
Taxonomy and Phylogeny of the Wood-Inhabiting Fungal Genus Hyphoderma with Descriptions of Three New Species from East Asia


1
Key Laboratory for Forest Resources Conservation and Utilization in the Southwest Mountains of China, Ministry of Education, Southwest Forestry University, Kunming 650224, China
2
College of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(4), 308; https://doi.org/10.3390/jof7040308
Submission received: 28 March 2021
/
Revised: 14 April 2021
/
Accepted: 15 April 2021
/
Published: 16 April 2021
(This article belongs to the Special Issue Diversity and Classification of Environmental Fungi)
Abstract
:Three new wood-inhabiting fungi, Hyphoderma crystallinum, H. membranaceum, and H. microporoides spp. nov., are proposed based on a combination of morphological features and molecular evidence. Hyphoderma crystallinum is characterized by the resupinate basidiomata with smooth hymenial surface scattering scattered nubby crystals, a monomitic hyphal system with clamped generative hyphae, and numerous encrusted cystidia present. Hyphoderma membranaceum is characterized by the resupinate basidiomata with tuberculate hymenial surface, presence of the moniliform cystidia, and ellipsoid to cylindrical basidiospores. Hyphoderma microporoides is characterized by the resupinate, cottony basidiomata distributing the scattered pinholes visible using hand lens on the hymenial surface, presence of halocystidia, and cylindrical to allantoid basidiospores. Sequences of ITS+nLSU rRNA gene regions of the studied samples were generated, and phylogenetic analyses were performed with maximum likelihood, maximum parsimony, and Bayesian inference methods. These phylogenetic analyses showed that three new species clustered into Hyphoderma, in which H. crystallinum was sister to H. variolosum, H. membranaceum was retrieved as a sister species of H. sinense, and H. microporoides was closely grouped with H. nemorale. In addition to new species, map to show global distribution of Hyphoderma species treated in the phylogenetic tree and an identification key to Chinese Hyphoderma are provided.
1. Introduction
Fungi are an ecologically important branch of the tree of life based on its distinct and diverse characteristics, in which these organisms play a vital role in ecosystems as diverse as soil, forest, rocks, and ocean, but their roles are primarily enacted behind the scenes, literally as hidden layers within their substrate [1]. On the basis of the nature of their intertwined partners in numerous symbiotic interactions, they have mostly marched via stepwise codiversification with the plants [2]. Fungi have evolved numerous strategies to degrade hard-to-digest substrates for outcompeting with other microbes, while combating competitors using an arsenal of bioactive metabolites, such as the familiar antibiotics, ethanol, and organic acids [3]. Taxonomy plays a central role in understanding the diversity of life, discovering into systems of names that capture the relationships between species, and translating the products of biological exploration [4]. Despite the early embrace of the molecular systematics by mycologists, both the discovery and classification of fungi are still in great flux, particularly among the more basal branches of the tree, in which the true diversity is only now coming to light from genomic analyses and environmental DNA surveys [1]. The researches revealed that perhaps less than 5% of the estimated two to four million species have been formally described, therefore, the hidden and microscopic nature of many fungi also means that their diversity is undersampled [5,6].
The genus Hyphoderma Wallr. is one of the most important fungal groups because of its key role in the carbon cycle and being the most efficient wood decomposers in the forest ecosystem [7]. This genus is typified by H. setigerum (Fr.) Donk [8]. Hyphoderma is characterized by the resupinate to effuse-reflexed basidiomata with ceraceous consistency, and smooth to tuberculate or hydnoid hymenophore and a monomitic hyphal structure (rarely dimitic) with clamp connections on generative hyphae, presence of cystidia or not, basidia suburniform to subcylindrical and cylindrical, ellipsoid to subglobose, smooth, thin-walled basidiospores [9,10]. Currently, about 100 species have been accepted in Hyphoderma worldwide [8,11,12,13,14,15]. Index Fungorum (http://www.indexfungorum.org; accessed on 16 April 2021) and MycoBank (https://www.mycobank.org; accessed on 16 April 2021) register 192 specific and infraspecific names in Hyphoderma.
Molecular systematics covering Hyphoderma revealed the classification of corticioid fungi and showed that H. obtusum J. Erikss. and H. setigerum clustered into Meruliaceae Rea and then grouped with Hypochnicium polonense (Bres.) Å. Strid, based on the internal transcribed spacer (ITS) regions and the large subunit nuclear ribosomal RNA gene (nLSU) sequences [16]. Telleria et al. [17] discussed the relationships between Hyphoderma and Peniophorella P. Karst., in which some species from Hyphoderma and Peniophorella are grouped and they proposed a new species, H. macaronesicum Tellería et al. The research on studying Hyphoderma setigerum complex showed that H. pinicola Yurch. and Sheng H. Wu represented a fifth species in this complex, which revealed that this complex was a white-rot wood-decaying corticoid fungal species and occurred worldwide from tropical to temperate regions [18]. A revised family-level classification of the Polyporales revealed that four Hyphoderma species nested into the residual polyporoid clade belonging to Hyphodermataceae, and then, they were grouped with three related genera Meripilus P. Karst., Physisporinus P. Karst., and Rigidoporus Murrill [19]. Chinese Hyphoderma species were compared with closely related taxa, and two new species were proposed, H. fissuratum C.L. Zhao and X. Ma and H. mopanshanense C.L. Zhao [15].
In this study, three undescribed species of corticioid fungi were collected from Yunnan Province, China. Morphological characteristics and molecular phylogenetic analyses of combined ITS+nLSU rRNA sequences supported the recognition of three new species within Hyphoderma.
2. Materials and Methods
2.1. Morphology
The studied specimens are deposited at the herbarium of Southwest Forestry University (SWFC), Kunming, Yunnan Province, China. Macromorphological descriptions are based on field notes and photos captured in the field and laboratory. Color terminology follows Petersen [20]. Micromorphological data were obtained from the dried specimens, which were observed under a light microscope following Dai [21]. The following abbreviations are used: KOH = 5% potassium hydroxide water solution, CB = Cotton Blue, CB– = acyanophilous, IKI = Melzer’s reagent, IKI– = both inamyloid and indextrinoid, L = mean spore length (arithmetic average for all spores), W = mean spore width (arithmetic average for all spores), Q = variation in the L/W ratios between the specimens studied, n = a/b (number of spores (a) measured from given number (b) of specimens).
2.2. Molecular Phylogeny
CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to obtain genomic DNA from dried specimens, according to the manufacturer’s instructions [22]. ITS region was amplified with primer pair ITS5 and ITS4 [23]. Nuclear nLSU region was amplified with primer pair LR0R and LR7 (http://lutzonilab.org/nuclear-ribosomal-dna/; accessed on 16 April 2021). The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min; followed by 35 cycles at 94 °C for 40 s, 58 °C for 45 s, and 72 °C for 1 min; and a final extension of 72 °C for 10 min. The PCR procedure for nLSU was as follows: initial denaturation at 94 °C for 1 min; followed by 35 cycles at 94 °C for 30 s, 48 °C for 1 min, and 72 °C for 1.5 min; and a final extension of 72 °C for 10 min. The PCR products were purified and directly sequenced at Kunming Tsingke Biological Technology Limited Company, Kunming, Yunnan Province, China. All newly generated sequences were deposited in NCBI GenBank (Table 1).
Sequences were aligned in MAFFT 7 (https://mafft.cbrc.jp/alignment/server/; accessed on 10 April 2021) using the “G-INS-i” strategy for ITS+nLSU and manually adjusted in BioEdit [32]. The dataset was aligned first and then, ITS and nLSU sequences were combined with Mesquite. Alignment datasets were deposited in TreeBASE (submission ID 27983). Climacocystis borealis (Fr.) Kotl. and Pouzar and Diplomitoporus crustulinus (Bres.) Domański were selected as an outgroup for phylogenetic analysis of ITS+nLSU phylogenetic tree (Figure 1) following a previous study [19].
Maximum parsimony analysis was applied to the combined (ITS+nLSU) dataset. Its approaches followed previous study [22], and the tree construction procedure was performed in PAUP* version 4.0b10 [33]. All characters were equally weighted and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1000 random sequence additions. Max-trees were set to 5000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using bootstrap (BT) analysis with 1000 replicates [34]. Descriptive tree statistics: tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI) were calculated for each maximum parsimonious tree generated. Datamatrix was also analyzed using maximum likelihood (ML) approach with RAxML-HPC2 through the CIPRES Science Gateway (www.phylo.org; accessed on 8 April 2021) [35]. Branch support (BS) for ML analysis was determined by 1000 bootstrap replicates.
MrModeltest 2.3 [36] was used to determine the best-fit evolution model for each data set for Bayesian inference (BI). BI was calculated with MrBayes 3.1.2 [37]. Four Markov chains were run for 2 runs from random starting trees for 6 million generations for ITS+nLSU (Figure 1). The first one-fourth of all generations was discarded as burn-in. The majority rule consensus tree of all remaining trees was calculated. Branches were considered as significantly supported if they received maximum likelihood bootstrap value (BS) > 70%, maximum parsimony bootstrap value (BT) > 70%, or Bayesian posterior probabilities (BPP) > 0.95.
3. Results
3.1. Molecular Phylogeny
The ITS+nLSU dataset (Figure 1) included sequences from 78 fungal specimens representing 44 taxa. The dataset had an aligned length of 2086 characters, of which 1245 characters are constant, 127 are variable and parsimony-uninformative, and 714 are parsimony-informative. Maximum parsimony analysis yielded 5000 equally parsimonious trees (TL = 3441, CI = 0.3787, HI = 0.6213, RI = 0.7178, RC = 0.2718). Best model for the ITS+nLSU dataset estimated and applied in the Bayesian analysis was GTR+I+G (lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1)). Bayesian analysis and ML analysis resulted in a similar topology to MP analysis with an average standard deviation of split frequencies = 0.007698 (BI).
The phylogram inferred from ITS+nLSU sequences (Figure 1) demonstrated that three new species are clustered into genus Hyphoderma, in which H. crystallinum was sister to H. variolosum Boidin, Lanq. and Gilles, H. membranaceum was retrieved as a sister species of H. sinense C.L. Zhao and Q.X. Guan, and H. microporoides was closely grouped with H. nemorale K.H. Larss. (100% BS, 100% BP, and 1.00 BPP).
3.2. Taxonomy
MycoBank no.: MB 839276.
Holotype—China, Yunnan Province, Puer, Jingdong County, the Forest of Pineapple, E 100°48′, N 24°21′, 2113 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 4 January 2019, C.L. Zhao 9338 (SWFC).
Etymology—crystallinum (Lat.): referring to the numerous and larger crystals on the hymenial surface.
Fruiting body—Basidiomata annual, resupinate, adnate, without odor and taste when fresh, membranaceous when fresh, becoming hard membranaceous upon drying, and up to 15 cm long, 3 cm wide, and 30–100 µm thick. Hymenial surface white to pale gray when fresh, pale gray to slightly cream upon drying, with scattered nubby crystals. Margin sterile indistinct and white.
Hyphal system—Monomitic, generative hyphae with clamp connections, colorless, thin-walled, frequently branched, interwoven, 2–3.5 µm in diameter, IKI-, CB-; tissues unchanged in KOH.
Hymenium—Cystidia of two types: (1) tubular cystidia, colorless, thin-walled, 32–51 µm × 5–10 µm and (2) encrusted cystidia, numerous, colorless, encrusted by crystals, 14–46 µm × 4–11 µm. Basidia clavate to subcylindrical, slightly constricted in the middle to somewhat sinuous, with 4 sterigmata and a basal clamp connection, 21.5–31 µm × 6–8.5 µm.
Spores—Basidiospores allantoid, colorless, thin-walled, smooth, with oil drops inside, IKI-, CB-, (10.5–)11–14.5(–15) µm × 4–5.5(–6) µm, L = 12.99 µm, W = 4.81 µm, Q = 2.47–2.98 (n = 90/3).
Additional specimens examined—China, Yunnan Province, Puer, Jingdong County, the Forest of Pineapple, E 100°48′, N 24°21′, 2113 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 4 January 2019, C.L. Zhao 9374 (SWFC); Dali, Nanjian County, Lingbaoshan National Forestry Park, E 100°30′, N 24°46′, 1963 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 9 January 2019, C.L. Zhao 10224 (SWFC); Wenshan, Funing County, Guying village, E 105°35′, N 23°36′, 976 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 20 January 2019, C.L. Zhao 11723 (SWFC); Wenshan, Xichou County, Jiguanshan Forestry Park, E 103°46′, N 23°33′, 1670 m asl., on fallen angiosperm trunk, leg. C.L. Zhao, 22 July 2019, C.L. Zhao 15841 (SWFC); Honghe, Pingbian County, Daweishan National Nature Reserve, E 103°35′, N 22°53′, 1990 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 3 August 2019, C.L. Zhao 18459 (SWFC).
MycoBank no.: MB 839278.
Holotype—China, Yunnan Province, Chuxiong, Zixishan Forestry Park, E 101°24′, N 25°01′, 2356 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 1 July 2018, C.L. Zhao 6971 (SWFC).
Etymology—membranaceum (Lat.): referring to the membranous hymenophore.
Fruiting body—Basidiomata annual, resupinate, adnate, membranous, without odor and taste when fresh, and up to 15 cm long, 2 cm wide, and 30–100 µm thick. Hymenial surface tuberculate, white to pale gray when fresh, pale gray to cream on drying, with cracking. Margin sterile, narrow, and gray.
Hyphal system—Monomitic, generative hyphae with clamp connections, colorless, thin-walled, frequently branched, interwoven, 2.5–4.5 µm in diameter; IKI-, CB-; tissues unchanged in KOH.
Hymenium—Cystidia moniliform, thin-walled, 28–60 µm × 6.5–10.5 µm; basidia clavate to subcylindrical, slightly constricted in the middle to somewhat sinuous, with 4 sterigmata and a basal clamp connection, 21.5–31 µm × 5–7.5 µm.
Spores—Basidiospores ellipsoid to cylindrical, colorless, thin-walled, smooth, with irregular vacuole inside, IKI-, CB-, (10.5–)11–13.5(–14) µm × 4.5–5.5(–6) µm, L = 12.52 µm, W = 5.18 µm, Q = 2.42 (n = 60/2).
Additional specimens examined—China, Yunnan Province, Puer, Zhenyuan County, Heping Town, Liangzizhai, E 101°25′, N 23°56′, 2246 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 15 Jan 2018, C.L. Zhao 5844 (SWFC).
MycoBank no.: MB 839277.
Holotype—China, Yunnan Province, Chuxiong, Zixishan Forestry Park, E 101°24′, N 25°01′, 2313 m asl., on fallen angiosperm trunk, leg. C.L. Zhao, 30 June 2018, C.L. Zhao 6857 (SWFC).
Etymology—microporoides (Lat.): referring to the scattered pinholes on the hymenophore that are visible under hand lens.
Fruiting body—Basidiomata annual, resupinate, adnate, without odor and taste when fresh, cottony when fresh, fragile upon drying, and up to 22 cm long, 2.5 cm wide, and 50–100 µm thick. Hymenial surface smooth with scattered pinholes visible under hand lens, cream to pale buff when fresh, and slightly buff upon drying. Margin sterile, indistinct, and white to cream.
Hyphal system—Monomitic, generative hyphae with clamp connections, colorless, thin-walled, frequently branched, interwoven, 3–5 µm in diameter, IKI-, CB-; tissues unchanged in KOH.
Hymenium—Halocystidia capitate, thin-walled, smooth, 18–51 µm × 4.5–7 µm; basidia clavate, constricted, somewhat sinuous, with 4 sterigmata and a basal clamp connection, 18.5–29.5 µm × 5–7 µm.
Spores—Basidiospores cylindrical to allantoid, colorless, thin-walled, smooth, with oil drops inside, IKI-, CB-, 8.5–10(–10.5) µm × 2.5–3.5(–4) µm, L = 9.29 µm, W = 3.24 µm, Q = 2.87 (n = 30/1).
Additional specimens examined—China, Yunnan Province, Puer, Jingdong County, Taizhong Town, Ailaoshan Ecological Station, E 100°56′, N 24°29′, 1938 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 25 August 2018, C.L. Zhao 8695 (SWFC).
4. Discussion
In the present study, three new species, Hyphoderma crystallinum, H. membranaceum, and H. microporoides are described based on phylogenetic analyses and morphological characteristics.
Phylogenetically, the family-level classification of the Polyporales (Basidiomycota) amplified nLSU, nITS, and rpb1 genes across the Polyporales, was employed, in which four species Hyphoderma macaronesicum, H. medioburiense (Burt) Donk, H. mutatum (Peck) Donk, and H. setigerum, nested into family Hyphodermataceae within the residual polyporoid clade [19]. In the present study, three new taxa clustered into Hyphoderma, in which Hyphoderma crystallinum was sister to H. variolosum, H. microporoides grouped closely with H. nemorale, and H. membranaceum grouped with H. sinense and H. transiens (Bres.) Parmasto (Figure 1). However, morphologically, H. variolosum differs from H. crystallinum by its narrower tubular cystidia (40–50 µm × 4–6 µm) [38]; H. nemorale is separated from H. microporoides by having the colliculose hymenial surface, wider moniliform cystidia (35–70 µm × 7–8 µm) and basidiospores (9.5–14 µm × 4–5 µm) [27]; H. sinense differs from H. membranaceum by having the encrusted cystidia and smaller basidiospores (8–11.5 µm × 3–5 µm) [25], and another species H. transiens differs in its odontioid hymenial surface and narrower basidiospores (9–13 µm × 3–4.5 µm) [39].
Morphologically, Hyphoderma ayresii (Berk. ex Cooke) Boidin and Gilles, H. cremeum Sheng H. Wu and H. rimulosum Sheng H. Wu are similar to H. crystallinum by having encrusted cystidia. However, H. ayresii differs in its larger encrusted cystidia (70–130 µm × 13–20 µm) and wider basidiospores (9.5–12.5 µm × 6–8 µm) [38]; H. cremeum differs from H. crystallinum by having both larger encrusted cystidia (40–90 µm × 10–15 µm) and basidia (35–50 µm × 6.5–8 µm) [40]; H. rimulosum is separated from H. crystallinum by smaller basidiospores (6–7 µm × 3.9–4.1 µm) [41]. Hyphoderma incrustatum K.H. Larss., H. medioburiense, H. multicystidium (Hjortstam and Ryvarden) Hjortstam and Tellería and H. roseocremeum (Bres.) Donk are similar to H. crystallinum by having tubular cystidia. However, H. incrustatum differs from H. crystallinum by the porulose hymenial surface and the larger tubular cystidia (50–80 µm × 6–10 µm) [42]; H. medioburiense is separated from H. crystallinum by the porulose hymenial surface and the larger tubular cystidia (60–100 µm × 7–10 µm) [8]; H. multicystidium differs in its larger tubular cystidia (60–80 µm × 5–7 µm), larger basidia (35–50 µm × 5–7 µm) and smaller basidiospores (8–10 µm × 4.5–5 µm) [43]; H. roseocremeum differs from H. crystallinum by having larger tubular cystidia (80–100 × 6–9 µm) and smaller basidiospores (8–12 µm × 3–4 µm) [8].
Hyphoderma litschaueri, H. moniliforme (P.H.B. Talbot) Manjón, G. Moreno and Hjortstam, H. paramacaronesicum Tellería et al., H. prosopidis (Burds.) Tellería et al. and H. sinense are similar to H. membranaceum by having moniliform or apically moniliform cystidia. However, H. litschaueri differs from H. membranaceum by having larger moniliform cystidia (60–100 µm × 6–8 µm) and narrower basidiospores (9–12 µm × 3–4 µm) [44]; H. moniliforme differs from H. membranaceum by having smaller basidiospores (8–9 µm × 3.5–4 µm) [27]; H. paramacaronesicum differs in its having both larger moniliform cystidia (70–124 µm × 8–13 µm) and basidia (40−48 µm × 6−9 µm), and wider basidiospores (12–15 µm × 5.5–7 µm) [14]; H. prosopidis differs from H. membranaceum by the arachnoid to farinaceous hymenial surface and larger basidia (40−45 µm × 8−11 µm) [17]; and H. sinense differs in its having encrusted cystidia (18.5–38 µm × 6–11 µm) and smaller basidiospores (8–11.5 µm × 3–5 µm) [25].
Hyphoderma clavatum Sheng H. Wu, H. etruriae Bernicchia, H. incrustatum, H. orphanellum (Bourdot & Galzin) Donk, and H. subclavatum Sheng H. Wu are similar to H. microporoides by having capitate cystidia. However, H. clavatum differs from H. microporoides by the tuberculate hymenial surface and larger basidiospores (10–13 µm × 4.2–5.2 µm) [41]; H. etruriae differs from H. microporoides by the grandinioid hymenial surface and wider basidiospores (9–11 µm × 5.5–6.5 µm) [45]; H. incrustatum differs in having larger basidiospores (11–14 µm × 4–5 µm) [42]; H. orphanellum differs from H. microporoides by having larger capitate cystidia (50–80 µm × 8–10 µm) and wider basidiospores (8–10 µm × 5–6 µm) [8]; H. subclavatum is separated from H. microporoides by having both larger basidia (40–55 µm × 7–8 µm) and basidiospores (10–12 µm × 4.2–5.3 µm) [41].
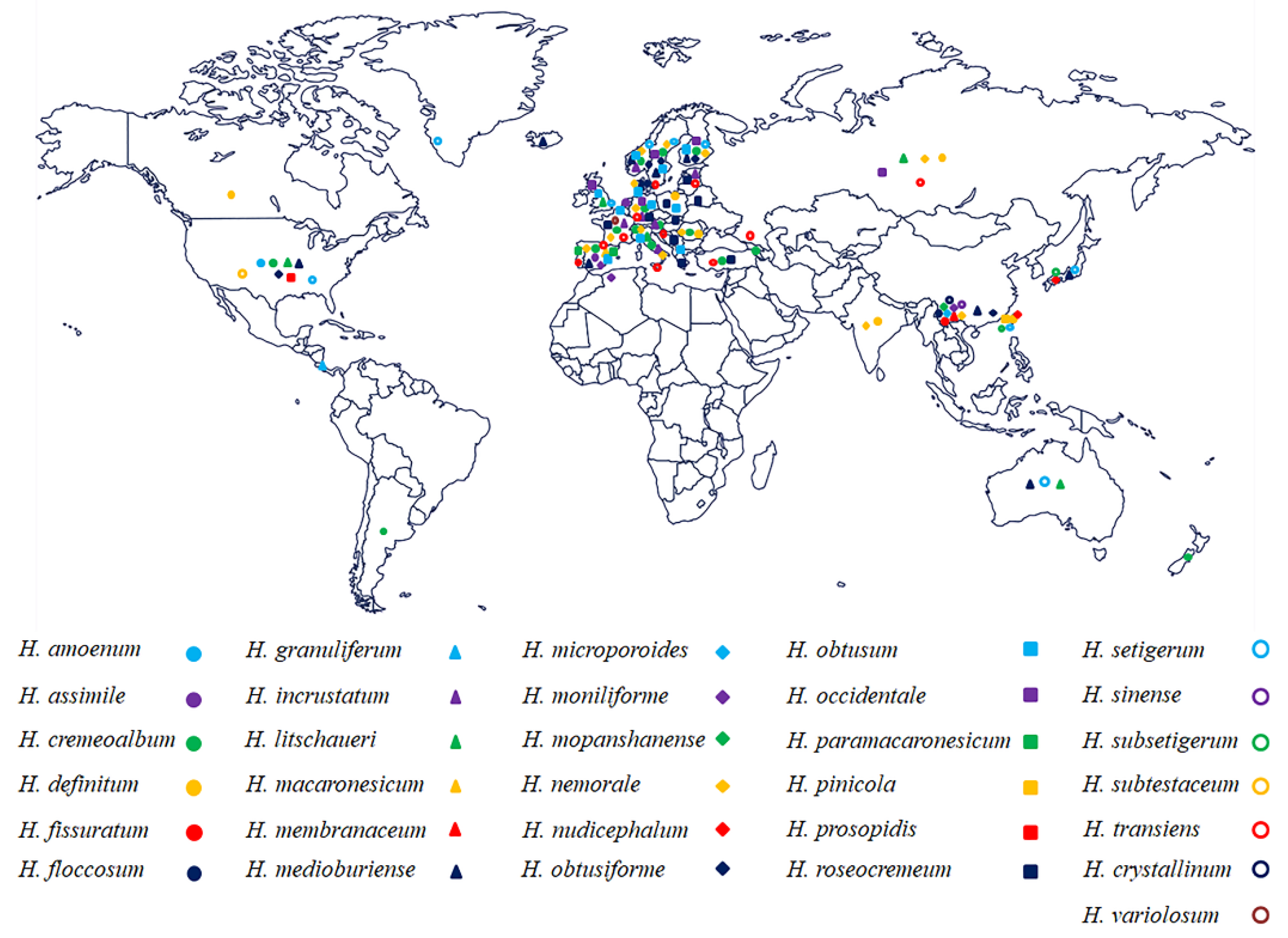
Hyphoderma species are an extensively studied group [10,46], mainly distributed in Europe (e.g., Austria, Russia, France, Germany, Poland, UK, The Netherlands, Portugal, Sweden, Italy, Denmark, Norway, Finland, Spain) (Figure 8) and mainly found on hardwood, although a few species grow on coniferous wood. Many species of Hyphoderma were found in Europe, but most of them have not been reported in northern China (Figure 8), in which we presumed that Hyphoderma are undersampled by mycologists. Several studies on new wood-decaying fungi of Hyphoderma from China have been reported [15,40,41,46], in which 26 Hyphoderma species were reported, H. acystidiatum Sheng H. Wu, H. clavatum, H. cremeoalbum (Höhn. and Litsch.) Jülich, H. cremeum, H. definitum (H.S. Jacks.) Donk, H. densum Sheng H. Wu, H. fissuratum, H. floccosum C.L. Zhao and Q.X. Guan, H. litschaueri, H. crystallinum, H. medioburiense, H. microcystidium Sheng H. Wu, H. microporoides, H. moniliforme, H. mopanshanense, H. nemorale, H. obtusiforme J. Erikss. and Å. Strid, H. pinicola, H. rimulosum, H. setigerum, H. sibiricum (Parmasto) J. Erikss. and Å. Strid, H. sinense, H. subclavatum, H. subsetigerum Sheng H. Wu, H. transiens, and H. membranaceum [8,18,25,27,29,40,41,46]. Further studies should focus on the relationships between the host and Hyphoderma species, as well as trying to better understand the evolutionary directions between plant and Hyphoderma species. The researches on the phylogeny of Hyphoderma, as well as many fungal studies on the molecular systematics [47,48,49], will be useful to push the further research on fundamental research and applied research of fungi. More species of Hyphoderma should be found in subtropical and tropical Asia as it was shown that wood-inhabiting fungi are rich in tropical China [50,51].
Key to 26 accepted species of Hyphoderma in China
- Cystidia absent 2
- Cystidia present 5
- Hymenial surface grandinioid H. acystidiatum
- Hymenial surface smooth 3
- Basidiospores > 10.5 µm in length H. densum
- Basidiospores < 10.5 µm in length 4
- Hymenophore cracked; basidiospores > 8.5 µm in length H. fissuratum
- Hymenophore uncracked; basidiospores < 8.5 µm in length H. sibiricum
- Hymenophore smooth 6
- Hymenophore tuberculate, porulose, grandinioid, or odontoid 14
- Two types of cystidia present 7
- One type of cystidia present 8
- Moniliform cystidia absent H. microcystidium
- Moniliform cystidia present H. sinense
- Hymenophore uncracked 9
- Hymenophore cracked 10
- Basidiospores > 11 µm in length H. definitum
- Basidiospores < 11 µm in length H. microporoides
- Cystidia moniliform 11
- Cystidia cylindrical 12
- Basidiospores > 9 µm in length H. litschaueri
- Basidiospores < 9 µm in length H. moniliforme
- Basidiospores ellipsoid < 10 μm in length H. rimulosum
- Basidiospores cylindrical > 10 μm in length 13
- Basidiospores > 12 µm in length H. cremeum
- Basidiospores < 12 µm in length H. subclavatum
- Hymenophore odontoid or grandinioid 15
- Hymenophore tuberculate, porulose 16
- Hymenophore odontoid, basidiospores > 9 µm in length H. transiens
- Hymenophore grandinioid, basidiospores < 9 µm in length H. subsetigerum
- Cystidia of two types 17
- Cystidia of one type 19
- Septate cystidia absent H. crystallinum
- Septate cystidia present 18
- Basidia 2-sterigmata, basidiospores > 13 µm in length H. pinicola
- Basidia 4-sterigmata, basidiospores < 13 µm in length H. floccosum
- Septate cystidia present 20
- Septate cystidia absent 21
- Hymenophore porulose to pilose, basidia < 5 µm in width H. mopanshanense
- Hymenophore tuberculate, basidia > 5 µm in width H. setigerum
- Hymenophore porulose H. obtusiforme
- Hymenophore tuberculate, colliculose 22
- Cystidia > 30 µm in length 23
- Cystidia < 30 µm in length H. cremeoalbum
- Basidia > 30 µm in length 24
- Basidia < 30 µm in length 25
- Hymenophore cracking, cystidia < 10 µm in width H. medioburiense
- Hymenophore not cracking, cystidia > 10 µm in width H. clavatum
- Hymenophore colliculose H. nemorale
- Hymenophore tuberculate H. membranaceum
Author Contributions
Conceptualization, C.-L.Z.; methodology, C.-L.Z. and Q.-X.G.; software, C.-L.Z. and Q.-X.G.; validation, C.-L.Z. and Q.-X.G.; formal analysis, C.-L.Z. and Q.-X.G.; investigation, C.-L.Z. and Q.-X.G.; resources, C.-L.Z.; writing—original draft preparation, C.-L.Z. and Q.-X.G.; writing—review and editing, C.-L.Z. and Q.-X.G.; visualization, C.-L.Z. and Q.-X.G.; supervision, C.-L.Z.; project administration, C.-L.Z.; funding acquisition, C.-L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Yunnan Fundamental Research Project, grant number 202001AS070043.
Institutional Review Board Statement
Not applicable for studies involving humans or animals.
Informed Consent Statement
Not applicable for studies involving humans.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: [https://www.ncbi.nlm.nih.gov/; https://www.mycobank.org/page/Simple%20names%20search; http://purl.org/phylo/treebase, submission ID 27983; accessed on 16 April 2021].
Conflicts of Interest
The authors declare no conflict of interest.
References
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 2020, 74, 1–23. [Google Scholar] [CrossRef]
- Lutzoni, F.; Nowak, M.D.; Alfaro, M.E.; Reeb, V.; Miadlikowska, J.; Krug, M.; Arnold, A.E.; Lewis, L.A.; Swofford, D.L.; Hibbett, D.; et al. Contemporaneous radiations of fungi and plants linked to symbiosis. Nat. Commun. 2018, 9, 5451. [Google Scholar] [CrossRef]
- Rokas, A.; Wisecaver, J.H.; Lind, A.L. The birth, evolution and death of metabolic gene clusters in fungi. Nat. Rev. Microbiol. 2018, 16, 731–744. [Google Scholar] [CrossRef]
- Hibbett, D.S. The invisible dimension of fungal diversity. Science 2016, 351, 1150–1151. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, M. The Fungi: 1, 2, 3... 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martínez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donk, M.A. Notes on resupinate Hymenomycetes IV. Fungus 1957, 27, 1–29. [Google Scholar]
- Wallroth, C.F.W. Flora Cryptogamica Germaniae; Nabu Press: Nürnberg, Germany, 1833; Volume 2, pp. 1–923. [Google Scholar]
- Bernicchia, A.; Gorjón, S.P. Fungi Europaei 12: Corticiaceae s.l.; Edizioni Candusso: Alassio, Italy, 2010. [Google Scholar]
- Nakasone, K.K. Type studies of corticioid Hymenomycetes described by Bresadola. Cryptogam. Mycol. 2008, 29, 231–257. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.H.; Nilsson, H.R.; Chen, C.T.; Yu, S.Y.; Hallenberg, N. The white-rotting genus Phanerochaete is polyphyletic and distributed throughout the phlebioid clade of the Polyporales (Basidiomycota). Fungal Divers. 2010, 42, 107–118. [Google Scholar] [CrossRef]
- Baltazar, J.M.; Silveira, R.M.B.; Rajchenberg, M. Type studies of J. Rick’s corticioid Homobasidiomycetes (Agaricomycetes, Basidiomycota) housed in the Herbarium Anchieta (PACA). Phytotaxa 2016, 255, 101–132. [Google Scholar] [CrossRef]
- Martín, M.P.; Zhang, L.F.; Fernández-López, J.; Dueñas, M.; Rodríguez-Armas, J.L.; Beltrán-Tejera, E.; Telleria, M.T. Hyphoderma paramacaronesicum sp. nov. (Meruliaceae, Polyporales, Basidiomycota), a cryptic lineage to H. macaronesicum. Fungal Syst. Evol. 2018, 2, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Huang, R.X.; Zhang, Y.; Zhao, C.L. Hyphoderma fissuratum and H. mopanshanense spp. nov. (Polyporales) from southern China. Mycoscience 2021, 62, 36–41. [Google Scholar] [CrossRef]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef] [PubMed]
- Telleria, M.T.; Dueñas, M.; Beltrán-Tejera, E.; Rodríguez-Armas, J.L.; Martín, M.P. A new species of Hyphoderma (Meruliaceae, Polyporales) and its discrimination from closely related taxa. Mycologia 2012, 104, 1121–1132. [Google Scholar] [CrossRef]
- Yurchenko, E.; Wu, S.H. Hyphoderma formosanum sp. nov. (Meruliaceae, Basidiomycota) from Taiwan. Sydowia 2014, 66, 19–23. [Google Scholar]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Linder, D.; Nakasone, K.; Niemelä, T.; Larsson, K.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996; pp. 1–6. [Google Scholar]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom Fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.X.; Zhao, C.L. Two new corticioid species: Hyphoderma sinense and H. floccosum spp. nov. (Hyphodermataceae, Polyporales) from southern China. Mycosystema 2021, in press. [Google Scholar] [CrossRef]
- Floudas, D.; Hibbett, D.S. Revisiting the taxonomy of Phanerochaete (Polyporales, Basidiomycota) using a four gene dataset and extensive ITS sampling. Fungal Biol. 2015, 119, 679–719. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, E.; Wu, S.H. Hyphoderma moniliforme and H. nemorale (Basidiomycota) newly recorded from China. Mycosphere 2015, 6, 113–121. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Hallenberg, N.; Nordén, B.; Maekawa, N.; Wu, S.H. Phylogeography of Hyphoderma setigerum (Basidiomycota) in the Northern Hemisphere. Mycol. Res. 2003, 107, 645–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurchenko, E.; Wu, S.H. Hyphoderma pinicola sp. nov. of H. setigerum complex (Basidiomycota) from Yunnan, China. Bot. Stud. 2014, 55, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Telleria, M.T.; Duenas, M.; Melo, I.; Hallenberg, N.; Martin, M.P. A re-evaluation of hypochnicium (polyporales) based on morphological and molecular characters. Mycologia 2010, 102, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chen, J.J.; Ji, X.H.; Vlasák, J.; Dai, Y.C. Phylogeny and diversity of the morphologically similar polypore genera Rigidoporus, Physisporinus, Oxyporus and Leucophellinus. Mycologia 2017, 109, 749–765. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. Available online: http://www.phylo.org/sub_sections/portal (accessed on 9 September 2020).
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boidin, J.; Gilles, G. Basidiomycètes Aphyllophorales de l’Île de La Réunion. XVI. Les genres Hyphoderma, Hyphodermopsis, Chrysoderma nov. gen. et Crustoderma. Cryptogam. Mycol. 1991, 12, 97–132. [Google Scholar]
- Parmasto, E. Conspectus Systematis Corticiacearum; Institutum Zoologicum et Botanicum Academiae Scientiarum RPSS Estonicae: Tartu, Estonia, 1968. [Google Scholar]
- Wu, S.H. New species of Hyphoderma from Taiwan. Mycologia 1997, 89, 132–140. [Google Scholar] [CrossRef]
- Wu, S.H. New species and new records of Hyphoderma (Basidiomycotina) from Taiwan. Bot. Bull. Acad. Sin. 1997, 38, 63–72. [Google Scholar]
- Larsson, K.H. Two new species in Hyphoderma. Nord. J. Bot. 1998, 18, 121–127. [Google Scholar] [CrossRef]
- Tellería, M.T. Annotated list of the Corticiaceae, sensu lato (Aphyllophorales, Basidiomycotina), for Peninsular Spain and Balearic Islands. Bibl. Mycol. 1990, 135, 1–152. [Google Scholar]
- Eriksson, J.; Ryvarden, L. The Corticiaceae of North Europe; Fungiflora: Oslo, Norway, 1975; Volume 3, pp. 288–546. [Google Scholar]
- Bernicchia, A. Hyphoderma etruriae sp. nov. (Corticiaceae, Basidiomycetes) from the natural reserve of Burano, Italy. Mycotaxon 1993, 46, 37–40. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Integrating different lines of evidence to establish a novel Ascomycete genus and family (anastomitrabeculia, anastomitrabeculiaceae) in Pleosporales. J. Fungi 2021, 7, 94. [Google Scholar] [CrossRef]
- Lambert, C.; Pourmoghaddam, M.J.; Cedeo-Sanchez, M.; Surup, F.; Stadler, M. Resolution of the hypoxylon fuscum complex (Hypoxylaceae, Xylariales) and discovery and biological characterization of two of its prominent secondary metabolites. J. Fungi 2021, 7, 131. [Google Scholar] [CrossRef]
- Zhou, M.; Dai, Y.C.; Vlasák, J.; Yuan, Y. Molecular phylogeny and global diversity of the genus Haploporus (Polyporales, Basidiomycota). J. Fungi 2021, 7, 96. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.K.; Li, H.J.; Ji, X.; Zhou, J.L.; Song, J.; Si, J.; Yang, Z.L.; Dai, Y.C. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers 2019, 97, 137–392. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, H.S.; Zhou, L.W.; Yuan, Y.; Cui, B.K.; Dai, Y.C. Polypore diversity in South China. Mycosystema 2020, 39, 653–681. [Google Scholar] [CrossRef]
Figure 1.
Maximum Parsimony strict consensus tree illustrating the phylogeny of three new species and related species in Hyphoderma within Polyporales based on internal transcribed spacer + nuclear ribosomal RNA gene (ITS+nLSU) sequences. Branches are labeled with maximum likelihood bootstrap values > 70%, parsimony bootstrap values > 50% and Bayesian posterior probabilities > 0.95. The new species are in bold. Clade names follow previous study [19].
Figure 1.
Maximum Parsimony strict consensus tree illustrating the phylogeny of three new species and related species in Hyphoderma within Polyporales based on internal transcribed spacer + nuclear ribosomal RNA gene (ITS+nLSU) sequences. Branches are labeled with maximum likelihood bootstrap values > 70%, parsimony bootstrap values > 50% and Bayesian posterior probabilities > 0.95. The new species are in bold. Clade names follow previous study [19].

Figure 2.
Hyphoderma crystallinum (holotype) (A): basidiomata on the substrate (B); scattered nubby crystals. Bars: A = 2 cm and B = 1 mm.
Figure 2.
Hyphoderma crystallinum (holotype) (A): basidiomata on the substrate (B); scattered nubby crystals. Bars: A = 2 cm and B = 1 mm.

Figure 3.
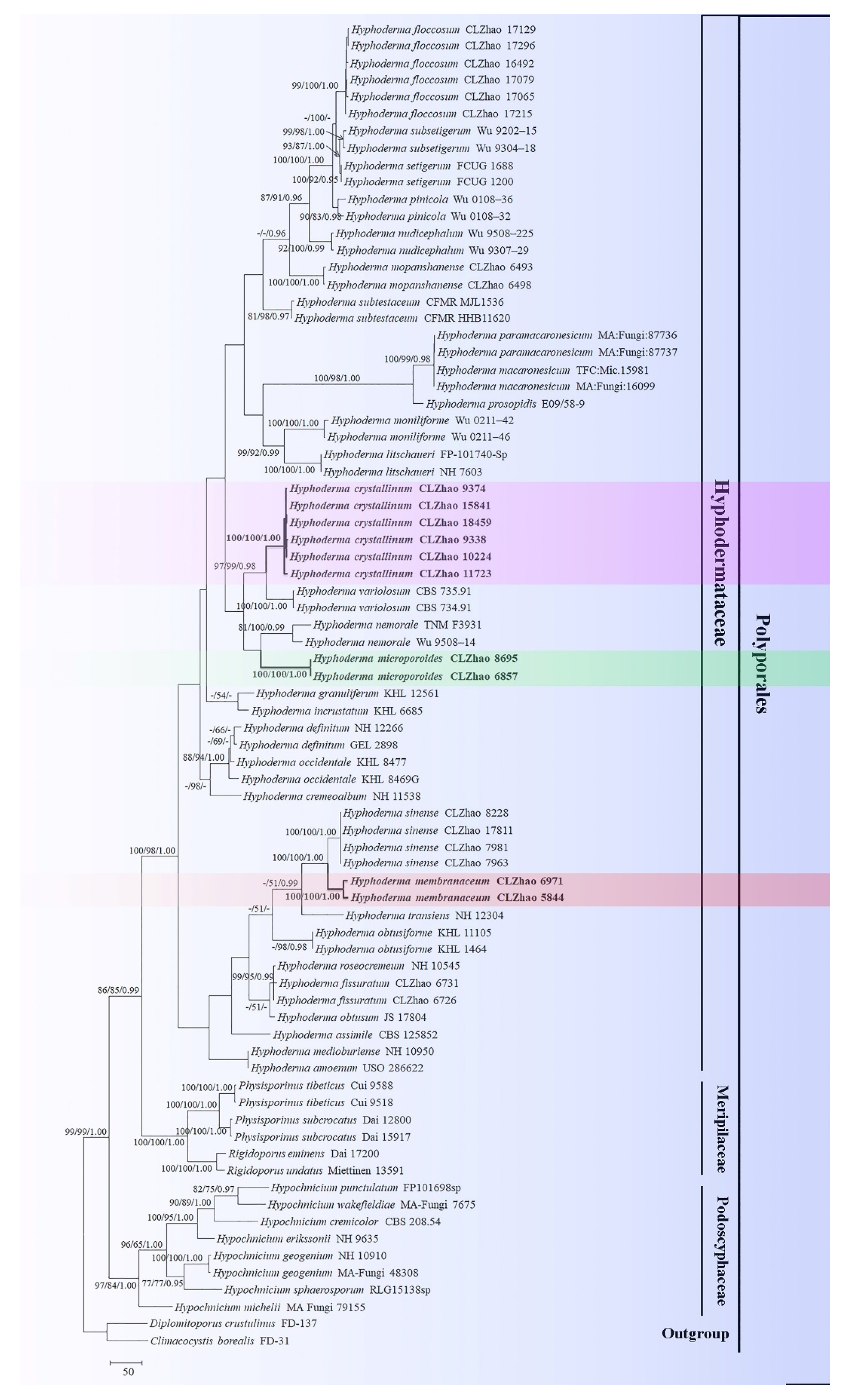
Microscopic structures of Hyphoderma crystallinum (holotype) (A): basidiospores (B), basidia and basidioles (C), tubular cystidia (D), and encrusted cystidia (E). A section of hymenium. Bars: A–E = 10 µm.
Figure 3.
Microscopic structures of Hyphoderma crystallinum (holotype) (A): basidiospores (B), basidia and basidioles (C), tubular cystidia (D), and encrusted cystidia (E). A section of hymenium. Bars: A–E = 10 µm.

Figure 4.
Basidiomata (A) of Hyphoderma membranaceum (B) (holotype). Bars: A = 2 cm and B = 1 mm.

Figure 5.
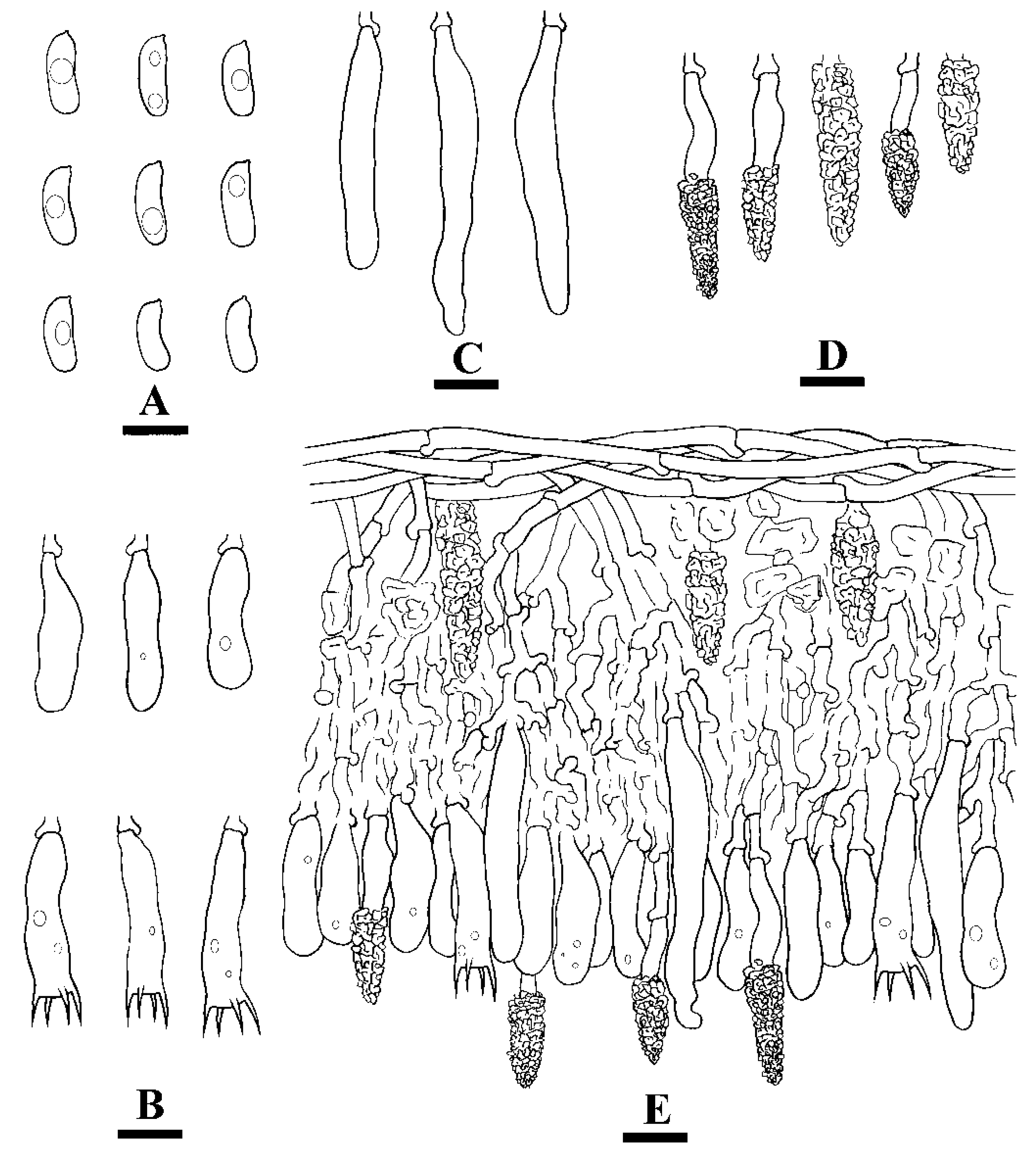
Microscopic structures of Hyphoderma membranaceum (holotype) (A): basidiospores (B), basidia and basidioles (C), and cystidia (D). A section of hymenium. Bars: A–D = 10 µm.
Figure 5.
Microscopic structures of Hyphoderma membranaceum (holotype) (A): basidiospores (B), basidia and basidioles (C), and cystidia (D). A section of hymenium. Bars: A–D = 10 µm.

Figure 6.
Hyphoderma microporoides (holotype) (A): basidiomata on the substrate (B) and scattered pinholes. Bars: A = 2 cm, B = 1 mm.
Figure 6.
Hyphoderma microporoides (holotype) (A): basidiomata on the substrate (B) and scattered pinholes. Bars: A = 2 cm, B = 1 mm.

Figure 7.
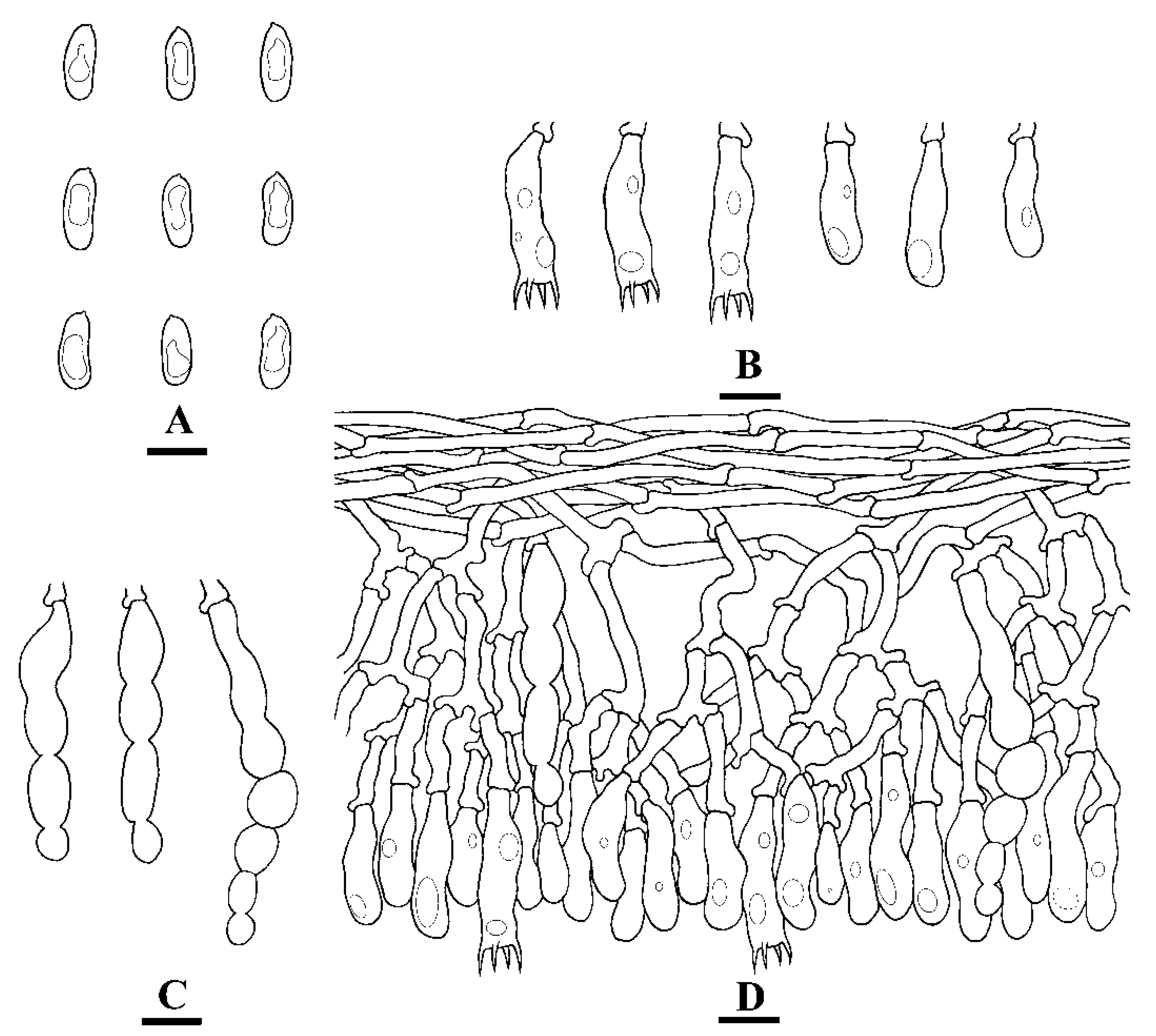
Microscopic structures of Hyphoderma microporoides (holotype) (A): basidiospores (B), basidia and basidioles (C), and cystidia (D). A section of hymenium. Bars: A = 5 µm, B–D = 10 µm.
Figure 7.
Microscopic structures of Hyphoderma microporoides (holotype) (A): basidiospores (B), basidia and basidioles (C), and cystidia (D). A section of hymenium. Bars: A = 5 µm, B–D = 10 µm.

Figure 8.
Geographic distribution of Hyphoderma species treated in the phylogenetic tree.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of species, specimens, and GenBank accession numbers of sequences used in this study.
| Species Name | Specimen No. | GenBank Accession No. | References | |
|---|---|---|---|---|
| ITS | LSU | |||
| Climacocystis borealis | FD-31 | KP135308 | KP135210 | [19] |
| Diplomitoporus crustulinus | FD-137 | KP135299 | KP135211 | [19] |
| Hyphoderma amoenum | USO 286622 | HE577030 | [17] | |
| H. assimile | CBS 125852 | MH863808 | MH875272 | [24] |
| H. cremeoalbum | NH 11538 | DQ677492 | DQ677492 | [16] |
| H. crystallinum | CLZhao 9338 | MW917161 | MW913414 | Present study |
| H. crystallinum | CLZhao 9374 | MW917162 | MW913415 | Present study |
| H. crystallinum | CLZhao 10224 | MW917163 | MW913416 | Present study |
| H. crystallinum | CLZhao 11723 | MW917164 | MW913417 | Present study |
| H. crystallinum | CLZhao 15841 | MW917165 | MW913418 | Present study |
| H. crystallinum | CLZhao 18459 | MW917166 | MW913419 | Present study |
| H. definitum | GEL 2898 | AJ406509 | [18] | |
| H. definitum | NH 12266 | DQ677493 | DQ677493 | [16] |
| H.fissuratum | CLZhao 6731 | MT791331 | MT791335 | [15] |
| H.fissuratum | CLZhao 6726 | MT791330 | MT791334 | [15] |
| H. floccosum | CLZhao 17129 | MW301683 | MW293733 | [25] |
| H. floccosum | CLZhao 17296 | MW301686 | MW293736 | [25] |
| H. floccosum | CLZhao 16492 | MW301688 | MW293734 | [25] |
| H. floccosum | CLZhao 17215 | MW301687 | MW293735 | [25] |
| H. floccosum | CLZhao 17079 | MW301685 | [25] | |
| H. floccosum | CLZhao 17065 | MW301684 | [25] | |
| H. granuliferum | KHL 12561 | JN710545 | JN710545 | [18] |
| H. incrustatum | KHL 6685 | AY586668 | [18] | |
| H. litschaueri | NH 7603 | DQ677496 | DQ677496 | [16] |
| H. litschaueri | FP-101740- sp | KP135295 | KP135219 | [26] |
| H. macaronesicum | MA:Fungi:16099 | HE577027 | [18] | |
| H. macaronesicum | TFC:Mic.15981 | HE577028 | [18] | |
| H. medioburiense | NH 10950 | DQ677497 | DQ677497 | [16] |
| H. membranaceum | CLZhao 5844 | MW917167 | MW913420 | Present study |
| H. membranaceum | CLZhao 6971 | MW917168 | MW913421 | Present study |
| H. microporoides | CLZhao 6857 | MW917169 | MW913422 | Present study |
| H. microporoides | CLZhao 8695 | MW917170 | MW913423 | Present study |
| H. moniliforme | Wu 0211-42 | KC928282 | KC928283 | [27] |
| H. moniliforme | Wu 0211-46 | KC928284 | KC928285 | [27] |
| H. mopanshanense | CLZhao 6498 | MT791329 | MT791333 | [15] |
| H. mopanshanense | CLZhao 6493 | MT791328 | MT791332 | [15] |
| H. nemorale | TNM F3931 | KJ885183 | KJ885184 | [27] |
| H. nemorale | Wu 9508-14 | KC928280 | KC928281 | [27] |
| H. nudicephalum | Wu 9307-29 | AJ534269 | [28] | |
| H. nudicephalum | Wu 9508-225 | AJ534268 | [28] | |
| H. obtusiforme | KHL 1464 | JN572909 | [29] | |
| H. obtusiforme | KHL 11105 | JN572910 | [29] | |
| H. obtusum | JS 17804 | AY586670 | [29] | |
| H. occidentale | KHL 8469 | AY586674 | [29] | |
| H. occidentale | KHL 8477 | DQ677499 | DQ677499 | [16] |
| H. paramacaronesicum | MA:Fungi:87736 | KC984399 | KF150074 | [14] |
| H. paramacaronesicum | MA:Fungi:87737 | KC984405 | KF150073 | [14] |
| H. pinicola | Wu 0108-32 | KJ885181 | KJ885182 | [29] |
| H. pinicola | Wu 0108-36 | KC928278 | KC928279 | [29] |
| H. prosopidis | E09/58-9 | HE577029 | [29] | |
| H. roseocremeum | NH 10545 | AY586672 | [29] | |
| H. setigerum | FCUG 1200 | AJ534273 | [28] | |
| H. setigerum | FCUG 1688 | AJ534272 | [28] | |
| H. sinense | CLZhao 7963 | MW301679 | MW293730 | [25] |
| H. sinense | CLZhao 17811 | MW301682 | MW293732 | [25] |
| H. sinense | CLZhao 8228 | MW301681 | [25] | |
| H. sinense | CLZhao 7981 | MW301680 | MW293731 | [25] |
| H. subsetigerum | Wu 9304-18 | AJ534277 | [28] | |
| H. subsetigerum | Wu 9202-15 | AJ534278 | [28] | |
| H. subtestaceum | HHB11620 | GQ409521 | [29] | |
| H. subtestaceum | CFMR MJL1536 | GQ409522 | [29] | |
| H. transiens | NH 12304 | DQ677504 | DQ677504 | [16] |
| H. variolosum | CBS 734.91 | MH862320 | MH873992 | [24] |
| H. variolosum | CBS 735.91 | MH862321 | MH873993 | [24] |
| Hypochnicium cremicolor | CBS 208.54 | MH857294 | MH868826 | [24] |
| H. erikssonii | NH 9635 | DQ677508 | DQ677508 | [16] |
| H. geogenium | NH 10910 | DQ677509 | DQ677509 | [16] |
| H. geogenium | MA-Fungi 48308 | FN552534 | JN939576 | [30] |
| H. michelii | MA-Fungi 79155 | NR119742 | NG060635 | [30] |
| H. punctulatum | FP101698sp | KY948827 | KY948860 | [19] |
| H. sphaerosporum | RLG15138sp | KY948803 | KY948861 | [19] |
| H. wakefieldiae | MA-Fungi 7675 | FN552531 | JN939577 | [30] |
| Physisporinus subcrocatus | Dai 15917 | KY131870 | KY131926 | [31] |
| P. subcrocatus | Dai 12800 | KY131869 | KY131925 | [31] |
| P. tibeticus | Cui 9588 | KY131873 | KY131929 | [31] |
| P. tibeticus | Cui 9518 | KY131872 | KY131928 | [31] |
| Rigidoporus eminens | Dai 17200 | MT279690 | MT279911 | [31] |
| R. undatus | Miettinen-13591 | KY948731 | KY948870 | [19] |
New sequences are shown in bold.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guan, Q.-X.; Zhao, C.-L. Taxonomy and Phylogeny of the Wood-Inhabiting Fungal Genus Hyphoderma with Descriptions of Three New Species from East Asia. J. Fungi 2021, 7, 308. https://doi.org/10.3390/jof7040308
AMA Style
Guan Q-X, Zhao C-L. Taxonomy and Phylogeny of the Wood-Inhabiting Fungal Genus Hyphoderma with Descriptions of Three New Species from East Asia. Journal of Fungi. 2021; 7(4):308. https://doi.org/10.3390/jof7040308
Chicago/Turabian StyleGuan, Qian-Xin, and Chang-Lin Zhao. 2021. "Taxonomy and Phylogeny of the Wood-Inhabiting Fungal Genus Hyphoderma with Descriptions of Three New Species from East Asia" Journal of Fungi 7, no. 4: 308. https://doi.org/10.3390/jof7040308
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.