: No sequence available.
| Species Name | Isolate Number | Location | Date | ITS | LSU | tef |
|---|---|---|---|---|---|---|
| Aplosporella hesperidica | GZCC 19-0095 | Fanjing mountain, Tongren District | July 2018 | MZ781423 | MZ781490 | MZ852496 |
| Barriopsis tectonae | GZCC 19-0266 | Maolan natural reserve, Libo District | July 2017 | MZ781424 | MZ781491 | MZ852497 |
| Botryobambusa guizhouensis | CGMCC 3. 20348 | Forest Park, Chishui District | July 2019 | MZ781425 | MZ781492 | MZ852498 |
| GZCC 19-0734 | Forest Park, Chishui District | July 2019 | MZ781426 | MZ781493 | MZ852499 | |
| Botryosphaeria dothidea | GZCC 20-0524 | Huaxi wetland park, Guiyang District | April 2017 | MZ781435 | N/A | MZ852508 |
| GZCC 20-0525 | Xingyi Wanfenglin | June 2019 | MZ781436 | N/A | MZ852509 | |
| GZCC 20-0526 | Xingyi Wanfenglin | June 2019 | MZ781437 | N/A | MZ852510 | |
| GZCC 20-0527 | Suiyang broad water nature reserve | April 2018 | MZ781438 | N/A | MZ852511 | |
| GZCC 20-0528 | Forest Park, Chishui District | July 2019 | MZ781439 | N/A | MZ852512 | |
| GZCC 20-0529 | Suiyang broad water nature reserve | April 2018 | MZ781440 | N/A | MZ852513 | |
| GZCC 20-0530 | Suiyang broad water nature reserve | April 2018 | MZ781441 | N/A | MZ852514 | |
| GZCC 20-0531 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781442 | N/A | MZ852515 | |
| GZCC 20-0532 | Forest Park, Chishui District | July 2019 | MZ781443 | N/A | MZ852516 | |
| GZCC 20-0533 | Huaxi wetland park, Guiyang District | April 2017 | MZ781444 | N/A | MZ852517 | |
| GZCC 20-0534 | Suiyang broad water nature reserve | April 2018 | MZ781445 | N/A | MZ852518 | |
| GZCC 20-0535 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781446 | N/A | MZ852519 | |
| GZCC 20-0536 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781447 | N/A | MZ852520 | |
| GZCC 20-0537 | Huaxi wetland park, Guiyang District | April 2017 | MZ781448 | N/A | MZ852521 | |
| GZCC 20-0538 | Forest Park, Chishui District | July 2019 | MZ781449 | N/A | MZ852522 | |
| GZCC 20-0539 | Forest Park, Chishui District | July 2019 | MZ781450 | N/A | MZ852523 | |
| GZCC 20-0540 | Huaxi wetland park, Guiyang District | April 2017 | MZ781451 | N/A | MZ852524 | |
| GZCC 20-0541 | Forest Park, Chishui District | July 2019 | MZ781452 | N/A | MZ852525 | |
| GZCC 20-0542 | Forest Park, Chishui District | July 2019 | MZ781453 | N/A | MZ852526 | |
| GZCC 20-0543 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781454 | N/A | MZ852527 | |
| GZCC 20-0544 | Suiyang broad water nature reserve | April 2018 | MZ781455 | N/A | MZ852528 | |
| GZCC 20-0545 | Xingyi Wanfenglin | June 2019 | MZ781456 | N/A | MZ852529 | |
| GZCC 20-0546 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781457 | N/A | MZ852530 | |
| Diplodia mutila | GZCC 20-0547 | Xingyi Wanfenglin | June 2019 | MZ781459 | N/A | MZ852531 |
| GZCC 20-0548 | Huaxi wetland park, Guiyang District | April 2017 | MZ781460 | N/A | MZ852532 | |
| Di. neojuniperi | GZCC 19-0191 | Maolan natural reserve, Libo District | July 2017 | MZ781463 | N/A | MZ852533 |
| Di. pseudoseriata | GZCC 19-0072 | Xingyi Wanfenglin | June 2019 | MZ781461 | N/A | MZ852534 |
| Di. sapinea | GZCC 19-0075 | Suiyang broad water nature reserve | April 2018 | MZ781462 | N/A | MZ852535 |
| Di. seriata | GZCC 19-0548 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781458 | N/A | MZ852536 |
| Dothiorella sarmentorum | GZCC 19-0092 | Xingyi Wanfenglin | June 2019 | MZ781464 | N/A | MZ852537 |
| Do. yunnana | GZCC 19-0175 | Suiyang broad water nature reserve | April 2018 | MZ781465 | N/A | MZ852538 |
| Lasiodiplodia pseudotheobromae | GZCC 19-0162 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781466 | N/A | MZ852539 |
| GZCC 19-0184 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781467 | N/A | MZ852540 | |
| GZCC 19-0227 | Huaxi wetland park, Guiyang District | April 2017 | MZ781468 | N/A | MZ852541 | |
| GZCC 20-1575 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781469 | N/A | MZ852542 | |
| GZCC 19-0063 | Suiyang broad water nature reserve | April 2018 | MZ781470 | N/A | MZ852543 | |
| Neofusicoccum parvum | GZCC 19-0123 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781471 | N/A | MZ852544 |
| GZCC 19-0198 | Xingyi Wanfenglin | June 2019 | MZ781472 | N/A | MZ852545 | |
| GZCC 19-0218 | Xingyi Wanfenglin | June 2019 | MZ781473 | N/A | MZ852546 | |
| GZCC 19-2013 | Maolan natural reserve, Libo District | July 2017 | MZ781474 | N/A | MZ852547 | |
| GZCC 20-0549 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781475 | N/A | MZ852548 | |
| GZCC 20-0550 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781476 | N/A | MZ852549 | |
| GZCC 20-0551 | Xingyi Wanfenglin | June 2019 | MZ781477 | N/A | MZ852550 | |
| GZCC 20-0552 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781478 | N/A | MZ852551 | |
| GZCC 20-0553 | Huaxi wetland park, Guiyang District | April 2017 | MZ781479 | N/A | MZ852552 | |
| GZCC 20-0554 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781480 | N/A | MZ852553 | |
| GZCC 20-0555 | Suiyang broad water nature reserve | April 2018 | MZ781481 | N/A | MZ852554 | |
| GZCC 20-0556 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781482 | N/A | MZ852555 | |
| Sardiniella celtidis | GZCC 19-0248 | Xingyi Wanfenglin | June 2019 | MZ781427 | MZ781494 | MZ852500 |
| Sa. elliptica | CGMCC 3.20349 | Huaxi wetland park, Guiyang District | April 2017 | MZ781429 | MZ781496 | MZ852502 |
| GZCC 19-0245 | Xingyi Wanfenglin | June 2019 | MZ781431 | MZ781498 | MZ852504 | |
| GZCC 19-0262 | Maolan natural reserve, Libo District | July 2017 | MZ781430 | MZ781497 | MZ852503 | |
| Sa. guizhouensis | GZCC 19-0229 | Maolan natural reserve, Libo District | July 2017 | MZ781428 | MZ781495 | MZ852501 |
| Sphaeropsis citrigena | GZCC 20-0557 | Xingyi Wanfenglin | June 2019 | MZ781432 | MZ781499 | MZ852505 |
| Sp. guizhouensis | CGMCC 3.20352 | Maolan natural reserve, Libo District | July 2017 | MZ781433 | MZ781500 | MZ852506 |
| GZCC 19-0273 | Xingyi Wanfenglin | June 2019 | MZ781434 | MZ781501 | MZ852507 |
Table 2.
Details of gene regions, respective primer pairs, primer sequences, and PCR conditions used in the study.
Table 2.
Details of gene regions, respective primer pairs, primer sequences, and PCR conditions used in the study.
| Gene Region | Primers | Sequence 5′-3′ | Optimized PCR Protocols | References |
|---|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG | 94 °C: 5 min, (94 °C: 30 s, 55 °C: 50 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | White et al. [26] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| LSU | LR0R | ACCCGCTGAACTTAAGC | 94 °C: 5 min, (94 °C: 30 s, 54 °C: 50 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | Vilgalys and Hester [27] |
| LR5 | TCCTGAGGGAAACTTCG | |||
| tef | EF1-728F | CATCGAGAAGTTCGAGAAGG | 95 °C: 5 min, (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | Carbone and Kohn [28] |
| EF1-986R | TACTTGAAGGAACCCTTACC |
Table 3.
Botryosphaeriales taxa used in the phylogenetic analysis. The culture collection accession number for the type strains are given in bold. N/A: No sequence available.
Table 3.
Botryosphaeriales taxa used in the phylogenetic analysis. The culture collection accession number for the type strains are given in bold. N/A: No sequence available.
| Species Name | Isolate Number | ITS | LSU | tef |
|---|---|---|---|---|
| Aplosporellaceae | ||||
| Alanomyces indica | CBS 134264 | HF563622 | HF563623 | AB872219 |
| Aplosporella africana | CBS 121777 | KF766196 | EU101380 | EU101360 |
| A. africana | CBS 121778 | EU101316 | EU101381 | EU101361 |
| A. artocarpi | CPC 22791 | KM006450 | N/A | KM006481 |
| A. ginkgonis | CFCC 52442 | MH133916 | MH133933 | MH133950 |
| A. ginkgonis | CFCC 89661 | KM030583 | KM030590 | KM030597 |
| A. hesperidica | CBS 732.79 | KX464083 | KX464239 | N/A |
| A. hesperidica | CBS 208.37 | JX681069 | MH867398 | N/A |
| A. javeedii | CFCC 50054 | KP208840 | KP208843 | KP208846 |
| A. javeedii | CFCC 50052 | KP208838 | KP208841 | KP208844 |
| A. macropycnidia | CGMCC 3.17725 | KT343648 | N/A | KX011176 |
| A. macropycnidia | CGMCC 3.17726 | KT343649 | N/A | KX011177 |
| A. papillata | CBS 121780 | EU101328 | EU101383 | EU101373 |
| A. papillata | CBS 121781 | EU101329 | EU101384 | EU101374 |
| A. prunicola | CBS 121167 | KF766147 | KF766315 | N/A |
| A. prunicola | STE-U 6326 | EF564375 | EF564377 | N/A |
| A. sophorae | CPC 29688 | KY173388 | KY173482 | N/A |
| A. thailandica | MFLU 16-0615 | KX423536 | N/A | KX423537 |
| A. yalgorensis | MUCC511 | EF591926 | EF591943 | EF591977 |
| A. yalgorensis | MUCC512 | EF591927 | EF591944 | EF591978 |
| Botryosphaeriaceae | ||||
| Alanphillipsia aloeicola | CBS 138896 | KP004444 | KP004472 | N/A |
| A. euphorbiae | CPC 21628 | KF777140 | KF777196 | N/A |
| Barriopsis fusca | CBS 174.26 | EU673330 | DQ377857 | EU673296 |
| Ba. iraniana | IRAN1448C | FJ919663 | KF766318 | FJ919652 |
| Ba. iraniana | IRAN1449C | FJ919665 | N/A | FJ919654 |
| Ba. tectonae | MFLUCC 12-0381 | KJ556515 | N/A | KJ556516 |
| Botryobambusa fusicoccum | MFLUCC 11-0143 | JX646792 | JX646809 | JX646857 |
| Bo. fusicoccum | MFLUCC 11-0657 | JX646793 | JX646810 | JX646858 |
| Botryosphaeria agaves | MFLUCC 11-0125 | JX646791 | JX646808 | JX646856 |
| B. agaves | MFLUCC 10-0051 | JX646790 | JX646807 | JX646855 |
| B. corticis | CBS 119047 | DQ299245 | EU673244 | EU017539 |
| B. corticis | ATCC 22927 | DQ299247 | EU673245 | EU673291 |
| B. dothidea | CMW8000 | AY236949 | DQ377852 | AY236898 |
| B. dothidea | CBS 110302 | AY259092 | EU673243 | AY573218 |
| B. dothidea | CMW 25413 | KF766167 | KF766332 | EU101348 |
| B. dothidea | GZCC 16-0013 | KX447675 | N/A | KX447678 |
| B. dothidea | GZCC 16-0014 | KX447676 | N/A | KX447679 |
| B. dothidea | CGMCC 3.17722 | KT343254 | N/A | KU221233 |
| B. dothidea | CGMCC 3.17724 | KT343256 | N/A | KU221234 |
| B. dothidea | CGMCC 3.18744 | KX278002 | N/A | KX278107 |
| B. dothidea | CGMCC 3.18745 | KX278003 | N/A | KX278108 |
| B. dothidea | CGMCC3.20094 | MT327839 | N/A | MT331606 |
| B. dothidea | GZCC 19-0186 | MT327832 | N/A | MT331600 |
| B. dothidea | GZCC 19-0188 | MT327833 | N/A | MT331601 |
| B. fabicerciana | CMW27094 | HQ332197 | N/A | HQ332213 |
| B. fabicerciana | CMW27108 | HQ332200 | N/A | HQ332216 |
| B. fabicerciana | MFLUCC 10-0098 | JX646789 | JX646806 | JX646854 |
| B. fabicerciana | MFLUCC 11-0507 | JX646788 | JX646805 | JX646853 |
| B. kuwatsukai | CBS 135219 | KJ433388 | N/A | KJ433410 |
| B. kuwatsukai | LSP5 | KJ433395 | N/A | KJ433417 |
| B. kuwatsukai | CGMCC 3.18007 | KX197074 | N/A | KX197094 |
| B. kuwatsukai | CGMCC 3.18008 | KX197075 | N/A | KX197095 |
| B. qingyuanensis | CGMCC 3.18742 | KX278000 | N/A | KX278105 |
| B. qingyuanensis | CGMCC 3.18743 | KX278001 | N/A | KX278106 |
| B. ramosa | CBS 122069 | EU144055 | N/A | EU144070 |
| B. ramosa | CGMCC 3.18739 | KX277988 | N/A | KX278093 |
| B. ramosa | CGMCC 3.18740 | KX277989 | N/A | KX278094 |
| B. scharifii | IRAN1529C | JQ772020 | N/A | JQ772057 |
| B. scharifii | IRAN1543C | JQ772019 | N/A | JQ772056 |
| Cophinforma eucalypti | MFLUCC 11-0655 | JX646801 | JX646818 | JX646866 |
| C. mamane | CBS 117444 | KF531822 | DQ377855 | KF531801 |
| Diplodia africana | CBS 120835 | EF445343 | N/A | EF445382 |
| Di. africana | CBS 121104 | EF445344 | N/A | EF445383 |
| Di. agrifolia | CBS 132777 | JN693507 | N/A | JQ517317 |
| Di. agrifolia | UCROK1429 | JQ411412 | N/A | JQ512121 |
| Di. allocellula | CBS 130408 | JQ239397 | JQ239410 | JQ239384 |
| Di. allocellula | CBS 130410 | JQ239399 | JQ239412 | JQ239386 |
| Di. arengae | MFLU 17-2769 | MG762771 | N/A | MG762774 |
| Di. bulgarica | CBS 124135 | GQ923853 | N/A | GQ923821 |
| Di. bulgarica | CBS 124254 | GQ923852 | N/A | GQ923820 |
| Di. corticola | CBS 112546 | AY259100 | AY928051 | AY573227 |
| Di. corticola | CBS 112549 | AY259110 | EU673262 | DQ458872 |
| Di. crataegicola | MFLU 15-1311 | KT290244 | N/A | KT290248 |
| Di. cupressi | CBS 168.87 | DQ458893 | EU673263 | DQ458878 |
| Di. cupressi | CBS 261.85 | DQ458894 | EU673264 | DQ458879 |
| Di. eriobotryicola | BN 21 | MT587342 | N/A | MT592047 |
| Di. estuarina | CMW41231 | KP860831 | N/A | KP860676 |
| Di. estuarina | CMW41230 | KP860830 | N/A | KP860675 |
| Di. fraxini | CBS 136010 | KF307700 | N/A | KF318747 |
| Di. galiicola | MFLU 15-1310 | KT290245 | N/A | KT290249 |
| Di. gallae | CBS 213.25 | KX464092 | N/A | KX464566 |
| Di. malorum | CBS 124130 | GQ923865 | N/A | GQ923833 |
| Di. malorum | CBS 112554 | AY259095 | N/A | DQ458870 |
| Di. mutila | CBS 112553 | AY259093 | AY928049 | AY573219 |
| Di. mutila | CBS 230.30 | DQ458886 | AY928049 | DQ458869 |
| Di. mutila | CBS 121862 | KX464093 | N/A | KX464567 |
| Di. neojuniperi | CPC22753 | KM006431 | N/A | KM006462 |
| Di. olivarum | CBS 121887 | EU392302 | N/A | EU392279 |
| Di. olivarum | CBS 121886 | EU392297 | N/A | EU392274 |
| Di. pseudoseriata | CBS 124906 | EU080927 | MH874931 | EU863181 |
| Di. pseudoseriata | CBS 124907 | EU080922 | N/A | EU863179 |
| Di. pseudoseriata | CBS 124931 | FJ888460 | MH874935 | FJ888444 |
| Di. pseudoseriata | CBS 124933 | FJ888478 | N/A | FJ888446 |
| Di. pseudoseriata | CBS 140350 | KX833072 | N/A | KX833073 |
| Di. quercivora | CBS 133852 | JX894205 | N/A | JX894229 |
| Di. rosulata | CBS 116470 | EU430265 | DQ377896 | EU430267 |
| Di. rosulata | CBS 116472 | EU430266 | DQ377897 | EU430268 |
| Di. sapinea | CBS 393.84 | DQ458895 | DQ377893 | DQ458880 |
| Di. sapinea | CBS 109725 | DQ458896 | EU673270 | DQ458881 |
| Di. sapinea | CBS 124462 | GQ923858 | MH874896 | GQ923826 |
| Di. sapinea | CBS 124134 | HM036528 | N/A | GQ923851 |
| Di. sapinea | CBS 141915 | KT956270 | N/A | KU378605 |
| Di. scrobiculata | CBS 118110 | AY253292 | KF766326 | AY624253 |
| Di. scrobiculata | CBS 109944 | DQ458899 | EU673268 | DQ458884 |
| Di. scrobiculata | CBS 113423 | DQ458900 | EU673267 | DQ458885 |
| Di. seriata | CBS 112555 | AY259094 | AY928050 | AY573220 |
| Di. seriata | CBS 119049 | DQ458889 | EU673266 | DQ458874 |
| Di. subglobosa | CBS 124133 | GQ923856 | N/A | GQ923824 |
| Di. tsugae | CBS 418.64 | DQ458888 | DQ377867 | DQ458873 |
| Dothiorella acacicola | CPC26349 | KX228269 | KX228320 | KX228376 |
| Do. acericola | KUMCC 18-0137 | KY385661 | N/A | KY393212 |
| Do. alpina | CGMCC 3.18001 | KX499645 | N/A | KX499651 |
| Do. brevicollis | CBS 130411 | JQ239403 | JQ239416 | JQ239390 |
| Do. capri-amissi | CBS 121763 | EU101323 | N/A | EU101368 |
| Do. capri-amissi | CMW25404 | EU101324 | N/A | EU101369 |
| Do. casuarinae | CBS 120688 | DQ846773 | N/A | DQ875331 |
| Do. casuarinae | CBS 120690 | DQ846774 | N/A | DQ875333 |
| Do. citricola | ICMP 16828 | EU673323 | N/A | EU673290 |
| Do. citricola | ICMP 16827 | EU673322 | N/A | EU673289 |
| Do. dulcispinae | CBS 130413 | JQ239400 | JQ239413 | JQ239387 |
| Do. dulcispinae | CBS 121765 | EU101300 | KX464317 | EU101345 |
| Do. iranica | IRAN1587C | KC898231 | N/A | KC898214 |
| Do. juglandis | CBS 188.87 | EU673316 | DQ377891 | EU673283 |
| Do. lampangensis | MFLUCC 18-0232 | MK347758 | N/A | MK340869 |
| Do. longicollis | CBS 122068 | EU144054 | MH874718 | EU144069 |
| Do. longicollis | CBS 122066 | EU144052 | KX464311 | EU144067 |
| Do. magnoliae | CFCC 51563 | KY111248 | N/A | KY213687 |
| Do. mangifericola | IRAN1584C | MT587407 | N/A | MT592119 |
| Do. mangifericola | IRAN1545C | KC898221 | N/A | KX464614 |
| Do. mangifericola | CBS 121760 | KF766227 | N/A | EU101335 |
| Do. mangifericola | CBS 121761 | EU101293 | N/A | EU101338 |
| Do. moneti | MUCC 505 | EF591920 | EF591937 | EF591971 |
| Do. moneti | MUCC 507 | EF591922 | EF591939 | EF591973 |
| Do. plurivora | IRAN1557C | KC898225 | N/A | KC898208 |
| Do. plurivora | IRAN1537C | KC898226 | N/A | KC898209 |
| Do. pretoriensis | CBS 130404 | JQ239405 | JQ239418 | JQ239392 |
| Do. pretoriensis | CBS 130403 | JQ239406 | JQ239419 | JQ239393 |
| Do. prunicola | IRAN1541 | EU673313 | EU673232 | EU673280 |
| Do. rhamni | MFLUCC 14-0902 | KU246381 | KU246382 | N/A |
| Do. rhamni | BN 81 | MT587399 | N/A | MT592111 |
| Do. santali | MUCC 509 | EF591924 | EF591941 | EF591975 |
| Do. santali | MUCC 508 | EF591923 | EF591940 | EF591974 |
| Do. sarmentorum | IMI63581b | AY573212 | AY928052 | AY573235 |
| Do. sarmentorum | CBS 115038 | AY573206 | DQ377860 | AY573223 |
| Do. sarmentorum | CBS 128309 | HQ288218 | MH876298 | HQ288262 |
| Do. sarmentorum | CBS 128310 | HQ288219 | MH876299 | HQ288263 |
| Do. sarmentorum | CBS 141587 | KX357188 | N/A | KX357211 |
| Do. sarmentorum | MFLUCC 17-0242 | KY797637 | KY815014 | KY815020 |
| Do. sarmentorum | CBS 115045 | AY573202 | AY928053 | AY573222 |
| Do. sarmentorum | CBS 113188 | AY573198 | EU673230 | EU673278 |
| Do. sarmentorum | MFLUCC 17-0951 | MF398891 | N/A | MF398943 |
| Do. sarmentorum | CBS 140349 | KP205497 | N/A | KP205470 |
| Do. sarmentorum | CBS 188.87 | EU673316 | DQ377891 | EU673283 |
| Do. sarmentorum | IRAN1579C | KC898234 | N/A | KC898217 |
| Do. sarmentorum | IRAN1585C | KC898235 | N/A | KC898218 |
| Do. sarmentorum | IRAN1583C | KC898236 | N/A | KC898219 |
| Do. sarmentorum | IRAN1581C | KC898237 | N/A | KC898220 |
| Do. sarmentorum | MFLUCC 13-0497 | KJ742378 | N/A | KJ742381 |
| Do. sarmentorum | MFLUCC 13-0196 | KU234782 | N/A | KU234796 |
| Do. sarmentorum | DAR78992 | EU768874 | N/A | EU768881 |
| Do. sarmentorum | DAR78993 | EU768876 | N/A | EU768882 |
| Do. striata | ICMP 16824 | EU673320 | EU673240 | EU673287 |
| Do. striata | ICMP 16819 | EU673320 | EU673239 | EU673287 |
| Do. striata | DAR 80992 | KJ573643 | N/A | KJ573640 |
| Do. styphnolobii | JZB3150013 | MH880849 | N/A | MK069594 |
| Do. tectonae | MFLUCC 12-0382 | KM396899 | N/A | KM409637 |
| Do. thailandica | MFLUCC 11-0438 | JX646796 | JX646813 | JX646861 |
| Do. thripsita | BRIP 51876 | FJ824738 | N/A | KJ573639 |
| Do. ulmacea | CBS 138855 | KR611881 | KR611899 | KR611910 |
| Do. uruguayensis | CBS 124908 | EU080923 | MH874932 | EU863180 |
| Do. vinea gemmae | DAR81012 | KJ573644 | N/A | KJ573641 |
| Do. viticola | CBS 117009 | AY905554 | MH874565 | AY905559 |
| Do. viticola | CBS 117006 | AY905555 | EU673236 | AY905562 |
| Do. viticola | DAR80529 | HM009376 | N/A | HM800511 |
| Do. viticola | DAR80530 | HM009378 | N/A | HM800513 |
| Do. yunnana | CGMCC 3.17999 | KX499643 | N/A | KX499649 |
| Endomelanconiopsis endophytica | CBS 120397 | EU683656 | EU683629 | EU683637 |
| E. microspora | CBS 353.97 | EU683655 | KF766330 | EU683636 |
| Lasiodiplodia americana | CERC1960 | KP217058 | N/A | KP217066 |
| L. americana | CERC1961 | KP217059 | N/A | KP217067 |
| L. avicenniae | CMW41467 | KP860835 | N/A | KP860680 |
| L. avicenniarum | MFLUCC 17-2591 | MK347777 | MK347994 | MK340867 |
| L. brasiliense | CMM4015 | JX464063 | N/A | JX464049 |
| L. brasiliense | CMM2185 | KC484800 | N/A | KC481530 |
| L. bruguierae | CMW42480 | KP860832 | N/A | KP860677 |
| L. chonburiensis | MFLUCC 16-0376 | N/A | ||
| L. cinnamomi | CFCC 51997 | MG866028 | N/A | MH236799 |
| L. citricola | CBS 124707 | GU945354 | N/A | GU945340 |
| L. citricola | IRAN1521C | GU945353 | N/A | GU945339 |
| L. crassispora | CBS 118741 | DQ103550 | DQ377901 | EU673303 |
| L. crassispora | WAC12534 | DQ103551 | N/A | DQ103558 |
| L. crassispora | CBS 110492 | EF622086 | EU673251 | EF622066 |
| L. crassispora | CBS 121770 | EU101307 | N/A | EU101352 |
| L. crassispora | CBS 121771 | EU101308 | N/A | EU101353 |
| L. euphorbicola | CMM3609 | KF234543 | N/A | KF226689 |
| L. euphorbicola | IBL329 | KT247490 | N/A | KT247492 |
| L. gilanensis | CBS 124704 | GU945351 | N/A | GU945342 |
| L. gilanensis | IRAN1501C | GU945352 | N/A | GU945341 |
| L. gilanensis | CBS 128311 | HQ288225 | N/A | HQ288267 |
| L. gilanensis | UCD2199MO | HQ288226 | N/A | HQ288268 |
| L. gonubiensis | CBS 115812 | AY639595 | DQ377902 | DQ103566 |
| L. gonubiensis | CBS 116355 | AY639594 | EU673252 | DQ103567 |
| L. gravistriata | CMM4564 | KT250949 | N/A | KT250950 |
| L. hormozganensis | CBS 124709 | GU945355 | N/A | GU945343 |
| L. hormozganensis | CBS 124708 | GU945356 | N/A | GU945344 |
| L. iraniensis | CBS 124710 | GU945346 | MH874918 | GU945334 |
| L. iraniensis | CBS 124711 | GU945347 | N/A | GU945335 |
| L. iraniensis | CMM3610 | MT587430 | N/A | MT592142 |
| L. iraniensis | CMM0247 | MT587431 | N/A | MT592143 |
| L. laeliocattleyae | CBS 167.28 | KU507487 | N/A | KU507454 |
| L. laeliocattleyae | CBS 130992 | JN814397 | N/A | JN814424 |
| L. laeliocattleyae | CMM3611 | JN814401 | N/A | JN814428 |
| L. lignicola | CBS 134112 | JX646797 | N/A | KU887003 |
| L. lignicola | MFLUCC 11-0656 | JX646797 | JX646815 | KU887003 |
| L. lignicola | CGMCC 3.18061 | KX499889 | N/A | KX499927 |
| L. lignicola | CBS 342.78 | KX464140 | N/A | KX464634 |
| L. macrospora | CMM3833 | KF234557 | N/A | KF226718 |
| L. mahajangana | CBS 124927 | FJ900597 | N/A | FJ900643 |
| L. mahajangana | CBS 124925 | FJ900595 | N/A | FJ900641 |
| L. mahajangana | IBL352 | KT154759 | N/A | KT154753 |
| L. mahajangana | CMM1325 | KT154760 | N/A | KT008006 |
| L. mahajangana | CBS 137785 | KJ638317 | N/A | KJ638336 |
| L. mahajangana | BL184 | KJ638318 | N/A | KJ638337 |
| L. mahajangana | MFLUCC 16-0265 | MH275068 | MH260301 | MH412774 |
| L. margaritacea | CBS 122519 | EU144050 | N/A | EU144065 |
| L. margaritacea | CBS 122065 | EU144051 | N/A | EU144066 |
| L. marypalme | CMM2275 | KC484843 | N/A | KC481567 |
| L. marypalme | CMM2271 | KC484844 | N/A | KC481568 |
| L. mediterranea | CBS 137783 | KJ638312 | N/A | KJ638331 |
| L. mediterranea | ALG36 | KJ638314 | N/A | KJ638333 |
| L. parva | CBS 456.78 | EF622083 | KF766362 | EF622063 |
| L. parva | CBS 494.78 | EF622084 | EU673258 | EF622064 |
| L. parva | CBS 356.59 | AY343482 | N/A | EF445396 |
| L. plurivora | CBS 120832 | EF445362 | KX464356 | EF445395 |
| L. plurivora | CBS 121103 | AY343482 | KX464357 | EF445396 |
| L. pontae | CMM1277 | KT151794 | N/A | KT151791 |
| L. pontae | IBL14 | KT151794 | N/A | KT151791 |
| L. pseudotheobromae | CBS 116459 | EF622077 | EU673256 | EF622057 |
| L. pseudotheobromae | CBS 447.62 | EF622081 | MH869806 | EF622060 |
| L. rubropurpurea | CBS 118740 | DQ103553 | DQ377903 | EU673304 |
| L. rubropurpurea | WAC12536 | DQ103554 | N/A | DQ103572 |
| L. subglobosa | CMM3872 | KF234558 | N/A | KF226721 |
| L. subglobosa | CMM4046 | KF234560 | N/A | KF226723 |
| L. thailandica | CPC22795 | KJ193637 | N/A | KJ193681 |
| L. thailandica | CGMCC 3.17975 | KX499879 | MG321677 | KX499917 |
| L. thailandica | MFLUCC 18-0244 | MK347789 | N/A | MK340870 |
| L. theobromae | CBS 164.96 | AY640255 | EU673253 | AY640258 |
| L. theobromae | CBS 124.13 | DQ458890 | AY928054 | DQ458875 |
| L. theobromae | CBS 111530 | EF622074 | N/A | EF622054 |
| L. theobromae | CAA006 | DQ458891 | EU673254 | DQ458876 |
| L. theobromae | CBS 164.96 | AY640255 | EU673253 | AY640258 |
| L. venezuelensis | CBS 118739 | DQ103547 | DQ377904 | EU673305 |
| L. venezuelensis | WAC12540 | DQ103548 | N/A | DQ103569 |
| L. viticola | CBS 128313 | HQ288227 | KX098286 | HQ288269 |
| L. viticola | CBS 128315 | HQ288228 | N/A | HQ288270 |
| L. vitis | CBS 124060 | KX464148 | N/A | MN938928 |
| Lecanosticta acicola | LNPV252 | JX901755 | JX901844 | JX901639 |
| Macrophomina phaseolina | CBS 227.33 | KF951627 | DQ377906 | KF952000 |
| Neodeightonia palmicola | MFLUCC 10-0822 | HQ199221 | HQ199222 | N/A |
| N. phoenicum | CBS 122528 | EU673340 | EU673261 | EU673309 |
| N. subglobosa | CBS 448.91 | EU673337 | DQ377866 | EU673306 |
| Neofusicoccum arbuti | CBS 116131 | AY819720 | DQ377915 | KF531792 |
| N. arbuti | UW13 | AY819724 | N/A | KF531791 |
| N. arbuti | CBS 117453 | AY693976 | DQ377914 | AY693977 |
| N. arbuti | CMW13446 | DQ306263 | N/A | DQ306264 |
| N. australe | CMW6837 | AY339262 | KF766367 | AY339270 |
| N. australe | CMW6853 | AY339263 | N/A | AY339271 |
| N. brasiliense | CMM1338 | JX513630 | N/A | JX513610 |
| N. buxi | CBS 116.75 | KX464164 | KX464406 | KX464677 |
| N. cordaticola | CBS 123634 | EU821898 | MH874849 | EU821868 |
| N. cordaticola | CBS 123635 | EU821903 | KX464410 | EU821873 |
| N. cryptoaustrale | CMW23785 | FJ752742 | N/A | FJ752713 |
| N. eucalypticola | CBS 115679 | AY615141 | N/A | AY615133 |
| N. eucalypticola | CBS 115766 | AY615143 | N/A | AY615135 |
| N. eucalyptorum | CBS 115791 | AF283686 | N/A | AY236891 |
| N. eucalyptorum | CMW10126 | AF283687 | N/A | AY236892 |
| N. grevilleae | CBS 129518 | JF951137 | N/A | N/A |
| N. hellenicum | CERC1947 | KP217053 | N/A | KP217061 |
| N. hongkongensis | CERC 2973 | KX278052 | MF410096 | KX278157 |
| N. illicii | CGMCC 3.18311 | KY350150 | N/A | KY817756 |
| N. kwambonambiense | EU821900 | EU821900 | N/A | EU821870 |
| N. kwambonambiense | CMW14140 | EU821919 | N/A | EU821889 |
| N. lumnitzerae | CMW41469 | KP860881 | N/A | KP860724 |
| N. luteum | CBS 110299 | AY259091 | AY928043 | AY573217 |
| N. luteum | CBS 110497 | EU673311 | N/A | EU673277 |
| N. luteum | CMW41365 | KP860859 | N/A | KP860702 |
| N. macroclavatum | CBS 118223 | DQ093196 | N/A | DQ093217 |
| N. macroclavatum | WAC12446 | DQ093197 | N/A | DQ093218 |
| N. mangiferae | CBS 118532 | AY615185 | DQ377921 | DQ093221 |
| N. mangiferae | CMW7797 | AY615186 | N/A | DQ093220 |
| N. mediterraneum | CBS 121718 | GU251176 | MH874696 | GU251308 |
| N. mediterraneum | CBS 121558 | GU799463 | N/A | GU799462 |
| N. mediterraneum | CBS 113083 | KX464186 | KX464465 | KX464712 |
| N. mediterraneum | CBS 113089 | KX464199 | N/A | KX464727 |
| N. microconidium | CERC 3497 | KX278053 | MF410097 | KX278158 |
| N. nonquaesitum | CBS 126655 | GU251163 | MH875645 | GU251295 |
| N. nonquaesitum | PD301 | GU251164 | N/A | GU251296 |
| N. occulatum | CBS 128008 | EU301030 | MH876179 | EU339509 |
| N. occulatum | MUCC286 | EU736947 | N/A | EU339511 |
| N. parvum | CMW9081 | AY236943 | AY928045 | AY236888 |
| N. parvum | CBS 110301 | AY259098 | AY928046 | AY573221 |
| N. parvum | CBS 137504 | KJ657702 | N/A | KJ657715 |
| N. parvum | ALG9 | KJ657704 | N/A | KJ657721 |
| N. parvum | MFLUCC 15-0900 | KY856755 | N/A | KY856754 |
| N. pennatisporum | WAC13153 | EF591925 | EF591942 | EF591976 |
| N. pistaciae | CBS 595.76 | KX464163 | KX464404 | KX464676 |
| N. protearum | STE-U 4361 | AF196295 | N/A | N/A |
| N. protearum | STE-U 1775 | AF452539 | N/A | N/A |
| N. ribis | CMW7772 | AY236935 | N/A | AY236877 |
| N. ribis | CBS 121.26 | AF241177 | N/A | AY236879 |
| N. ribis | CBS 124923 | FJ900607 | N/A | FJ900653 |
| N. ribis | CBS 124924 | FJ900608 | N/A | FJ900654 |
| N. ribis | CMW14058 | EU821904 | N/A | EU821874 |
| N. ribis | CMW14060 | EU821905 | N/A | EU821875 |
| N. sinense | CGMCC 3.18315 | KY350148 | N/A | KY817755 |
| N. sinoeucalypti | CERC 2005 | KX278061 | MF410105 | KX278166 |
| N. stellenboschiana | CBS 110864 | AY343407 | KX464513 | AY343348 |
| N. terminaliae | CBS 125264 | GQ471802 | N/A | GQ471780 |
| N. ursorum | CBS 122811 | FJ752746 | MH874765 | FJ752709 |
| N. viticlavatum | CBS 112878 | AY343381 | MH874474 | AY343342 |
| N. viticlavatum | CBS 112977 | AY343380 | KX464528 | AY343341 |
| N. vitifusiforme | CBS 110887 | AY343383 | MH874455 | AY343343 |
| N. vitifusiforme | CBS 110880 | AY343382 | KX464475 | AY343344 |
| N. vitifusiforme | CBS 120081 | DQ923533 | MN162190 | KX464682 |
| N. vitifusiforme | CBS 121112 | EF445349 | N/A | EF445391 |
| Oblongocollomyces variabile | CBS 121774 | NR136994 | KX464536 | EU101357 |
| O. variabile | CMW25420 | EU101313 | N/A | EU101358 |
| O. variabile | CMW25421 | EU101314 | N/A | EU101359 |
| Phaeobotryon mamane | CPC 12264 | EU673331 | DQ377898 | EU673297 |
| P. mamane | CBS 122980 | EU673332 | EU673248 | EU673298 |
| Sardiniella celtidis | MFLUCC 17-981 | MF443249 | N/A | MF443248 |
| Sa. urbana | BL180 | KX379677 | KX379679 | KX379678 |
| Sa. urbana | BL181 | KX379680 | KX379682 | KX379681 |
| Sa. urbana | CBS 141580 | KX379674 | KX379676 | KX379675 |
| Sphaeropsis citrigena | ICMP16812 | EU673328 | EU673246 | EU673294 |
| Sp. citrigena | ICMP16818 | EU673329 | EU673247 | EU673295 |
| Sp. eucalypticola | MFLUCC 11-0579 | JX646802 | JX646819 | JX646867 |
| Sp. eucalypticola | MFLUCC 11-0654 | JX646803 | JX646820 | JX646868 |
| Sp. porosa | CBS 110496 | AY343379 | DQ377894 | AY343340 |
| Sp. porosa | CBS 110574 | AY343378 | DQ377895 | AY343339 |
| Sp. visci | CBS 100163 | EU673324 | N/A | EU673292 |
| Sp. visci | CBS 186.97 | EU673325 | DQ377868 | EU673293 |
| Tiarosporella graminis-karoo | CBS 118718 | KF531828 | DQ377939 | KF531807 |
| Melanopsaceae | ||||
| Melanops sp. | CBS 118.39 | FJ824771 | DQ377856 | FJ824776 |
| M. tulasnei | CBS 116805 | FJ824769 | KF766365 | FJ824774 |
| M. tulasnei | CBS 116806 | FJ824770 | FJ824765 | KX464644 |
| Phyllostictaceae | ||||
| Phyllosticta citricarpa | CBS 102374 | FJ538313 | DQ377877 | FJ538376 |
| P. minima | CBS 111635 | KF766215 | EU754194 | KF766433 |
| P. parthenocissi | CBS 111645 | EU683672 | DQ377876 | EU683653 |
| P. podocarpi | CBS 111647 | KF766217 | KF766383 | KF766434 |
| Pseudofusicoccum adansoniae | CMW 26147 | KF766220 | KF766386 | N/A |
| P. ardesiacum | CMW 26159 | EU144060 | KF766387 | N/A |
| P. kimberleyensis | CMW 26156 | EU144057 | KF766388 | N/A |
| Planistromellaceae | ||||
| Kellermania agaves | CPC 21713 | KF777164 | KF777217 | N/A |
| K. confusa | CBS 131723 | KF766174 | KF766344 | KF766405 |
| K. micranthae | CBS 131724 | KF766179 | NG042706 | KF766410 |
| K. plurilocularis | CBS 131719 | KF766181 | KF766351 | KF766412 |
| K. yuccifoliorum | CBS 131726 | KF766185 | KF766355 | KF766416 |
| Umthunziomyces hagahagensis | CPC 29917 | KY173472 | KY173561 | N/A |
| Saccharataceae | ||||
| Neoseptorioides eucalypti | CBS 140665 | KT950857 | KT950871 | KT950882 |
| Saccharata banksiae | CPC 27698 | KY173449 | KY173539 | KY173596 |
| S. daviesiae | CPC 29174 | KY173450 | KY173540 | N/A |
| S. proteae | CBS 115206 | KF766226 | DQ377882 | GU349030 |
| Septorioides pini-thunbergi | CBS 473.91 | NR145234 | KF251746 | N/A |
| S. strobi | CBS 141443 | KT884699 | KT884685 | KT884713 |
ALG: Personal culture collection of A. Berraf-Tebbal; ATCC: American Type Culture Collection, Virginia, USA; BL: Personal collection of B.T. Linaldeddu; BRIP: Culture collection, Queensland Department of Agriculture and Fisheries, Queensland, Australia; CAA: Personal culture collection of Artur Alves, Universidade de Aveiro, Portuga; CBS: CBS-KNAW Fungal Biodiversity Centre, Utrecht, The Netherlands; CERC: Culture collection of China Eucalypt Research Centre, Chinese Academy of Forestry, ZhanJiang, GuangDong, China; CFCC: China Forestry Culture Collection Center, Beijing, China; CGMCC: China General Microbiological Culture Collection Center; CMM: Culture Collection of Phytopathogenic Fungi “Prof. Maria Menezes”, Universidade Federal Rural de Pernambuco, Recife, Brazil; CMW: Tree Patholgy Co-operative Program, Forestry and Agricultural Biotechnology Institute, University of Pretoria, South Africa; CPC: Working collection of P.W. Crous, housed at CBS; DAR: Plant Pathology Herbarium, Orange Agricultural Institute, Forest Road, Orange. NSW 2800, Australia; GZCC: Guizhou Academy of Agricultural Sciences Culture Collection, GuiZhou, China; IBL: Personal culture collection of I.B.L. Coutinho; ICMP: International Collection of Microorganisms from Plants, Landcare Research, Aukland, New Zealand; IMI: International Mycological Institute, CBI-Bioscience, Egham, Bakeham Lane, UK; IRAN: Iranian Fungal Culture Collection, Iranian Research Institute of Plant Protection, Iran; KUMCC: Kunming University Culture Collection, Yunnan, China. MFLU: Mae Fah Luang University Herbarium Collection, Chiang Rai, Thailand; MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand; MUCC: Murdoch University Culture Collection, Murdoch, Australia; PD: Culture collection, University of California, Davis, USA; STE-U: Culture collection of the Department of Plant Pathology, University of Stellenbosch, South Africa; UCD: University of California, Davis, Plant Pathology Department Culture Collection; UCROK: Culture collection, University of Riverside, California, USA; WAC: Department of Agriculture, Western Australia Plant Pathogen Collection, South Perth, Western Australia.
Table 4.
Alignment details and comparison of MP, ML, and BI analyses results of each phylogenetic tree constructed in this study.
Table 4.
Alignment details and comparison of MP, ML, and BI analyses results of each phylogenetic tree constructed in this study.
| Character | Botryosphaeriales (Figure 1) | Botryosphaeria (Figure 10) | Diplodia (Figure 11) | Dothiorella (Figure 12) | Lasiodiplodia (Figure 13) | Neofusicoccum (Figure 14) | ||
|---|---|---|---|---|---|---|---|---|
| Number of base pairs in each gene region (including the gaps after alignment) | ITS (680), LSU (803), tef (374) | ITS (565), tef (265) | ITS (551), tef (331) | ITS (504), tef (343) | ITS (490), tef (344) | ITS (549), tef (253) | ||
| Number of isolates obtained in this study | 12 | 23 | 6 | 2 | 5 | 12 | ||
| Number of taxa originated from GenBank | 95 | 32 | 49 | 68 | 78 | 65 | ||
| Outgroup taxon | Lecanosticta acicula | Macrophomina phaseolina | Lasiodiplodia theobromae | Neofusicoccum parvum | Diplodia mutila | Dothiorella viticola | ||
| MP | Total number of characters | 1857 | 830 | 882 | 847 | 834 | 802 | |
| Constant characters | 954 | 690 | 688 | 578 | 592 | 565 | ||
| Variable/parsimony uninformative characters | 204 | 75 | 52 | 77 | 54 | 107 | ||
| Parsimony informative characters | 704 | 70 | 146 | 202 | 192 | 134 | ||
| Number of parsimonious trees obtained | 77 | 5 | 10 | 10 | 10 | 10 | ||
| Tree length (TL) | 4199 | 180 | 385 | 725 | 559 | 463 | ||
| (CI) | 0.393 | 0.911 | 0.644 | 0.568 | 0.612 | 0.646 | ||
| (RI) | 0.762 | 0.938 | 0.865 | 0.835 | 0.857 | 0.865 | ||
| (RC) | 0.300 | 0.854 | 0.557 | 0.475 | 0.524 | 0.559 | ||
| (HI) | 0.607 | 0.089 | 0.356 | 0.432 | 0.388 | 0.354 | ||
| ML | Final likelihood value | −22,205.648157 | −2130.358718 | −3417.346418 | −4843.109271 | −4183.640855 | −3665.812286 | |
| Number of distinct alignment patterns | 1096 | 212 | 297 | 390 | 293 | 310 | ||
| Percentage of undetermined characters or gaps | 26.52% | 7.31% | 11.38% | 21.84% | 8.52% | 9.63% | ||
| Base frequencies | A | 0.223450 | 0.206023 | 0.207760 | 0.210373 | 0.213076 | 0.201476 | |
| C | 0.259348 | 0.298959 | 0.297528 | 0.293067 | 0.286249 | 0.297204 | ||
| G | 0.288244 | 0.259350 | 0.261525 | 0.252521 | 0.258157 | 0.273045 | ||
| T | 0.228958 | 0.235668 | 0.233187 | 0.244039 | 0.242519 | 0.228274 | ||
| Substitution rates | AC | 1.251462 | 0.503912 | 0.982007 | 1.364862 | 0.729024 | 1.047005 | |
| AG | 2.644446 | 1.747452 | 3.644983 | 2.268590 | 2.724108 | 4.839820 | ||
| AT | 1.415897 | 1.056533 | 0.863841 | 1.240011 | 1.053880 | 1.265619 | ||
| CG | 1.353737 | 0.612519 | 1.681164 | 1.176948 | 0.817807 | 0.870409 | ||
| CT | 4.677551 | 3.387698 | 4.528302 | 4.552526 | 4.062263 | 8.746955 | ||
| GT | 1.000000 | 1.000000 | 1.000000 | 1.000000 | 1.000000 | 1.000000 | ||
| Gamma distribution rate parameter (alpha) | 0.286904 | 0.231276 | 0.174245 | 0.234867 | 0.223197 | 0.268848 | ||
| BI (model of each gene region) | ITS | SYM+I+G | GTR+I | GTR+I+G | HKY+I+G | SYM+I+G | GTR+I+G | |
| tef | HKY+G | GTR+G | HKY+I+G | GTR+G | HKY+I+G | HKY+G | ||
| Tree base ID | 28690 | 28685 | 28686 | 28687 | 28688 | 28689 | ||
| Reviewer access URL | http://purl.org/phylo/treebase/phylows/study/TB2:S28690?x-access-code=fe183dad30514d2dfbbfb8087dbbe53a&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28685?x-access-code=ac877963f1b00fed7de4920660ebc78f&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28686?x-access-code=9df25bff3d5c7c2faeb0029110c6675d&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28687?x-access-code=382c7e58f8f7e3b815b9ae27dbe6f639&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28688?x-access-code=e58fc08c292227a8965dd978203b449a&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28689?x-access-code=7152a6819a1dd4ef1e743d4db1e0240c&format=html | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dissanayake, A.J.; Chen, Y.-Y.; Cheewangkoon, R.; Liu, J.-K. Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. J. Fungi 2021, 7, 893. https://doi.org/10.3390/jof7110893
AMA Style
Dissanayake AJ, Chen Y-Y, Cheewangkoon R, Liu J-K. Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. Journal of Fungi. 2021; 7(11):893. https://doi.org/10.3390/jof7110893
Chicago/Turabian StyleDissanayake, Asha J., Ya-Ya Chen, Ratchadawan Cheewangkoon, and Jian-Kui Liu. 2021. "Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China" Journal of Fungi 7, no. 11: 893. https://doi.org/10.3390/jof7110893
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.