
Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies

 ,
,
Abstract
:1. Introduction
2. Bioactive New Metabolites Isolated from Endophytic Fungi and Their Biological Activities
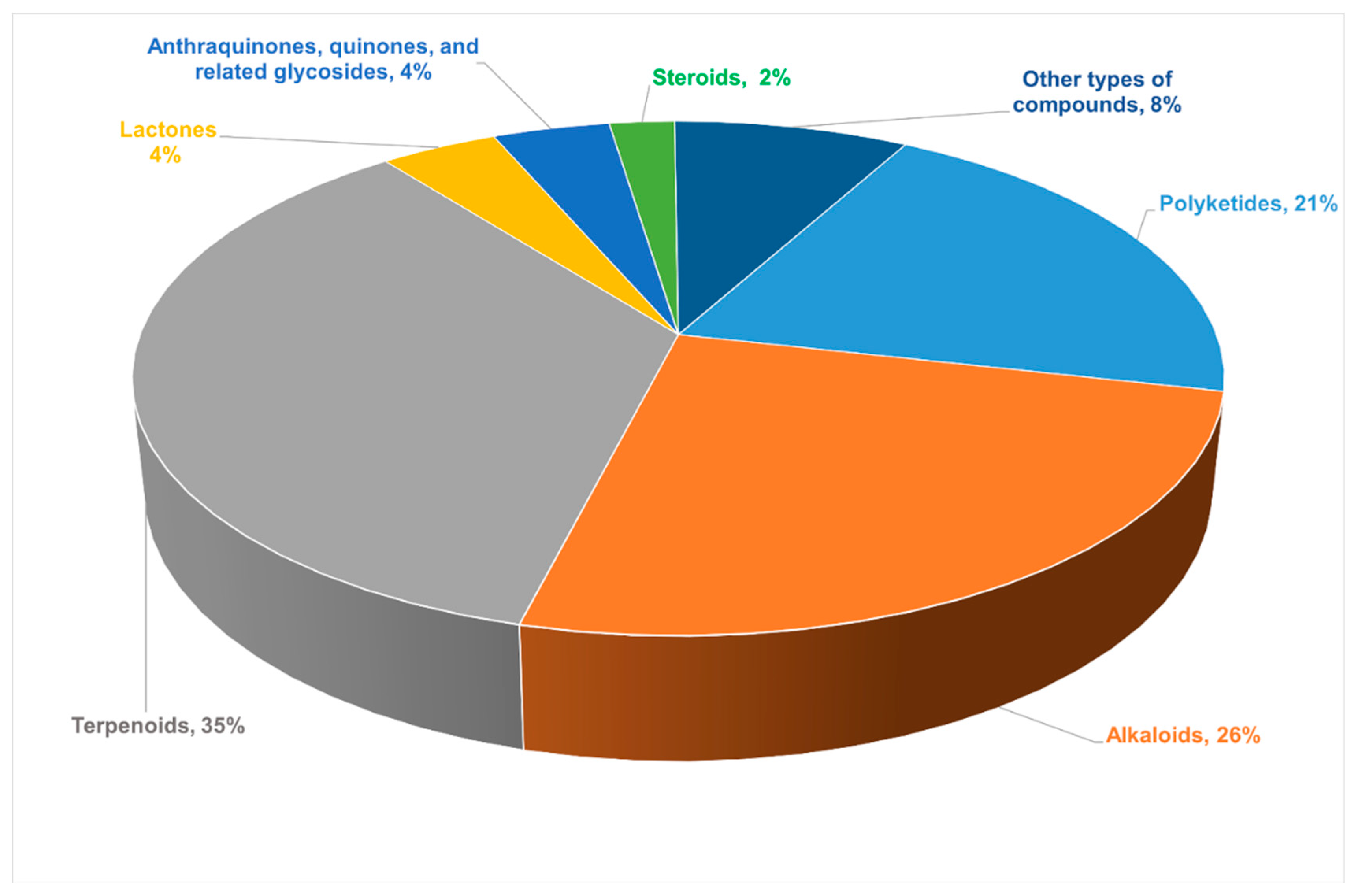
2.1. Polyketides
2.1.1. Chromones
2.1.2. α-Pyrones
2.1.3. Other Polyketides
2.2. Alkaloids
2.2.1. Cytochalasin
2.2.2. Indole Alkaloids
2.2.3. Diketopiperazine Derivatives
2.2.4. Other Types of Alkaloids
2.3. Terpenoids
2.3.1. Sesquiterpenoids and Their Derivatives
2.3.2. Diterpenoids
2.3.3. Triterpenoids
2.3.4. Meroterpenoids
2.4. Lactones
2.5. Anthraquinones, Quinones, and Related Glycosides
2.6. Steroids
2.7. Other Types of Compounds
3. Future Prospects and Challenges of Using Endophytic Fungi as an Alternative Source of Plant Bioactive Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hyde, K.D.; Xu, J.C.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Patchett, A.; Newman, J.A. Comparison of Plant Metabolites in Root Exudates of Lolium perenne Infected with Different Strains of the Fungal Endophyte Epichlo festucae var. lolii. J. Fungi 2021, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Odelade, K.A.; Babalola, O.O. Bacteria, fungi and archaea domains in rhizospheric soil and their effects in enhancing agricultural productivity. Int. J. Environ. Res. Public Health 2019, 16, 3873. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O.; Cheseto, X.; Torto, B. Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiol. Res. 2021, 242, 126640. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhao, Q.; Zhang, X.M.; Li, W.J. Research progress on bioactive products from endophytes. J. Microbiol. 2018, 38, 103–113. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic fungi: A Source of Potential Antifungal Compounds. J. Fungi. 2018, 4, 77. [Google Scholar] [CrossRef]
- Kouipou, R.M.T.; Boyom, F.F. Endophytic fungi from Terminalia species: A comprehensive review. J. Fungi 2019, 5, 43. [Google Scholar] [CrossRef]
- Popli, D.; Anila, V.; Subramanyama, A.B.; Namratha, M.N.; Ranjitha, V.R.; Raoa, S.N.; Ravishankar, V.; Govindappaa, M. Endophyte fungi, Cladosporium species-mediated synthesis of silver nanoparticles possessing in vitro antioxidant, anti-diabetic and anti-Alzheimer activity. Artif. Cell Nanomed. Biotechnol. 2018, 46, 676–683. [Google Scholar] [CrossRef]
- Zheng, R.H.; Li, S.J.; Zhang, X.; Zhao, C.Q. Biological activities of some new secondary metabolites isolated from endophytic fungi: A review study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef]
- Rustamova, N.; Bozorov, K.; Efferth, T.; Yili, A. Novel secondary metabolites from endophytic fungi: Synthesis and biological properties. Phytochem. Rev. 2020, 19, 425–448. [Google Scholar] [CrossRef]
- Ding, W.J.; Wang, S.S.; Ren, J.Q.; Li, G.; Zhan, J.P. Progress on plant endophyte. Curr. Biotechnol. 2015, 5, 425–428. [Google Scholar] [CrossRef]
- Jin, L.R.; Yang, L.; Li, W.J.; Xu, D.; Yang, N.N.; Li, G.Q.; Wan, P. Diversity and Biocontrol Potential of Culturable Endophytic Fungi in Cotton. Front. Microbiol. 2021, 12, 698930. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Liu, J.M.; Sun, J.; Huang, Y.T.; Jin, N.; Li, M.M.; Liang, Y.T.; Fan, B.; Wang, F.Z. Analysis of Endophytic Bacterial Diversity From Different Dendrobium Stems and Discovery of an Endophyte Produced Dendrobine-Type Sesquiterpenoid Alkaloids. Front. Microbiol. 2020, 12, 775665. [Google Scholar] [CrossRef]
- Kharwar, R.N.; Mishra, A.; Stierle, A.; Kharwar, R.N.; Gond, S.K.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Wu, B.; Hussain, M.; Zhang, W.W.; Stadler, M.; Liu, X.Z.; Xiang, M.C. Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Vogl, A. Mehl und die andiron mehlprodukt der cerealien und leguminosen. Nahrunsm. Unters. Hug. Waren. 1898, 12, 25–29. [Google Scholar]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.S.; Silva, C.A.; Hamerski, L. Natural Products from Endophytic Fungi associated with Rubiaceae Species. J. Fungi 2020, 6, 128. [Google Scholar] [CrossRef]
- Gokhale, M.; Gupta, D.; Gupta, U.; Faraz, R.; Sandhu, S.S. Patents on endophytic fungi. Curr. Med. Chem. 2017, 11, 120–140. [Google Scholar] [CrossRef]
- Oberhofer, M.; Wackerlig, J.; Zehl, M.; Büyük, H.; Cao, J.J.; Urban, E.; Zotchev, S.B. Endophytic Akanthomyces sp. LN303 from edelweiss produces emestrin and two new 2-hydroxy-4 pyridone alkaloids. ACS Omega 2021, 6, 2184–2191. [Google Scholar] [CrossRef]
- Mao, Z.L.; Zhang, W.H.; Wu, C.Y.; Feng, H.; Peng, Y.H.; Shahid, H.; Cui, Z.N.; Ding, P.; Shan, T.J. Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbiol. 2021, 21, 155. [Google Scholar] [CrossRef]
- Pereira, C.B.; Oliveira, D.M.; Hughes, A.F.; Kohlhoff, M.; Vieira1, M.L. Endophytic fungal compounds active against Cryptococcus neoformans and C. gattii. J. Antibiot. 2015, 68, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Abdou, R.; Shabana, S.; Rateb, M.E. Terezine E, bioactive prenylated tryptophan analogue from an endophyte of Centaurea stoebe. Nat. Prod. Res. 2020, 34, 503–510. [Google Scholar] [CrossRef]
- Carvalho, C.R.D.; Vieira, M.D.L.A.; Cantrell, C.L.; Wedge, D.E.; Alves, T.M.; Zani, C.L.; Pimenta, R.S.; Rosaa, L.H. Biological activities of ophiobolin k and 6-epi-ophiobolin k produced by the endophytic fungus Aspergillus calidoustus. Nat. Prod. Res. 2016, 30, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Jouda, J.B.; Tamokou, J.; Mbazoa, C.D.; Douala-Meli, C.; Sarkar, P.; Bag, P.K.; Wandji, J. Antibacterial and cytotoxic cytochalasins from the endophytic fungus Phomopsis sp. harbored in garcinia kola (heckel) nut. BMC Complement. Altern. Med. 2016, 16, 462. [Google Scholar] [CrossRef] [PubMed]
- Sana, T.; Siddiqui, B.S.; Shahzad, S.; Farooq, A.D.; Siddiqui, F.; Sattar, S.; Begumet, S. Antiproliferative activity and characterization of metabolites of Aspergillus nidulans: An endophytic fungus from Nyctanthes arbor-tristis linn. against three human cancer cell lines. Med. Chem. 2019, 15, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Sahar, L.; Doustmorad, Z. Antiproliferative and antimicrobial activities of secondary metabolites and phylogenetic study of endophytic Trichoderma species from Vinca plants. Front. Microbiol. 2018, 9, 1484. [Google Scholar] [CrossRef]
- Dhakshinamoorthy, M.; Ponnusamy, S.K.; Kannaian, U.P.; Srinivasan, B.; Shankar, S.N.; Packiam, K.K. Plant-microbe interactions implicated in the production of camptothecin—An anticancer biometabolite from Phyllosticta elongata MH458897 a novel endophytic strain isolated from medicinal plant of Western Ghats of India. Environ. Res. 2021, 201, 111564. [Google Scholar] [CrossRef]
- Han, W.X.; Li, W.Z.; Li, X.F.; Zhang, H.; Yang, S.Z.; Du, J.F.; Jing, H.; Cheng, C.B. Isolation and identification of endophytic fungus producing Huperzine A from Huperzia serrata. Microbiol. China 2017, 44, 2153–2160. [Google Scholar] [CrossRef]
- Khan, N.; Afroz, F.; Begum, M.N.; Rony, S.R.; Sharmin, S.; Moni, F.; Hasan, C.M.; Shaha, K.; Sohrab, M.H. Endophytic Fusarium solani: A rich source of cytotoxic and antimicrobial napthaquinone and aza-anthraquinone derivatives. Toxicol. Rep. 2018, 5, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Mazlan, N.W.; Tate, R.; Yusoff, Y.M.; Clements, C.; Edrada-Ebel, R. Metabolomics-guided isolation of anti-trypanosomal compounds from endophytic fungi of the mangrove plant Avicennia lanata. Curr. Med. Chem. 2020, 27, 1815–1835. [Google Scholar] [CrossRef]
- Okoye, F.B.; Lu, S.; Nworu, C.S.; Esimone, C.O.; Proksch, P.; Chadlic, A.; Debbaba, A. Depsidone and diaryl ether derivatives from the fun gus Corynespora cassiicola, an endophyte of Gongronema latifolium. Tetrahedron Lett. 2013, 54, 4210–4214. [Google Scholar] [CrossRef]
- Liu, G.R.; Huo, R.Y.; Zhai, Y.N.; Liu, L. New bioactive sesquiterpeniods from the plant endophytic fungus Pestalotiopsis theae. Front. Microbiol. 2021, 12, 641504. [Google Scholar] [CrossRef] [PubMed]
- Seetharaman, P.; Gnanasekar, S.; Chandrasekaran, R.; Chandrakasan, G.; Kadarkarai, M.; Sivaperumal, S. Isolation and characterization of anticancer flavone chrysin (5,7-dihydroxy flavone)-producing endophytic fungi from Passiflora incarnata L. leaves. Ann. Microbiol. 2017, 67, 321–331. [Google Scholar] [CrossRef]
- Hu, Z.B.; Wu, Z.H.; Su, Q.H.; Li, M.Z.; Wu, S.S.; Meng, R.Q.; Ding, W.J.; Li, C.Y. Metabolites with phytopathogenic fungi inhibitory activities from the mangrove endophytic fungus Botryosphaeria ramose. Bioorg. Chem. 2020, 104, 104300. [Google Scholar] [CrossRef]
- Hashad, N.; Ibrahim, R.; Mady, M.; Abdel-Aziz, M.S.; Moharram, F.A. Review: Bioactive metabolites and host-specific toxins from endophytic fungi, alternaria alternate. Vietnam. J. Chem. 2021, 59, 733–759. [Google Scholar] [CrossRef]
- Li, H.; Tian, J.M.; Tang, H.Y.; Pan, S.Y.; Zhang, A.L.; Gao, J.M. Chaetosemins A–E, new chromones isolated from an Ascomycete Chaetomium seminudum and their biological activities. RSC Adv. 2015, 37, 29185–29192. [Google Scholar] [CrossRef]
- Liu, S.C.; Liu, L. Isoprenylated chromones from the plant endophytic fungus Pestalotiopsis fici. Mycosystema 2011, 29, 585–587. [Google Scholar] [CrossRef]
- Apgma, B.; Ajcc, A.; Anb, C.; Ma, C.; Ba, C.; Laep, A. Tetrasubstituted α-pyrone derivatives from the endophytic fungus, Neurospora udagawae. Phytochem. Lett. 2020, 35, 147–151. [Google Scholar] [CrossRef]
- Dong, L.; Li, X.M.; Li, C.S.; Wang, B.G. Nigerasterols A and B, Antiproliferative Sterols from the Mangrove-Derived Endophytic Fungus Aspergillus niger MA-132. Helv. Chim. Acta 2013, 96, 1055–1061. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Guo, L.; Che, Y.S.; Liu, L. 2H-Pyran-2-one and 2H-Furan-2-one Derivatives from the plant Endophytic Fungus Pestalotiopsis fici. Chem. Biodivers. 2013, 10, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.M.; Hou, Y.T.; Zhu, L.J.; Chen, Y.J.; Dong, M.; Zhou, M.; Shen, D.; Chen, M.H. Two new α-pyronoids from endophytic fungus Aspergillus oryzae derived from Paris polyphylla var. yunnanensis. Chin. Tradit. Herb. Drugs 2020, 51, 4891–4895. [Google Scholar] [CrossRef]
- Guo, D.L.; Qiu, L.; Feng, D.; He, X.; Li, X.H.; Cao, Z.X.; Gu, Y.C.; Mei, L.; Deng, F.; Deng, Y. Three new α-pyrone derivatives induced by chemical epigenetic manipulation of Penicillium herquei, an endophytic fungus isolated from Cordyceps sinensis. Nat. Prod. Res. 2020, 34, 958–964. [Google Scholar] [CrossRef]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.Y.; Lin, R.K.; Lee, T.H. Angiogenesis Inhibitors and Anti-Inflammatory Agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef] [PubMed]
- Anoumedem, G.; Mountessou, B.; Kouam, S.F.; Narmani, A.; Surup, F. Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3. Antibiotics 2020, 9, 753. [Google Scholar] [CrossRef]
- Zhang, F.Z.; Li, X.M.; Li, X.; Yang, S.Q.; Meng, L.H.; Wang, B.G. Polyketides from the Mangrove-Derived Endophytic Fungus Cladosporium cladosporioides. Mar. Drugs 2019, 17, 296. [Google Scholar] [CrossRef]
- Ola, A.R.; Tawo, B.D.; Belli, H.L.; Proksch, P.; Tommy, D.; Hakim, E.H. A new antibacterial polyketide from the endophytic fungi Aspergillus fumigatiaffinis. Nat. Prod. Commun. 2018, 13, 1573–1574. [Google Scholar] [CrossRef]
- Zhao, S.S.; Wang, B.; Tian, K.L.; Ji, W.X.; Zhang, T.Y.; Ping, C.; Yan, W.; Ye, Y.H. Novel metabolites from the Cercis chinensis derived endophytic fungus Alternaria alternata ZHJG5 and their antibacterial activities. Pest. Manag. Sci. 2021, 77, 2264–2271. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Sarotti, A.M.; Wu, X.H.; Yang, B.J.; Turkson, J.; Chen, Y.F.; Liu, Q.S.; Cao, S.G. An Unusual Benzoisoquinoline-9-one Derivative and Other Related Compounds with Antiproliferative Activity from Hawaiian Endophytic Fungus Peyronellaea sp. FT431. Molecules 2019, 24, 196. [Google Scholar] [CrossRef]
- Wu, J.C.; Hou, Y.; Xu, Q.H.; Jin, X.J.; Chen, Y.X.; Fang, J.G.; Hu, B.R.; Wu, Q.X. (±)-Alternamgin, a Pair of Enantiomeric Polyketides, from the Endophytic Fungi Alternaria sp. MG1. Org. Lett. 2019, 21, 1551–1554. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.W.; Hu, K.; Su, X.Z.; Li, X.N.; Puno, P.T. Phomopsisins A-C: Three new cytochalasans from the plant endophytic fungus Phomopsis sp. sh917. Tetrahedron 2020, 76, 131475. [Google Scholar] [CrossRef]
- Chen, C.M.; Tong, Q.Y.; Zhu, H.C.; Tan, D.D.; Zhang, J.W.; Xue, Y.B.; Yao, G.M.; Luo, Z.W.; Wang, J.P.; Wang, Y.Y. Nine new cytochalasan alkaloids from Chaetomium globosum TW1-1 (Ascomycota, Sordariales). Sci. Rep. 2016, 6, 18711. [Google Scholar] [CrossRef]
- Yang, X.L.; Wu, P.; Xue, J.H.; Li, H.X.; Wei, X.Y. Cytochalasans from endophytic fungus Diaporthe sp. SC-J0138. Fitoterapia 2020, 145, 104611. [Google Scholar] [CrossRef] [PubMed]
- Mou, Q.L.; Yang, S.X.; Xiang, T.; Liu, W.W.; Yang, J.; Guo, L.P.; Wang, W.J.; Yang, X.L. New cytochalasan alkaloids and cyclobutane dimer from an endophytic fungus Cytospora chrysosperma in Hippophae rhamnoides and their antimicrobial activities. Tetrahedron Lett. 2021, 87, 153207. [Google Scholar] [CrossRef]
- Yang, J.; Gong, L.Z.; Guo, M.M.; Jiang, Y.; Ding, Y.; Wang, Z.J.; Xin, X.J.; An, F.L. Bioactive Indole Diketopiperazine Alkaloids from the Marine Endophytic Fungus Aspergillus sp. YJ191021. Mar. Drugs 2021, 19, 157. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wei, X.L.; Xue, L.; Zhang, Z.F.; Zhang, P. Antimicrobial Meroterpenoids and Erythritol Derivatives Isolated from the Marine-Algal-Derived Endophytic Fungus Penicillium chrysogenum XNM-12. Mar. Drugs 2020, 18, 578. [Google Scholar] [CrossRef] [PubMed]
- Lutfia, L.; Munir, E.; Yurnaliza, Y.; Basyuni, M. Chemical analysis and anticancer activity of sesterterpenoid from an endophytic fungus Hypomontagnella monticulosa Zg15SU and its host Zingiber griffithii Baker. Heliyon 2021, 7, e06292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Jiang, Y.; Xin, X.J.; An, F.L. Bioactive indole alkaloids from insect derived endophytic Aspergillus lentulus. Fitoterapia 2021, 153, 104973. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.J.; Huo, G.M.; Zhang, Q.; Li, D.; Wang, D.C.; Qi, J.Z.; Gao, J.M. Phaeosphaones: Tyrosinase Inhibitory Thiodiketopiperazines from an Endophytic Phaeosphaeria fuckelii. J. Nat. Prod. 2020, 83, 1592–1597. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Wang, J.N. Oxepine-Containing Diketopiperazine Alkaloids from the Algal-Derived Endophytic Fungus Paecilomyces variotii EN-291. Helv. Chim. Acta 2015, 98, 800–804. [Google Scholar] [CrossRef]
- Ye, G.T.; Huang, C.Y.; Li, J.L.; Chen, T.; Tang, J.; Liu, W.B.; Long, Y.H. Isolation, Structural Characterization and Antidiabetic Activity of New Diketopiperazine Alkaloids from Mangrove Endophytic Fungus Aspergillus sp. 16-5c. Mar. Drugs 2021, 19, 402. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.H.; Peng, Z.; Li, X.M.; Wang, B.G. Penicibrocazines A–E, Five New Sulfide Diketopiperazines from the Marine-Derived Endophytic Fungus Penicillium brocae. Mar. Drugs. 2015, 13, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.H.; Wang, C.Y.; Mándi, A.; Li, X.M.; Hu, X.Y.; Kassack, M.U. Three Diketopiperazine Alkaloids with Spirocyclic Skeletons and One Bisthiodiketopiperazine Derivative from the Mangrove-Derived Endophytic Fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef]
- Zheng, C.J.; Li, L.; Zou, J.P.; Han, T.; Qin, L.P. Identification of a quinazoline alkaloid produced by Penicillium vinaceum, an endophytic fungus from Crocus sativus. Pharm. Biol. 2012, 50, 129–133. [Google Scholar] [CrossRef]
- Sirimangkalakitti, N.; Yokoya, M.; Chamni, S.; Chanvorachote, P.; Plubrukrn, A.; Saito, N.; Suwanborirux, K. Synthesis and Absolute Configuration of Acanthodendrilline, a New Cytotoxic Bromotyrosine Alkaloid from the Thai Marine Sponge Acanthodendrilla sp. Chem. Pharm. Bull. 2016, 64, 258–262. [Google Scholar] [CrossRef]
- Xu, Y.C.; Wang, L.P.; Zhu, G.L.; Zuo, M.X.; Gong, Q.Y.; He, W.W.; Li, M.P.; Yuan, C.M.; Hao, X.J.; Zhu, W.M. New phenylpyridone derivatives from the Penicillium sumatrense GZWMJZ-313, a fungal endophyte of Garcinia multiflora. Chin. Chem. Lett. 2019, 30, 431–434. [Google Scholar] [CrossRef]
- Cui, H.; Lin, Y.; Luo, M.C.; Lu, Y.J.; Huang, X.S.; She, Z.G. Diaporisoindoles A−C: Three Isoprenylisoindole Alkaloid Derivatives from the Mangrove Endophytic Fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef]
- Wu, S.H.; Zhao, L.X.; Chen, Y.W.; Huang, R.; Miao, C.P.; Wang, J. Sesquiterpenoids from the Endophytic Fungus Trichoderma sp. PR-35 of Paeonia delavayi. Chem. Biodivers. 2011, 9, 1717–1723. [Google Scholar] [CrossRef]
- Wang, A.; Yin, R.Y.; Zhou, Z.Y.; Gu, G.; Dai, J.G.; Lai, D.W.; Zhou, L.G. Eremophilane-Type Sesquiterpenoids from the Endophytic Fungus Rhizopycnis vagum and Their Antibacterial, Cytotoxic, and Phytotoxic Activities. Front. Chem. 2020, 26, 596889. [Google Scholar] [CrossRef]
- Jiang, C.X.; Li, J.; Zhang, J.M.; Jin, X.J.; Yu, B.; Fang, J.G.; Wu, Q.X. Isolation, Identification, and Activity Evaluation of Chemical Constituents from Soil Fungus Fusarium avenaceum SF-1502 and Endophytic Fungus Fusarium proliferatum AF-04. J. Agric. Food Chem. 2019, 67, 839–1846. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y.; Li, X.M.; Meng, L.H.; Wang, B.G. Antioxidant bisabolane-type sesquiterpenoids from algal-derived fungus Aspergillus sydowii EN-434. J. Oceanol. Limnol. 2020, 38, 1532–1536. [Google Scholar] [CrossRef]
- Wang, Q.X.; Bao, L.; Yang, X.L.; Liu, D.L.; Guo, H.; Dai, H.Q.; Song, F.H.; Zhang, L.X.; Guo, L.D.; Li, S.J.; et al. Ophiobolins P–T, five new cytotoxic and antibacterial sesterterpenes from the endolichenic fungus Ulocladium sp. Fitoterapia 2013, 90, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.S.; Song, Y.P.; Meng, L.H.; Yang, S.Q.; Li, X.M. Isolation and Characterization of Antibacterial Carotane Sesquiterpenes from Artemisia argyi Associated Endophytic Trichoderma virens QA-8. Antibiotics 2021, 10, 213. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zou, G.; Yang, W.C.; Zhao, Y.Y.; Tan, Q.; Chen, L.; Wang, J.M.; Ma, C.Y.; Kang, W.Y.; She, Z.G. Metabolites with Anti-Inflammatory Activity from the Mangrove Endophytic Fungus Diaporthe sp. QYM12. Mar. Drugs 2021, 19, 56. [Google Scholar] [CrossRef]
- Fan, M.M.; Xiang, G.; Chen, J.W.; Gao, J.; Xue, W.X.; Wang, Y.X.; Li, W.H.; Zhou, L.; Jiao, R.H.; Shen, Y.; et al. Libertellenone M, a diterpene derived from an endophytic fungus Phomopsis sp. S12, protects against DSS-induced colitis via inhibiting both nuclear translocation of NF-κB and NLRP3 inflammasome activation. Int. Immunopharmacol. 2020, 80, 106144. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, X.; Chen, J.W.; Shen, Y.; Jiang, N.; Tan, R.X.; Jiao, R.H.; Ge, H.M. Anti-inflammatory diterpenoids from an endophytic fungus Phomopsis sp. S12. Tetrahedron Lett. 2019, 60, 151045. [Google Scholar] [CrossRef]
- Chen, Y.M.; Yang, Y.H.; Li, X.N.; Zou, C.; Zhao, P.J. Diterpenoids from the Endophytic Fungus Botryosphaeria sp. P483 of the Chinese Herbal Medicine Huperzia serrata. Molecules 2015, 20, 16924–16932. [Google Scholar] [CrossRef]
- Wang, X.N.; Bashyal, B.P.; Wijeratne, E.M.; Ren, J.M.; Liu, M.X.; Gunatilaka, M.K. Smardaesidins A-G, Isopimarane and 20-Nor-isopimarane Diterpenoids from Smardaea sp., a Fungal Endophyte of the Moss Ceratodon purpureus. J. Nat. Prod. 2011, 74, 2052–2061. [Google Scholar] [CrossRef]
- Xu, M.F.; Jia, O.Y.; Wang, S.J.; Zhu, Q. A new bioactive diterpenoid from pestalotiopsis adusta, an endophytic fungus from Clerodendrum canescens. Nat. Prod. Res. 2016, 30, 2642–2647. [Google Scholar] [CrossRef]
- Sun, P.X.; Zheng, C.J.; Li, W.C.; Jin, G.L.; Huang, F.; Qin, L.P. Trichodermanin A, a novel diterpenoid from endophytic fungus culture. J. Nat. Med. 2011, 65, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.F.; Li, X.M.; Meng, L.; Cui, C.M.; Gao, S.S.; Li, C.S.; Huang, C.G.; Wang, B.G. Asperolides A−C, Tetranorlabdane Diterpenoids from the Marine Alga-Derived Endophytic Fungus Aspergillus wentii EN-48. J. Nat. Med. 2012, 75, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Fang, H.Q.; Gui, F.; Wang, Y.Z.; Xu, Q.Y.; Deng, X.M. Two New Ring A-Cleaved Lanostane-Type Triterpenoids and Four Known Steroids Isolated from Endophytic Fungus Glomerella sp. F00244. Helv. Chim. Acta 2016, 99, 601–607. [Google Scholar] [CrossRef]
- Morandini, L.; Neto, A.T.; Pedroso, M.; Antoniolli, Z.I.; Burrow, R.; Morel, A.F. Lanostane-type triterpenes from the fungal endophyte Scleroderma UFSMSc1 (Persoon) Fries. Bioorg. Med. Chem. Lett. 2016, 47, 1173–1176. [Google Scholar] [CrossRef]
- Tian, C.; Gao, H.; Peng, X.P.; Li, G.; Lou, H.X. Fusidic acid derivatives from the endophytic fungus Acremonium pilosum F47. J. Asian Nat. Prod. Res. 2021, 23, 1148–1155. [Google Scholar] [CrossRef]
- Qin, D.; Shen, W.Y.; Wang, Y.Q.; Han, M.J.; Chai, F.N.; Duan, X.X.; Yan, X.; Guo, J.L.; Gao, T.C.; Zuo, S.H.; et al. Enhanced production of unusual triterpenoids from Kadsura angustifolia fermented by a symbiont endophytic fungus, Penicillium sp. SWUKD4.1850. Phytochemistry 2019, 158, 56–66. [Google Scholar] [CrossRef]
- Liang, H.Q.; Zhang, D.W.; Guo, S.X.; Jie, Y. Two new tetracyclic triterpenoids from the endophytic fungus Hypoxylon sp. 6269. J. Asian Nat. Prod. Res. 2018, 20, 1485662. [Google Scholar] [CrossRef]
- Ibrahim, R.M.; Abdallah, H.M.; Mohamed, G.A.; Ross, S.A. Integracides H-J: New tetracyclic triterpenoids from the endophytic fungus Fusarium sp. Fitoterapia 2016, 112, 161–167. [Google Scholar] [CrossRef]
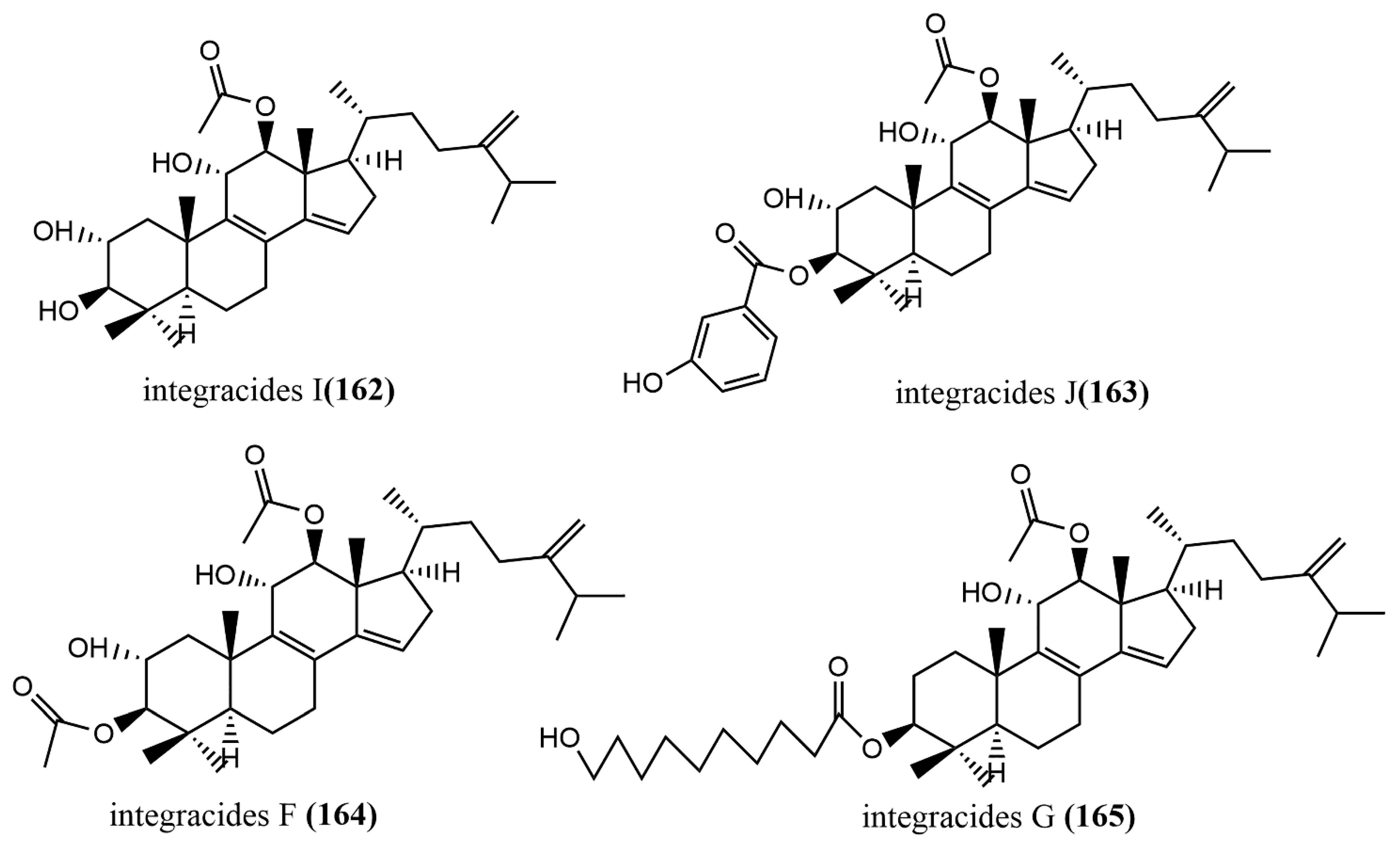
- Ibrahim, R.M.; Abdallah, H.M.; Mohamed, G.A.; Ross, S.A. Integracides F and G: New tetracyclic triterpenoids from the endophytic fungus Fusarium sp. Phytochem. Lett. 2016, 15, 125–130. [Google Scholar] [CrossRef]
- Sun, Z.H.; Liang, F.L.; Wu, W.; Chen, Y.C.; Pan, Q.L.; Li, H.H.; Ye, W.; Liu, H.X.; Li, S.N.; Tan, G.H.; et al. Guignardones P-S, New Meroterpenoids from the Endophytic Fungus Guignardia mangi ferae A348 Derived from the Medicinal Plant Smilax glabra. Molecules 2015, 20, 22900–22907. [Google Scholar] [CrossRef]
- Li, Q.; Chen, C.M.; Cheng, L.; Wei, M.S.; Dai, C.; He, Y.; Gong, J.J.; Zhu, R.Q.; Li, X.N.; Liu, J.J.; et al. Emeridones A−F, a Series of 3,5-Demethylorsellinic Acid-Based Meroterpenoids with Rearranged Skeletons from an Endophytic Fungus Emericella sp. TJ29. J. Org. Chem. 2019, 84, 1534–1541. [Google Scholar] [CrossRef]
- Yang, H.G.; Zhao, H.; Li, J.J.; Chen, S.M.; Mou, L.M.; Zou, J.; Chen, G.D.; Qin, S.Y.; Wang, C.X.; Hu, D.; et al. Phyllomeroterpenoids A-C, Multi-biosynthetic Pathway Derived Meroterpenoids from the TCM Endophytic Fungus Phyllosticta sp. and their Antimicrobial Activities. Sci. Rep. 2017, 7, 12925. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.; Silva, F.M.; Magallanes-Noguera, C.A.; Kurina-Sanz, M.; Silva, E. Natural trypanocidal product produced by endophytic fungi through co-culturing. Folia Microbiol. 2020, 65, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Yi, J.L.; Cai, J.; Zhou, X.M.; Chen, L.; Zhuo, X.; Lai, X.Y. Two new bioactive secondary metabolites from the endophytic fungus Talaromyces assiutensis JTY2. Nat. Prod. Res. 2021, 17, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.Z.; Lv, Y.B.; Hao, J.; Chen, H.; Yang, X.; Huang, Y.; Liu, C.; Huang, H.Q.; Ma, Y.R.; Yang, X.Z. Two new compounds of Penicillium polonicum, an endophytic fungus from Camptotheca acuminata Decne. Nat. Prod. Res. 2019, 34, 1879–1883. [Google Scholar] [CrossRef]
- Bussey, R.O.; Kaur, A.; Todd, D.A.; Egan, J.M.; El-Elimat, T.; Graf, T.N.; Raja, H.A.; Oberlies, N.H.; Cech, N.J. Comparison of the chemistry and diversity of endophytes isolated from wild-harvested and greenhouse-cultivated yerba mansa (Anemopsis californica). Phytochem. Lett. 2015, 11, 202–208. [Google Scholar] [CrossRef]
- Tchoukoua, A.; Ota, T.; Akanuma, R.; Ju, Y.M.; Supratman, U.; Murayama, T.; Koseki, T.; Shiono, Y. A phytotoxic bicyclic lactone and other compounds from endophyte Xylaria curta. Nat. Prod. Res. 2017, 31, 2113–2118. [Google Scholar] [CrossRef]
- Chen, S.H.; Liu, Z.M.; Liu, H.J.; Long, Y.H.; Chen, D.N.; Lu, Y.J.; She, Z.G. Lasiodiplactone A, a novel lactone from the mangrove endophytic fungus Lasiodiplodia theobromae ZJ-HQ1. Org. Biomol. Chem. 2017, 15, 6338–6341. [Google Scholar] [CrossRef]
- Gusmao, A.S.; Abreu, L.S.; Tavares, J.F.; Freitas, H.F.; Pita, S.; Santos, E.; Caldas, I.; Vieira, A.A.; Silva, E.O. Computer-guided trypanocidal activity of natural lactones produced by endophytic fungus of Euphorbia umbellata. Chem. Biodivers. 2021, 18, e2100493. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.L.; Wang, X.F.; Xu, K.; Li, W.; Chen, D.; Zhang, P. Characterization of a New Insecticidal Anthraquinone Derivative from an Endophyte of Acremonium vitellinum against Helicoverpa armigera. J. Agric. Food Chem. 2020, 68, 11480–11487. [Google Scholar] [CrossRef]
- Yang, Y.C.; Yang, H.Y.; Li, Y.; Ye, Y.Q.; Hu, Q.F.; Gao, X.M.; Du, G.A. New Xanthone from the Fermentation Products of Endophytic Fungus of Phomopsis Species. Asian J. Chem. 2014, 26, 4591–4593. [Google Scholar] [CrossRef]
- Du, F.Y.; Li, X.M.; Song, J.Y.; Li, C.S.; Wang, B.G. Anthraquinone Derivatives and an Orsellinic Acid Ester from the Marine Alga-Derived Endophytic Fungus Eurotium cristatum EN-220. Helv. Chim. Acta 2014, 97, 973–978. [Google Scholar] [CrossRef]
- Hawas, U.W.; El-Beih, A.A.; El-Halawany, A.M. Bioactive Anthraquinones from Endophytic Fungus Aspergillus versicolor Isolated from Red Sea Algae. Arch. Pharm. Res. 2012, 35, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Teiten, M.H.; Mack, F.; Debbab, A.; Aly, A.H.; Dicato, M.; Proksch, P.; Diederich, M. Anticancer effect of altersolanol A, a metabolite produced by the endophytic fungus Stemphylium globuliferum, mediated by its proapoptotic and antiinvasive potential via the inhibition of NF-kB activity. Bioorg. Med. Chem. 2013, 21, 3850–3858. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, W.C.; Zou, G.; Yan, Z.Y.; Qiu, P.; Long, Z.G.; She, Z.G. Metabolites with anti-inflammatory and α-glucosidase inhibitory activities from the mangrove endophytic fungus Phoma sp. SYSU-SK-7. Tetrahedron Lett. 2020, 61, 152578. [Google Scholar] [CrossRef]
- Ibrahim, S.; Mohamed, G.A.; Haidari, R.; El-Kholy, A.; Zayed, M.F. Fusaristerol A: A New Cytotoxic and Antifungal Ergosterol Fatty Acid Ester from the Endophytic Fungus Fusarium sp. Associated with Mentha longifolia Roots. Pharmacogn. Mag. 2018, 56, 308–311. [Google Scholar] [CrossRef]
- Wu, S.H.; Huang, R.; Miao, C.P.; Chen, Y.W.; Chen, Y.W. Two New Steroids from an Endophytic Fungus Phomopsis sp. Chem. Biodivers. 2013, 10, 1276–1283. [Google Scholar] [CrossRef]
- Qi, C.X.; Gao, W.X.; Wang, J.P.; Liu, M.T.; Zhang, J.W.; Chen, C.M.; Hu, Z.X.; Xue, Y.B.; Li, D.X.; Zhang, Y.H. Errusnolides A-D, new butenolides with anti-inflammatory activities from an endophytic Aspergillus from Tripterygium wilfordii. Fitoterapia 2018, 130, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Akhter, N.; Pan, C.Q.; Liu, Y.Q.; Shi, Y.T.; Wu, B. Isolation and structure determination of a new indene derivative from endophytic fungus Aspergillus flavipes Y-62. Nat. Prod. Res. 2019, 33, 2939–2944. [Google Scholar] [CrossRef]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harrasi, A.; Ahmad, M.; Mabood, F. Microsphaerol and Seimatorone: Two New Compounds Isolated from the Endophytic Fungi, Microsphaeropsis sp. and Seimatosporium sp. Chem. Biodivers. 2015, 46, 289–294. [Google Scholar] [CrossRef]
- Wu, X.; Pang, X.J.; Xu, L.L.; Zhao, T.; Long, X.Y.; Zhang, Q.Y.; Qin, H.L.; Yang, D.F.; Yang, X.L. Two new alkylated furan derivatives with antifungal and antibacterial activities from the plant endophytic fungus Emericella sp. XL029. Nat. Prod. Res. 2017, 32, 2625–2631. [Google Scholar] [CrossRef] [PubMed]
- Niaz, S.I.; Khan, D.; Naz, R.; Safdar, K.; Zain, S.; Abidin, U.; Khan, I.U.; Gul, R.; Khan, W.U.; Aman, M.; et al. Antimicrobial and antioxidant chlorinated azaphilones from mangrove Diaporthe perseae sp. isolated from the stem of Chinese mangrove Pongamia pinnata. J. Asian Nat. Prod. Res. 2020, 23, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.X.; Wang, N.N.; Kang, Y.F.; Ma, Y.M. New furan derivative from an endophytic Aspergillus tubingensis of Decaisnea insignis (Griff.) Hook.f. & Thomson. Nat. Prod. Res. 2019, 33, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.W.; Chen, J.Q.; Zhu, S.R.; He, Y.Y.; Ding, W.J.; Li, C.Y. Two new compounds from Nigrospora sphaerica ZMT05, a fungus derivated from Oxya chinensis Thunber. Nat. Prod. Res. 2018, 32, 2375–2381. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Li, X.M.; Yang, S.Q.; Cao, J.; Li, Y.H.; Wang, B.G. Induced terreins production from marine red algal-derived endophytic fungus Aspergillus terreus EN-539 co-cultured with symbiotic fungus Paecilomyces lilacinus EN-531. J. Antibiot. 2020, 73, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yuan, L.Y.; Guo, D.L.; Ye, Y.; Da-Wa, Z.M.; Wang, X.L.; Ma, F.W.; Chen, L. Bioactive halogenated dihydroisocoumarins produced by the endophytic fungus Lachnum palmae isolated from Przewalskia tangutica. Phytochemistry 2018, 148, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, D.K.; Kharwar, R.N.; White, J.F.; Gond, S.K. Fungal Endophytes as Efficient Sources of Plant-Derived Bioactive Compounds and Their Prospective Applications in Natural Product Drug Discovery: Insights, Avenues, and Challenges. Microorganisms 2021, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S. Endophytic fungi: Novel sources of anticancer lead molecules. Appl. Microbiol. Biotechnol. 2012, 95, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Zhang, L.; Li, L.; Zheng, C.J.; Guo, L.; Li, W.C.; Sun, P.X.; Qin, L.P. Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol. Res. 2010, 165, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G. The Emergence of Endophytic Microbes and Their Biological Promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef]
- Hastuti, U.S.; Asna, P.M.A.; Rahmawati, D. Histologic observation, identification, and secondary metabolites analysis of endophytic fungi isolated from a medicinal plant, hedychium acuminatum Roscoe. AIP Conf. Proc. 2018, 2002, 020070. [Google Scholar] [CrossRef]
- El-Sayed, A.; Khalaf, S.A.; Azez, H.A.; Hussein, H.A.; El-Baz, A.F. Production, bioprocess optimization and anticancer activity of camptothecin from Aspergillus terreus and Aspergillus flavus, endophytes of ficus elastica. Process Biochem. 2021, 107, 59–73. [Google Scholar] [CrossRef]
- Xiong, Z.Q.; Yang, Y.Y.; Zhao, N.; Wang, Y. Diversity of endophytic fungi and screening of fungal paclitaxel producer from anglojap yew, Taxus x media. BMC Microbiol. 2013, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yin, H.; Sun, Y.; Zhang, Z.; Cui, Y. Mutagenesis of a berberine-producing endophytic fungus. J. Fungal Res. 2008, 6, 216–225. [Google Scholar]
- Xu, B.; Wang, M.R.; Xia, Y.; Yang, K.; Zhang, C.Y. Improvement of the output of teicoplanin by genome shuffling. Chin. J. Antibiot. 2006, 31, 237–242. [Google Scholar] [CrossRef]
- Eyberger, A.L.; Dondapati, R.; Porter, J.R. Endophyte Fungal Isolates from Podophyllum peltatum Produce Podophyllotoxin. J. Nat. Prod. 2006, 68, 1121–1124. [Google Scholar] [CrossRef] [PubMed]
- Venugopalan, A.; Srivastava, S. Endophytes as in vitro production platforms of high value plant secondary metabolites. Biotechnol. Adv. 2015, 33, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Vigneshwari, A.; Rakk, D.; Németh, A.; Kocsubé, S. Host metabolite producing endophytic fungi isolated from Hypericum perforatum. PLoS ONE 2019, 14, e0217060. [Google Scholar] [CrossRef]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Staden, J.V. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef]
- Wei, Q.; Bai, J.; Yan, D.J.; Bao, X.Q.; Li, W.T.; Hu, Y.C. Genome mining combined metabolic shunting and osmac strategy of an endophytic fungus leads to the production of diverse natural products. Acta Pharm. Sin. B 2021, 11, 572–587. [Google Scholar] [CrossRef]
- Zhang, L.H.; Niaz, S.I.; Khan, D.; Wang, Z.; Zhu, Y.H.; Zhou, H.Y.; Lin, Y.C.; Li, J.; Liu, L. Induction of Diverse Bioactive Secondary Metabolites from the Mangrove Endophytic Fungus Trichoderma sp. (Strain 307) by Co-Cultivation with Acinetobacter johnsonii (Strain B2). Mar. Drugs 2017, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Vagish, D.; Shreya, K.; Sanjai, S. Isolation and enhancement of resveratrol production in Xylaria psidii by exploring the phenomenon of epigenetics: Using DNA methyltransferases and Histone deacetylase as epigenetic modifiers. Mol. Biol. Rep. 2019, 46, 4123–4137. [Google Scholar] [CrossRef]
- Pu, X.; Qu, X.X.; Chen, F.; Bao, J.K.; Zhang, G.L.; Luo, Y.G. Camptothecin-producing endophytic fungus Trichoderma atroviride ly357: Isolation, identification, and fermentation conditions optimization for camptothecin production. Appl. Microbiol. Biotechnol. 2013, 97, 9365–9375. [Google Scholar] [CrossRef] [PubMed]
- Pelo, S.P.; Adebo, O.A.; Green, E. Chemotaxonomic profiling of fungal endophytes of Solanum mauritianum (alien weed) using gas chromatography high resolution time-of-flight mass spectrometry (GC-HRTOF-MS). Metabolomics 2021, 17, 43. [Google Scholar] [CrossRef] [PubMed]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Endophytic Fungus | Host Plant | Regions/Countries | Compound | Biological Activity | Ref. |
|---|---|---|---|---|---|---|
| 1 | Lophiostoma sp. | Eucalyptus exserta | Guangzhou, China. | Scorpinone | Antibacterial | [22] |
| 2 | Mycosphaerella sp. | Myrciaria floribunda | Amazon rainforest, Brazil. | Myriocin | Antifungal | [23] |
| 3 | Mucor sp. | Centaurea stoebe | Idaho, USA | Terezine E | Antifungal and cytotoxicity | [24] |
| 4 | Aspergillus calidoustus | Acanthospermum australe | Jalapao State Park, Tocantins, Brazil. | Ophiobolin K 6-epi-ophiobolin K | Antifungal, trypanocidal and cytotoxicity | [25] |
| 5 | Phomopsis sp. | Garcinia kola (Heckel) nut | Yaounde, Cameroon | Cytochalasins H | Antibacterial and cytotoxicity | [26] |
| 6 | Aspergillus nidulans | Nyctanthes arbor-tristis Linn | Karachi, Pakistan | Sterigmatocystin | Antiproliferative activity | [27] |
| 7 | Trichoderma asperellum and Trichoderma brevicompactum | Vinca herbacea | Hamedan, Iran | 4b-hydroxy-12,13-epoxytrichothec-9-ene | Antimicrobial and antiproliferative activity | [28] |
| 8 | Phyllosticta elongata | Cipadessa baccifera | Western Ghats, India | Camptothecin | Anticancer agent | [29] |
| 9 | Fusarium verticillioides | Huperzia serrata | Gucheng Mountain, Sichuan, China | Huperzine A | Treatment of Alzheimer’s disease | [30] |
| 10 | Fusarium solani | Cassia alata | Bangladesh | Napthaquinones Azaanthraquinones | Cytotoxicity, antimicrobial and antioxidant activity | [31] |
| 11 | Fusarium sp. and Lasiodiplodia theobromae | Avicennia lanata | Terengganu, Malaysia | Anhydrofusarubin dihydrojavanicin | Antitrypanosomal | [32] |
| 12 | Corynespora cassiicola | Gongronema latifolium | Nigeria | Corynesidone D | Anti-inflammatory/anticancer agent | [33] |
| 13 | Pestalotiopsis theae | Camellia sinensis Theaceae | Hangzhou, China | punctaporonin H | Antibacterial and cytotoxicity | [34] |
| 14 | Phialocephala fortinii | Podophyllum peltatum | Tamilnadu, India | Podophyllotoxin | Antiviral, antioxidant, and antirheumatic activities | [35] |
| Compound | Molecular Formula | Color and Morphology | Endophytic Fungus | Host Plant | Site and Nation | Pharmacological Activity | Ref. |
|---|---|---|---|---|---|---|---|
| Polyketides Chromones | |||||||
| 1 | C12H13O6 | colorless powder | Botryosphaeria ramosa L29 | leaf of Myoporum bontioides | Leizhou Peninsula, China | Displayed acceptable antimicrobial activities against Fusarium oxysporum | [36] |
| 2 | C14H15O6 | white powder | |||||
| 3 | C11H11O5 | ||||||
| 4 | C17H19N3O3S2 | yellow crystals | Phaeosphaeria fuckelii | Phlomis umbrosa | Mount Hua, China | Mushroom tyrosinase inhibitory activity | [37] |
| 5 | C15H16O7S | yellow powder | Chaetomium seminudum | Showed antifungal activity (5–6); Exhibited radical scavenging activity against DPPH; Showed significant antioxidant activity ((5) | [38] | ||
| 6 | C16H18O7S | ||||||
| 7 | C16H24O5 | colorless oil | Pestalotiopsis fici W106-1 | Camellia sinensis | Hangzhou, China | Displayed inhibitory effects on HIV-1 replication in C8166 cells ((7–8); Showed low to moderate cytotoxic activity (9–10); Displayed significant antifungal activity (9) | [39] |
| 8 | |||||||
| 9 | C32H54O6 | ||||||
| 10 | C32H54O6Na | ||||||
| Polyketides α-pyrones | |||||||
| 11 | C11H14O4 | colorless crystals | Neurospora udagawae | shoot of Quercus macranthera | Exhibited moderate antifungal (vs. Rhodoturula glutinis) activity and cytotoxicity against KB3.1 cells (12) | [40] | |
| 12 | C10H10O4 | colorless oil | |||||
| 13 | C28H42O4 | colorless amorphous powder | Aspergillus niger MA-132 | Avicennia marina | Hainan Province, China | Showed potent antifungal and cytotoxic activities | [41] |
| 14 | |||||||
| 15 | C14H22O5 | yellow oil | Pestalotiopsis fici | branches of Camellia sinensis (Theaceae) | Hangzhou, China | Displayed significant antifungal against Gibberella zeae | [42] |
| 17 | C10H14O4 | yellow oil | Aspergillus oryzae | Paris polyphylla var. yunnanensis | Dali, Yunnan Province, China | The biological activities of compounds 17–18 were not tested | [43] |
| 18 | |||||||
| 19 | C11H16O4 | yellow gum | Penicillium herque | Cordyceps sinensis | Xiahe, China | Weak cytotoxic activity | [44] |
| 20 | C12H16O5 | ||||||
| 21 | |||||||
| Polyketides: Other polyketides | |||||||
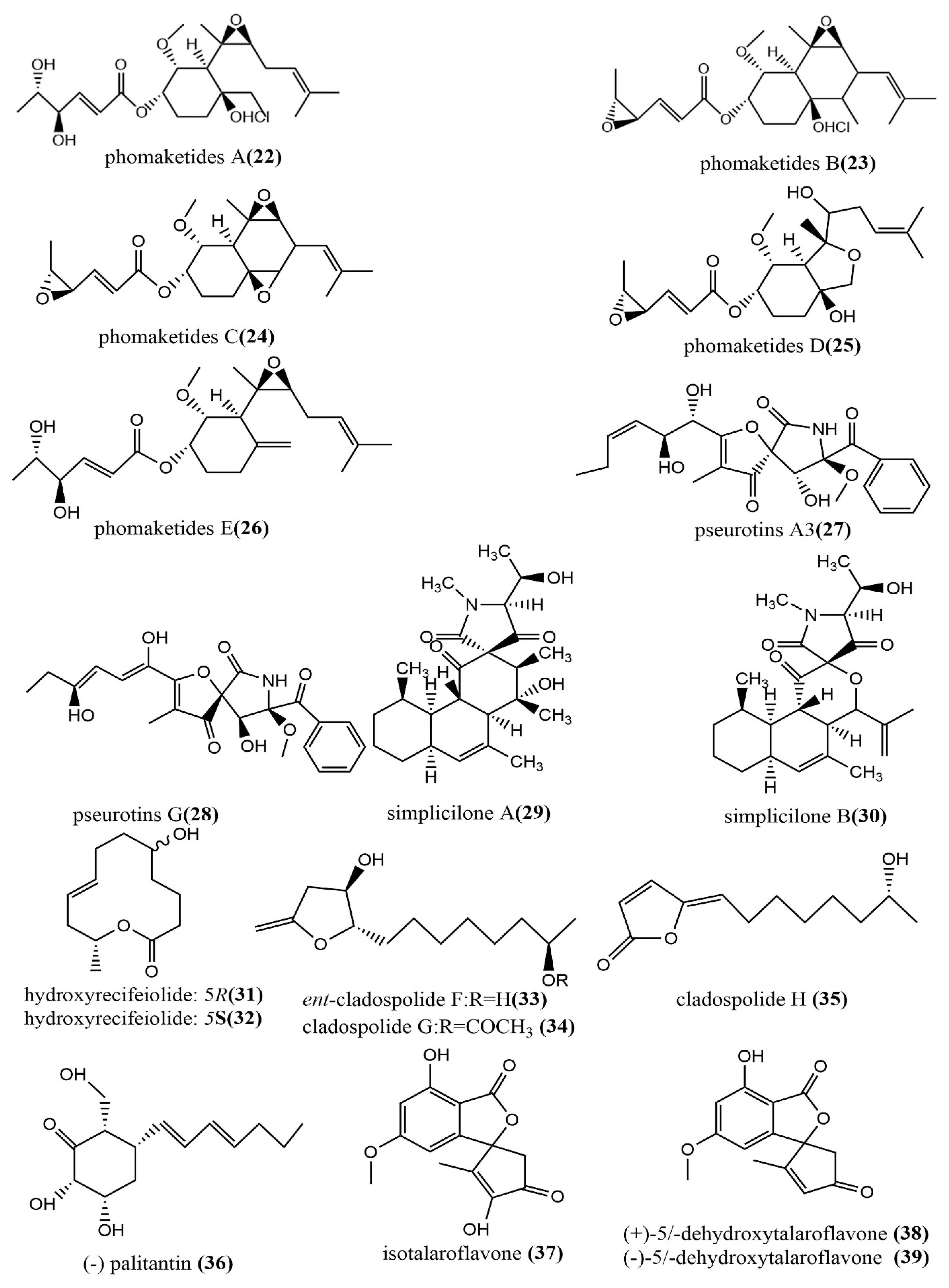
| 22 | C22H35ClO7 | white powder | Phoma sp. NTOU4195 | Pterocladiella capillacea | Taiwan, China | Showed potent anti-angiogenic activity (22); Exhibited inhibition of nitric oxide production in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophage cells (24) | [45] |
| 23 | |||||||
| 24 | |||||||
| 25 | |||||||
| 26 | |||||||
| 27 | C22H34O6 | ||||||
| 28 | C22H26NO8 | ||||||
| 29 | C24H35NO5 | colorless oil | Simplicillium subtropicum SPC3 | fresh bark of Duguetia staudtii | Cameroon | Weak cytotoxic activity | [46] |
| 30 | C24H35NO6 | ||||||
| 31 | C12H20O3 | colorless oil | Cladosporium cladosporioides MA-299 | leaves of the mangrove plant Bruguiera gymnorrhiza | Hainan Island, China | Showed potent antimicrobial((vs. Escherichia coli and Staphylococcus aureus) activity and moderate inhibition activity against acetylcholinesterase (33) | [47] |
| 32 | |||||||
| 33 | C12H22O4 | pale yellow powder | |||||
| 34 | C14H24O5 | pale yellow oil | |||||
| 35 | C12H20O4 | colorless crystals | |||||
| 36 | C14H20O5 | colorless powder | Aspergillus fumigatiaffnis | Tribulus terestris | Weak antimicrobial activities | [48] | |
| 37 | C14H12O6Na | white amorphous solid | Alternariaalternata ZHJG5 | leaf of Cercis hinensis | Nanjing, China | Exhibited potent antimicrobial activity; Showed significant protective effect against the bacterial blight of rice (37) | [49] |
| 38 | C14H12O5Na | white powder | |||||
| 39 | C14H12O5Na | ||||||
| 40 | C29H22O12Na | ||||||
| 41 | C24H27NO5 | brown solid | Peyronellaea sp. FT431 | healthy leaf of a Hawaiian indigenous plant, Verbena sp. | Lyon, France | Showed weak to moderate cytotoxic activity (41–42) | [50] |
| 42 | C24H26O7 | ||||||
| 43 | C24H26O7 | ||||||
| 44 | C18H20O5 | ||||||
| 45 | C29H22O9 | red wine colored lump crystal | Alternaria sp. MG1 | Vitis quinquangularis | Showed weak cytotoxicity | [51] | |
| Alkaloids Cytochalasin | |||||||
| 46 | C22H32N2O5 | white amorphous solid | Phomopsis sp. sh917 | Fresh stems of I. eriocalyx var. laxiflora | Kunming, China | Significant inhibitory activity against NO production in LPS-induced RAW264.7 cells (46) | [52] |
| 47 | C22H33NO4 | ||||||
| 48 | |||||||
| 49 | C32H40N2O6 | colorless amorphous powder | Chaetomium globosum TW1-1 | Armadillidium vulgare | Hubei Province, China | Showed potential cytotoxic activities against cancer cell lines (HL-60, A-549, SMMC-7721, MCF-7, and SW-480) | [53] |
| 50 | C32H38N2O6 | ||||||
| 51 | |||||||
| 52 | C32H40N2O6 | ||||||
| 53 | C32H38N2O5 | white amorphous powder | |||||
| 54 | C32H83N2O6 | ||||||
| 55 | C32H36N2O4 | ||||||
| 56 | colorless amorphous powder | ||||||
| 57 | C34H42N2O7Na | ||||||
| 58 | C28H37NO3 | white amorphous solid | Diaporthe sp. SC-J0138 | Cyclosorus parasiticus (Thelypteridaceae) | Guangdong Province, China | Showed significant cytotoxic activities against four human cancer cell lines (A549, HeLa, HepG2, and MCF-7) (58); Exhibited selective cytotoxic activity (59–62) | [54] |
| 59 | C28H37NO | ||||||
| 60 | |||||||
| 61 | C28H37NO4 | ||||||
| 62 | |||||||
| 63 | C25H37NO4 | colorless crystal | Cytospora chrysosperma HYQZ-931 | Hippophae rhamnoides | Exhibited significant antibacterial activity (63,65) | [55] | |
| 64 | white amorphous powder | ||||||
| 65 | C26H41NO5 | ||||||
| Alkaloids Indole alkaloids | |||||||
| 66 | C26H28N3O4 | brilliant yellowish powder | Aspergillus sp. YJ191021 | Zhejiang Province, China | Exhibited moderate antibacterial activity (66); Displayed notable anti-inflammatory; Exhibited notable cytotoxicity (66–69) | [56] | |
| 67 | C26H29N3O5 | white powder | |||||
| 68 | C27H31N3O6Na | ||||||
| 69 | C27H31N3O6 | ||||||
| 70 | C28H31N3O6 | ||||||
| 71 | C26H31N3O6 | ||||||
| 72 | C30H33NO7 | white amorphous powder | Penicillium chrysogenum XNM-12 | Leathesia nana | Shandong Province, China | Exhibited moderate antibacterial effects against Ralstonia solanacearum | [57] |
| 73 | C23H38N1NaO3 | amorphous powder | Hypomontagnella monticulosa Zg15SU | fresh rhizome of Zingiber griffithii Baker | Indonesia | Showed potent cytotoxic activity | [58] |
| 74 | C20H22N2NaO4 | yellowish powder | Aspergillus lentulus DTO 327G5 | Caenagrion | Shanghai, China | Displayed weak to moderate antibacterial activity | [59] |
| 75 | C19H21O4N2 | white powder | |||||
| 76 | C24H25N3NaO3 | ||||||
| Alkaloids Diketopiperazine derivatives | |||||||
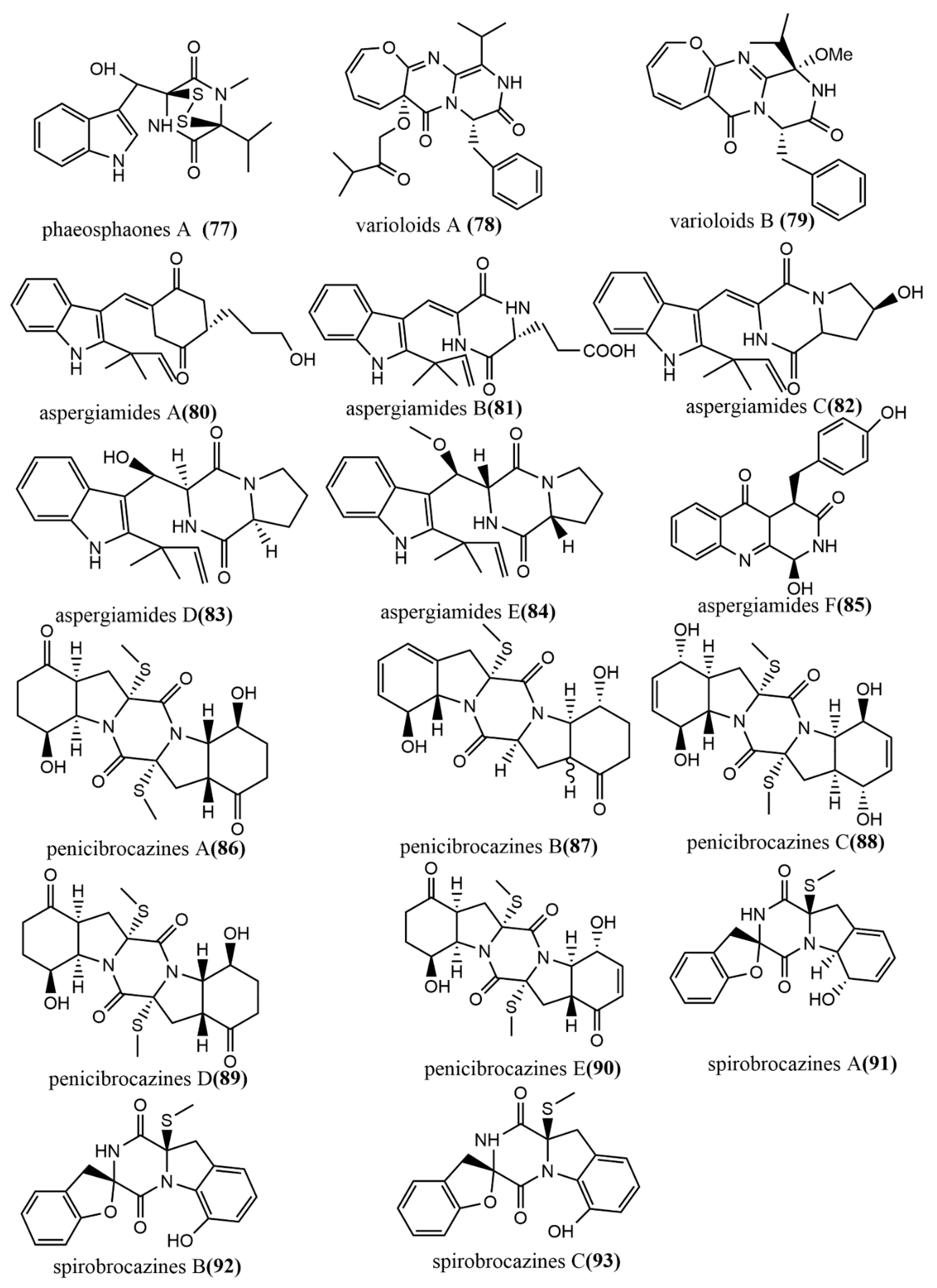
| 77 | C20H27N3O3S2Na | white solid powder | Phaeosphaeria fuckelii | Phlomis umbrosa | Mount Hua, China | Showed strong inhibitory effects on mushroom tyrosinase | [60] |
| 78 | C26H29N3O5 | colorless oil | Paecilomyces variotii EN-291 | Grateloupia turuturu | Qingdao Province, China | Exhibited potent antifungal effects | [61] |
| 79 | C22H23N3O4 | ||||||
| 80 | C21H25O3N3 | yellow powder | Aspergillus sp. 16-5c | leaf of S. apetala | Hainan Island, China | Showed potent to moderate α-glucosidase inhibitory activity (80–81) | [62] |
| 81 | C21H23O4N3 | white powder | |||||
| 82 | C21H23O3N3 | yellow powder | |||||
| 83 | C21H25O3N3 | ||||||
| 84 | C22H27O3N3 | ||||||
| 85 | C18H15O4N3 | white powder | |||||
| 86 | C19H24N2O6S | colorless crystals | Penicillium brocae MA-231 | Avicennia marina | Displayed moderate to high activities against Staphylococcus aureus | [63] | |
| 87 | C19H22N2O5S | yellowish solid | |||||
| 88 | C20H26N2O6S2 | colorless crystals | |||||
| 89 | C20H26N2O6S2 | colorless solid | |||||
| 90 | C20H24N2O6S2 | ||||||
| 91 | C19H18N2O4S | colorless crystals | Penicillium brocae MA-231 | Showed moderate antimicrobial activities against S. aureus and Aeromonas hydrophilia | [64] | ||
| 92 | C19H16N2O4S | ||||||
| 93 | C18H14N2O4 | ||||||
| Alkaloids: Other types of alkaloids | |||||||
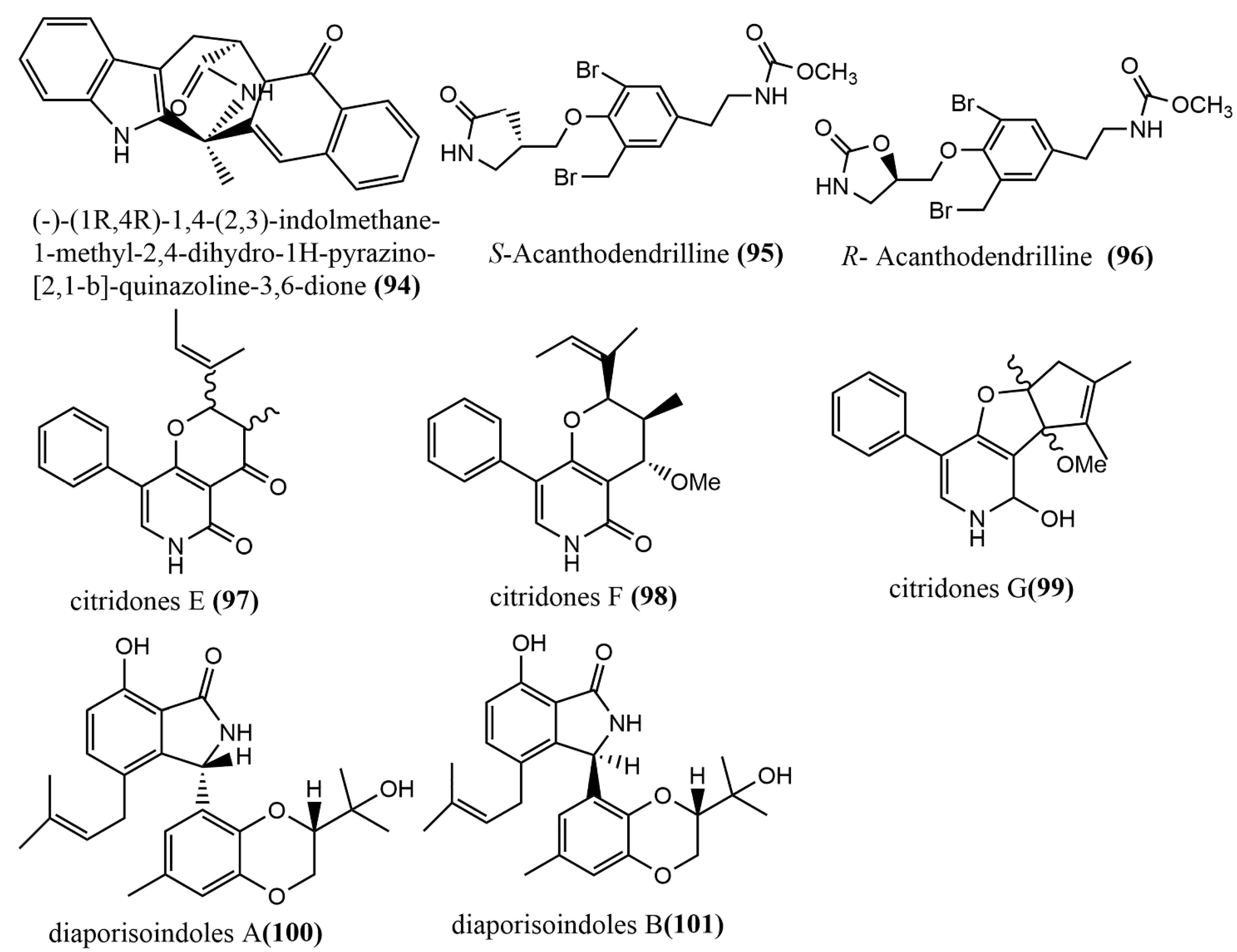
| 94 | C21H16N4O2 | colorless needles | Penicillium vinaceum (X17) | corm of Crocus sativus | Shanghai, China | Showed weak cytotoxic activities against three human tumor cell lines (A549, LOVO, and MCF-7) | [65] |
| 95 | C14H16Br2N2O5 | colorless amorphous powder | Acanthodendrilla sp. | Thailand | Exhibited efficient and selective cytotoxic activities against two human tumor cell lines (H292 and HaCaT) | [66] | |
| 96 | |||||||
| 97 | C19H20NO3 | colorless needles crystal | Penicillium sumatrense GZWMJZ-313 | leaf of Garcinia multiflora | Guizhou, China | Showed moderate to weak antimicrobial activities against Staphylococcus aureus, Pseudomonas aeruginosa, and Escherichia coli | [67] |
| 98 | C20H24NO3 | white powder | |||||
| 99 | C20H21NO3 | ||||||
| 100 | C25H29O5N | white powder | Diaporthe sp. SYSUHQ3 | fresh branch of the mangrove plant Excoecaria agallocha | Showed potent inhibition activity against Mycobacterium tuberculosis protein-tyrosine phosphatase B | [68] | |
| 101 | C25H29O5N | ||||||
| Terpenoids Sesquiterpenoids and their derivatives | |||||||
| 102 | C15H22O3Na | colorless oil | Rhizopycnis vagum Nitaf22 | Nicotiana tabacum | Exhibited high selective cytotoxicity against NCI-H1650 and BGC823 cell lines (115); Showed strong phytotoxic activities against radicle growth in rice seedlings (106–107, 113–114) | [70] | |
| 103 | C15H23O3 | ||||||
| 104 | C15H22O4Na | colorless amorphous solid | |||||
| 105 | C15H20O3Na | colorless oil | |||||
| 106 | C15H24O3Na | ||||||
| 107 | C15H22NaO3 | ||||||
| 108 | C15H21O4 | ||||||
| 109 | C15H25O4 | ||||||
| 110 | C15H22O3Na | ||||||
| 111 | C15H19O5 | colorless amorphous solid | |||||
| 112 | C15H13O5 | brown amorphous solid | Rhizopycnis vagum Nitaf22 Rhizopycnis vagum Nitaf22 | Nicotiana tabacum Nicotiana tabacum | Exhibited high selective cytotoxicity against NCI-H1650 and BGC823 cell lines (115); Showed strong phytotoxic activities against radicle growth in rice seedlings (106–107, 113–114) | [70] | |
| 113 | C15H14O4Na | yellowish oil | |||||
| 114 | C17H18NO4 | greenish-yellow amorphous solid | |||||
| 115 | C14H15NO4 | light-yellowish amorphous solid | |||||
| 116 | C15H22O3 | colorless oil | Trichoderma sp. PR-35 | healthy stem of Paeonia delavayi | Yunnan Province, China | Showed moderate to weak antimicrobial activities against Escherichia coli and Shigella sonnei | [69] |
| 117 | C15H26O2 | ||||||
| 118 | C15H28O3Na | colorless oil | Fusarium proliferatum AF-04 | Lanzhou, China | Displayed weak antimicrobial against Bacillus subtilis, Clostridium perfringens, E. coli, and MRSA | [71] | |
| 119 | C15H26O2Na | ||||||
| 120 | C15 H 20 O5 | colorless oil | Aspergillus sydowii EN-434 | Symphyocladialatiuscula | Qingdao Province, China | Exhibited radical scavenging activity against DPPH | [72] |
| 121 | C25H37O4 | amorphous powder | Ulocladium sp. | Yunnan Province, China | Showed moderate antimicrobial activities against B. subtilis and multi-drug-resistant S. aureus (121–125); Exhibited high selective cytotoxicity against the HepG2 cell line (125) | [73] | |
| 122 | |||||||
| 123 | C26H38O4Na | ||||||
| 124 | C26H40O5Na | ||||||
| 125 | C25H34O3Na | ||||||
| 126 | C15H26O3 | colorless crystals | Trichoderma virens QA-8 | fresh inner root tissue of the grown medicinal herb Artemisia argyi H. Lév. and Vaniot | Hubei Province, China | Showed significant antimicrobial activities against E. coli | [74] |
| 127 | amorphous powder | ||||||
| 128 | colorless oil | ||||||
| 129 | C15H24O2 | ||||||
| 130 | C15H26O3 | colorless waxy solid | |||||
| Terpenoids Diterpenoids | |||||||
| 131 | C20H32O6Na | colorless oil | Diaporthe sp. QYM12 | healthy leaves of Kandelia candel | Hainan Province, China | Showed significant anti-inflammatory effects through the inhibition of NO production | [75] |
| 132 | C21H28O6 | colorless crystals | Phomopsis sp. S12 | seed of Illigera rhodantha | Showed excellent inhibitory effects on the production of IL-1β and IL-18; Effects on the NF-κB signaling pathway | [76] | |
| 133 | C20H26O5 | colorless needle crystal | Phomopsis sp. S12 | seed of Illigera rhodantha | Exhibited anti-inflammatory activity against the production of IL-1b and IL-6 induced by lipopolysaccharide (LPS) in macrophages | [77] | |
| 134 | C20H28O4 | colorless oil | |||||
| 135 | C20H26O6 | ||||||
| 136 | C16H20O5 | colorless needles | Botryosphaeria sp. P483 | Chinese Herbal Medicine Huperzia serrata | Kunming, China | Showed effective antifungal antifungal activities against Gaeumannomyces graminis, Fusarium solani, and Pyricularia oryza (136); Showed weak nematicidal activities | [78] |
| 137 | C16H20O6 | colorless solid | |||||
| 138 | C20H28O6 | white amorphous solid | Smardaea sp. AZ0432 | photosynthetic tissue of the moss Ceratodon purpureus | Chiricahua Mountains of southeastern Arizona, USA | Exhibited selective cytotoxicity | [79] |
| 139 | C20H16O5 | yellowish needles | Pestalotiopsisadusta | Fresh, healthy stems of Clerodendrum canescens | Yandang, Zhejiang Province, China | Demonstrated cytotoxic activities against the HL-60 tumor cell line | [80] |
| 140 | C20H34O2 | colorless needles | Trichoderma atroviride S361 | Cephalotaxus fortunei | Jiande, Zhejiang, China | Bioactivity tests were not performed | [81] |
| 141 | C16H16O5 | colorless needles | Aspergillus wentii EN-48 | unidentified marine brown algal species of the genus Sargassum | Qingdao Province, China | Showed moderate cytotoxic activities against seven human tumor cell lines (NCI-H460, MDA-MB-231, HeLa, MCF-7, SMMC-7721, HepG2, and SW1990) | [82] |
| 142 | C16H16O5 | ||||||
| 143 | C16H24O5 | ||||||
| Terpenoids Triterpenoids | |||||||
| 144 | C30H48O | colorless solid | Scleroderma UFSMSc1 | Eucalyptus grandis | Showed moderate to weak antifungal activities against Candida albicans and Candida parapsolosis | [84] | |
| 145 | |||||||
| 146 | |||||||
| 147 | C29H46O5 | white powder | Acremonium pilosum F47 | pedicel of the Chinese medicinal plant Mahonia fortunei | Qingdao Province, China | Displayed effective antimicrobial activities against S. aureus and B. subtili | [85] |
| 148 | C30H50O6 | yellow amorphous powder | Glomerella sp. F00244 | stem of mason pine | Fujian Province, China | Showed weak cytotoxic activity (148) | [83] |
| 149 | C31H52O6 | white amorphous powder | |||||
| 150 | C30H40O6 | yellowish needle crystals | Penicillium sp. SWUKD4.1850 | healthy branches of Kadsura angustifolia | Yunnan Province, China | Exhibited moderate in vitro cytotoxic activities | [86] |
| 151 | C30H40O6 | white needle crystals | |||||
| 152 | C30H40O6 | white amorphous solid | |||||
| 153 | C30H41O6 | ||||||
| 154 | C32H44O7 | white amorphous powder | |||||
| 155 | C30H42O6 | white powder | |||||
| 156 | C34H46O8 | yellow amorphous solid | |||||
| 157 | C31H44O6 | ||||||
| 158 | C30H46O6 | white amorphous powder | |||||
| 159 | C32H50O5 | white amorphous powder | Hypoxylon sp. 6269 | Artemisia annua | Weak inhibition activity against the HIV-1 integrase (159) | [87] | |
| 160 | C29H44O4 | ||||||
| 161 | C36H55O7 | white amorphous powder | Fusarium sp. | roots of Mentha longifolia | Saudi Arabia | Showed significant antileishmanial activity (161) | [88] |
| 162 | C32H51O5 | ||||||
| 163 | C39H55O7 | ||||||
| 164 | C34H53O6 | colorless powder | Fusarium sp. | roots of Mentha longifolia | Saudi Arabia | Displayed potent cytotoxic activity towards BT-549 and SKOV-3; Showed potent antileishmanial activities against L. donovani promastigotes | [89] |
| 165 | C42H68O7 | white amorphous powder | |||||
| Terpenoids Meroterpenoids | |||||||
| 166 | C18H26O5 | colorless crystal | Guignardia mangiferae A348 | Medicinal Plant Smilax glabra | Luofu Mountain Natural Reservation, Guangdong Province, China | Showed weak cytotoxic activities against MCF-7 cell lines(167,169) | [90] |
| 167 | C17H22O4 | ||||||
| 168 | C18H28O5 | white powder | |||||
| 169 | C17H24O4 | ||||||
| 170 | C25H30O5 | colorless amorphous powder | Emericella sp. TJ29 | root of the plant Hypericum perforatum | the Shennongjia areas of Hubei Province, China | Showed moderate cytotoxic activities against five human tumor cell lines (HL-60, SMMC7721, A549, MCF-7, and SW-480) (172, 173, 175) | [91] |
| 171 | C27H34O6 | white powder | |||||
| 172 | C26H32O6 | colorless crystals | |||||
| 173 | C25H30O6 | colorless crystals | |||||
| 174 | C25H32O6 | white powder | |||||
| 175 | C25H28O6 | colorless crystals | |||||
| 176 | C31H35O9 | yellowish oil | Phyllosticta sp. J13-2-12Y | leaf of Acorus tatarinowii | Guangxi Province, China | Exhibited moderate antimicrobial activities against Staphylococcus aureus 209P, Candida aureus 209P, and Candida albicans FIM709 | [92] |
| 177 | C31H37O9 | ||||||
| 178 | C31H34O9 | ||||||
| 179 | C27H32O9 | white powder | Co-culture Talaromyces purpurogenus H4 and Phanerochaete sp. H2 | Handroanthus impetiginosus | Alfenas, Minas Gerais, Brazil. | Showed moderate trypanocidal activity against T. cruzi | [93] |
| Lactones | |||||||
| 180 | C13H22O3 | colorless gum | Talaromyces assiutensis JTY2 | leaf of Ceriops tagal | South China Sea, China | Showed moderate cytotoxic activities against three human cancer cell lines (HeLa, MCF-7, and A549) | [94] |
| 181 | C21H34O5 | yellow oil | Penicillum polonicum | fruits of Camptotheca acuminata Decne | Wuhan, China | Showed effective glucose uptake activity on rat skeletal muscle myoblast L6 (181); Significantly promoted GLUT4 translocation in L6 cells | [95] |
| 182 | C16H28O5 | light red oil | |||||
| 183 | C24H33NO8 | colorless crystal | Chaetomium cupreum | Anemopsis californica | New Mexico, U.S.A. | Showed weak antimicrobial activity against S. aureus | [96] |
| 184 | Xylaria curta 92092022 | Taiwan, China | Showed moderate antimicrobial activities against Pseudomonas aeruginosa and Staphylococcus aureus; Displayed strongly inhibited lettuce seed germination | [97] | |||
| 185 | C24H34O5 | white powder | Lasiodiplodia theobromae ZJ-HQ1 | healthy leaves of the marine mangrove Acanthus ilicifolius | South China Sea, China | Exhibited inhibitory effects on lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophage cells; Showed moderate inhibitory activity against α-glucosidase | [98] |
| 186 | C8H10O3 | Aspergillus pseudonomiae J1 | Euphorbia umbellata (Pax) Bruyns (Euphorbiaceae) | Bahia, Brazil | Showed moderate to weak anti-trypanosomal activity | [99] | |
| 187 | C10H12O4 | ||||||
| 188 | C10H12O3 | ||||||
| Anthraquinones, quinones, and related glycosides | |||||||
| 189 | C20H19O7 | Brilliant yellowish oil | Acremonium vitellinum | Acanthus ilicifolius Linn | Qingdao Province, China | Showed moderate insecticidal activities against the third-instar larvae of Helicoverpa ar-migera | [100] |
| 190 | C20H16O7 | yellow solid | |||||
| 191 | C22H21O7 | ||||||
| 192 | C18H18O8Na | yellow amorphous powder | Phomopsis sp. | Paris polyphyllavar | Yunnan Province, China | Showed significant cytotoxic activities against A549 and PC3 cell lines | [101] |
| 193 | C21H20O10 | red amorphous powder | Eurotium cristatum EN-220 | Sargassum thunbergii | Qingdao Province, China | Showed weak antimicrobial activity against E. coli only; Showed moderate fatal activity against brine shrimp larvae | [102] |
| 194 | C18H15O6 | orange yellow powder | Aspergillus versicolor | Halimeda opuntia | South Sinai, Egypt | Weak inhibitory activity against hepatitis C virus NS3/4A protease | [103] |
| 195 | C12H11O4 | red powder | |||||
| 196 | C16H11O5 | orange powder | |||||
| 197 | C16H21O7 | red powder | Stemphylium globuliferum | healthy stems of Mentha pulegium | Beni Mellal, Morocco | Showed significant inhibition of proliferation of K562 and A549 cells | [104] |
| Steroids | |||||||
| 198 | C27H40O3 | white crystals | Phoma sp. SYSU-SK-7 | Guangdong Province, China | Exhibited inhibitory effects on lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophage cells; Showed moderate inhibitory activity against α-glucosidase | [105] | |
| 199 | C28H41O3 | white solid | |||||
| 200 | C38H64O4 | white amorphous powder | Fusarium sp. | Mentha longifolia | Egypt | Showed moderate cytotoxic activity against human colorectal cancer cell line HCT 116 | [106] |
| 201 | C28H40O4 | Phomopsis sp. | Aconitum carmichaeli | Huize County, Yunnan Province, China | Showed weak antifungal activities against C. albicans and F. avenaceum | [107] | |
| 202 | C30H48O4 | ||||||
| Other types of compounds | |||||||
| 203 | C20H22O3 | yellow oil | Aspergillus sp. | root of Tripterygium wilfordii | Wuhan, China | Showed significant inhibition of LPS-induced IL-1β, TNF-α, and NO production in RAW264.7 cells | [108] |
| 204 | C24H26O6 | ||||||
| 205 | C24H26O6 | colorless oil | |||||
| 206 | C23H24O6 | ||||||
| 207 | C23H24O5 | brown powder | Aspergillus flavipes Y-62 | stems of plant Suaeda glauca Bunge | Zhoushan coast, Zhejiang province, China | Exhibited antimicrobial activities against the Gram-negative pathogens Pseudomonas aeruginosa and Klebsiella pneumoniae | [109] |
| 208 | C23H21O5 | white powder | Microsphaeropsis sp. | Showed effective antimicrobial activities against B. megaterium and E. coli | [110] | ||
| 209 | C15H20O2 | brown amorphous powder | Emericella sp. XL029 | leaf of Panax notoginseng | Shijiazhuang, Hebei province, China | Showed potent antifungal activities against six tested plant pathogenic fungi (Rhi-zoctorzia solani, Verticillium dahliae Kleb, Helminthosporium maydis, Fusarium oxysporum, Fusarium tricinctum, and Botryosphaeria dothidea) | [111] |
| 210 | C16H20O4 | ||||||
| 211 | C18H18O6Cl | yellow powder | Diaporthe perseae sp. | stem of Chinese mangrove Pongamia pinnata | Hainan city, China | Showed significant DPPH and ABTS radical scavenging activities | [112] |
| 212 | C7H7O4 | colorless flake crystal | Aspergillus tubingensis DS37 | Decaisnea insignis (Griff.) Hook. f. and Thomson | Showed significant inhibition activities against Fusarium graminearum and Streptococcus lactis | [113] | |
| 213 | C13H15NO4Na | amorphous powder | Nigrospora sphaerica ZMT05 | Oxya chinensisThunber | Guangdong Province, China. | Showed significant α-glucosidase inhibitory activity | [114] |
| 214 | C9H14O2 | colorless oil | Co-culture Aspergillus terreus EN-539 & Paecilomyces lilacinus EN-531 | Laurencia okamurai | Qingdao, China | Showed weak antimicrobial activities against Physalospora piricola and Staphylococcus aureus | [115] |
| 215 | C11H11BrO4 | white amorphous powder | Lachnum palmae | Przewalskia tangutica | Exhibited potent to weak antimicrobial activities against Cryptococcus neoformans, Penicillium sp., Candida albicans, Bacillus subtilis, and Staphy-lococcus aureus (215–220); Showed moderate inhibitory effects on NO production in LPS-induced RAW 264.7 cells (215,219) | [116] | |
| 216 | |||||||
| 217 | C12H13BrO4 | ||||||
| 218 | C10H9BrO4 | ||||||
| 219 | C11H11BrO5 | ||||||
| 220 | C11H11ClO4 | ||||||
| No. | Endophytic Fungus | Host Plant | Culture Conditions | Secondary Metabolites | Yield | Ref. |
|---|---|---|---|---|---|---|
| 1 | Hansfordia biophila | Hedychium acuminatum Roscoe | Inoculated in potato glucose broth (PDB) medium and shaken at 120 rpm at 25 °C for 7 days. | Tannin | 41.6 μm·mL−1 | [121] |
| 2 | Aspergillus terreus | Ficus elastica | Inoculated into PDB medium and incubated at 30 °C for 20 days on a rotatory shaker incubator at 140 rpm. | Camptothecin | 320 μg/L | [122] |
| 3 | Guignardia mangiferae HAA11 | Taxus x media | Inoculated into (PDB) medium and incubated at 200 rpm at 28 °C for 5 days. | Paclitaxel | 720 ng/L | [123] |
| 4 | Papulasora sp.S6 | Phellodendron amurense Rupr | Mutagenesis by UV, X-ray rays, and NaNO2, inoculated in PDB medium, and shaken at 100 rpm at 28 °C for 7 days. | Berberine | 12.28 mg/L | [124] |
| 5 | Actinoplanes teichomyceticus | Improvement of the output of teicoplanin by genome shuffling; Inoculated teicoplanin medium and cultured at 28 °C for 15–20 days. | Teicoplanin | 3016 μm·mL−1 | [125] | |
| 6 | Phialocephala fortinii | Podophyllum peltatum | Inoculated in Sabouraud’s dextrose agar (SDA) and cultured at 23 °C for 4–6 weeks. | Podophyllotoxin | 189 µg/L | [126] |
| 7 | Entrophospora infrequens RJMEF001 | Nothapodytes foetida | Inoculated into wheat bran containing Sabouraud’s broth, and incubation was carried out at 28 ± 2 °C for 28 days. | Camptothecin | 503 ± 25 μg/100 g dry cell mass (in Sabouraud broth) | [127] |
| 8 | Epicoccum nigrum SZMC 23769 | Hypericum perforatum | Fungal isolates were grown in potato dextrose broth (PDB) for 7 days at 25 °C. | Hypericin, Emodin | 117.1 μg/mL, 87.7 μg/mL | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.; Okyere, S.K.; Wang, S.; Wang, J.; Xie, L.; Ran, Y.; Hu, Y. Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies. J. Fungi 2022, 8, 205. https://doi.org/10.3390/jof8020205
Wen J, Okyere SK, Wang S, Wang J, Xie L, Ran Y, Hu Y. Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies. Journal of Fungi. 2022; 8(2):205. https://doi.org/10.3390/jof8020205
Chicago/Turabian StyleWen, Juan, Samuel Kumi Okyere, Shu Wang, Jianchen Wang, Lei Xie, Yinan Ran, and Yanchun Hu. 2022. "Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies" Journal of Fungi 8, no. 2: 205. https://doi.org/10.3390/jof8020205