
Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
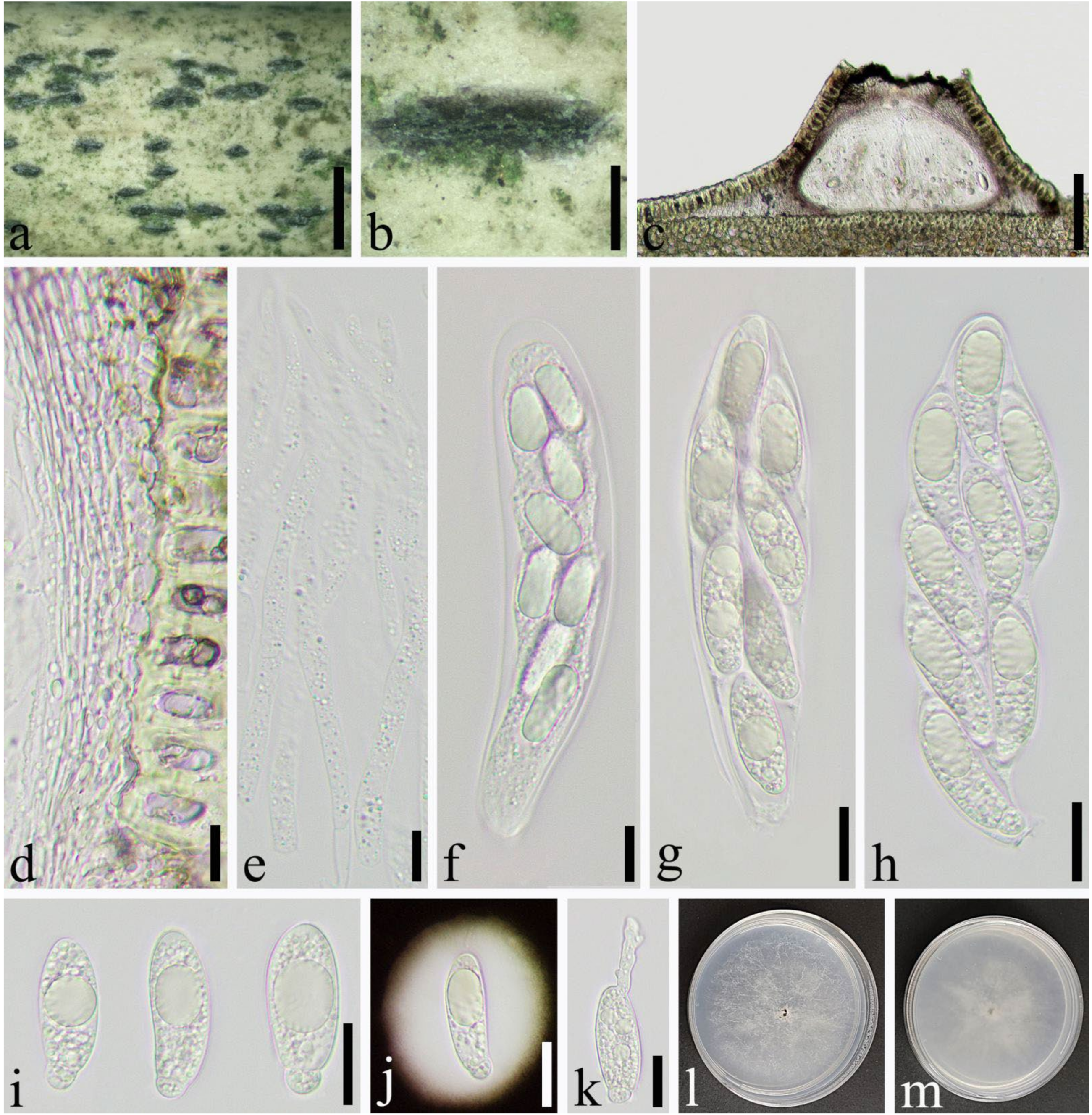
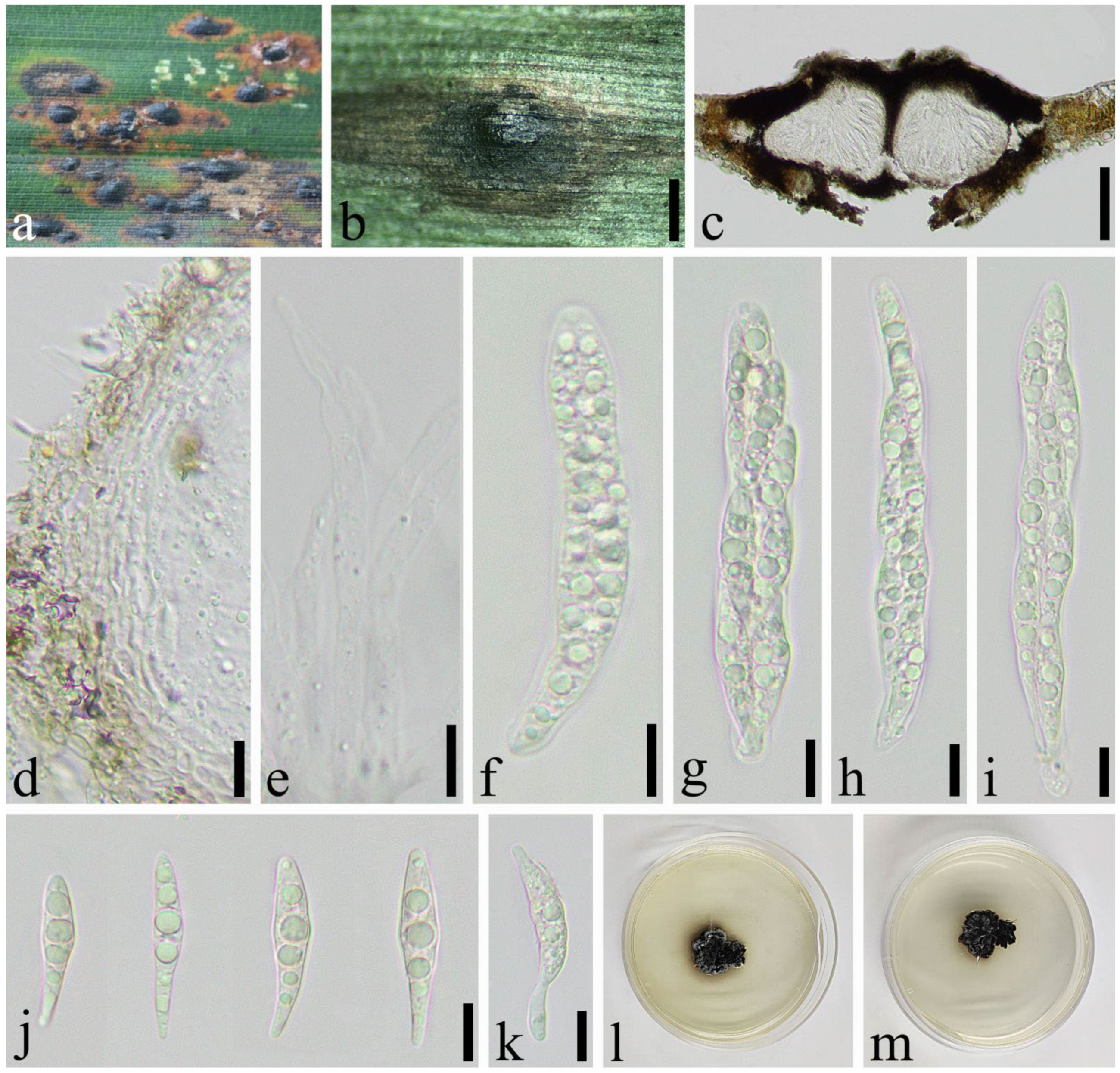
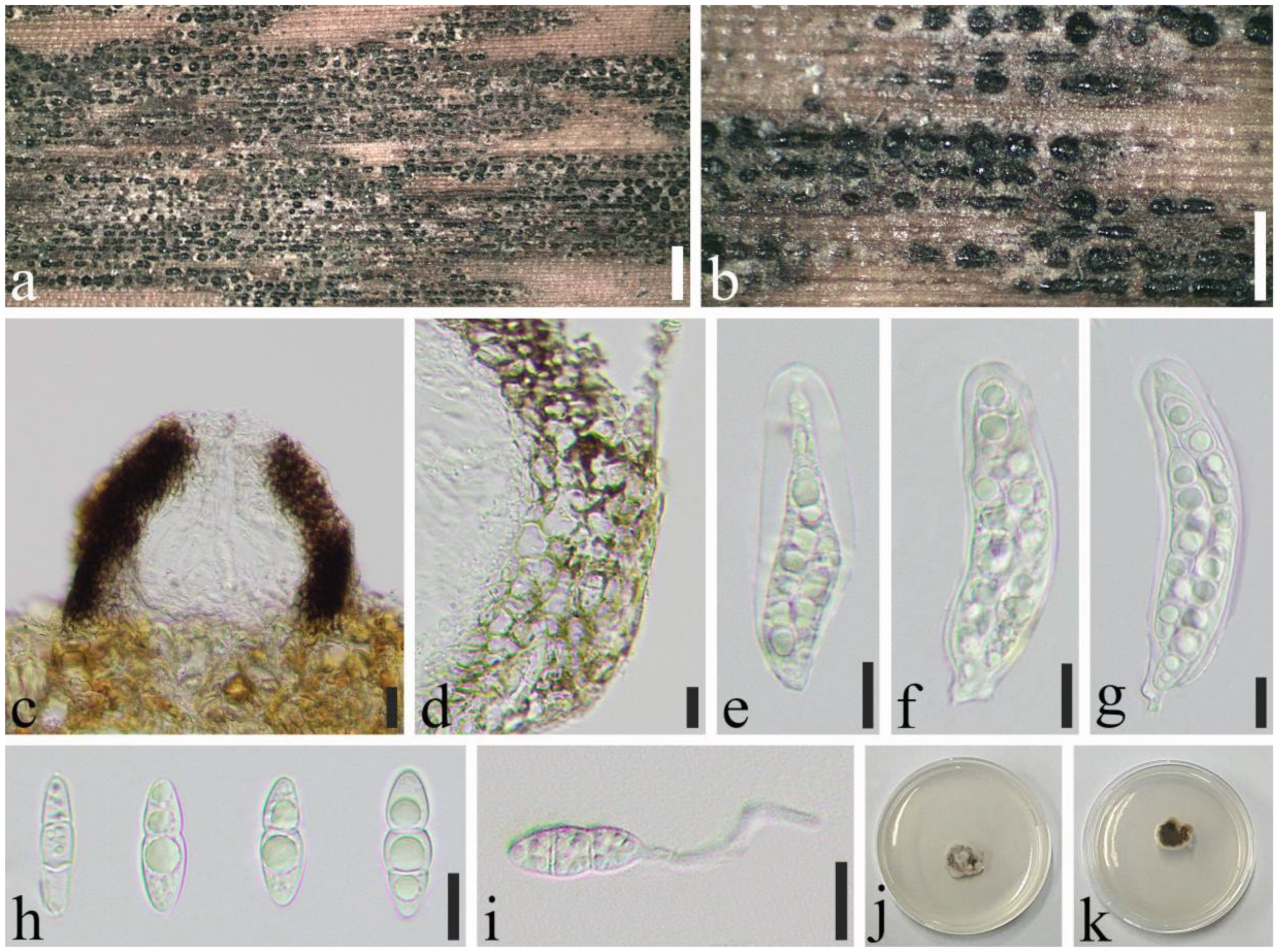
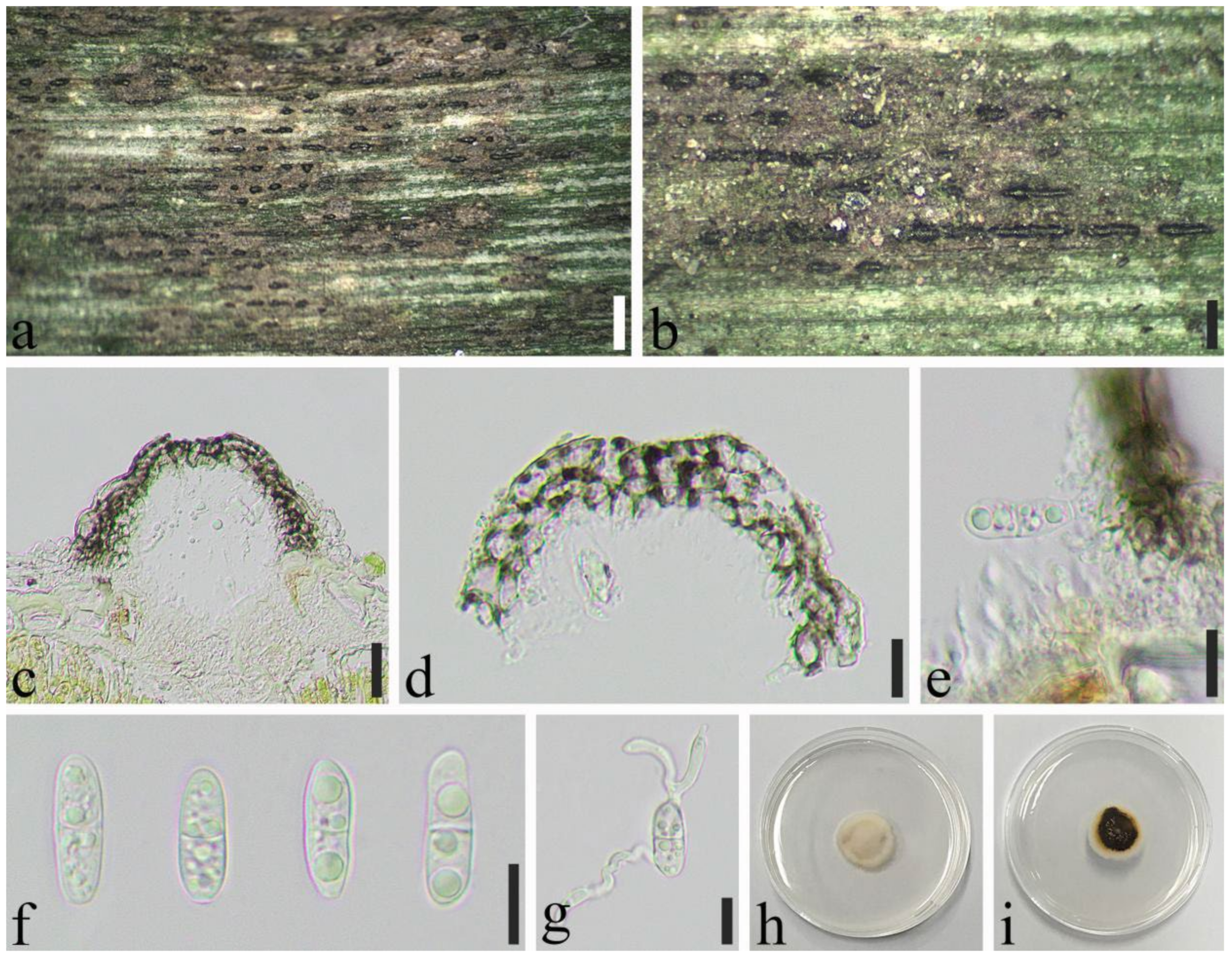
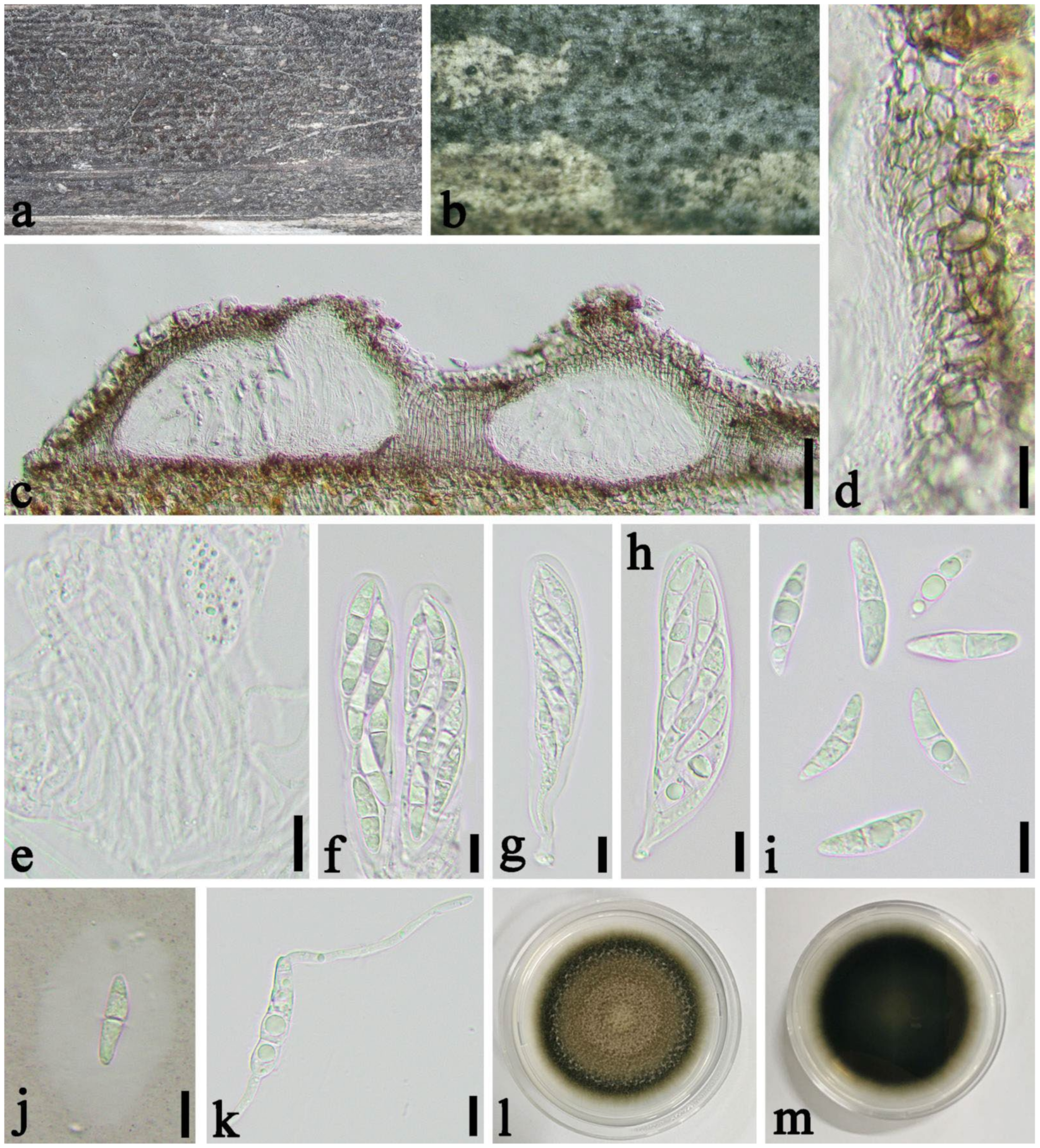
2.1. Specimen Collection and Morphological Study
2.2. DNA Extraction, PCR Amplification, and Nucleotide Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
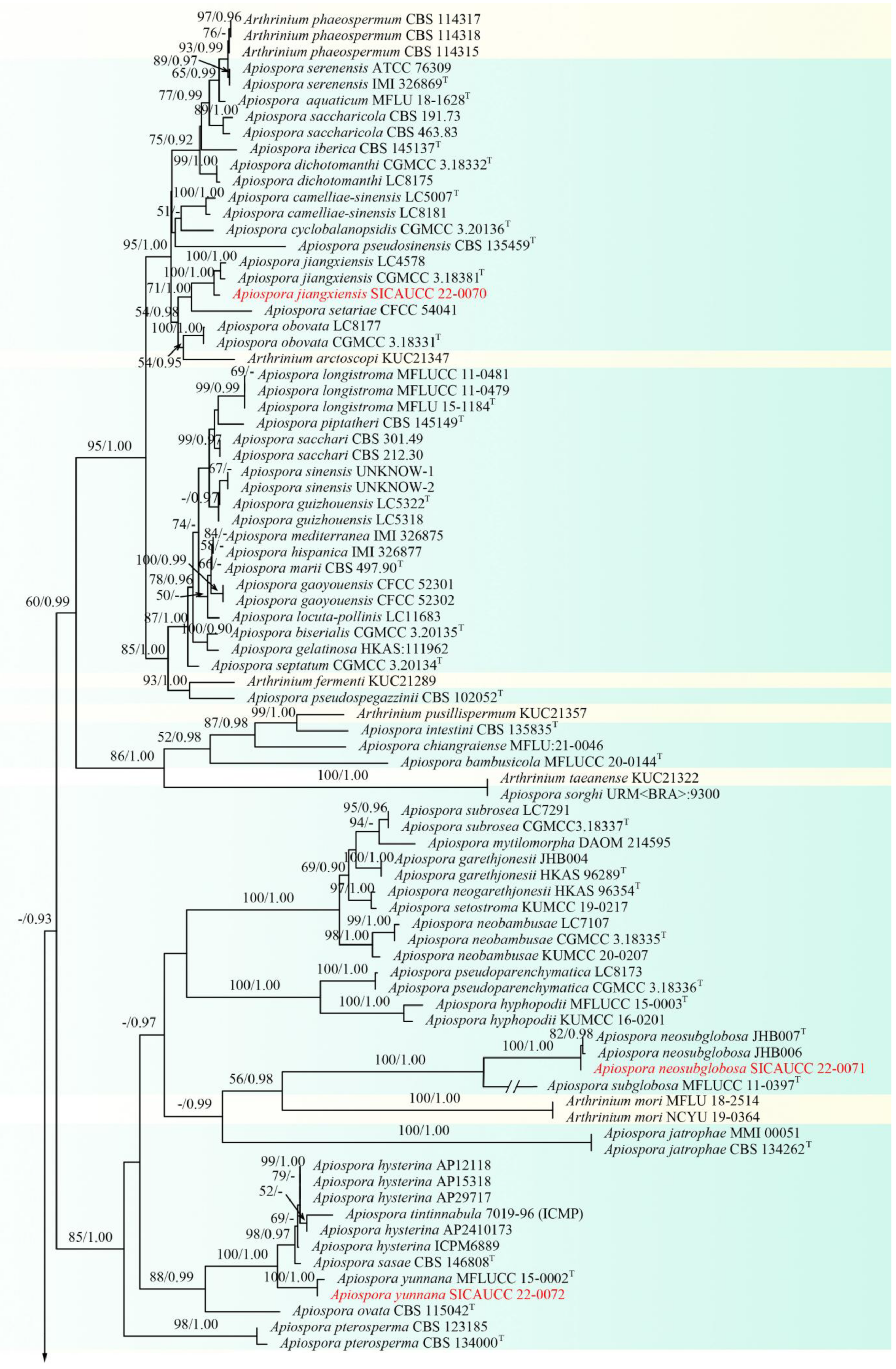
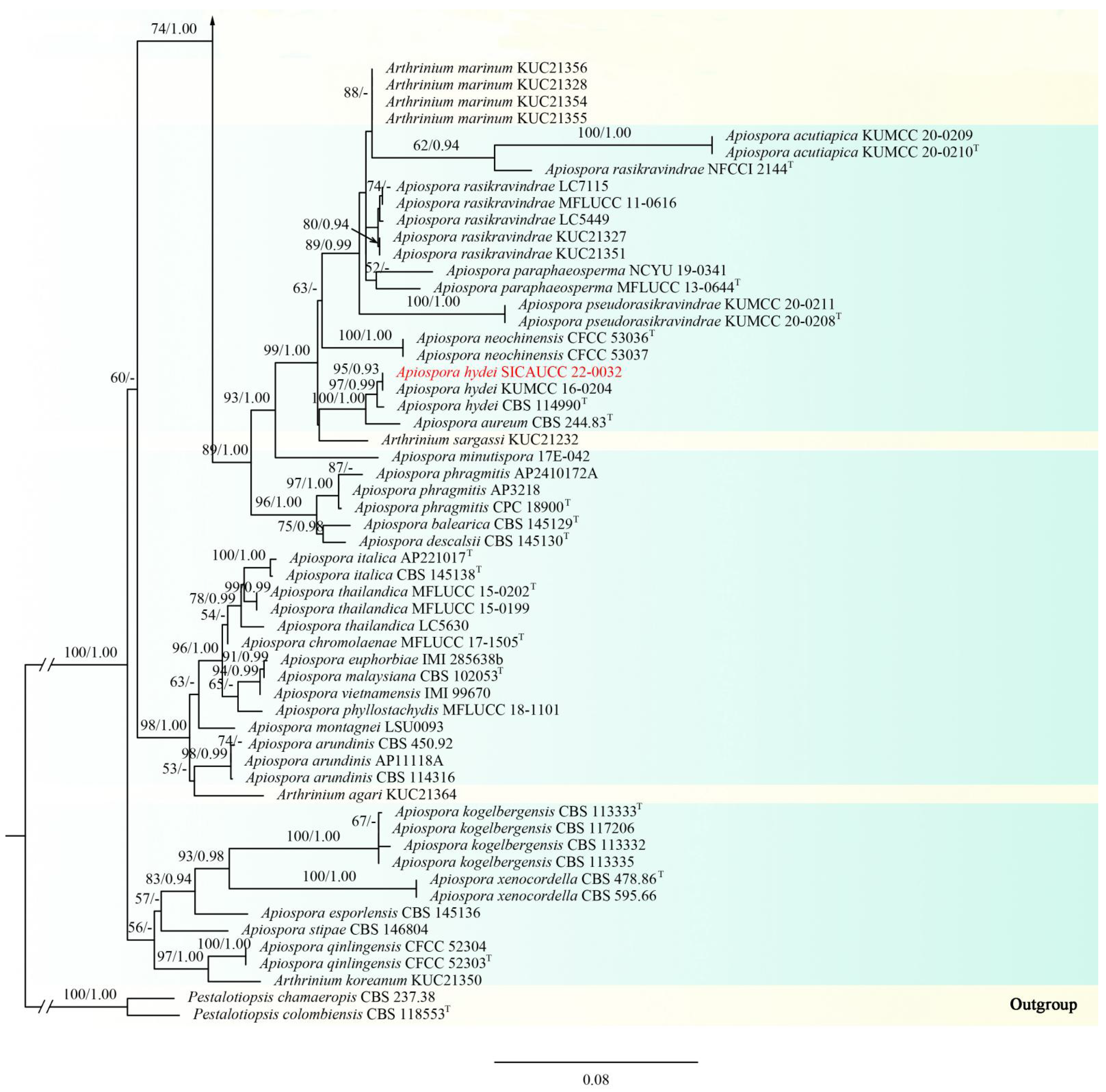
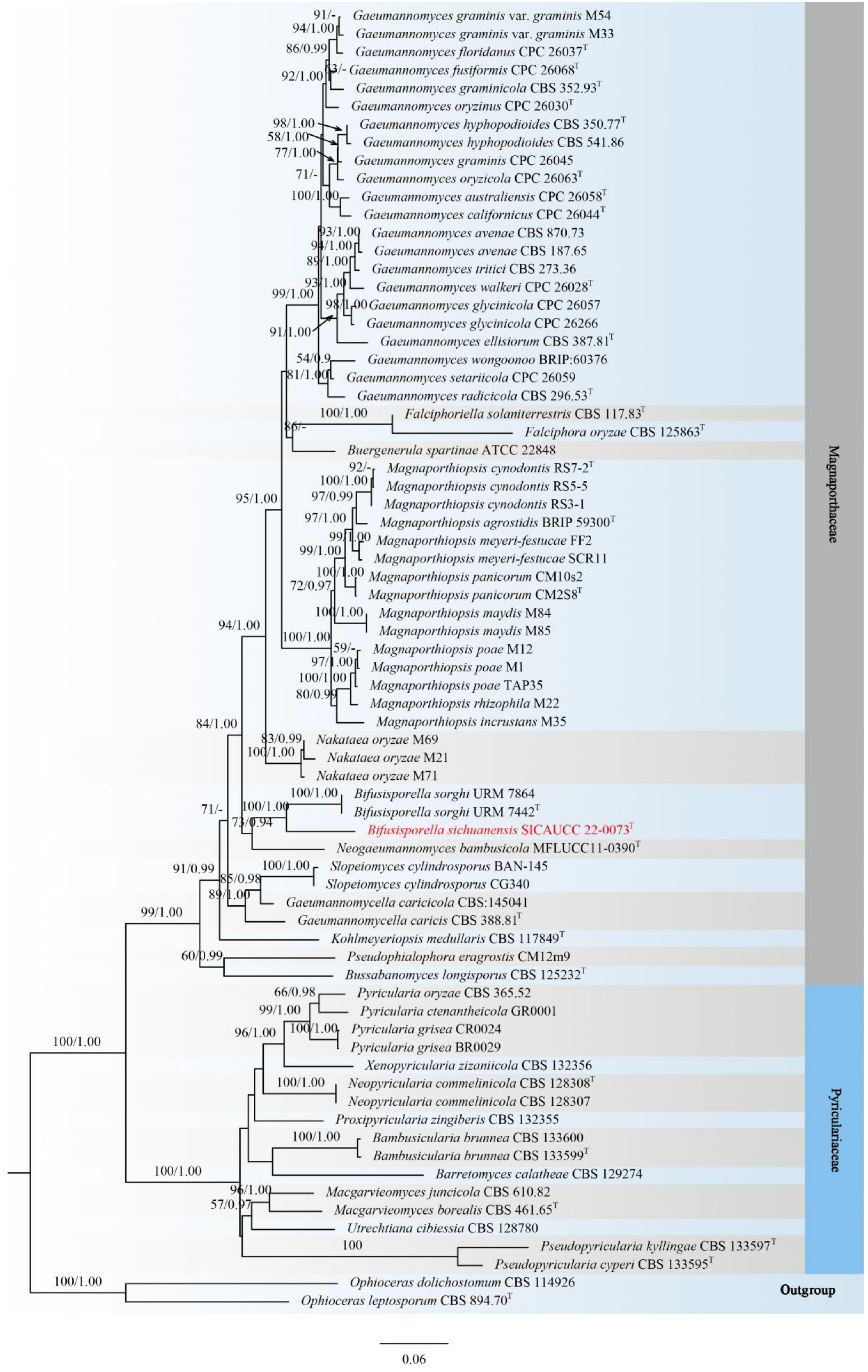
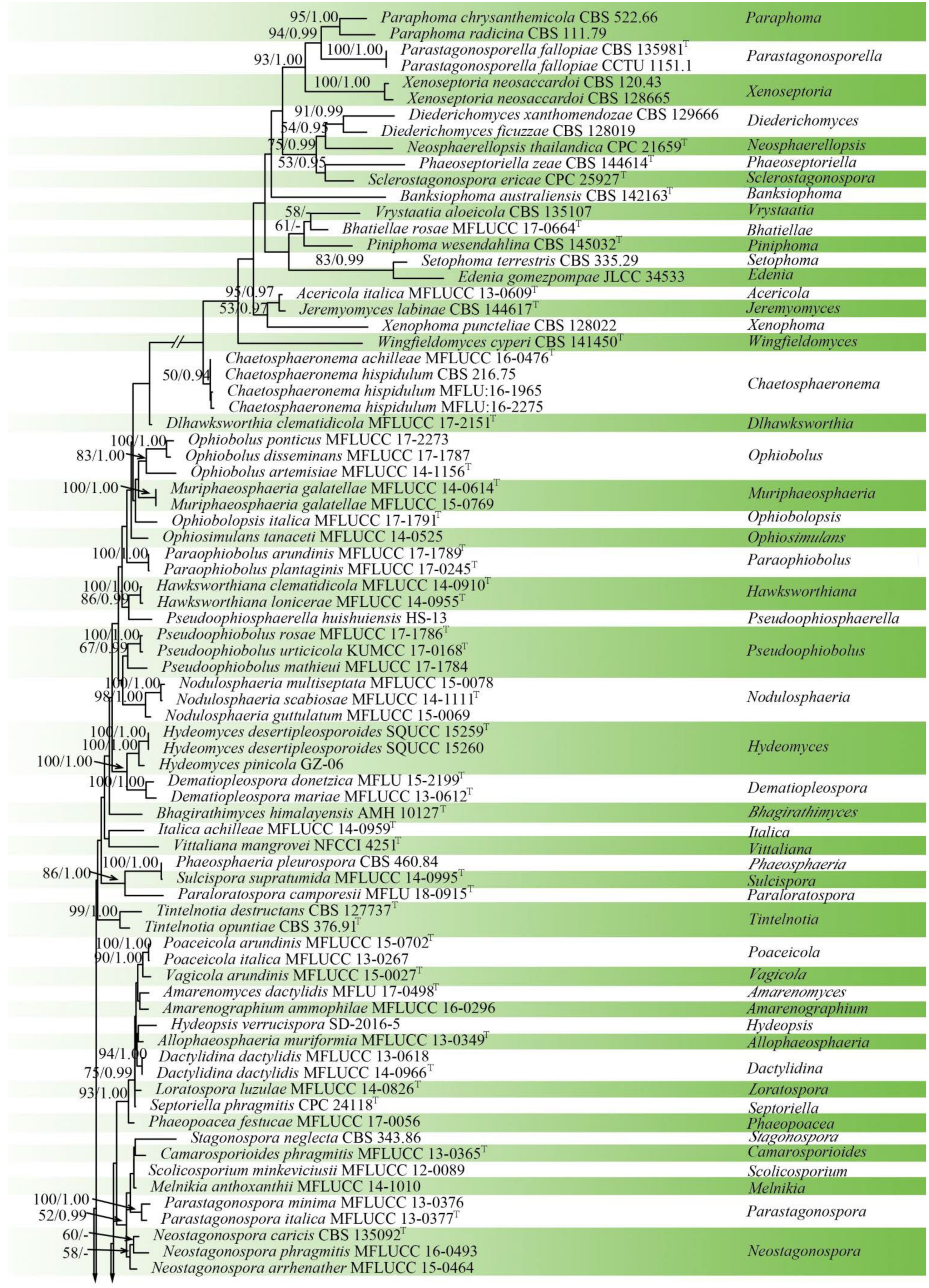
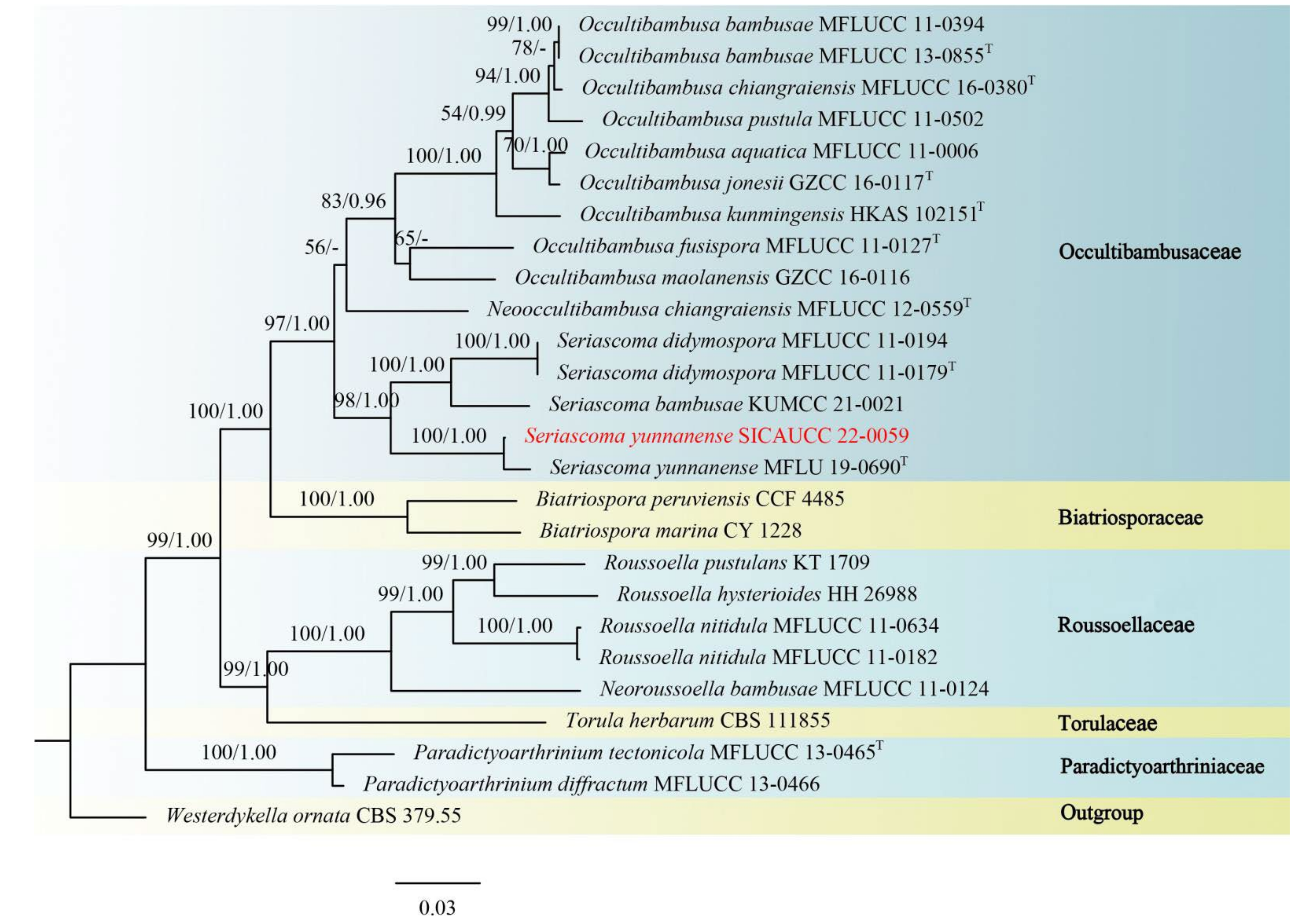
3.1. Phylogenetic Analyses
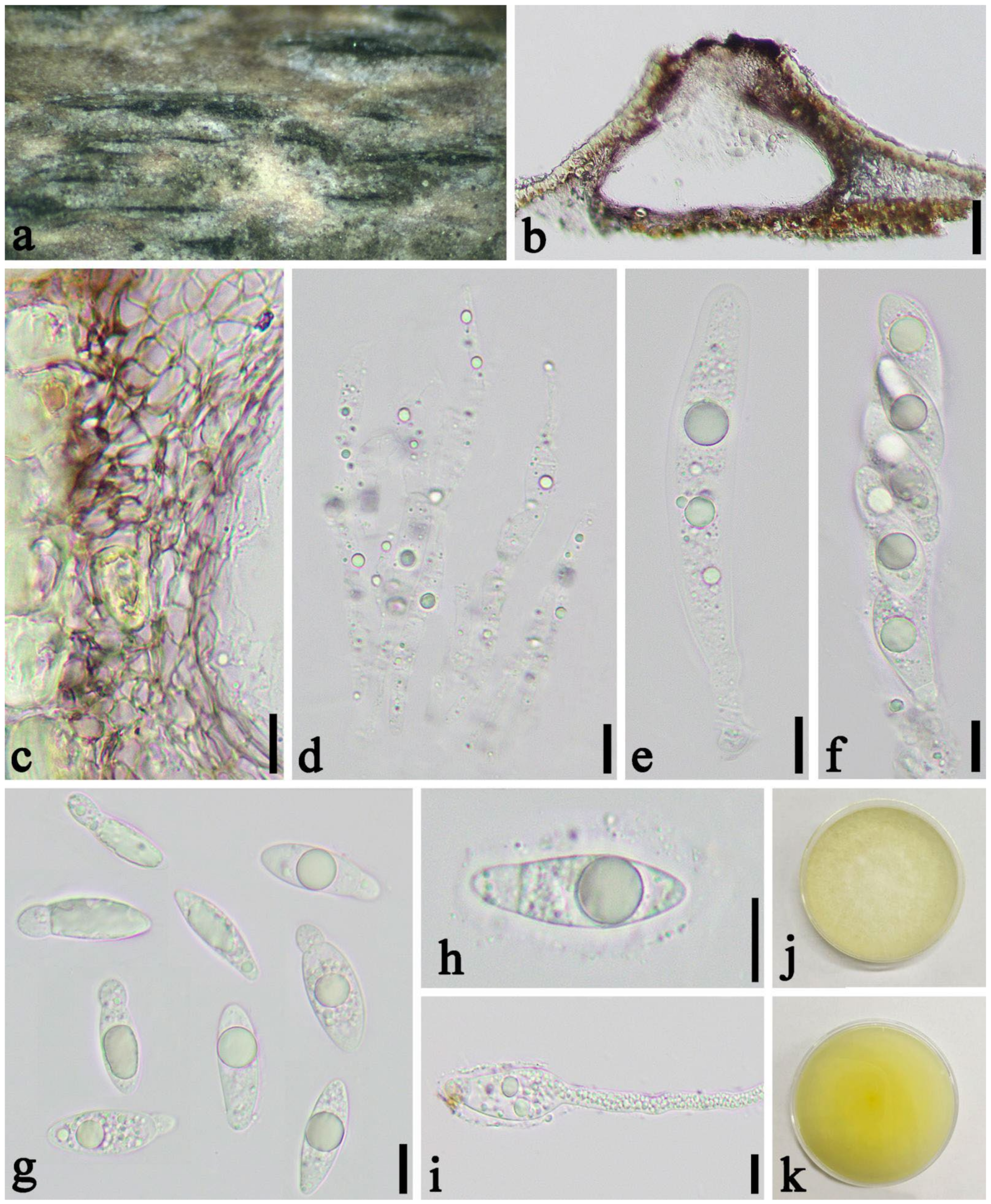
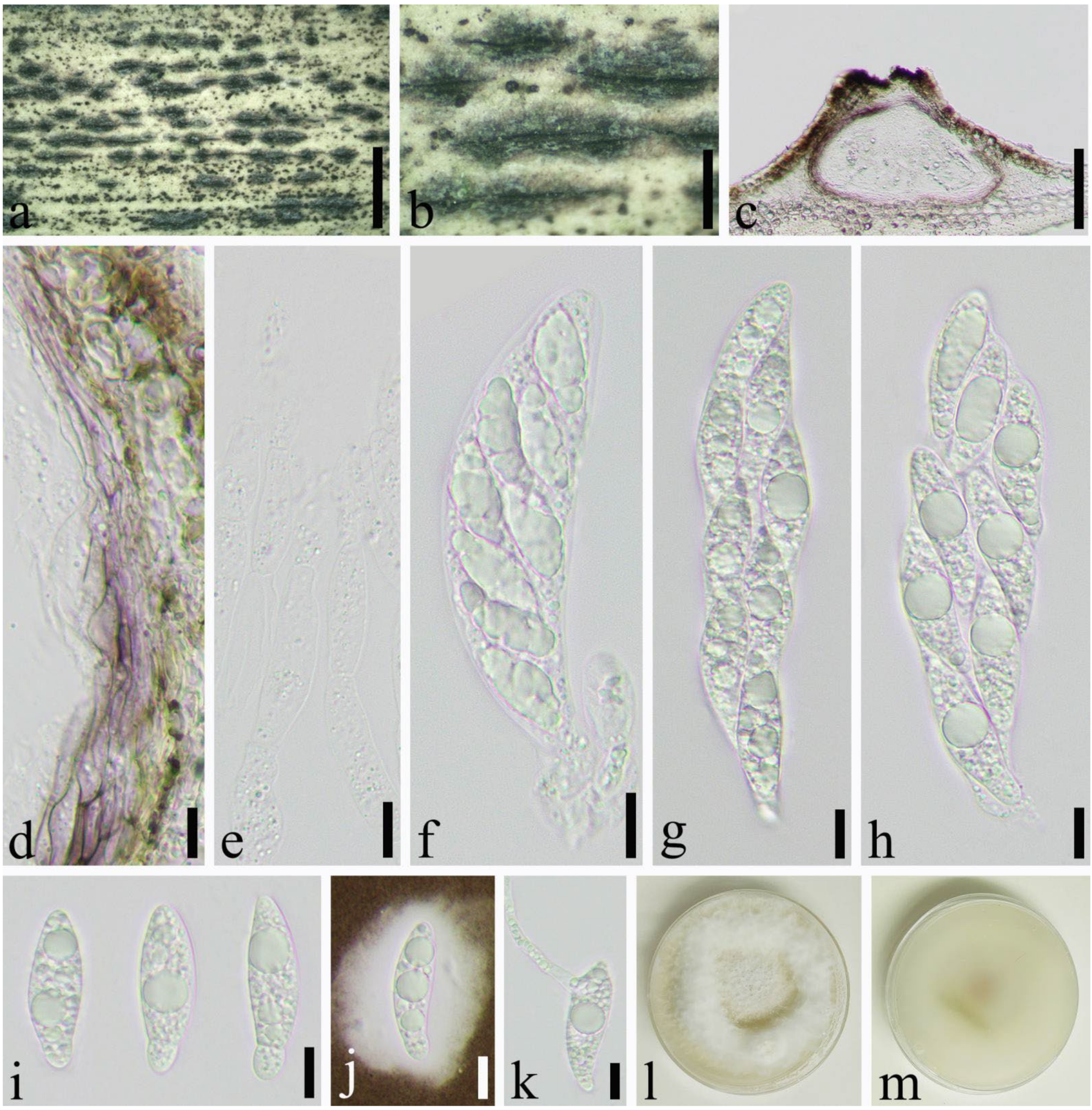
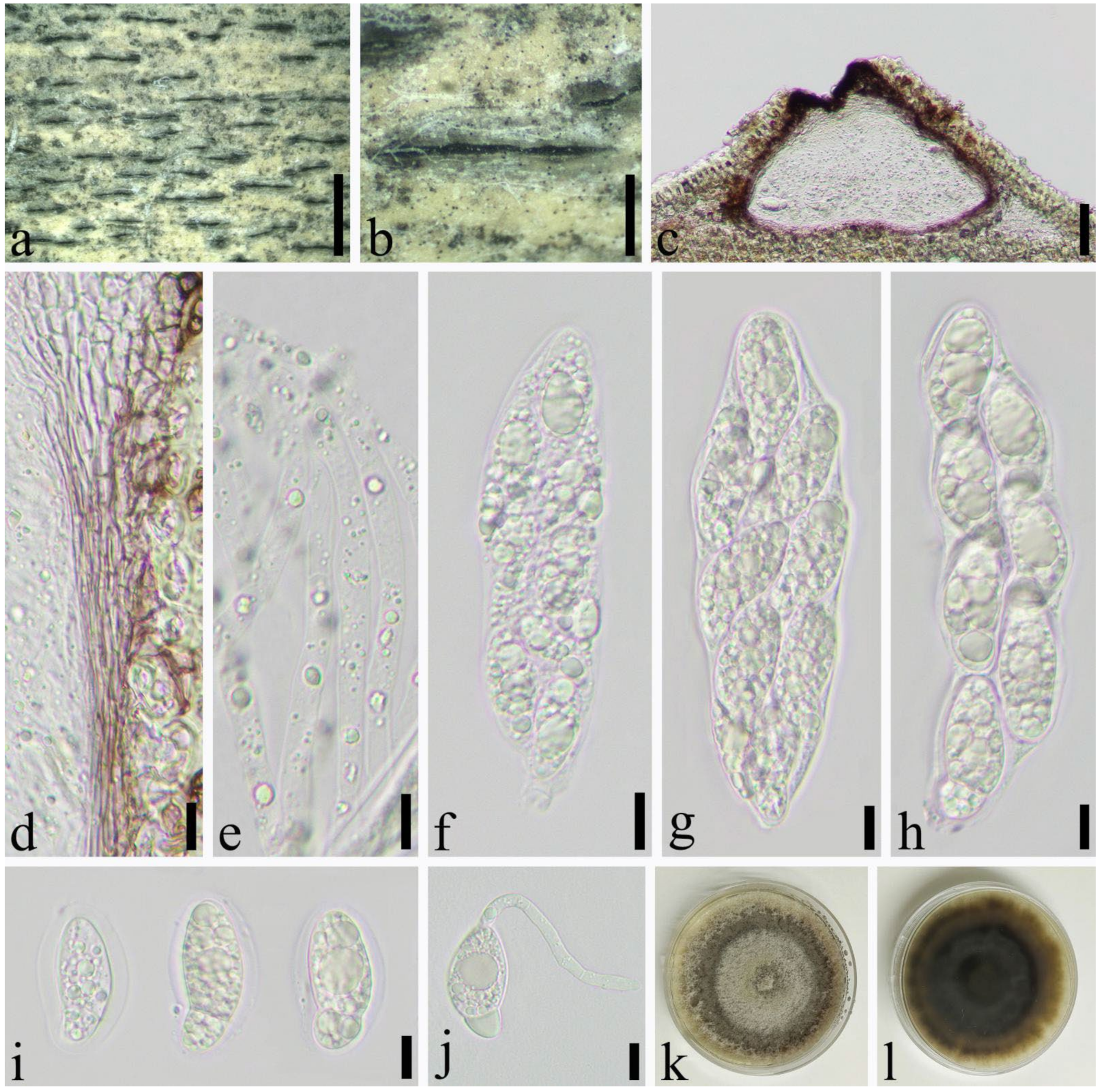
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Molecular Data Used in This Study and GenBank Accession Numbers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | GenBank Accession Numbers | |||||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | LSU | tub2 | tef1-a | rpb1 | SSU | rpb2 | References | ||
| Apiospora acutiapica | KUMCC 20-0209 | MT946342 | MT946338 | – | – | – | – | – | [71] |
| Apiospora acutiapica | KUMCC 20-0210 T | MT946343 | MT946339 | – | – | – | – | [71] | |
| Apiospora aquaticum | MFLU 18-1628 T | MK828608 | MK835806 | – | – | – | – | – | [55] |
| Apiospora arundinis | CBS 114316 | KF144884 | KF144928 | KF144974 | KF145016 | – | – | – | [40] |
| Apiospora arundinis | CBS 450.92 | AB220259 | – | AB220306 | – | – | – | – | [71] |
| Apiospora arundinis | AP11118A | MK014868 | MK014835 | MK017974 | MK017945 | – | – | – | [72] |
| Apiospora aureum | CBS 244.83 T | AB220251 | KF144935 | KF144981 | KF145023 | – | – | – | NCBI |
| Apiospora balearica | CBS 145129 T | MK014869 | MK014836 | MK017975 | MK017946 | – | – | – | [72] |
| Apiospora bambusicola | MFLUCC 20-0144 T | MW173030 | MW173087 | – | MW183262 | – | – | – | [73] |
| Apiospora biserialis | CGMCC 3.20135 T | MW481708 | – | MW522955 | MW522938 | – | – | – | [52] |
| Apiospora camelliae-sinensis | LC5007 T | KY494704 | KY494780 | KY705173 | KY705103 | – | – | – | [36] |
| Apiospora camelliae-sinensis | LC8181 | KY494761 | KY494837 | KY705229 | KY705157 | – | – | – | [36] |
| Apiospora chiangraiense | MFLU:21-0046 | MZ542520 | MZ542524 | MZ546409 | – | – | – | – | [49] |
| Apiospora chromolaenae | MFLUCC 17-1505 T | MT214342 | MT214436 | – | – | – | – | – | [74] |
| Apiospora cyclobalanopsidis | CGMCC 3.20136 T | MW481713 | – | MW522962 | MW522945 | – | – | – | [52] |
| Apiospora descalsii | CBS 145130 T | MK014870 | MK014837 | MK017976 | MK017947 | – | – | – | [72] |
| Apiospora dichotomanthi | CGMCC 3.18332 T | KY494697 | KY494832 | KY705167 | KY705096 | – | – | – | [36] |
| Apiospora dichotomanthi | LC8175 | KY494755 | KY494831 | KY705223 | KY705151 | – | – | – | [36] |
| Apiospora esporlensis | CBS 145136 T | MK014878 | MK014845 | MK017983 | MK017954 | – | – | – | [72] |
| Apiospora euphorbiae | IMI 285638b | AB220241 | – | AB220288 | – | – | – | – | [71] |
| Apiospora gaoyouensis | CFCC 52301 | MH197124 | – | MH236789 | MH236793 | – | – | – | [53] |
| Apiospora gaoyouensis | CFCC 52302 | MH197125 | – | MH236790 | MH236794 | – | – | – | [53] |
| Apiospora garethjonesii | JHB004 | KY356086 | KY356091 | – | – | – | – | – | [41] |
| Apiospora garethjonesii | HKAS 96289 T | NR_154736 | NG_057131 | – | – | – | – | – | [41] |
| Apiospora gelatinosa | HKAS:111962 | – | – | MW5229 | MW522941 | – | – | – | [52] |
| Apiospora guizhouensis | LC5318 | KY494708 | KY494784 | KY705177 | KY705107 | – | – | – | [36] |
| Apiospora guizhouensis | CGMCC 3.18334 T = LC5322 | KY494709 | KY494785 | KY705178 | KY705108 | – | – | – | [36] |
| Apiospora hispanica | IMI 326877 | AB220242 | AB220336 | AB220289 | – | – | – | – | [71] |
| Apiospora hydei | CBS 114990 T | KF144890 | KF144936 | KF144982 | KF145024 | – | – | – | [40] |
| Apiospora hydei | KUMCC 16-0204 | KY356087 | KY356092 | – | – | – | – | – | [41] |
| Apiospora hydei | SICAUCC 22-0032 | ON183998 | ON185553 | ON221313 | ON221312 | – | – | – | This study |
| Apiospora hyphopodii | MFLUCC 15-003 T | KR069110 | KR069111 | – | – | – | – | – | [75] |
| Apiospora hyphopodii | KUMCC 16-0201 | KY356088 | KY356093 | – | – | – | – | – | [41] |
| Apiospora hysterina | AP15318 | MK014873 | MK014840 | MK017979 | MK017950 | – | – | – | [72] |
| Apiospora hysterina | ICPM6889 | MK014874 | MK014841 | MK017980 | MK017951 | – | – | – | [72] |
| Apiospora hysterina | AP29717 | MK014875 | MK014842 | MK017981 | MK017952 | – | – | – | [72] |
| Apiospora hysterina | AP2410173 | MK014876 | MK014843 | – | – | – | – | – | [72] |
| Apiospora hysterina | AP12118 | MK014877 | MK014844 | MK017982 | MK017953 | – | – | – | [72] |
| Apiospora iberica | CBS 145137 T | MK014879 | MK014846 | MK017984 | MK017955 | – | – | – | [72] |
| Apiospora intestini | CBS 135835 T | KR011352 | MH877577 | KR011350 | KR011351 | – | – | – | [76] |
| Apiospora italica | CBS 145138 T | MK014880 | MK014847 | MK017985 | MK017956 | – | – | – | [72] |
| Apiospora italica | AP221017 T | MK014881 | MK014848 | MK017986 | MK017957 | – | – | – | [72] |
| Apiospora jatrophae | CBS 134262 T | NR_154675 | – | – | – | – | – | – | [77] |
| Apiospora jatrophae | MMI 00051 = CBS:134262 | JQ246355 | – | – | – | – | – | – | [77] |
| Apiospora jiangxiensis | CGMCC 3.18381 T | KY494693 | – | KY705163 | KY705092 | – | – | – | [36] |
| Apiospora jiangxiensis | LC4578 | KY494694 | KY494770 | KY705164 | KY705093 | – | – | – | [36] |
| Apiospora jiangxiensis | SICAUCC 22-0070 | ON227094 | ON227098 | ON244432 | ON244431 | – | – | – | This study |
| Apiospora kogelbergensis | CBS 113332 | KF144891 | KF144937 | KF144983 | KF145025 | – | – | – | [40] |
| Apiospora kogelbergensis | CBS 113333 T | KF144892 | KF144938 | KF144984 | KF145026 | – | – | – | [40] |
| Apiospora kogelbergensis | CBS 113335 | KF144893 | KF144939 | KF144985 | KF145027 | – | – | – | [40] |
| Apiospora kogelbergensis | CBS 117206 | KF144895 | KF144941 | KF144987 | KF145029 | – | – | – | [40] |
| Apiospora locuta-pollinis | LC11683 | MF939595 | – | MF939622 | MF939616 | – | – | – | [78] |
| Apiospora longistroma | MFLUCC 11-0479 | KU940142 | KU863130 | – | – | – | – | – | [16] |
| Apiospora longistroma | MFLUCC 11-0481 | KU940141 | KU863129 | – | – | – | – | – | [16] |
| Apiospora longistroma | MFLU 15-1184 T | NR_154716 | – | – | – | – | – | – | [16] |
| Apiospora malaysiana | CBS 102053 T | KF144896 | KF144942 | KF144988 | KF145030 | – | – | – | [40] |
| Apiospora marii | CBS 497.90 T | AB220252 | KF144947 | KF144993 | KF145035 | – | – | – | [40] |
| Apiospora mediterranea | IMI 326875 | AB220243 | – | AB220290 | – | – | – | – | [71] |
| Apiospora minutispora | 17E-042 | LC517882 | – | LC518888 | LC518889 | – | – | – | [79] |
| Apiospora montagnei | LSU0093 | MT000394 | MT000490 | – | – | – | – | – | [80] |
| Apiospora mytilomorpha | DAOM 214595 | KY494685 | – | – | – | – | – | – | [36] |
| Apiospora neobambusae | CGMCC 3.18335 T | KY494718 | KY494794 | KY705186 | KY806204 | – | – | – | [36] |
| Apiospora neobambusae | KUMCC 20-0207 | MT946346 | MT946340 | – | – | – | – | – | [71] |
| Apiospora neobambusae | LC7107 | KY494719 | KY494795 | KY705187 | KY705117 | – | – | – | [36] |
| Apiospora neochinensis | CFCC 53037 | MK819292 | – | MK818548 | MK818546 | – | – | – | [81] |
| Apiospora neochinensis | CFCC 53036 T | MK819291 | – | MK818547 | MK818545 | – | – | – | [81] |
| Apiospora neogarethjonesii | HKAS 96354 T | MK070897 | MK070898 | – | – | – | – | – | [82] |
| Apiospora neosubglobosa | JHB006 | KY356089 | KY356094 | – | – | – | – | – | [41] |
| Apiospora neosubglobosa | JHB007 T | KY356090 | KY356095 | – | – | – | – | – | [41] |
| Apiospora neosubglobosa | SICAUCC 22-0071 | ON227095 | ON227099 | ON244430 | ON244429 | – | – | – | This study |
| Apiospora obovata | CGMCC 3.18331 T | KY494696 | KY494834 | KY705166 | KY705095 | – | – | – | [41] |
| Apiospora obovata | LC8177 | KY494757 | KY494833 | KY705225 | KY705153 | – | – | – | [41] |
| Apiospora ovata | CBS 115042 T | KF144903 | KF144950 | KF144995 | KF145037 | – | – | – | [40] |
| Arthrinium paraphaeospermum | NCYU 19-0341 | MW114315 | MW293936 | – | MW288020 | – | – | – | NCBI |
| Apiospora paraphaeosperma | MFLUCC 13-0644 T | KX822128 | KX822124 | – | – | – | – | – | [71] |
| Apiospora phragmitis | CPC 18900 T | KF144909 | – | KF145001 | KF145043 | – | – | – | [40] |
| Apiospora phragmitis | AP3218 | MK014891 | MK014858 | MK017996 | MK017967 | – | – | – | [72] |
| Apiospora phragmitis | AP2410172A | MK014890 | MK014857 | MK017995 | MK017966 | – | – | – | [72] |
| Apiospora phyllostachydis | MFLUCC 18-1101 | – | – | MK291949 | – | – | – | – | [65] |
| Apiospora piptatheri | CBS 145149 T | MK014893 | MK014860 | – | MK017969 | – | – | – | [72] |
| Apiospora pseudoparenchymatica | CGMCC 3.18336 T | KY494743 | KY494819 | KY705211 | KY705139 | – | – | – | [36] |
| Apiospora pseudoparenchymatica | LC8173 | KY494753 | KY494829 | KY705221 | KY705149 | – | – | – | [36] |
| Apiospora pseudorasikravindrae | KUMCC 20-0208 T | MT946344 | – | – | – | – | – | – | [71] |
| Apiospora pseudorasikravindrae | KUMCC 20-0211 | MT946345 | – | – | – | – | – | – | [71] |
| Apiospora pseudosinensis | CBS 135459 T | KF144910 | KF144957 | – | KF145044 | – | – | – | [40] |
| Apiospora pseudospegazzinii | CBS 102052 T | KF144911 | KF144958 | KF145002 | KF145045 | – | – | – | [40] |
| Apiospora pterosperma | CBS 123185 | KF144912 | KF144959 | KF145003 | – | – | – | [40] | |
| Apiospora pterosperma | CBS 134000 T | KF144913 | KF144960 | KF145004 | KF145046 | – | – | – | [40] |
| Apiospora qinlingensis | CFCC 52303 T | MH197120 | – | MH236791 | MH236795 | – | – | – | [53] |
| Apiospora qinlingensis | CFCC 52304 | MH197121 | – | MH236792 | MH236796 | – | – | – | [53] |
| Apiospora rasikravindrae | NFCCI 2144 T | KF144914 | – | – | – | – | – | – | [83] |
| Apiospora rasikravindrae | MFLUCC 11-0616 | KU940144 | KU863132 | – | – | – | – | – | [16] |
| Apiospora rasikravindrae | LC5449 | KY494713 | KY494789 | KY705182 | KY705112 | – | – | – | [36] |
| Apiospora rasikravindrae | LC7115 | KY494721 | KY494797 | KY705189 | KY705118 | – | – | – | [36] |
| Apiospora rasikravindrae | KUC21351 | MH498540 | MH498498 | MN868932 | – | – | – | [84] | |
| Apiospora rasikravindrae | KUC21327 | MH498541 | MH498499 | MH544670 | – | – | – | [84] | |
| Apiospora sacchari | CBS 212.30 | KF144916 | KF144962 | KF145005 | KF145047 | – | – | – | [40] |
| Apiospora sacchari | CBS 301.49 | KF144917 | KF144963 | KF145006 | KF145048 | – | – | – | [40] |
| Apiospora saccharicola | CBS 191.73 | KF144920 | KF144966 | KF145009 | KF145051 | – | – | – | [40] |
| Apiospora saccharicola | CBS 463.83 | KF144921 | KF144968 | KF145010 | KF145052 | – | – | – | [40] |
| Apiospora sasae | CBS 146808 T | MW883402 | MW883797 | MW890120 | MW890104 | – | – | – | [85] |
| Apiospora septatum | CGMCC 3.20134 T | MW481711 | – | MW522960 | MW522943 | – | – | – | [52] |
| Apiospora serenensis | IMI 326869 T | AB220250 | – | AB220297 | – | – | – | – | [71] |
| Apiospora serenensis | ATCC 76309 | AB220240 | – | AB220287 | – | – | – | – | [71] |
| Apiospora setariae | CFCC 54041 | MT492004 | – | MT497466 | MW118456 | – | – | – | [86] |
| Apiospora setostroma | KUMCC 19-0217 | MN528012 | MN528011 | – | MN527357 | – | – | – | [87] |
| Apiospora sinensis | UNKNOW-1 = HKUCC 3143 | – | AY083831 | – | – | – | – | – | NCBI |
| Apiospora sinensis | UNKNOW-2 | – | DQ810215 | – | – | – | – | – | NCBI |
| Apiospora sorghi | URM<BRA>:9300 | MK371706 | – | – | – | – | – | – | NCBI |
| Apiospora stipae | CBS 146804 | MW883403 | MW883798 | MW890121 | MW890105 | – | – | – | [85] |
| Apiospora subglobosa | MFLUCC 11-0397 T | KR069112 | KR069113 | – | – | – | – | – | [75] |
| Apiospora subrosea | LC7291 | KY494751 | KY494827 | KY705219 | KY705147 | – | – | – | [36] |
| Apiospora subrosea | CGMCC3.18337 T | KY494752 | KY494828 | KY705220 | KY705148 | – | – | – | [36] |
| Apiospora thailandica | MFLUCC 15-0199 | KU940146 | KU863134 | – | – | – | – | – | [16] |
| Apiospora thailandica | MFLUCC 15-0202 T | KU940145 | KU863133 | – | – | – | – | – | [16] |
| Apiospora thailandica | LC5630 | KY494714 | KY494790 | KY806200 | KY705113 | – | – | – | [36] |
| Apiospora tintinnabula | 7019-96 (ICMP) | – | DQ810216 | – | – | – | – | – | [71] |
| Apiospora vietnamensis | IMI 99670 | KX986096 | KX986111 | KY019466 | – | – | – | – | [88] |
| Apiospora xenocordella | CBS 478.86 T | KF144925 | KY494763 | – | – | – | – | – | [40] |
| Apiospora xenocordella | CBS 595.66 | KF144926 | KF144971 | KF145013 | KF145055 | – | – | – | [40] |
| Apiospora yunnana | MFLUCC 15-0002 T | KU940147 | KU863135 | – | – | – | – | – | [16] |
| Apiospora yunnana | SICAUCC 22-0072 | ON227096 | ON227100 | ON244426 | ON244425 | – | – | – | This study |
| Arthrinium agari | KUC21364 | MH498516 | – | MH498474 | MN868917 | – | – | – | [84] |
| Arthrinium arctoscopi | KUC21347 | MH498525 | – | MH498483 | MN868922 | – | – | – | [84] |
| Arthrinium fermenti | KUC21289 | MF615226 | – | MF615231 | MH544667 | – | – | – | [84] |
| Arthrinium koreanum | KUC21350 | MH498521 | – | MH498479 | MN868929 | – | – | – | [84] |
| Arthrinium marinum | KUC21328 | MH498538 | MH498496 | MH544669 | [84] | ||||
| Arthrinium marinum | KUC21356 | MH498534 | – | MH498492 | MN868926 | – | – | – | [84] |
| Arthrinium marinum | KUC21355 | MH498535 | MH498493 | MN868925 | [84] | ||||
| Arthrinium marinum | KUC21354 | MH498536 | MH498494 | MN868924 | [84] | ||||
| Arthrinium mori | MFLU 18-2514 | MW114313 | MW114393 | – | – | – | – | – | [89] |
| Arthrinium mori | NCYU 19-0364 | MW114314 | MW114394 | – | – | – | – | – | [89] |
| Arthrinium phaeospermum | CBS 114315 | KF144905 | KF144952 | KF144997 | KF145039 | – | – | – | [40] |
| Arthrinium phaeospermum | CBS 114317 | KF144906 | KF144953 | KF144998 | KF145040 | – | – | – | [40] |
| Arthrinium phaeospermum | CBS 114318 | KF144907 | KF144954 | KF144999 | KF145041 | – | – | – | [40] |
| Arthrinium pusillispermum | KUC21357 | MH498532 | – | MH498490 | MN868931 | – | – | – | [84] |
| Arthrinium sargassi | KUC21232 | KT207750 | – | KT207648 | MH544676 | – | – | – | [84] |
| Arthrinium taeanense | KUC21322 | MH498515 | – | MH498473 | MH544662 | – | – | – | [84] |
| Pestalotiopsis chamaeropis | CBS 237.38 | MH855954 | MH867450 | KM199392 | KM199474 | – | – | – | [76] |
| Pestalotiopsis colombiensis | CBS 118553 T | KM199307 | KM116222 | KM199421 | KM199488 | – | – | – | [90] |
| Bambusicularia brunnea | CBS 133599 T | KM484830 | KM484948 | – | – | KM485043 | – | – | [91] |
| Bambusicularia brunnea | CBS 133600 | AB274436 | KM484949 | – | – | KM485044 | – | – | [91,92] |
| Barretomyces calatheae | CBS 129274 = CPC 18464 | KM484831 | KM484950 | – | – | KM485045 | – | – | [76] |
| Bifusisporella sichuanensis | SICAUCC 22-0073T | ON227097 | ON227101 | – | ON244427 | ON244428 | – | – | This study |
| Bifusisporella sorghi | URM 7442 T | MK060155 | MK060153 | – | MK060157 | MK060159 | – | – | [42] |
| Bifusisporella sorghi | URM 7864 | MK060156 | MK060154 | – | MK060158 | MK060160 | – | – | [42] |
| Buergenerula spartinae | ATCC 22848 | JX134666 | DQ341492 | – | JX134692 | JX134720 | – | – | [93] |
| Bussabanomyces longisporus | CBS 125232 T | KM484832 | KM484951 | – | KM009202 | KM485046 | – | – | [94] |
| Falciphora oryzae | CBS 125863 T | EU636699 | KJ026705 | – | JN857963 | KJ026706 | – | – | [95] |
| Falciphoriella solaniterrestris | CBS 117.83 T | KM484842 | KM484959 | – | – | KM485058 | – | – | [91] |
| Gaeumannomycella caricicola | CBS:145041 | MK442584 | MK442526 | – | – | – | – | – | [96] |
| Gaeumannomycella caricis | CBS 388.81 T | KM484843 | KM484960 | – | KX306674 | – | – | – | [91] |
| Gaeumannomyces australiensis | CPC 26058 T | KX306480 | KX306550 | – | KX306683 | KX306619 | – | – | [97] |
| Gaeumannomyces avenae | CBS 187.65 | JX134668 | JX134680 | – | – | JX134722 | – | – | [93] |
| Gaeumannomyces avenae | CBS 870.73 = DAR 20999 | KM484833 | DQ341495 | – | – | KM485048 | – | – | [91] |
| Gaeumannomyces californicus | CPC 26044 T | KX306490 | KX306560 | – | KX306691 | KX306625 | – | – | [97] |
| Gaeumannomyces ellisiorum | CBS 387.81 T | KM484835 | KM484952 | – | KX306692 | KM485051 | – | – | [91] |
| Gaeumannomyces floridanus | CPC 26037 T | KX306491 | KX306561 | – | KX306693 | KX306626 | – | – | [97] |
| Gaeumannomyces fusiformis | CPC 26068 T | KX306492 | KX306562 | – | KX306694 | KX306627 | – | – | [97] |
| Gaeumannomyces glycinicola | CPC 26266 | KX306494 | KX306564 | – | KX306696 | KX306629 | – | – | [97] |
| Gaeumannomyces glycinicola | CPC 26057 | KX306493 | KX306563 | – | KX306695 | KX306628 | – | – | [97] |
| Gaeumannomyces graminicola | CBS 352.93 T | KM484834 | DQ341496 | – | KX306697 | KM485050 | – | – | [91] |
| Gaeumannomyces graminis | CPC 26045 | KX306505 | KX306575 | – | KX306708 | KX306640 | – | – | [97] |
| Gaeumannomyces graminis var. graminis | M33 | JF710374 | JF414896 | – | JF710411 | JF710442 | – | – | [98] |
| Gaeumannomyces graminis var. graminis | M54 | JF414848 | JF414898 | – | JF710419 | JF710444 | – | – | [98] |
| Gaeumannomyces hyphopodioides | CBS 350.77 T | KX306506 | KX306576 | – | – | – | – | – | [97] |
| Gaeumannomyces hyphopodioides | CBS 541.86 | KX306507 | KX306577 | – | KX306709 | – | – | – | [97] |
| Gaeumannomyces oryzicola | CPC 26063 T | KX306516 | KX306586 | – | KX306717 | KX306646 | – | – | [97] |
| Gaeumannomyces oryzinus | CPC 26030 T | KX306517 | KX306587 | – | KX306718 | KX306647 | – | – | [97] |
| Gaeumannomyces radicicola | CBS 296.53 T | KM009170 | KM009158 | – | KM009206 | KM009194 | – | – | [94] |
| Gaeumannomyces setariicola | CPC 26059 | KX306524 | KX306594 | – | KX306725 | KX306654 | – | – | [97] |
| Gaeumannomyces tritici | CBS 273.36 | KX306525 | KX306595 | – | KX306729 | KX306655 | – | – | [97] |
| Gaeumannomyces walkeri | CPC 26028 T | KX306543 | KX306613 | – | KX306746 | KX306670 | – | – | [97] |
| Gaeumannomyces wongoonoo | BRIP:60376 | KP162137 | KP162146 | – | – | – | – | – | [99] |
| Kohlmeyeriopsis medullaris | CBS 117849 T = JK5528S | KM484852 | KM484968 | – | – | KM485068 | – | – | [91] |
| Macgarvieomyces borealis | CBS 461.65 T | MH858669 | DQ341511 | – | KM009198 | KM485070 | – | – | [94] |
| Macgarvieomyces juncicola | CBS 610.82 | KM484855 | KM484970 | – | KM009201 | KM485071 | – | – | [91] |
| Magnaporthiopsis agrostidis | BRIP 59300 T | KT364753 | KT364754 | – | KT364756 | KT364755 | – | – | [100] |
| Magnaporthiopsis cynodontis | RS7-2 = CBS 141700 T | KJ855508 | KM401648 | – | KP282714 | KP268930 | – | – | [101] |
| Magnaporthiopsis cynodontis | RS5-5 | KJ855506 | KM401646 | – | KP282712 | KP268928 | – | – | [101] |
| Magnaporthiopsis cynodontis | RS3-1 | KJ855505 | KM401645 | – | KP282711 | KP268927 | – | – | [101] |
| Magnaporthiopsis incrustans | M35 | JF414843 | JF414892 | – | JF710412 | JF710437 | – | – | [98] |
| Magnaporthiopsis maydis | M84 | KM009160 | KM009148 | – | KM009196 | KM009184 | – | – | [94] |
| Magnaporthiopsis maydis | M85 | KM009161 | KM009149 | – | KM009197 | KM009185 | – | – | [94] |
| Magnaporthiopsis meyeri-festucae | FF2 | MF178146 | MF178151 | – | MF178167 | MF178162 | – | – | [102] |
| Magnaporthiopsis meyeri-festucae | SCR11 | MF178150 | MF178155 | – | MF178171 | MF178166 | – | – | [102] |
| Magnaporthiopsis panicorum | CM2S8 T | KF689643 | KF689633 | – | KF689623 | KF689613 | – | – | [103] |
| Magnaporthiopsis panicorum | CM10s2 | KF689644 | KF689634 | – | KF689624 | KF689614 | – | – | [103] |
| Magnaporthiopsis poae | TAP35 | KJ855511 | KM401651 | – | KP282717 | KP268933 | – | – | [104] |
| Magnaporthiopsis poae | M1 | JF414827 | JF414876 | – | JF710400 | JF710425 | – | – | [98] |
| Magnaporthiopsis poae | M12 | JF414828 | JF414877 | – | JF710401 | JF710426 | – | – | [98] |
| Magnaporthiopsis rhizophila | M22 | JF414833 | JF414882 | – | JF710407 | JF710431 | – | – | [98] |
| Nakataea oryzae | M21 | JF414838 | JF414887 | – | JF710406 | JF710441 | – | – | [98] |
| Nakataea oryzae | M69 | JX134672 | JX134685 | – | JX134698 | JX134726 | – | – | [93] |
| Nakataea oryzae | M71 | JX134673 | JX134686 | – | JX134699 | JX134727 | – | – | [93] |
| Neogaeumannomyces bambusicola | MFLUCC11-0390 T | KP744449 | KP744492 | – | – | – | – | – | [105] |
| Neopyricularia commelinicola | CBS 128307 = KACC 44083 | FJ850125 | KM484984 | – | KM009199 | KM485086 | – | – | [91,106] |
| Neopyricularia commelinicola | CBS 128308 T | FJ850122 | KM484985 | – | – | KM485087 | – | – | [91,106] |
| Ophioceras dolichostomum | CBS 114926 = HKUCC 3936 = KM 8 | JX134677 | JX134689 | – | JX134703 | JX134731 | – | – | [93] |
| Ophioceras leptosporum | CBS 894.70 T = ATCC 24161 = HME 2955 | JX134678 | JX134690 | – | JX134704 | JX134732 | – | – | [83] |
| Proxipyricularia zingiberis | CBS 132355 = MAFF 240221 | AB274433 | KM484987 | – | – | KM485090 | – | – | [91] |
| Pseudophialophora eragrostis | CM12m9 | KF689648 | KF689638 | – | KF689628 | KF689618 | – | – | [103] |
| Pseudopyricularia cyperi | CBS 133595 T = MAFF 240229 | KM484872 | KM484990 | – | – | AB818013 | – | – | [91] |
| Pseudopyricularia kyllingae | CBS 133597 T = MAFF 240227 | KM484876 | KM484992 | – | KT950880 | KM485096 | – | – | [91] |
| Pyricularia ctenantheicola | GR0001 = Ct-4 = ATCC 200218 | KM484878 | KM484994 | – | – | KM485098 | – | – | [91] |
| Pyricularia grisea | BR0029 | KM484880 | KM484995 | – | – | KM485100 | – | – | [91] |
| Pyricularia grisea | CR0024 | KM484882 | KM484997 | – | – | KM485102 | – | – | [91] |
| Pyricularia oryzae | CBS 365.52 = MUCL 9451 | KM484890 | KM485000 | – | – | KM485110 | – | – | [76] |
| Slopeiomyces cylindrosporus | BAN-145 | JF508361 | – | – | – | – | – | – | [107] |
| Slopeiomyces cylindrosporus | CG340 | AY428776 | – | – | – | – | – | – | [108] |
| Utrechtiana cibiessia | CBS 128780 = CPC 18916 | JF951153 | JF951176 | – | – | KM485047 | – | – | [76] |
| Xenopyricularia zizaniicola | CBS 132356 | KM484946 | KM485042 | – | KM009203 | KM485160 | – | – | [91] |
| Acericola italica | MFLUCC 13-0609 T | MF167428 | MF167429 | – | – | – | MF167430 | – | [109] |
| Alloneottiosporina thailandica | MFLUCC 15-0576 T | MT177913 | MT177940 | – | MT454002 | – | MT177968 | – | [43] |
| Allophaeosphaeria muriformia | MFLUCC 13-0349 T | KP765680 | KP765681 | – | – | – | KP765682 | – | [105] |
| Amarenographium ammophilae | MFLUCC 16-0296 | KU848196 | KU848197 | – | MG520894 | – | KU848198 | – | [109] |
| Amarenomyces dactylidis | MFLU 17-0498 T | KY775577 | KY775575 | – | – | – | – | – | [110] |
| Ampelomyces quisqualis | CBS 129.79 T | – | EU754128 | – | – | – | EU754029 | – | [111] |
| Banksiophoma australiensis | CBS 142163 T | KY979739 | KY979794 | – | KY979889 | – | – | – | [112] |
| Bhagirathimyces himalayensis | AMH 10127 T = NFCCI 4580 | MK836021 | MK836020 | – | – | – | MN121697 | – | [113] |
| Bhatiellae rosae | MFLUCC 17-0664 T | MG828873 | MG828989 | – | – | – | MG829101 | – | [114] |
| Brunneomurispora lonicerae | KUMCC 18-0157 T | MK356373 | MK356346 | – | MK359065 | – | MK356360 | – | [59] |
| Camarosporioides phragmitis | MFLUCC 13-0365 T | KX572340 | KX572345 | – | KX572354 | – | KX572350 | – | [115] |
| Chaetosphaeronema achilleae | MFLUCC 16-0476 T | KX765265 | KX765266 | – | – | – | – | [115] | |
| Chaetosphaeronema hispidulum | MFLU:16-1965 | MT177915 | MT177942 | – | – | – | MT177970 | – | [43] |
| Chaetosphaeronema hispidulum | MFLU:16-2275 | MT177914 | MT177941 | – | MT454003 | – | MT177969 | – | [43] |
| Chaetosphaeronema hispidulum | CBS 216.75 | KF251148 | KF251652 | – | KF253108 | – | – | [116] | |
| Dactylidina dactylidis | MFLUCC 13-0618 | KP744432 | KP744473 | – | – | – | KP753946 | – | [105] |
| Dactylidina dactylidis | MFLUCC 14-0966 T | MG828886 | MG829002 | – | MG829199 | – | MG829113 | – | [114] |
| Dematiopleospora donetzica | MFLU 15-2199 T | – | MG829005 | – | – | – | MG829116 | – | [114] |
| Dematiopleospora mariae | MFLUCC 13-0612 T | KJ749654 | KJ749653 | – | KJ749655 | – | KJ749652 | – | [117] |
| Diederichomyces ficuzzae | CBS 128019 | KP170647 | – | – | KP170673 | – | – | – | [118] |
| Diederichomyces xanthomendozae | CBS 129666 | KP170651 | – | – | KP170677 | – | – | – | [118] |
| Dlhawksworthia clematidicola | MFLUCC 17-2151 T | MT310619 | MT214574 | – | MT394633 | – | MT226687 | – | [119] |
| Edenia gomezpompae | JLCC 34533 | KC193601 | – | – | – | – | – | – | [120] |
| Elongaticollum hedychii | MFLUCC 18-1638 T | MT321796 | MT321810 | – | MT328753 | – | MT321803 | – | [115] |
| Elongaticollum hedychii | NCYUCC 19-0286 | MT321797 | MT321811 | – | MT328754 | – | MT321804 | – | [115] |
| Embarria clematidis | MFLUCC 14-0652 | KT306949 | KT306953 | – | – | – | KT306956 | – | [121] |
| Embarria clematidis | MFLUCC 14-0976 | MG828871 | MG828987 | – | MG829194 | – | MG829099 | – | [114] |
| Equiseticola fusispora | MFLUCC 14-0522 T | KU987668 | KU987669 | – | MG520895 | – | KU987670 | – | [122] |
| Galliicola pseudophaeosphaeria | MFLUCC 14-0524 | – | – | – | MG520896 | – | – | – | [109] |
| Hawksworthiana clematidicola | MFLUCC 14-0910 T | MG828901 | MG829011 | – | MG829202 | – | MG829120 | – | [114] |
| Hawksworthiana lonicerae | MFLUCC 14-0955 T | MG828902 | MG829012 | – | MG829203 | – | MG829121 | – | [114] |
| Hydeomyces desertipleosporoides | SQUCC 15260 | MK290842 | MK290840 | – | MK290849 | – | MK290844 | – | [123] |
| Hydeomyces desertipleosporoides | SQUCC 15259 T | MK290841 | MK290839 | – | MK290848 | – | MK290843 | – | [123] |
| Hydeomyces pinicola | GZ-06 | MK522506 | MK522496 | – | MK523386 | – | MK522502 | – | [124] |
| Hydeopsis verrucispora | SD-2016-5 | MK522508 | MK522498 | – | MK523388 | – | MK522504 | – | [124] |
| Italica achilleae | MFLUCC 14-0959 T | MG828903 | MG829013 | – | MG829204 | – | MG829122 | – | [114] |
| Jeremyomyces labinae | CBS 144617 T | MK442589 | – | – | MK442695 | – | – | – | [96] |
| Juncaceicola italica | MFLUCC 13-0750 | KX500110 | KX500107 | – | MG520897 | – | KX500108 | – | [109] |
| Juncaceicola luzulae | MFLUCC 13-0780 | KX449529 | KX449530 | – | MG520898 | – | KX449531 | – | [125] |
| Kwanghwana miscanthi | FU31017 | MK503817 | MK503823 | – | MT009126 | – | MK503829 | – | [126] |
| Leptosphaeria doliolum | CBS 505.75 T | JF740205 | GU301827 | – | GU349069 | – | GU296159 | – | [127,128] |
| Leptospora galii | KUMCC 15-0521 T | KX599547 | KX599548 | – | MG520899 | – | KX599549 | – | [109] |
| Leptospora rubella | CPC 11006 | DQ195780 | DQ195792 | – | – | – | DQ195803 | – | [129] |
| Leptospora thailandica | MFLUCC 16-0385 T | KX655559 | KX655549 | – | KX655564 | – | KX655554 | – | [130] |
| Loratospora luzulae | MFLUCC 14-0826 T | KT328497 | KT328495 | – | – | – | KT328496 | – | [121] |
| Mauginiella scaettae | CBS 239.58 | MH857770 | MH869303 | – | – | – | – | – | [76] |
| Melnikia anthoxanthii | MFLUCC 14-1010 | – | KU848204 | – | – | – | KU848205 | – | [131] |
| Murichromolaenicola chiangraiensis | MFLUCC 17-1488 T | MN994582 | MN994559 | – | MN998163 | – | MN994605 | – | [74] |
| Muriphaeosphaeria galatellae | MFLUCC 15-0769 | – | KT438330 | – | – | – | KT438332 | – | [132] |
| Muriphaeosphaeria galatellae | MFLUCC 14-0614 T | KT438333 | KT438329 | – | MG520900 | – | KT438331 | – | [132] |
| Neoophiobolus chromolaenae | MFLUCC 17-1467 T | MN994583 | MN994562 | – | MN998164 | – | MN994606 | – | [74] |
| Neosetophoma garethjonesii | MFLUCC 14-0528 | – | – | – | KY514402 | – | KY501126 | – | [133] |
| Neosetophoma rosigena | MFLUCC 17-0768 T | MG828928 | MG829037 | – | – | – | MG829143 | – | [114] |
| Neosphaerellopsis thailandica | CPC 21659 T | KP170652 | KP170721 | – | KP170678 | – | – | – | [118] |
| Neostagonospora arrhenather | MFLUCC 15-0464 | KX926417 | KX910091 | – | MG520901 | – | KX950402 | – | [134] |
| Neostagonospora caricis | CBS 135092 T | KF251163 | KF251667 | – | – | – | – | – | [76] |
| Neostagonospora phragmitis | MFLUCC 16-0493 | KX926416 | KX910090 | – | MG520902 | – | KX950401 | – | [134] |
| Neostagonosporella sichuanensis | MFLUCC 18-1223 | MH394690 | MH394687 | – | MK313854 | – | MK296469 | – | [58] |
| Neostagonosporella sichuanensis | MFLUCC 18-1228 T | MH368073 | MH368079 | – | MK313851 | – | MH368088 | – | [58] |
| Neosulcatispora strelitziae | CPC 25657 | KX228253 | KX228305 | – | – | – | – | – | [112] |
| Nodulosphaeria guttulatum | MFLUCC 15-0069 | – | – | – | KY514394 | – | KY501115 | – | [133] |
| Nodulosphaeria multiseptata | MFLUCC 15-0078 | KY496748 | KY496728 | – | – | – | – | – | [133] |
| Nodulosphaeria scabiosae | MFLUCC 14-1111 T | KU708850 | KU708846 | – | KU708854 | – | KU708842 | – | [135] |
| Ophiobolopsis italica | MFLUCC 17-1791 T | MG520939 | MG520959 | – | MG520903 | – | MG520977 | – | [109] |
| Ophiobolus artemisiae | MFLUCC 14-1156 T | KT315508 | KT315509 | – | MG520905 | – | MG520979 | – | [109] |
| Ophiobolus disseminans | MFLUCC 17-1787 | MG520941 | MG520961 | – | MG520906 | – | MG520980 | – | [109] |
| Ophiobolus ponticus | MFLUCC 17-2273 | MG520943 | MG520963 | – | MG520908 | – | MG520982 | – | [109] |
| Ophiosimulans tanaceti | MFLUCC 14-0525 | KU738890 | KU738891 | – | MG520910 | – | KU738892 | – | [109] |
| Ophiosphaerella herpotricha | KY423 | KP690989 | – | – | KP691011 | – | – | – | [136] |
| Ophiosphaerella korrae | ATCC 56289 | KC848509 | – | – | KC848515 | – | – | – | [136] |
| Ophiosphaerella narmari | ATCC 64688 | KC848510 | – | – | KC848516 | – | – | – | [136] |
| Paraleptosphaeria dryadis | CBS 643.86 | JF740213 | GU301828 | – | GU349009 | – | KC584632 | – | [127,128] |
| Paraleptospora chromolaenae | MFLUCC 17-1481 T | MN994587 | MN994563 | – | MN998167 | – | MN994609 | – | [74] |
| Paralloneottiosporina sichuanensis | SICAUCC 22-0074 T | ON226746 | ON227102 | – | ON244423 | – | ON227129 | – | This study |
| Paralloneottiosporina sichuanensis | SICAUCC 22-0075 | ON226747 | ON227103 | – | ON244424 | – | ON227130 | – | This study |
| Paraloratospora camporesii | MFLU 18-0915 T | MN756639 | MN756637 | – | – | – | MN756635 | – | [113] |
| Paraophiobolus arundinis | MFLUCC 17-1789 T | MG520945 | MG520965 | – | MG520912 | – | MG520984 | – | [109] |
| Paraophiobolus plantaginis | MFLUCC 17-0245 T | KY797641 | KY815010 | – | – | – | KY815012 | – | [109] |
| Paraphoma chrysanthemicola | CBS 522.66 | KF251166 | KF251670 | – | KF253124 | – | – | – | [116] |
| Paraphoma radicina | CBS 111.79 | KF251172 | KF251676 | – | KF253130 | – | – | – | [116] |
| Parastagonospora italica | MFLUCC 13-0377 T | KU058714 | KU058724 | – | MG520915 | – | MG520985 | – | [109,137] |
| Parastagonospora minima | MFLUCC 13-0376 | KU058713 | KU058723 | – | MG520916 | – | MG520986 | – | [109,137] |
| Parastagonosporella fallopiae | CCTU 1151.1 | MH460544 | MH460546 | – | MH460550 | – | – | – | [138] |
| Parastagonosporella fallopiae | CBS 135981 T | MH460543 | MH460545 | – | MH460549 | – | – | – | [138] |
| Phaeopoacea festucae | MFLUCC 17-0056 | KY824766 | KY824767 | – | – | – | KY824769 | – | [134] |
| Phaeoseptoriella zeae | CBS 144614 T | MK442611 | MK442547 | – | MK442702 | – | – | – | [96] |
| Phaeosphaeria chiangraina | MFLUCC 13-0231 T | KM434270 | KM434280 | – | KM434298 | – | KM434289 | – | [57] |
| Phaeosphaeria oryzae | CBS 110110 T | KF251186 | KF251689 | – | – | – | GQ387530 | – | [139] |
| Phaeosphaeria pleurospora | CBS 460.84 | AF439498 | – | – | – | – | – | – | [140] |
| Phaeosphaeriopsis glaucopunctata | MFLUCC 13-0265 | KJ522473 | KJ522477 | – | MG520918 | – | KJ522481 | – | [109,141] |
| Phaeosphaeriopsis triseptata | MFLUCC 13-0271 | KJ522475 | KJ522479 | – | MG520919 | – | KJ522484 | – | [109,141] |
| Phaeosphaeriopsis yuccae | MFLUCC 16-0558 | KY554482 | KY554481 | – | MG520920 | – | KY554480 | – | [109] |
| Piniphoma wesendahlina | CBS 145032 T | MK442615 | MK442551 | – | MK442706 | – | – | – | [96] |
| Poaceicola arundinis | MFLUCC 15-0702 T | KU058716 | KU058726 | – | MG520921 | – | MG520988 | – | [109] |
| Poaceicola italica | MFLUCC 13-0267 | KX926421 | KX910094 | – | MG520924 | – | KX950409 | – | [109,134] |
| Populocrescentia ammophilae | MFLUCC 17-0665 T | MG828949 | MG829059 | – | MG829231 | – | MG829164 | – | [114] |
| Populocrescentia forlicesenensis | MFLUCC 14-0651 T | KT306948 | KT306952 | – | MG520925 | – | KT306955 | – | [121] |
| Populocrescentia rosae | TASM 6125 T | – | MG829060 | – | MG829232 | – | MG829165 | – | [114] |
| Pseudoophiobolus mathieui | MFLUCC 17-1784 | MG520949 | MG520969 | – | MG520928 | – | MG520991 | – | [109] |
| Pseudoophiobolus rosae | MFLUCC 17-1786 T | MG520952 | MG520972 | – | MG520930 | – | MG520993 | – | [109] |
| Pseudoophiobolus urticicola | KUMCC 17-0168 T | MG520955 | MG520975 | – | MG520933 | – | MG520996 | – | [109] |
| Pseudoophiosphaerella huishuiensis | HS-13 | MK522509 | MK522499 | – | MK523389 | – | MK522505 | – | [124] |
| Pseudophaeosphaeria rubi | MFLUCC 14-0259 T | KX765298 | KX765299 | – | MG520934 | – | KX765300 | – | [130] |
| Sclerostagonospora ericae | CPC 25927 T | KX228268 | KX228319 | – | KX228375 | – | – | – | [112] |
| Scolicosporium minkeviciusii | MFLUCC 12-0089 | – | KF366382 | – | – | – | KF366383 | – | [142] |
| Septoriella phragmitis | CPC 24118 T | KR873251 | KR873279 | – | – | – | – | – | [143] |
| Setomelanomma holmii | CBS 110217 | KT389542 | GU301871 | – | GU349028 | – | GU296196 | – | [127,144] |
| Setophoma terrestris | CBS 335.29 | KF251246 | KF251749 | – | KF253196 | – | – | – | [116] |
| Stagonospora neglecta | CBS 343.86 | AJ496630 | – | – | – | – | – | – | [145] |
| Sulcispora supratumida | MFLUCC 14-0995 T | KP271443 | KP271444 | – | MH665366 | – | KP271445 | – | [146] |
| Tintelnotia destructans | CBS 127737 T | KY090652 | KY090664 | – | – | – | KY090698 | – | [147] |
| Tintelnotia opuntiae | CBS 376.91 T | KY090651 | GU238123 | – | – | – | GU238226 | – | [147,148] |
| Vagicola arundinis | MFLUCC 15-0027 T | KY706139 | KY706129 | – | MG520936 | – | KY706134 | – | [109] |
| Vittaliana mangrovei | NFCCI 4251 T | MG767311 | MG767312 | – | MG767314 | – | MG767313 | – | [149] |
| Vrystaatia aloeicola | CBS 135107 | KF251278 | KF251781 | – | – | – | – | – | [116] |
| Wingfieldomyces cyperi | CBS 141450 T | KX228286 | KX228337 | – | MK540163 | – | – | – | [150] |
| Wojnowicia italica | MFLUCC 13-0447 T | KX342923 | KX430001 | – | KX430003 | – | KX430002 | – | [130] |
| Wojnowicia rosicola | MFLUCC 15-0128 T | MG828979 | MG829091 | – | – | – | MG829191 | – | [114] |
| Wojnowiciella eucalypti | CBS 139904 T | KR476741 | KR476774 | – | – | – | – | – | [76] |
| Xenophoma puncteliae | CBS 128022 | – | JQ238619 | – | KP170686 | – | – | – | [118,151] |
| Xenoseptoria neosaccardoi | CBS 120.43 | KF251280 | KF251783 | – | KF253227 | – | – | – | [116] |
| Xenoseptoria neosaccardoi | CBS 128665 | KF251281 | KF251784 | – | KF253228 | – | – | – | [116] |
| Yunnanensis phragmitis | MFLUCC 17-1361 T | MF684869 | MF684865 | – | – | – | MF684864 | – | [152] |
| Yunnanensis phragmitis | MFLUCC 17-0315 | MF684862 | MF684863 | – | MF683624 | – | MF684867 | – | [152] |
| Biatriospora marina | CY 1228 | – | GQ925848 | – | GU479848 | – | GQ925835 | GU479823 | [153] |
| Biatriospora peruviensis | CCF 4485 | – | LN626683 | – | LN626671 | – | LN626677 | LN626665 | [154] |
| Neooccultibambusa chiangraiensis | MFLUCC 12-0559 T | – | KU764699 | – | – | – | KU712458 | – | [155] |
| Neoroussoella bambusae | MFLUCC 11-0124 | – | KJ474839 | – | KJ474848 | – | – | KJ474856 | [156] |
| Occultibambusa aquatica | MFLUCC 11-0006 | – | KX698110 | – | – | – | KX698112 | – | [130] |
| Occultibambusa bambusae | MFLUCC 11-0394 | – | KU863113 | – | KU940194 | – | KU872117 | KU940171 | [16] |
| Occultibambusa bambusae | MFLUCC 13-0855 T | – | KU863112 | – | KU940193 | – | KU872116 | KU940170 | [16] |
| Occultibambusa chiangraiensis | MFLUCC 16-0380 T | – | KX655546 | – | – | – | KX655551 | KX655566 | [130] |
| Occultibambusa fusispora | MFLUCC 11-0127 T | – | KU863114 | – | KU940195 | – | – | KU940172 | [16] |
| Occultibambusa jonesii | GZCC 16-0117 T | – | KY628322 | – | KY814756 | – | KY628324 | KY814758 | [157] |
| Occultibambusa kunmingensis | HKAS 102151 T | – | MN913733 | – | MT954407 | – | MT864342 | MT878453 | [61] |
| Occultibambusa maolanensis | GZCC 16-0116 | – | KY628323 | – | KY814757 | – | KY628325 | KY814759 | [157] |
| Occultibambusa pustula | MFLUCC 11-0502 | – | KU863115 | – | – | – | KU872118 | – | [16] |
| Paradictyoarthrinium diffractum | MFLUCC 13-0466 | – | KP744498 | – | – | – | KP753960 | KX437764 | [105,158] |
| Paradictyoarthrinium tectonicola | MFLUCC 13-0465 T | – | KP744500 | – | – | – | KP753961 | KX437763 | [105,158] |
| Roussoella hysterioides | HH 26988 | – | AB524622 | – | AB539115 | – | AB524481 | AB539102 | [127] |
| Roussoella nitidula | MFLUCC 11-0182 | – | KJ474843 | – | KJ474852 | – | – | KJ474859 | [156] |
| Roussoella nitidula | MFLUCC 11-0634 | – | KJ474842 | – | KJ474851 | – | – | KJ474858 | [156] |
| Roussoella pustulans | KT 1709 | – | AB524623 | – | AB539116 | – | AB524482 | AB539103 | [1,127] |
| Seriascoma bambusae | KUMCC 21-0021 | – | MZ329035 | – | MZ325468 | – | MZ329031 | MZ325470 | [159] |
| Seriascoma didymospora | MFLUCC 11-0179 T | – | KU863116 | – | KU940196 | – | – | KU940173 | [16] |
| Seriascoma didymospora | MFLUCC 11-0194 | – | KU863117 | – | KU940197 | – | – | KU940174 | [16] |
| Seriascoma yunnanense | MFLU 19-0690 T | – | NG_068303 | – | MN381858 | – | MN174694 | MN210324 | [44] |
| Seriascoma yunnanense | SICAUCC 22-0059 | – | ON226771 | – | ON567182 | – | ON227356 | ON567183 | This study |
| Torula herbarum | CBS 111855 | – | KF443386 | – | KF443403 | – | KF443391 | KF443396 | [160] |
| Westerdykella ornata | CBS 379.55 | – | GU301880 | – | GU349021 | – | GU296208 | GU371803 | [127] |
References
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Hatakeyama, S.; Harada, Y.; Sano, T.; Shirouzu, T.; Hosoya, T. Molecular taxonomy of bambusicolous fungi: Tetraplosphaeriaceae, a new pleosporalean family with Tetraploa-like anamorphs. Stud. Mycol. 2009, 64, 175–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.Y.; Zhou, D.Q.; Ma, L.S.; Yao, J.; Zhang, D.M. Diversity of bamboo species in China. World Bamboo Ratt. 2020, 18, 55–65. [Google Scholar] [CrossRef]
- Du, S.S. Classification and arrangement of Phyllostachys (Poaceae: Bambusoideae), China. J. Fujian Forestry Sci. Technol. 2020, 47, 120–123. [Google Scholar] [CrossRef]
- Li, Y.G.; Xue, L.; Fan, L.L.; Ye, L.T.; Cheng, L.Y.; He, T.Y.; Zheng, Y.S. Research progress in germplasm resources and applications of Phyllostachy. J. Sic. For. Sci. Technol. 2019, 40, 117–122. [Google Scholar] [CrossRef]
- Bystriakova, N.; Kapos, V.; Lysenko, I.; Stapleton, C.M.A. Distribution and conservation status of forest bamboo biodiversity in the Asia-Pacific Region. Biodivers. Conserv. 2003, 12, 1833–1841. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Dayton, D.C.; Hames, B. Bamboo: An overlooked biomass resource? Biomass. Bioener. 2000, 19, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Idris, M.A.; Mohamad, A. Bamboo shoot utilization in peninsular Malaysia: A case study in Pahang. J. Bamboo Rattan. 2002, 1, 141–155. [Google Scholar] [CrossRef]
- Shi, J.Y.; Chen, Q.B.; Huang, J.Y.; Zhou, D.Q.; Ma, L.S.; Yao, J. Biodiversity of the staple food bamboos of giant panda and its important value. World Bamboo Rattan. 2020, 18, 10–19. [Google Scholar] [CrossRef]
- Wang, X.J.; Wang, T.; Chi, M.; Li, L.B. Research progress of ornamental bamboos in China. J. Bamboo Res. 2019, 38, 3–9. [Google Scholar] [CrossRef]
- Hyde, K.D.; Zhou, D.Q.; Mckenzie, E.H.C.; Ho, W.H.; Dalisay, T. Vertical distribution of saprobic fungi on bamboo culms. Fungal Divers. 2002, 11, 109–118. [Google Scholar]
- Tanaka, E.; Shimizu, K.; Imanishi, Y.; Yasuda, F.; Tanaka, C. Isolation of basidiomycetous anamorphic yeast-like fungus Meira argovae found on Japanese bamboo. Mycoscience 2008, 49, 329–333. [Google Scholar] [CrossRef]
- Dai, D.Q.; Bhat, D.J.; Liu, J.K.; Chukeatirote, E.; Zhao, R.L.; Hyde, K.D. Bambusicola, a new genus from bamboo with asexual and sexual morphs. Cryptogamie Mycol. 2012, 33, 363–379. [Google Scholar] [CrossRef]
- Doungporn, M.; Hiroko, K.; Tatsuji, S. Molecular diversity of bamboo–associated fungi isolated from Japan. FEMS Microbiol. Lett. 2007, 266, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Zhou, D.; Dalisay, T. Bambusicolous fungi: A review. Fungal Divers. 2002, 9, 1–14. [Google Scholar]
- Tanaka, K.; Harada, Y. Bambusicolous fungi in Japan (1): Four Phaeosphaeria species. Mycoscience 2004, 45, 377–382. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Tanaka, K.; Harada, Y. Bambusicolous fungi in Japan (7): A new coelomycetous genus, Versicolorisporium. Mycoscience 2008, 49, 211–214. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Tanaka, K.; Harada, Y. Bambusicolous fungi in Japan (5): Three species of Tetraploa. Mycoscience 2005, 46, 196–200. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Zhang, X. Potentials of bamboo in traditioanl Chinese medicine and development of heath products. World Sci. Technol. 2000, 3, 54–56. [Google Scholar] [CrossRef]
- Zhou, B.Z.; Fu, M.Y.; Xie, J.Z.; Yang, X.S.; Li, Z.C. Ecological functions of bamboo forest: Research and application. J. Forestry Res. 2005, 16, 143–147. [Google Scholar] [CrossRef]
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S.N. Bamboo shoots: A novel source of nutrition and medicine. Crit. Rev. Food Sci. Nutr. 2013, 53, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Teng, S.C. Fungi of China; Mycotaxon, Ltd.: New York, NY, USA, 1996; pp. 1–728. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press, Academica Sinica: Beijing, China, 1979; pp. 1–1527. [Google Scholar]
- Chen, M.M. Forest Fungi Phytogeography: Forest Fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research and Education Center: Sacramento, CA, USA, 2002; pp. 1–469. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Hudson, B.A.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplifcation and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfaud, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identifification and mapping of enzymatically amplifified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef]
- Castlebury, L.; Rossman, A.; Sung, G.; Hyten, A.; Spatafora, J. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Wang, M.; Tan, X.M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. MycoKeys 2018, 34, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Xu, X.L.; Yang, C.L.; Jeewon, R.; Wanasinghe, D.N.; Xiao, Q.G. Morpho-molecular diversity of Linocarpaceae (Chaetosphaeriales): Claviformispora gen. nov. from decaying branches of Phyllostachys heteroclada. MycoKeys 2020, 69, 113–129. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of Arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, D.Q.; Jiang, H.B.; Tang, L.Z.; Bhat, D.J. Two new species of Arthrinium (Apiosporaceae, Xylariales) associated with bamboo from Yunnan, China. Mycosphere 2016, 7, 1332–1345. [Google Scholar] [CrossRef]
- Silva, R.M.; Oliveira, R.J.; Bezerra, J.D.; Bezerra, J.L.; Souza–Motta, C.M.; Silva, G.A. Bifusisporella sorghi gen. et sp. nov. (Magnaporthaceae) to accommodate an endophytic fungus from Brazil. Mycol. Prog. 2019, 18, 847–854. [Google Scholar] [CrossRef]
- Li, W.J.; McKenzie, E.H.C.; Liu, J.K.; Bhat, D.J.; Dai, D.Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.L.; Shang, Q.J.; et al. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Rathnayaka, A.R.; Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Liu, J.K.; Tennakoon, D.S.; Hyde, K.D. Introducing Seriascoma yunnanense sp. nov. (Occultibambusaceae, Pleosporales) based on evidence from morphology and phylogeny. Asian J. Mycol. 2019, 2, 245–253. [Google Scholar] [CrossRef]
- Dai, D.Q.; Tang, L.Z.; Wang, H.B. A Review of Bambusicolous Ascomycetes. Bamboo Curr. Future Prospect. 2018, 165–183. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P. Conspectus generum pyrenomycetum italicorum additis speciebus fungorum Venetorum novis vel criticis, systemate carpologico dispositorum. Atti Soc. Veneziana-Trent. Istriana Sci. Nat. 1875, 4, 77–100. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes VI. Commonw. Mycol. Inst. 1965, 103, 1–46. [Google Scholar]
- Pintos, Á.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2021, 7, 197–221. [Google Scholar] [CrossRef]
- Tian, X.G.; Karunarathna, S.C.; Mapook, A.; Promputtha, I.; Xu, J.C.; Bao, D.F.; Tibpromma, S. One new species and two new host records of Apiospora from bamboo and maize in Northern Thailand with thirteen new combinations. Life 2021, 11, 1071. [Google Scholar] [CrossRef]
- Chen, K.; Wu, X.Q.; Huang, M.X.; Han, Y.Y. First report of brown culm ctreak of Phyllostachys praecox caused by Arthrinium arundinis in Nanjing, China. Plant Dis. 2014, 98, 1274. [Google Scholar] [CrossRef]
- Yin, C.W.; Luo, F.Y.; Zhang, H.; Fang, X.M.; Zhu, T.H.; Li, S.J. First report of Arthrinium kogelbergense causing blight disease of Bambusa intermedia in Sichuan Province, China. Plant Dis. 2021, 105, 214. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, J.K.; Lin, C.G.; Chen, Y.Y.; Xiang, M.M.; Liu, Z.Y. Additions to the genus Arthrinium (Apiosporaceae) from bamboos in China. Front. Microbiol. 2021, 12, 661281. [Google Scholar] [CrossRef]
- Jiang, N.; Li, J.; Tian, C.M. Arthrinium species associated with bamboo and reed plants in China. Fungal Syst. Evol. 2018, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.L.; Xu, X.L.; Liu, Y.G.; Xu, X.L. First report of bamboo blight disease caused by Arthrinium yunnanum on Phyllostachys heteroclada in Sichuan, China. Plant Dis. 2018, 103, 1026. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Tennakoon, D.S.; Jeewon, R.; Gentekaki, E.; Kuo, C.H.; Hyde, K.D. Multi-gene phylogeny and morphotaxonomy of Phaeosphaeria ampeli sp. nov. from Ficus ampelas and a new record of P. musae from Roystonea regia. Phytotaxa 2019, 406, 111–128. [Google Scholar] [CrossRef]
- Phookamsak, R.; Liu, J.K.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.; Thambugala, K.M.; Dai, D.Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Wanasinghe, D.N.; Jeewon, R.; Phookamsak, R.; Liu, Y.G.; Liu, L.J.; Hyde, K.D. Neostagonosporellasichuanensis gen. et sp. nov. (Phaeosphaeriaceae, Pleosporales) on Phyllostachys heteroclada (Poaceae) from Sichuan Province, China. MycoKeys 2019, 46, 119–150. [Google Scholar] [CrossRef] [PubMed]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.J.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungal. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–135. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Liu, Y.G. Two new species of Bambusicola (Bambusicolaceae, Pleosporales) on Phyllostachys heteroclada from Sichuan, China. Nova Hedwigia. 2019, 108, 527–545. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Liu, Y.G.; Hyde, K.D.; Mckenzie, E.H.C. A new species of Phyllachora (Phyllachoraceae, Phyllachorales) on Phyllostachys heteroclada from Sichuan, China. Phytotaxa 2019, 392, 186–196. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Dong, W.; Wanasinghe, D.N.; Liu, Y.G.; Hyde, K.D. Introducing Arthrinium phyllostachium sp. nov. (Apiosporaceae, Xylariales) on Phyllostachys heteroclada from Sichuan Province, China. Phytotaxa 2019, 406, 91–110. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Liu, Y.G. Podonectria sichuanensis, a potentially mycopathogenic fungus from Sichuan Province in China. Phytotaxa 2019, 402, 219–231. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Jeewon, R.; Boonmee, S.; Liu, Y.G.; Hyde, K.D. Acremonium arthrinii sp. Nov., a mycopathogenic fungus on Arthrinium yunnanum. Phytotaxa 2019, 420, 283–299. [Google Scholar] [CrossRef]
- Yang, C.L.; Baral, H.O.; Xu, X.L.; Liu, Y.G. Parakarstenia phyllostachydis, a new genus and species of non-lichenized Odontotremataceae (Ostropales, Ascomycota). Mycol. Prog. 2019, 18, 833–845. [Google Scholar] [CrossRef]
- Yan, H.; Jiang, N.; Liang, L.Y.; Yang, Q.; Tian, C.M. Arthriniumtrachycarpum sp. nov. from Trachycarpus fortunei in China. Phytotaxa 2019, 400, 203–210. [Google Scholar] [CrossRef]
- Alves-Silva, G.; Drechsler-Santos, E.R.; da Silveira, R.M.B. Bambusicolous Fomitiporia revisited: Multilocus phylogeny reveals a clade of host-exclusive species. Mycologia 2020, 112, 633–648. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Bhat, J.D.; Cheewangkoon, R.; Xie, N. Bambusicolous Arthrinium species in Guangdong Province, China. Front. Microbiol. 2020, 11, 2981. [Google Scholar] [CrossRef]
- Pintos, A.; Alvarado, P.; Planas, J.; Jarling, R. Six new species of Arthrinium from Europe and notes about A.caricicola and other species found in Carex spp. hosts. MycoKeys 2019, 49, 15–48. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Goonasekara, I.D.; Jayawardena, R.S.; Jiang, H.B.; Li, J.F.; Hyde, K.D.; Kang, J.C. Arthrinium bambusicola (Fungi, Sordariomycetes), a new species from Schizostachyum brachycladum in northern Thailand. Biodivers Data J. 2020, 8, e58755. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde., K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2018, 91, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Kulkarni, G.; Sonawane, M.S.; Shouche, Y.S. A new endophytic species of Arthrinium (Apiosporaceae) from Jatropha podagrica. Mycoscience 2014, 55, 118–123. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Zhang, Z.F.; Cai, L.; Peng, W.J.; Liu, F. Four new filamentous fungal species from newly-collected and hive-stored bee pollen. Mycosphere 2018, 9, 1089–1116. [Google Scholar] [CrossRef]
- Das, K.; Lee, S.Y.; Choi, H.W.; Eom, A.H.; Choe, Y.J.; Jung, H.Y. Taxonomy of Arthrinium minutisporum sp. nov., Pezicula neosporulosa, and Acrocalymma pterocarpi: New records from soil in Korea. Mycobiology 2020, 48, 450–463. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.J.; de Vries, A.E.; Bologna, N.J.; Bickford, W.A.; Kowalski, K.P.; Meyerson, L.A.; Cronin, J.T. Intraspecific and biogeographical variation in foliar fungal communities and pathogen damage of native and invasive Phragmites australis. Glob. Ecol. Biogeogr. 2020, 29, 1199–1211. [Google Scholar] [CrossRef]
- Jiang, N.; Liang, Y.M.; Tian, C.M. A novel bambusicolous fungus from China, Arthrinium chinense (Xylariales). Sydowia 2020, 72, 77–83. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for fungi. Database 2014, 336–341. [Google Scholar] [CrossRef]
- Kwon, S.L.; Park, M.S.; Jang, S.; Lee, Y.M.; Heo, Y.M.; Hong, J.H.; Lee, H.; Jang, Y.; Park, J.H.; Kim, C.; et al. The genus Arthrinium (Ascomycota, Sordariomycetes, Apiosporaceae) from marine habitats from Korea, with eight new species. IMA Fungus 2021, 12, 13. [Google Scholar] [CrossRef]
- Crous, P.; Hernandez-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Koelling, G.; Marais, E.; Wingfield, M.J.; Yilmaz, N.; Adan, O.C.G.; Akulov, A.; et al. New and interesting fungi. 4. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Jiang, N.; Tian, C.M. The holomorph of Arthrinium setariae sp. nov. (Apiosporaceae, Xylariales) from China. Phytotaxa 2021, 483, 149–159. [Google Scholar] [CrossRef]
- Jiang, H.B.; Hyde, K.D.; Doilom, M.; Karunarathna, S.C.; Xu, J.C.; Phookamsak, R. Arthrinium setostromum (Apiosporaceae, Xylariales), a novel species associated with dead bamboo from Yunnan, China. Asian J. Mycol. 2019, 2, 254–268. [Google Scholar] [CrossRef]
- Wang, M.; Liu, F.; Crous, P.W.; Cai, L. Phylogenetic reassessment of Nigrospora: Ubiquitous endophytes, plant and human pathogens. Persoonia 2017, 39, 118–142. [Google Scholar] [CrossRef] [PubMed]
- Tennakoon, D.S.; Kuo, C.H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef]
- Hirata, K.; Kusaba, M.; Chuma, I.; Osue, J.; Nakayashiki, H.; Mayama, S.; Tosa, Y. Speciation in Pyricularia inferred from multilocus phylogenetic analysis. Mycol. Res. 2007, 111, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Zhang, N. Magnaporthiopsis, a new genus in Magnaporthaceae (Ascomycota). Mycologia 2013, 105, 1019–1029. [Google Scholar] [CrossRef]
- Luo, J.; Walsh, E.; Zhang, N. Toward monophyletic generic concepts in Magnaporthales: Species with Harpophora asexual states. Mycologia 2015, 107, 641–646. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Lin, F.C.; Zhang, C.L.; Kubicek, C.P. A new species of Harpophora (Magnaporthaceae) recovered from healthy wild rice (Oryza granulata) roots, representing a novel member of a beneficial dark septate endophyte. FEMS Microbiol. Lett. 2010, 307, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernandez-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Restrepo, M.; Groenewald, J.Z.; Elliott, M.L.; Canning, G.; Mcmillan, V.E.; Crous, P.W. Take-all or nothing. Stud. Mycol. 2016, 83, 19–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Zhao, S.; Shen, Q. A six-gene phylogeny reveals the evolution of mode of infection in the rice blast fungus and allied species. Mycologia 2011, 103, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.T.W. Gaeumannomyces wongoonoo sp. nov., the cause of a patch disease of buffalo grass (St Augustine grass). Mycol. Res. 2002, 106, 857–862. [Google Scholar] [CrossRef]
- Khemmuk, W.; Geering, A.D.; Shivas, R.G. Wongia gen. nov. (Papulosaceae, Sordariomycetes), a new generic name for two root-infecting fungi from Australia. IMA Fungus 2016, 7, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Vines, P.L.; Hoffmann, F.G.; Meyer, F.; Allen, T.W.; Luo, J.; Zhang, N.; Tomaso-Peterson, M. Magnaporthiopsis cynodontis, a novel turfgrass pathogen with widespread distribution in the United States. Mycologia 2020, 112, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Vines, P.L.; Grimshaw, A.; Hoffman, L.; Walsh, E.; Bonos, S.A.; Clarke, B.B.; Murphy, J.A.; Meyer, W.A.; Zhang, N. Magnaporthiopsis meyeri-festucae, sp. nov., associated with a summer patch-like disease of fine fescue turfgrasses. Mycologia 2017, 109, 780–789. [Google Scholar] [CrossRef]
- Luo, J.; Walsh, E.; Zhang, N. Four new species in Magnaporthaceae from grass roots in New Jersey Pine Barrens. Mycologia 2014, 106, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Vines, P.L. Evaluation of Ultradwarf Bermudagrass Cultural Management Practices and Identification, characterization, and Pathogenicity of Ectotrophic Rootinfecting Fungi Associated with Summer Decline of Ultradwarf Bermudagrass Putting Greens. Master Thesis, Mississippi State University, Starkville, MI, USA, 2015. [Google Scholar]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Park, M.J.; Shin, H.D. A new species of Pyricularia on Commelina communis. Mycotaxon 2009, 108, 449–456. [Google Scholar] [CrossRef]
- Ban, Y.; Tang, M.; Chen, H.; Xu, Z.; Zhang, H.; Yang, Y. The response of dark septate endophytes (DSE) to heavy metals in pure culture. PLoS ONE 2018. 7, e47968. [CrossRef]
- Saleh, A.A.; Leslie, J.F. Cephalosporium maydis is a distinct species in the Gaeumannomyces-Harpophora species complex. Mycologia 2004, 96, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Phookamsak, R.; Wanasinghe, D.N.; Hongsanan, S.; Phukhamsakda, C.; Huang, S.K.; Tennakoon, D.S.; Norphanphoun, C.; Camporesi, E.; Bulgakov, T.S.; Promputtha, I.; et al. Towards a natural classification of Ophiobolus and ophiobolus-like taxa; introducing three novel genera Ophiobolopsis, Paraophiobolus and Pseudoophiobolus in Phaeosphaeriaceae (Pleosporales). Fungal Divers. 2017, 87, 299–339. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- De Gruyter, J.; Aveskamp, M.M.; Woudenberg, J.H.; Verkley, G.J.; Groenewald, J.Z.; Crous, P.W. Molecular phylogeny of Phoma and allied anamorph genera: Towards a reclassification of the Phoma complex. Mycol. Res. 2009, 113, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal planet description sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Pem, D.; Bundhun, D.; Karunarathna, A.; Ekanayaka, A.H.; Bao, D.F.; Li, J.F.; Samarakoon, M.C.; Chaiwan, N.; Lin, C.G.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, G.E.B.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Thambugala, K.M.; Wanasinghe, D.N.; Gentekaki, E.; Promputtha, I.; Kuo, C.H.; Hyde, K.D. Additions to Phaeosphaeriaceae (Pleosporales): Elongaticollum gen. nov., Ophiosphaerella taiwanensis sp. nov., Phaeosphaeriopsis beaucarneae sp. nov. and a new host record of Neosetophoma poaceicola from Musaceae. MycoKeys 2020, 70, 59–88. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.J.; Shin, H.D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanasinghe, D.N.; Jones, E.B.G.; Camporesi, E.; Boonmee, S.; Karunarathna, S.C.; Thines, M.; Mortimer, P.E.; Xu, J.; Hyde, K.D. Dematiopleospora mariae gen. sp. nov., from Ononis Spinosa in Italy. Cryptogamie. Mycol. 2014, 35, 105–117. [Google Scholar] [CrossRef]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Alfenas, A.C.; Crous, P.W. Mycoparasitic species of Sphaerellopsis, and allied lichenicolous and other genera. IMA Fungus 2014, 5, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Cui, Y.; Jia, H.; He, D.; Yu, H.; Gao, S.; Yokoyama, K.; Li, J.; Wang, L. Characterization of Edenia gomezpompae isolated from a patient with keratitis. Mycopathologia 2013, 176, 75–81. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Tibpromma, S.; Wanasinghe, D.N.; Camporesi, E.; Hyde, K.D. Equiseticola gen. nov. (Phaeosphaeriaceae), from Equisetum sp. in Italy. Phytotaxa 2016, 284, 169–180. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Wanasinghe, D.N.; Dayarathne, M.C.; Al-Saady, N.A.; Al-Sadi, A.M. Phylogenetic classification and generic delineation of Hydeomyces desertipleosporoides gen. et sp. nov., (Phaeosphaeriaceae) from Jebel Akhdar Mountain in Oman. Phytotaxa 2019, 391, 28–38. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, J.K.; Jeewon, R.; Wanasinghe, D.N.; Liu, Z.Y. Fungi from Asian Karst formations III. Molecular and morphological characterization reveal new taxa in Phaeosphaeriaceae. Mycosphere 2019, 10, 202–220. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; Camporesi, E.; Promputtha, I. Taxonomy and phylogeny of Juncaceicola gen. nov. (Phaeosphaeriaceae, Pleosporinae, Pleosporales). Cryptogamie Mycol. 2016, 37, 135–156. [Google Scholar] [CrossRef]
- Karunarathna, A.; Phookamsak, R.; Jayawardena, R.S.; Hyde, K.D.; Kuo, C.H. Kwanghwana miscanthi Karun., C.H. Kuo & K.D. Hyde, gen. et sp. nov. (Phaeosphaeriaceae, Pleosporales) on Miscanthus floridulus (Labill.) Warb. ex K. Schum. & Lauterb. (Poaceae). Cryptogamie. Mycol. 2020, 41, 119–132. [Google Scholar] [CrossRef]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef] [PubMed]
- De Gruyter, J.; Woudenberg, J.H.; Aveskamp, M.M.; Verkley, G.J.; Groenewald, J.Z.; Crous, P.W. Redisposition of phoma-like anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Verkley, G.J.; Groenewald, J.Z. Eucalyptus microfungi known from culture. 1. Cladoriella and Fulvoflamma genera nova, with notes on some other poorly known taxa. Stud. Mycol. 2006, 55, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Wanasinghe, D.N.; Papizadeh, M.; Goonasekara, I.D.; Camporesi, E.; Bhat, D.J.; McKenzie, E.H.C.; Phillips, A.J.L.; Diederich, P.; et al. Taxonomy and phylogeny of dematiaceous coelomycetes. Fungal Divers 2016, 77, 1–316. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Ariyawansa, H.A.; Phookamsak, R.; Chomnunti, P.; Bulgakov, T.S.; Yang, J.B.; Bhat, D.J.; Bahkali, A.H.; Hyde, K.D. Muriphaeosphaeria galatellae gen. et sp. nov. in Phaeosphaeriaceae (Pleosporales). Phytotaxa 2015, 227, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1–50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Mapook, A.; Boonmee, S.; Ariyawansa, H.A.; Tibpromma, S.; Campesori, E.; Jones, E.B.G.; Bahkali, A.H.; Hyde, K.D. Taxonomic and phylogenetic placement of Nodulosphaeria. Mycol. Progs. 2016, 15, 1–15. [Google Scholar] [CrossRef]
- Flores, F.J.; Marek, S.M.; Orquera, G.; Walker, N.R. Molecular identification and multilocus phylogeny of Ophiosphaerella species associated with spring dead spot of bermudagrass. Crop. Sci. 2017, 57, 249. [Google Scholar] [CrossRef]
- Li, W.J.; Bhat, D.J.; Camporesi, E.; Tian, Q.; Wijayawardene, N.N.; Dai, D.Q.; Phookamsak, R.; Chomnunti, P.; Bahkali, A.H.; Hyde, K.D. New asexual morph taxa in Phaeosphaeriaceae. Mycosphere 2015, 6, 681–708. [Google Scholar] [CrossRef]
- Bakhshi, M.; Arzanlou, M.; Groenewald, J.Z.; Quaedvlieg, W.; Crous, P.W. Parastagonosporella fallopiae gen. et sp. nov. (Phaeosphaeriaceae) on Fallopia convolvulus from Iran. Mycol. Prog. 2019, 18, 203–214. [Google Scholar] [CrossRef]
- De Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010, 102, 1066–1081. [Google Scholar] [CrossRef]
- Camara, M.P.S.; Palm, M.E.; van Berkum, P.; O’Neill, N.R. Molecular phylogeny of Leptosphaeria and Phaeosphaeria. Mycologia 2002, 94, 630–640. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Camporesi, E.; Ariyawansa, H.A.; Phookamsak, R.; Liu, Z.Y.; Hyde, K.D. Phylogeny and morphology of Phaeosphaeriopsis triseptata sp. nov., and Phaeosphaeriopsis glaucopunctata. Phytotaxa 2014, 176, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Camporesi, E.; Song, Y.; Dai, D.Q.; Hyde, K.D. Multi-gene analyses reveal taxonomic placement of Scolicosporium minkeviciusii in Phaeosphaeriaceae (Pleosporales). Cryptogamie Mycol. 2013, 34, 357–366. [Google Scholar] [CrossRef]
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hernandez-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.H.; Schumacher, R.K.; Stielow, J.B.; et al. The genera of fungi—Fixing the application of the type species of generic names—G 2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [Green Version]
- Ernst, M.; Mendgen, K.W.; Wirsel, S.G. Endophytic fungal mutualists: Seed-borne Stagonospora spp. enhance reed biomass production in axenic microcosms. Mol. Plant-Microb. MPMI 2003, 16, 580–587. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Jeewon, R.; Camporesi, E.; Hyde, K.D.; Zeng, Y.J.; Tian, S.L.; Xie, N. Sulcisporasupratumida sp. nov. (Phaeosphaeriaceae, Pleosporales) on Anthoxanthumodoratum from Italy. MycoKeys 2018, 38, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Hofmuller, W.; Seibold, M.; de Hoog, G.S.; Harak, H.; Tammer, I.; van Diepeningen, A.D.; Behrens-Baumann, W. Tintelnotia, a new genus in Phaeosphaeriaceae harbouring agents of cornea and nail infections in humans. Mycoses. 2016, 60, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.; Verkley, G.J.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef]
- Devadatha, B.; Mehta, N.; Wanasinghe, D.N.; Baghela, A.; Venkateswara, V.; Vittaliana, S. Vittaliana mangrovei Devadatha, Nikita, A. Baghela & V.V. Sarma, gen. nov, sp. nov. (Phaeosphaeriaceae), from mangroves near Pondicherry (India), based on morphology and multigene phylogeny. Cryptogamie Mycol. 2019, 40, 117–132. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef] [PubMed]
- Lawrey, J.D.; Diederich, P.; Nelsen, M.P.; Freebury, C.; Van den Broeck, D.; Sikaroodi, M.; Ertz, D. Phylogenetic placement of lichenicolous Phoma species in the Phaeosphaeriaceae (Pleosporales, Dothideomycetes). Fungal Divers. 2012, 55, 195–213. [Google Scholar] [CrossRef]
- Karunarathna, A.; Papizadeh, M.; Senanayake, I.C.; Jeewon, R.; Phookamsak, R.; Goonasekara, I.D.; Wanasinghe, D.N.; Wijayawardene, N.N.; Amoozegar, M.A.; Shahzadeh-Fazeli, S.A.; et al. Novel fungal species of Phaeosphaeriaceae with an asexual/sexual morph connection. Mycosphere 2017, 8, 1818–1834. [Google Scholar] [CrossRef]
- Suetrong, S.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; Sakayaroj, J.; Phongpaichit, S.; Tanaka, K.; Hirayama, K.; Jones, E.B.G. Molecular systematics of the marine Dothideomycetes. Stud. Mycol. 2009, 64, 155–173. [Google Scholar] [CrossRef]
- Kolarik, M.; Spakowicz, D.; Gazis, R.; Shaw, J.; Novakova, A.; Chudickova, M.; Forcina, G.C.; Kang, K.W.; Kelnarova, I.; Skaltsas, D.; et al. Biatriospora (Ascomycota: Pleosporales) is an ecologically diverse genus including facultative marine fungi and endophytes with biotechnological potential. Plant Syst. Evol. 2017, 303, 35–50. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Liu, J.K.; Phookamsak, R.; Dai, D.Q.; Tanaka, K.; Jones, E.B.G.; Xu, J.C.; Chukeatirote, E.; Hyde, K.D. Roussoellaceae, a new pleosporalean family to accommodate the genera Neoroussoella gen. nov., Roussoella and Roussoellopsis. Phytotaxa 2014, 181, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Yang, W.; Liu, Z.Y. Fungi from Asian Karst formations II. Two new species of Occultibambusa (Occultibambusaceae, Dothideomycetes) from karst landforms of China. Mycosphere 2017, 8, 550–559. [Google Scholar] [CrossRef]
- Li, J.; Bhat, D.J.; Phookamsak, R.; Mapook, A.; Lumyong, S.; Hyde, K.D. Sporidesmioides thailandica gen. et sp nov (Dothideomycetes) from northern Thailand. Mycol. Prog. 2016, 15, 1169–1178. [Google Scholar] [CrossRef]
- Jiang, H.B.; Phookamsak, R.; Hyde, K.D.; Mortimer, P.E.; Xu, J.C.; Kakumyan, P.; Karunarathna, S.C.; Kumla, J. A taxonomic appraisal of bambusicolous fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with new collections from Yunnan Province, China. Life 2021, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; van de Sande, W.W.; Stevens, D.A.; Fahal, A.; van Diepeningen, A.D.; Menken, S.B.; de Hoog, G.S. Revision of agents of black-grain eumycetoma in the order Pleosporales. Persoonia 2014, 33, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]














| Genera | Sequences Dataset |
|---|---|
| Apiospora | ITS, LSU, tub2, tef1-α |
| Bifusisporella | ITS, LSU, tef1-α, rpb1 |
| Paralloneottiosporina | ITS, LSU, SSU, tef1-α |
| Seriascom | ITS, LSU, SSU, tef1-α, rpb2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Q.; Lv, Y.-C.; Xu, X.-L.; Deng, Y.; Wang, F.-H.; Liu, S.-Y.; Liu, L.-J.; Yang, C.-L.; Liu, Y.-G. Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China. J. Fungi 2022, 8, 702. https://doi.org/10.3390/jof8070702
Zeng Q, Lv Y-C, Xu X-L, Deng Y, Wang F-H, Liu S-Y, Liu L-J, Yang C-L, Liu Y-G. Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China. Journal of Fungi. 2022; 8(7):702. https://doi.org/10.3390/jof8070702
Chicago/Turabian StyleZeng, Qian, Yi-Cong Lv, Xiu-Lan Xu, Yu Deng, Fei-Hu Wang, Si-Yi Liu, Li-Juan Liu, Chun-Lin Yang, and Ying-Gao Liu. 2022. "Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China" Journal of Fungi 8, no. 7: 702. https://doi.org/10.3390/jof8070702