
Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China



The Key Laboratory for Silviculture, Beijing Forestry University, Conservation of Ministry of Education, Beijing 100083, China
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(4), 377; https://doi.org/10.3390/jof8040377
Submission received: 13 March 2022
/
Revised: 3 April 2022
/
Accepted: 5 April 2022
/
Published: 7 April 2022
(This article belongs to the Special Issue Ascomycota: Diversity, Taxonomy and Phylogeny)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Species of Cytospora are considered important plant pathogens of a wide range of plant hosts, especially Salicaceae plants. Salix (Salicaceae, Malpighiales) has been widely cultivated in China because of its strong ecological adaptability, fast growth, and easy reproduction. In this study, a total of eight species of Cytospora were discovered on Salix in China, including C. ailanthicola, C. alba, C. chrysosperma, C. gigaspora, C. nivea, C. paracinnamomea, C. rostrata, and C. sophoriopsis. Among them, C. alba and C. paracinnamomea were identified as novel species based on morphology and phylogenetic analyses of ITS, act, rpb2, tef1-α, and tub2 gene sequences and were confirmed as pathogens of willow canker disease by pathogenicity tests. The mycelial growth rates of strains from these two novel species (C. alba and C. paracinnamomea) had optimum temperatures of 21 to 22 °C and an optimum pH value of 5 to 6. The effectiveness of six carbon sources on the mycelial growth showed that fructose and maltose had the highest influence. Cytospora species richness was significantly positively correlated with dry and wet areas. This study represents a significant evaluation of Cytospora associated with willow canker disease in China and provides a theoretical basis for predicting the potential risk of willow canker disease.
1. Introduction
Willow (Salix) trees have been widely cultivated in China because of their strong ecological adaptability, fast growth, easy reproduction, and short rotation period [1]. In terms of horizontal distribution, they are distributed all over the country, with the most concentrated distribution in the northeast, northwest, and southwest regions. These regions have higher elevations or latitudes, and the horizontal distribution of species is extremely rich [2]. However, a large number of willow trees suffer from diseases caused by several fungal pathogens, such as over 20 species of Cytospora causing canker disease [3,4] (https://nt.arsgrin.gov/fungaldatabases, accessed on 6 April 2022); over 50 species of Melampsora causing rust disease [5,6,7,8,9] (https://nt.ars-grin.gov/fungaldatabases, accessed on 6 April 2022); Colletotrichum salicis causing anthracnose disease [10]; and Rhytisma filamentosum causing tar-spot disease [11,12]. Among various diseases that affect willow trees, canker and dieback disease caused by Cytospora are the main branch and stem diseases in China, especially in the north (Figure 1) [4,13,14,15,16]. The pathogens in Cytospora are host-dominant, and usually invade weak trees. There are many factors that cause tree weakness, such as meteorological factors including temperature, humidity, rainfall, sunshine, airflow, etc. Meanwhile, tree age, tree species, slope aspect, soil, nursery management, planting density, and stand tending management technology are also factors that cause Cytospora canker disease. These factors influence each other and work together [17].
Symptoms of Cytospora canker disease associated with willow trees vary according to the stage of disease development. The disease infects the inner bark and causes the sapwood to sink slightly and discolor [18]. Several prominent fruiting bodies form and are buried or semi-buried under the bark, solitary, or in clusters [4]. Under moist conditions, the conidia emerge from the conidiomata in the form of colored and coiled tendrils [4]. Usually, these conidiomata develop in the cankers on bark but may also exist in healthy plant tissue and can be isolated from sound bark, xylem, and leaves of many tree species [19,20]. The genus Cytospora (Cytosporaceae, Diaporthales) was established by Ehrenberg [21], which comprises important phytopathogenic, saprobic, and endophytic fungi [4,18]. Species of Cytospora inhabit a wide variety of hosts that include economically and ecologically important trees (e.g., Elaeagnaceae, Juglandaceae, Rosaceae, Salicaceae, Ulmaceae) [22,23,24,25,26]. About 150 species of Cytospora in total have been discovered on dieback and stem canker in over 130 species of woody hosts [18,27,28,29,30,31,32,33].
Currently, use of multiphase approaches such as morphological and phylogenetic analyses to define species of Cytospora has been proposed [4]. In morphology, presence or absence of conceptacle, quantity and arrangement of locule(s), shape and size of conidiophores, and conidial size are significantly taxonomic [3,4]. In phylogeny, the current studies use multigene approaches such as the internal transcribed spacer (ITS), partial actin (act), RNA polymerase II subunit (rpb2), translation elongation factor 1-α (tef1-α), and beta-tubulin (tub2) to perform phylogenetic analysis [4,29,32,33].
Over 20 Cytospora species infect willow trees all over the world (https://nt.ars-grin.gov/fungaldatabases, accessed on 6 April 2022). An increase in the number of Cytospora species isolated from willow cankers has also been reported in China recently. Fan et al. [22] concluded that six species of Cytospora from Salix were observed in northern China: C. chrysosperma, C. fugax, C. leucosperma, C. nivea, C. populina, and C. rostrata. Wang et al. [3] identified six Cytospora species occurring on poplar and willow: C. atrocirrhata, C. chrysosperma, C. davidiana, C. fugax, C. kantschavelii, and C. translucens. The diversity of Cytospora on willows deserves further exploration for better disease management. There is an urgent need for systematic studies and re-evaluation to facilitate the identification of Cytospora from Salix in China. The objectives of this study were (1) to determine species of Cytospora from willow trees in China; (2) to confirm the pathogenicity of two new Cytospora species; (3) to determine the effects of temperature, pH, and carbon source on the mycelial growth of the new Cytospora species; and (4) to evaluate the geographical distribution of Cytospora species associated with willow canker disease in China.
2. Materials and Methods
2.1. Specimens and Strains
Sixty-two specimens were collected from diseased branches or twigs of Salix matsudana, Salix psammophila, and Salix cupularis distributed in China. Sixty-six Cytospora strains were isolated by removing a mucoid spore mass from conidiomata and/or ascomata. The suspension was spread over the surface of potato dextrose agar (PDA) (potato, 200 g; glucose, 20 g; agar, 20 g; distilled water, to complete 1000 mL) media, which were incubated at 25 °C in darkness until spores germinated. Hyphal tips were removed to a new PDA plate twice to obtain a pure culture. Specimens were preserved in the Museum of Beijing Forestry University (BJFC). Strains were deposited in the China Forestry Culture Collection Centre (CFCC).
2.2. Microscopic Observations
The ascomata and conidiomata formed on the branches and/or twigs of Salix were photographed using the Leica stereomicroscope (M205) (Leica Microsystems, Wetzlar, Germany). Over 30 conidiomata (and/or ascomata) and 50 conidia and/or ascospores were selected randomly to measure their lengths and widths using a Nikon Eclipse 80i microscope (Nikon Corporation, Tokyo, Japan) equipped with a Nikon digital sight DS-Ri2 high-definition color camera with differential interference contrast (DIC). Cultural characteristics of strains plated onto PDA and incubated at 25 °C in darkness were recorded after seven and thirty days according to the color charts of Rayner [34].
2.3. DNA Extraction, Amplification, and Phylogeny
Total genomic DNA was harvested from axenic cultures on PDA using CTAB method [35]. PCR amplifications and sequencing of ITS, act, rpb2, tef1-α, and tub2 genes were performed. The PCR conditions and primers are listed in Table S1. PCR products were electrophoresed in 1% agarose gel and the DNA was sequenced by the Tsingke Biotechnology Company Limited. A preliminary identification of strains was based on BLAST using the ITS sequences to confirm the strains obtained in the current study grouped in the genus Cytospora. Twenty-two representative strains were selected for the following analyses excluding the strains having parallel ITS sequence, morphological features, host, and location for each species. The sequences generated from this study were supplemented with other sequences obtained from GenBank (Table S2) based on recent publications of Cytospora [4,26,32]. For the five individual datasets (ITS, act, rpb2, tef1-α, and tub2), sequences alignments were made using MAFFT v. 6.0 [36] and adjusted using MEGA v. 6.0 [37]. Ambiguous regions were excluded from alignments.
Phylogenetic analyses were carried out with maximum parsimony (MP), maximum likelihood (ML), and Bayesian Inference (BI) analyses. MP analysis was computed with PAUP v. 4.0b10 [38] using a heuristic search option of 1000 random-addition sequences with a tree bisection and reconnection (TBR) branch swapping algorithm. ML analysis was computed with GTR+G+I model of site substitution using PhyML v. 3.0 [39] following the instruction of Fan et al. [4]. BI analysis was computed using the best-fit evolutionary models for each partitioned locus estimated in MrModeltest v. 2.3 [40], with a Markov Chain Monte Carlo (MCMC) algorithm in MrBayes v. 3.1.2 [41]. Phylograms were viewed in Figtree v. 1.3.1 [42]. Sequence data were deposited in GenBank. The multilocus sequence alignment and the trees obtained were deposited in TreeBASE (study ID S29245).
2.4. Pathogenicity Test
After species identification, healthy cutting plants of Salix matsudana inoculated with two novel species (C. alba and C. paracinnamomea) to determine the relative pathogenicity. For mycelial inoculation, the ex-holotype strains (C. alba: CFCC 55462; C. paracinnamomea: CFCC 55453) were cultured for a duration of five days at room temperature (25 °C) in darkness. Cuttings of plants were scalded using a 5-mm-diameter sterile inoculation loop, putting a PDA mycelial plug on each burn wound. Moistened cotton was placed directly above each disc, and all were covered with parafilm. Ten inoculation dots were set for each isolate. Ten replicates were inoculated using uncolonized PDA plugs to serve as controls. After a week, the parafilm and cotton wool were removed. The inoculated plants were maintained in the field to observe the symptoms of willow canker and expansion of lesion length. Re-isolation and morphological observation of fungal pathogens were performed to verify Koch’s postulates [43].
2.5. Temperature, pH, and Carbon Source Tests
Ex-holotype strains (C. alba: CFCC 55462; C. paracinnamomea: CFCC 55453) were selected to evaluate the effects of temperature, pH, and carbon source on mycelial growth of Cytospora. For the temperature variation test, a 5-mm-diameter mycelial plug was inoculated onto PDA media in triplicate and incubated in the dark at 0–40 °C with a 5 °C gradient. For the pH variation test, the PDA was adjusted with 1 mol/L NaOH and 1 mol/L HCl to obtain pH values ranging from 3.0 to 11.0 at an interval of 1.0. A 5-mm-diameter mycelial plug was inoculated in the medium in triplicate and the cultures were incubated in the dark at 25 °C [44,45]. For the investigation of the utilization of carbon sources, the strains were incubated in triplicate in the dark at 25 °C and the PDA was used as the base medium replacing glucose by the tested carbon sources. The 20 g of glucose was replaced by 20 g of fructose, galactose, maltose, sucrose, and xylose [44,45]. Measurements of colony diameter were made every 24 h for four days. Data were analyzed using IBM SPSS Statistics v. 19.0 by one-way analysis of variance (ANOVA). Linear Regression was used to estimate optimum growth temperature and pH value. Figures were output using Microsoft Excel.
2.6. Geographic Distribution
The administrative map of China used in this study includes 34 provincial administrative regions (http://www.tianditu.gov.cn, accessed on 1 March 2022). The dry and wet conditions of China’s land surface followed Wu et al. [46] to divide China into four dry and wet regions, namely, humid, semi-humid, semi-arid, and arid. Salix species distribution information was mainly obtained from the Flora of China [47], the Sylva Sinica [48], the National Specimen Information Infrastructure (NSII) (http://www.nsii.org.cn/, accessed on 6 April 2022), and the Atlas of Woody Plants in China [49]. Cytospora species distribution information was mainly obtained from the current study (66 strains, 8 species), original records (633 records, 7 species) [13,50,51], and recent publications (63 strains, 11 species) [3,4,16,22]. A total of 13 geographically distributed Cytospora species were studied, including eight collected in this study (C. ailanthicola, C. alba, C. chrysosperma, C. gigaspora, C. nivea, C. paracinnamomea, C. rostrata, and C. sophoriopsis) and five previously reported (C. atrocirrhata, C. fugax, C. leucosperma, C. populina, and C. translucens). Statistical tests were performed with Poisson regression in R [52].
3. Results
3.1. Phylogenetic Analyses
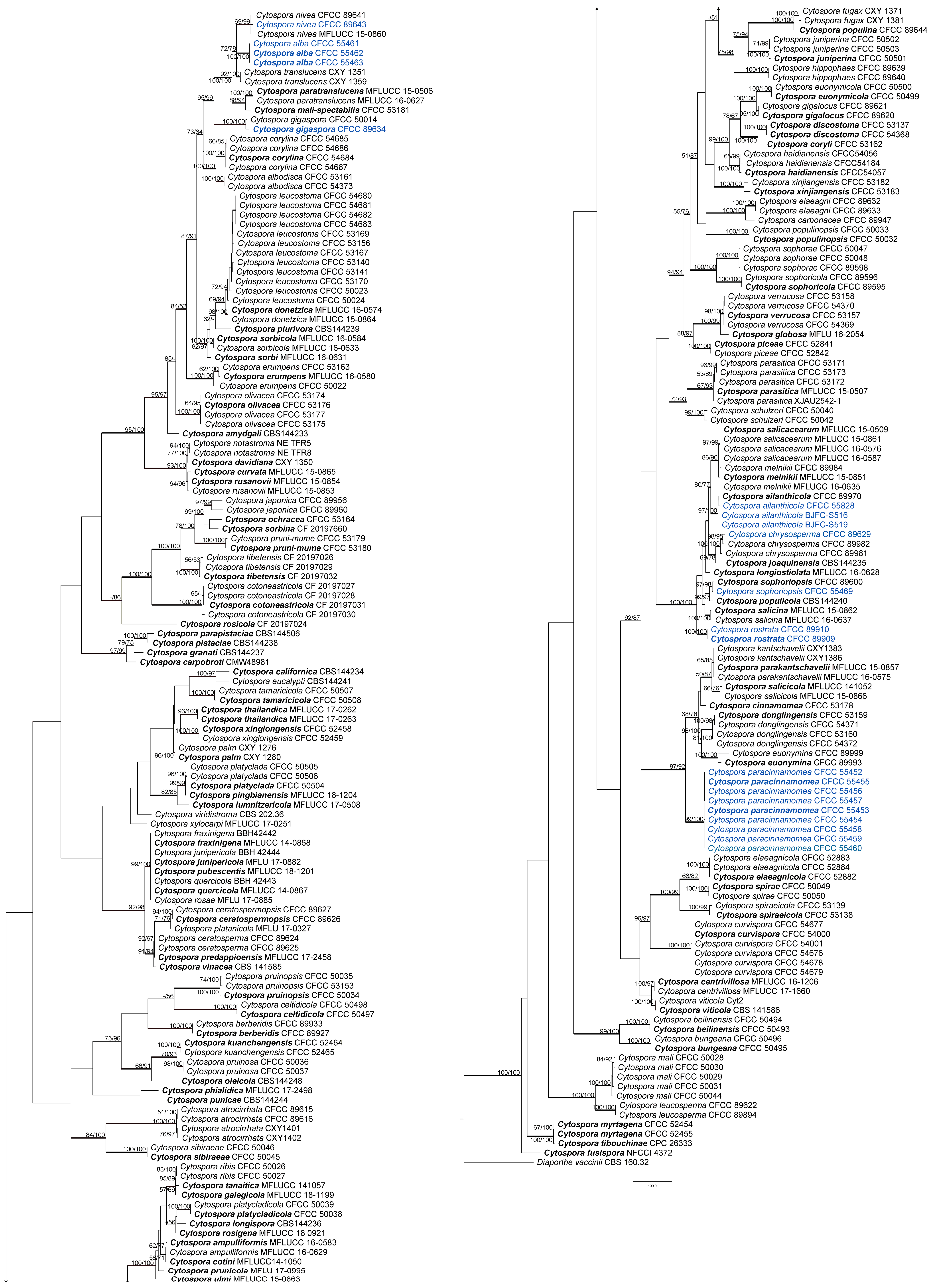
The combined matrix of five genes of Cytospora included 3189 characters with gaps (ITS, act, rpb2, tef1-α, and tub2, included 604, 351, 726, 843, and 665, respectively). The concatenated alignment comprised sequences from 255 strains and Diaporthe vaccinii CBS 160.32 was selected as the outgroup. The alignment contained 189 parsimony-uninformative characters, 1445 characters were variable and parsimony-informative. MP analysis generated 200 equally parsimonious trees with similar clade topologies, one of which was presented in Figure 2 (TL = 9814, CI = 0.305, RI = 0.835, RC = 0.254). In ML analysis, the matrix had 1967 distinct alignment patterns. Estimated base frequencies are as follows: A = 0.244805, C = 0.285887, G = 0.239268, T = 0.230041; substitution rates: AC = 1.258949, AG = 3.497398, AT = 1.467433, CG = 0.961514, CT = 6.191047, GT = 1.000000; gamma distribution shape parameter: α = 0.366687. For BI analysis, the best-fit model of nucleotide evolution was deduced on the AIC (ITS and act: GTR+I+G; rpb2 and tef1-α: TrN+I+G; tub2: HKY+I+G). ML and Bayesian analyses did not significantly differ from the MP analysis.
The 21 strains obtained in this study clustered in eight clades based on the combined ITS/act/rpb2/tef1-α/tub2 tree as well as each gene tree except for ITS tree (Figures S1–S5 and Figure 2). The three strains CFCC 55461–55463 grouped as one clade were phylogenetically distinct from all Cytospora species with a high statistical support (MP/ML/BI = 100/100/1). Nine strains (CFCC 55452–CFCC 55460) were grouped together (MP/ML/BI = 99/100/1) and distinguished from other Cytospora species. These two separate clades are herein described as novel species.
3.2. Taxonomy
Cytospora species on Salix herein include two novel species (C. alba and C. paracinnamomea) and six known species collected from Salix (C. ailanthicola, C. chrysosperma, C. gigaspora, C. nivea, C. rostrata, and C. sophoriopsis). The asexual morph of C. alba as well as the sexual and asexual morph of C. paracinnamomea were described in this study.
Cytospora alba L. Lin and X.L. Fan, sp. nov. (Figure 3)
MycoBank: MB 841281
Etymology: The name reflects the white ectostromatic disc.
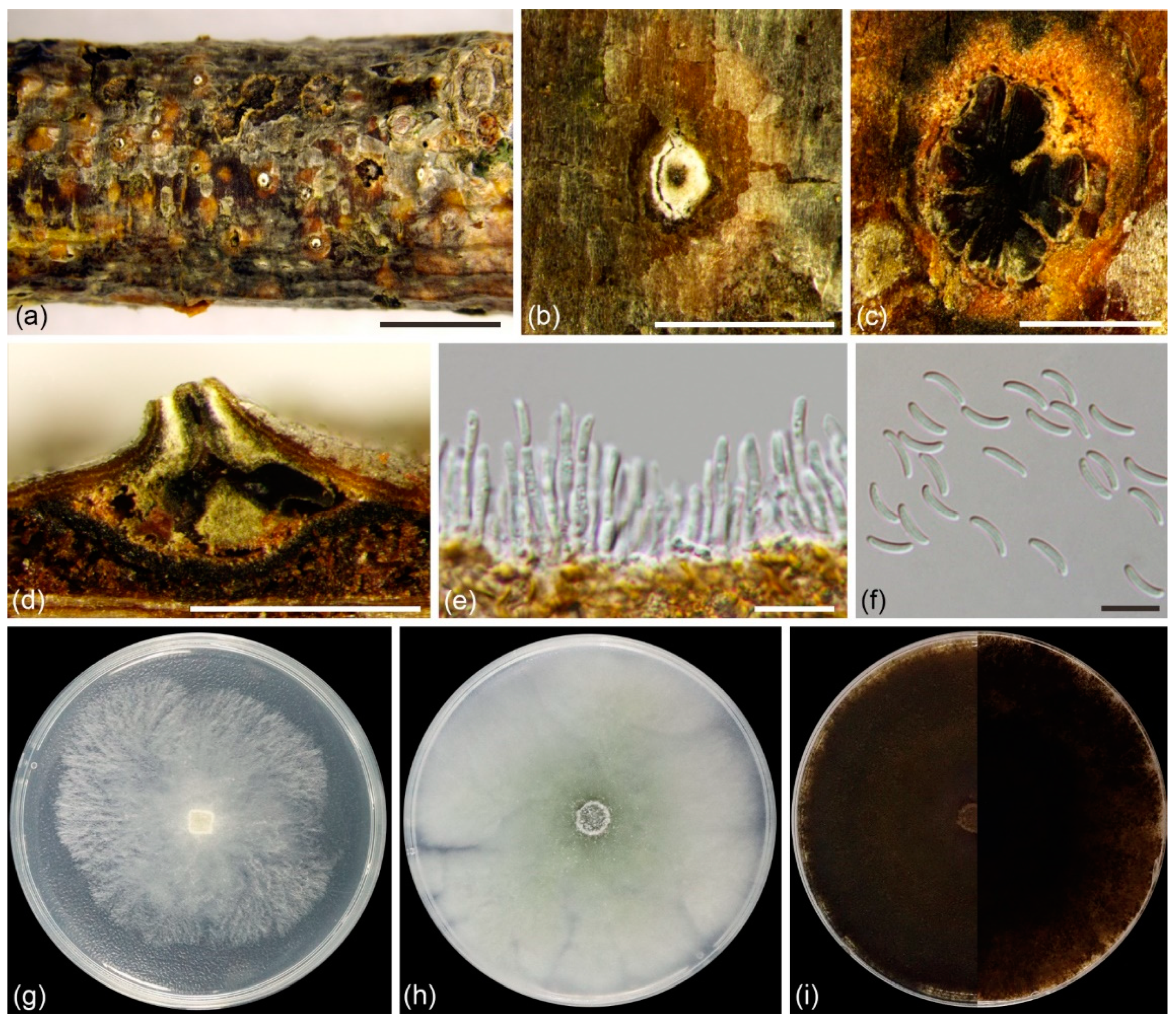
Description: Asexual morph: Conidiomata labyrinthine cytosporoid, immersed in bark, erumpent when mature, discoid to conical, 990–1930 µm in diam, with multiple locules. Conceptacle present. Ectostromatic disc white to primrose, circular to ovoid, 270–570 µm in diam, with single ostiole per disc in center. Ostiole circular to ovoid, hazel to olivaceous at the same level as disc surface, 93–216 µm in diam. Locules complex with irregular shapes, subdivided by invaginations with common walls. Conidiophores hyaline, unbranched, or occasionally branched at bases, 10–18 × 1.0–2.5 µm (av. = 13.8 ± 1.6 × 1.7 ± 0.2 µm, n = 30). Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical, 3.0–6.5 × 1.0–2.0 µm (av. = 5.1 ± 0.9 × 1.5 ± 0.2 µm, n = 30). Conidia hyaline, unicellular, eguttulate, elongate-allantoid, 6.0–9.0 × 1.5–2.0 µm (av. = 8.2 ± 0.5 × 1.7 ± 0.1 µm, n = 50). Sexual morph: not observed.
Culture characteristics: Cultures on PDA initially white, growing to 6.5 cm after 2 days, entirely covering the 9-cm Petri dish after 3 days and becoming greyish yellow green after 7 days. Colonies pale mouse grey and flat with a uniform texture after one month.
Specimens examined: CHINA, Gansu Province: Lanzhou City, Yongdeng County, 102°49′27″ E, 36°44′20″ N, from branches of Salix matsudana, X.L. Fan, N. Jiang, and C. Peng, 20 October 2020 (holotype BJFC CF20201001, ex-holotype living culture CFCC 55462; ibid., living culture CFCC 55461). Gansu Province: Lanzhou City, Yongdeng County, 102°49′26.7″ E, 36°44′23.2″ N, from branches of Salix matsudana, X.L. Fan, N. Jiang, and C. Peng, 20 October 2020 (paratype BJFC CF20201011, ex-paratype living culture CFCC 55463).
Notes: Cytospora alba is associated with canker disease of Salix matsudana in China. It is close to C. mali-spectabilis in the phylogenetic diagram (Figure 2). It can be distinguished from C. mali-spectabilis by smaller conidia (6.0–9.0 × 1.5–2.0 vs. 9.0–10.0 × 1.5–2 µm in C. mali-spectabilis) and the lack of a central column in the conidiomata [26]. Cytospora alba has black conceptacle surrounding the asexual stroma, whereas C. mali-spectabilis lacks a conceptacle [26]. Furthermore, C. alba has multiloculate conidiomata sharing a larger single ostiole (93–216 vs. 60–84 µm) than C. mali-spectabilis [26].
Cytospora ailanthicola X.L. Fan and C.M. Tian, Persoonia 45: 13, 2020.
Descriptions: See Fan et al. [4].
Materials examined: CHINA, Gansu Province: Gannan Autonomous Prefecture, Lintan County, 103°21′20.23″ E, 34°41′33.9″ N, from branches of Salix matsudana, 5 August 2012, X.L. Fan (BJFC CF20210502, living culture CFCC 55828). Ningxia Autonomous Region: Zhongwei City, 105°02′17.41″ E, 37°28′43.00′′ N, from branches of Salix matsudana, 3 June 2012, X.L. Fan, BJFC-S516. Ningxia Autonomous Region: Yinchuan City, 106°12′28.52″ E, 38°14′07.11″ N, from branches of Salix matsudana, 5 June 2012, X.L. Fan, BJFC-S519.
Notes: Cytospora ailanthicola was introduced by Fan et al. [4] causing canker disease on Ailanthus altissima in China. In the current study, we collected specimens of this species associated with twigs and branches of Salix matsudana. Although C. ailanthicola was closely related to C. salicacearum and C. melnikii in the phylogram, they can be distinguished by morphological characters. Morphologically, C. ailanthicola has multiple locules subdivided frequently by invaginations with common walls, whereas C. salicacearum and C. melnikii have conidiomata with 1–2 locule(s) and a single locule, respectively [4,22]. In addition, this is the first time that C. ailanthicola has been reported from willow tree.
Cytospora chrysosperma (Pers.) Fr., Syst. Mycol. 2: 542, 1823.
Basionym. Sphaeria chrysosperma Pers., Neues Mag. Bot. 1: 82, 1794.
Synonym. Naemaspora chrysosperma (Pers.) Pers., Observ. Mycol. (Lipsiae) 1: 80, 1796.
=Valsa sordida Nitschke, Pyrenomyc. Germ. 2: 203, 1870.
Descriptions: See Fan et al. [22].
Material examined: CHINA, Shaanxi Province: Yulin City, Airport East Road, 109°39′54.73″ E, 38°19′21.16″ N, from branches of Salix psammophila, 1 August 2013, X.L. Fan (BJFC-S975, living culture CFCC 89629).
Notes: Cytospora chrysosperma, the type species of Cytospora [53], is considered a significant plant pathogen causing dieback and canker disease [4,18]. Previous studies have concluded that many species of Cytospora have a similar morphology (such as conidiomata cytosporoid and conceptacle absent) as C. chrysosperma, which indicated that these species should be regarded as a species complex [4,18]. Recently, Fan et al. [4] revisited ten related Cytospora species as belonging to the C. chrysosperma complex based on multigene analyses.
Cytospora gigaspora C.M. Tian, X.L. Fan, and K.D. Hyde, Phytotaxa 197: 232, 2015.
Description: See Fan et al. [24].
Material examined: CHINA, Shaanxi Province: Yulin City, Airport East Road, 109°39′54.73″ E, 38°19′21.16″ N, on twigs and branches of Salix psammophila, 1 August 2013, X.L. Fan (BJFC-S975, living culture CFCC 89634).
Notes: Cytospora gigaspora was originally observed on twigs and branches of Salix psammophila in Shaanxi Province [24]. After that, Fan et al. [4] added specimens from Shanxi. Cytospora gigaspora differs from Cytospora nivea by having flat locules and larger conidia [4].
Cytospora nivea (Hoffm.) Sacc., Michelia 2: 264, 1881.
Basionym. Sphaeria nivea Hoffm., Veg. Crypt. 1: 28, 1787.
Synonyms. Leucostoma niveum (Hoffm.) Höhn., Mitt. Bot. Inst. Tech. Hochsch. Wien 5: 58, 1928.
=Valsa nivea (Hoffm.) Fr., Summa Veg. Scand., Section Post. (Stockholm): 411, 1849.
Descriptions: See Fan et al. [4].
Material examined: CHINA, Shaanxi Province: Yulin City, Hongshi Gorge, 109°42′00.69″ E, 38°19′32.43″ N, on twigs and branches of Salix psammophila, 29 July 2013, X.L. Fan (BJFC-S979, living culture CFCC 89643).
Notes: Cytospora nivea commonly occurred along with C. chrysosperma in association with hosts in the Salicaceae [4,22]. Cytospora nivea has a unique characteristic of immersed conidiomata with black conceptacle, multiple locules, and gelatinous conidial tendrils under wet conditions [22]. Cytospora nivea have been recorded in Gansu, Ningxia, Shaanxi, and Xinjiang of China [4,13,22,51].
Cytospora paracinnamomea L. Lin and X.L. Fan sp. nov. (Figure 4)
MycoBank: MB 841282
Etymology: The name reflects the similar culture characteristics with C. cinnamomea.
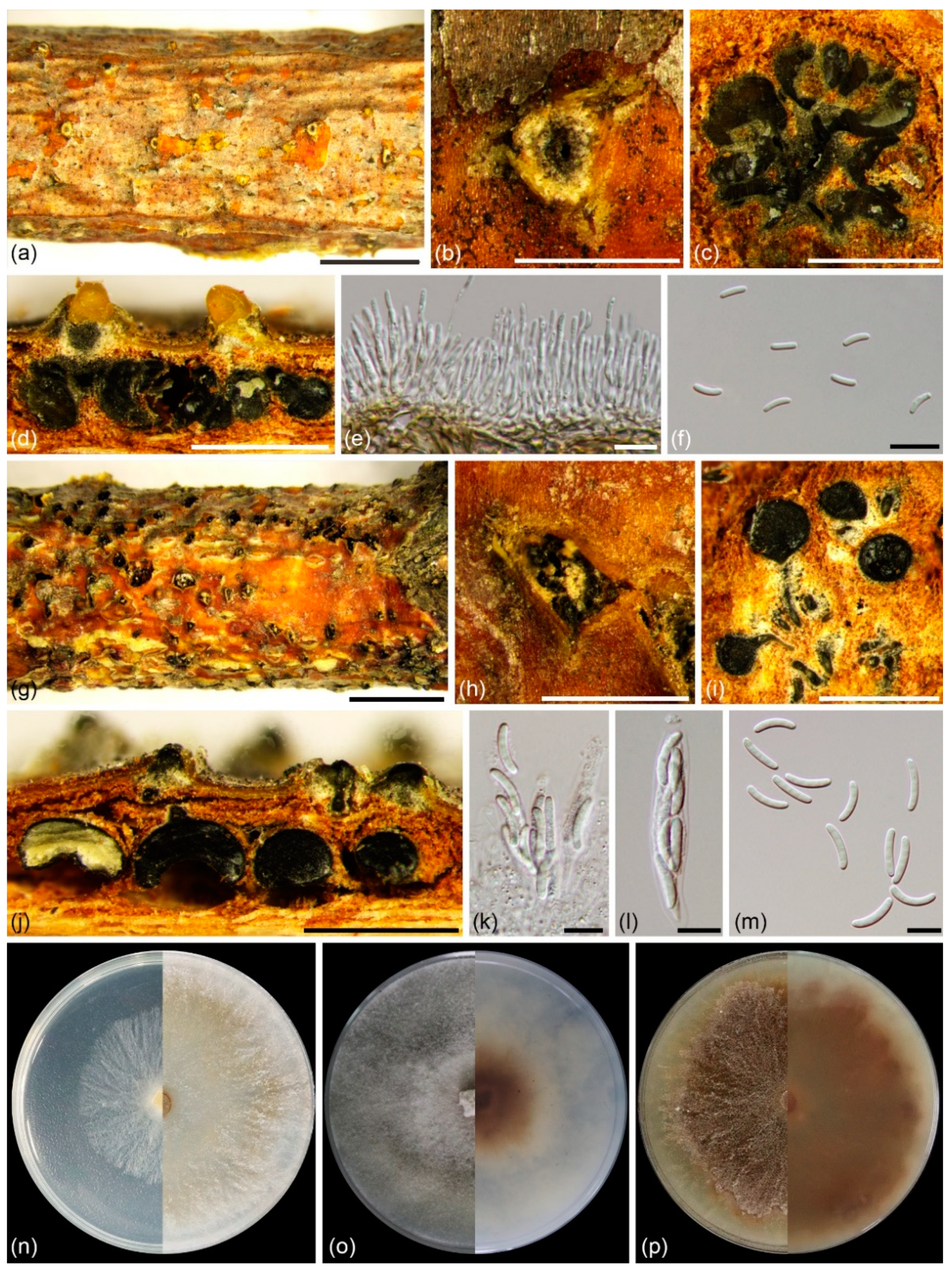
Description: Asexual morph: Conidiomata labyrinthine cytosporoid, immersed in bark, erumpent when mature, discoid to conical, 620–2910 µm in diam, with multiple locules. Conceptacle absent. Ectostromatic disc pale luteous to amber, nearly flat, ovoid to ellipsoid, 380–745 µm diam, with single ostiole per disc in center. Ostiole conspicuous, circular to ovoid, umber to black at the same or lower level as disc surface, 235–585 µm diam. Locules numerous, irregular arrangement with individual walls. Conidiophores borne along locules, hyaline, unbranched, or occasionally branched at base, 12.5–20.0(–28.0) × 1.0–1.5 µm (av. = 16.7 ± 3.1 × 1.4 ± 0.1 µm, n = 30). Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical, 4.0–8.5 × 1.0–2.0 µm (av. = 6.0 ± 1.1 × 1.4 ± 0.2 µm, n = 30). Conidia hyaline, allantoid, smooth, aseptate, thin-walled, (4.0–)5.0–6.2(–6.6) × 1.3–1.7(–2.0) µm (av. = 5.6 ± 0.4 × 1.5 ± 0.1 µm, n = 50). Sexual morph: Ascomata euvalsoid, monostichous, immersed in bark, erumpent through bark surface in a large area, scattered, with 4–7 perithecia arranged irregularly. Conceptacle absent. Ectostromatic disc pale luteous, usually surrounded by ostiolar necks, triangular to circular, 530–1020 µm in diam., with 7–17 ostioles arranged irregularly. Ostioles numerous, greenish olivaceous to black when mature, at the same or below level as disc, arranged irregularly in a disc, 50–90 µm in diam. Perithecia primrose to black when mature, flask-shaped to spherical, arranged irregularly, 355–695 µm in diam. Asci hyaline, with refractive, chitinoid ring in nonamyloid apical apparatus, clavate to elongate-obovoid, 40.0–50.0 × 8.0–10.0 (av. = 46.5 ± 3.2 × 9.0 ± 0.4, n = 30) µm, 8-spored. Ascospores hyaline, biseriate to multiseriate, elongate to allantoid, thin-walled, aseptate, 12.0–16.0 × 2.5–4.0 µm (av. = 14.2 ± 0.7 × 3.0 ± 0.2 µm, n = 50).
Culture characteristics: Cultures on PDA initially white, growing up to 9 cm after 2 days, and becoming umber after 14 days in center. Colonies thick, concentric circles with a uniform texture, lacking aerial mycelium.
Specimens examined: CHINA, Gansu Province: Lanzhou City, Yongdeng County, 102°49′28″ E, 36°44′21″ N, from branches of Salix matsudana, X.L. Fan and L. Lin, 20 October 2020 (holotype BJFC CF20201003, ex-holotype living culture CFCC 55453; ibid., living culture CFCC 55452, CFCC 55454). Gansu Province: Lanzhou City, Yongdeng County, 102°49′26.7″ E, 36°44′22.4″ N, from branches of Salix matsudana, X.L. Fan and L. Lin, 20 October 2020 (paratype BJFC CF20201002, ex-paratype living culture CFCC 55455; ibid., living culture CFCC 55456–55460).
Notes: Cytospora paracinnamomea is associated with canker disease of Salix matsudana in China. It can be distinguished from its phylogenetically closely related species C. cinnamomea (ex-holotype isolate CFCC 53178) and C. donglingensis (ex-type isolate CFCC 53159). Morphologically, it differs from C. cinnamomea by larger ostiole (235–585 vs. 50–63 µm) [26]. It differs from C. donglingensis by cultures on PDA becoming umber after 14 days in the center while cultures of C. donglingensis on PDA become straw after 10 days. Phylogenetically, the current nine strains (CFCC 55452–CFCC 55460) grouped in a separate clade with high statistical support (MP/ML/BI = 99/100/1).
Cytospora rostrata C.M. Tian and X.L. Fan, Mycotaxon 129: 307, 2014.
Descriptions: See Fan et al. [22]
Materials examined: CHINA, Gansu Province: Gannan Autonomous Prefecture, Diebu County, 103°23′34.20″ E, 34°04′48.85″ N, from stems of Salix cupularis, 9 August 2012, Y.M. Liang and X.L. Fan (BJFC S726, living culture CFCC 89909); Gansu Province: Ganan City, Diebu County, 103°23′36.60″ E, 34°04′48.35″ N, from stems of Salix cupularis, 9 August 2012, Y.M. Liang and X.L. Fan (BJFC S727, living culture CFCC 89910).
Notes: Cytospora rostrata can be distinguished from other species by having thornlike ostiolar beak [22]. This species has only been found in Salix cupularis in Gansu Province.
Cytospora sophoriopsis X.L. Fan and C.M. Tian, Persoonia 45: 39, 2020.
Descriptions: See Fan et al. [4]
Material examined: CHINA, Gansu Province: Lanzhou City, Yongdeng County, 102°49′29″ E, 36°44′22″ N, from branches of Salix matsudana, X.L. Fan, N. Jiang, and C. Peng, 20 October 2020 (BJFC CF20201005, living culture CFCC 55469).
Notes: Cytospora sophoriopsis has scattered conidiomata without conceptacles, numerous locules subdivided by invaginations with common wall, and conidia measuring 4.0–4.5 × 1.0–1.5 µm [4]. Phylogenetically, the isolate in the current study (CFCC 55469) formed a fully supported clade with sequences from the ex-type isolate of C. sophoriopsis. In addition, the current study extends the host range of C. sophoriopsis to Salix matsudana in China.
3.3. Pathogenicity Test
Through pathogenicity tests, both C. alba (CFCC 55462) and C. paracinnamomea (CFCC 55453) were confirmed as pathogens on the willow stems (Figure 5). Sunken and resinous cankers were obvious on the stems when the parafilm was removed after 7 days. The enlarged cankers lead further to wilting and consequent death of the plants. No symptoms were observed in the non-inoculated controls. Cytospora alba (CFCC 55462) was found to be more virulent than C. paracinnamomea (CFCC 55453) with the lesion presence frequency reaching 100% and canker length averaging 53 mm after 7 days. The average lesion caused by C. paracinnamomea (CFCC 55453) was 36 mm in length after 7 days. All Cytospora species were reisolated from the lesions or reproductive structures on inoculated plants.
3.4. Temperature, pH, and Carbon Source Tests
The effects of temperature, pH, and carbon source tests were performed with the ex-holotype strains (C. alba: CFCC 55461 and C. paracinnamomea: CFCC 55453). The regression equations and the estimated optimum growth temperature and pH value are presented in Table S3.
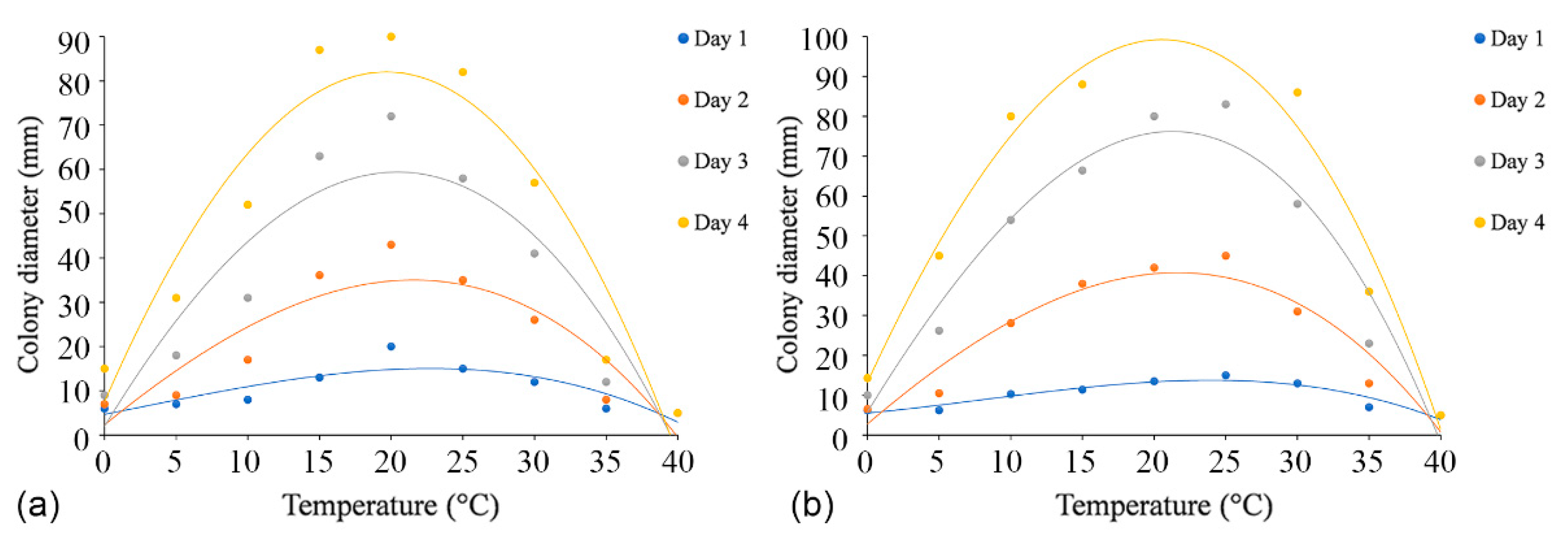
Colonies of both C. alba (CFCC 55461) and C. paracinnamomea (CFCC 55453) grew in the temperature range of 0–30 °C, but not at 35 °C or 40 °C. The fastest mycelial growth of C. alba (CFCC 55461) occurred at 20 °C, and C. paracinnamomea (CFCC 55453) occurred at 20 °C or 25 °C, with colonies of both reaching a diameter of 90 mm after four days. The least mycelial growth of C. alba (CFCC 55461) and C. paracinnamomea (CFCC 55453) all occurred at 0 °C, reaching only 15 mm and 14 mm, respectively, after four days. Based on the regression analysis, the optimal growth after incubation was estimated to occur at 21.1 °C for C. alba (CFCC 55461) and at 21.9 °C for C. paracinnamomea (CFCC 55453) (Figure 6).
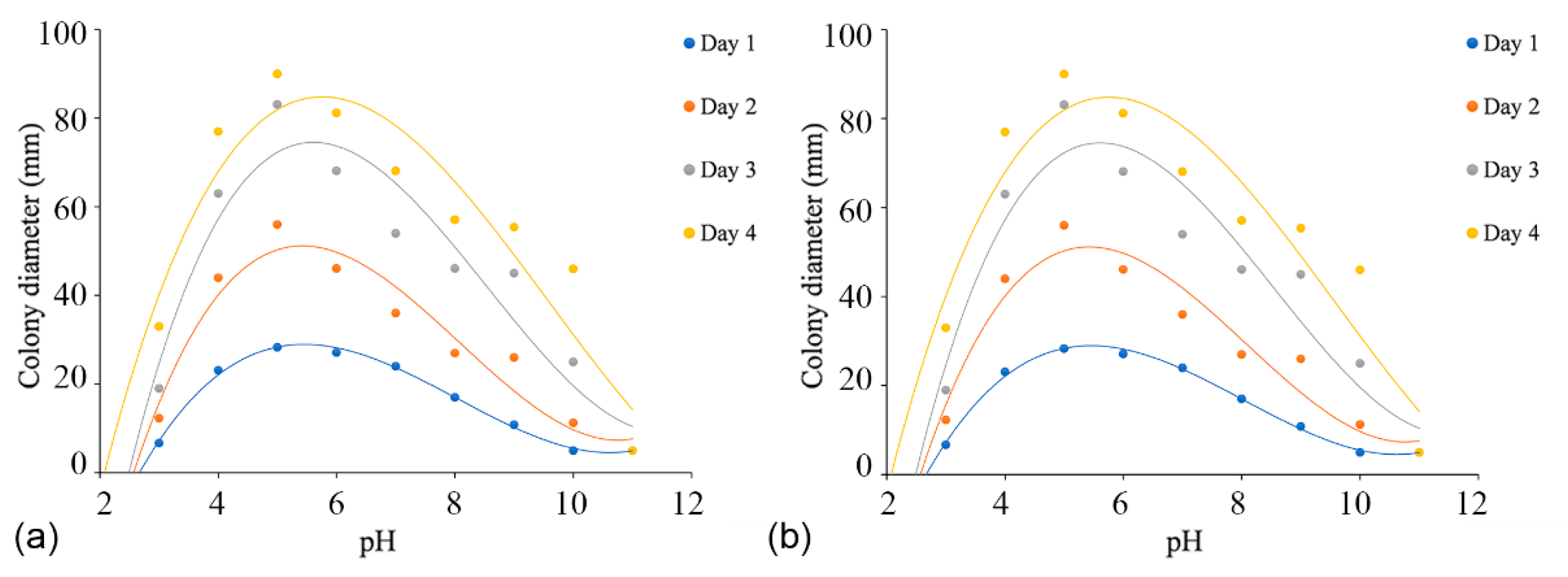
Colonies of both C. alba (CFCC 55461) and C. paracinnamomea (CFCC 55453) grew on PDA in the pH range of 3.0–10.0, but not at a pH of 11.0. For C. alba (CFCC 55461), mycelium grew most rapidly at pH 5.0, reaching 90 mm after four days, followed by pH 4.0 and 6.0. Mycelium grew slowly at pH values of 10.0 and 11.0, attaining colony diameters of no more than 30 mm after four days. For C. paracinnamomea (CFCC 55453), mycelium grew most rapidly at pH 6.0, reaching 90 mm after three days, followed by pH 5.0 and 7.0. Mycelium grew slowly at pH values of 10.0, attaining colony diameters of no more than 14 mm after four days. Based on the regression analysis, the optimal growth after incubation was estimated to occur at pH 5.6 for C. alba (CFCC 55461) and at pH 5.4 for C. paracinnamomea (CFCC 55453) (Figure 7).
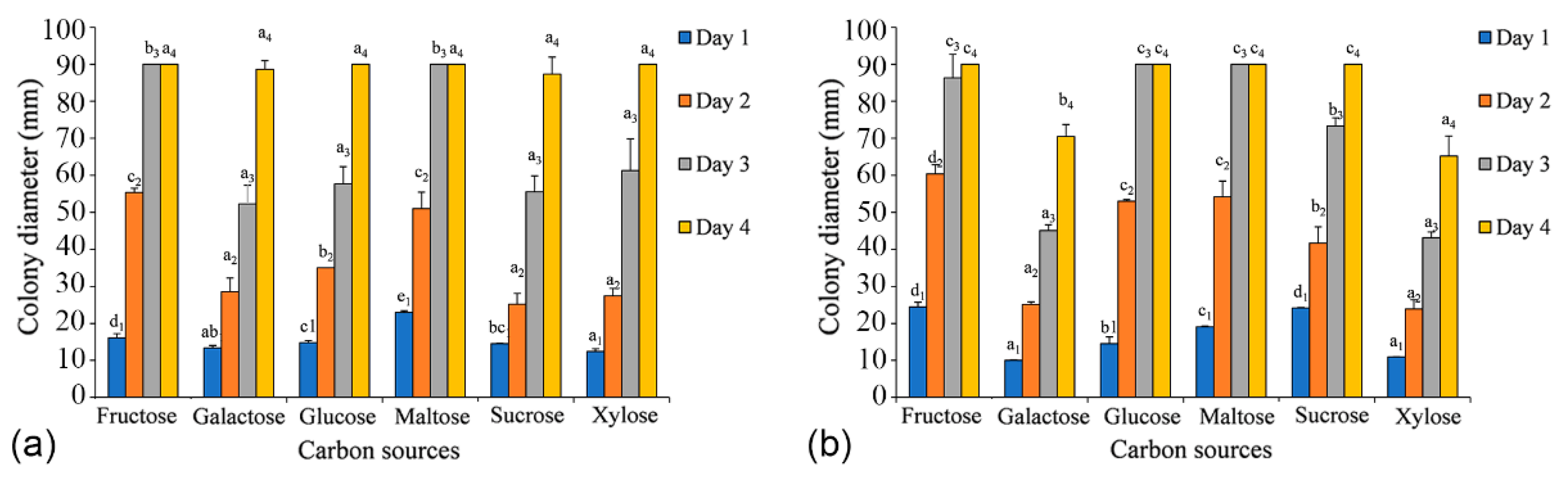
Both C. alba (CFCC 55461) and C. paracinnamomea (CFCC 55453) have the ability to grow on all six carbon sources tested. After two days, the growth of mycelia of C. alba (CFCC 55461) on the fructose and maltose media was apparently faster than that on other media (p-value = 0.0001), and the growth of mycelia of C. paracinnamomea (CFCC 55453) on the fructose, glucose, and maltose media was apparently faster than that on other media after two days (p-value = 0.0001). The colonies grown on other carbon sources were not significantly different from one another. After three days, mycelium growing on fructose and maltose were the first cultures to reach 90 mm in diameter. For C. alba (CFCC 55461), the utilization of sucrose was apparently lower than that of the others, reaching no more than 87 mm growth in diameter after four days. For C. paracinnamomea (CFCC 55453), the utilization of xylose was apparently lower than that of the others, reaching no more than 65 mm growth in diameter after four days (Figure 8).
3.5. Geographic Distribution
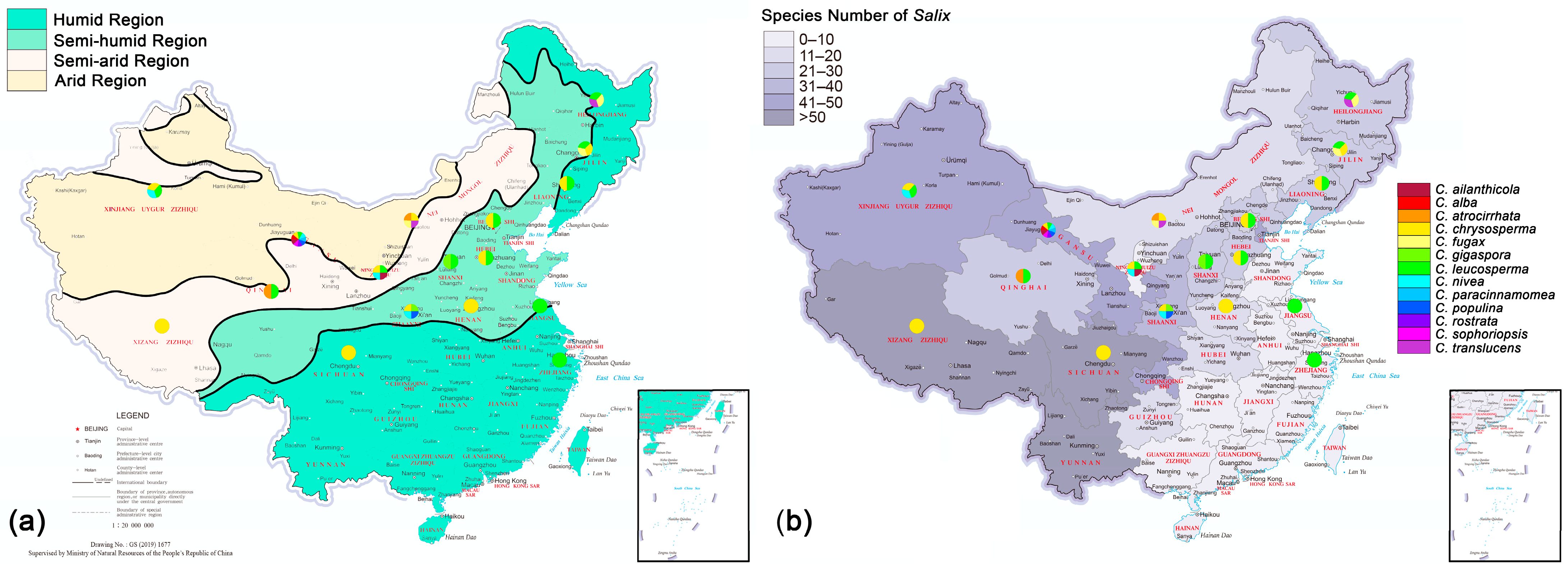
The geographical distribution of Cytospora species from Salix shows that C. chrysosperma and C. leucosperma are the main species observed on Salix, followed by C. nivea (Figure 9). Cytospora chrysosperma and C. leucosperma can be found almost in the entire distribution area of Salix (12 provinces); C. nivea and C. populina are mainly distributed in the Yellow River Basin (four and two provinces, respectively). Gansu province has the highest species diversity of Cytospora (nine species). The results show that the number of Cytospora species from Salix in China is more abundant in the northwest and less in the southeast (Figure 9). Poisson regression analysis showed that Cytospora species richness was significantly positively correlated with dry and wet areas (p-value = 0.024 < 0.05, Estimate = 0.33296). The more drought conditions there were, the more Cytospora species richness was recorded. There was no correlation with host Salix species richness (p-value = 0.303).
4. Discussion
As an afforestation tree species with important economic and ecological value, Salix can be infected by a variety of fungi [4,6,10,12]. Cytospora is a genus in Diaporthales that includes many plant pathogens and endophytes [4,18,27,28,29,54,55]. In the current study, a total of eight Cytospora species from Salix in China were re-evaluated, including C. ailanthicola, C. alba, C. chrysosperma, C. gigaspora, C. nivea, C. paracinnamomea, C. rostrata, C. sophoriopsis, and C. translucens [3,4,13,16,50,51,56,57]. Among these, C. alba and C. paracinnamomea were described as novel species in this study. Cytospora ailanthicola and C. sophoriopsis represent the first record from willow trees in China.
In previous studies, five species (C. atrocirrhata, C. fugax, C. leucosperma, C. populina, and C. translucens) associated with willow canker disease have also been reported in China [3,4,13,24,51,57]. Cytospora atrocirrhata was first reported on branches of willow in Georgia [58], which was described as erumpent conidiomata with distinct conceptacles and single locules and ostiole [24]. Subsequently, this species was reported on poplar and walnut in Inner Mongolia and Qinghai [3,24]. Spielman [53] re-described the sexual and asexual morph of C. fugax, which provided important information for the accurate identification of this species. In China, Cytospora fugax was previously recorded in Heilongjiang, Inner Mongolia, and Jilin [3]. Cytospora leucosperma was recorded in Beijing, Gansu, Hebei, Heilongjiang, Jiangsu, Jilin, Liaoning, Ningxia, Qinghai, Shanxi, Xinjiang, and Zhejiang Provinces in China [4,13,51,57]. Cytospora populina was redescribed by Fan et al. [24] on branches of Salix psammophila in China. It can be distinguished from other Cytospora species by its asci with four ascospores [24]. Cytospora translucens was previously discovered on willow and poplar in Beijing, Heilongjiang, Inner Mongolia, and Jilin [3]. Vu et al. [59] added their DNA data; however, they still require a modern illustration and description.
Cytospora atrocirrhata, C. chrysosperma, C. fugax, C. leucosperma, C. nivea, and C. translucens were also reported to infect Salix spp. in other countries, e.g., Canada, England, Iran, New Zealand, Poland (https://nt.ars-grin.gov/fungaldatabases/index.cfm, accessed on 6 April 2022). Among these species, C. chrysosperma can infect over 110 host species, which is a prominent species of Cytospora in the world [4,18]. Cytospora ailanthicola, C. gigaspora, C. rostrata, C. sophoriopsis have only been found in China, of which C. rostrata has been only found in Salix cupularis [3,4,22,23]. Although C. alba and C. paracinnamomea have been confirmed as pathogens of willow canker disease, they have only been found in Gansu province. Whether they are distributed elsewhere and infect local willow trees are unknown.
The results of pathogenicity tests showed that C. alba and C. paracinnamomea were pathogens to Salix matsudana, which had a high degree of damage. The optimal growth was estimated to occur at 21.1 °C for C. alba (CFCC 55461) and at 21.9 °C for C. paracinnamomea (CFCC 55453); and at pH 5.6 for C. alba (CFCC 55461) and at pH 5.4 for C. paracinnamomea (CFCC 55453). Fructose and maltose were utilized better than other tested carbon sources. Based on the biological characterization of these two novel Cytospora species, more attention is needed to prevent the occurrence of Cytospora canker disease in the region with an average monthly temperature of approximately 20 °C, such as April, May, September, and October in Southwest, Northwest, and North China. This indicates that both spring and autumn may be a time of high disease risk. Therefore, the understanding of biological characteristics of these Cytospora species can be used for distribution prediction. Preventive measures can be taken in advance.
The geographical distribution analysis showed that Cytospora on Salix was less commonly known in the humid regions than in the arid region, semiarid region, and semi-humid region. Previous studies showed that drought-stressed plants are more prone to Cytospora cankers [60,61]. However, Kristen et al. [62] demonstrated that reduced water availability and a 2–3 °C increase in temperature did not significantly increase the incidence or severity of cankers in inoculated willow plants. Therefore, willows that occur in dry areas are more likely to be infected by Cytospora because the trees are weakened by drought stress and poor soil conditions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8040377/s1, Refs. [63,64,65,66,67], Figure S1: Phylogenetic tree of Cytospora species based on ML analyses of the ITS gene sequences; Figure S2: Phylogenetic tree of Cytospora species based on ML analyses of the act gene sequences; Figure S3: Phylogenetic tree of Cytospora species based on ML analyses of the rpb2 gene sequences; Figure S4: Phylogenetic tree of Cytospora species based on ML analyses of the tef1-α gene sequences; Figure S5: Phylogenetic tree of Cytospora species based on ML analyses of the tub2 gene sequences; Table S1: Genes used in this study with PCR primers, primer DNA sequence, optimal annealing temperature; Table S2: Strains of Cytospora used in the molecular analyses in this study; Table S3: The regression equations and the estimated optimum growth temperature, pH value, and carbon source.
Author Contributions
Conceptualization, L.L. and X.F.; methodology, L.L. and M.P.; software, L.L. and X.F.; validation, L.L. and M.P.; formal analysis, L.L. and M.P.; investigation, L.L., M.P. and X.F.; resources, L.L., X.F. and C.T.; writing—original draft preparation, L.L.; writing—review and editing, X.F. and C.T.; visualization, L.L.; supervision, X.F.; funding acquisition, X.F. and C.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32101533), National Science and Technology Fundamental Resources Investigation Program of China (2021FY100900) and College Student Research and Career-Creation Program of Beijing (X202110022006).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Alignments generated during the current study are available in Tree-BASE (accession http://purl.org/phylo/treebase/phylows/study/TB2:S29245, accessed on 6 April 2022). All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Acknowledgments
We are grateful to Yingmei Liang (the Museum of Beijing Forestry University) for providing relevant specimens, also to Xiaohong Liang, Jing Han (the Experimental Teaching Centre, College of Forestry, Beijing Forestry University) for providing installed scientific equipment in the whole process. We are also grateful for the assistance of Ce Shang (School of Ecology and Nature Conservation, Beijing Forestry University) during this study. Xinlei Fan would like to acknowledge the supporting of strain preservation of Chungen Piao and Minwei Guo (China Forestry Culture Collection Centre, Chinese Academy of Forestry, Beijing).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kuzovkina, Y.A.; Volk, T.A. The characterization of willow (Salix L.) varieties for use in ecological engineering applications: Co–ordination of structure, function and autecology. Ecol. Eng. 2009, 358, 1178–1189. [Google Scholar] [CrossRef]
- Yu, Z.X.; Qin, G.H.; Jiang, Y.Z.; Song, Y.M.; Qiao, Y.L. Analysis of the reasons for the difficulties in willow classification based on the research status of willow in China. J. Agric. Sci. 2018, 8, 22–25. (In Chinese) [Google Scholar]
- Wang, Y.L.; Lu, Q.; Decock, C.; Li, Y.X.; Zhang, X.Y. Cytospora species from Populus and Salix in China with C. davidiana sp. nov. Fungal Biol. 2015, 119, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Persoonia 2020, 451, 1–45. [Google Scholar] [CrossRef]
- Bubner, B.; Wunder, S.; Zaspel, I.; Zander, M.; Gloger, J.; Fehrenz, S.; Ulrichs, C. Melampsora rust species on biomass willows in central and north-eastern Germany. Fungal Biol. 2014, 118, 910–923. [Google Scholar] [CrossRef]
- Zhao, P.; Tian, C.M.; Yao, Y.J.; Wang, Q.; Yamaoka, Y.; Kakishima, M. Two new species and one new record of Melampsora on willows from China. Mycol. Prog. 2015, 14, 66. [Google Scholar] [CrossRef]
- Yun, Y.H.; Ahn, G.R.; Yoon, S.K.; Kim, H.H.; Son, S.Y.; Kim, S.H. New rust disease of Korean willow (Salix koreensis) caused by Melampsora yezoensis, unrecorded pathogen in Korea. Mycobiology 2016, 444, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Ishaq, A.; Afshan, N.U.S.; Khalid, A.N. First report of Melampsora dimorphospora on white willow in Pakistan. J. Plant Pathol. 2019, 101, 197. [Google Scholar] [CrossRef]
- Stupar, M.; Ljaljevic Grbic, M.; Vukojevic, J.; Lakusic, D. New reports of Melampsora rust (Pucciniomycetes) on the Salix retusa complex in Balkan countries. Bot. Serbica 2020, 44, 89–93. [Google Scholar] [CrossRef]
- Okorski, A.; Pszczolkowska, A.; Sulima, P.; Paukszto, L.; Jastrzebski, J.P.; Przyborowski, J.; Makowczenko, K.G. First report of willow anthracnose caused by Colletotrichum salicis in Poland. Plant Dis. 2018, 102, 2036–2037. [Google Scholar] [CrossRef]
- Masumoto, S.; Tojo, M.; Uchida, M.; Imura, S. Morphological and molecular characterization of Rhytisma filamentosum sp. nov. from Nagano Prefecture, Japan. Mycol. Prog. 2015, 14, 44. [Google Scholar] [CrossRef]
- Jung, B.N.; Park, J.H.; Shin, H.D. First report of Rhytisma filamentosum causing tar-spot disease on Salix koriyanagi. For. Pathol. 2020, 50, e12577. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- Wu, C.L. Cytospora canker outbreak in the main city of northern China. China J. For. Pest Dis. 1999, 2, 36. (In Chinese) [Google Scholar]
- Zhang, X.Y.; Luo, Y.Q. Major Forest Diseases and Insect Pests in China; China Forest Press: Beijing, China, 2003. (In Chinese) [Google Scholar]
- Wang, Y.L.; Lu, Q.; Jia, X.Z.; Liang, J.; Zhang, X.Y. First report of branch canker caused by Cytospora atrocirrhata on Populus sp. and Salix sp. in China. Plant Dis. 2013, 97, 426. [Google Scholar] [CrossRef]
- Xu, Y.L. Synthetic Cure and Management of Valsa Sordida Nits of Main Virescence Wood Species in Harbin; Master-Northeast Forestry University: Harbin, China, 2003. (In Chinese) [Google Scholar]
- Adams, G.C.; Wingfield, M.J.; Common, R.; Roux, J. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Stud. Mycol. 2005, 52, 1–144. [Google Scholar]
- Christensen, C.M. Studies on the biology of Valsa sordida and Cytospora chrysosperma. Phytopathology 1940, 30, 459–475. [Google Scholar]
- Bills, G.F. Isolation and analysis of endophytic fungal communities from woody plants. In Endophytic Fungi in Grasses and Woody Plants; Redlin, S.C., Carris, L.M., Eds.; APS Press: St. Paul, MN, USA, 1996; pp. 31–65. [Google Scholar]
- Ehrenberg, C.G. Sylvae Mycologicae Berolinenses; Formis Theophili Bruschcke: Berlin, Germany, 1818. [Google Scholar]
- Fan, X.L.; Tian, C.M.; Yang, Q.; Liang, Y.M.; You, C.J.; Zhang, Y.B. Cytospora from Salix in northern China. Mycotaxon 2014, 129, 303–315. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Liu, M.; Liang, Y.M.; Tian, C.M. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 1195, 310–319. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Ma, R.; Tian, C.M. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 1974, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Fan, X.L.; Crous, P.W.; Liang, Y.M.; Tian, C.M. Cytospora from Ulmus pumila in Northern China. Mycol. Prog. 2015, 14, 74. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Bonthond, G.; Tian, C.M.; Fan, X.L. High diversity of Cytospora associated with canker and dieback of Rosaceae in China, with 10 new species described. Front. Plant Sci. 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Norphanphoun, C.; Doilom, M.; Daranagama, D.A.; Phookamsak, R.; Wen, T.C.; Bulgakov, T.S.; Hyde, K.D. Revisiting the genus Cytospora and allied species. Mycosphere 2017, 8, 51–97. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Raspé, O.; Jeewon, R.; Wen, T.C.; Hyde, K.D. Morphological and phylogenetic characterisation of novel Cytospora species associated with mangroves. MycoKeys 2018, 38, 93–120. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Holland, L.A.; Nouri, M.T.; Travadon, R.; Abramians, A.; Michailides, T.J.; Trouillas, F.P. Molecular phylogeny of Cytospora species associated with canker diseases of fruit and nut crops in California, with the descriptions of ten new species and one new combination. IMA Fungus 2018, 92, 333–369. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Pan, M.; Bonthond, G.; Tian, C.M.; Fan, X.L. Diaporthalean fungi associated with canker and dieback of trees from Mount Dongling in Beijing, China. MycoKeys 2019, 59, 67–94. [Google Scholar] [CrossRef]
- Jiang, N.; Yang, Q.; Fan, X.L.; Tian, C.M. Identification of six Cytospora species on Chinese chestnut in China. MycoKeys 2020, 62, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhu, H.Y.; Tian, C.M.; Huang, M.R.; Fan, X.L. Assessment of Cytospora strains from conifer cankers in China, with the descriptions of four new Cytospora species. Front. Plant Sci. 2021, 12, 636460. [Google Scholar] [CrossRef]
- Shang, Q.J.; Hyde, K.D.; Camporesi, E.; Maharachchikumbura, S.S.N.; Norphanphoun, C.; Brooks, S.; Liu, J.K. Additions to the genus Cytospora with sexual morph in Cytosporaceae. Mycosphere 2020, 11, 189–224. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: London, UK, 1970. [Google Scholar]
- Doyle, J.J. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 304, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony, * and Other Methods, version 4.0b10; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, H.O. New algorithms and methods to estimate maximum–likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 593, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. Mr. Bayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A. FigTree, version 1.3.1; Institute of Evolutionary Biology: Edinburgh, UK, 2010. [Google Scholar]
- Koch, R. Uber bakteriologische forschung. In Proceedings of the Xth International Congress of Medicine, Berlin, Germany, 4 August 1890; Volume 1, p. 35. [Google Scholar]
- Zhou, X.; Pan, M.; Li, H.Y.; Tian, C.M.; Fan, X.L. Dieback of Euonymus alatus (Celastraceae) caused by Cytospora haidianensis sp. nov. in China. Forests 2020, 115, 524. [Google Scholar] [CrossRef]
- Gao, H.; Pan, M.; Tian, C.M.; Fan, X.L. Cytospora and Diaporthe species associated with hazelnut canker and dieback in Beijing, China. Front. Cell. Infect. Microbiol. 2021, 11, 664366. [Google Scholar] [CrossRef]
- Wu, S.H.; Yin, Y.H.; Zheng, D.; Yang, Q.Y. Study on dry and wet conditions of land surface in China in recent 30 years. Sci. China 2005, 3, 276–283. (In Chinese) [Google Scholar]
- Fang, Z.F.; Zhao, S.D.; Alexei, K.S. Salicaceae. In Cochairs of the Editorial Committee, Flora of China; Wu, Z.Y., Peter, H.R., Eds.; Science Press: Beijing, China, 2004; Volume 4. [Google Scholar]
- Zheng, W.J. Sylva Sinica; China Forestry Publishing House: Beijing, China, 1985. (In Chinese) [Google Scholar]
- Fang, J.Y.; Wang, Z.H.; Tang, Z.Y. Atlas of Woody Plant in China—Distribution and Climate; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963. (In Chinese) [Google Scholar]
- Wei, J.C. Identification of Fungus Handbook; Science Press: Shanghai, China, 1979. (In Chinese) [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Spielman, L.J. A monograph of Valsa on hardwoods in North America. Can. J. Bot. 1985, 63, 1355–1378. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from fagaceae leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in the Netherlands. MycoKeys 2021, 85, 31. [Google Scholar] [CrossRef]
- Chen, M.M. Forest fungi Phytogeography: Forest fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research & Education Center: Sacramento, CA, USA, 2003. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Ithaca: New York, NY, USA, 2005. (In Chinese) [Google Scholar]
- Gvritishvili, M.N. New species of fungi belonging to genus Cytospora Fr. II. Mikol. Fitopatol. 1973, 7, 544–549. (In Russian) [Google Scholar]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 9, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Kepley, J.; Jacobi, W.R. Pathogenicity of Cytospora fungi on six hardwood species. J. Arboric. 2000, 26, 326–333. [Google Scholar] [CrossRef]
- McIntyre, G.A.; Jacobi, W.R.; Ramaley, A.W. Factors affecting Cytospora canker occurrence on aspen. J. Arboric. 1996, 22, 229–233. [Google Scholar]
- Kaczynski, K.M.; Cooper, D.J. Susceptibility of Salix monticola to Cytospora canker under increased temperatures and decreased water levels. For. Ecol. Manag. 2013, 305, 223–228. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
Figure 1.
Field sampling in Lanzhou City, Gansu Province: (a) death of the willow trees caused by Cytospora species; (b) symptoms of branches canker.
Figure 1.
Field sampling in Lanzhou City, Gansu Province: (a) death of the willow trees caused by Cytospora species; (b) symptoms of branches canker.

Figure 2.
Maximum parsimony phylogram of Cytospora based on combined ITS, act, rpb2, tef1-α, and tub2 genes. MP and ML bootstrap support values above 50% are shown at the first and second position. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type strains are in bold. Strains from the current study are in blue.
Figure 2.
Maximum parsimony phylogram of Cytospora based on combined ITS, act, rpb2, tef1-α, and tub2 genes. MP and ML bootstrap support values above 50% are shown at the first and second position. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type strains are in bold. Strains from the current study are in blue.

Figure 3.
Cytospora alba (BJFC CF20201001) from Salix matsudana: (a,b) habit of conidiomata on twig; (c) transverse section of conidioma; (d) longitudinal section through conidioma; (e) conidiophores and conidiogenous cells; (f) conidia. Scale bars: 5 mm (a); 1 mm (b–d); 10 µm (e,f). Cultures on PDA (g–i, CFCC 55402): (g) 2 days; (h) 7 days; (i) 30 days, front side at left and back side at right.
Figure 3.
Cytospora alba (BJFC CF20201001) from Salix matsudana: (a,b) habit of conidiomata on twig; (c) transverse section of conidioma; (d) longitudinal section through conidioma; (e) conidiophores and conidiogenous cells; (f) conidia. Scale bars: 5 mm (a); 1 mm (b–d); 10 µm (e,f). Cultures on PDA (g–i, CFCC 55402): (g) 2 days; (h) 7 days; (i) 30 days, front side at left and back side at right.

Figure 4.
Cytospora paracinnamomea (BJFC CF20201003) from Salix matsudana: (a,b) habit of conidiomata on twig; (c) transverse section of conidioma; (d) longitudinal section through conidiomata; (e) conidiophores and conidiogenous cells; (f) conidia; (g,h) habit of ascomata on twig; (i) transverse section of ascoma; (j) longitudinal section through ascomata; (k,l) asci; (m) ascospores. Scale bars: 5 mm (a,g); 1 mm (b–d,g,h); 10 µm (e,f,k–m). Cultures on PDA (n–p, CFCC 55453): (n) 2 days at left and 7 days at right; (o) 14 days, front side at left and back side at right; (p) 30 days, front side at left and back side at right.
Figure 4.
Cytospora paracinnamomea (BJFC CF20201003) from Salix matsudana: (a,b) habit of conidiomata on twig; (c) transverse section of conidioma; (d) longitudinal section through conidiomata; (e) conidiophores and conidiogenous cells; (f) conidia; (g,h) habit of ascomata on twig; (i) transverse section of ascoma; (j) longitudinal section through ascomata; (k,l) asci; (m) ascospores. Scale bars: 5 mm (a,g); 1 mm (b–d,g,h); 10 µm (e,f,k–m). Cultures on PDA (n–p, CFCC 55453): (n) 2 days at left and 7 days at right; (o) 14 days, front side at left and back side at right; (p) 30 days, front side at left and back side at right.

Figure 5.
Stem blight symptoms on Salix matsudana caused by Cytospora species: (a–d) stem blight symptoms on Salix matsudana caused by Cytospora alba (CFCC 55462). Symptoms after (a) one week and (b) two weeks. (c) Resinous canker formed after inoculation with C. alba. (d) Longitudinal section through pathogenic site. (e,f) Stem blight symptoms on Salix matsudana caused by Cytospora paracinnamomea (CFCC 55453). Symptoms after (e) one week and (f) two weeks. (g,h) Blank control group, no symptoms on Salix matsudana twigs after three weeks of inoculation with agar block.
Figure 5.
Stem blight symptoms on Salix matsudana caused by Cytospora species: (a–d) stem blight symptoms on Salix matsudana caused by Cytospora alba (CFCC 55462). Symptoms after (a) one week and (b) two weeks. (c) Resinous canker formed after inoculation with C. alba. (d) Longitudinal section through pathogenic site. (e,f) Stem blight symptoms on Salix matsudana caused by Cytospora paracinnamomea (CFCC 55453). Symptoms after (e) one week and (f) two weeks. (g,h) Blank control group, no symptoms on Salix matsudana twigs after three weeks of inoculation with agar block.

Figure 6.
Effects of temperature on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462): yDay1 = −0.0005x3 + 0.0035x2 + 0.6438x + 4.6768 (R2 = 0.7348); yDay2 = −0.0009x3 − 0.033x2 + 2.6289x + 2.3212 (R2 = 0.8171); yDay3 = −0.0007x3 − 0.1103x2 + 5.3437x + 1.9394 (R2 = 0.8513); yDay4 = −0.0004x3 − 0.1762x2 + 7.4157x + 7.4949 (R2 = 0.9055). (b) Cytospora paracinnamomea (CFCC 55453): yDay1 = −0.0006x3 + 0.0147x2 + 0.3385x + 5.5222 (R2 = 0.8937); yDay2 = −0.0009x3 − 0.0406x2 + 3.0735x + 2.7606 (R2 = 0.9161); yDay3 = −0.0017x3 − 0.0843x2 + 5.8622x + 5.7758 (R2 = 0.944); yDay4 = −0.0014x3 − 0.1473x2 + 7.7947x + 13.211 (R2 = 0.9641).
Figure 6.
Effects of temperature on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462): yDay1 = −0.0005x3 + 0.0035x2 + 0.6438x + 4.6768 (R2 = 0.7348); yDay2 = −0.0009x3 − 0.033x2 + 2.6289x + 2.3212 (R2 = 0.8171); yDay3 = −0.0007x3 − 0.1103x2 + 5.3437x + 1.9394 (R2 = 0.8513); yDay4 = −0.0004x3 − 0.1762x2 + 7.4157x + 7.4949 (R2 = 0.9055). (b) Cytospora paracinnamomea (CFCC 55453): yDay1 = −0.0006x3 + 0.0147x2 + 0.3385x + 5.5222 (R2 = 0.8937); yDay2 = −0.0009x3 − 0.0406x2 + 3.0735x + 2.7606 (R2 = 0.9161); yDay3 = −0.0017x3 − 0.0843x2 + 5.8622x + 5.7758 (R2 = 0.944); yDay4 = −0.0014x3 − 0.1473x2 + 7.7947x + 13.211 (R2 = 0.9641).

Figure 7.
Effects of pH on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462): yDay1 = 0.3546x3 − 8.5492x2 + 61.603x − 110.27 (R2 = 0.9957); yDay2 = 0.5799x3 − 14.068x2 + 101.43x − 177.67 (R2 = 0.9252); yDay3 = 0.6425x3 − 16.478x2 + 124.19x − 217.02 (R2 = 0.8945); yDay4 = 0.4139x3 − 11.886x2 + 95.703x − 151.21 (R2 = 0.8693). (b) Cytospora paracinnamomea (CFCC 55453): yDay1 = 0.2383x3 − 5.3296x2 + 35.599x − 57.712 (R2 = 0.9028); yDay2 = 0.9923x3 − 23.044x2 + 160.21x − 287.09 (R2 = 0.9761); yDay3 = 1.0142x3 − 25.35x2 + 188.44x − 350.98 (R2 = 0.9806); yDay4 = 1.037x3 − 27.082x2 + 208.41x − 391.11 (R2 = 0.9361).
Figure 7.
Effects of pH on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462): yDay1 = 0.3546x3 − 8.5492x2 + 61.603x − 110.27 (R2 = 0.9957); yDay2 = 0.5799x3 − 14.068x2 + 101.43x − 177.67 (R2 = 0.9252); yDay3 = 0.6425x3 − 16.478x2 + 124.19x − 217.02 (R2 = 0.8945); yDay4 = 0.4139x3 − 11.886x2 + 95.703x − 151.21 (R2 = 0.8693). (b) Cytospora paracinnamomea (CFCC 55453): yDay1 = 0.2383x3 − 5.3296x2 + 35.599x − 57.712 (R2 = 0.9028); yDay2 = 0.9923x3 − 23.044x2 + 160.21x − 287.09 (R2 = 0.9761); yDay3 = 1.0142x3 − 25.35x2 + 188.44x − 350.98 (R2 = 0.9806); yDay4 = 1.037x3 − 27.082x2 + 208.41x − 391.11 (R2 = 0.9361).

Figure 8.
Effects of carbon sources on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462). (b) Cytospora paracinnamomea (CFCC 55453). A p-value < 0.05 was considered significant. Graphs with SE-bars showed the difference in mycelium growth under different conditions.
Figure 8.
Effects of carbon sources on the growth of Cytospora species. (a) Cytospora alba (CFCC 55462). (b) Cytospora paracinnamomea (CFCC 55453). A p-value < 0.05 was considered significant. Graphs with SE-bars showed the difference in mycelium growth under different conditions.

Figure 9.
Geographical distribution of Cytospora species associated with willow canker disease in China: (a) drought conditions and Cytospora species richness of each province; (b) Salix species richness and Cytospora species richness of each province.
Figure 9.
Geographical distribution of Cytospora species associated with willow canker disease in China: (a) drought conditions and Cytospora species richness of each province; (b) Salix species richness and Cytospora species richness of each province.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, L.; Pan, M.; Tian, C.; Fan, X. Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. J. Fungi 2022, 8, 377. https://doi.org/10.3390/jof8040377
AMA Style
Lin L, Pan M, Tian C, Fan X. Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. Journal of Fungi. 2022; 8(4):377. https://doi.org/10.3390/jof8040377
Chicago/Turabian StyleLin, Lu, Meng Pan, Chengming Tian, and Xinlei Fan. 2022. "Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China" Journal of Fungi 8, no. 4: 377. https://doi.org/10.3390/jof8040377
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.