
Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water
by
Huang Zhang
1,
Rong Zhu
2,
Yun Qing
3,
Hao Yang
3,4,
Chunxue Li
3,
Gennuo Wang
2,
Di Zhang
1,* and
Ping Ning
2,* 1
Shandong Provincial Key Laboratory of Water and Soil Conservation and Environmental Protection, College of Resources and Environment, Linyi University, West Side of North Section of Industrial Avenue, Linyi 276000, China
2
Faculty of Environmental Science and Engineering, Kunming University of Science & Technology, Kunming 650500, China
3
Faculty of Food Science and Engineering, Kunming University of Science & Technology, Kunming 650500, China
4
Key Laboratory for Humid Subtropical Eco-Geographical Processes of the Ministry of Education, Fujian Normal University, Fuzhou 350007, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(10), 1063; https://doi.org/10.3390/jof8101063
Submission received: 20 July 2022
/
Revised: 14 August 2022
/
Accepted: 22 August 2022
/
Published: 10 October 2022
(This article belongs to the Special Issue Polyphasic Identification of Fungi)
Abstract
:Twelve new specimens of sporidesmium-like taxa were collected from freshwater habitats in China and Thailand. Phylogenetic analysis of nuc 28S rDNA (LSU), internal transcribed spacer (ITS), translation elongation factor 1-alpha (TEF1-α) and second-largest subunit of RNA polymerase II (RPB2) sequence data, combined with morphological data, revealed that they are Distoseptispora species. Among them, six new species, including D. aqualignicola, D. aquamyces, D. crassispora, D. curvularia, D. nonrostrata and D. pachyconidia, are introduced. Two new combinations, D. adscendens and D. leonensis, are transferred from Ellisembia. A new habitat and geographical record are reported for D. clematidis, collected from a freshwater habitat in China. New RPB2 sequence data for D. dehongensis are provided.
1. Introduction
Freshwater fungi have been reported in the Chytridiomycota, Zygomycota, Ascomycota and Basidiomycota, and their asexual morphs are the most widely distributed groups in fresh water [1]. In previous studies, sporidesmium-like hyphomycetes were commonly found on submerged wood in streams [2,3,4,5,6,7,8,9]. Sporidesmium was established by Link [10] in the early 1800s, with S. atrum as the type species. Ellis [11,12] retrieved and redescribed the genus as having unbranched conidiophores; integrated, terminal, monoblastic, determinate or percurrent conidiogenous cells; and acrogenous, solitary, transversely septate or distoseptate conidia. According to the broad circumscription, more than 400 Sporidesmium species were introduced, mostly based on morphology alone. To deal with a heterogeneous assemblage of species under Sporidesmium, Subramanian [13] reassessed the genus and introduced seven new genera, namely, Acarocybellina, Ellisembia, Gangliophora, Hemicorynesporella, Penzigomyces, Repetophragma and Stanjehughesia, to accommodate some species on the basis of conidial septation (euseptate vs. distoseptate) and conidiogenesis. Wu and Zhuang [14] placed Penzigomyces in Sporidesmium and Imicles in Ellisembia, thereby expanding the generic concepts of Sporidesmium (euseptate) and Ellisembia (distoseptate) to include fungi with no percurrent conidiophores or lageniform, doliiform or nodose percurrently extending conidiophores.
With increasing molecular data, multi-locus phylogenetic relationships between Sporidesmium and Ellisembia species have been investigated to gain a better understanding of their taxonomy, and it turned out that these species were distributed in different subclasses in Sordariomycetes, including Diaporthomycetidae, Hypocreomycetidae, Sordariomycetidae and Xylariomycetidae [5,7]. Therefore, some previously described species were revised and transferred to newly established genera, such as Pseudosporidesmium. The newly collected sporidesmium-like taxa were placed in new genera, such as Pseudostanjehughesia and Distoseptispora, and Ellisembia was synonymized with Sporidesmium [7].
Distoseptispora was introduced for sporidesmium-like species with macronematous, unbranched, olive-green, cylindrical conidiophores; monoblastic, integrated, determinate, terminal, cylindrical conidiogenous cells; and acrogenous, distoseptate, cylindrical, smooth, darker conidia with slightly paler, but not hyaline, rounded apices of indeterminate length. Su et al. [7] introduced two new species, D. fluminicola (type) and D. aquatica, found on submerged wood in China, and also included two taxa, Ellisembia adscendens and E. Leonensis, named Distoseptispora adscendens and D. leonensis in their phylogenetic tree, but did not validly introduce them as new combinations. Subsequently, the generic concept of Distoseptispora was emended to include percurrently elongating conidiogenous cells and euseptate, verrucose conidia with percurrent proliferation, based on the newly introduced species D. guttulata and D. suoluoensis [5]. In the past five years, more than 40 new species (excluding D. submersa) were successively reported in Thailand and China [3,5,7,8,15,16,17,18,19,20,21,22,23,24,25]. Distoseptispora submersa, found on submerged wood in China, was introduced by Luo et al. [17]. The authors stated that it was phylogenetically close to D. tectonae but had larger conidiophores and shorter conidia. However, Dong et al. [9] synonymized D. submersa with D. tectonae due to the small nucleotide differences discovered between these two species and identification as D. tectonae extending to larger conidiophores.
Recently, Yang et al. [24] used multi-locus analysis and reported, for the first time, a sexual morph in Distoseptispora, namely, D. hyalina, which was characterized by an immersed to semi-immersed, subglobose to ellipsoidal, dark-brown perithecial with a short neck; a relatively thick peridium; sparse, persistent, hyaline paraphyses; cylindrical asci with non-amyloid apical annuli; and hyaline, filiform, 0–3-septate ascospores with mucilaginous sheaths. To date, it is the only known sexual morph in Distoseptispora. The hosts of Distoseptispora species are diverse, including Carex sp., Pandanus sp. and Tectona grandis, as well as bamboo [3,18,19]. Distoseptispora is the only genus in Distoseptisporaceae, Distoseptisporales, Diaporthomycetidae and Sordariomycetes.
In this study, 12 fresh specimens of sporidesmium-like taxa were collected from freshwater habitats in China and Thailand and were preliminarily identified as Distoseptispora species. In order to clarify the classification of these specimens, an updated phylogeny of Distoseptisporales and their relatives is provided based on a concatenated nuc 28S rRNA (LSU)–internal transcribed spacer (ITS)–translation elongation factor 1-alpha (TEF1-α)–second-largest subunit of RNA polymerase II (RPB2) dataset. Six new species and two new combinations are introduced in this study based on phylogenetic analyses combined with morphological data. The characters of species are discussed.
2. Materials and Methods
2.1. Sample Collection, Morphological Studies and Isolation
In this study, decaying wood samples were collected from various regions of Thailand (Chiang Rai Province) and China (Sichuan and Yunnan Provinces). Decaying wood samples were place in zip-lock plastic bags with sterile wet cotton. The locations, collectors, dates of collection and countries were recorded for all collections, and they were taken to the laboratory for detailed observation and the isolation of pure cultures.
Fungal structures were observed and examined using a Nikon SMZ–171 dissecting microscope. Micro-morphological characters were examined using a Nikon ECLIPSE Ni compound microscope and photographed with a Canon 600D digital camera fitted to the microscope. The Tarosoft (R) Image Frame Work program and the Oplenic program were used to measure the fungal structures. Adobe Photoshop CS6 extended v. 13.0 software (Adobe Systems, San Jose, CA, USA) was used to process images. Single-spore isolation was obtained following the method described in Chomnunti et al. [26]. Germinated spores were aseptically transferred to potato dextrose agar (PDA) and incubated at 20–25 °C for 2–4 weeks. Colony characteristics were recorded, including color and shape. Dry specimens were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand, and the Kunming Institute of Botany Academia Sinica (HKAS), Kunming, China. Living cultures were deposited in the Mae Fah Luang University Culture Collection (MFLUCC) and the Kunming Institute of Botany Culture Collection (KUMCC). Facesoffungi and Index Fungorum numbers were acquired according to Jayasiri et al. [27] and Index Fungorum (http://www.indexfungorum.org/names/names.asp, accessed on 15 March 2021), respectively.
2.2. DNA Extraction, PCR Amplification and Sequencing
Fresh fungal mycelia were scraped off with a needle and transferred into a 1.5 mL centrifuge tube. A Plant genomic DNA extraction kit (generic) (TreliefJM, Kunming, China) was used to extract DNA from fresh mycelia, according to the manufacturer’s instructions.
DNA amplification for LSU, ITS, TEF1-α and RPB2 genes was performed by polymerase chain reaction (PCR). The primers and PCR thermal cycle programs are shown in Table 1. PCR products were confirmed on 1% agarose electrophoresis gels stained with ethidium bromide. PCR products were sent to TsingKe Biological Technology (Kunming, China) for purification and sequencing.
2.3. Phylogenetic Analyses
The raw sequences were initially checked with Finch TV v. 1.4.0, and each gene was subjected to a BlastN search in NCBI’s GenBank to assess the confidence level. Sequences of Distoseptisporaceae species, along with the newly generated sequences and related taxa, following Dong et al. [9] and Phukhamsakda et al. [25], were downloaded from GenBank. The gene sequences were aligned using the online multiple alignment program MAFFT v. 7 (http://mafft.cbrc.jp/alignment/server/index.html, accessed on 7 April 2022) [32] and manually improved, where necessary, in BioEdit v. 7.2 [33], then concatenated with Mesquite v. 3.11.
Maximum likelihood (ML) and Bayesian inference (BI) methods were used for the phylogenetic analyses. Maximum likelihood analysis was performed with RAxML-HPC v. 8 on XSEDE in the CIPRES Science Gateway (https://www.phylo.org/portal2/home.action/, accessed on 8 April 2022) [34,35], with the following changes made to the default settings: maximum hours to run: 10; model for bootstrapping phase: GTRGAMMA; analysis type: rapid bootstrap analysis/search for best-scoring ML tree (-f a); bootstrapping type: rapid bootstrapping (-x); bootstrap iterations: 1000 (the maximum value allowed).
For BI analysis, the best model for each gene was selected using MrModelTest 2.3 [36], and GTR+I+G was selected as the best-fitting model for LSU, ITS TEF1-α and RPB2. BI analysis was conducted using MrBayes 3.2.6 for posterior probabilities (PPs) by Markov chain Monte Carlo sampling (BMCMC) [37,38]. Six simultaneous Markov chains were run for one million generations, and trees were sampled every 100 generations. The first 2500 trees, representing the burn-in phase of the analyses, were discarded and the remaining trees were used for the calculation of PPs in the majority rule consensus tree (the critical value for the topological convergence diagnostic was 0.01). The phylogenetic tree was visualized in FigTree v. 1.4.2 [39] and edited in Adobe Illustrator CS6 (Adobe Systems, USA). Newly generated sequences were submitted to GenBank. All of the sequences used in this study are listed in Table 2.
3. Results
3.1. Phylogenetic Analyses
The combined LSU, ITS, TEF1-α and RPB2 dataset was analyzed for species of Distoseptisporales and their relatives, using Myrmecridium banksiae (CPC 19852) and Myrmecridium schulzeri (CBS 100.54) as outgroup taxa. The dataset consists of 94 strains with an alignment length of 3743 total characters. The RAxML analysis resulted in a best-scoring likelihood tree selected with a final ML optimization likelihood value of −36376.671888. The matrix had 1984 distinct alignment patterns, with 35.27% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.241400, C = 0.263989, G = 0.281489, T = 0.213122; substitution rates AC = 1.230237, AG = 3.213547, AT = 1.297497, CG = 0.852333, CT = 6.671646, GT = 1.000000; gamma distribution shape parameter α = 0.254430. The phylogenetic trees generated from BI analyses were topologically similar to the one generated via the ML analyses, and the latter is shown in Figure 1.
The genus Distoseptispora consists of three well-supported clades: nine of our new isolates clustered in Clade 1, and another three clustered in Clade 3. Six of them were identified as new species which combined the morphological characteristics described below, viz., D. aquamyces, D. aqualignicola, D. crassispora, D. curvularia, D. nonrostrata and D. pachyconidia. MFLU 19–0544 and MFLU 17–1671 clustered with D. dehongensis, with strong support (100% MLBS/1.00 BIPP). KUNCC 21–10732, KUNCC 21–10727, MFLU 19–0543 and KUNCC 21–10728 clustered with D. bambusae, D. clematidis, D. rayongensis and D. tectonae, respectively, with high bootstrap support.
3.2. Taxonomy in Distoseptispora
Distoseptispora adscendens (Berk.) R. Zhu & H. Zhang, comb. nov. (Figure 2).
Basionym: Sporidesmium adscendens Berk., Ann. nat. Hist., Mag. Zool. Bot. Geol. 4: 292 (1840).
Synonym: Clasterosporium adscendens (Berk.) Sacc., Syll. fung. (Abellini) 4: 394 (1886).
Ellisembia adscendens (Berk.) Subram., Proc. Indian natn Sci. Acad., Part B. Biol. Sci. 58(4): 183 (1992).
Sexual morph: Miyoshiella triseptata (Shoemaker & G.P. White) Réblová, Mycotaxon 71: 33 (1999).
Synonym: Lasiosphaeria triseptata Shoemaker & G.P. White, Sydowia 38: 282 (1986).
Index Fungorum number: 559917; Facesoffungi number: 12574.
Notes: Sporidesmium adscendens was synonymized as Ellisembia adscendens on the basis of pseudoseptate conidia and conidiophores with irregular percurrent proliferation [13]. An ascomycetes, Miyoshiella triseptata (≡Lasiosphaeria triseptata), collected in Hungary, was found to be associated with S. adscendens in the host of Platanus orientalis and therefore reported as its sexual morph [40,41]. Shenoy et al. [42] reported a strain, HKUCC 10820, of E. adscendens with LSU and RPB2 sequence data. The strain was later proved to phylogenetically cluster in the genus Distoseptispora by Su et al. [7] and was therefore named Distoseptispora adscendens in their phylogenetic tree but not validly introduced as a new combination. Many subsequent authors followed them in using the name D. adscendens [5,8,17,23].
The morphology of strain HKUCC 10820 was referred to Wu and Zhuang [14, P107], who described it as having macronematous, mononematous, cylindrical, straight or slightly flexuous, smooth, 1–4-septate, brown to dark-brown conidiophores 20–50 μm long, 7–10 μm wide; integrated, terminal, cylindrical, smooth, brown to dark-brown conidiogenous cells with a size of 5–10 × 3–4.5 μm; and acrogenous, cylindrical to obclavate, rostrate, 16–62-distoseptate conidia 110–375 μm long, 14–20 μm wide at the broadest part, with rounded apexes 5–10 μm wide and subcylindrical to conical–truncate basal cells 5–7 μm wide at the base. This description is consistent with that of S. adscendens given in Ellis [12], even with respect to the sizes of the conidiophores and conidia. The above characters fit the generic features of Distoseptispora well. Therefore, we formally place E. adscendens in Distoseptispora, as D. adscendens.
Distoseptispora aqualignicola C.X. Li & H. Zhang, sp. nov. (Figure 3).
Index Fungorum number: 559918; Facesoffungi number: 12575.
Holotype: HKAS 122184.
Etymology: “Aqua-” refers to the aquatic habitat in which the fungus was found, while “lignicola” refers to the fact that the fungus was found on a lignicolous substrate.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, scattered, hairy, brown to dark-brown. Mycelium partly superficial, partly immersed, composed of branched, septate, smooth, hyaline to pale-brown hyphae. Conidiophores 90–190(–240) μm long ( = 162 µm, n = 15) and 5–8 μm wide ( = 6 µm, n = 15), macronematous, mononematous, unbranched , multi-septate, single or in groups of two or three, cylindrical, straight or slightly flexuous, smooth, brown, rounded at the apex. Conidiogenous cells 13–21 μm long ( = 18 µm, n = 10), 4–5.5 μm wide ( = 4.5 µm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, brown, smooth. Conidia 41–94(–104) μm long ( = 73 µm, n = 20) (rostrum included), 10.5–12.5 µm at the widest part ( = 11.5 µm, n = 20), 2–5 µm wide at the apex ( = 3.5 µm, n = 20), acrogenous, dry, obclavate, rostrate, straight or curved, 4–8-euseptate, mostly 6–7-euseptate, tapering towards the rounded apex, brown at the base, smooth, thin-walled, subhyaline to pale-brown at the apex.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the conidial apex. Colonies on PDA reaching 34 mm diameter after 22 days at 20–25 °C, circular, dry, dark-brown to black on surface and reverse, raised, with entire margin.
Material examined: China, Sichuan Province, Yibin City, Southern Sichuan Bamboo Sea, Qicai Lake, found on dead, submerged, decaying wood of unidentified plants, 16 June 2019, Chunxue Li, S1–10 (HKAS 122184, holotype), ex-type living culture KUNCC 21–10729.
Notes: Distoseptispora aqualignicola, D. aquamyces,D. lancangjiangensis,D. meilingensis, D. suoluoensis, D. yongxiuensis and D. verrucose form a strongly supported clade (99% MLBS/1.00 BIPP) in Clade 2 of Distoseptispora (Figure 1). Morphologically, D. aqualignicola possesses smooth-walled conidia, which are distinct from the verrucose conidia of D. aquamyces, D. suoluoensis and D. verrucosa [5,24]. Distoseptispora aqualignicola differs from D. meilingensis in having euseptate, thin-walled conidia as compared with the distoseptate, thick-walled conidia of D. MEILINGENSIS [43]. Distoseptispora lancangjiangensis and D. yongxiuensis resemble D. aqualignicola with respect to their smooth-walled conidia but they are phylogenetically distinct.
Distoseptispora aquamyces R. Zhu & H. Zhang, sp. nov. (Figure 4).
Index Fungorum number: 559919; Facesoffungi number: 12576.
Holotype: HKAS 122186.
Etymology: “Aqua-” refers to the aquatic habitat in which the fungus was found.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum: superficial, effuse, scattered, hairy, brown to dark-brown. Mycelium partly superficial, partly immersed, composed of branched, septate, smooth, hyaline to pale-brown hyphae. Conidiophores (78–)91–198 μm long ( = 127 µm, n = 10), 4–7 μm wide ( = 5 µm, n = 10), macronematous, mononematous, unbranched, multi-septate, single or in groups of two or three, cylindrical, straight or slightly flexuous, smooth, pale-brown. Conidiogenous cells 10–20 μm long ( = 16 µm, n = 10), 4.5–5.5 μm wide ( = 5 µm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, pale-brown, smooth. Conidia 30–95 μm long ( = 73 µm, n = 20) (rostrum included), 7–12 µm at the widest part ( = 8 µm, n = 20), 2–5 µm wide at the apex ( = 3 µm, n = 20), acrogenous, dry, obclavate to obpyriform, mostly rostrate, straight or curved, 4–10-euseptate, mostly 6–9-euseptate, tapering towards the rounded apex, verrucose, thin-walled, pale-brown to brown at the base, subhyaline to pale-brown at the apex, verrucose.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced at end of conidia. Colonies on PDA reaching 40 mm diameter after 45 days at 20–25 °C, circular, flat, surface rough, grey from above, dark-brown from below, edge entire.
Material examined: China, Sichuan Province, Yibin City, Southern Sichuan Bamboo Sea, Qicai Lake, found on dead, submerged, decaying wood of unidentified plants, 3 November 2020, Yun Qing, SN–18 (HKAS 122186, holotype), ex-type living culture KUNCC 21–10731.
Notes: Distoseptispora aquamyces differs from D. suoluoensis in having pale-brown to brown, shorter conidia (30–95 μm vs. 80–125 μm), sometimes with percurrent proliferation, while D. aquamyces has yellowish-brown or dark-olivaceous conidia without proliferation. In our phylogenetic tree, Distoseptispora aquamyces is close to D. suoluoensis, with low support. Two species in Distoseptispora possess verrucose conidia, i.e., D. suoluoensis and D. verrucose. Distoseptispora verrucose has percurrently proliferating conidiogenous cells and olivaceous conidia which differ from the non-proliferating conidiogenous cells and pale-brown to brown conidia of D. aquamyces. More importantly, D. aquamyces is distinct from D. suoluoensis and D. verrucose in our phylogenetic tree. Therefore, D. aquamyces is introduced as a new species.
Distoseptispora bambusae Y.R. Sun, I.D. Goonasekara, Yong. Wang bis & K.D. Hyde, Biodiversity Data Journal 8(e53678): 6 (2020) (Figure 5).
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, hairy, black. Mycelium mostly immersed, composed of branched, septate, smooth, pale-brown hyphae. Conidiophores (39–)45–90 μm long ( = 64 µm, n = 10), 3.5–5 μm wide ( = 4.5 µm, n = 10), macronematous, mononematous, septate, unbranched, single or in groups of two or three, cylindrical, straight or slightly flexuous, smooth, pale-brown, robust at the base. Conidiogenous cells 12–20 μm long ( = 15 µm, n = 10), 4–5 μm wide ( = 4 µm, n = 10), monoblastic or polyblastic, integrated, determinate, terminal, cylindrical, pale-brown, smooth. Conidia (29–)43–94(–105) μm long ( = 71 μm, n = 25) (rostrum included), 6–8.5 μm at the widest part ( = 7 μm, n = 25), 2–3 µm wide at the apex ( = 2.5 µm, n = 25), acrogenous, solitary, obclavate, rostrate, straight or curved, (4–)7–13-euseptate, rounded at the apex, truncate at the base, tapering towards apex, smooth, thin-walled, pale-brown or brown.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the two sides of the conidia. Colonies on PDA reaching 40 mm diameter after 40 days at 20–25 °C, circular, flat, rough surface, grey from above, dark-brown from below, edge entire.
Material examined: China, Sichuan Province, Yinbin City, Southern Sichuan Bamboo Sea, Qicai Lake, found on dead, submerged, decaying wood of unidentified plants, 13 November 2020, Yun Qing, SN–34 (HKAS 122187), living culture KUNCC 21–10732.
Notes: Distoseptispora bambusae was initially collected from culms of bamboo by Sun et al. [22] in Guizhou, China, and Chiangrai, Thailand. The morphological characters and size of our new collection are consistent with the holotype, except for the wider range of conidial length ((29–)43–94(–105) μm vs. 45–74 μm). Phylogenetically, the new collection clusters with D. bambusae, with strong support (100% MLBS/1.00 BIPP; Figure 1). Our isolate was collected from dead, submerged, decaying wood, which constitutes a new habitat record.
Distoseptispora clematidis Phukhams., M.V. de Bult & K.D. Hyde, Fungal Diversity 102(1): 168 (2020) (Figure 6).
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, scattered, hairy, dark-brown. Mycelium mostly immersed, composed of branched, septate, smooth, dark-brown hyphae. Conidiophores (8–)15–25 μm long ( = 20 µm, n = 10), 5–8 μm wide ( = 7 µm, n = 10), macronematous, mononematous, solitary, unbranched, single or in groups of two or three, 2–5-septate, cylindrical, straight or slightly flexuous, smooth, dark-brown to brown, robust at the base. Conidiogenous cells 4–7 μm long ( = 5 µm, n = 10), 4–6 μm wide ( = 5 µm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, dark-brown, smooth. Conidia (90–)126–245(–303) µm long (rostrum included) ( = 185 µm, n = 30), 14–20 µm at the widest part ( = 16 µm, n = 30), 7–10 µm wide at the apex ( = 8.5 µm, n = 30), acrogenous, dry, obclavate, elongated, straight or curved, (17–)20–41(–49)-distoseptate, truncate at the base, rounded at the apex, smooth, thick-walled, brown with a green tinge, sometimes with percurrent proliferation and forming another conidium.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the conidial base. Colonies on PDA reaching 34 mm diameter after 3 weeks at 20–25 °C, brown with sparse mycelium from above, dark-brown from below, rough surface, dry, flat, entire at edge.
Material examined: China, Yunnan Province, Xishuangbanna, Man Feilong Reservoir, found on dead, submerged, decaying wood of unidentified plants, 7 November 2020, Rong Zhu, N76 (HKAS 122182), living culture KUNCC 21–10727.
Notes: Distoseptispora clematidis was introduced by Phukhamsakda et al. [21] and was found on a dried stem of Clematis sikkimensis in Thailand. The new isolate KUNCC 21–10727 clusters with the type strain of D. clematidis, with strong support (98% MLBS/1.00 BIPP; Figure 1). Morphologically, our isolate has the same characters as the holotype of D. clematidis, including conidial size and conidial septa number, except for slightly shorter conidiophores ((8–)15–25 μm vs. 22–40 μm)) [21]. The conidia of the new isolate sometimes have percurrent proliferation and form another conidium from the conidial apex, which was not reported in the original description of D. clematidis, though it is obvious in picture f of Figure 109 in Phukhamsakda et al.’s [21]. Comparisons of sequence data for KUNCC 21–10727 and the ex-type strain showed nine (1%, including one gap), nine (1.6%, no gaps) and five (0.7%, including one gap) nucleotide differences in the LSU, ITS and RPB2 regions, respectively. Although the nucleotide differences in the ITS regions were more than 1.5%, which is high enough to establish a new species, according to Jeewon and Hyde [44], there were no significant morphological differences between our isolate and the holotype. Therefore, we propose to identify the new isolate as D. clematidis until more strains have been examined. The new isolate was collected from submerged, decaying wood in China, which constitutes a new habitat and geographical record.
Distoseptispora crassispora R. Zhu & H. Zhang, sp. nov. (Figure 7).
Index Fungorum number: 559920; Facesoffungi number: 12577.
Holotype: HKAS 122181.
Etymology: “Crass” means thick and refers to the thick-walled conidia.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, scattered, hairy, dark-brown. Mycelium mostly immersed, composed of branched, septate, smooth, dark-brown hyphae. Conidiophores 14–27 μm long, 6–10 μm wide, macronematous, mononematous, solitary, unbranched, septate, cylindrical, straight or flexuous, smooth, brown to dark-brown, robust at the base. Conidiogenous cells 2.5–7 μm long ( = 4.5 μm, n = 10), 5.5–8 μm wide ( = 6 μm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, dark-brown, smooth. Conidia 95–197(–214) µm long (rostrum included) ( = 141 μm, n = 30), 13–24 μm at the widest part ( = 17 μm, n = 30), 6–12 μm wide at the apex ( = 9 μm, n = 30), acrogenous, dry, obclavate, rostrate, mostly curved, 15–36(–41)-distoseptate, truncate at the base, rounded at the apex, smooth, thick-walled, brown with a green tinge.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the two sides of the conidia. Colonies on PDA reaching 20 mm diameter after 20 days at 20–25 °C, circular, raised, velvety, aerial, medium–sparse, dark-brown from both above and below.
Material examined: China, Yunnan Province, Xishuangbanna, Man Feilong Reservoir, found on dead, submerged, decaying wood of unidentified plants, 7 November 2020, Rong Zhu, N66 (HKAS 122181, holotype), ex-type living culture KUNCC 12–10726.
Notes: Multi-gene phylogenetic analyses showed that D. crassispora is a distinct species in Distoseptispora and clusters with D. chinense and D. tectonigena (97% MLBS/1.00 BIPP; Figure 1). Distoseptispora crassispora is morphologically similar to the latter two species in having straight to slightly curved, septate conidiophores and obclavate, distoseptate, rostrate, straight or slightly curved conidia. However, D. crassispora possesses shorter conidiophores (up to 27 μm vs. up to 110 μm) and wider conidia (13–24 μm vs. 10–13 μm) than D. tectonigena. Additionally, the conidia of D. tectonigena are sometimes percurrently proliferating 5–10 times at the apex, which was not observed in D. crassispora [15]. Distoseptispora crassispora can be distinguished from D. chinense on the basis of molecular data. Comparisons of nucleotides between D. crassispora and D. chinense revealed 12 (2.3%, including four gaps) and 23 (2.6%) nucleotide differences in the ITS region and TEF1-α, respectively, which follows the generally accepted norm, according to which nucleotide differences of more than 1.5% in the ITS region are likely to indicate a new species [44]. We therefore introduce D. crassispora as a new species in Distoseptispora.
Distoseptispora curvularia R. Zhu & H. Zhang, sp. nov. (Figure 8).
Index Fungorum number: 559921; Facesoffungi number: 12578.
Holotype: HKAS 122180.
Etymology: In reference to the mostly curved conidia.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, scattered, hairy, dark-brown. Mycelium mostly immersed, composed of branched, septate, smooth, dark-brown hyphae. Conidiophores 11–28 μm long ( = 18 µm, n = 10), 5–9 μm wide ( = 7 µm, n = 10), macronematous, mononematous, unbranched, septate, cylindrical, straight or flexuous, smooth, brown to dark-brown, robust at the base. Conidiogenous cells 5–6 μm long, 5–5.5 μm wide, monoblastic, integrated, determinate, terminal, cylindrical, dark-brown, smooth. Conidia (60–)100–200(–314) µm long (rostrum included) ( = 155 μm, n = 30), 12–19 μm at the widest part ( = 15 μm, n = 30), 7–9.5 μm wide at the apex ( = 9 μm, n = 30), acrogenous, dry, obclavate, rarely oblong, rostrate, mostly curved, (9–)16–48(–59)-distoseptate, truncate at the base, rounded at the apex, smooth, thick-walled, brown with a green tinge.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the conidial base. Colonies on PDA reaching 30 mm in diameter after 20 days at 20–25 °C, circular, raised, convex or dome-shaped with dark-brown papillate surface, brown at the margins, dark-brown in reverse.
Material examined: China, Yunnan Province, Xishuangbanna, Man Feilong Reservoir, found on dead, submerged, decaying wood of unidentified plants, 7 November 2020, Rong Zhu, N53 (HKAS 122180, holotype), ex-type living culture KUNCC 21–10725.
Notes: The new species has a close phylogenetic affinity to D. clematidis (98% MLBS/1.00 BIPP; Figure 1). Morphologically, D. curvularia is similar to D. clematidis in having acrogenous, obclavate, rostrate conidia and being brown with a green tinge. However, D. curvularia differs from the holotype of D. clematidis (MFLU 17–1501) in having longer conidia (up to 314 μm vs. up to 210 μm) and more conidial septa (up to 59 vs. up to 35) [15]. Additionally, our new collection of D. clematidis (KUNCC 21–10725) possesses conidia sometimes with percurrent proliferation and the formation of another conidium from conidial apices. A comparison of sequence data for these two species showed a difference of 15 (2.81%, no gaps) noticeable nucleotides in ITS gene regions. Therefore, we introduce D. curvularia as a new species.
Distoseptispora dehongensis W. Dong, H. Zhang & K.D. Hyde, Fungal Diversity 96, 145 (2019) (Figure 9 and Figure 10).
Specimen MFLU 19–0544: Conidiophores 70–110 μm long, 3–4.5 μm wide, 5–7-septate, pale-brown to mid-brown. Conidiogenous cells (12–)18–23 μm long, 3–4.5 μm wide, monoblastic or polyblastic, pale-brown. Conidia 20–57 μm long ( = 31 µm, n = 30), 6–15 μm at the widest part ( = 10 µm, n = 30), 5.5–7 μm wide at the apex ( = 6 μm, n = 20), acrogenous or intercalary, solitary or catenate, obpyriform to obclavate, broad cylindrical or irregular, non-rostrate, straight or curved, 4–7-distoseptate, mostly 6-distoseptate, truncate at the base, rounded at the apex, smooth, thick-walled, pale-brown, sometimes with percurrent proliferation and forming another conidium from the conidial apex.
Specimen MFLU 17–1671: Conidiophores 47–128 μm long, 3–5.5 μm wide, 4–7-septate, pale-olivaceous. Conidiogenous cells 18–25 μm long, 4–5 μm wide, monoblastic or polyblastic, pale-olivaceous. Conidia 17–80 μm long ( = 51 µm, n = 30), 8–13 μm at the widest part ( = 13 μm, n = 30), 4–6 μm wide at the apex ( = 5 μm, n = 30), acrogenous or intercalary, solitary, obpyriform, non-rostrate, straight or curved, 4–10-distoseptate, mostly 6–8-distoseptate, truncate at the base, smooth, thick-walled, olivaceous, sometimes with percurrent proliferation and forming another conidium from the conidial apex.
Material examined: Thailand, Mukdahan Province, small river of Nong Bo Na Kae, found on dead, submerged, decaying wood of unidentified plants, 13 December 2018, Hao Yang, t34 (MFLU 19–0544), ex-type living culture MFLUCC 19–0335; Thailand, Long Khot subdistrict, found on submerged wood in a stream, 1 September 2017, Gennuo Wang, 4.7 (MFLU 17–1671), living culture MFLUCC 17–2326.
Notes: Our new isolates, MFLUCC 19–0335 (from MFLU 19–0544) and MFLUCC 17–2326 (from MFLU 17–1671), cluster with ex-type strain KUMCC 18–0090 of D. dehongensis, with strong support (100% MLBS/1.00 BIPP; Figure 1). Morphologically, the two new isolates fit well with the characters of D. dehongensis in having monoblastic or polyblastic conidiogenous cells and obpyriform to obclavate, straight or curved conidia with fewer than ten distosepta [45], except that the conidia of MFLU 19–0544 and MFLU 17–1671 show some percurrent proliferation, which is not observed in the holotype [45]. Additionally, the conidia of MFLU 19–0544 are pale-brown, while those of the holotype and MFLU 17–1671 are olivaceous. Comparisons of sequence data between the three strains showed no differences in LSU genes and three to six nucleotide differences in ITS genes. The holotype was collected from Yunan Provice, China [45], while our isolates were collected from Thailand, their locations constituting new geographic records. We provide new RPB2 sequence data for D. dehongensis in this study.
Distoseptispora leonensis (M.B. Ellis) R. Zhu & H. Zhang, comb. nov. (Figure 11).
Basionym: Sporidesmium leonense M.B. Ellis, Mycol. Pap. 70: 28 (1958).
Synonym: Ellisembia leonensis (M.B. Ellis) McKenzie, Mycotaxon 56: 13 (1995).
Index Fungorum number: 559922; Facesoffungi number: 12579.
Notes: The case of Distoseptispora leonensis is the same as that of D. adscendens, that is, it is the only strain of D. leonensis (HKUCC 10822) reported by Shenoy et al. [42], with LSU and RPB2 sequence data, which has been proven to phylogenetically cluster in the genus Distoseptispora [7] and has been named as D. adscendens in the phylogenetic tree, though it has not been validly introduced as a new combination. The morphology of strain HKUCC 10822 was also referred to Wu and Zhuang [14] (P137), who described it as having macronematous, mononematous, cylindrical, straight or slightly flexuous, smooth, 5–7-septate, mid-brown to brown conidiophores 110–130 μm long, 6–8 μm wide; integrated, terminal, cylindrical, smooth, pale-brown to brown conidiogenous cells 5–15 × 4.5–5 μm in size; and acrogenous, obclavate, fusiform or ellipsoidal, rostrate, 8–10-distoseptate conidia, 50–75 μm long, 15–18 μm wide at the broadest part, with conical–truncate basal cells 3–4.5 μm wide at the base. This description is consistent with the description and illustration of the holotype of S. leonensis given in Ellis [11], even regarding the sizes of conidiophores and conidia, though the conidiophore and conidial proliferations are points of difference. The above characters fit the generic features of Distoseptispora well.
Index Fungorum number: 559923; Facesoffungi number: 12580.
Holotype: HKAS 122185.
Etymology: In reference to the mostly non-rostrate conidia.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, hairy or velvety, dark-brown. Mycelium mostly immersed, consisting of branched, septate, smooth, subhyaline to pale-brown hyphae. Conidiophores 105–160 μm long ( = 129 µm, n = 10), 4.5–7 μm wide ( = 5.5 µm, n = 10), macronematous, mononematous, solitary, unbranched, 5–10-septate, cylindrical, straight or slightly flexuous, smooth, brown, becoming pale-brown towards apex. Conidiogenous cells 15–20 μm long ( = 18 µm, n = 10), 5–5.5 μm wide ( = 5 µm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, pale-brown, smooth. Conidia 22–51 μm long ( = 36 μm, n = 20), 8–14 μm at the widest part ( = 12 μm, n = 20), 5.5–10 μm wide at the apex ( = 8 μm, n = 20), acrogenous, solitary, oblong, obclavate or narrowly obpyriform, mostly non-rostrate, rarely rostrate, straight or curved, 4–10-distoseptate, truncate at the base, smooth, thick-walled, pale-olivaceous or pale-brown.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from conidial bases. Colonies on PDA reaching 30–35 mm diameter after 5 weeks at 20–25 °C, circular, raised, fluffy, dense, convex or dome-shaped, with dark-brown papillate surface, brown in the center, lighter on the outside, dark-brown at the margins, consistently dark-brown in reverse.
Material examined: China, Sichuan Province, Yibin City, Southern Sichuan Bamboo Sea, Qicai Lake, found on dead, submerged, decaying bamboo, 13 November 2020, Yun Qing, SN–16 (HKAS 122185, holotype), ex-type living culture KUNCC 21–10730.
Notes: In our phylogenetic analyses, Distoseptispora nonrostrata cluster with D. effusa. Morphologically, D. nonrostrata possesses oblong, obclavate or narrowly obpyriform, mostly non-rostrate, pale-olivaceous or pale-brown conidia, while D. effusa is clearly different in having obclavate, rostrate, olivaceous-brown to dark-brown, longer conidia (35.5–113 μm vs. 22–51 μm) [24]. Comparisons of sequence data between these two species showed differences of 27 (4.9%, four gaps) and 34 (3.8%, no gaps) noticeable nucleotides in ITS and TEF gene regions, respectively. Therefore, we introduce D. nonrostrata as a new species.
Distoseptispora pachyconidia R. Zhu & H. Zhang, sp. nov. (Figure 13).
Index Fungorum number: 559924; Facesoffungi number: 12581.
Holotype: HKAS 122179.
Etymology: “Pachy-” means thick in Latin, referring to the thick-walled conidia.
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, hairy or velvety, dark-brown or black. Mycelium mostly immersed, consisting of branched, septate, smooth, hyaline to pale-brown hyphae. Conidiophores 11–27 μm long ( = 19 µm, n = 10), 4–9 μm wide ( = 6 µm, n = 10), macronematous, mononematous, solitary, unbranched, 2–4-septate, cylindrical, straight or flexuous, smooth, pale-brown to mid-brown, slightly tapering distally, truncate at the apex. Conidiogenous cells 5–9 μm long ( = 7 µm, n = 10), 4–6.5 μm wide ( = 5 µm, n = 10), monoblastic, integrated, determinate, terminal, cylindrical, pale-brown, smooth. Conidia 42–136 μm long ( = 84 μm, n = 30), 14–22 μm at the widest part ( = 17 μm, n = 30), 8–14 μm wide at the apex ( = 11 μm, n = 30), acrogenous, solitary, obclavate, lanceolate, rostrate or not, straight or curved, 8–21-distoseptate, truncate at the base, tapering towards the rounded apex, smooth, thick-walled, pale-brown with a green tinge.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from both ends. Colonies on PDA reaching 35–38 mm diameter after 5 weeks at 20–25 °C, dark-brown at the margins, greenish-glaucous at the center, olivaceous-brown in reverse, with smooth margin.
Material examined: China, Yunnan Province, Xishuangbanna, Na Dameng Reservoir, found on dead, submerged, decaying wood of unidentified plants, 7 November 2020, Rong Zhu, N37 (HKAS 122179, holotype), ex-type living culture KUNCC 21–10724.
Notes: Distoseptispora pachyconidia clusters as an independent branch in Distoseptispora based on a concatenated LSU–ITS–TEF1-α–RPB2 phylogeny. It is phylogenetically close to D. crassispora, D. chinense and D. tectonigena. Compared to D. tectonigena, D. pachyconidia has shorter conidiophores (11–27 μm vs. up to 110 μm) and shorter conidia (42–136 μm vs. 148–225(–360) μm), as well as fewer conidial septa (8–21 vs. 20–46) [3]. In addition, the conidia of D. tectonigena were sometimes percurrently proliferating 5–10 times at the apex, which was not observed in D. pachyconidia. The conidia in D. pachyconidia are light-colored and shorter than those in D. crassispora and D. chinense (42–136 μm vs. 95–197 μm vs. 81–283) [46]. More importantly, D. pachyconidia is phylogenetically distinct from D. crassispora, D. chinense and D. tectonigena.
Distoseptispora rayongensis J. Yang & K.D. Hyde, Mycosphere 11(1): 579 (2020) (Figure 14).
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, hairy or velvety, black. Mycelium mostly immersed, consisting of branched, septate, smooth, brown hyphae. Conidiophores 62–76 μm long ( = 69 µm, n = 10), 4–6 μm wide ( = 5.5 µm, n = 10), macronematous, mononematous, solitary, unbranched, 2–4-septate, cylindrical, straight or slightly flexuous, smooth, brown, rounded at apex. Conidiogenous cells 17–23 μm long, 5–6 μm wide, monoblastic, integrated, determinate, terminal, cylindrical, brown, smooth. Conidia 90–180(–261) μm long (rostrum included) ( = 134 µm, n = 20), 10–15 µm at the widest part ( = 13 µm, n = 20), 2–5 µm wide at the apex ( = 4 µm, n = 20), acrogenous, obclavate or obspathulate, rostrate, straight or slight curved, 12–17-distoseptate, truncate at the base, rounded at the apex, guttulate, smooth, thick-walled, pale-brown or pale-olivaceous, becoming paler or hyaline towards the apex.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the conidial apices. Colonies on PDA fast-growing, reaching 16 mm diameter after 15 days at 25 °C, circular, aerial mycelium dense, brown, reverse dark-brown, with entire, white margin.
Material examined: Thailand, Mukdahan Province, small river of Nong Bo Na Kae, found on dead, submerged, decaying wood of unidentified plants, 13 December 2018, Hao Yang, t33 (MFLU 19–0543), living culture MFLUCC 19–0334.
Notes: Distoseptispora rayongensis was introduced by Hyde et al. [47] and collected from a freshwater habitat in Rayong Province, Thailand. In our phylogenetic analysis, our new isolate was grouped with D. rayongensis, with strong support (99% MLBS/1.00 BIPP; Figure 1). Morphologically, our isolate was the same as D. rayongensi, except that it had shorter conidiophores (62–76 μm vs. 75–125 µm). Additionally, in the holotype, conidia showed percurrent proliferation and conidia forming from the conidial apices, which was not observed in our isolate. Comparisons of nucleotides between the holotype and our isolate showed no differences in ITS and TEF1-α regions and 1 and 35 (3.5%, including 21 gaps) nucleotide differences in LSU and RPB2 regions, respectively.
Distoseptispora tectonae Doilom & K.D. Hyde, Fungal Diversity 80: 222 (2016) (Figure 15).
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on the substratum superficial, effuse, scattered, hairy, black. Mycelium partly superficial, partly immersed, composed of branched, septate, smooth, pale-brown hyphae. Conidiophores 9–35 μm long, 5–7 μm wide, macronematous, mononematous, unbranched, 1–3-septate, cylindrical, straight or slightly flexuous, smooth, brown to dark-brown, robust at the base. Conidiogenous cells 5–10 μm long, 4.5–5.5 μm wide, holoblastic, monoblastic, integrated, terminal, cylindrical, brown, smooth. Conidia 46–192 μm long ( = 111 µm, n = 30) (rostrum included), 12–18 µm at the widest part ( = 15 µm, n = 30), 8–12 µm wide at the apex ( = 10 µm, n = 30), acrogenous, solitary, cylindric–obclavate, elongate, rostrate, straight or curved, 9–39-distoseptate, rounded at the apex, truncate at the base, smooth, thick-walled, olivaceous-green when young, dark-brown at maturity, with a wedge-shaped base, 3.5–6 μm wide.
Culture characteristics: Conidia germinating on PDA within 24 h. Germ tubes produced from the conidial apices. Colonies on PDA reaching 35 mm diameter after 36 days at 20–25 °C, circular, dry, dark-olivaceous-green to black on surface and reverse, raised, with entire margin.
Material examined: China, Sichuan Province, Yibin City, Southern Sichuan Bamboo Sea, Qicai Lake, found on dead, submerged, decaying wood of unidentified plants, 16 June 2019, Chunxue Li, S1–4 (HKAS 122183), living culture KUNCC 21–10728.
Notes: Distoseptispora tectonae was initially collected from a dead twig of Tectona grandis [3]. Subsequently, an isolate which was collected from submerged water and identified as D. submerse was synonymized as D. tectonae [9]. More recently, two new isolates collected from saprobic and submerged water in Thailand were reported [9,22]. Our new isolate clustered with the above strains, with strong support (100% MLBS/1.00 BIPP; Figure 1). It has a wider range of conidial lengths than the holotype (46–192 μm vs. (90–)130–140(–170) μm) [3]. The conidia of our isolate and MFLU 15–2693 are olivaceous, while those of the holotype (MFLU 15–3417) and MFLU 20–0262 are brown.
4. Discussion
To date, a total of 54 Distoseptispora species have been reported. They have been found to cluster into three well-supported clades, which is in agreement with all previous studies on Distoseptispora [5,8,9,23,24,25]. The morphologies of Distoseptispora species are similar, and taxonomic identifications mainly rely on phylogeny. To reappraising generic and specific delimitations, we summarized the characters of all Distoseptispora species in Figure 1, including the length of conidiophores; proliferation and conidiogenesis (monoblastic or polyblastic) in conidiogenous cells; the type (distoseptate or euseptate) and number of septa, shape, length, color, proliferation, rostrateness and wall thickness of conidia; as well as habitats. With respect to the characters listed in Figure 1, we can summarize that:
(1) Proliferation and conidiogenesis (monoblastic or polyblastic) in conidiogenous cells
Proliferation of conidiogenous cells has been reported in all clades: D. amniculi, D. guttulata, D. effusa and D. rostrata in Clade 1; D. atroviridis and D. fusiformis in Clade 2; and D. verrucosa and D. yunnanensis in Clade 3. Although no proliferation has been reported in D. palmarum and D. yunnanensis, we can see obvious proliferation in the illustrations of Hyde et al. [45] (Figure 104d) and Li et al. [23] (Figure 3b,h). All species possess monoblastic conidiogenous cells, except for D. palmarum, which possesses polyblastic ones, and D. dehongensis in Clade 1, D. saprophytica in Clade 2, and D. bambusae and D. meilingensis in Clade 3, which possess mono- or polyblastic ones.
(2) Type of conidial septa—distoseptate or euseptate
Regarding the definitions of “distoseptate” and “euseptate”, in Dictionary of the Fungi (9th edition), “euseptate (of conidial septation)” is defined as “having cells separated by multilayered walls of similar structure to lateral walls”, while “distoseptate (of conidial septation)” is defined as “having the individual cells each surrounded by a sac-like wall distinct from the outer wall”. Given these definitions and actual observations in Distoseptispora species, we can conclude that the euseptate condition appears as a single line or thicker region in the middle of septa, while the distoseptate condition is characterized by sac-like septa, usually narrower in the middle of septa or exhibiting cracks, as in D. clematidis, D. crassispora and D. nonrostrata (indicated by the arrows in Figure 6f, Figure 7d and Figure 12g in this paper).
According to Figure 1, most species in Clades 1 and 2 have distoseptate conidia, just like the generic type D. fluminicola. The species with euseptate conidia are all grouped in Clade 3, except for D. guttulata and D. martinii. Distoseptispora martinii was first described as Acrodictys martinii, then was transferred to Junewangia and Rhexoacrodictys [48,49,50]. Recently, it was proved to be a new combination in Distoseptispora on the basis of phylogenetic analysis [16]. Morphologically, it has ellipsoid, oblate to subglobose, muriform conidia instead of obclavate to obpyriform, transversely distoseptate conidia. Distoseptispora uttulate is another exception in Distoseptispora, as it has 11–14-euseptate conidia. Distoseptispora lignicola was described as euseptate, but it can be seen from the illustrations that it is euseptate only at the rostra. At the main part of the conidia, it should be distoseptate, due to the obvious narrowing in the middle of the septa [8] (Figure 5g,h). Distoseptispora rayongensis has the same characteristics, such that its septa should be recognized as distoseptate ([47], Figure 99k; this study, Figure 10g). Distoseptispora meilingensis is the only species in Clade 3 that has distoseptate conidia.
(3) Type of conidial wall—thick or thin, smooth or verrucose
All species in Clades 1 and 2 have thick-walled conidia, except for D. guttulata and D. martini, which have thin-walled conidia. The species with euseptate and thin-walled conidia are all grouped in Clade 3, except for D. meilingensis, which has thick-walled conidia. Distoseptispora bambusae and D. euseptata were described as thick-walled, with wall thicknesses less than 1 µm—thicker than those in D. Suoluoensis and D. Lancangjiangensis but not as thick as the ones in Clades 1 and 2 (ca. 2–4 µm). Thus, rather than thick-walled, it is more proper to call them medium–thick-walled. All Distoseptispora species have smooth conidial walls, except for D. aquamyces, D. suoluoensis and D. verrucose, which are verrucose-walled.
(4) Conidial shapes and color
Conidial shapes in Clades 1 and 2 are mostly obclavate, sometimes obpyriform, oblong, subcylindrical/cylindrical, rarely fusiform or ellipsoid/oblate/subglobose, while those in Clade 3 are obclavate. D. euseptata also has obpyriform conidia. Conidial color in most species is brown with a green tinge, and a few have a single color. For example, in Clade 1, conidia are brown in D. tectonigena and D. longispora, and olivaceous green in D. multiseptata. Conidial color may vary within a species. For example, in D. tectonae, conidia are brown in the holotype (MFLU 15–3417) and specimen MFLU 20–0262 and olivaceous in specimen MFLU 15–2693. Another example is D. dehongensis: conidial color is pale brown in specimen MFLU 19–0544 and olivaceous in the holotype and specimen MFLU 17–1671.
(5) The length of conidiophores and conidia, and numbers of septa
The lengths of conidiophores in the three clades are irregular, including 0 (maximum < 100 µm), 1 (minimum < 100 µm and maximum > 100 µm) and 2 (minimum > 100 µm). The conidial lengths of species in Clade 1 are relatively long, mostly longer than 100 µm, even up to 700 µm. Therefore, they have more conidial septa—many more than 20 and even up to 80. The conidial lengths of species in Clade 3 are relatively short, all mostly shorter than 100 µm. Conidial length may vary within a species, as in D. multiseptata. The conidial lengths of collection MFLU 15–1144 (mostly 300–600 µm long, up to 700 µm long) are significantly longer than those of the holotype (up to 380 µm long) [5]. Yang et al. [5] suspected that the length of conidia may depend on the length of incubation.
(6) Conidial rostrateness
Conidia are described as rostrate when they have a beak-like extension or appearance, according to Dictionary of the Fungi (9th edition), where “beak” means a pointed part on the tip (https://dictionary.cambridge.org, accessed on 8 May 2022). Given this definition, we re-checked all species from the published illustrations and listed those with rostrate conidia in Figure 1. Species with rostrate conidia that were not reported as such in their original descriptions are labeled in red. From Figure 1, we can see that most species possess rostrate conidia. Species with only non-rostrate conidia are D. atroviridis, D. dehongensis, D. fusiformis, D. hydei, D. martinii and D. saprophytica.
(7) Conidial proliferation
Conidial proliferation has been described in Clade 1 (D. amniculi, D. bangkokensis, D. tectonigena, D. rayongensis, D. yunjushanensis) and Clade 3 (D. suoluoensis). Distoseptispora saprophytica was thought to produce catenate conidia occasionally [9]. However, it is more proper to use the term conidial proliferation, because the upper conidia were obviously immature [9] (Figure 14g). Although no conidial proliferation was reported in D. xishuangbannaensis and D. thailandica, we can see obvious proliferation in their illustrations ([18], Figure 65c,d and Figure 64c). Conidial proliferation is not a species-delimiting factor because an individual species may exhibit variation in this character, as in D. clematidis, D. dehongensis and D. multiseptata. These species were not reported as being characterized by conidial proliferation with respect to their holotype, though proliferation was reported in the new collections ([18], Figure 3; this study, Figure 3 and Figure 7).
(8) Habitats
Most Distoseptispora species were reported from freshwater habitats, except for D. adscendens, D. hydei, D. martinii, D. tectonigena, D. thailandica and D. xishuangbannaensis in Clade 1, and D. caricis, D. leonensis and D. palmarum in Clade 2.
To sum up, no significant morphological differences were found among Clades 1, 2 and 3. Therefore, there is not enough evidence to segregate Clades 2 and 3 from Clade 1, in which the type species D. fluminicola is included, although Clades 1, 2 and 3 all received strong support (100% MLBS/1.00 BIPP; Figure 1). Conidial differentiation on the basis of euseptate and distoseptate does not receive molecular support, as such differences are found in both Clades 1 and 3. This is the same as the situation for euseptate/distoseptate conidia in Sporidesmium. Subramanian [13] segregated the species with distoseptate conidia from Sporidesmium and introduced the genus Ellisembia to accommodate them. However, Su et al. [7] proved that the euseptate/distoseptate difference between Sporidesmium and Ellisembia does not have molecular support and proposed Ellisembia as a synonym of Sporidesmium. Yang et al. [24] also inferred that euseptate/distoseptate differences have no taxonomic significance for generic delimitation of sporidesmium-like taxa, though they are informative at the species level.
Author Contributions
Conceptualization, D.Z. and P.N.; methodology, H.Z. and R.Z.; formal analysis and investigation, H.Z. and R.Z.; resources, Y.Q., H.Y., C.L. and G.W.; writing—original draft preparation, H.Z. and R.Z.; writing—review and editing, H.Z. and D.Z.; funding acquisition, P.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was mainly supported by the National Natural Science Foundation of China (Project ID: NSF 31500017 to Huang Zhang).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated for this study can be found in the NCBI database.
Acknowledgments
This work was mainly supported by the National Natural Science Foundation of China (Project ID: NSF 31500017 to Huang Zhang) and the Graduate Research and International Exchange Program of Kunming University of Science and Technology 2018. Hao Yang thanks Mae Fah Luang University (Chiang Rai, Thailand) for experimental support. Wei Dong is thanked for species identification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Hyde, K.; Goh, T. Fungi on submerged wood in Lake Barrine, north Queensland, Australia. Mycol. Res. 1998, 102, 739–749. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, W.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Hongsanan, S.; Jayarama Bhat, D.; Al-Sadi, A.M.; Zhang, D. Towards a natural classification of Annulatascaceae-like taxa: Introducing Atractosporales ord. nov. and six new families. Fungal Divers. 2017, 85, 75–110. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, K.Q.; McKenzie, E.H.C.; Hyde, K.D. Freshwater fungi from bamboo and wood submerged in the Liput River in the Philippines. Fungal Divers. 2003, 13, 1–12. [Google Scholar]
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.J.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Hyde, K.D.; Jeewon, R.; Doilom, M.; Yu, X.-D.; Wang, G.-N.; Liu, N.G.; Hu, D.M.; Nalumpang, S.; Zhang, H. Towards a natural classification of annulatascaceae-like taxa II: Introducing five new genera and eighteen new species from freshwater. Mycosphere 2021, 12, 1–88. [Google Scholar] [CrossRef]
- Link, H.F. Observationes in ordines plantarumnaturales. Diss. I. Mag. Ges. Naturf. Freunde Berl. 1809, 3, 3–42. [Google Scholar]
- Ellis, M.B. Clasterosporium and some allied Dematiaceae—Phragmosporae, I. Mycol. Pap. 1958, 70, 1–89. [Google Scholar]
- Ellis, M.B. (Ed.) Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: London, UK, 1971. [Google Scholar]
- Subramanian, C.V. A reassessment of Sporidesmium (Hyphomycetes) and some related taxa. Proc. Indian Natl. Sci. Acad. USA 1992, 58, 179–190. [Google Scholar]
- Wu, W.; Zhuang, W. Sporidesmium, Endophragmiella and Related Genera from China; Fungal Diversity Press: Hong Kong, China, 2005. [Google Scholar]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep.-UK 2017, 7, 7888. [Google Scholar] [CrossRef]
- Luo, Z.; Hyde, K.; Liu, J.; Bhat, D.; Bao, D.; Li, W.; Su, H. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsaard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Marc, S.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Sun, Y.; Goonasekara, I.D.; Thambugala, K.M.; Jayawardena, R.S.; Wang, Y.; Hyde, K.D. Distoseptispora bambusae sp. nov.(Distoseptisporaceae) on bamboo from China and Thailand. Biodivers. Data J. 2020, 8, e53678. [Google Scholar] [CrossRef]
- Li, W.-L.; Liu, Z.-P.; Zhang, T.; Dissanayake, A.J.; Luo, Z.-L.; Su, H.-Y.; Liu, J.-K. Additions to Distoseptispora (Distoseptisporaceae) associated with submerged decaying wood in China. Phytotaxa 2021, 520, 75–86. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.-L.; Jones, E.B.G.; Li, W.-L.; Hyde, K.D.; Liu, Z.-Y. Morphological variety in Distoseptispora and introduction of six novel species. J. Fungi 2021, 7, 945. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols a Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 19, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; San Diego Supercomputer Center: La Jolla, CA, USA, 2010; pp. 1–8. [Google Scholar]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef]
- Nylander, J.A.A. (Ed.) MrModeltest V2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1. 4.2, A Graphical Viewer of Phylogenetic Trees. Available from Left Angle Bracket. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/rightanglebracket (accessed on 5 May 2022).
- Shoemaker, R.A. Lasiosphaeria caesariata with Sporidesmium hormiscioides and L. triseptata with S. adscendens. Sydowia 1985, 38, 278–283. [Google Scholar]
- Réblová, M. Studies in Chaetosphaeria sensu lato III. Umbrinosphaeria gen. nov. and Miyoshiella with Sporidesmium anamorphs. Mycotaxon 1999, 71, 13–43. [Google Scholar]
- Shenoy, B.D.; Jeewon, R.; Wu, W.P.; Bhat, D.J.; Hyde, K.D. Ribosomal and RPB2 DNA sequence analyses suggest that Sporidesmium and morphologically similar genera are polyphyletic. Mycol. Res. 2006, 110, 916–928. [Google Scholar] [CrossRef]
- Zhai, Z.-J.; Yan, J.-Q.; Li, W.-W.; Gao, Y.; Hu, H.-J.; Zhou, J.-P.; Song, H.-Y.; Hu, D.-M. Three novel species of Distoseptispora (Distoseptisporaceae) isolated from bamboo in Jiangxi Province, China. Mycokeys 2021, 88, 35–54. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Hyde, K.D.; Suwannarach, N.; Jayawardena, R.S.; Manawasinghe, I.S.; Liao, C.F.; Doilom, M.; Cai, L.; Zhao, P.; Buyck, B.; Phukhamsakda, C.; et al. Mycosphere notes 325–344—Novel species and records of fungal taxa from around the world. Mycosphere 2021, 12, 1101–1156. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Ba, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Crane, J.L.; Dumont, K.P. Hyphomycetes from the West Indies and Venezuela. Can. J. Bot. 1975, 53, 843–851. [Google Scholar] [CrossRef]
- Baker, W.A.; Partridge, C.; Morganjones, G. Notes on hypomycetes. LXXXVII. Rhexoacrodictys, a new segregate genus to accomodate four species previously classified n acrodictys. Mycotaxon 2002, 82, 95–113. [Google Scholar]
- Delgado, G. South Florida microfungi: Veramycella bispora, a new palmicolous, anamorphic genus and species, with some new records for the continental USA. Mycotaxon 2009, 107, 357–373. [Google Scholar] [CrossRef]
Figure 1.
The best-scoring maximum likelihood (RAxML) tree for the family Distoseptisporaceae, based on the LSU, ITS, TEF1-α and RPB2 sequence dataset. The tree is rooted with Myrmecridium schulzeri (CBS 100.54) and Myrmecridium banksiae (CPC 19852). Maximum likelihood support values greater than 70% and Bayesian posterior probabilities greater than 0.95 are shown near the nodes. New strains identified in this study are shown in blue; new species are shown in red. Ex-type strains are shown in bold. The characters of all Distoseptispora species are summarized, including conidiophores (size: S (maximum length < 100 µm), L (minimum length > 100 µm), M (minimum length < 100 µm and maximum length > 100 µm)); conidiogenous cells (proliferation: Y (yes), N (no); type: mono (monophyletic), poly (polyphyletic)); and conidia (septa: D (distospetate), E (euseptate); wall: TK (thick), TN (thin), MT (medium thick); conidial shape: 0 (obpyriform), 1 (obclavate), 2 (oblong), 3 (subcylindrical/cylindrical), 4 (fusiform), 5 (ellipsoid, oblate or subglobose); color: B (brown), O (olivaceous or green); septa number; size: S, M and L (the same as for conidiophore size); rostrateness and proliferation: Y (yes), N (no); habitats: T (terrestrial), F (freshwater)).
Figure 1.
The best-scoring maximum likelihood (RAxML) tree for the family Distoseptisporaceae, based on the LSU, ITS, TEF1-α and RPB2 sequence dataset. The tree is rooted with Myrmecridium schulzeri (CBS 100.54) and Myrmecridium banksiae (CPC 19852). Maximum likelihood support values greater than 70% and Bayesian posterior probabilities greater than 0.95 are shown near the nodes. New strains identified in this study are shown in blue; new species are shown in red. Ex-type strains are shown in bold. The characters of all Distoseptispora species are summarized, including conidiophores (size: S (maximum length < 100 µm), L (minimum length > 100 µm), M (minimum length < 100 µm and maximum length > 100 µm)); conidiogenous cells (proliferation: Y (yes), N (no); type: mono (monophyletic), poly (polyphyletic)); and conidia (septa: D (distospetate), E (euseptate); wall: TK (thick), TN (thin), MT (medium thick); conidial shape: 0 (obpyriform), 1 (obclavate), 2 (oblong), 3 (subcylindrical/cylindrical), 4 (fusiform), 5 (ellipsoid, oblate or subglobose); color: B (brown), O (olivaceous or green); septa number; size: S, M and L (the same as for conidiophore size); rostrateness and proliferation: Y (yes), N (no); habitats: T (terrestrial), F (freshwater)).

Figure 2.
Distoseptispora adscendens. (redraw according to Wu and Zhuang [14]). (a) Conidiophores with conidia. (b) Conidia.
Figure 2.
Distoseptispora adscendens. (redraw according to Wu and Zhuang [14]). (a) Conidiophores with conidia. (b) Conidia.

Figure 3.
Distoseptispora aqualignicola (HKAS 122184, holotype). (a,b) Colonies on natural substrate. (c,d) Conidiophores with conidia. (e–g) Conidiogenous cells bearing conidia. (h−m) Conidia. (n) Germinated conidium. (o) Colony on PDA (from front). (p) Colony on PDA (from reverse). Scale bars: (c,d) 30 μm; (e,h–i) 10 μm; (f,g,j–n) 20 μm.
Figure 3.
Distoseptispora aqualignicola (HKAS 122184, holotype). (a,b) Colonies on natural substrate. (c,d) Conidiophores with conidia. (e–g) Conidiogenous cells bearing conidia. (h−m) Conidia. (n) Germinated conidium. (o) Colony on PDA (from front). (p) Colony on PDA (from reverse). Scale bars: (c,d) 30 μm; (e,h–i) 10 μm; (f,g,j–n) 20 μm.

Figure 4.
Distoseptispora aquamyces (HKAS 122186, holotype). (a,b) Colonies on natural substrate. (c,d) Conidiophores with conidia. (e,f) Conidiogenous cells bearing conidia. (g−l) Conidia. (m) Germinated conidium. (n) Colony on PDA (from front). (o) Colony on PDA (from reverse). Scale bars: (c,d) 40 μm; (e,f,j–l) 20 μm; (g–i,m) 10 μm.
Figure 4.
Distoseptispora aquamyces (HKAS 122186, holotype). (a,b) Colonies on natural substrate. (c,d) Conidiophores with conidia. (e,f) Conidiogenous cells bearing conidia. (g−l) Conidia. (m) Germinated conidium. (n) Colony on PDA (from front). (o) Colony on PDA (from reverse). Scale bars: (c,d) 40 μm; (e,f,j–l) 20 μm; (g–i,m) 10 μm.

Figure 5.
Distoseptispora bambusae. (a) Colonies on natural substrate. (b–d) Conidiophores with conidia. (e–k) Conidia. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b–d) 30 μm; (e–k) 20 μm.
Figure 5.
Distoseptispora bambusae. (a) Colonies on natural substrate. (b–d) Conidiophores with conidia. (e–k) Conidia. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b–d) 30 μm; (e–k) 20 μm.

Figure 6.
Distoseptispora clematidis. (a) Colonies on natural substrate. (b) Conidiophores with conidia. (c–h) Conidia. The arrow in f points to a gap in the middle of the septum, which indicates the distosepta. (i,j) Portions of conidia with percurrent proliferation in the middle of conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b,g–h) 50 μm; (c,d) 10 μm; (i,j) 5 μm.
Figure 6.
Distoseptispora clematidis. (a) Colonies on natural substrate. (b) Conidiophores with conidia. (c–h) Conidia. The arrow in f points to a gap in the middle of the septum, which indicates the distosepta. (i,j) Portions of conidia with percurrent proliferation in the middle of conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b,g–h) 50 μm; (c,d) 10 μm; (i,j) 5 μm.

Figure 7.
Distoseptispora crassispora (HKAS 122181, holotype). (a,b) Colonies on natural substrate. (c) Conidiophores with conidium. (d–j) Conidia. The arrow in d points to a gap in the middle of the septum, which indicates the distosepta. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c) 40 μm; (d–f) 20 μm; (g–j) 30 μm; (k) 2 μm.
Figure 7.
Distoseptispora crassispora (HKAS 122181, holotype). (a,b) Colonies on natural substrate. (c) Conidiophores with conidium. (d–j) Conidia. The arrow in d points to a gap in the middle of the septum, which indicates the distosepta. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c) 40 μm; (d–f) 20 μm; (g–j) 30 μm; (k) 2 μm.

Figure 8.
Distoseptispora curvularia (HKAS 122180, holotype). (a) Colonies on natural substrate. (b) Conidiophores with conidium. (c–k) Conidia. The arrow in i points to a gap in the middle of the septum, which indicates the distosepta. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b–g) 20 μm; (h–i) 30 μm; (j–k) 50 μm.
Figure 8.
Distoseptispora curvularia (HKAS 122180, holotype). (a) Colonies on natural substrate. (b) Conidiophores with conidium. (c–k) Conidia. The arrow in i points to a gap in the middle of the septum, which indicates the distosepta. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b–g) 20 μm; (h–i) 30 μm; (j–k) 50 μm.

Figure 9.
Distoseptispora dehongensis (from MFLU 19–0544). (a) Colonies on natural substrate. (b) Conidiophore. (c,d) Conidiophores with conidia. (e–g) Conidiogenous cells bearing conidia. Note the conidium with percurrent proliferation that is forming another conidium in g. (h–m) Conidia. Arrow in k points a gap in the middle of septum, which indicateds the distoseptate, note the percurrently proliferating conidium in m; (n) Germinated conidium. (o) Colony on PDA (from front). (p) Colony on PDA (from reverse). Scale bars: (b–c,n) 20 μm; (d,g) 30 μm; (e,f,h–m) 10 μm.
Figure 9.
Distoseptispora dehongensis (from MFLU 19–0544). (a) Colonies on natural substrate. (b) Conidiophore. (c,d) Conidiophores with conidia. (e–g) Conidiogenous cells bearing conidia. Note the conidium with percurrent proliferation that is forming another conidium in g. (h–m) Conidia. Arrow in k points a gap in the middle of septum, which indicateds the distoseptate, note the percurrently proliferating conidium in m; (n) Germinated conidium. (o) Colony on PDA (from front). (p) Colony on PDA (from reverse). Scale bars: (b–c,n) 20 μm; (d,g) 30 μm; (e,f,h–m) 10 μm.

Figure 10.
Distoseptispora dehongensis (from MFLU 17–1671). (a,b) Colonies on natural substrate. (c–f) Conidiophores with conidia. (g–l) Conidia. The arrows in g and h indicate conidial percurrent proliferating points. The arrow in l points to a gap in the middle of the septum, which indicates the distosepta. (m) Germinated conidium. (n) Colony on PDA (from front). (o) Colony on PDA (from reverse). Scale bars: (c–f) 20 μm; (g–l) 15 μm; (m) 10 μm.
Figure 10.
Distoseptispora dehongensis (from MFLU 17–1671). (a,b) Colonies on natural substrate. (c–f) Conidiophores with conidia. (g–l) Conidia. The arrows in g and h indicate conidial percurrent proliferating points. The arrow in l points to a gap in the middle of the septum, which indicates the distosepta. (m) Germinated conidium. (n) Colony on PDA (from front). (o) Colony on PDA (from reverse). Scale bars: (c–f) 20 μm; (g–l) 15 μm; (m) 10 μm.

Figure 11.
Distoseptispora leonensis (redraw according to Wu and Zhuang [14]). (a,b) Conidiophores with conidia.
Figure 11.
Distoseptispora leonensis (redraw according to Wu and Zhuang [14]). (a,b) Conidiophores with conidia.

Figure 12.
Distoseptispora nonrostrata (HKAS 122185, holotype). (a,b) Colonies on natural substrate. (c) Conidiophore with conidium. (d,e) Conidiogenous cells bearing conidia. (f–k) Conidia. The arrow in g points to a gap in the middle of the septum, which indicates the distosepta. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c) 30 μm; (d–k) 10 μm.
Figure 12.
Distoseptispora nonrostrata (HKAS 122185, holotype). (a,b) Colonies on natural substrate. (c) Conidiophore with conidium. (d,e) Conidiogenous cells bearing conidia. (f–k) Conidia. The arrow in g points to a gap in the middle of the septum, which indicates the distosepta. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c) 30 μm; (d–k) 10 μm.

Figure 13.
Distoseptispora pachyconidia (HKAS 122179, holotype). (a,b) Colonies on natural substrate. (c) Conidiophores with conidia. (d,e) Conidiogenous cells with conidia. (f–l) Conidia. The arrow in h points to a gap in the middle of the septum, which indicates the distosepta. (m) Colony on PDA (from front). (n) Colony on PDA (from reverse). Scale bars: (c) 40 μm; (d–l) 20 μm.
Figure 13.
Distoseptispora pachyconidia (HKAS 122179, holotype). (a,b) Colonies on natural substrate. (c) Conidiophores with conidia. (d,e) Conidiogenous cells with conidia. (f–l) Conidia. The arrow in h points to a gap in the middle of the septum, which indicates the distosepta. (m) Colony on PDA (from front). (n) Colony on PDA (from reverse). Scale bars: (c) 40 μm; (d–l) 20 μm.

Figure 14.
Distoseptispora rayongensis. (a) Colonies on natural substrate. (b) Conidiophore. (c) Conidiogenous cell with conidium. The arrow in c points to a gap in the middle of the septum, which indicates the distosepta. (d) Conidiophores with conidia. (e–j) Conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b) 20 μm; (c,e–k) 30 μm; (d) 40 μm.
Figure 14.
Distoseptispora rayongensis. (a) Colonies on natural substrate. (b) Conidiophore. (c) Conidiogenous cell with conidium. The arrow in c points to a gap in the middle of the septum, which indicates the distosepta. (d) Conidiophores with conidia. (e–j) Conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (b) 20 μm; (c,e–k) 30 μm; (d) 40 μm.

Figure 15.
Distoseptispora tectonae. (a,b) Colonies on natural substrate. (c,f,g) Conidiophores with conidia. The arrow in f points to a gap in the middle of the septum, which indicates the distosepta. (d,e,h–j) Conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c,e) 20 μm; (d) 10 μm; (f–h) 50 μm; (i,k) 30 μm; (j) 40 μm.
Figure 15.
Distoseptispora tectonae. (a,b) Colonies on natural substrate. (c,f,g) Conidiophores with conidia. The arrow in f points to a gap in the middle of the septum, which indicates the distosepta. (d,e,h–j) Conidia. (k) Germinated conidium. (l) Colony on PDA (from front). (m) Colony on PDA (from reverse). Scale bars: (c,e) 20 μm; (d) 10 μm; (f–h) 50 μm; (i,k) 30 μm; (j) 40 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
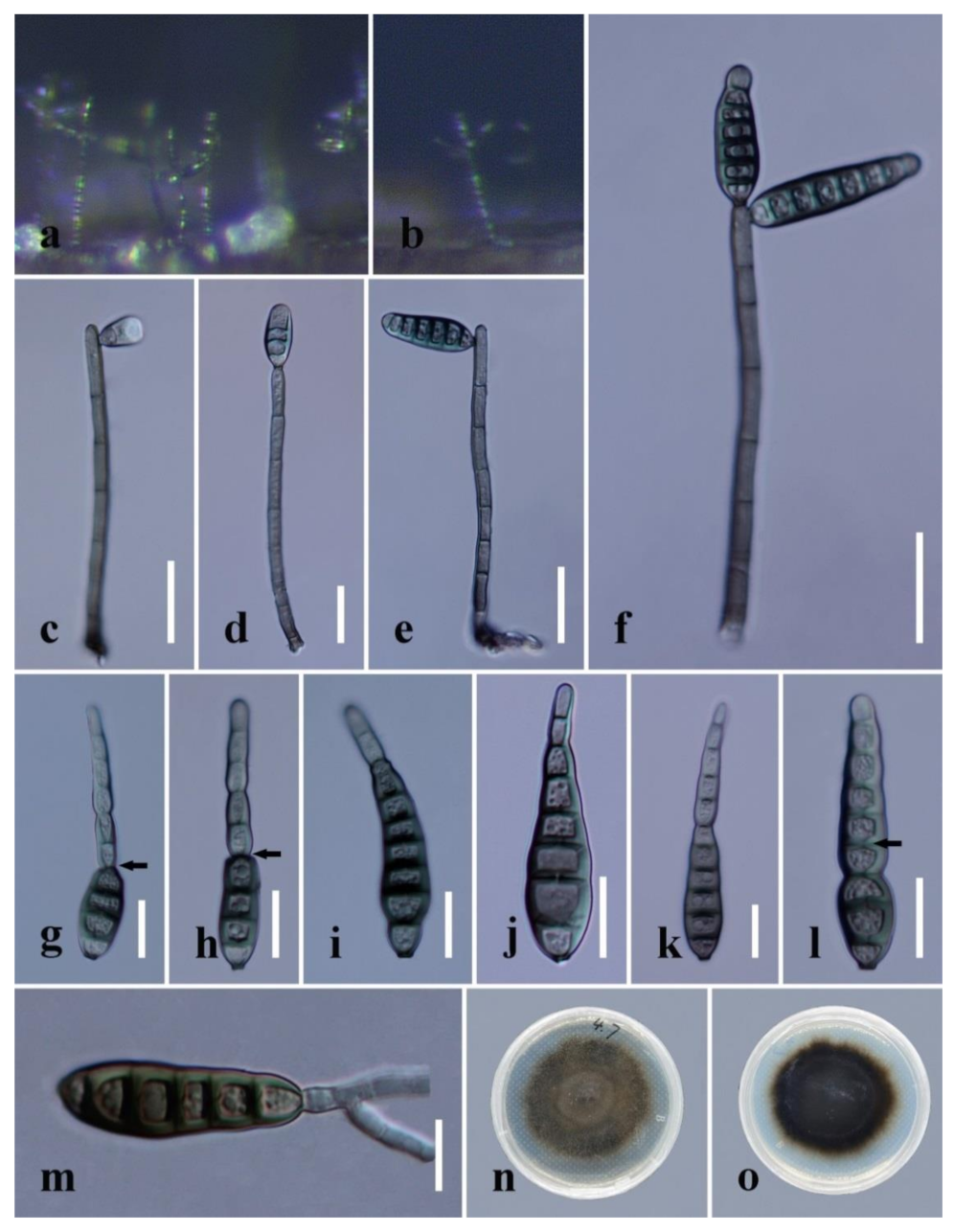{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The primers for the LSU, ITS, TEF1-α and RPB2 genes and the PCR thermal cycle programs used in this study.
Table 1.
The primers for the LSU, ITS, TEF1-α and RPB2 genes and the PCR thermal cycle programs used in this study.
| Gene | PCR Primers | PCR Amplification Reaction System | PCR Conditions |
|---|---|---|---|
| LSU, ITS | LR5/LROR [28], ITS5/ITS4 [29] | Total: 25 μL 12.5 μL Master Mix 9.5 μL ddH2O 1 μL of each primer (10 μM) 1 μL DNA template | 1. 94 °C, 3 min 2. 94 °C, 45 s 3. 56 °C, 50 s 4. 72 °C, 1 min 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 10 min 7. Keep at 4 °C |
| TEF1-α | 983F/2218R [30] | Total: 50 μL 45 μL T3 Mix 2 μL of each primer (10 μM) 1 μL DNA template | 1. 98 °C, 2 min 2. 98 °C, 10 s 3. 56 °C, 10 s 4. 72 °C, 20 s 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 5 min 7. Keep at 4 °C |
| RPB2 | 5F/7cR [31] | Total: 50 μL 45 μL T3 Mix 2 μL of each primer (10 μM) 1 μL DNA template | 1. 98 °C, 2 min 2. 98 °C, 10 s 3. 51 °C, 10 s 4. 72 °C, 20 s 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 5 min 7. Keep at 4 °C |
Table 2.
Taxa used in the phylogenetic analyses and their corresponding GenBank accession numbers.
| Species | Voucher/Culture | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| LSU | ITS | TEF1-α | RPB2 | ||
| Acrodictys bambusicola | CGMCC 3.18641 | KX033564 | KU999973 | – | – |
| Acrodictys elaeidicola | CGMCC 3.18642 | KX033569 | KU999978 | – | – |
| Aquapteridospora aquatica | MFLUCC 17–2371T | MW287767 | MW286493 | – | – |
| Aquapteridospora fusiformis | MFLU 18–1601T | MK849798 | MK828652 | MN194056 | – |
| Aquapteridospora lignicola | MFLUCC 15–0377T | KU221018 | MZ868774 | MZ892980 | MZ892986 |
| Bullimyces aurisporus | AF316–1bT | JF775590 | – | – | – |
| Bullimyces communis | AF281–5 | JF775587 | – | – | – |
| Cancellidium applanatum | CBS 337.76 | MH872755 | NR159777 | _ | _ |
| Cancellidium cinereum | MFLUCC 18–0424T | MT370363 | MT370353 | MT370488 | MT370486 |
| Distoseptispora adscendens | HKUCC 10820 | DQ408561 | – | – | DQ435092 |
| Distoseptispora amniculi | MFLUCC17–2129T | MZ868761 | MZ868770 | – | MZ892982 |
| Distoseptispora appendiculata | MFLUCC 18–0259T | MN163023 | MN163009 | MN174866 | – |
| Distoseptispora aqualignicola | KUNCC 21–10729T | ON40845 | OK341186 | OP413480 | OP413474 |
| Distoseptispora aquamyces | KUNCC 21–10732T | OK341199 | OK341187 | OP413482 | OP413476 |
| Distoseptispora aquatica | MFLUCC 15–0374T | KU376268 | MF077552 | – | – |
| Distoseptispora aquatica | MFLUCC 16–0904 | MK849794 | MK828649 | MN194053 | – |
| Distoseptispora aquatica | MFLUCC 18–0646 | MK849793 | MK828648 | MN194052 | – |
| Distoseptispora aquatica | S-965 | MK849792 | MK828647 | MN194051 | MN124537 |
| Distoseptispora atroviridis | GZCC 20–0511T | MZ868763 | MZ868772 | MZ892978 | MZ892984 |
| Distoseptispora atroviridis | GZCC 19–0531 | MZ227223 | MW133915 | – | – |
| Distoseptispora bambusae | MFLUCC 20–0091T | MT232718 | MT232713 | MT232880 | MT232881 |
| Distoseptispora bambusae | MFLUCC 14–0583 | MT232717 | MT232712 | – | MT232882 |
| Distoseptispora bambusae | KUNCC 21–10732 | OK341200 | OK341188 | OP413492 | OP413487 |
| Distoseptispora bangkokensis | MFLUCC 18–0262T | MZ518206 | MZ518205 | – | – |
| Distoseptispora cangshanensis | MFLUCC 16–0970 T | MG979761 | MG979754 | MG988419 | – |
| Distoseptispora caricis | CPC 36498T | MN567632 | MN562124 | – | MN556805 |
| Distoseptispora caricis | CPC 36442T | – | MN562125 | – | MN556806 |
| Distoseptispora chinense | GZCC 21–0665 T | MZ474867 | MZ474871 | MZ501609 | – |
| Distoseptispora clematidis | MFLUCC 17–2145T | MT214617 | MT310661 | – | MT394721 |
| Distoseptispora clematidis | KUMCC 21–10727 | OK341197 | OK341184 | OP413488 | OP413483 |
| Distoseptispora crassispora | KUMCC 21–10726T | OK341196 | OK310698 | OP413479 | OP413473 |
| Distoseptispora curvularia | KUMCC 21–10725T | OK341195 | OK310697 | OP413478 | OP413472 |
| Distoseptispora dehongensis | KUMCC 18–0090 T | MK079662 | MK085061 | MK087659 | – |
| Distoseptispora dehongensis | MFLUCC 19–0335 | OK341201 | OK341189 | OP413491 | OP413486 |
| Distoseptispora dehongensis | MFLUCC 17–2326 | OK341193 | OK341183 | OP413493 | – |
| Distoseptispora effusa | GZCC 19–0532T | MZ227224 | MW133916 | MZ206156 | – |
| Distoseptispora euseptata | MFLUCC 20–0154T | MW081544 | MW081539 | – | MW151860 |
| Distoseptispora euseptata | DLUCC S2024 | MW081545 | MW081540 | MW084994 | MW084996 |
| Distoseptispora fasciculata | KUMCC 19–0081T | MW287775 | MW286501 | MW396656 | – |
| Distoseptispora fluminicola | MFLUCC 15–0417T | KU376270 | NR154041 | – | – |
| Distoseptispora fusiformis | GZCC 20–0512T | MZ868764 | MZ868773 | MZ892979 | MZ892985 |
| Distoseptispora guizhouensis | GZCC 21–0666T | MZ474869 | MZ474868 | MZ501610 | MZ501611 |
| Distoseptispora guttulata | MFLUCC 16–0183T | MF077554 | MF077543 | MF135651 | – |
| Distoseptispora hyalina | MFLUCC 17–2128 T | MZ868760 | MZ868769 | MZ892976 | MZ892981 |
| Distoseptispora hydei | MFLUCC 20–0115T | MT742830 | MT734661 | – | MT767128 |
| Distoseptispora lancangjiangensis | DLUCC 1864T | MW879522 | MW723055 | – | – |
| Distoseptispora leonensis | HKUCC 10822 | DQ408566 | – | – | DQ435089 |
| Distoseptispora lignicola | MFLUCC 18–0198T | MK849797 | MK828651 | – | – |
| Distoseptispora longispora | HFJAU 0705T | MH555357 | MH555359 | – | – |
| Distoseptispora martinii | CGMCC 3.18651 | KX033566 | KU999975 | – | – |
| Distoseptispora meilingensis | JAUCC 4727T | OK562396 | OK562390 | OK562408 | – |
| Distoseptispora meilingensis | JAUCC 4728T | OK562397 | OK562391 | OK562409 | – |
| Distoseptispora multiseptata | MFLUCC 16–1044 | MF077555 | MF077544 | MF135652 | MF135644 |
| Distoseptispora multiseptata | MFLUCC 15–0609T | KX710140 | KX710145 | MF135659 | – |
| Distoseptispora neorostrata | MFLUCC 18–0376T | MN163017 | MN163008 | – | – |
| Distoseptispora nonrostrata | KUNCC 21–10730T | OK341198 | OK310699 | OP413481 | OP413475 |
| Distoseptispora obclavata | MFLUCC 18–0329T | MN163010 | MN163012 | – | – |
| Distoseptispora obpyriformis | DLUCC 0867 | MG979765 | MG979757 | MG988423 | MG988416 |
| Distoseptispora obpyriformis | MFLUCC 17–1694T | MG979764 | – | MG988422 | MG988415 |
| Distoseptispora pachyconidia | KUMCC 21–10724T | OK341194 | OK310696 | OP413477 | OP413471 |
| Distoseptispora palmarum | MFLUCC 18–1446T | MK079663 | MK085062 | MK087660 | MK087670 |
| Distoseptispora phangngaensis | MFLUCC 16–0857T | MF077556 | MF077545 | MF135653 | – |
| Distoseptispora rayongensis | MFLUCC 18–0417 | MH457138 | MH457173 | MH463254 | MH463256 |
| Distoseptispora rayongensis | MFLUCC 18–0415T | MH457137 | MH457172 | MH463253 | MH463255 |
| Distoseptispora rayongensis | MFLU 19–0543 | MN163010 | MN513037 | OP413490 | OP413485 |
| Distoseptispora rostrata | DLUCC 0885 | MG979767 | MG979759 | MG988425 | – |
| Distoseptispora rostrata | MFLUCC 16–0969T | MG979766 | MG979758 | MG988424 | MG988417 |
| Distoseptispora saprophytica | MFLUCC 18–1238T | MW287780 | MW286506 | MW396651 | MW504069 |
| Distoseptispora songkhlaensis | MFLUCC 18–1234T | MW287755 | MW286482 | MW396642 | – |
| Distoseptispora suoluoensis | MFLUCC 17–1305 | MF077558 | MF077547 | – | MZ945510 |
| Distoseptispora suoluoensis | MFLUCC 17–0224T | MF077557 | MF077546 | MF135654 | – |
| Distoseptispora tectonae | MFLUCC 12–0291T | KX751713 | KX751711 | KX751710 | KX751708 |
| Distoseptispora tectonae | MFLUCC 15–0981 | MW287763 | MW286489 | MW396641 | – |
| Distoseptispora tectonae | MFLUCC 16–0946 | MG979768 | MG979760 | MG988426 | MG988418 |
| Distoseptispora tectonae | KUNCC 21–10728 | OK348852 | OK341185 | OP413489 | OP413484 |
| Distoseptispora tectonigena | MFLUCC 12–0292T | KX751714 | KX751712 | – | KX751709 |
| Distoseptispora thailandica | MFLUCC 16–0270T | MH260292 | MH275060 | MH412767 | – |
| Distoseptispora thysanolaenae | KUMCC 18–0182T | MK064091 | MK045851 | MK086031 | – |
| Distoseptispora thysanolaenae | HKAS 112710 | MW879524 | MW723057 | MW729783 | – |
| Distoseptispora verrucosa | GZCC 20–0434T | MZ868762 | MZ868771 | MZ892977 | MZ892983 |
| Distoseptispora xishuangbannaensis | KUMCC 17–0290T | MH260293 | MH275061 | MH412768 | MH412754 |
| Distoseptispora yongxiuensis | JAUCC 4725T | OK562394 | OK562388 | OK562406 | – |
| Distoseptispora yongxiuensis | JAUCC 4726T | OK562395 | OK562389 | OK562407 | – |
| Distoseptispora yunjushanensis | JAUCC 4724T | OK562398 | OK562392 | OK562410 | – |
| Distoseptispora yunjushanensis | JAUCC 4723T | OK562399 | OK562393 | OK562411 | – |
| Distoseptispora yunnanensis | MFLUCC 20–0153T | MW081546 | MW081541 | MW081541 | MW151861 |
| Fluminicola saprophytica | MFLUCC 15–0976T | MF374367 | MF374358 | MF370956 | MF370954 |
| Myrmecridium banksiae | CPC 19852T | JX069855 | JX069871 | – | – |
| Myrmecridium schulzeri | CBS 100.54 | EU041826 | EU041769 | – | – |
| Papulosa amerospora | AFTOL-ID 748 | DQ470950 | – | DQ471069 | DQ470901 |
| Pleurophragmium bambusinum | MFLUCC 12–0850 | KU863149 | KU940161 | KU940213 | – |
| Pseudostanjehughesia aquitropica | MFLUCC 16–0569T | MF077559 | MF077548 | MF135655 | – |
| Pseudostanjehughesia lignicola | MFLUCC 15–0352T | MK849787 | MK828643 | MN194047 | MN124534 |
| Wongia griffinii | DAR 80512T | KU850471 | KU850473 | – | – |
Notes: The ex-type cultures are indicated using “T” after strain numbers; newly generated sequences are indicated in bold. “–” stands for no sequence data in GenBank.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, H.; Zhu, R.; Qing, Y.; Yang, H.; Li, C.; Wang, G.; Zhang, D.; Ning, P. Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water. J. Fungi 2022, 8, 1063. https://doi.org/10.3390/jof8101063
AMA Style
Zhang H, Zhu R, Qing Y, Yang H, Li C, Wang G, Zhang D, Ning P. Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water. Journal of Fungi. 2022; 8(10):1063. https://doi.org/10.3390/jof8101063
Chicago/Turabian StyleZhang, Huang, Rong Zhu, Yun Qing, Hao Yang, Chunxue Li, Gennuo Wang, Di Zhang, and Ping Ning. 2022. "Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water" Journal of Fungi 8, no. 10: 1063. https://doi.org/10.3390/jof8101063
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.