| Abortiporus biennis (Bull.) Singer | √ | | | | | | SS | Medicinal | PHT fungi | |
| Agaricus abruptibulbus Peck | √ | | | | | | SS | Edible, Poisonous | Agarics | ON683434 |
| Agaricus arvensis Schaeff. | | | | | √ | | SS | Edible, Medicinal | Agarics | |
| Agaricus bresadolanus Bohus | √ | | | | | | SS | Edible, Medicinal | Agarics | |
| Agaricus campestris L. | √ | | √ | | √ | √ | SS | Edible, Medicinal | Agarics | |
| Agaricus comtulus Fr. | √ | | | √ | | | SS | Edible | Agarics | |
| Agaricus micromegethus Peck | √ | | | | √ | √ | SS | Edible, Medicinal | Agarics | |
| Agaricus moelleri Wasser | | | √ | | | √ | SS | Edible, Poisonous | Agarics | ON683435 |
| Agaricus perrarus Fr. | | | | | | √ | SS | Others | Agarics | |
| Agaricus placomyces Peck | | | | | √ | | SS | Edible, Medicinal | Agarics | |
| Agaricus purpurellus F.H. Møller | | | | | | √ | SS | Others | Agarics | |
| Agaricus silvaticus Schaeff. | | | √ | √ | | | SS | Edible, Medicinal | Agarics | |
| Agaricus silvicolae-similis Bohus and Locsmándi | √ | | | | √ | | SS | Others | Agarics | |
| Agaricus subrufescens Peck | √ | | | | √ | | SS | Edible, Medicinal | Agarics | |
| Agaricus subrutilescens (Kauffman) Hotson and D.E. Stuntz | √ | | √ | | | √ | SS | Edible, Medicinal | Agarics | |
| Agaricus sylvaticus Schaeff. | √ | | | | | √ | SS | Edible, Medicinal | Agarics | |
| Agrocybe pediades (Fr.) Fayod | | | | | √ | √ | SS | Edible, Medicinal | Agarics | |
| Agrocybe praecox (Pers.) Fayod | √ | | √ | | √ | √ | SS | Edible, Medicinal | Agarics | ON683416 |
| Aleuria aurantia (Pers.) Fuckel | √ | | | | √ | | SS | Edible, Medicinal, Poisonous | Larger Ascomycetes | |
| Aleurodiscus stereoides Yasuda | | | | | | √ | SS | Others | PHT fungi | |
| Amanita ceciliae (Berk. and Broome) Bas | √ | | | √ | | | EM | Others | Agarics | ON683421 |
| Amanita fulva Fr. | | | | | √ | √ | EM | Others | Agarics | |
| Amanita hemibapha (Berk. and Broome) Sacc. | | | √ | | √ | | EM | Edible, Medicinal | Agarics | |
| Amanita imazekii T. Oda, C. Tanaka and Tsuda | √ | | | √ | | | EM | Edible | Agarics | |
| Amanita longistriata S. Imai | √ | | √ | | √ | | EM | Poisonous | Agarics | |
| Amanita nivalis Grev. | √ | | | √ | | | EM | Edible | Agarics | |
| Amanita pantherina (DC.) Krombh | √ | | √ | √ | √ | | EM | Poisonous | Agarics | |
| Amanita phalloides (Vaill. ex Fr.) Link | √ | | | | | √ | EM | Others | Agarics | ON683436 |
| Amanita porphyria Alb. and Schwein. | | | √ | | | | EM | Others | Agarics | ON683437 |
| Amanita spreta (Peck) Sacc. | | | | | √ | | EM | Others | Agarics | |
| Amanita subjunquillea S. Imai | √ | | √ | √ | | √ | EM | Poisonous | Agarics | ON683438 |
| Amanita vaginata (Bull.) Lam. | √ | | √ | √ | √ | √ | EM | Edible, Medicinal, Poisonous | Agarics | ON683439 |
| Amanita virosa Secr. | √ | | | √ | | | EM | Medicinal, Poisonous | Agarics | |
| Amanitopsis fulva (Fr.) W.G. Sm. | | | | | √ | | EM | Others | Agarics | |
| Ampulloclitocybe clavipes (Pers.) Redhead, Lutzoni, Moncalvo and Vilgalys | √ | √ | √ | √ | √ | √ | LS | Edible, Medicinal, Poisonous | Agarics | |
| Apioperdon pyriforme (Schaeff.) Vizzini | √ | | | √ | √ | | EM | Edible, Medicinal | Gasteroid fungi | |
| Armillaria borealis Marxm. and Korhonen | √ | | | √ | | | WS | Edible, Medicinal | Agarics | |
| Armillaria cepistipes Velen. | √ | | | | √ | | WS | Others | Agarics | |
| Armillaria gallica Marxm. and Romagn. | √ | | | | | | WS | Edible, Medicinal | Agarics | ON683440 |
| Armillaria mellea (Vahl) P. Kumm. | √ | √ | | √ | √ | | WS | Edible, Medicinal | Agarics | |
| Armillaria ostoyae (Romagn.) Herink | √ | √ | | | | | WS | Edible, Medicinal | Agarics | |
| Armillaria sinapina Bérubé and Dessur. | √ | | | | | | WS | Edible, Medicinal | Agarics | ON683422 |
| Armillariella mellea (Vahl) P. Karst. | | √ | √ | | √ | √ | WS | Edible, Medicinal | Agarics | |
| Artomyces pyxidatus (Pers.) Jülich | √ | √ | √ | | √ | | WS | Edible | Coral fungi | |
| Ascocoryne cylichnium (Tul.) Korf | | √ | √ | | √ | | WS | Others | Larger Ascomycetes | |
| Astraeus hygrometricus (Pers.) Morgan | √ | | | | | | SS | Medicinal | Gasteroid fungi | |
| Atheniella adonis (Bull.) Redhead, Moncalvo, Vilgalys, Desjardin, and B.A. Perry | √ | | | √ | | | LS | Others | Agarics | |
| Auricularia cornea Ehrenb. | | √ | √ | √ | √ | √ | WS | Edible, Medicinal | Jelly fungi | |
| Auricularia heimuer F. Wu, B.K. Cui, and Y.C. Dai | | √ | √ | √ | | | WS | Edible, Medicinal | Jelly fungi | |
| Auricularia mesenterica (Dicks.) Pers. | | √ | | | √ | √ | WS | Others | Jelly fungi | |
| Auricularia nigricans (Sw.) Birkebak, Looney and Sánchez-García | | | √ | | √ | √ | WS | Edible, Medicinal | Jelly fungi | |
| Auriscalpium vulgare Gray | | | √ | | √ | √ | WS | Others | PHT fungi | |
| Baeospora myriadophylla (Peck) Singer | | √ | | | | | LS | Others | Agarics | |
| Bisporella sulfurina (Quél.) S.E. Carp. | √ | | √ | | | | WS | Others | Larger Ascomycetes | |
| Bjerkandera adusta (Willd.) P. Karst. | √ | | | | √ | | WS | Medicinal | PHT fungi | |
| Bolbitius vitellinus (Pers.) Fr. | | | | | √ | | SS | Others | Agarics | |
| Bolbitius vitellinus (Pers.) Fr. | √ | | | √ | | | EM | Others | Boletes | |
| Boletinellus merulioides (Schwein.) Murrill | | | √ | | | √ | EM | Others | Boletes | |
| Boletus edulis Bull. | √ | | √ | √ | √ | | EM | Edible | Boletes | |
| Boletus subtomentosus J.A. Palmer | √ | | | | | | EM | Others | Boletes | |
| Boletus subvelutipes Peck | | | | | | √ | EM | Others | Boletes | |
| Boletus yunnanensis W.F. Chiu | | | | | √ | | EM | Poisonous | Boletes | |
| Bovista pusilla (Batsch) Pers. | √ | | | | | | SS | Others | Gasteroid fungi | |
| Bovista pusilliformis (Kreisel) Kreisel | | | | | | √ | SS | Others | Gasteroid fungi | |
| Bulgaria inquinans (Pers.) Fr. | | √ | | | | | WS | Edible, Medicinal, Poisonous | Larger Ascomycetes | |
| Byssomerulius corium (Pers.) Parmasto | | | | | | √ | WS | Others | PHT fungi | |
| Calocera cornea (Batsch) Fr. | √ | √ | | | | | WS | Others | Jelly fungi | |
| Calocera viscosa (Pers.) Fr. | | | √ | | | | WS | Medicinal, Poisonous | Jelly fungi | |
| Calocybe gambosa (Fr.) Donk | √ | | | | | √ | SS | Edible, Medicinal | Agarics | |
| Calocybe ionides (Bull.) Donk | | | | | | √ | SS | Edible | Agarics | |
| Calvatia craniiformis (Schwein.) Fr. ex De Toni | √ | | | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Calvatia lilacina (Mont. and Berk.) Henn. | | | | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Calvatia tatrensis Hollós | | | √ | | | | SS | Medicinal | Gasteroid fungi | |
| Camarophyllus pratensis (Pers.) P. Kumm. | | | | | √ | | SS | Others | Agarics | |
| Campanella tristis (G. Stev.) Segedin | √ | √ | | | | | WS | Others | Agarics | |
| Cantharellus cibarius Fr. | √ | √ | √ | | √ | √ | EM | Edible, Medicinal | Cantharelloid fungi | ON683495 |
| Cantharellus minor Peck | √ | | | | √ | | EM | Edible, Medicinal | Cantharelloid fungi | |
| Cerioporus squamosus (Huds.) Quél. | √ | | √ | | √ | √ | WS | Medicinal | PHT fungi | |
| Cerioporus varius (Pers.) Zmitr. and Kovalenko | | | | | √ | | WS | Edible | PHT fungi | |
| Ceriporia tarda (Berk.) Ginns | √ | | | √ | | | WS | Others | PHT fungi | |
| Cheilymenia coprinaria (Cooke) Boud. | | | | | √ | √ | WS | Others | Larger Ascomycetes | |
| Chlorociboria aeruginascens (Nyl.) Kanouse | | | | √ | √ | | WS | Others | Larger Ascomycetes | |
| Chroogomphus purpurascens (Lj.N. Vassiljeva) M.M. Nazarova | | √ | | √ | | | EM | Edible | Boletes | |
| Chroogomphus roseolus Y.C. Li and Zhu L. Yang | | | | | √ | | EM | Edible, Medicinal | Boletes | |
| Chroogomphus rutilus (Schaeff.) O.K. Mill. | | √ | √ | | √ | √ | EM | Edible, Medicinal | Boletes | ON683423 |
| Chroogomphus tomentosus (Murrill) O.K. Mill. | | | | | √ | √ | EM | Others | Boletes | |
| Chrysomphalina aurantiaca (Peck) Redhead | √ | | | | √ | √ | WS | Others | Agarics | |
| Clavaria fragilis Holmsk. | | | | √ | | √ | SS | Medicinal, Poisonous | Coral fungi | |
| Clavaria vermicularis Sw. | | | √ | | | | SS | Edible | Coral fungi | |
| Clavariadelphus pistillaris (L.) Donk | | | √ | | √ | | LS | Edible, Poisonous | Coral fungi | ON683441 |
| Clavicorona pyxidata (Pers.) Doty | | | | | | √ | WS | Others | Coral fungi | |
| Clavulina coralloides (L.) J. Schröt. | √ | | | | √ | | LS | Edible | Coral fungi | ON683424 |
| Clavulinopsis corniculata (Schaeff.) Corner | | | √ | | | √ | LS | Edible | Coral fungi | |
| Clavulinopsis fusiformis (Sowerby) Corner | | | √ | √ | | | LS | Edible | Coral fungi | |
| Clavulinopsis helvola (Pers.) Corner | | | | | | √ | LS | Edible | Coral fungi | |
| Clitocybe infundibuliformis (Schaeff.) Quél. | | | | √ | √ | √ | LS | Edible, Medicinal | Agarics | |
| Clitocybe nebularis (Batsch) P. Kumm. | | √ | | | | | LS | Edible, Medicinal | Agarics | |
| Clitocybe odora (Bull.) P. Kumm. | | | | | | √ | LS | Edible, Medicinal | Agarics | ON683417 |
| Clitocybe phyllophila (Pers.) P. Kumm. | √ | | | √ | √ | | LS | Poisonous | Agarics | |
| Clitopilus prunulus (Scop.) P. Kumm. | √ | √ | | √ | | | LS | Edible | Agarics | |
| Collybia nivea (Mont.) Dennis | | | | | | √ | SS | Others | Agarics | |
| Collybiopsis confluens (Pers.) R.H. Petersen | | √ | √ | | √ | √ | SS | Others | Agarics | ON683418 |
| Collybiopsis peronata (Bolton) R.H. Petersen | √ | | √ | | √ | | SS | Others | Agarics | |
| Coltricia cinnamomea (Jacq.) Murrill | | √ | √ | | √ | √ | SS | Others | PHT fungi | |
| Coltricia perennis (L.) Murrill | √ | √ | | | | | SS | Others | PHT fungi | |
| Connopus acervatus (Fr.) K.W. Hughes, Mather and R.H. Petersen | | √ | √ | √ | √ | √ | SS | Edible | Agarics | |
| Conocybe lactea (J.E. Lange) Métrod | | | √ | | √ | √ | SS | Medicinal | Agarics | |
| Conocybe tenera (Schaeff.) Kühner | | | | | √ | | SS | Poisonous | Agarics | |
| Coprinellus disseminatus (Pers.) J.E. Lange | √ | | √ | √ | √ | √ | SS | Poisonous | Agarics | |
| Coprinellus micaceus (Bull.) Vilgalys, Hopple and Jacq. Johnson | √ | √ | √ | √ | √ | | SS | Medicinal, Poisonous | Agarics | |
| Coprinopsis atramentaria (Bull.) Redhead, Vilgalys and Moncalvo | | √ | √ | | √ | | SS | Edible, Medicinal, Poisonous | Agarics | |
| Coprinopsis cinerea (Schaeff.) Redhead, Vilgalys and Moncalvo | | | | | √ | √ | SS | Medicinal | Agarics | |
| Coprinopsis insignis (Peck) Redhead, Vilgalys & Moncalvo | | | √ | | | | SS | Medicinal | Agarics | |
| Coprinopsis lagopus (Fr.) Redhead, Vilgalys and Moncalvo | √ | √ | √ | | | | SS | Medicinal | Agarics | |
| Coprinopsis picacea (Bull.) Redhead, Vilgalys and Moncalvo | | | | | √ | | SS | Poisonous | Agarics | |
| Coprinus comatus (O.F. Müll.) Pers. | | √ | √ | | | √ | DS | Edible, Medicinal, Poisonous | Agarics | |
| Coprinus micaceus (Bull.) Fr. | | | | | | √ | DS | Medicinal, Poisonous | Agarics | |
| Coprinus plicatilis (Curtis) Fr. | | √ | √ | | | | DS | Others | Agarics | |
| Coprinus sterquilinus (Fr.) Fr. | √ | | √ | √ | | | DS | Medicinal | Agarics | |
| Cordyceps farinosa (Holmsk.) Kepler, B. Shrestha and Spatafora | | | | | | √ | EI | Others | Larger Ascomycetes | |
| Cordyceps militaris (L.) Fr. | | √ | √ | | √ | √ | EI | Edible, Medicinal | Larger Ascomycetes | |
| Cordyceps nutans Pat. | | √ | | | | | EI | Others | Larger Ascomycetes | |
| Coriolopsis gallica (Fr.) Ryvarden | | | √ | | √ | √ | WS | Others | PHT fungi | |
| Cortinarius alboviolaceus (Pers.) Fr. | | √ | | | | | EM | Edible | Agarics | |
| Cortinarius armillatus (Fr.) Fr. | | √ | | | | | EM | Others | Agarics | |
| Cortinarius bovinus Fr. | | | | | √ | | EM | Edible, Medicinal | Agarics | |
| Cortinarius caerulescens (Schaeff.) Fr. | | | √ | | √ | | EM | Others | Agarics | |
| Cortinarius callochrous (Pers.) Gray | √ | | √ | √ | | | EM | Edible, Poisonous | Agarics | |
| Cortinarius caperatus (Pers.) Fr. | | | √ | | | | EM | Edible, Medicinal | Agarics | |
| Cortinarius cinnamomeus (L.) Gray | | | √ | √ | | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Cortinarius collinitus (Sowerby) Gray | | | | | √ | | EM | Edible, Medicinal | Agarics | |
| Cortinarius colymbadinus Fr. | | | | | √ | | EM | Others | Agarics | |
| Cortinarius elatior Fr. | √ | | | | √ | | EM | Others | Agarics | |
| Cortinarius galeroides Hongo | √ | | √ | | | | EM | Others | Agarics | |
| Cortinarius lilacinus Peck | | | | √ | | | EM | Poisonous | Agarics | |
| Cortinarius longipes Peck | | | | | | √ | EM | Others | Agarics | |
| Cortinarius multiformis Fr. | √ | | | | | | EM | Edible | Agarics | |
| Cortinarius orellanus Fr. | √ | | | | | | EM | Poisonous | Agarics | |
| Cortinarius purpurascens Fr. | | | √ | | | | EM | Edible | Agarics | ON683425 |
| Cortinarius sanguineus (Wulfen) Gray | | | √ | | | √ | EM | Medicinal, Poisonous | Agarics | |
| Cortinarius trivialis J.E. Lange | √ | | | | | | EM | Edible, Poisonous | Agarics | |
| Cotylidia diaphana (Cooke) Lentz | √ | | | | | | LS | Others | PHT fungi | |
| Cotylidia komabensis (Henn.) D.A. Reid | | | √ | | √ | √ | LS | Others | PHT fungi | |
| Craterellus undulatus (Pers.) E. Campo and Papetti | √ | | | | √ | √ | EM | Edible | Agarics | |
| Crepidotus applanatus (Pers.) P. Kumm. | | √ | √ | | | | WS | Others | Agarics | |
| Crepidotus badiofloccosus S. Imai | | √ | | | | | WS | Others | Agarics | |
| Crepidotus betulae Murrill | | √ | √ | | | | WS | Others | Agarics | |
| Crepidotus brasiliensis Rick | | √ | | √ | | | WS | Medicinal | Agarics | |
| Crepidotus epibryus (Fr.) Quél. | √ | | | | | | WS | Others | Agarics | |
| Crepidotus fulvotomentosus (Peck) Peck | | | √ | | | | WS | Others | Agarics | |
| Crepidotus herbarum Sacc. | | | √ | | | | WS | Others | Agarics | |
| Crepidotus mollis (Schaeff.) Staude | √ | √ | | | √ | √ | WS | Others | Agarics | |
| Crepidotus nephrode (Berk. and M.A. Curtis) Sacc. | | | √ | | | | WS | Others | Agarics | |
| Crepidotus nephrodes (Berk. and M.A. Curtis) Sacc. | √ | | | | | | WS | Others | Agarics | |
| Crepidotus palmularis (Berk. and M.A. Curtis) Sacc. | √ | | √ | | | | WS | Others | Agarics | |
| Crepidotus sulphurinus Imazeki and Toki | √ | √ | √ | | | | WS | Others | Agarics | |
| Crepidotus variabilis (Pers.) P. Kumm. | | | √ | | | | WS | Others | Agarics | |
| Crinipellis stipitaria (Fr.) Pat. | | | | | √ | | LS | Others | Agarics | |
| Cuphophyllus pratensis (Pers.) Bon | | √ | √ | | | | SS | Edible | Agarics | ON683442 |
| Cuphophyllus virgineus (Wulfen) Kovalenko | √ | | √ | | √ | | SS | Others | Agarics | |
| Cyanosporus caesius (Schrad.) McGinty | | √ | √ | | √ | √ | WS | Others | PHT fungi | |
| Cyathus stercoreus (Schwein.) De Toni | | | | | √ | | WS | Medicinal | Gasteroid fungi | |
| Cyathus striatus Willd. | | √ | | | | | WS | Medicinal | Gasteroid fungi | |
| Cyclocybe cylindracea (DC.) Vizzini and Angelini | | | | | √ | | SS | Edible, Medicinal | Agarics | |
| Cyclocybe erebia (Fr.) Vizzini and Matheny | | | | | | √ | SS | Edible, Medicinal | Agarics | |
| Cystoderma amianthinum (Scop.) Fayod | √ | √ | √ | | | | SS | Edible | Agarics | |
| Cystoderma fallax A.H. Sm. and Singer | | √ | | | | | SS | Edible | Agarics | |
| Cystodermella granulosa (Batsch) Harmaja | | √ | | | | | SS | Edible | Agarics | |
| Dacrymyces chrysospermus Berk. and M.A. Curtis | √ | √ | | | | √ | WS | Others | Jelly fungi | |
| Dacrymyces palmatus Bres. | | | √ | | | | WS | Medicinal | Jelly fungi | ON683443 |
| Dacryopinax spathularia (Schwein.) G.W. Martin | √ | √ | | √ | √ | √ | WS | Others | Jelly fungi | |
| Daedalea dickinsii Yasuda | | √ | √ | | √ | | WS | Medicinal | PHT fungi | ON683444 |
| Daedaleopsis confragosa (Bolton) J. Schröt. | | √ | | | √ | | WS | Others | PHT fungi | |
| Daedaleopsis tricolor (Bull.) Bondartsev and Singer | √ | √ | | | √ | | WS | Medicinal | PHT fungi | ON683426 |
| Daldinia concentrica (Bolton) Ces. and De Not. | √ | √ | √ | √ | √ | √ | WS | Medicinal, Poisonous | Larger Ascomycetes | |
| Daldinia grandis Child | | | | √ | | | WS | Others | Larger Ascomycetes | |
| Deconica coprophila (Bull.) P. Karst. | | | | | √ | √ | DS | Poisonous | Agarics | |
| Deconica merdaria (Fr.) Noordel. | | | | | √ | | DS | Poisonous | Agarics | |
| Desarmillaria tabescens (Scop.) R.A. Koch and Aime | √ | √ | √ | | | | SS | Edible, Medicinal, Poisonous | Agarics | |
| Descolea flavoannulata (Lj.N. Vassiljeva) E. Horak | | | | √ | √ | | SS | Edible | Agarics | |
| Dumontinia tuberosa (Bull.) L.M. Kohn | | | √ | | √ | √ | SS | Others | Larger Ascomycetes | |
| Entoloma abortivum (Berk. and M.A. Curtis) Donk | | √ | | | | | SS | Edible, Medicinal | Agarics | |
| Entoloma albipes Hesler | √ | | | | √ | | SS | Others | Agarics | |
| Entoloma chamaecyparidis (Hongo) Hongo | | | | | | √ | SS | Others | Agarics | |
| Entoloma clypeatum (L.) P. Kumm. | √ | √ | √ | | √ | √ | SS | Medicinal, Poisonous | Agarics | |
| Entoloma japonicum (Hongo) Hongo and Izawa | | | | | | √ | SS | Others | Agarics | |
| Entoloma lividum Quél. | | | √ | | | | SS | Others | Agarics | |
| Entoloma murinipes (Murrill) Hesler | √ | | | √ | | | SS | Others | Agarics | |
| Entoloma murrillii Hesler | √ | | | | | | SS | Others | Agarics | |
| Entoloma politum (Pers.) Noordel. | √ | | | √ | | | SS | Others | Agarics | |
| Entoloma rhodopolium (Fr.) P. Kumm. | √ | | √ | √ | √ | √ | SS | Medicinal, Poisonous | Agarics | |
| Entoloma sinuatum (Bull.) P. Kumm. | √ | √ | | | √ | | SS | Medicinal, Poisonous | Agarics | |
| Entoloma speculum (Fr.) Quél. | √ | | √ | √ | | | SS | Others | Agarics | |
| Entoloma umbilicatum Dennis | √ | | √ | | | | SS | Others | Agarics | |
| Entonaema liquescens Möller | √ | √ | | √ | | | WS | Others | Larger Ascomycetes | |
| Exidia glandulosa (Bull.) Fr. | √ | | √ | √ | √ | | WS | Edible, Poisonous | Jelly fungi | |
| Flammulaster erinaceellus (Peck) Watling | √ | √ | | | √ | √ | WS | Others | Agarics | |
| Flammulina filiformis (Z.W. Ge, X.B. Liu, and Zhu L. Yang) P.M. Wang, Y.C. Dai, E. Horak, and Zhu L. Yang | √ | √ | √ | √ | √ | √ | WS | Edible | Agarics | |
| Fomes fomentarius (L.) Fr. | √ | √ | | √ | | | WS | Medicinal | PHT fungi | ON683445 |
| Fomitopsis betulina (Bull.) B.K. Cui, M.L. Han, and Y.C. Dai | | | √ | | | | WS | Medicinal | PHT fungi | |
| Fomitopsis officinalis (Vill.) Bondartsev and Singer | | √ | √ | | | | WS | Medicinal | PHT fungi | |
| Fomitopsis pinicola (Sw.) P. Karst. | | √ | | | | | WS | Medicinal | PHT fungi | ON683446 |
| Galerina helvoliceps (Berk. and M.A. Curtis) Singer | | | | | | √ | WS | Poisonous | PHT fungi | |
| Galerina marginata (Batsch) Kühner | | √ | | | | √ | SS | Poisonous | PHT fungi | |
| Galerina vittaeformis (Fr.) Singer | | | | | √ | | SS | Others | Agarics | |
| Galiella amurensis (Lj.N. Vassiljeva) Raitv. | | √ | | | | | SS | Others | Larger Ascomycetes | |
| Ganoderma applanatum (Pers.) Pat. | √ | √ | √ | √ | √ | √ | WS | Medicinal | PHT fungi | |
| Ganoderma tsugae Murrill | √ | √ | | | √ | | WS | Medicinal | PHT fungi | |
| Geastrum fimbriatum Fr. | | √ | | √ | | | LS | Medicinal | Gasteroid fungi | |
| Geastrum pectinatum Pers. | | | | | | √ | LS | Others | Gasteroid fungi | |
| Geastrum saccatum Fr. | | | √ | √ | √ | √ | LS | Medicinal | Gasteroid fungi | |
| Geastrum triplex Jungh. | | √ | | | √ | √ | LS | Medicinal | Gasteroid fungi | |
| Gerronema albidum (Fr.) Singer | √ | | | | | √ | SS | Edible | Agarics | |
| Gliophorus psittacinu (Schaeff.) Herink | | | | | | √ | SS | Edible, Poisonous | Agarics | |
| Gloeophyllum sepiarium (Wulfen) P. Karst. | | √ | | | | | WS | Medicinal | PHT fungi | |
| Gloeophyllum subferrugineum (Berk.) Bondartsev and Singer | √ | | | √ | | | WS | Others | PHT fungi | |
| Gloeophyllum trabeum (Pers.) Murrill | | | | | √ | | WS | Medicinal | PHT fungi | |
| Gloeostereum incarnatum S. Ito and S. Imai | | √ | | | √ | √ | WS | Edible, Medicinal | PHT fungi | |
| Gomphidius maculatus (Scop.) Fr. | | √ | | | | | EM | Edible | Boletes | |
| Gomphus clavatus (Pers.) Gray | | | | | √ | | EM | Edible, Medicinal | Cantharelloid fungi | |
| Guepinia helvelloides (DC.) Fr. | √ | | | √ | | | SS | Edible | Jelly fungi | |
| Gymnopilus aeruginosus (Peck) Singer | | √ | √ | | | | WS | Others | Agarics | |
| Gymnopilus junonius (Fr.) P.D. Orton | | √ | | | | | WS | Others | Agarics | |
| Gymnopilus liquiritiae (Pers.) P. Karst. | | √ | | | √ | | WS | Medicinal, Poisonous | Agarics | |
| Gymnopilus penetrans (Fr.) Murrill | √ | | | | | | WS | Poisonous | Agarics | ON683447 |
| Gymnopus alkalivirens (Singer) Halling | | | | | √ | | LS | Others | Agarics | |
| Gymnopus androsaceus (L.) J.L. Mata and R.H. Petersen | | | √ | | √ | √ | LS | Medicinal | Agarics | |
| Gymnopus dryophilus (Bull.) Murril | √ | √ | √ | √ | √ | √ | LS | Edible, Poisonous | Agarics | ON683448 |
| Gymnopus erythropus (Pers.) Antonín, Halling and Noordel. | | | | √ | | | LS | Edible | Agarics | ON683449 |
| Gymnopus fusipes (Bull.) Gray | | | | | √ | √ | LS | Edible | Agarics | |
| Gymnopus ocior (Pers.) Antonín and Noordel. | | | | | | √ | LS | Edible | Agarics | |
| Gymnopus polyphyllus (Peck) Halling | √ | | | | | | LS | Others | Agarics | |
| Harrya chromipes (Frost) Halling, Nuhn, Osmundson and Manfr. Binder | | | | | √ | | EM | Others | Boletes | |
| Hebeloma hiemale Bres. | | | √ | | | | SS | Others | Agarics | |
| Hebeloma radicosum (Bull.) Ricken | | | √ | | | | SS | Edible, Poisonous | Agarics | |
| Helvella atra J. König | | | | | | √ | SS | Edible | Larger Ascomycetes | |
| Helvella crispa (Scop.) Fr. | √ | √ | √ | √ | √ | √ | SS | Edible, Poisonous | Larger Ascomycetes | |
| Helvella elastica Bull. | √ | | √ | √ | √ | √ | SS | Edible, Poisonous | Larger Ascomycetes | |
| Helvella ephippium Lév. | | | | | √ | √ | SS | Edible | Larger Ascomycetes | |
| Helvella lacunosa Afzel. | | | √ | | | √ | SS | Edible, Medicinal | Larger Ascomycetes | |
| Hemistropharia albocrenulata (Peck) Jacobsson and E. Larss. | | | | | | √ | WS | Edible, Poisonous | Agarics | |
| Hericium coralloides (Scop.) Pers. | | √ | | | | | WS | Edible, Medicinal | PHT fungi | |
| Hericium erinaceus (Bull.) Pers. | √ | √ | √ | | | | WS | Edible, Medicinal | PHT fungi | ON683494 |
| Heterobasidion insulare (Murrill) Ryvarden | | √ | √ | | | | WS | Others | PHT fungi | |
| Hohenbuehelia reniformis (G. Mey.) Singer | | √ | | | | √ | WS | Edible | Agarics | |
| Hohenbuehelia serotina (Pers.) Singer | | | | √ | | | WS | Others | Agarics | |
| Hortiboletus rubellus (Krombh.) Simonini, Vizzini and Gelardi | √ | | | | | √ | EM | Edible, Medicinal | Boletes | |
| Humaria hemisphaerica (F.H. Wigg.) Fuckel | √ | | √ | | √ | √ | SS | Others | Larger Ascomycetes | |
| Hydnum repandum L. | | | √ | √ | √ | √ | EM | Edible, Medicinal | PHT fungi | |
| Hygrocybe ceracea (Sowerby) P. Kumm. | √ | | | √ | | | SS | Edible | Agarics | |
| Hygrocybe chlorophana (Fr.) Wünsche | | | √ | | | | SS | Edible | Agarics | |
| Hygrocybe coccinea (Schaeff.) P. Kumm. | | | √ | | | | SS | Edible | Agarics | |
| Hygrocybe conica (Schaeff.) P. Kumm. | | √ | √ | √ | √ | | SS | Medicinal, Poisonous | Agarics | |
| Hygrocybe flavescens (Kauffman) Singer | | | √ | | | | SS | Poisonous | Agarics | |
| Hygrocybe marchii (Bres.) Singer | √ | | | √ | | | SS | Others | Agarics | |
| Hygrocybe miniata (Fr.) P. Kumm. | √ | | √ | | | √ | SS | Edible | Agarics | ON683450 |
| Hygrophorus chrysodon (Batsch) Fr. | √ | | | | √ | √ | SS | Edible | Agarics | |
| Hygrophorus eburneus (Bull.) Fr. | | | √ | √ | | | SS | Edible | Agarics | |
| Hygrophorus laurae Morgan | √ | | | | | √ | SS | Others | Agarics | |
| Hygrophorus lucorum Kalchbr. | √ | √ | | | √ | | SS | Edible, Medicinal | Agarics | |
| Hygrophorus occidentalis A.H. Sm. and Hesler | √ | | √ | | | | SS | Others | Agarics | |
| Hygrophorus olivaceo-albus (Fr.) Fr. | | | √ | | | | SS | Edible | Agarics | |
| Hygrophorus piceae Kühner | √ | | | √ | | | SS | Others | Agarics | |
| Hygrophorus pseudochrysaspis Hesler and A.H. Sm. | √ | | √ | | √ | | SS | Others | Agarics | |
| Hygrophorus russula (Schaeff. ex Fr.) Kauffman | √ | | √ | | | | SS | Edible | Agarics | ON683451 |
| Hymenochaete adusta (Lév.) Har. and Pat. | | | | | | √ | WS | Others | PHT fungi | |
| Hymenochaete corrugata (Fr.) Lév. | √ | √ | | | | √ | WS | Others | PHT fungi | |
| Hymenochaete xerantica (Berk.) S.H. He and Y.C. Dai | | | | | | √ | WS | Others | PHT fungi | |
| Hymenopellis radicata (Relhan) R.H. Petersen | | | | | √ | √ | SS | Others | Agarics | |
| Hymenoscyphus fructigenus (Bull.) Gray | | | | | | √ | WS | Others | Larger Ascomycetes | |
| Hypholoma capnoides (Fr.) P. Kumm. | | √ | | | | | WS | Medicinal, Poisonous | Agarics | |
| Hypholoma fasciculare (Huds.) P. Kumm. | | √ | √ | √ | √ | √ | WS | Medicinal, Poisonous | Agarics | |
| Hypholoma lateritium (Schaeff.) P. Kumm. | | √ | √ | | | | WS | Poisonous | Agarics | |
| Hypsizygus marmoreus (Peck) H.E. Bigelow | | √ | | | | | WS | Others | Agarics | ON683452 |
| Hypsizygus ulmarius (Bull.) Redhead | | √ | | √ | | | WS | Edible, Medicinal | Agarics | |
| Infundibulicybe geotropa (Bull.) Harmaja | √ | | | | | | SS | Edible, Medicinal, Poisonous | Agarics | |
| Infundibulicybe gibba (Pers.) P. Kumm. | √ | √ | | √ | √ | | LS | Edible | Agarics | ON683453 |
| Inocybe asterospora Quél. | | | | | √ | √ | EM | Poisonous | Agarics | |
| Inocybe calamistrata (Fr.) Gillet | | | | | √ | | EM | Poisonous | Agarics | |
| Inocybe changbaiensis T. Bau and Y.G. Fan | √ | | | √ | | | EM | Others | Agarics | |
| Inocybe cookei Bres. | | | | √ | | | EM | Poisonous | Agarics | |
| Inocybe earleana Kauffma | √ | | | √ | | | EM | Others | Agarics | |
| Inocybe euviolacea E. Ludw. | | | | | | √ | EM | Others | Agarics | |
| Inocybe fulvella Bres. | √ | | | | | | EM | Others | Agarics | |
| Inocybe geophylla P. Kumm. | √ | | √ | | √ | √ | EM | Poisonous | Agarics | |
| Inocybe napipes J.E. Lange | √ | | | | √ | | EM | Poisonous | Agarics | |
| Inocybe praetervisa Quél. | | | | | √ | | EM | Poisonous | Agarics | |
| Inocybe subalbidodisca Stangl and J. Veselský | √ | | | | √ | | EM | Others | Agarics | |
| Inocybe umbrinella Bres. | √ | | | √ | √ | | EM | Others | Agarics | |
| Inonotus hispidus (Bull.) P. Karst. | √ | | | | | | WS | Medicinal | PHT fungi | ON683454 |
| Inonotus obliquus (Fr.) Pilát | | | | √ | | | WS | Medicinal | PHT fungi | |
| Inosperma calamistratum (Fr.) Matheny and Esteve-Rav. | | | | | | √ | LS | Others | Agarics | |
| Irpex lacteus (Fr.) Fr. | | √ | | | √ | | WS | Others | PHT fungi | |
| Isaria japonica Yasuda | | | | | | √ | EI | Others | Larger Ascomycetes | |
| Junghuhnia nitida (Pers.) Ryvarden | | | | | | √ | WS | Edible | PHT fungi | |
| Kuehneromyces mutabilis (Schaeff.) Singer and A.H. Sm. | √ | √ | √ | | √ | | WS | Edible | Agarics | ON683455 |
| Laccaria alba Zhu L. Yang and Lan Wang | √ | | | √ | | | EM | Edible | Agarics | ON683456 |
| Laccaria amethystea (Bull.) Murrill | | | √ | √ | | √ | EM | Others | Agarics | ON683457 |
| Laccaria amethystina Cooke | √ | | | | | | EM | Others | Agarics | |
| Laccaria laccata (Scop.) Cooke | √ | | √ | √ | √ | | EM | Edible, Medicinal | Agarics | ON683458 |
| Laccaria proxima (Boud.) Pat. | √ | | √ | | | | EM | Others | Agarics | |
| Laccaria purpureobadia D.A. Reid | | | | √ | | | EM | Others | Agarics | |
| Laccaria tortilis (Bolton) Cooke | √ | | | | | | EM | Edible, Medicinal | Agarics | ON683459 |
| Lacrymaria lacrymabunda (Bull.) Pat. | | | | | √ | | SS | Poisonous | Agarics | |
| Lactarius acris (Bolton) Gray | | | | √ | | | EM | Others | Agarics | |
| Lactarius aurantiacus (Pers.) Gray | √ | | √ | | | | EM | Edible, Medicinal | Agarics | |
| Lactarius camphoratus (Bull.) Fr. | | | √ | | √ | | EM | Edible | Agarics | |
| Lactarius circellatus Fr. | | | √ | | | | EM | Edible | Agarics | |
| Lactarius deliciosus (L.) Gray | √ | | | | √ | √ | EM | Edible, Medicinal | Agarics | |
| Lactarius fuliginosus (Fr.) Fr. | | | √ | | | | EM | Edible | Agarics | |
| Lactarius hatsudake Nobuj. Tanaka | √ | | √ | | | √ | EM | Edible, Medicinal | Agarics | |
| Lactarius lignyotus Fr. | | | | | √ | | EM | Medicinal, Poisonous | Agarics | |
| Lactarius mitissimus (Fr.) Fr. | | | √ | | | | EM | Others | Agarics | |
| Lactarius piperatus (L.) Pers. | √ | | √ | √ | √ | √ | EM | Edible, Medicinal, Poisonous | Agarics | |
| Lactarius subdulcis (Pers.) Gray | √ | | | | | | EM | Edible | Agarics | |
| Lactarius theiogalus (Bull.) Gray | √ | | | | | | EM | Others | Agarics | |
| Lactarius torminosus (Schaeff.) Pers. | | | √ | | | √ | EM | Poisonous | Agarics | |
| Lactarius trivialis (Fr.) Fr. | | | | | √ | | EM | Others | Agarics | |
| Lactarius vellereus (Fr.) Fr. | √ | | √ | | | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Lactarius vietus (Fr.) Fr. | √ | | √ | √ | | | EM | Edible | Agarics | |
| Lactarius volemus (Fr.) Fr. | √ | | √ | | | | EM | Edible, Medicinal | Agarics | ON683460 |
| Lactarius zonarius (Bull.) Fr. | √ | | | | √ | | EM | Medicinal, Poisonous | Agarics | |
| Lactifluus subpiperatus (Hongo) Verbeken | √ | | | | √ | | EM | Others | Agarics | |
| Laetiporus cremeiporus Y. Ota and T. Hatt. | | √ | | | | | WS | Edible, Medicinal | PHT fungi | |
| Laetiporus montanus Černý ex Tomšovský and Jankovský | | √ | | | | | WS | Edible | PHT fungi | |
| Laetiporus sulphureus (Bull.) Murrill | | | √ | √ | √ | | WS | Edible, Medicinal | PHT fungi | ON683461 |
| Leccinum chromapes (Frost) Singer | | | √ | | | √ | EM | Others | Boletes | |
| Leccinum scabrum (Bull.) Gray | √ | | √ | | √ | √ | EM | Edible, Poisonous | Boletes | |
| Lentinellus flabelliformis (Bolton) S. Ito | √ | √ | | | | | WS | Others | Agarics | |
| Lentinellus ursinus (Fr.) Kühner | | √ | | | √ | √ | WS | Edible | Agarics | |
| Lentinula edodes (Berk.) Pegler | | √ | | | | | WS | Edible, Medicinal | Agarics | |
| Lentinus arcularius (Batsch) Zmitr. | √ | √ | √ | | √ | √ | WS | Medicinal | PHT fungi | |
| Lentinus brumalis (Pers.) Zmitr. | √ | | | | | | WS | Edible | PHT fungi | |
| Lentinus elmeri Bres. | | | √ | | | | WS | Others | PHT fungi | |
| Lentinus substrictus (Bolton) Zmitr. and Kovalenko | | √ | | √ | | | WS | Others | PHT fungi | |
| Lentinus tigrinus (Bull.) Fr. | | √ | | √ | | | WS | Edible, Medicinal | PHT fungi | |
| Lenzites albida (Fr.) Fr. | | | √ | | | | WS | Edible | PHT fungi | |
| Lenzites betulinus (L.) Fr. | | √ | | | √ | √ | WS | Medicinal | PHT fungi | |
| Lenzites repanda Fr. | | | √ | | | | WS | Poisonous | PHT fungi | |
| Leotia lubrica (Scop.) Pers. | | | √ | | | √ | SS | Others | Larger Ascomycetes | |
| Lepiota brunneoincarnata Chodat and C. Martín | | | | | | √ | LS | Poisonous | Agarics | |
| Lepiota castanea Quél. | √ | | | √ | √ | √ | LS | Poisonous | Agarics | |
| Lepiota clypeolaria (Bull.) P. Kumm. | √ | | | | √ | √ | LS | Poisonous | Agarics | ON683462 |
| Lepiota cristata (Bolton) P. Kumm. | √ | | √ | √ | √ | √ | LS | Poisonous | Agarics | |
| Lepiota erminea (Fr.) P. Kumm. | √ | | | | | √ | LS | Edible | Agarics | |
| Lepiota felina (Pers.) P. Karst. | √ | | √ | | √ | | LS | Others | Agarics | |
| Lepiota fusciceps Hongo | | | | | | √ | LS | Others | Agarics | |
| Lepiota magnispora Murrill | | | | √ | | | LS | Others | Agarics | |
| Lepista glaucocana (Bres.) Singer | √ | | | | | √ | LS | Edible | Agarics | |
| Lepista irina (Fr.) H.E. Bigelow | √ | √ | √ | | | | LS | Medicinal, Poisonous | Agarics | |
| Lepista nuda (Bull.) Cooke | √ | √ | √ | √ | √ | √ | LS | Edible, Medicinal | Agarics | ON683463 |
| Lepista personata (Fr.) Cooke | √ | | | | √ | | LS | Edible, Medicinal | Agarics | |
| Lepista sordida (Schumach.) Singer | | √ | | | √ | √ | LS | Edible, Medicinal | Agarics | |
| Leucoagaricus leucothites (Vittad.) Wasser | √ | | | | | √ | SS | Edible, Medicinal, Poisonous | Agarics | |
| Leucoagaricus rubrotinctus (Peck) Singer | √ | | √ | | √ | | SS | Others | Agarics | ON683464 |
| Leucocoprinus brebissonii (Godey) Locq. | √ | | | √ | | | SS | Others | Agarics | |
| Leucocybe candicans (Pers.) Vizzini, P. Alvarado, G. Moreno, and Consiglio | | | | | | √ | SS | Medicinal, Poisonous | Agarics | |
| Leucocybe connata (Schumach.) Vizzini, P. Alvarado, G. Moreno, and Consiglio | | | √ | | √ | | SS | Edible, Poisonous | Agarics | |
| Leucocybe houghtonii (W. Phillips) Halama and Pencak. | | | | √ | | | SS | Others | Agarics | |
| Lopharia cinerascens (Schwein.) G. Cunn. | | | | | | √ | WS | Others | PHT fungi | |
| Lycoperdon fuscum Huds. | | | | | | √ | SS | Edible, Medicinal | Gasteroid fungi | |
| Lycoperdon mammaeforme Pers. | | | √ | | √ | √ | SS | Others | Gasteroid fungi | |
| Lycoperdon pedicellatum Batsch | | | | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Lycoperdon perlatum Pers. | √ | √ | | | √ | √ | SS | Edible, Medicinal | Gasteroid fungi | ON683465 |
| Lycoperdon pusillum Hedw. | | | | | √ | | SS | Others | Gasteroid fungi | |
| Lycoperdon pyriforme Schaeff. | | √ | | | | | SS | Edible, Medicinal | Gasteroid fungi | |
| Lycoperdon umbrinum Pers. | √ | | √ | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Lyophyllum decastes (Fr.) Singer | | √ | | | √ | | EM | Edible, Medicinal | Agarics | |
| Lysurus mokusin (L.) Fr. | | | | | √ | √ | SS | Medicinal | Gasteroid fungi | |
| Macrocystidia cucumis (Pers.) Joss. | | | √ | | | √ | SS | Others | Agarics | |
| Macrolepiota procera (Scop.) Singer | √ | √ | √ | √ | √ | √ | SS | Edible, Medicinal, Poisonous | Agarics | |
| Mallocybe terrigena (Fr.) Matheny, Vizzini, and Esteve-Rav. | | | | √ | | | SS | Edible | Agarics | |
| Marasmiellus candidus (Fr.) Singer | | | | | √ | | WS | Others | Agarics | |
| Marasmiellus eburneus (Theiss.) Singer | √ | | √ | | | | WS | Others | Agarics | |
| Marasmiellus ramealis (Bull.) Singer | √ | | | √ | √ | √ | WS | Medicinal | Agarics | |
| Marasmius aurantiacus I. Hino | | | | | √ | | LS | Edible, Medicinal | Agarics | |
| Marasmius beniensis Singer | | | √ | | | | LS | Others | Agarics | |
| Marasmius chordalis Fr. | √ | | | | √ | | LS | Others | Agarics | |
| Marasmius cohaerens (Pers.) Cooke and Quél. | √ | | √ | | √ | √ | LS | Others | Agarics | |
| Marasmius epiphyllus (Pers.) Fr. | | | √ | | | | LS | Others | Agarics | |
| Marasmius floriceps Berk. and M.A. Curtis | √ | | | | | | LS | Others | Agarics | |
| Marasmius maximus Hongo | √ | | √ | | √ | | LS | Edible | Agarics | ON683427 |
| Marasmius occultatiformis Antonín, Ryoo, and H.D. Shin | √ | | | | √ | | LS | Others | Agarics | ON683419 |
| Marasmius oreades (Bolton) Fr. | √ | | | | √ | √ | LS | Edible, Medicinal | Agarics | |
| Marasmius pallidocephalus Gilliam | √ | | | √ | | | LS | Others | Agarics | |
| Marasmius polylepidis Dennis | √ | | | √ | | | LS | Others | Agarics | |
| Marasmius pseudoniveus Singer | | | √ | | | | LS | Others | Agarics | |
| Marasmius pulcherripes Peck | | | | | | √ | LS | Others | Agarics | |
| Marasmius riparius Singer | √ | | √ | | √ | | LS | Others | Agarics | |
| Marasmius rotuloides Dennis | √ | | | √ | | | LS | Others | Agarics | |
| Marasmius sessiliaffinis Singer | | | √ | | | | LS | Others | Agarics | |
| Marasmius siccus (Schwein.) Fr. | √ | | √ | | | √ | LS | Others | Agarics | |
| Megacollybia platyphylla (Pers.) Kotl. and Pouzar | | | √ | | | | WS | Others | Agarics | |
| Melanoleuca brevipes (Bull.) Pat. | | | | | | √ | SS | Edible | Agarics | |
| Melanoleuca grammopodia (Bull.) Murrill | √ | | √ | | | | SS | Edible | Agarics | |
| Melanoleuca melaleuca (Pers.) Murrill | √ | | | | √ | √ | SS | Edible | Agarics | ON683466 |
| Melanoleuca strictipes (P. Karst.) Jul. Schäff. | √ | | | | | | SS | Edible | Agarics | |
| Melanoleuca stridula (Fr.) Singer | | | | | √ | √ | SS | Others | Agarics | |
| Melanoleuca verrucipes (Fr.) Singer | √ | | | √ | | √ | SS | Edible | Agarics | |
| Melastiza chateri (W.G. Sm.) Boud. | | | | | | √ | SS | Others | Larger Ascomycetes | |
| Microporus affinis (Blume and T. Nees) Kuntze | √ | | | | | | WS | Others | PHT fungi | |
| Microstoma floccosum (Sacc.) Raitv. | | √ | | | | | WS | Others | Larger Ascomycetes | |
| Morchella conica Pers. | | | | | | √ | SS | Edible | Larger Ascomycetes | |
| Morchella crassipes (Vent.) Pers. | | | | | √ | √ | SS | Edible, Medicinal | Larger Ascomycetes | |
| Morchella esculenta (L.) Pers. | √ | | √ | | √ | √ | SS | Edible, Medicinal | Larger Ascomycetes | |
| Morchella vulgaris (Pers.) Gray | | | √ | | | | SS | Edible, Medicinal | Larger Ascomycetes | |
| Mucidula brunneomarginata (Lj.N. Vassiljeva) R.H. Petersen | | √ | √ | √ | | | WS | Edible | Agarics | |
| Mucidula mucida (Schrad.) Pat. | √ | √ | √ | √ | | | WS | Others | Agarics | ON683467 |
| Mutinus caninus (Huds.) Fr. | | | √ | | | √ | SS | Poisonous | Gasteroid fungi | |
| Mycena alphitophora (Berk.) Sacc. | | | | | | √ | LS | Others | Agarics | |
| Mycena collybiiformis (Murrill) Murrill | √ | | | | | | LS | Others | Agarics | |
| Mycena debilis (Fr.) Quél. | √ | | √ | | | | LS | Others | Agarics | |
| Mycena epipterygia (Scop.) Gray | √ | | | | | | LS | Others | Agarics | |
| Mycena filopes (Bull.) P. Kumm. | | | | | √ | | LS | Others | Agarics | |
| Mycena flavescens Velen. | √ | | | √ | | | LS | Poisonous | Agarics | |
| Mycena fuliginella A.H. Sm. | √ | | √ | | | | LS | Others | Agarics | |
| Mycena galericulata (Scop.) Gray | √ | | √ | | √ | | LS | Edible, Medicinal | Agarics | ON683420 |
| Mycena haematopus (Pers.) P. Kumm. | √ | √ | √ | √ | √ | | LS | Medicinal, Poisonous | Agarics | ON683468 |
| Mycena inclinata (Fr.) Quél. | √ | | | | | | LS | Others | Agarics | |
| Mycena nucleata X. He and X.D. Fang | √ | | | √ | | | LS | Others | Agarics | |
| Mycena osmundicola J.E. Lange | | | √ | | | | LS | Others | Agarics | |
| Mycena pseudoandrosacea (Bull.) Z.S. Bi | √ | | √ | | | | LS | Others | Agarics | |
| Mycena pura (Pers.) P. Kumm. | √ | √ | √ | √ | √ | √ | LS | Medicinal, Poisonous | Agarics | ON683469 |
| Mycena rosea Gramberg | √ | | | √ | | | LS | Others | Agarics | |
| Mycena sanguinolenta (Alb. and Schwein.) P. Kumm. | √ | | | | | | LS | Others | Agarics | |
| Mycena subcana A.H. Sm. | √ | | | | | | LS | Others | Agarics | |
| Mycena subgracilis Métrod | √ | | | √ | | | LS | Others | Agarics | |
| Mycoleptodonoides pergamenea (Yasuda) Aoshima and H. Furuk. | | √ | | √ | | | WS | Others | PHT fungi | |
| Myxarium nucleatum Wallr. | √ | | √ | √ | | | WS | Others | Larger Ascomycetes | |
| Naematelia aurantialba (Bandoni and M. Zang) Millanes and Wedin | √ | | √ | | | | WS | Edible, Medicinal | Jelly fungi | |
| Neofavolus alveolaris (DC.) Sotome and T. Hatt. | | | √ | | | | WS | Others | PHT fungi | ON683470 |
| Neolentinus adhaerens (Alb. and Schwein.) Redhead and Ginns | | √ | | | | | WS | Edible, Medicinal | PHT fungi | |
| Neolentinus cyathiformis (Schaeff.) Della Magg. and Trassin. | | √ | | | | | WS | Others | PHT fungi | |
| Neolentinus lepideus (Fr.) Redhead and Ginns | √ | √ | √ | √ | √ | √ | WS | Edible, Medicinal, Poisonous | PHT fungi | ON683471 |
| Nidula niveotomentosa (Henn.) Lloyd | | | | | | √ | WS | Medicinal | Agarics | |
| Omphalina lilaceorosea Svrček and Kubička | | √ | | | | | LS | Others | Agarics | |
| Omphalotus guepiniformis (Berk.) Neda | √ | √ | √ | | | | WS | Poisonous | Agarics | |
| Onnia tomentosa (Fr.) P. Karst. | | | | | √ | | WS | Others | PHT fungi | |
| Ophiocordyceps nutans (Pat.) G.H. Sung, J.M. Sung, Hywel-Jones, and Spatafora | | | | √ | | | EI | Medicinal | Larger Ascomycetes | |
| Ophiocordyceps sphecocephala (Klotzsch ex Berk.) G.H. Sung, J.M. Sung, Hywel-Jones, and Spatafora | | √ | | | | √ | EI | Medicinal | Larger Ascomycetes | |
| Ossicaulis lignatilis (Pers.) Redhead and Ginns | | √ | | | | √ | WS | Edible | Agarics | |
| Osteina undosa (Peck) Zmitr. | | | | | | √ | WS | Others | PHT fungi | |
| Otidea cochleata (L.) Fuckel | √ | | √ | √ | √ | √ | SS | Poisonous | Larger Ascomycetes | |
| Otidea leporina (Batsch) Fuckel | | | | √ | | | SS | Others | Larger Ascomycetes | |
| Oudemansiella radicata (Relhan) Singer | | √ | | | | | SS | Others | Agarics | |
| Panaeolus campanulatus (L.) Quél. | | | √ | | | | DS | Others | Agarics | |
| Panaeolus fimicola (Pers.) Gillet | √ | | √ | | | | DS | Poisonous | Agarics | |
| Panaeolus papilionaceus (Bull.) Quél. | √ | | | | √ | √ | DS | Poisonous | Agarics | |
| Panellus stipticus (Bull.) P. Karst. | √ | √ | | | √ | | WS | Medicinal, Poisonous | Agarics | ON683472 |
| Panus conchatus (Bull.) Fr. | | √ | | | | | WS | Edible, Medicinal | Agarics | ON673473 |
| Panus rudis Fr. | | | | | √ | √ | WS | Others | Agarics | |
| Paralepista splendens (Pers.) Vizzini | √ | | √ | | √ | | SS | Others | Agarics | |
| Paralepistopsis acromelalga (Ichimura) Vizzini | | | | | | √ | SS | Poisonous | Agarics | |
| Parasola plicatilis (Curtis) Redhead, Vilgalys, and Hopple | √ | | | √ | | | SS | Medicinal | Agarics | |
| Parasola leiocephala (P.D. Orton) Redhead, Vilgalys and Hopple | √ | √ | | | | | SS | Others | Agarics | |
| Paxillus involutus (Batsch) Fr. | | √ | √ | | √ | √ | EM | Medicinal, Poisonous | Boletes | |
| Perenniporia inflexibilis (Berk.) Ryvarden | √ | | | √ | | | WS | Others | PHT fungi | |
| Perenniporia medulla-panis (Jacq.) Donk | | | √ | | | √ | WS | Others | PHT fungi | |
| Perenniporia subacida (Peck) Donk | | | √ | | | √ | WS | Medicinal | PHT fungi | |
| Peziza ampliata Pers. | | | √ | | | | SS | Others | Larger Ascomycetes | |
| Peziza badia Pers. | | √ | | √ | √ | √ | SS | Poisonous | Larger Ascomycetes | |
| Peziza praetervisa Bres. | √ | | √ | | | √ | SS | Others | Larger Ascomycetes | |
| Peziza sylvestris (Boud.) Sacc. and Traverso | | | | | √ | | SS | Others | Larger Ascomycetes | |
| Peziza vesiculosa Bolton | | √ | √ | | √ | √ | SS | Edible, Poisonous | Larger Ascomycetes | |
| Phaeoclavulina abietina (Pers.) Giachini | | | √ | | | | LS | Others | Coral fungi | |
| Phaeoclavulina flaccida (Fr.) Giachini | | | √ | | | | LS | Poisonous | Coral fungi | |
| Phaeolepiota aurea (Matt.) Maire | | | | √ | √ | | SS | Edible, Medicinal, Poisonous | Agarics | |
| Phaeolus schweinitzii (Fr.) Pat. | | | | | √ | | WS | Medicinal | PHT fungi | |
| Phaeotremella foliacea (Pers.) Wedin, J.C. Zamora and Millanes | | | √ | | √ | | EM | Edible, Medicinal | Jelly fungi | |
| Phallus flavocostatus Kreisel | √ | | √ | | | | SS | Others | Gasteroid fungi | ON683474 |
| Phallus hadriani Vent. | | | | | | √ | SS | Others | Gasteroid fungi | |
| Phallus impudicus L. | √ | | | | √ | √ | SS | Edible, Medicinal | Gasteroid fungi | |
| Phallus indusiatus Vent. | | | | | √ | | SS | Others | Gasteroid fungi | |
| Phallus rubicundus (Bosc) Fr. | √ | | | | | | SS | Medicinal, Poisonous | Gasteroid fungi | |
| Phallus rugulosus (E. Fisch.) Lloyd | | | | | √ | √ | SS | Others | Gasteroid fungi | |
| Phellinus gilvus (Schwein.) Pat. | | | | | √ | √ | WS | Others | PHT fungi | |
| Phellinus igniarius (L.) Quél. | | √ | | | | | WS | Medicinal | PHT fungi | |
| Phellodon fuligineoalbus (J.C. Schmidt) R.E. Baird | | | √ | | | | WS | Others | PHT fungi | |
| Phellodon tomentosus (L.) Banker | √ | | | | | | WS | Edible | PHT fungi | |
| Phillipsia domingensis (Berk.) Berk. ex Denison | | √ | √ | | | | WS | Others | Larger Ascomycetes | |
| Phlebia tremellosa (Schrad.) Nakasone and Burds. | | | | | √ | | WS | Medicinal | PHT fungi | |
| Phloeomana minutula (Sacc.) Redhead | √ | | | | | | WS | Others | Agarics | |
| Phloeomana speirea (Fr.) Redhead | | | √ | | √ | √ | WS | Others | Agarics | |
| Pholiota adiposa (Batsch) P. Kumm. | √ | √ | | √ | | √ | WS | Edible, Medicinal | Agarics | |
| Pholiota aurivella (Batsch) P. Kumm. | | √ | √ | √ | √ | √ | WS | Edible, Medicinal | Agarics | |
| Pholiota flammans (Batsch) P. Kumm. | √ | √ | √ | | √ | | WS | Edible, Medicinal, Poisonous | Agarics | |
| Pholiota hiemalis A.H. Sm. and Hesler | | √ | | √ | | | WS | Others | Agarics | |
| Pholiota lenta (Pers.) Singer | | | √ | | | | WS | Edible, Medicinal | Agarics | |
| Pholiota lubrica (Pers.) Singer | √ | | | | | | WS | Edible, Medicinal, Poisonous | Agarics | ON683475 |
| Pholiota mutabilis (Schaeff.) P. Kumm. | | | | | √ | | WS | Others | Agarics | |
| Pholiota spumosa (Fr.) Singer | √ | | | √ | √ | | WS | Edible, Medicinal | Agarics | ON683476 |
| Pholiota squarrosa (Vahl) P. Kumm. | √ | √ | √ | | √ | | WS | Edible, Medicinal, Poisonous | Agarics | |
| Pholiota squarrosoides (Peck) Sacc. | √ | √ | √ | | √ | | WS | Edible, Poisonous | Agarics | ON683477 |
| Pholiota subflavida (Murrill) A.H. Sm. and Hesler | √ | | | | √ | | WS | Others | Agarics | |
| Pholiota tuberculosa (Schaeff.) P. Kumm. | | | √ | | | | WS | Others | Agarics | |
| Pholiota veris A.H. Sm. and Hesler | | | | | | √ | WS | Others | Agarics | |
| Pholiota vinaceobrunnea A.H. Sm. and Hesler | | √ | | | | | WS | Others | Agarics | |
| Pholiota terrestris Overh. | √ | √ | √ | | √ | | WS | Edible, Medicinal, Poisonous | Agarics | |
| Phyllotopsis nidulans (Pers.) Singer | | | √ | | | √ | WS | Edible | Agarics | |
| Physalacria lateriparies X. He and F.Z. Xue | | | | | √ | | WS | Others | Agarics | |
| Picipes badius (Pers.) Zmitr. and Kovalenko | | | √ | | | √ | WS | Others | PHT fungi | ON683478 |
| Picipes melanopus (Pers.) Zmitr. and Kovalenko | | | √ | | | | WS | Others | PHT fungi | |
| Piptoporus betulinus (Bull.) P. Karst. | | √ | | √ | | | WS | Medicinal | PHT fungi | |
| Plectania melastoma (Sowerby) Fuckel | √ | | | √ | | | WS | Others | Larger Ascomycetes | |
| Pleuroflammula flammea (Murrill) Singer | | √ | | | | | SS | Others | Agarics | |
| Pleuroflammula multifolia (Peck) E. Horak | √ | | | | | | SS | Others | Agarics | |
| Pleurotus citrinopileatus Singer | √ | √ | √ | | | | WS | Edible, Medicinal | Agarics | ON683479 |
| Pleurotus cornucopiae (Paulet) Quél. | √ | √ | √ | | √ | | WS | Edible, Medicinal | Agarics | |
| Pleurotus corticatus (Fr.) P. Kumm. | | | √ | | | | WS | Others | Agarics | |
| Pleurotus dryinus (Pers.) P. Kumm. | √ | | | √ | | | WS | Edible, Medicinal | Agarics | |
| Pleurotus limpidus (Fr.) P. Karst. | √ | | | | √ | | WS | Edible | Agarics | |
| Pleurotus ostreatus (Jacq.) P. Kumm. | √ | √ | √ | √ | √ | √ | WS | Edible, Medicinal | Agarics | ON683480 |
| Pleurotus pulmonarius (Fr.) Quél. | √ | √ | √ | | √ | √ | WS | Edible, Medicinal | Agarics | ON683481 |
| Pleurotus spodoleucus (Fr.) Quél. | √ | | | √ | | | WS | Edible, Medicinal | Agarics | |
| Pluteus atricapillus (Batsch) Fayod | | | √ | | | | WS | Others | Agarics | |
| Pluteus atromarginatus (Konrad) Kühner | | | | | | √ | WS | Edible | Agarics | |
| Pluteus aurantiorugosus (Trog) Sacc. | | √ | √ | | √ | | WS | Edible | Agarics | ON683482 |
| Pluteus cervinus (Schaeff.) P. Kumm. | √ | √ | | √ | √ | √ | WS | Edible | Agarics | |
| Pluteus depauperatus Romagn. | | | √ | | | | WS | Others | Agarics | |
| Pluteus leoninus (Schaeff.) P. Kumm. | | √ | √ | | √ | | WS | Edible | Agarics | |
| Pluteus nanus (Pers.) P. Kumm. | | | √ | | √ | | WS | Others | Agarics | |
| Pluteus petasatus (Fr.) Gillet | | | | | √ | √ | WS | Edible | Agarics | |
| Pluteus plautus (Weinm.) Gillet | √ | √ | | | | | WS | Others | Agarics | |
| Pluteus salicinus (Pers.) P. Kumm. | | √ | √ | | | | WS | Edible | Agarics | |
| Pluteus umbrosus (Pers.) P. Kumm. | √ | | | | | | WS | Edible | Agarics | |
| Podosordaria pedunculata (Dicks.) Dennis | | √ | | √ | | | WS | Others | Larger Ascomycetes | |
| Polyporus alveolaris (DC.) Bondartsev and Singer | | | | | √ | √ | WS | Others | PHT fungi | |
| Polyporus badius (Pers.) Schwein. | | | | | √ | | WS | Others | PHT fungi | |
| Polyporus brumalis (Pers.) Fr. | | √ | | | | | WS | Others | PHT fungi | |
| Polyporus conifericola H.J. Xue and L.W. Zhou | | √ | | | | | WS | Others | PHT fungi | ON683483 |
| Polyporus squamosus (Huds.) Fr. | | √ | | | | | WS | Medicinal | PHT fungi | |
| Polyporus tuberaster (Jacq. ex Pers.) Fr. | | √ | | √ | | | WS | Others | PHT fungi | |
| Polyporus varius (Pers.) Fr. | | √ | | | | | WS | Medicinal | PHT fungi | |
| Polystictus unicolor Rick | √ | | | | √ | | WS | Others | PHT fungi | |
| Polystictus versicolor (L.) Fr. | | | | | √ | | WS | Others | PHT fungi | |
| Psathyrella candolleana (Fr.) Maire | √ | √ | | | √ | √ | WS | Medicinal, Poisonous | Agarics | ON683484 |
| Psathyrella hydrophila (Bull.) Maire | | | | | √ | | WS | Others | Agarics | |
| Psathyrella multissima (S. Imai) Hongo | | √ | √ | | √ | √ | WS | Others | Agarics | |
| Psathyrella subnuda (P. Karst.) A.H. Sm. | | √ | √ | | | | WS | Others | Agarics | |
| Pseudoclitocybe cyathiformis (Bull.) Singer | √ | | | | √ | √ | SS | Edible, Medicinal | Agarics | ON683485 |
| Pseudofavolus tenuis (Fr.) G. Cunn. | √ | | | √ | | | WS | Medicinal, Poisonous | PHT fungi | |
| Pseudohydnum gelatinosum (Scop.) P. Karst. | | | | √ | | | WS | Edible, Medicinal | Jelly fungi | |
| Pseudosperma avellaneum (Kobayasi) Matheny and Esteve-Rav. | √ | | | | | | SS | Others | Agarics | |
| Pseudosperma rimosum (Bull.) Matheny and Esteve-Rav. | √ | | √ | √ | √ | √ | SS | Others | Agarics | |
| Pseudosperma umbrinellum (Bres.) Matheny and Esteve-Rav. | | | | | | √ | SS | Others | Agarics | |
| Pterula multifida (Chevall.) Fr. | | √ | √ | | √ | √ | SS | Others | Coral fungi | ON683486 |
| Pycnoporus cinnabarinus (Jacq.) P. Karst. | √ | √ | | | | √ | WS | Medicinal | PHT fungi | |
| Pycnoporus sanguineus (L.) Murrill | | | | | √ | | WS | Medicinal | PHT fungi | |
| Radulodon copelandii (Pat.) N. Maek. | | | √ | | | √ | WS | Others | PHT fungi | |
| Radulomyces copelandii (Pat.) Hjortstam and Spooner | | √ | √ | | | | WS | Others | PHT fungi | |
| Ramaria apiculata (Fr.) Donk | √ | √ | √ | | √ | √ | EM | Edible, Medicinal | Coral fungi | |
| Ramaria botrytis (Pers.) Bourdot | | | | | √ | | EM | Edible, Medicinal | Coral fungi | |
| Ramaria bourdotiana Maire | | | √ | | | | EM | Edible | Coral fungi | |
| Ramaria flava (Schaeff.) Quél. | √ | | √ | | √ | √ | EM | Edible, Medicinal, Poisonous | Coral fungi | |
| Ramaria formosa (Pers.) Quél. | | | √ | | | | EM | Edible, Medicinal, Poisonous | Coral fungi | |
| Ramaria lutea Schild | | | | | | √ | EM | Edible | Coral fungi | |
| Ramaria madagascariensis (Henn.) Corner | | | √ | | | | EM | Others | Coral fungi | |
| Ramaria stricta (Pers.) Quél. | √ | | √ | | √ | √ | EM | Edible | Coral fungi | |
| Ramaria subbotrytis (Coker) Corner | | | √ | | | | EM | Edible | Coral fungi | |
| Ramariopsis kunzei (Fr.) Corner | | | | | | √ | EM | Edible | Coral fungi | |
| Resupinatus applicatus (Batsch) Gray | | √ | | | | √ | WS | Others | Agarics | |
| Rhizocybe vermicularis (Fr.) Vizzini, P. Alvarado, G. Moreno, and Consiglio | | | √ | | | | SS | Others | Agarics | |
| Rhizomarasmius undatus (Berk.) R.H. Petersen | | | | | | √ | SS | Others | Agarics | |
| Rhodocollybia butyracea (Bull.) Lennox | | √ | | | | | SS | Edible | Agarics | |
| Rhodocollybia prolixa (Fr.) Antonín and Noordel. | | | | | | √ | SS | Others | Agarics | |
| Rickenella fibula (Bull.) Raithelh. | √ | | √ | | | | EM | Others | Cantharelloid fungi | |
| Ripartites tricholoma (Alb. and Schwein.) P. Karst. | √ | | | | | | LS | Others | Agarics | ON683428 |
| Russula adusta (Pers.) Fr. | √ | | | | | | EM | Edible, Medicinal | Agarics | |
| Russula aeruginea Lindblad ex Fr. | √ | | √ | | √ | √ | EM | Edible | Agarics | |
| Russula albida A. Blytt | √ | | √ | | | | EM | Edible | Agarics | |
| Russula alutacea (Fr.) Fr. | | | | | √ | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Russula amoena Quél. | √ | | √ | √ | | √ | EM | Others | Agarics | |
| Russula aurata Fr. | | | √ | | √ | | EM | Others | Agarics | |
| Russula aurea Pers. | √ | | | | | | EM | Others | Agarics | |
| Russula chloroides (Krombh.) Bres. | √ | | √ | | | | EM | Edible | Agarics | |
| Russula crustosa Peck | √ | | √ | | | | EM | Edible, Medicinal | Agarics | ON683429 |
| Russula cyanoxantha (Schaeff.) Fr. | √ | | √ | | | √ | EM | Edible, Medicinal | Agarics | ON683430 |
| Russula delica Fr. | √ | | | | √ | √ | EM | Edible, Medicinal | Agarics | |
| Russula densifolia Secr. ex Gillet | | | √ | | | √ | EM | Edible, Medicinal | Agarics | ON683487 |
| Russula emetica (Schaeff.) Pers. | √ | | √ | | √ | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Russula exalbicans (Pers.) Melzer and Zvára | | | | √ | | | EM | Edible | Agarics | |
| Russula faginea Romagn. | | | √ | | | | EM | Edible | Agarics | |
| Russula flavida Frost ex Peck | | | √ | | √ | √ | EM | Poisonous | Agarics | |
| Russula foetens Pers. | √ | | √ | √ | √ | √ | EM | Medicinal, Poisonous | Agarics | ON683488 |
| Russula fragilis Fr. | √ | | √ | | | | EM | Medicinal, Poisonous | Agarics | ON683489 |
| Russula furcata Pers. | | | √ | | | | EM | Edible | Agarics | |
| Russula grata Britzelm. | | | √ | | √ | √ | EM | Others | Agarics | |
| Russula integra (L.) Fr. | √ | | √ | | | | EM | Edible, Medicinal | Agarics | |
| Russula lilacea Quél. | √ | | | √ | | | EM | Edible, Medicinal | Agarics | |
| Russula mariae Peck | | | | | | √ | EM | Edible | Agarics | |
| Russula mustelina Fr. | √ | | | | √ | | EM | Edible | Agarics | |
| Russula nauseosa (Pers.) Fr. | √ | | √ | | | | EM | Edible | Agarics | |
| Russula paludosa Britzelm. | | | √ | √ | | | EM | Edible | Agarics | |
| Russula pectinata Fr. | | | | | √ | | EM | Poisonous | Agarics | |
| Russula pseudodelica J.E. Lange | √ | | √ | | | | EM | Edible, Medicinal | Agarics | |
| Russula puellaris Fr. | √ | | √ | | | | EM | Edible | Agarics | |
| Russula pungens Beardslee | | | | | | √ | EM | Others | Agarics | |
| Russula risigallina (Batsch) Sacc. | √ | | | √ | | | EM | Edible | Agarics | |
| Russula rosea Pers. | √ | | | | | | EM | Edible, Medicinal | Agarics | |
| Russula rubra (Lam.) Fr. | √ | | √ | | | | EM | Edible | Agarics | |
| Russula sanguinaria (Schumach.) Rauschert | √ | | √ | √ | | | EM | Edible | Agarics | |
| Russula sanguinea Fr. | √ | | | | √ | √ | EM | Others | Agarics | |
| Russula sororia (Fr.) Romell | | | | √ | √ | √ | EM | Medicinal | Agarics | ON683431 |
| Russula squalida Peck | | | | √ | | | EM | Others | Agarics | |
| Russula subdepallens Peck | √ | | √ | | | | EM | Edible | Agarics | |
| Russula vinosa Lindblad | | | | √ | | | EM | Others | Agarics | ON683432 |
| Sarcodontia spumea (Sowerby) Spirin | √ | √ | √ | √ | √ | | EM | Others | PHT fungi | |
| Sarcomyxa edulis (Y.C. Dai, Niemelä, and G.F. Qin) T. Saito, Tonouchi, and T. Harada | √ | √ | | | √ | | WS | Medicinal | Agarics | |
| Sarcoscypha coccinea (Gray) Boud. | | √ | √ | √ | √ | | WS | Poisonous | Larger Ascomycetes | |
| Schizophyllum commune Fr. | √ | √ | √ | √ | √ | √ | WS | Edible, Medicinal | Agarics | |
| Scleroderma areolatum Ehrenb. | | √ | √ | | | √ | SS | Edible, Medicinal, Poisonous | Gasteroid fungi | |
| Scleroderma bovista Fr. | | √ | √ | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Scleroderma polyrhizum (J.F. Gmel.) Pers. | | | | | √ | | SS | Edible, Medicinal | Gasteroid fungi | |
| Scutellinia pseudovitreola W.Y. Zhuang and Zhu L. Yang | | √ | | | | | WS | Others | Larger Ascomycetes | |
| Scutellinia scutellata (L.) Lambotte | √ | √ | √ | | √ | √ | WS | Others | Larger Ascomycetes | |
| Sparassis latifolia Y.C. Dai and Zheng Wang | | √ | | | | | WS | Edible, Medicinal | PHT fungi | ON683490 |
| Spathularia flavida Pers. | | | | √ | √ | √ | SS | Others | Larger Ascomycetes | |
| Sphaerobolus stellatus Tode | √ | | | | | √ | WS | Others | Gasteroid fungi | |
| Spongiporus zebra (Y.L. Wei and W.M. Qin) B.K. Cui, L.L. Shen, and Y.C. Dai | √ | | | | | √ | WS | Others | PHT fungi | |
| Steccherinum ochraceum (Pers. ex J.F. Gmel.) Gray | | | √ | | | | WS | Others | PHT fungi | |
| Steccherinum rawakense (Pers.) Banker | √ | | | √ | | | WS | Others | PHT fungi | |
| Stereum hirsutum (Willd.) Pers. | √ | √ | | | √ | √ | WS | Medicinal | PHT fungi | |
| Stereum rugosum Pers. | √ | | | | √ | | WS | Others | PHT fungi | |
| Stereum subtomentosum Pouzar | | √ | | | | | WS | Others | PHT fungi | |
| Stereum ostrea (Blume and T. Nees) Fr. | √ | √ | √ | | √ | | WS | Others | PHT fungi | |
| Strobilurus stephanocystis (Kühner and Romagn. ex Hora) Singer | | | | | | √ | WS | Others | Agarics | |
| Stropharia aeruginosa (Curtis) Quél. | | √ | √ | | √ | | SS | Edible, Poisonous | Agarics | |
| Stropharia rugosoannulata Farl. ex Murrill | √ | √ | | | √ | | SS | Edible, Medicinal | Agarics | |
| Suillellus luridus (Schaeff.) Murrill | | | | | √ | | EM | Edible, Medicinal | Boletes | |
| Suillus bovinus (L.) Roussel | | | | | √ | √ | EM | Edible, Medicinal, Poisonous | Boletes | |
| Suillus flavus (Quél.) Singer | √ | | | | √ | | EM | Others | Boletes | |
| Suillus granulatus (L.) Roussel | √ | | √ | √ | √ | √ | EM | Edible, Medicinal, Poisonous | Boletes | ON683433 |
| Suillus grevillei (Klotzsch) Singer | √ | | | √ | √ | √ | EM | Edible, Medicinal | Boletes | |
| Suillus lactifluus (With.) A.H. Sm. and Thiers | | | | | | √ | EM | Edible | Boletes | |
| Suillus laricinus (Berk.) Kuntze | | | | | √ | √ | EM | Others | Boletes | |
| Suillus luteus (L.) Roussel | √ | | | | √ | | EM | Edible, Medicinal, Poisonous | Boletes | |
| Suillus spraguei (Berk. and M.A. Curtis) Kuntze | √ | | | | | | EM | Others | Boletes | |
| Suillus subaureus (Peck) Snell | | | √ | | | | EM | Edible, Medicinal | Boletes | |
| Suillus viscidus (L.) Roussel | √ | | | | √ | | EM | Edible, Medicinal | Boletes | |
| Tapinella atrotomentosa (Batsch) Šutara | | | √ | | √ | √ | EM | Medicinal, Poisonous | Boletes | |
| Tapinella panuoides (Fr.) E.-J. Gilbert | | √ | | √ | | | EM | Poisonous | Boletes | |
| Terana caerulea (Lam.) Kuntze | | | √ | | | √ | WS | Others | PHT fungi | |
| Tetrapyrgos nigripes (Fr.) E. Horak | √ | √ | | | | √ | WS | Others | Agarics | |
| Thelephora anthocephala (Bull.) Fr. | | | √ | √ | | | SS | Others | PHT fungi | |
| Thelephora palmata (Scop.) Fr. | | | | | √ | √ | SS | Others | PHT fungi | |
| Tolypocladium capitatum (Holmsk.) C.A. Quandt, Kepler, and Spatafora | | | | | | √ | EI | Others | Larger Ascomycetes | |
| Trametes coccinea (Fr.) Hai J. Li and S.H. He | | √ | √ | | | | WS | Others | PHT fungi | |
| Trametes conchifer (Schwein.) Pilát | | √ | | √ | | | WS | Others | PHT fungi | |
| Trametes gibbosa (Pers.) Fr. | √ | | | | √ | | WS | Medicinal | PHT fungi | ON683491 |
| Trametes hirsuta (Wulfen) Lloyd | | | √ | | √ | √ | WS | Medicinal | PHT fungi | |
| Trametes membranacea (Sw.) Kreisel | | | | | | √ | WS | Others | PHT fungi | |
| Trametes pubescens (Schumach.) Pilát | √ | | | √ | | | WS | Edible, Medicinal, Poisonous | PHT fungi | |
| Trametes suaveolens (L.) Fr. | √ | | | | √ | √ | WS | Medicinal | PHT fungi | |
| Trametes trogi Berk. | | | | | √ | | WS | Poisonous | PHT fungi | |
| Trametes versicolor (L.) Lloyd | √ | √ | √ | √ | | √ | WS | Medicinal | PHT fungi | ON683492 |
| Tremella aurantia Schwein. | | | | | √ | | WS | Edible | Jelly fungi | |
| Tremella foliacea Pers. | | √ | | | | | WS | Others | Jelly fungi | ON683493 |
| Tremella fuciformis Berk. | | √ | √ | | | | WS | Edible, Medicinal | Boletes | |
| Tremella mesenterica (Schaeff.) Pers. | √ | √ | | | | √ | WS | Edible, Medicinal | Jelly fungi | |
| Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden | √ | | | √ | | | WS | Medicinal | PHT fungi | |
| Trichaptum biforme (Fr.) Ryvarden | | | √ | | | | WS | Medicinal | PHT fungi | |
| Trichaptum pargamenum (Fr.) G. Cunn. | | √ | | | | √ | WS | Others | PHT fungi | |
| Tricholoma acerbum (Bull.) Quél. | √ | | | | | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Tricholoma album (Schaeff.) P. Kumm. | √ | | √ | | | | EM | Edible, Medicinal, Poisonous | Agarics | |
| Tricholoma aurantium (Schaeff.) Ricken | √ | | | | √ | | EM | Others | Agarics | |
| Tricholoma equestre (L.) P. Kumm. | | | | √ | | | EM | Edible, Poisonous | Agarics | |
| Tricholoma matsutake (S. Ito and S. Imai) Singer | √ | √ | | | | | EM | Edible, Medicinal | Agarics | |
| Tricholoma scalpturatum (Fr.) Quél. | | | | √ | | | EM | Edible, Poisonous | Agarics | |
| Tricholoma terreum (Schaeff.) P. Kumm. | | | √ | | √ | √ | EM | Others | Agarics | |
| Tricholoma tigrinum (Schaeff.) Gillet | √ | | | √ | | | EM | Poisonous | Agarics | |
| Tricholoma vaccinum (Schaeff.) P. Kumm. | √ | | | √ | | | EM | Edible, Medicinal | Agarics | |
| Tricholomopsis decora (Fr.) Singer | √ | | | | | | WS | Edible | Agarics | |
| Tricholomopsis rutilans (Schaeff.) Singer | √ | √ | √ | √ | √ | √ | WS | Poisonous | Agarics | |
| Tulostoma bonianum Pat. | | | √ | | √ | √ | SS | Others | Gasteroid fungi | |
| Turbinellus floccosus (Schwein.) Earle ex Giachini and Castellano | | | √ | √ | | | EM | Poisonous | Cantharelloid fungi | |
| Verpa bohemica (Krombh.) J. Schröt. | | | | | | √ | SS | Edible, Medicinal, Poisonous | Larger Ascomycetes | |
| Verpa digitaliformis Pers. | | | | | | √ | SS | Edible | Larger Ascomycetes | |
| Vitreoporus dichrous (Fr.) Zmitr. | | | | | √ | | WS | Others | PHT fungi | |
| Volvariella bombycina (Schaeff.) Singer | | | √ | | √ | | WS | Edible, Medicinal | Agarics | |
| Volvariella pusilla (Pers.) Singer | | | | | | √ | SS | Edible, Medicinal | Agarics | |
| Volvopluteus gloiocephalus (DC.) Vizzini, Contu and Justo | | | √ | | √ | | SS | Poisonous | Agarics | |
| Xanthochrous gilvicolor (Lloyd) Teng | √ | | | | | √ | WS | Others | PHT fungi | |
| Xerocomellus chrysenteron (Bull.) Šutara | | | | | √ | | EM | Others | Boletes | |
| Xerocomus chrysenteron (Bull.) Quél. | √ | | | | √ | √ | EM | Others | Boletes | |
| Xeromphalina campanella (Batsch) Kühner and Maire | √ | √ | √ | √ | √ | | WS | Medicinal | Agarics | |
| Xerula pudens (Pers.) Singer | √ | | | | | | WS | Others | Agarics | |
| Xylaria carpophila (Pers.) Fr. | | | √ | | | √ | WS | Medicinal | Larger Ascomycetes | |
| Xylaria hypoxylon (L.) Grev. | √ | √ | √ | | √ | √ | WS | Others | Larger Ascomycetes | |
| Xylaria polymorpha (Pers.) Grev. | | √ | √ | √ | | | WS | Others | Larger Ascomycetes | |


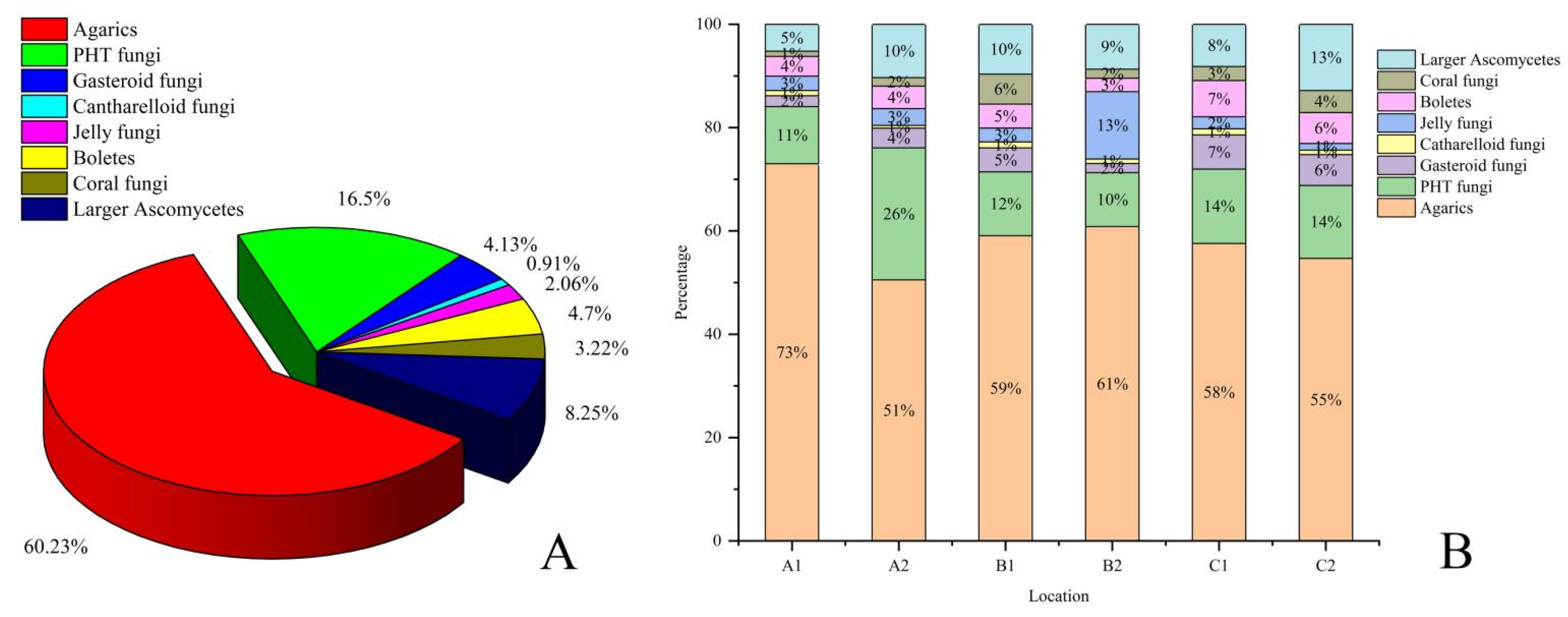
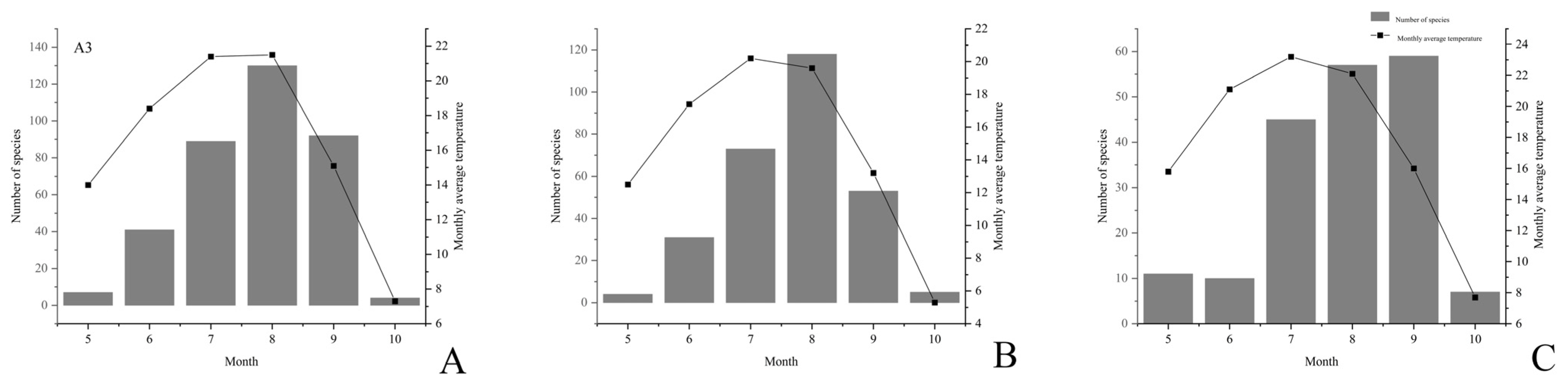
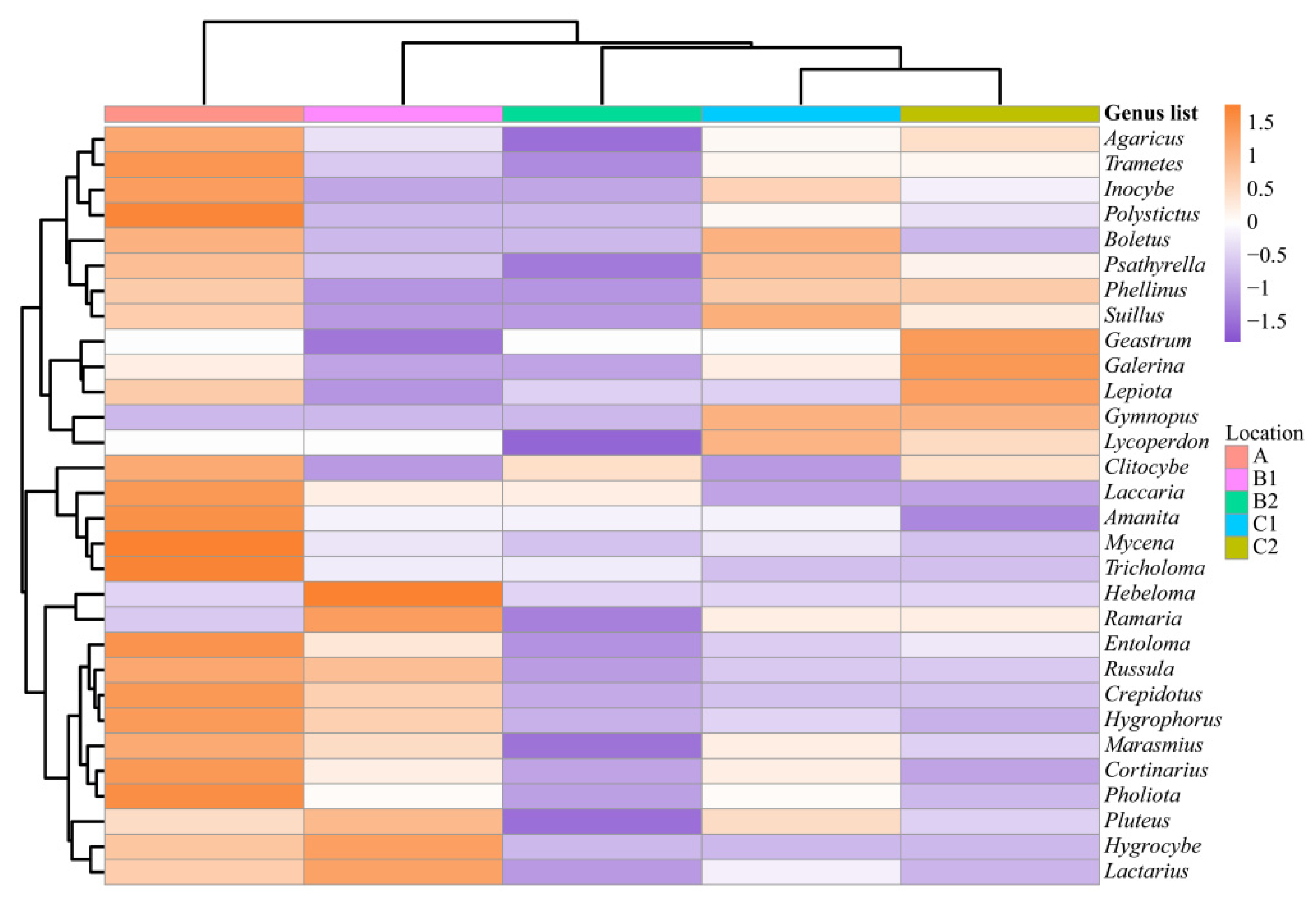
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













