
Morphological Variety in Distoseptispora and Introduction of Six Novel Species
by
 ,
,
Jing Yang
1,2,3 ,
,
Ling-Ling Liu
1,4,*,
E. B. Gareth Jones
5,6,
Wen-Li Li
3,
Kevin D. Hyde
2 and
Zuo-Yi Liu
4 1
Guizhou Institute of Soil and Fertilizer, Guizhou Academy of Agricultural Sciences, Guiyang 550006, China
2
Center of Excellence in Fungal Research, Mae Fah Luang University, Chiang Rai 57100, Thailand
3
School of Life Science and Technology, University of Electronic Science and Technology of China, Chengdu 611731, China
4
Guizhou Key Laboratory of Agricultural Biotechnology, Guizhou Academy of Agricultural Sciences, Guiyang 550006, China
5
Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
6
Department of Botany and Microbiology, College of Science, King Saud University, Riyadh 11451, Saudi Arabia
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(11), 945; https://doi.org/10.3390/jof7110945
Submission received: 3 October 2021
/
Revised: 26 October 2021
/
Accepted: 4 November 2021
/
Published: 9 November 2021
(This article belongs to the Special Issue Fungal Biodiversity and Ecology 2.0)
Abstract
:Distoseptispora is one of the sporidesmium-like taxa with great variation in asexual morphology and delineation of species. Phylogenetic analyses of four gene regions LSU, ITS, TEF1α, and RPB2 revealed the placement of several sporidesmium-like species in Distoseptispora (Distoseptisporaceae, Distoseptisporales, Sordariomycetes), collected on submerged decaying twigs from streams in China and Thailand. Based on morphological examination and molecular DNA data, six new species, Distoseptispora amniculi, D. atroviridis, D. effusa, D. fusiformis, D. hyalina, and D. verrucosa, are proposed. Among them, D. hyalina is the first sexual morph confirmed in the genus. A new geographical record is reported for D. lignicola in China. Conidial length proved to be of less taxonomic significance for some Distoseptispora species, whereas the type of conidial septa is informative at species level.
1. Introduction
The dematiaceous sporidesmium-like hyphomycetes are common saprobes on decaying wood from terrestrial and freshwater habitats and distributed worldwide. They are characterized by holoblastic phragmoconidia produced on macronematous, proliferating or non-proliferating conidiophores or reduced to conidiogenous cells [1,2]. Considering the taxonomic value of euseptate and distoseptate conidia, absence or presence of conidiophores, conidiophore proliferation, and the shape of conidiogenous cells, several genera segregated from Sporidesmium sensu lato and allied genera were introduced, e.g., Ellisembia, Imicles, Penzigomyces, Polydesmus, Repetophragma, Sporidesmiella, and Stanjehughesia [1,3,4,5,6]. Later, with the incorporation of molecular data in phylogenetic studies, Sporidesmium sensu lato and morphologically similar genera were revealed to be polyphyletic mainly within Dothideomycetes and Sordariomycetes. Many of the morphological characters used to delimit the genera did not appear phylogenetically significant [7,8,9,10,11].
Distoseptispora is one of the sporidesmium-like genera introduced by Su et al. [9] with the type species D. fluminicola and the second species D. aquatica having long, cylindrical, distoseptate conidia. Yang et al. [10] emended the generic concept of Distoseptispora based on the morphological features of D. guttulata and D. suoluoensis, namely taxa with conspicuously longer conidiophores elongating percurrently and obclavate, rostrate, euseptate conidia. The generic circumscription was later expanded since more species with new morphological traits were described in the genus, such as D. palmarum [12] having polyblastic conidiogenous cells, D. appendiculata [13] and D. hydei [14] characterized by a mucilaginous conidial sheath, and D. martinii (Basionym: Acrodictys martinii) with muriform, transverse conidia [15]. Members in Distoseptispora are commonly found in freshwater habitats, and many new species have been discovered from China and Thailand in recent years [10,11,14,16,17,18,19,20,21,22,23,24,25]. Currently, more than 30 species are accepted in the genus but without a known sexual morph.
It is challenging to classify Distoseptispora species based on morphology alone because of the high morphological similarity to Sporidesmium and Ellisembia with euseptate and distoseptate conidia, respectively. Distoseptispora forms a monophyletic clade that is distinct from other sporidesmium-like taxa [9,10,13]. The genus was the only member belonging to Distoseptisporaceae and together with the monotypic Aquapteridosporaceae in Distoseptisporales [26,27].
During our survey of the taxonomy and diversity of freshwater fungi along a north–south gradient in the Asian/Australasian region [28], several sporidesmium-like taxa were collected from freshwater streams in China and Thailand. Using multi-gene loci of LSU, ITS, TEF1α, and RPB2 gene regions, the systematic placement of these collections revealed several Distoseptispora species. Based on the morphology and molecular evidence, we introduce six novel species in Distoseptispora including a sexual morph for D. hyalina and a new geographical record for D. lignicola in China. An updated generic description and backbone tree of Distoseptispora is provided and its generic delimitation discussed.
2. Materials and Methods
2.1. Collection and Examination of Specimens
Specimens of submerged decaying twigs were collected from streams in China and Thailand. Samples were taken to the laboratory in zip-lock plastic bags and incubated in plastic boxes lined with moistened tissue at room temperature for one week. Motic SMZ 168 Series (Motic, Xiamen, China) and Nikon SMZ-171 (Nikon, Tokyo, Japan) dissecting microscopes were used to observe the fungal colonies and fruiting bodies. Fungal structures were examined and photographed using a Nikon ECLIPSE 80i (Nikon, Tokyo, Japan) compound microscope fitted with a Canon 600D/70D (Canon, Tokyo, Japan) digital camera. Single spore isolations were made onto freshwater agar (WA) or potato dextrose agar (PDA) and germinated spores were transferred onto malt extract agar (MEA) or PDA following the method in Luo et al. [17]. Tarosoft Image Frame Work (Tarosoft, Nontha Buri, Thailand) was used for measurement and images used for figures were processed with Adobe Photoshop CC 2019 (Adobe Systems, San Jose, CA, USA) software. Herbarium specimens (dry wood with fungal material) were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand and herbarium of Cryptogams, Kunming Institute of Botany Academia Sinica (HKAS), Kunming, China. Axenic cultures were deposited in Mae Fah Luang University Culture Collection (MFLUCC) and Guizhou Culture Collection (GZCC). Facesoffungi and Index Fungorum numbers were registered as outlined in Jayasiri et al. [29] and Index Fungorum [30].
2.2. DNA Extraction, PCR Amplification, and Sequencing
Germinated spores were grown on MEA/PDA medium at 25 °C for one month. Fungal mycelium was scraped off using a sterilized scalpel and transferred to a 1.5 mL microcentrifuge tube for genomic DNA extraction. A Ezup Column Fungi Genomic DNA Purification Kit (Sangon Biotech, Shanghai, China) was used to extract DNA following the manufacturer’s instructions. DNA amplification was performed by polymerase chain reaction (PCR). LSU, SSU, ITS, TEF1α, and RPB2 gene regions were amplified using the primer pairs LR0R/LR5, NS1/NS4, ITS5/ITS4, 983F/2218R, and fRPB2-5F/fRPB2-7cR [31,32,33,34]. The amplifications were performed in a 25 μL reaction volume containing 9.5 μL ddH2O, 12.5 μL 2 × Taq PCR Master Mix with blue dye (Sangon Biotech, China), 1 μL of DNA template, and 1 μL of each primer (10 μM). The amplification condition for LSU, SSU, ITS, and TEF1α consisted of initial denaturation at 94 °C for 3 min, followed by 40 cycles of 45 s at 94 °C, 50 s at 56 °C, and 1 min at 72 °C, and a final extension period of 10 min at 72 °C. The amplification condition for RPB2 gene consisted of initial denaturation at 95 °C for 5 min, followed by 37 cycles of 15 s at 95 °C, 50 s at 56 °C, and 2 min at 72 °C, final extension period of 10 min at 72 °C. Purification and sequencing of PCR products were carried out by Shanghai Sangon Biological Engineering Technology and Services Co., Shanghai, China.
2.3. Phylogenetic Analyses
The ex-type and additional strains of Distoseptisporaceae species and related families (Acrodictyaceae, Aquapteridosporaceae, Bullimycetaceae, Cancellidiaceae, Papulosaceae, and Pseudostanjehughesiaceae) were selected in the phylogenetic analyses (Table 1). Four gene regions LSU, ITS, TEF1α, and RPB2 were used for the multi-gene analyses. Sequences were optimized manually to allow maximum alignment and maximum sequence similarity. The sequences were aligned using the online multiple alignment program MAFFT v.7 (Available online: http://mafft.cbrc.jp/alignment/server/ (accessed on 3 August 2021)) [35]. The alignments were checked visually and improved manually where necessary.
Maximum likelihood (ML), Bayesian inference (BI), and maximum parsimony (MP) analyses were employed to assess phylogenetic relationships. ML and BI analyses were performed through the CIPRES science Gateway V. 3.3 [36]. ML analyses were conducted with RAxML-HPC v. 8.2.12 [37] using a GTRGAMMA approximation with rapid bootstrap analysis followed by 1000 bootstrap replicates. For the BI approach, MrModeltest2 v. 2.3 [38] was used to infer the appropriate substitution model that would best fit the model of DNA evolution for the combined dataset. GTR + G + I substitution model was selected for LSU, ITS, TEF1α, and RPB2 partitions. BI analyses were performed in a likelihood framework as implemented in MrBayes 3.2.6 [39]. Six simultaneous Markov chains were run until the average standard deviation of split frequencies was below 0.01, with trees saved every 1000 generations. The first 25% of saved trees, representing the burn-in phase of the analysis, were discarded. The remaining trees were used for calculating posterior probabilities of recovered branches [40]. MP analyses were conducted with PAUP v. 4.0a167 [41]. A heuristic search was performed with the stepwise-addition option with 1000 random taxon addition replicates and tree bisection and reconnection branch swapping. All characters were unordered and of equal weight and gaps were treated as missing data. Maxtrees were unlimited, branches of zero length were collapsed, and all multiple, equally parsimonious trees were saved. Clade stability was assessed using a bootstrap analysis with 1000 replicates, each with 10 replicates of random stepwise addition of taxa [42].
The resulting trees were printed with FigTree v. 1.4.4 and the layout was created in Adobe Illustrator 2019 (Adobe Systems, San Jose, CA, USA). Sequences generated in this study were deposited in GenBank (Table 1).
3. Phylogenetic Results
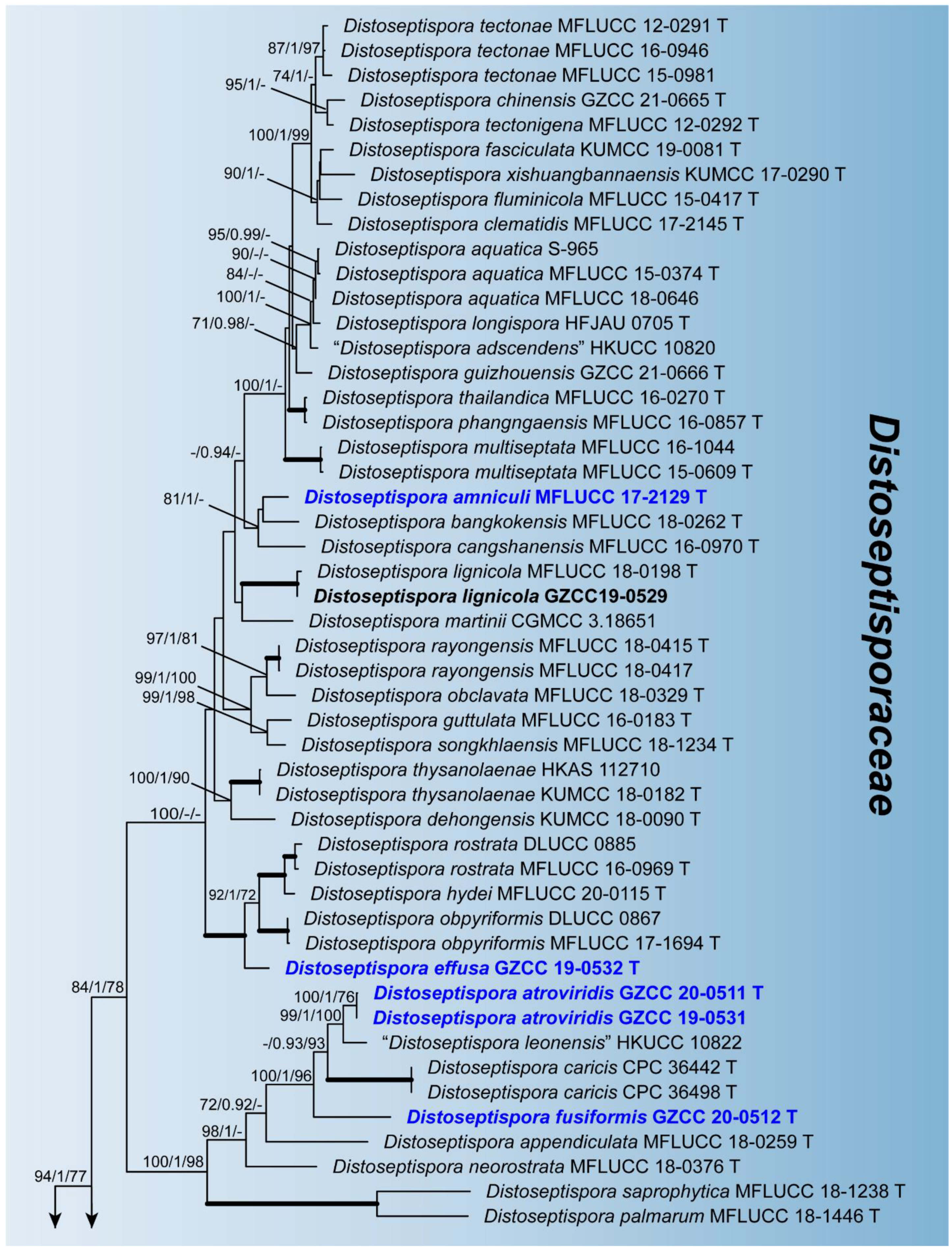
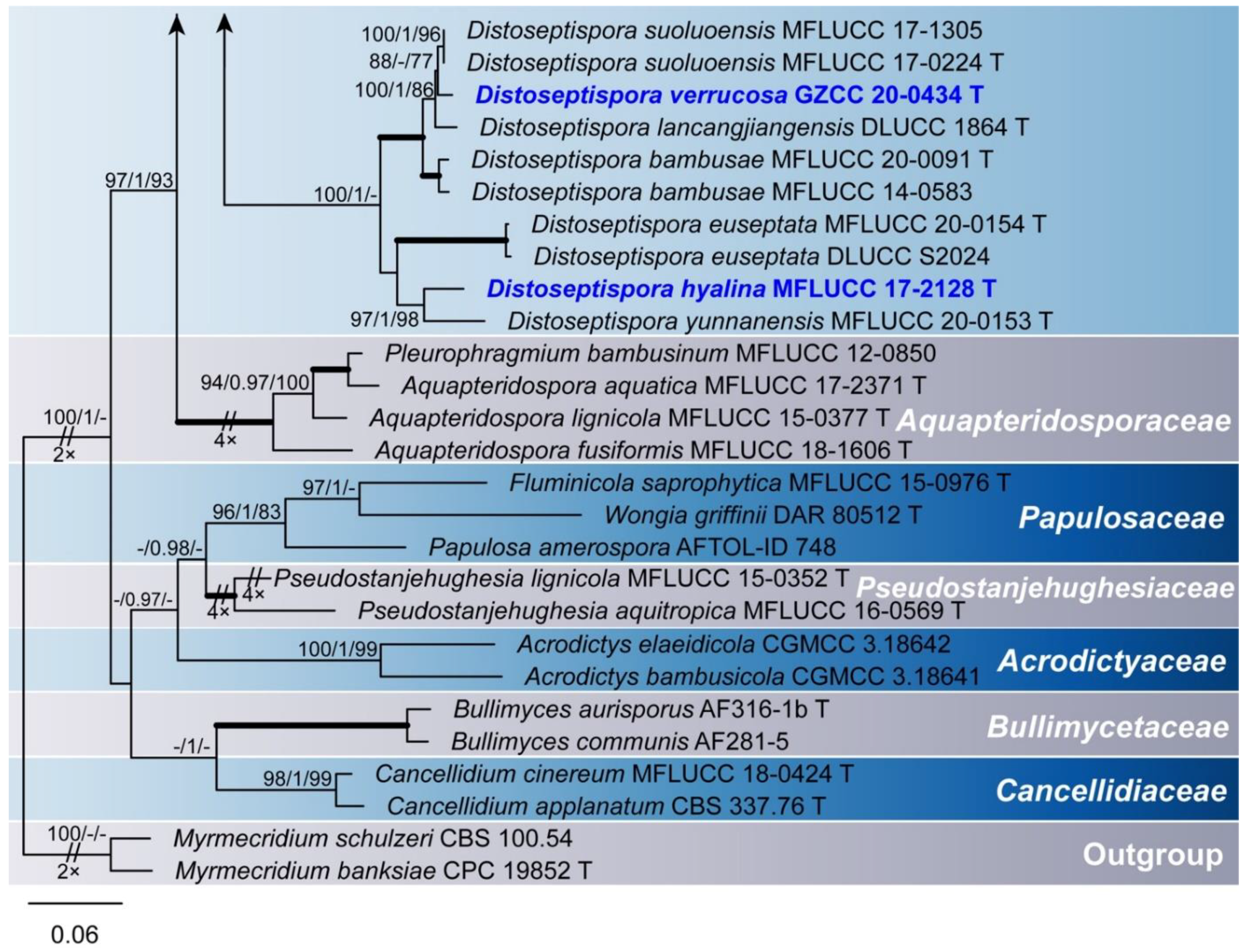
Phylogenetic relationships of seven Distoseptispora species were assessed in the combined analysis using four gene regions of 76 strains representing 61 species in Distoseptisporaceae and related families (Acrodictyaceae, Aquapteridosporaceae, Bullimycetaceae, Cancellidiaceae, Papulosaceae, and Pseudostanjehughesiaceae). The analyzed alignment consisted of combined LSU (1–858 bp), ITS (859–1522 bp), TEF1α (1523–2432 bp), and RPB2 (2433–3490 bp) sequence data including gaps. Myrmecridium schulzeri (CBS 100.54) and Myrmecridium banksiae (CPC 19852) served as outgroup taxa. The best scoring RAxML tree is shown in Figure 1. The analyzed ML, MP, and Bayesian trees were similar in topology and did not conflict significantly. Distoseptispora was resolved as a monophyletic clade. Our eight strains nested within the genus representing seven species. Distoseptispora amniculi (MFLUCC 17-2129) clustered as sister taxon to D. bangkokensis (MFLUCC 18-0262) but with weak support. Distoseptispora effusa (GZCC 19-0532) formed a distinct clade sister to the clade containing D. hydei (MFLUCC 20-0115), D. rostrata (MFLUCC 16-0969 and DLUCC 0885), and D. obpyriformis (MFLUCC 17-1694 and DLUCC 0867) with strong statistical support (100% ML BS/1.0 PP/100% MP BS). Distoseptispora atroviridis (GZCC 20-0511 and GZCC 19-0531), based on two strains, grouped with “D. leonensis” (HKUCC 10822) and was close to D. fusiformis (GZCC 20-0512). Distoseptispora verrucosa (GZCC 20-0434) grouped with D. suoluoensis (MFLUCC 17-1305 and MFLUCC 17-0224), D. lancangjiangensis (DLUCC 1864), and D. bambusae (MFLUCC 20-0091 and MFLUCC 14-0583), in a sister group of D. euseptata (MFLUCC 20-0154 and DLUCC S2024), D. hyalina (MFLUCC 17-2128), and D. yunnanensis (MFLUCC 20-0153), forming a statistically well supported clade by ML and BI analyses. Members of this clade have similar conidial morphology except D. hyalina in sexual stage. The strain GZCC 19-0529 positioned sister to the ex-type strain (MFLUCC 18-0198) of D. lignicola with identical LSU sequences and three base pair differences in ITS sequences, and therefore is identified as D. lignicola based on the morphology and molecular evidence.
4. Taxonomy
Distoseptispora K.D. Hyde, McKenzie, and Maharachch., Fungal Diversity 80: 402 (2016).
Index Fungorum number: IF551833; Facesoffungi number: FoF01755.
Sexual morph: Ascomata solitary or gregarious, immersed to semi-immersed, perithecial, subglobose to ellipsoidal, dark brown, ostiolate, with a short neck. Ostiole periphysate. Ascomatal wall coriaceous, 2-layered, outer layer consisting of multi-rows of brown, thick-walled, polyhedral cells of textura angularis, inner layer comprising multi-rows of pale brown to hyaline, thin-walled, elongated cells of textura angularis or taxtura prismatica. Paraphyses sparse, persistent, septate, hyaline, tapering towards the apex, constricted at the septa. Asci unitunicate, 8-spored, cylindrical, pedicellate, obtuse at the apex, apex with a non-amyloid apical annulus. Ascospores overlapping, uniseriate, fusiform, hyaline, 0–3-septate, smooth-walled, guttulate, thin-walled, with a mucilaginous sheath. Asexual morph: Hyphomycetous. Colonies effuse, hairy, velvety, olivaceous to dark brown. Mycelium mostly immersed, composed of branched, septate, smooth, pale brown hyphae. Conidiophores macronematous, mononematous, erect, single or in groups or fasciculate, septate, unbranched, straight or slightly flexuous, smooth, olivaceous to brown, cylindrical, rounded or truncate at the apex, sometimes elongating percurrently, rarely reduced to conidiogenous cells. Conidiogenous cells mostly monoblastic, sometimes polyblastic, integrated, determinate, terminal, cylindrical or clavate with flared apex. Conidia acrogenous, solitary, cylindrical or obclavate, rostrate, ellipsoidal, obovoid to fusiform, subhyaline, olivaceous, dark green, brown or yellowish-brown to reddish-brown, euseptate or distoseptate, rarely muriform, truncate at base, sometimes indeterminate in length or producing a secondary conidium, septal pore and mucilaginous sheath present or absent.
Type species—Distoseptispora fluminicola McKenzie, H.Y. Su, Z.L. Luo, and K.D. Hyde
Notes: Hyde et al. [11] provided the family description for the monotypic Distoseptisporaceae. The diagnosis of the sexual morph in the family was based on Miyoshiella triseptata, which was associated with “Distospetispora adscendens” (as Ellisembia adscendens) in the same collection [7,43]. However, neither the cultural study nor molecular data has proved their connection. We prefer to treat Miyoshiella triseptata as a possible sexual morph of sporidesmium-like taxa. The sexual description here is based on Distoseptispora hyalina. Distoseptispora martinii is unique in the genus by transverse ellipsoid or subglobose, muriform conidia [15]. Additional collections and further molecular evidence are needed to confirm its taxonomy.
Distoseptispora amniculi J. Yang and K.D. Hyde, sp. nov., Figure 2.
Index Fungorum number: IF558670; Facesoffungi number: FoF10250.
Etymology: referring to the collecting site of a small stream.
Holotype: MFLU 21-0138.
Saprobic on submerged decaying wood in a freshwater habitat. Asexual morph: Colonies on wood effuse, hairy, dark brown, scattered or in small groups, glistening, usually retiform. Mycelium partly immersed, partly superficial, composed of septate, smooth-walled, pale brown to hyaline hyphae. Conidiophores macronematous, mononematous, erect, solitary or caespitose, straight or flexuous, cylindrical, rounded at the apex, smooth-walled, septate, unbranched, grayish brown, 90–180 × 3–4.5 µm ( = 125 × 4 µm, n = 20). Conidiogenous cells monoblastic, integrated, terminal, determinate, cylindrical, pale brown, rounded and darkened at the apex, sometimes elongating percurrently. Conidia acrogenous, obclavate, rostrate, olivaceous brown, grayish brown or mid brown, paler towards the apex, (7–)12–24-septate, 85–167 × 9–11.8 µm ( = 120 × 10 µm, n = 20), smooth-walled, truncate and darkened at the base, sometimes with a secondary conidium. Sexual morph: undetermined.
Culture characteristics: conidia germinating on PDA within 24 h and germ tubes produced from both ends. Colonies growing on PDA slow growing, reaching 10–15 mm in a month at 25 °C in natural light, circular, with dense, gray mycelium in the middle, darker of the inner ring, with sparser, white mycelium of the outer ring on the surface, in reverse dark brown to black with smooth margin.
Material examined: Thailand, Trat Province, Amphoe Ko Chang, 12°7.98′ N, 102°37.98′ E, on decaying wood submerged in a freshwater stream, 27 April 2017, YZ Lu, YJT 26-2 (MFLU 21-0138, holotype); ex-type living culture MFLUCC 17-2129; additional sequence, SSU: MZ868766.
Notes: Distoseptispora amniculi is similar to D. neorostrata and D. rostrata in the relative long conidiophores (more than 80 µm long) and obclavate, rostrate, distoseptate conidia. Distoseptispora neorostrata [13] can be distinguished from the present species in having truncate apex to the conidiophores and dark and wider conidia (109–147 × 13–15 µm); D. rostrata [17] has olivaceous to pale brown conidia that are 115–155 × 9–11 µm. Conidiophores of D. amniculi are longer than that in D. neorostrata (90–180 µm vs. 93–117 µm) and D. rostrata (90–180 µm vs. 82–126 μm). In the multi-gene phylogenetic tree (Figure 1), D. amniculi clustered with D. bangkokensis. They have distoseptate conidia and rounded apex of conidiophores. Distoseptispora amniculi is well distinguishable from D. bangkokensis [44] by longer conidiophores (90–180 µm vs. 37–55 µm) and shorter conidia (85–167 µm vs. 400–568 µm). Comparison of the LSU, ITS, and SSU sequences of D. amniculi and D. bangkokensis showed 99.75% (798/800bp), 92.91% (485/522bp including 11bp of gaps), and 99.44% (893/898bp) sequence identity, respectively.
Distoseptispora atroviridis J. Yang and K.D. Hyde, sp. nov., Figure 3.
Index Fungorum number: IF558671; Facesoffungi number: FoF10252.
Etymology: referring to the dark green conidia.
Holotype: HKAS 112616.
Saprobic on submerged decaying wood in a freshwater habitat. Asexual morph: Colonies on wood effuse, dark brown to black, scattered or in small groups, glistening. Mycelium mostly immersed, composed of septate, smooth-walled, brown to hyaline hyphae. Conidiophores macronematous, fasciculate, loosely compact, erect, straight or slightly flexuous, cylindrical, wider and truncate at the apex, smooth-walled, septate, unbranched, pale grayish brown, slightly paler at the apical cell, 100–167 × 2.7–4 µm ( = 124.5 × 3.5 µm, n = 30), 4.7–6.9 µm wide at the apex. Conidiogenous cells monoblastic, integrated, terminal, determinate, sometimes elongating percurrently, flared, pale grayish brown. Conidia acrogenous, solitary, ellipsoidal to obovoid, dark green, subhyaline at the basal cell, 6-septate, 31–43 × 13–20 µm ( = 39 × 18 µm, n = 40), smooth-walled, guttulate, truncate at the base, sometimes released with part of conidiogenous cells. Sexual morph: Undetermined.
Culture characteristics: conidia germinating on PDA within 24 h and swollen germ tubes produced from both ends. Colonies growing on PDA reaching 5–10 mm in two weeks at 25 °C in the dark, with dense, velvety, dark green mycelium on the surface; in reverse dark green with entire margin.
Material examined: China, Guizhou Province, Chishui City, Sidonggou Waterfall, 25°27.38′ N, 107°39.85′ E, on submerged decaying twig in a freshwater stream, 11 July 2019, J Yang, CS 40-1 (HKAS 112616, holotype); ex-type living culture GZCC 20-0511; ibid, near 28°25′ N, 106°0′ E, at an altitude of 500 m, on submerged decaying wood in a freshwater stream, 16 July 2019, LL Liu, CS 1-4-2 (GZAAS 20-0426, paratype); living culture GZCC 19-0531 (additional sequence, SSU: MW134695).
Notes: Distoseptispora atroviridis is well distinguishable among other species of the genus by fasciculate, synnematous-like conidiophores, flared conidiogenous cells and ellipsoidal to obovoid, dark green, 6-septate conidia with paler to subhyaline basal cell. Distoseptispora atroviridis resembles some Phragmocephala species, such as P. atra, P. elliptica, P. hughesii, and P. garethjonesii, in having loosely to compactly fasciculate conidiophores aggregated at the base, clavate, flared conidiogenous cells sometimes elongating percurrently and ellipsoidal to obovoid conidia that secedes rhexolyticly [6,45]. However, Phragmocephala species differ from D. atroviridis by brown conidia with thickened and darkened bands. Molecular analyses revealed the placement of Phragmocephala in Melanommataceae (Pleosporales, Dothideomycetes) distinct from D. atroviridis [45,46]. In our phylogenetic tree, D. atroviridis (GZCC 20-0511 and GZCC 19-0531) was sister to “Distoseptispora leonensis” HKUCC10822 (99% ML BS/1.0 PP/100% MP BS, Figure 1), but they are distinguishable in morphology. “Distoseptispora leonensis” was characterized by solitary, mid to dark brown conidiophores with up to three successive proliferations [47] while D. atroviridis has fasciculate, loosely compact, longer but narrower conidiophores (100–167 × 2.7–4 µm vs. 70–120 × 4.5–8 µm) rarely percurrently proliferating. Conidiogenous cells are cylindrical and slightly narrower at the apex in “D. leonensis” [47] while those are flared with wider apex in D. atroviridis. The former species has fusiform or rostrate, mid to dark brown, 9–17-septate conidia with tapering apical cells [47] while the latter has ellipsoidal to obovoid, dark green, 6-septate, smaller conidia (31–43 × 13–20 µm vs. 45–90 × 15–18 µm).
Distoseptispora effusa L.L. Liu and Z.Y. Liu, sp. nov., Figure 4.
Index Fungorum number: IF558406; Facesoffungi number: FoF09863.
Etymology: referring to the effuse colonies.
Holotype: GZAAS 20-0427.
Saprobic on decaying wood in a freshwater habitat. Asexual morph: Colonies on natural substrates superficial, effuse, dark brown to black, hairy, velvety. Mycelium mostly immersed, consisting of branched, hyaline to pale brown, smooth, septate hyphae. Conidiophores macronematous, mononematous, erect, solitary or in small groups, cylindrical, dark brown, 5–14-septate, straight or slightly curved, smooth, 72–171 × 5–6.5 μm ( = 104.5 × 5.5 μm, n = 15), rounded at the apex. Conidiogenous cells monoblastic, integrated, terminal, determinate, cylindrical, brown, sometimes elongating percurrently, darkened at the rounded apex and percurrent loci. Conidia acrogenous, solitary, obclavate, rostrate, smooth-walled, olivaceous brown to dark brown, sometimes slightly paler at the apex, straight or slightly curved, 4–9-distoseptate, 35.5–113 × 7–12.5 μm ( = 71 × 10 μm, n = 20), truncate and darkened at the base. Sexual morph: Undetermined.
Cultural characteristics: conidia germinated on WA within 24 h and germ tube produced from the apex. Colonies on PDA reaching 15–20 mm diam. after 2 weeks at 25 °C in dark, circular, with fluffy, dense, dark olivaceous brown aerial mycelium on the surface; in reverse dark brown with entire margin.
Material examined: China, Guizhou Province, Chishui City, Chishui river basin, near 28°25′ N, 106°0′ E, at an altitude of 500 m, on submerged decaying wood in a freshwater stream, July 2019, LL Liu, CS 3-7 (GZAAS 20-0427, holotype); ex-type living culture GZCC 19-0532; additional sequences, SSU: MW134696.
Notes: Phylogenetically, Distoseptispora effusa (GZCC 19-0532) nested within Distoseptispora and formed a distinct clade sister to the clade containing D. rostrata, D. hydei, and D. obpyriformis (Figure 1). Morphologically, D. effusa is similar to D. rostrata [17] with percurrently elongate conidiophores and obclavate, distoseptate conidia but differs by longer conidiophores (72–171 μm vs. 82–126 μm) and shorter conidia (35.5–113 μm vs. 115–155 μm). Distoseptispora hydei [14] is distinguished by fusiform to obpyriform, light olivaceous to brown conidia. Distoseptispora obpyriformis [17] has obclavate to obpyriform, olivaceous to dark brown conidia that are shorter but wider than D. effusa (53–71 × 12–16 μm vs. 35.5–113 × 7–12.5 μm).
Distoseptispora fusiformis J. Yang and K.D. Hyde, sp. nov., Figure 5.
Index Fungorum number: IF558672; Facesoffungi number: FoF10253.
Etymology: referring to the fusiform conidia.
Holotype: HKAS 112617.
Saprobic on submerged decaying wood in a freshwater habitat. Asexual morph: Colonies on wood effuse, hairy, dark brown to black, scattered or in small groups, glistening. Mycelium mostly immersed, composed of septate, smooth-walled, brown to hyaline hyphae. Conidiophores macronematous, mononematous, erect, straight or slightly flexuous, cylindrical, slightly tapering towards the apex, smooth-walled, septate, unbranched, pale to dark brown, slightly paler at the apical cells, 40–110 × 3.5–5.8 µm ( = 86 × 4.6 µm, n = 20). Conidiogenous cells monoblastic, integrated, terminal, determinate, sometimes elongating percurrently, cylindrical, brown. Conidia acrogenous, solitary, ellipsoidal to fusiform, dark olivaceous brown to dark brown, pale brown at both ends, 6–8-septate, 35–52 × 13.5–22 µm ( = 42 × 18.5 µm, n = 30), smooth-walled, guttulate, truncate at the base, with an obconical basal cell. Sexual morph: Undetermined.
Culture characteristics: conidia germinating on PDA within 24 h and swollen germ tubes produced from both ends. Colonies growing on PDA reaching 5–10 mm in two weeks at 25 °C in dark, circular, with velvety, dark olivaceous brown mycelium on the surface; in reverse dark brown with filiform margin.
Material examined: China, Guizhou Province, Chishui City, Sidonggou Waterfall, 25°27.38′ N, 107°39.85′ E, on submerged decaying twig in a freshwater stream, 11 July 2019, J Yang, CS 40-2 (HKAS 112617, holotype); ex-type living culture GZCC 20-0512; additional sequence, SSU: MZ868768.
Notes: Distoseptispora fusiformis can be distinguished from other species in the genus by relatively long conidiophores with truncate apex and ellipsoidal to broadly fusiform, 6–8-septate, dark olivaceous brown to dark brown conidia with paler polar cells. Distoseptispora fusiformis was collected on the same material as D. atroviridis but they are distinct in morphology and phylogeny. Distoseptispora atroviridis can be distinguished from D. fusiformis by fasciculate or loosely compact conidiophores, trapezoidal conidiogenous cells and ellipsoidal to obovoid, dark green conidia. Conidiophores and conidia of D. atroviridis are longer and slightly smaller than those in D. fusiformis (100–167 µm vs. 40–110 µm; 31–43 × 13–20 µm vs. 35–52 × 13.5–22 µm), respectively. Comparing the LSU, ITS, TEF1α, and RPB2 sequences of D. atroviridis and D. fusiformis showed 96.07% (32 bp differences), 90.21% (55 bp differences), 93.77% (56 bp differences), and 89.56% (114 bp differences) sequence similarity, respectively.
Distoseptispora hyalina J. Yang and K.D. Hyde, sp. nov., Figure 6.
Index Fungorum number: IF558673; Facesoffungi number: FoF10249.
Etymology: referring to the hyaline conidia.
Holotype: MFLU 21-0137.
Saprobic on decaying submerged wood in a freshwater habitat. Sexual morph: Ascomata solitary or gregarious, immersed to semi-immersed, perithecial, 150–200 μm high, 120–190 μm diam., subglobose to ellipsoidal, dark brown, ostiolate, with a short neck erumpent through host surface. Ostiole periphysate. Ascomatal wall coriaceous, 20–31 μm thick, 2-layered; outer layer consisting of multi-rows of brown, thick-walled, polyhedral cells of textura angularis, inner layer comprising multi-rows of pale brown to hyaline, thin-walled, elongated cells of textura angularis or taxtura prismatica. Paraphyses sparse, persistent, septate, hyaline, tapering towards the apex, c. 4–7 μm wide near the base, constricted at the septa. Asci 145–190 × 8–11 µm ( = 165 × 9.8 µm, n = 20), cylindrical, with a short pedicel, obtuse at the apex, 8-spored, apex with a non-amyloid apical annulus. Ascospores (19.5–)23–26(–28.5) × 4.5–7 µm ( = 25 × 6 µm, n = 30), overlapping, uniseriate, fusiform, straight, rarely slightly curved, hyaline, 0–3-septate, smooth-walled, guttulate, thin-walled, with a mucilaginous sheath. Asexual morph: Undetermined.
Culture characteristics: conidia germinating on PDA within 24 h. Germ tubes produced from both ends. Colonies on PDA reaching 10–15 mm diam. after 2 weeks at 25 °C in natural light, with dense mycelium on the surface, gray in the middle, dark grayish green of the inner ring, and grayish green of the outer ring; in reverse dark olivaceous green with entire margin.
Material examined: Thailand, Trat Province, Amphoe Ko Chang, 12°7.98′ N, 102°37.98′ E, on decaying wood submerged in a freshwater stream, 27 April 2017, YZ Lu, YJT 26-1 (MFLU 21-0137, holotype); ex-type living culture MFLUCC 17-2128; additional sequence, SSU: MZ868765.
Notes: Distoseptispora hyalina is the first sexual morph reported in the genus based on molecular DNA data. Distoseptispora hyalina resembles Sporidesmium thailandense in possessing cylindrical, pedicellate asci with a non-amyloid apical annulus and obliquely uniseriate, fusiform, hyaline ascospores with a mucilaginous sheath. However, D. hyalina possesses immersed to semi-immersed, erumpent ascomata, larger brown cells of ascomatal wall and 3-septate ascospores when mature while S. thailandense has immersed ascomata with compact layers of cells of the peridium that is undifferentiated from host tissue and 3–4-septate ascospores [10,48]. In addition, D. hyalina differs from S. thailandense by the smaller asci (145–190 × 8–11 µm vs. 160–220 × 11–14 µm) and narrower ascospores but similar in length (4.5–7 µm wide vs. 8–10 µm wide). Distoseptispora hyalina is similar to Miyoshiella triseptata in having a non-amyloid apical annulus of asci and fusiform, 3-septate ascospores with comparable dimension (20–25 × 5–7 µm) [43], but Miyoshiella triseptata is distinguished by carbonaceous, papillate ascomata, shorter and broader asci (90–110 × 12–17 µm) and hyaline to light yellowish brown ascospores lacking a sheath [43].
In the phylogenetic analyses, Distoseptispora hyalina was sister to D. yunnanensis with good support (97% ML BS/1.0 PP/98% MP BS, Figure 1). The LSU, ITS, TEF1α, and RPB2 sequence identity of D. hyalina and D. yunnanensis was 95.05% (807/849bp including 4bp of gaps), 94.59% (525/555bp including 9bp of gaps), 96.66% (898/929bp), and 93.41% (1035/1108bp), respectively.
Distoseptispora hyalina were colonized close to D. amniculi on the same twig in sexual and asexual stages, respectively. However, they are separate taxa based on molecular evidence.
Distoseptispora lignicola D.F. Bao, Z.L. Luo, H.Y. Su, and K.D. Hyde, Fungal Diversity 99: 487 (2019) Figure 7.
Index Fungorum number: IF555641, Facesoffungi number: FoF05413.
Cultural characteristics: conidia germinating on WA within 24 h and germ tube produced from both ends. Colonies on PDA reaching about 30 mm diam. after 3 weeks at 25 °C in dark, circular, with dense, velvety, whitish brown mycelium on the surface, uneven in the middle; in reverse dark brown at the entire margin, rimous in the middle.
Material examined: China, Guizhou Province, Chishui City, Chishui river basin, near 28°25′ N, 106°0′ E, at an altitude of 500 m, on submerged decaying wood in a freshwater stream, July 2019, LL Liu, CS 1-5-1 (GZAAS 20-0424), living culture GZCC 19-0529.
Notes: Our collection GZAAS 20-0424 matches the original diagnosis of the holotype of Distoseptispora lignicola (MFLU 18-1458) well [13]. Comparison of their LSU and ITS sequences showed 100% and 99.43% (526/529bp) similarity, respectively. We therefore identify our two collections as D. lignicola and report a new geographical record of this species in China.
Distoseptispora verrucosa J. Yang and K.D. Hyde, sp. nov., Figure 8.
Index Fungorum number: IF558674; Facesoffungi number: FoF10251.
Etymology: referring to the verrucose conidia.
Holotype: HKAS 112652.
Saprobic on submerged decaying wood in a freshwater habitat. Asexual morph: Colonies on wood effuse, hairy, dark brown to black, scattered or in small groups, glistening. Mycelium partly immersed, partly superficial, composed of septate, smooth-walled, pale brown to hyaline hyphae. Conidiophores macronematous, mononematous, erect, solitary or caespitose, straight or flexuous, cylindrical, rounded and usually darkened at the apex, smooth-walled, septate, unbranched, dark brown, slightly paler at the upper part, 92–250 × 4.7–6.3 µm ( = 162 × 5.7 µm, n = 20). Conidiogenous cells monoblastic, integrated, terminal, determinate, sometimes percurrently proliferating, cylindrical, brown. Conidia acrogenous, solitary, obclavate, rostrate, upper part tapering towards the apex, olivaceous brown, becoming paler at the apex, 6–8-septate, 41–63 × 8.8–12.6 µm ( = 51.5 × 10.8 µm, n = 30), verrucose, guttulate, truncate with a darkened scar at the base. Sexual morph: Undetermined.
Cultural characteristics: conidia germinating on PDA within 24 h and germ tubes produced from both ends. Colonies on PDA reaching 5–10 mm diam. after 14 days at 25 °C, in natural light, circular, with velvety, dense, grayish brown mycelium on the surface with entire margin; in reverse dark brown.
Material examined: China, Guizhou Province, Dushan District, 25°57.9′ N, 107°39′ E, on decaying wood submerged in a freshwater stream, 26 Aug 2017, J Yang, SG 75-1 (HKAS 112652, holotype), ex-type living culture GZCC 20-0434; additional sequence, SSU: MZ868767.
Notes: In the phylogenetic analyses, Distoseptispora verrucosa clustered with D. lancangjiangensis, D. suoluoensis, and D. bambusae. Their systematic placement is correlated to the highly similar morphological characters in having dark brown conidiophores with rounded apex and narrowly obclavate, rostrate, euseptate, verrucose conidia with a dark scar at the base. They differ in conidial color and dimensions: D. verrucosa has the smallest, olivaceous brown conidia 41–63 × 8.8–12.6 µm; D. suoluoensis [10] has the largest, dark yellowish brown to dark olivaceous brown conidia (65–)80–125(–145) × 8–13 μm usually with a secondary conidium; D. bambusae [23] and D. lancangjiangensis [44] possess brown or olivaceous conidia measuring 45–74 × 5.5–9.5 µm and 64–84 × 9–10 μm, respectively. The length of conidiophores of D. verrucosa (92–250 µm) and D. suoluoensis (80–250 µm) are comparable and longer than that in D. bambusae (40–96 µm) and D. lancangjiangensis (144–204 µm). Phylogenetically, D. verrucosa was sister to D. suoluoensis, showing 99.88% (838/839 bp), 98.65% (513/520 bp), 98.44% (884/898 bp), and 98.25% (1015/1033 bp) sequence identity of LSU, ITS, TEF1α, and RPB2 sequences, respectively. Distoseptispora verrucosa resembles Sporidesmium tengii and S. tunicatum with long conidiophores and obclavate, rostrate, euseptate conidia. However, conidiophores in the latter two species are truncate at the apex and conidia lack a basal darkened scar. Sporidesmium tengii [6] differs by shorter conidiophores (60–100 µm) and smooth-walled, smaller conidia (45–50 × 7–8.5 µm). Sporidesmium tunicatum [49] has shorter conidiophores (110–180 µm), verrucose but slightly longer conidia (43–75 × 9–13 µm) with an apical sheath. Molecular DNA data are not available for S. tengii and S. tunicatum.
5. Discussion
The establishment of Distoseptispora [9] was based on morphology and molecular DNA data. More than 30 species in the genus are supported by sequence data. The genus forms monophyletic clade (Figure 1) distinct from other sporidesmium-like taxa. Members in the genus mainly occur in the asexual morph, forming effuse, hairy colonies on decaying wood, bamboo culms, plant stems, rachis, and fallen leaves from terrestrial and freshwater habitats. The morphological concept of Distoseptispora is macronematous, mononematous, solitary or fasciculate conidiophores, sometimes elongating percurrently and rarely reduced to conidiogenous cells; monoblastic or polyblastic, cylindrical or clavate conidiogenous cells; conidia are cylindrical, obclavate, rostrate, ellipsoidal, obovoid or fusiform, subhyaline, olivaceous, dark green, brown or yellowish-brown to reddish-brown, euseptate or distoseptate, rarely muriform, sometimes born a secondary conidium, with or without septal pore and mucilaginous sheath. The characters delineating the genus Distoseptispora also cover the criteria of Ellisembia and Sporidesmium. Ellisembia was segregated from the widely circumscribed Sporidesmium by Subramanian [1] based on the septal type. However, given the non-taxonomic value of euseptate and distoseptate conidia among sporidesmium-like species, Su et al. [9] recognized the distoseptate Ellisembia as a synonym of the redefined Sporidesmium sensu stricto which forms a robust monoclade and accommodates species with distoseptate/euseptate, obclavate or subcylindrical conidia, and conidiophores with or without percurrent extensions. In this study, we follow Hyde et al. [12] in treating them as separate taxa because of their unclear relationship due to the absence of molecular DNA data from their type species.
Several sexual morphs have been linked to Ellisembia and Sporidesmium through cultural and/or molecular studies. Ellisembia folliculata (sexual morph: Lecythothecium duriligni) [50] and E. aurea [12] differ from the sexual morph D. hyalina by versicolorous ascospores and position within Chaetosphaeriaceae (Chaetosphaeriales, Sordariomycetes). Sporidesmium thailandense [10,48] and S. lignicola [13] can be distinguished by brown ascomata with a hyaline neck and compact, elongated cells of the ascomatal wall, outer layer undifferentiated from host tissue, and their systematic placement in Sporidesmiaceae (Sporidesmiales, Sordariomycetes). The morphology of sexual morphs of sporidesmium-like genera along with molecular DNA data characterizes their identification although they have similar asexual morphs.
It is challenging to identify some Distoseptispora species with highly similar morphology, such as those of D. multiseptata, D. phangngaensis, and D. xishuangbannaensis. Still, they can be well separated by molecular DNA data [10,16,18]. Some Distoseptispora species have a wide range of conidial length, for example, conidia of D. multiseptata are 95–290 µm long of the holotype but 300–700 µm long in the additional collection; those in D. phangngaensis 165–350 µm long and D. tectonigena 83–360 µm long [9,10,16]. The indeterminate conidial length may depend on the incubation period. Thus, conidial length is less taxonomically informative in separating some Distoseptispora species. The type of septa, however, is proven to have no taxonomic significance for generic delimitation of sporidesmium-like taxa, but it is informative at the species level [10,14].
Su et al. [9] accepted two former Ellisembia species E. adscendens and E. leonensis in Distoseptispora that were not validly published, based on the non-type strains “Distoseptispora adscendens” HKUCC 10820 and “D. leonensis” HKUCC 10822 lacking associated morphology. Ellisembia adscendens was initially introduced as Sporidesmium adscendens forming elongated black patches on the pileus of Polyporus versicolor Fr. (No. 1345) which was collected on the underside of timber in the Falkland Islands [51]. Ellisembia adscendens is a widespread species. It is similar to E. vaga [52] but differs by wider conidia. Ellisembia adscendens and E. vaga highly resemble a group of morphologically indistinguishable species in Distoseptispora, e.g., D. multiseptata and D. phangngaensis. They are probably members of Distoseptispora because of the typical Distoseptispora morphology in having short conidiophores and subcylindrical long conidia. On the other hand, the large number of specimens of Ellisembia adscendens may be a complex comprising several separate taxa. Ellisembia leonensis [47] is characterized by relatively long conidiophores with percurrent extensions and fusiform to rostrate, distoseptate conidia. It matches the morphological concept of both Distoseptispora and Sporidesmium sensu stricto. At present, we avoid reassigning E. adscendens and E. leonensis in Distoseptispora until their systematic placement is confirmed by molecular data from type materials or resolved by epitypifications.
Author Contributions
Conceptualization: J.Y., K.D.H. and Z.-Y.L.; methodology: J.Y. and L.-L.L.; formal analysis and investigation: J.Y., L.-L.L. and W.-L.L.; resources: K.D.H., Z.-Y.L. and E.B.G.J.; writing—original draft preparation, J.Y.; writing—review and editing, J.Y., E.B.G.J., K.D.H., L.-L.L., Z.-Y.L. and W.-L.L.; supervision, E.B.G.J., K.D.H. and Z.-Y.L.; funding acquisition, L.-L.L. and Z.-Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Foundation of Key Laboratory of Microbial Resources Collection and Preservation, Ministry of Agriculture and Rural Affairs (Project no. KLMRCP2021-08).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated for this study can be found in the NCBI database.
Acknowledgments
E.B.G.J. is supported under the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Saudi Arabia. J.Y. is grateful to Shaun Pennycook for corrections on the Latin names of the novel taxa. Jian-Kui Liu is thanked for the corrections on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Subramanian, C.V. A reassessment of Sporidesmium (hyphomycetes) and some related taxa. Proc. Indian Nat. Sci. Acad. 1992, 58, 179–190. [Google Scholar]
- Seifert, K.A.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; pp. 1–997. [Google Scholar]
- Kirk, P. New or interesting microfungi VI. Sporidesmiella gen. nov. (Hyphomycetes). Trans. Br. Mycol. Soc. 1982, 79, 479–489. [Google Scholar] [CrossRef]
- Hernández-Gutiérrez, A.; Sutton, B.C. Imimyces and Linkosia, two new genera segregated from Sporidesmium sensu lato, and redescription of Polydesmus. Mycol. Res. 1997, 101, 201–209. [Google Scholar] [CrossRef]
- Shoemaker, R.A.; Hambleton, S. “Helminthosporium” asterinum, Polydesmus elegans, Imimyces, and allies. Can. J. Bot. 2001, 79, 592–599. [Google Scholar] [CrossRef]
- Wu, W.P.; Zhuang, W.Y. Sporidesmium, Endophragmiella and related genera from China. Fungal Divers. Res. Ser. 2005, 15, 1–351. [Google Scholar]
- Réblová, M. Studies in Chaetosphaeria sensu lato III. Umbrinosphaeria gen. nov. and Miyoshiella with Sporidesmium anamorphs. Mycotaxon 1999, 71, 13–43. [Google Scholar]
- Shenoy, B.D.; Jeewon, R.; Wu, W.P.; Bhat, D.J.; Hyde, K.D. Ribosomal and RPB2 DNA sequence analyses suggest that Sporidesmium and morphologically similar genera are polyphyletic. Mycol. Res. 2006, 110, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.-K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.-Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2017, 17, 591–616. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Monkai, J.; Boonmee, S.; Ren, G.-C.; Wei, D.-P.; Phookamsak, R.; Mortimer, P.E. Distoseptispora hydei sp. nov. (Distoseptisporaceae), a novel lignicolous fungus on decaying bamboo in Thailand. Phytotaxa 2020, 459, 93–107. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 1–21. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.Y. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsa-Ard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Pers. Mol. Phylogeny Evol. Fungi 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Song, H.-Y.; El Sheikha, A.F.; Zhai, Z.-J.; Zhou, J.-P.; Chen, M.-H.; Huo, G.-H.; Huang, X.-G.; Hu, D.-M. Distoseptispora longispora sp. nov. from freshwater habitats in China. Mycotaxon 2020, 135, 513–523. [Google Scholar] [CrossRef]
- Sun, Y.; Goonasekara, I.D.; Thambugala, K.M.; Jayawardena, R.S.; Wang, Y.; Hyde, K.D. Distoseptispora bambusae sp. nov. (Distoseptisporaceae) on bamboo from China and Thailand. Biodivers. Data J. 2020, 8, e53678. [Google Scholar] [CrossRef]
- Dong, W.; Hyde, K.D.; Jeewon, R.; Doilom, M.; Yu, X.D.; Wang, G.N.; Liu, N.G.; Hu, D.M.; Nalumpang, S.; Zhang, H. Towards a natural classification of annulatascaceae-like taxa Ⅱ: Introducing five new genera and eighteen new species from freshwater. Mycosphere 2021, 12, 1–88. [Google Scholar] [CrossRef]
- Li, W.-L.; Liu, Z.-P.; Zhang, T.; Dissanayake, A.J.; Luo, Z.-L.; Su, H.-Y.; Liu, J.-K. Additions to Distoseptispora (Distoseptisporaceae) associated with submerged decaying wood in China. Phytotaxa 2021, 520, 75–86. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Hyde, K.D.; Bao, D.-F.; Hongsanan, S.; Chethana, K.W.T.; Yang, J.; Suwannarach, N. Evolution of freshwater Diaporthomycetidae (Sordariomycetes) provides evidence for five new orders and six new families. Fungal Divers. 2021, 107, 71–105. [Google Scholar] [CrossRef]
- Hyde, K.D.; Fryar, S.; Tian, Q.; Bahkali, A.H.; Xu, J. Lignicolous freshwater fungi along a north–south latitudinal gradient in the Asian/Australian region; can we predict the impact of global warming on biodiversity and function? Fungal Ecol. 2016, 19, 190–200. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.-C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 3 August 2021).
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols—A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. MrModeltest2 v. 2.3 (Program for Selecting DNA Substitution Models Using PAUP*); Evolutionary Biology Centre: Uppsala, Sweden, 2008. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larget, B.; Simon, D.L. Markov Chasin Monte Carlo Algorithms for the Bayesian Analysis of Phylogenetic Trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Shoemaker, R.A.; White, G.P. Lasiosphaeria caesariata with Sporidesmium hormiscioides and L. triseptata with Sporidesmium adscendens. Sydowia 1985, 38, 278–283. [Google Scholar]
- Shen, H.W.; Bao, D.F.; Hyde, K.D.; Su, H.Y.; Bhat, D.J.; Luo, Z.L. New species and records of Distoseptispora (Distoseptisporaceae) from freshwater habitats in China and Thailand. MycoKeys 2021, in press. [Google Scholar] [CrossRef]
- Su, H.-Y.; Udayanga, D.; Luo, Z.-L.; Manamgoda, D.; Zhao, Y.-C.; Yang, J.; Liu, X.-Y.; McKenzie, E.H.C.; Zhou, D.-Q.; Hyde, K.D. Hyphomycetes from aquatic habitats in Southern China: Species of Curvularia (Pleosporaceae) and Phragmocephala (Melannomataceae). Phytotaxa 2015, 226, 201–216. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Ellis, M.B. Clasterosporium and some allied Dematiaceae-Phragmosporae: I. Mycol. Pap. 1958, 70, 1–89. [Google Scholar]
- Zhang, H.; Dong, W.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Hongsanan, S.; Bhat, D.J.; Al-Sadi, A.M.; Zhang, D. Towards a natural classification of Annulatascaceae-like taxa: Introducing Atractosporales ord. nov. and six new families. Fungal Divers. 2017, 85, 75–110. [Google Scholar] [CrossRef]
- Mena-Portales, J.; Hernández-Restrepo, M.; Guarro, J.; Minter, D.W.; Gené, J. New species of Penzigomyces, Sporidesmium and Stanjehughesia from plant debris in Spain. Nova Hedwig. 2016, 103, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Réblová, M.; Winka, K. Generic concepts and correlations in ascomycetes based on molecular and morphological data: Lecy-thothecium duriligni gen. et sp. nov. with a Sporidesmium anamorph, and Ascolacicola austriaca sp. nov. Mycologia 2001, 93, 478–493. [Google Scholar] [CrossRef]
- Berkeley, M. XXXII.—Notice of some Fungi collected by C. Darwin, Esq., during the expedition of H. M. Ship Beagle. J. Nat. Hist. 1840, 4, 291–293. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew, Commonwealth Mycological Institute: Surrey, UK, 1971; pp. 117–118. [Google Scholar]
Figure 1.
Maximum likelihood majority rule consensus tree for Distoseptisporaceae and related families using LSU, ITS, TEF1α, and RPB2 sequence data. Bootstrap support values for maximum likelihood (ML) and maximum parsimony (MP) greater than 70% and Bayesian posterior probabilities greater than 0.90 are indicated above branches as ML BS/PP/MP BS. The scale bar represents the expected number of changes per site. The tree is rooted with Myrmecridium schulzeri (CBS 100.54) and Myrmecridium banksiae (CPC 19852). Ex-type strains are indicated with T. The new collections are in bold with new taxa in blue. Branches with 100% ML BS, 1.0 PP, and 100% MP BS are thickened. Families are indicated as colored blocks.
Figure 1.
Maximum likelihood majority rule consensus tree for Distoseptisporaceae and related families using LSU, ITS, TEF1α, and RPB2 sequence data. Bootstrap support values for maximum likelihood (ML) and maximum parsimony (MP) greater than 70% and Bayesian posterior probabilities greater than 0.90 are indicated above branches as ML BS/PP/MP BS. The scale bar represents the expected number of changes per site. The tree is rooted with Myrmecridium schulzeri (CBS 100.54) and Myrmecridium banksiae (CPC 19852). Ex-type strains are indicated with T. The new collections are in bold with new taxa in blue. Branches with 100% ML BS, 1.0 PP, and 100% MP BS are thickened. Families are indicated as colored blocks.


Figure 2.
Distoseptispora amniculi (MFLU 21-0138, holotype). (a) Colonies on wood. (b,c) Conidiophores. (d) Conidiogenous cell. (e–j) Conidia. (k) Germinated conidium. (l,m) Colony on PDA, l from above, m from below. Scale bars: (a) 200 µm, (b,c,e–k) 30 µm, (d) 20 µm.
Figure 2.
Distoseptispora amniculi (MFLU 21-0138, holotype). (a) Colonies on wood. (b,c) Conidiophores. (d) Conidiogenous cell. (e–j) Conidia. (k) Germinated conidium. (l,m) Colony on PDA, l from above, m from below. Scale bars: (a) 200 µm, (b,c,e–k) 30 µm, (d) 20 µm.

Figure 3.
Distoseptispora atroviridis (HKAS 112616, holotype). (a,b) Colonies on woody substrates. (c–e,g) Fasciculate conidiophores with conidia. (f) Conidiophores and conidiogenous cells. (h–m) Conidia. (n) Germinated conidium. (o,p) Culture, (o) from above, (p) from below. Scale bars: (c–e) 50 μm, (f,g,n) 30 μm, (h–m) 20 μm.
Figure 3.
Distoseptispora atroviridis (HKAS 112616, holotype). (a,b) Colonies on woody substrates. (c–e,g) Fasciculate conidiophores with conidia. (f) Conidiophores and conidiogenous cells. (h–m) Conidia. (n) Germinated conidium. (o,p) Culture, (o) from above, (p) from below. Scale bars: (c–e) 50 μm, (f,g,n) 30 μm, (h–m) 20 μm.

Figure 4.
Distoseptispora effusa (GZAAS 20-0427, holotype). (a) Colonies on natural substrate. (b,c) Conidiophores. (d–i) Conidia. (j) Conidiogenous cell with a conidium. (k) Germinated conidium. (l,m) Culture, l from above, m from below. Scale bars: (b–k) 50 μm.
Figure 4.
Distoseptispora effusa (GZAAS 20-0427, holotype). (a) Colonies on natural substrate. (b,c) Conidiophores. (d–i) Conidia. (j) Conidiogenous cell with a conidium. (k) Germinated conidium. (l,m) Culture, l from above, m from below. Scale bars: (b–k) 50 μm.

Figure 5.
Distoseptispora fusiformis (HKAS 112617, holotype). (a,b) Colonies on woody substrates. (c–h) Conidiophores with conidia. (i,j) Conidiogenous cells. (k–n) Conidia. (o) Germinated conidium. (p,q) Culture, p from above, q from below. Scale bars: (c–h) 40 μm, (i,j,o) 30 μm, (k–n) 20 μm.
Figure 5.
Distoseptispora fusiformis (HKAS 112617, holotype). (a,b) Colonies on woody substrates. (c–h) Conidiophores with conidia. (i,j) Conidiogenous cells. (k–n) Conidia. (o) Germinated conidium. (p,q) Culture, p from above, q from below. Scale bars: (c–h) 40 μm, (i,j,o) 30 μm, (k–n) 20 μm.

Figure 6.
Distoseptispora hyalina (MFLU 21-0137, holotype). (a) Ascomata on woody substrate. (b) Vertical section of an ascoma. (c) Section of the bottom wall. (d) Section of the lateral wall near the beak. (e) Paraphyses. (f–i) Asci. (j–o) Ascospores, n and o using Indian ink. (p) Apical ring. (q) Germinated spore. (r,s) Culture, r from above, s from below. Scale bars: (a) 200 μm, (b) 50 μm, (f–i,q) 30 μm, (c,e,n,p) 20 μm, (j–m,o) 15 μm, (d) 10 μm.
Figure 6.
Distoseptispora hyalina (MFLU 21-0137, holotype). (a) Ascomata on woody substrate. (b) Vertical section of an ascoma. (c) Section of the bottom wall. (d) Section of the lateral wall near the beak. (e) Paraphyses. (f–i) Asci. (j–o) Ascospores, n and o using Indian ink. (p) Apical ring. (q) Germinated spore. (r,s) Culture, r from above, s from below. Scale bars: (a) 200 μm, (b) 50 μm, (f–i,q) 30 μm, (c,e,n,p) 20 μm, (j–m,o) 15 μm, (d) 10 μm.

Figure 7.
Distoseptispora lignicola (GZAAS20-0424). (a,b) Colonies on natural substrate. (c–f) Conidia. (g) Conidiophores. (h) Conidiophore and a conidium. (i) Germinated conidium. (j,k) Culture, j from above, k from below. Scale bars: (c–i) 30 μm.
Figure 7.
Distoseptispora lignicola (GZAAS20-0424). (a,b) Colonies on natural substrate. (c–f) Conidia. (g) Conidiophores. (h) Conidiophore and a conidium. (i) Germinated conidium. (j,k) Culture, j from above, k from below. Scale bars: (c–i) 30 μm.

Figure 8.
Distoseptispora verrucosa (HKAS 112652, holotype). (a) Conidiophore with a conidium. (b,c) Conidiophores. (d) Germinated conidium. (e–g) Conidia. (h,i) Culture, h from above, i from below. Scale bars: (a) 50 μm, (b–d) 30 μm, (e–g) 20 μm.
Figure 8.
Distoseptispora verrucosa (HKAS 112652, holotype). (a) Conidiophore with a conidium. (b,c) Conidiophores. (d) Germinated conidium. (e–g) Conidia. (h,i) Culture, h from above, i from below. Scale bars: (a) 50 μm, (b–d) 30 μm, (e–g) 20 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa used in the phylogenetic analyses and their GenBank accession numbers. T denotes ex-type strains. Newly generated sequences are in bold.
Table 1.
Taxa used in the phylogenetic analyses and their GenBank accession numbers. T denotes ex-type strains. Newly generated sequences are in bold.
| Taxon | Voucher/Strain Number | GenBank Accession Number | |||
|---|---|---|---|---|---|
| LSU | ITS | TEF1α | RPB2 | ||
| Acrodictys bambusicola | CGMCC 3.18641 | KX033564 | KU999973 | – | – |
| Acrodictys elaeidicola | CGMCC 3.18642 | KX033569 | KU999978 | – | – |
| Aquapteridospora aquatica | MFLUCC 17-2371T | MW287767 | MW286493 | – | – |
| Aquapteridospora fusiformis | MFLUCC 18-1606T | MK849798 | MK828652 | MN194056 | – |
| Aquapteridospora lignicola | MFLUCC 15-0377T | KU221018 | MZ868774 | MZ892980 | MZ892986 |
| Bullimyces aurisporus | AF316-1bT | JF775590 | – | – | – |
| Bullimyces communis | AF281-5 | JF775587 | – | – | – |
| Cancellidium applanatum | CBS 337.76T | MH872755 | MH860985 | – | – |
| Cancellidium cinereum | MFLUCC 18-0424T | MT370363 | MT370353 | MT370488 | MT370486 |
| “Distoseptispora adscendens” | HKUCC 10820 | DQ408561 | – | – | DQ435092 |
| Distoseptispora amniculi | MFLUCC 17-2129T | MZ868761 | MZ868770 | – | MZ892982 |
| Distoseptispora appendiculata | MFLUCC 18-0259T | MN163023 | MN163009 | MN174866 | – |
| Distoseptispora aquatica | MFLUCC 15-0374T | KU376268 | MF077552 | – | – |
| Distoseptispora aquatica | MFLUCC 18-0646 | MK849793 | MK828648 | MN194052 | – |
| Distoseptispora aquatica | S-965 | MK849792 | MK828647 | MN194051 | MN124537 |
| Distoseptispora atroviridis | GZCC 20-0511T | MZ868763 | MZ868772 | MZ892978 | MZ892984 |
| Distoseptispora atroviridis | GZCC 19-0531 | MZ227223 | MW133915 | – | – |
| Distoseptispora bambusae | MFLUCC 20-0091T | MT232718 | MT232713 | MT232880 | MT232881 |
| Distoseptispora bambusae | MFLUCC 14-0583 | MT232717 | MT232712 | – | MT232882 |
| Distoseptispora bangkokensis | MFLUCC 18-0262T | MZ518206 | MZ518205 | – | – |
| Distoseptispora cangshanensis | MFLUCC 16-0970T | MG979761 | MG979754 | MG988419 | – |
| Distoseptispora caricis | CPC 36498T | MN567632 | MN562124 | – | MN556805 |
| Distoseptispora caricis | CPC 36442T | – | MN562125 | – | MN556806 |
| Distoseptispora chinensis | GZCC 21-0665T | MZ474867 | MZ474871 | MZ501609 | – |
| Distoseptispora clematidis | MFLUCC 17-2145T | MT214617 | MT310661 | – | MT394721 |
| Distoseptispora dehongensis | KUMCC 18-0090T | MK079662 | MK085061 | MK087659 | – |
| Distoseptispora effusa | GZCC 19-0532T | MZ227224 | MW133916 | MZ206156 | – |
| Distoseptispora euseptata | MFLUCC 20-0154T | MW081544 | MW081539 | – | MW151860 |
| Distoseptispora euseptata | DLUCC S2024 | MW081545 | MW081540 | MW084994 | MW084996 |
| Distoseptispora fasciculata | KUMCC 19-0081T | MW287775 | MW286501 | MW396656 | – |
| Distoseptispora fluminicola | MFLUCC 15-0417T | KU376270 | MF077553 | – | – |
| Distoseptispora fusiformis | GZCC 20-0512T | MZ868764 | MZ868773 | MZ892979 | MZ892985 |
| Distoseptispora guizhouensis | GZCC 21-0666T | MZ474869 | MZ474868 | MZ501610 | MZ501611 |
| Distoseptispora guttulata | MFLUCC 16-0183T | MF077554 | MF077543 | MF135651 | – |
| Distoseptispora hyalina | MFLUCC 17-2128T | MZ868760 | MZ868769 | MZ892976 | MZ892981 |
| Distoseptispora hydei | MFLUCC 20-0115T | MT742830 | MT734661 | – | MT767128 |
| Distoseptispora lancangjiangensis | DLUCC 1864T | MW879522 | MW723055 | – | – |
| “Distoseptispora leonensis” | HKUCC 10822 | DQ408566 | – | – | DQ435089 |
| Distoseptispora lignicola | MFLUCC 18-0198T | MK849797 | MK828651 | – | – |
| Distoseptispora lignicola | GZCC 19-0529 | MZ227219 | MW133911 | – | – |
| Distoseptispora longispora | HFJAU 0705T | MH555357 | MH555359 | – | – |
| Distoseptispora martinii | CGMCC 3.18651 | KX033566 | KU999975 | – | – |
| Distoseptispora multiseptata | MFLUCC 15-0609T | KX710140 | KX710145 | MF135659 | – |
| Distoseptispora multiseptata | MFLUCC 16-1044 | MF077555 | MF077544 | MF135652 | MF135644 |
| Distoseptispora neorostrata | MFLUCC 18-0376T | MN163017 | MN163008 | – | – |
| Distoseptispora obclavata | MFLUCC 18-0329T | MN163010 | MN163012 | – | – |
| Distoseptispora obpyriformis | MFLUCC 17-1694T | MG979764 | – | MG988422 | MG988415 |
| Distoseptispora obpyriformis | DLUCC 0867 | MG979765 | MG979757 | MG988423 | MG988416 |
| Distoseptispora palmarum | MFLUCC 18-1446T | MK079663 | MK085062 | MK087660 | MK087670 |
| Distoseptispora phangngaensis | MFLUCC 16-0857T | MF077556 | MF077545 | MF135653 | – |
| Distoseptispora rayongensis | MFLUCC 18-0415T | MH457137 | MH457172 | MH463253 | MH463255 |
| Distoseptispora rayongensis | MFLUCC 18-0417 | MH457138 | MH457173 | MH463254 | MH463256 |
| Distoseptispora rostrata | MFLUCC 16-0969T | MG979766 | MG979758 | MG988424 | MG988417 |
| Distoseptispora rostrata | DLUCC 0885 | MG979767 | MG979759 | MG988425 | – |
| Distoseptispora saprophytica | MFLUCC 18-1238T | MW287780 | MW286506 | MW396651 | MW504069 |
| Distoseptispora songkhlaensis | MFLUCC 18-1234T | MW287755 | MW286482 | MW396642 | – |
| Distoseptispora suoluoensis | MFLUCC 17-0224T | MF077557 | MF077546 | MF135654 | – |
| Distoseptispora suoluoensis | MFLUCC 17-1305 | MF077558 | MF077547 | – | MZ945510 |
| Distoseptispora tectonae | MFLUCC 12-0291T | KX751713 | KX751711 | KX751710 | KX751708 |
| Distoseptispora tectonae | MFLUCC 15-0981 | MW287763 | MW286489 | MW396641 | – |
| Distoseptispora tectonae | MFLUCC 16-0946 | MG979768 | MG979760 | MG988426 | MG988418 |
| Distoseptispora tectonigena | MFLUCC 12-0292T | KX751714 | KX751712 | – | KX751709 |
| Distoseptispora thailandica | MFLUCC 16-0270T | MH260292 | MH275060 | MH412767 | – |
| Distoseptispora thysanolaenae | KUMCC 18-0182T | MK064091 | MK045851 | MK086031 | – |
| Distoseptispora thysanolaenae | HKAS 112710 | MW879524 | MW723057 | MW729783 | – |
| Distoseptispora verrucosa | GZCC 20-0434T | MZ868762 | MZ868771 | MZ892977 | MZ892983 |
| Distoseptispora xishuangbannaensis | KUMCC 17-0290T | MH260293 | MH275061 | MH412768 | MH412754 |
| Distoseptispora yunnanensis | MFLUCC 20-0153T | MW081546 | MW081541 | MW084995 | MW151861 |
| Fluminicola saprophytica | MFLUCC 15-0976T | MF374367 | MF374358 | MF370956 | MF370954 |
| Myrmecridium banksiae | CPC 19852T | JX069855 | JX069871 | – | – |
| Myrmecridium schulzeri | CBS 100.54 | EU041826 | EU041769 | – | – |
| Papulosa amerospora | AFTOL-ID 748 | DQ470950 | – | DQ471069 | DQ470901 |
| Pleurophragmium bambusinum | MFLUCC 12-0850 | KU863149 | KU940161 | KU940213 | – |
| Pseudostanjehughesia aquitropica | MFLUCC 16-0569T | MF077559 | MF077548 | MF135655 | – |
| Pseudostanjehughesia lignicola | MFLUCC 15-0352T | MK849787 | MK828643 | MN194047 | MN124534 |
| Wongia griffinii | DAR 80512T | KU850471 | KU850473 | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, J.; Liu, L.-L.; Jones, E.B.G.; Li, W.-L.; Hyde, K.D.; Liu, Z.-Y. Morphological Variety in Distoseptispora and Introduction of Six Novel Species. J. Fungi 2021, 7, 945. https://doi.org/10.3390/jof7110945
AMA Style
Yang J, Liu L-L, Jones EBG, Li W-L, Hyde KD, Liu Z-Y. Morphological Variety in Distoseptispora and Introduction of Six Novel Species. Journal of Fungi. 2021; 7(11):945. https://doi.org/10.3390/jof7110945
Chicago/Turabian StyleYang, Jing, Ling-Ling Liu, E. B. Gareth Jones, Wen-Li Li, Kevin D. Hyde, and Zuo-Yi Liu. 2021. "Morphological Variety in Distoseptispora and Introduction of Six Novel Species" Journal of Fungi 7, no. 11: 945. https://doi.org/10.3390/jof7110945
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.